



www.bba-direct.com
Biochimica et Biophysica Acta 1640 (2003) 105–112
Effect of tributyltin on trout blood cells: changes in mitochondrial
morphology and functionality
Luca Tianoa,*, Donatella Fedelia, Giorgio Santonib, Ian Daviesc, Giancarlo Falcionia
aDepartment of Biology MCA, University of Camerino, Camerino (MC), ItalybDepartment of Pharmacology and Experimental Medicine, University of Camerino, Camerino (MC), Italy
cFisheries Research Services, Marine Laboratory, PO Box 101, 375 Victoria Road, Aberdeen AB11 9DB, UK
Received 16 May 2002; received in revised form 19 November 2002; accepted 14 February 2003
Abstract
The aquatic environment is the largest sink for the highly toxic organotin compounds, particularly as one of the main sources is the direct
release of organotins from marine antifouling paints. The aim of this study was to investigate the mitochondrial toxicity and proapoptotic
activity of tributyltin chloride (TBTC) in teleost leukocytes and nucleated erythrocytes, by means of electron microscopy investigation and
mitochondrial membrane potential evaluation, in order to provide an early indicator of aquatic environmental pollution. Erythrocytes and
leukocytes were obtained from an inbred strain of rainbow trout (Oncorhynchus mykiss). Transmission electronic micrographs of trout red
blood cells (RBC) incubated in the presence of TBTC at 1 and 5 AM for 60 min showed remarkable mitochondrial morphological changes.
TBTC-mediated toxicity involved alteration of the cristae ultrastructure and mitochondrial swelling, in a dose-dependent manner. Both
erythrocytes and leukocytes displayed a consistent drop in mitochondrial membrane potential following TBTC exposure at concentrations >1
AM. The proapoptotic effect of TBTC on fish blood cells, and involvement of mitochondrial pathways was also investigated by verifying the
release of cytochrome c, activation of caspase-3 and the presence of ‘‘DNA laddering’’. Although mitochondrial activity was much more
strongly affected in erythrocytes, leukocytes incubated in the presence of TBTC showed the characteristic features of apoptosis after only 1 h
of incubation. Longer exposures, up to 12 h, were required to trigger an apoptotic response in erythrocytes.
Crown Copyright D 2003 Published by Elsevier Science B.V. All rights reserved.
Keywords: Tributyltin chloride; Organotin compound; Rainbow trout
1. Introduction
Organotin compounds are pollutants of primarily anthro-
pogenic origin [1]. Their presence in the environment is due
to their use in many industrial applications [2] and as
agricultural biocides. Originally, organotin compounds (par-
ticularly butylated tin compounds) were developed as ther-
mal stabilisers in the synthesis of chlorinated polymers such
as PVC. However, their biocidal properties lead to new uses.
As a consequence of this expansion, concern increased over
their possible environmental and health effects.
The aquatic environment represents the largest sink for
accumulation of these xenobiotics, particularly from their
use in marine antifouling paints. Alkyltin compounds are a
0167-4889/03/$ - see front matter. Crown Copyright D 2003 Published by Elsev
doi:10.1016/S0167-4889(03)00025-9
* Corresponding author. Tel.: +39-737-403-208; fax: +39-737-636-
216.
E-mail address: [email protected] (L. Tiano).
significant hazard to aquatic organisms, through neurotoxic,
hepatotoxic, immunosuppressive and hormone disruptive
activities [3].
In the last few years, we have investigated the effects of
different organotin compounds on aquatic biota, using
nucleated trout erythrocytes as an experimental model [4].
As well as playing a central role in the physiology of fish
respiration, these cells represent an outstanding model to
study xenobiotic-induced damage to different cellular com-
partments. Despite their structural simplicity, the erythro-
cytes of lower vertebrates preserve the nucleus and mito-
chondria, unlike the anucleated erythrocytes of mammals.
The effects of alkyltin on trout red blood cells (RBCs) were
studied by monitoring the hemolytic process, by measuring
steady-state fluorescence anisotropy of different probes on
isolated membranes, by evaluating the stability of trout
hemoglobins and lastly by investigating their genotoxic
effects using the single-cell gel electrophoresis ‘‘Comet
Assay’’. The results obtained [5,6] indicated that incubation
ier Science B.V. All rights reserved.

L. Tiano et al. / Biochimica et Biophy106
in the presence of triorganotin induced a plasma membrane
perturbation and accelerated the precipitation process of Hb
IV, which represent 60% of the whole pigment and is
characterised by pH-dependent oxygen affinity (Root effect)
[7]. Moreover, experimental data demonstrated the ability of
tributyltin chloride (TBTC) to readily induce DNA single-
strand breaks [8]. Although DNA damage in nucleated
erythrocytes can be related to a metHb-dependent (i.e.
product of Hb oxidation or autoxidation) oxidative stress
[9], this was not the case in TBTC-mediated DNA damage.
In fact, single-strand breaks were not prevented by stabili-
sation of Hb in the carbomonoxy derivative.
In this paper, we report an attempt to investigate in
greater detail the mechanism of toxicity of alkyltin com-
pounds, particularly the effects of these xenobiotics on
mitochondria, because of the key role of these organelles
in the mechanism of cell death [10–15]. This may be due
to mitochondria being both the source and the final target
of free radicals. Mitochondrial ultrastructure was inves-
tigated using transmission electron microscopy (TEM).
Mitochondrial functionality was also monitored, at the
single-cell level, by means of flow cytometric mitochon-
drial membrane potential (Dcm) determination using the
fluorescent probe JC-1 [16–20], a delocalized lipophilic
cation which accumulates in charged membranes. JC-1
has advantages over other potential-sensitive probes, such
as rhodamines and other carbocyanines, since it changes
colour from green to orange as the membrane potential
increases to values greater than about 80–100 mV. This
property arises from the reversible formation of JC-1
aggregates on membrane polarisation, which causes shifts
in the emitted light from 530 nm (i.e. emission of JC-1
monomeric form) to 590 nm (i.e. emission of J-aggre-
gate).
Mitochondria-related features of apoptosis, such as cyto-
chrome c release and caspase-3 activation were also eval-
uated by means of Western blot analysis. The aim of this
study was to follow the effects of organotin compounds, at
sublethal concentrations, on mitochondria of peripheral
blood cells in relation to their involvement in triggering
DNA fragmentation and apoptotic cell death. The results
provide new information on the effect of organotins in
inducing apoptosis, and could be useful for developing an
early biosensor for aquatic pollution.
2. Materials and methods
All reagents were of analytical grade. Organotin com-
pounds were obtained from Aldrich, lymphoprep for sepa-
ration of RBCs and leukocytes was obtained from (Nycomed
Pharma AS, Oslo, Norway) and JC-1 was purchased from
Molecular Probes (Eugene, OR) and stored at � 20 jC as a
1-mM stock solution in DMSO. Cell culture media and
reagents were obtained from GIBCO, and molecular biology
reagents were obtained from SIGMA.
2.1. Samples
The cells used in this study were obtained from rainbow
trout (Oncorhynchus mykiss). The fish were kept in tanks
containing water from the Scarsito River, Italy, and fed
with commercial fish food. Experiments were performed
using blood from fish of the same age (f 24 months old),
and weighing between 180 and 300 g. Blood was with-
drawn from the lateral tail vein with a syringe into an
isotonic medium (0.1 M phosphate buffer, 0.1 M NaCl,
0.2% citrate, 1 mM EDTA, pH 7.8). Blood was held at
4jC and any further treatment was conducted within 2 h.
The whole blood was diluted 1:1 with the isotonic buffer
and this suspension was stratified on a solution of lym-
phoprep prior to being centrifuged for 20 min at 3000 rpm
at 4 jC. Peripheral blood lymphocytes (PBLs) were
separated from erythrocytes and both were washed with
RPMI medium. RBC and PBL suspensions were adjusted
to a density of 3.1�106 cells/ml in complete medium
supplemented with 10% foetal calf serum and incubated
for 1 h at 15 jC in the absence (control) or presence of
TBTC. A second set of experiments was conducted by
prolonging the incubation of RBCs up to 12 h at 15 jC in
the presence of TBTC at 0.1 and 1 AM. Organotin
compounds dissolved in 100% ethanol were added to the
erythrocytes (10 Al/ml of erythrocyte suspension) to final
concentrations of 0.1, 0.5, 1, 5 and 10 AM. Ten micro-
meters was used as the maximum concentration because,
in our experience, both hemolysis and met-Hb formation
are virtually absent at this concentration. Solvent control
experiments were performed by incubating the cells with
medium containing only ethanol.
An aliquot of treated erythrocytes was fixed in 2.5%
glutaraldehyde/0.1 M sodium cacodylate buffer pH 7.4 for 4
h at 4 jC, and postfixed in 1% osmium tetroxide in distilled
water for 1 h at room temperature. Cells were subsequently
dehydrated in ethanol and embedded in TAAB embedding
resin (medium grade). Ultrathin sections were cut using a
Reichert-Jung Ultracut ultramicrotome and then stained
with uranyl acetate and lead citrate. Stained sections were
observed using a Philips 301 transmission electron micro-
scope at 80 kV.
After exposure to pollutants, the erythrocyte suspension
was adjusted to a density of 1.5� 105 cells/ml and
incubated with 10 Ag/ml of JC-1 for 10 min at room
temperature in the dark for Dcm determination. A suspen-
sion of 1�106 cells/ml from each subfraction was ana-
lysed for relative fluorescence intensity using a FACScan
flow cytometer (Becton Dickinson, Mountain View, CA,
USA) equipped with a single 488 argon laser. The filter in
front of the fluorescence 1 (FL1) photomultiplier transmits
at 530 nm, and the filter used in the FL2 channel transmits
at 617 nm. The values of PMT were logarithmically set.
Red fluorescence (FL2) corresponds to the J-aggregate
form of JC-1 and is proportional to Dcm. Compensation
FL1–FL2 was 4% and compensation FL2–FL1 9.5%. A
sica Acta 1640 (2003) 105–112

L. Tiano et al. / Biochimica et Biophysica Acta 1640 (2003) 105–112 107
minimum of 13,000 cells per sample were acquired and
analysed using WINMDI 2.8 software on an IBM compat-
ible computer.
For Western blot analysis, leukocytes and erythrocytes
were treated with organotin 1 AM at 15 jC for up to 12 h.
Cells (1.5� 107) were washed once in 5 ml ice-cold phos-
phate buffer saline (PBS). For the determination of cyto-
chrome c release, cells were further centrifuged and the pellet
was resuspended in 250 Al of ice-cold homogenisation buffer
(20 mM Hepes pH 7.4, 10 mM KCl, 1.5 mM MgCl2, 1 mM
EDTA, 1 mM EGTA, 1 mM DTT) supplemented with
protease inhibitors (0.1 mM PMSF, 5 Ag/ml pepstatine, 10
Ag/ml leupeptine, 2 Ag/ml aprotinine). After being kept on ice
for 15 min, cells were lysed by douncing 20 times in an
Eppendorf douncer. After centrifugation at 1000� g for 5
min at 4 jC, the supernatants were further ultracentrifuged at24 c (90,000� g) for 40 min at 4 jC in an airfuge ultra-
centrifuge (Beckman). For caspase-3 immunodetection, after
Fig. 1. Transmission electronic micrographs of mitochondria from trout erythrocyt
foetal calf serum at 15 jC and in the presence of 1 AM (c, d), or 5 AM (e, f) TBTC
f = 0.4 Am; d = 2.7 Am.
washing in ice-cold PBS, 1.5� 107 cells were lysed in 200 Alof RIPA buffer (0.1% nonidet-p40, 1 mM CaCl2, 1 mM
MgCl2, 0.1% sodium azide, 1 mM PMSF, 0.03 mg/ml
aprotinin, 1 mM NaVO4). Protein concentration in lysates
was measured using the Bio-Rad protein assay with albumin
as a standard. Protein extracts (20 Ag) were loaded onto 14%SDS polyacrilamide gels, electrophoresed at 200 V and than
transferred to immobilion-p membranes (Millipore, Bedford,
MA, USA) at 250 mA for 1 h. Membranes were blocked in
5% BSA and than incubated with a mouse anti-cytochrome c
monoclonal antibody (1/400) or mouse anti-caspase-3 mono-
clonal antibody (1/500) (Medical and Biological Lab, Japan)
overnight at 4 jC, followed by incubation with HRP-con-
jugated sheep antimouse antibody (1/10,000) (Amersham).
Immunoreactivity was detected with the enhanced chemilu-
minescence system (Amersham) using the CL detector
Chemidoc (Bio-Rad) and analysis software Quantity One
(Bio-Rad). As a reference for the amount of proteins, blots
es. Control (a, b), erythrocytes suspended in RPMI supplemented with 10%
, for 1 h. N, nucleus; m, mitochondria; pm, plasma membrane. Bar a, b, c, e,
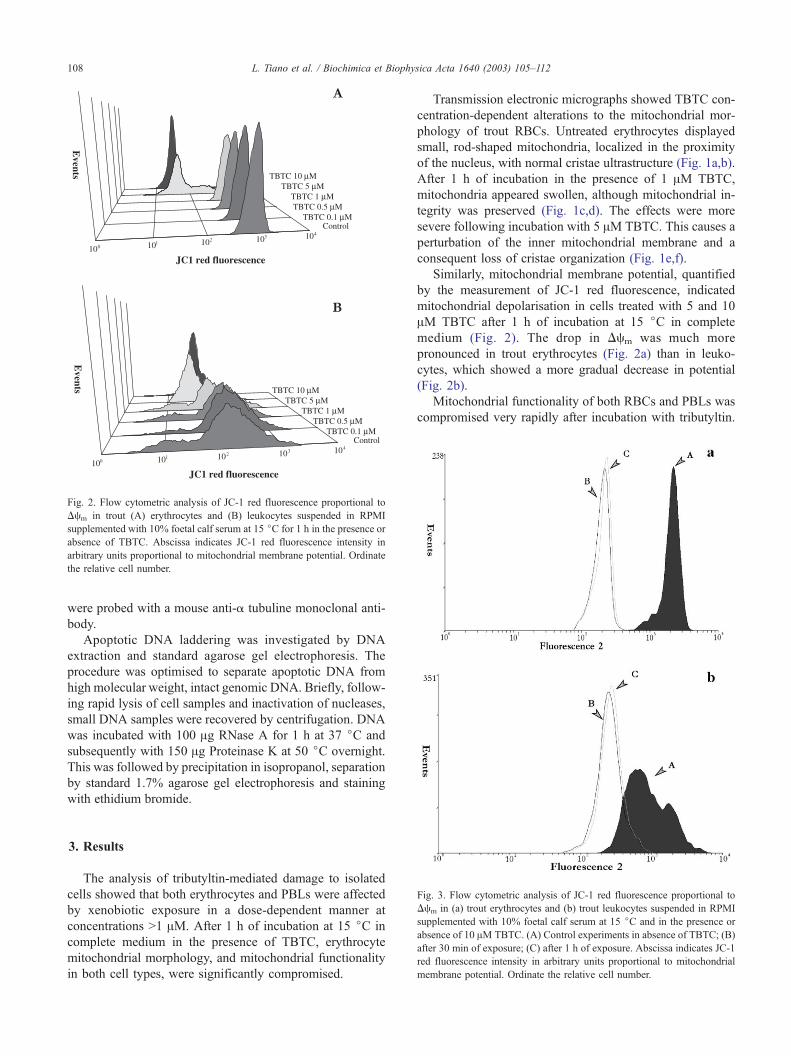
Fig. 2. Flow cytometric analysis of JC-1 red fluorescence proportional to
Dcm in trout (A) erythrocytes and (B) leukocytes suspended in RPMI
supplemented with 10% foetal calf serum at 15 jC for 1 h in the presence or
absence of TBTC. Abscissa indicates JC-1 red fluorescence intensity in
arbitrary units proportional to mitochondrial membrane potential. Ordinate
the relative cell number.
L. Tiano et al. / Biochimica et Biophysica Acta 1640 (2003) 105–112108
were probed with a mouse anti-a tubuline monoclonal anti-
body.
Apoptotic DNA laddering was investigated by DNA
extraction and standard agarose gel electrophoresis. The
procedure was optimised to separate apoptotic DNA from
high molecular weight, intact genomic DNA. Briefly, follow-
ing rapid lysis of cell samples and inactivation of nucleases,
small DNA samples were recovered by centrifugation. DNA
was incubated with 100 Ag RNase A for 1 h at 37 jC and
subsequently with 150 Ag Proteinase K at 50 jC overnight.
This was followed by precipitation in isopropanol, separation
by standard 1.7% agarose gel electrophoresis and staining
with ethidium bromide.
Fig. 3. Flow cytometric analysis of JC-1 red fluorescence proportional to
Dcm in (a) trout erythrocytes and (b) trout leukocytes suspended in RPMI
supplemented with 10% foetal calf serum at 15 jC and in the presence or
absence of 10 AM TBTC. (A) Control experiments in absence of TBTC; (B)
after 30 min of exposure; (C) after 1 h of exposure. Abscissa indicates JC-1
red fluorescence intensity in arbitrary units proportional to mitochondrial
membrane potential. Ordinate the relative cell number.
3. Results
The analysis of tributyltin-mediated damage to isolated
cells showed that both erythrocytes and PBLs were affected
by xenobiotic exposure in a dose-dependent manner at
concentrations >1 AM. After 1 h of incubation at 15 jC in
complete medium in the presence of TBTC, erythrocyte
mitochondrial morphology, and mitochondrial functionality
in both cell types, were significantly compromised.
Transmission electronic micrographs showed TBTC con-
centration-dependent alterations to the mitochondrial mor-
phology of trout RBCs. Untreated erythrocytes displayed
small, rod-shaped mitochondria, localized in the proximity
of the nucleus, with normal cristae ultrastructure (Fig. 1a,b).
After 1 h of incubation in the presence of 1 AM TBTC,
mitochondria appeared swollen, although mitochondrial in-
tegrity was preserved (Fig. 1c,d). The effects were more
severe following incubation with 5 AM TBTC. This causes a
perturbation of the inner mitochondrial membrane and a
consequent loss of cristae organization (Fig. 1e,f).
Similarly, mitochondrial membrane potential, quantified
by the measurement of JC-1 red fluorescence, indicated
mitochondrial depolarisation in cells treated with 5 and 10
AM TBTC after 1 h of incubation at 15 jC in complete
medium (Fig. 2). The drop in Dcm was much more
pronounced in trout erythrocytes (Fig. 2a) than in leuko-
cytes, which showed a more gradual decrease in potential
(Fig. 2b).
Mitochondrial functionality of both RBCs and PBLs was
compromised very rapidly after incubation with tributyltin.

Fig. 4. Increased release of cytochrome c to the cytosol in trout erythrocytes and leukocytes upon apoptotic stimulation with TBTC 1 AM in RPMI medium at
15 jC up to 6 and 12 h for leukocytes and erythrocytes, respectively. After ultracentrifugation normalized samples were subjected to 14% SDS-PAGE and
transferred to nitrocellulose filter. The filter was probed with a monoclonal anti-cytochrome c antibody. As a reference for the amount of proteins, blots were
probed with a mouse anti-a tubuline monoclonal antibody as described in Materials and methods.
L. Tiano et al. / Biochimica et Biophysica Acta 1640 (2003) 105–112 109
There were no significant changes in Dcm between 30 and
60 min incubation with 10 AM TBTC (Fig. 3).
Mitochondrial impairment and mitochondrial membrane
depolarisation, clearly evident in trout erythrocytes in our
experimental conditions, represent early events in apopto-
sis. It has been shown in isolated mitochondria [21] and
Jurkat cells [22], that TBTC induced a rapid loss of Dcm,
which is associated with the release of cytochrome c,
important for subsequent apoptosis. Here, we tested
whether TBTC induced the release of cytochrome c from
mitochondrial membrane into the cytosol of erythrocytes
and leukocytes. As shown by Western blot analysis of
cytosolic extracts, 1 AM TBTC induced cytochrome c
release in both cell types (Fig. 4). Interestingly, the release
of cytochrome c exhibited different kinetics in the two
cell types: in leukocytes cytosolic cytochrome c content
increased significantly after 2 h of incubation and peaked
after 6 h of exposure, whereas in erythrocytes a gradual
increase was detected up to 12 h of exposure. In accord-
ance with cytochrome c levels, caspase-3 activation was
sustained only 12 h after exposure of trout erythrocytes
(Fig. 5). In mammalian cells, caspase-3 is synthesised as a
32 kDa zymogen that is processed to mature 20/17 kDa
and 12 kDa subunits by cleavage at Asp9, Asp28 and
Asp175 [23]. In trout erythrocytes we found that the size
Fig. 5. Caspase-3 activation in trout erythrocytes by TBTC 1 AM. Cells
were incubated in complete medium at 15 jC in the presence of TBTC,
harvested at different time periods and lysed. Normalized samples were
subjected to 14% SDS-PAGE and transferred to nitrocellulose filter. The
filter was probed with a monoclonal anti-caspase-3 antibody, as described
in Materials and methods. Caspase-3 is synthesised as a 27 kDa zymogen
that is processed to mature 16.8 kDa subunit.
of zymogen and activated caspase were slightly different
from those in mammals, 27 and 16.8 kDa, respectively.
This difference is not surprising taking into account that
sequences of caspase-3 reported for other teleost (Acces-
sion No. BAC00949.1, BAC00948.1) are significantly
smaller than for mouse caspase-3. Nevertheless, the spe-
cificity of antibody reaction was tested on rat thymocyte
submitted to apoptotic stimuli (data not shown). Finally,
we isolated the low molecular weight DNA fraction and
used this to verify the presence of internucleosomal cleav-
age, a characteristic late feature of apoptosis. After 1 h of
incubation in the presence of TBTC, PBLs displayed DNA
laddering at xenobiotic concentrations of 1 AM or greater
(Fig. 6a). In contrast, a fragmented, low molecular weight
DNA fraction was absent from RBCs exposed in the same
experimental conditions (Fig. 6b). Nevertheless, on a lon-
ger time scale, TBTC was able to trigger apoptotic cell
death in erythrocytes in accordance with cytochrome c, and
caspase-3 results. DNA laddering was evident after 8–12 h
Fig. 6. Agarose gel electrophoresis of low molecular weight DNA isolated
from (a) trout leukocytes and (b) erythrocytes treated for 1 h with TBTC at
0.1 AM (lane 1/1V), 0.5 AM (lane 2/2V), 1 AM (lane 3/3V), 5 AM (lane 4/4V), or10 AM (lane 5/5V). Exposures were conducted at 15 jC in RPMI
supplemented with 10% foetal calf serum complete medium. DNA markers
(M/MV) were electrophoresed as a base pair reference.

Fig. 7. Agarose gel electrophoresis of low molecular weight DNA isolated
from (a) trout erythrocytes exposed to 1 AM TBTC in complete medium at
15 jC for up to 12 h; lane 1 = 1 h, lane 2 = 4 h, lane 3 = 8 h, lane 4 = 12 h.
(b) DNA isolated from trout erythrocytes incubated in complete medium at
15 jC for 12 h in the presence or absence of TBTC; lane 1V= control, lane2V= 0.1 AM TBTC, lane 3V= 1 AM TBTC.
L. Tiano et al. / Biochimica et Biophysica Acta 1640 (2003) 105–112110
of incubation in the presence of 0.1 and 1 AM TBTC
(Fig. 7).
4. Discussion
In the present study, we evaluated the impairment of
mitochondrial functionality and its relationship to apop-
totic DNA fragmentation in fish blood cells following
exposure to TBTC, a highly toxic organotin compound.
Our data show that exposure to TBTC at sublethal
concentrations, in the range 5–10 AM, can induce rapid
and clear mitochondrial impairment in both erythrocytes
and leukocytes. This is in agreement with other reports
that indicate that trialkyltin compounds can inhibit oxida-
tive phosphorylation. This inhibition is provoked by high-
affinity binding of alkyltins to sites on the mitochondrial
membrane [24–26]. It is coupled with the inhibition of a
step in the energy transfer process [26], leading to
incomplete oxygen reduction and subsequent formation
of reactive oxygen species (ROS). Therefore, an increase
in the cellular level of oxygen free radicals following
TBTC-mitochondria interactions, as documented in liter-
ature in other cellular models [27], may be the cause of
the TBT-induced DNA damage previously reported by us
in fish erythrocytes [8].
The formation of ROS, and the impairment of mito-
chondria, are known to be involved in apoptotic cell death
[28]. Apoptotic effects of organotins are well-documented
for several cellular models [29–33], even though the
molecular mechanisms involved are still unclear. Several
mechanisms have been proposed and mainly concern
disturbance of Ca2 + homeostasis [34,35]. This is a basic
mode of action of alkyltins, whereby an increase in
cytosolic calcium can activate a number of intracellular
reactions including release of neurotransmitters, protein
phosphorylation, protease activity, and, eventually, cell
death. ROS have also been proposed as possible causes
of these effects [27,35,36].
TBTC (5–10 AM) was able to destabilise mitochondrial
functionality in both cell types. However, after 1 h of
exposure, TBTC induced apoptotic DNA laddering only
in leukocytes, whereas longer exposure times (8–12 h) to
TBTC < 1 AM were required to trigger apoptosis in eryth-
rocytes.
Mitochondria may represent the initial target of orga-
notins, but they seem to be involved differently in the
death response of different cell types. In PBLs, mitochon-
drial depolarisation was significant, but gradual, in the
population. Subsequent production of ROS, induction of
mitochondrial permeability transition and cytochrome c
release in the cytosol would be responsible for the
induction of apoptosis, as indicated by DNA laddering.
By contrast, TBTC-mediated mitochondrial toxicity in
RBCs was more pronounced, but the apoptotic cascade
was activated only at low concentrations of TBTC (V1 AM)
after several hours of incubation. This may suggest that, in
heavily contaminated environments, mitochondrial impair-
ment and DNA damage are not the principal effects of
organotins on RBCs. TBTC toxicity to RBCs is manifested
earlier by membrane perturbation leading to hemolysis. On
the contrary, lower concentrations of TBTC enable RBCs
to react and carry out the programmed cell death pro-
cess.
Differences in the kinetics of cell death are related to
early events in the apoptotic cascade, as indicated by the
differences in mitochondrial depolarisation. Taking into
account membrane-directed toxicity of TBTC and the differ-
ent physicochemical properties of membranes of the differ-
ent cell type studied, it is likely that the different behaviour
observed in erythrocytes and leukocytes could be due to
differences in the uptake of the triorganotin derivative by the
cellular membrane.
Organotin derivatives are assumed to be taken up across
the plasma membrane via passive diffusion processes by
partitioning into hydrophobic biological membranes. Phys-
icochemical properties of membranes play an important role
as indicated by differential accumulation in different cell
types [37] and in cells of different ages [38]. Consequently,
different amounts of alkyltin available in the cytosol may
affect mitochondria in different ways. This is in agreement
with recent observations [21] which indicate that TBTC
may effect mitochondria by two discrete mechanisms
depending on its concentration. At low concentrations,
TBT induces gradual loss of mitochondrial membrane
potential and mitochondrial swelling, whereas at higher
concentration a rapid induction of MPT was observed.
Moreover, at high concentrations of TBT, cells are forced
into a necrotic death as TBT inhibits the catalytic activity of
caspase-3 [22].

L. Tiano et al. / Biochimica et Biophysica Acta 1640 (2003) 105–112 111
In addition to the toxicological relevance of our obser-
vations, a particular consideration can be made of the
modalities of cell death in nucleated erythrocytes, in com-
parison to mammalian erythrocytes. In fact, as mature
erythrocytes lack cellular organelles and because they can
survive conditions that induce programmed cell death,
RBCs have been considered to be the only mammalian cells
that lack programmed cell death processes. However, the
senescence involved in erythrocyte death and removal is
characterised by distinct morphological changes very sim-
ilar to the morphological characteristics of programmed cell
death, including cell shrinkage, plasma membrane micro-
vesiculation and phosphatidilserine externalisation. Follow-
ing the recent work of Bratosin et al. [39] and Berg et al.
[40], aspects of the programmed death of mammalian
erythrocytes has been reinvestigated in light of the discov-
ery of components of the cell death machinery in mature
RBCs. Mammalian erythrocytes contain procaspase-3 and
procaspase-8 at levels comparable with those found in
nucleated cells. However, their role is ambiguous, since
erythrocytes lack other elements of the apoptotic machinery,
such as APAF 1 cytochrome c and caspase-2, -6, -7 and -9.
An alternative role has been proposed for caspase-3 in
mammalian RBCs as inhibitor of flippase activity: it has
been shown that under oxidative stress, procaspase-3 is
activated leading to impairment of aminophospholipid trans-
locase, phosphatidilserine externalisation and increased
erythrophagocytosis [41].
On the contrary, nucleated erythrocytes of lower verte-
brates are less specialised than their non-nucleated counter-
parts, by retaining nucleus and organelles after the
reticulocyte stage. These organelles are not cellular relicts,
they are active and can influence the physiology of the cell.
Little is known about programmed cell death of nucleated
erythrocytes. Chicken erythrocyte are reported to be able to
react to proapoptotic stimuli such as serum deprivation,
staurosporine and cycloheximide by showing characteristics
of apoptosis [42], but caspase activation in this cell type is
uncertain and apparently decreases with cell age. In trout
erythrocytes Moyes et al. [43] recently reported that pro-
oxidants and mitochondrial inhibitors failed to induce pro-
grammed cell death, suggesting that mitochondrial pathways
are not able to trigger apoptosis in this cell type.
Our study demonstrates, for the first time, that nucleated
trout erythrocyte are able to commit apoptotic cell death via
a mitochondria-mediated route. Tributyltinchloride is
known to be mitochondria-active and to be able to induce
cytochrome c release in isolated mitochondria. TBTC
expresses toxicity not only through the induction of formal
membrane permeability transition, but also in a relatively
non-specific manner that involves mild loss of Dcm and
induction of swelling [21]. Our data indicate that the low
mitochondrial content of erythrocytes may release sufficient
amounts of proapoptotic proteins, and consequently that
mitochondrial pathways are able to trigger apoptosis in this
cell type.
Acknowledgements
This work was supported by CNR (National Research
Council) fund to G.F.
References
[1] R.J. Maguire, Review: environmental aspects of tributyltin, Appl.
Organomet. Chem. 1 (1987) 475–490.
[2] R.J. Maguire, Aquatic environmental aspects of non-pesticidal orga-
notin compounds, Water Pollut. Res. J. Can. 26 (1991) 243–248.
[3] G. Falcioni, G. Zolese, Effect of organotin compounds on trout eryth-
rocyte, Recent Res. Dev. Comp. Biochem. Physiol. 1 (2000) 67–76.
[4] K. Fent, Ecotoxicology of organotin compounds, Crit. Rev. Toxicol.
26 (1996) 1–117.
[5] G. Falcioni, R. Gabbianelli, A.M. Santroni, G. Zolese, D.E. Griffiths,
E. Bertoli, Plasma membrane perturbation induced by organotin on
erythrocytes from Salmo irideus trout, Appl. Organomet. Chem. 10
(1996) 451–457.
[6] A.M. Santroni, D. Fedeli, G. Zolese, R. Gabbianelli, G. Falcioni,
Plasma membrane perturbation induced by tributyltin chloride on
density separated trout erythrocytes, Appl. Organomet. Chem. 13
(1999) 777–781.
[7] M. Brunori, Molecular adaptation to physiological requirements: the
hemoglobin system of trout, Curr. Top. Cell. Regul. 9 (1975) 1–39.
[8] L. Tiano, D. Fedeli, M. Moretti, G. Falcioni, DNA damage induced
by organotins on trout-nucleated erythrocytes, Appl. Organomet.
Chem. 15 (2001) 575–580.
[9] M. Villarini, M. Moretti, E. Damiani, L. Greci, A.M. Santroni, D.
Fedeli, G. Falcioni, Detection of DNA damage in stressed trout
nucleated erythrocytes using the comet assay: protection by nitro-
xide radicals, Free Radic. Biol. Med. 24 (1998) 1310–1315.
[10] S.A. Susin, N. Zamzami, G. Kroemer, Mitochondria as regulators of
apoptosis: doubt no more, Biochim. Biophys. Acta 1366 (1998)
151–165.
[11] G. Lenaz, Role of mitochondria in oxidative stress and ageing, Bio-
chim. Biophys. Acta 1366 (1998) 53–67.
[12] B. Mignotte, J.L. Vayssiere, Mitochondria and apoptosis, Eur. J. Bio-
chem. 252 (1998) 1–15.
[13] V.P. Skulachev, Why are mitochondria involved in apoptosis? Perme-
ability transition pores and apoptosis as selective mechanisms to elim-
inate superoxide-producing mitochondria and cell, FEBS Lett. 397
(1996) 7–10.
[14] P.X. Petit, H. Lecoeur, E. Zorn, C. Dauguet, B. Mignotte, M.L. Gou-
geon, Alterations in mitochondrial structure and function are early
events of dexamethasone-induced thymocyte apoptosis, J. Cell Biol.
130 (1995) 157–167.
[15] N. Zamzami, P. Marchetti, M. Castedo, C. Zanin, J.L. Vayssiere, P.X.
Petit, G. Kroemer, Reduction in mitochondrial potential constitutes
an early irreversible step of programmed lymphocyte death in vivo,
J. Exp. Med. 181 (1995) 1661–1672.
[16] M. Poot, Y.Z. Zhang, J.A. Kramer, K.S. Wells, L.J. Jones, D.K.
Hanzel, A.G. Lugade, V.L. Singer, R.P. Haugland, Analysis of
mitochondrial morphology and function with novel fixable fluo-
rescent stains, J. Histochem. Cytochem. 44 (1996) 1363–1372.
[17] S. Salvioli, A. Ardizzoni, C. Franceschi, A. Cossarizza, JC-1, but not
DiOC6(3) or rhodamine 123, is a reliable fluorescent probe to assess
delta psi changes in intact cells: implications for studies on mitochon-
drial functionality during apoptosis, FEBS Lett. 411 (1997) 77–82.
[18] S.T. Smiley, M. Reers, C. Mottola-Hartshorn, M. Lin, A. Chen,
T.W. Smith, G.D.J. Steele, L.B. Chen, Intracellular heterogeneity
in mitochondrial membrane potentials revealed by a J-aggregate-
forming lipophilic cation JC-1, Proc. Natl. Acad. Sci. U. S. A. 88
(1991) 3671–3675.

L. Tiano et al. / Biochimica et Biophysica Acta 1640 (2003) 105–112112
[19] M. Reers, T.W. Smith, L.B. Chen, J-aggregate formation of a carbo-
cyanine as a quantitative fluorescent indicator of membrane potential,
Biochemistry 30 (1991) 4480–4486.
[20] A. Cossarizza, M. Baccarani-Contri, G. Kalashnikova, C. Franceschi,
A new method for the cytofluorimetric analysis of mitochondrial
membrane potential using the J-aggregate forming lipophilic cation
5,5V,6,6V-tetrachloro-1,1V,3,3V-tetraethylbenzimidazolcarbocyanine io-
dide (JC-1), Biochem. Biophys. Res. Commun. 197 (1993) 40–45.
[21] V. Gogvadze, H. Stridh, S. Orrenius, I. Cotgreave, Tributyltin causes
cytochrome C release from isolated mitochondria by two discrete
mechanisms, Biochem. Biophys. Res. Commun. 292 (2002) 904–908.
[22] H. Stridh, D. Gigliotti, S. Orrenius, I. Cotgreave, The role of calcium
in pre- and postmitochondrial events in tributyltin-induced T-cell
apoptosis, Biochem. Biophys. Res. Commun. 266 (1999) 460–465.
[23] D.W. Nicholson, A. Ali, N.A. Thornberry, J.P. Vaillancourt, C.K.
Ding, M. Gallant, Y. Gareau, P.R. Griffin, M. Labelle, Y.A. Lazebnik,
Identification and inhibition of the ICE/CED-3 protease necessary for
mammalian apoptosis, Nature 376 (1995) 37–43.
[24] W.N. Aldridge, B.W. Street, Oxidative phosphorylation. The relation
between the specific binding of trimethylytin and triethyltin to mito-
chondria and their effects on various mitochondrial functions, Bio-
chem. J. 124 (1971) 221–234.
[25] W.N. Aldridge, B.W. Street, Oxidative phosphorylation. The specific
binding of trimethyltin and triethyltin to rat liver mitochondria, Bio-
chem. J. 118 (1970) 171–179.
[26] W.N. Aldridge, B.W. Street, Oxidative phosphorylation. Biochemical
effects and properties of trialkyltins, Biochem. J. 91 (1964) 287–297.
[27] A. Gennari, B. Viviani, C.L. Galli, M. Marinovich, R. Pieters, E.
Corsini, Organotins induce apoptosis by disturbance of [Ca(2+)](i)
and mitochondrial activity, causing oxidative stress and activation of
caspases in rat thymocytes, Toxicol. Appl. Pharmacol. 169 (2000)
185–190.
[28] H.U. Simon, A. Haj-Yehia, F. Levi-Schaffer, Role of reactive oxygen
species (ROS) in apoptosis induction, Apoptosis 5 (2000) 415–418.
[29] R. Pieters, M. Bol, A.H. Penninks, Immunotoxic organotins as
possible model compounds in studying apoptosis and thymocyte
differentiation, Toxicology 91 (1994) 189–202.
[30] O. Yamanoshita, M. Kurasaki, T. Saito, K. Takahasi, H. Sasaki, T.
Hosokawa, M. Okabe, J. Mochida, T. Iwakuma, Diverse effect of
tributyltin on apoptosis in PC12 cells, Biochem. Biophys. Res.
Commun. 272 (2000) 557–562.
[31] F. Cima, L. Ballarin, TBT-induced apoptosis in tunicate haemocytes,
Appl. Organomet. Chem. 13 (1999) 697–703.
[32] H. Stridh, S. Orrenius, M.B. Hampton, Caspase involvement in the
induction of apoptosis by the environmental toxicants tributyltin and
triphenyltin, Toxicol. Appl. Pharmacol. 156 (1999) 141–146.
[33] J.D. Robertson, S. Orrenius, Molecular mechanisms of apoptosis in-
duced by cytotoxic chemicals, Crit. Rev. Toxicol. 30 (2000) 609–627.
[34] B. Viviani, A.D. Rossi, S.C. Chow, P. Nicotera, Organotin com-
pounds induce calcium overload and apoptosis in PC12 cells, Neuro-
toxicology 16 (1995) 19–25.
[35] S. Mizuhashi, Y. Ikegaya, N. Matsuki, Cytotoxicity of tributyltin in rat
hippocampal slice cultures, Neurosci. Res. 38 (2000) 35–42.
[36] M. Marinovich, B. Viviani, E. Corsini, F. Ghilardi, C.L. Galli, NF-
kappaB activation by triphenyltin triggers apoptosis in HL-60 cells,
Exp. Cell Res. 226 (1996) 98–104.
[37] R. Gabbianelli, M. Villarini, G. Falcioni, G. Lupidi, Effect of different
organotin compounds on DNA of gilthead sea bream (Sparus aurata)
erythrocytes assessed by the comet assay, Appl. Organomet. Chem.
16 (2002) 163–168.
[38] A.M. Santroni, D. Fedeli, G. Zolese, R. Gabbianelli, G. Falcioni,
Plasma membrane perturbation induced by trubutyltin chloride on
density separated trout erythrocytes, Appl. Organomet. Chem. 13
(1999) 777–781.
[39] D. Bratosin, J. Estaquier, F. Petit, D. Arnoult, B. Quatannens, J.P.
Tissier, C. Slomianny, C. Sartiaux, C. Alonso, J.J. Huart J. Mon-
treuil, J.C. Ameisen, Programmed cell death in mature erythro-
cytes: a model for investigating death effector pathways operating in
the absence of mitochondria, Cell Death Differ. 8 (2001) 1143–1156.
[40] C.P. Berg, I.H. Engels, A. Rothbart, K. Lauber, A. Renz, S.F.
Schlosser, K. Schulze-Osthoff, S. Wesselborg, Human mature red
blood cells express caspase-3 and caspase-8, but are devoid of mi-
tochondrial regulators of apoptosis, Cell Death Differ. 8 (2001)
1197–1206.
[41] D. Mandal, P.K. Moitra, S. Saha, J. Basu, Caspase 3 regulates phos-
phatidylserine externalization and phagocytosis of oxidatively stressed
erythrocytes, FEBS Lett. 513 (2002) 184–188.
[42] M. Weil, M.D. Jacobson, M.C. Raff, Are caspases involved in the
death of cells with a transcriptionally inactive nucleus? Sperm and
chicken erythrocytes, J. Cell Sci. 111 (Pt. 18) (1998) 2707–2715.
[43] C.D. Moyes, M.L. Sharma, C. Lyons, S.C. Leary, M. Leon, A. Petrie,
S.G. Lund, B.L. Tufts, Origins and consequences of mitochondrial
decline in nucleated erythrocytes, Biochim. Biophys. Acta 1591
(2002) 11–20.






























