



b i om a s s a n d b i o e n e r g y 3 5 ( 2 0 1 1 ) 3 0 7 9e3 0 8 5
Avai lab le a t www.sc iencedi rec t .com
ht tp : / /www.e lsev ier . com/ loca te /b iombioe
Characterization and identification of lipid-producingmicroalgae species isolated from a freshwater lake
Reda A.I. Abou-Shanab a,b, Ibrahim A. Matter a, Su-Nam Kim c, You-Kwan Oh d,Jaeyoung Choi c, Byong-Hun Jeon a,*aDepartment of Environmental Engineering, Yonsei University, Wonju, Gangwon-do 220-710, South KoreabDepartment of Environmental Biotechnology, Mubarak City for Scientific Research and Technology Applications, New Borg El Arab City,
Alexandria 21934, EgyptcKIST Gangneung Institute, Gangneung 210-340, South KoreadBioenergy Center, Korea Institute of Energy Research, Daejeon 305-343, South Korea
a r t i c l e i n f o
Article history:
Received 31 March 2010
Received in revised form
10 April 2011
Accepted 15 April 2011
Available online 5 May 2011
Keywords:
Biodiesel
Freshwater
Isolation
LSU rDNA (D1-D2)
Microalgae
* Corresponding author. Tel.: þ82 33 760 244E-mail address: [email protected] (B.-H
0961-9534/$ e see front matter ª 2011 Elsevdoi:10.1016/j.biombioe.2011.04.021
a b s t r a c t
Microalgal lipids are the oils of the future for sustainable biodiesel production. One of the
most important decisions in obtaining oil from microalgae is the choice of species. A total
of 45 algal cultures were isolated from a freshwater lake at Wonju, South Korea. Five
microalgal isolates were selected based on their morphology and ease of cultivation
under our test conditions. These cultures were identified as strains of Scenedesmus obli-
quus YSL02, Chlamydomonas pitschmannii YSL03, Chlorella vulgaris YSL04, S. obliquus YSL05,
and Chlamydomonas mexicana YSL07 based on microscopic examination and LSU rDNA
(D1-D2) sequence analysis. S. obliquus YSL02 reached a higher biomass concentration
(1.84 � 0.30 g L�1) with a lower lipid content (29% w/w), than did Chla. pitschmannii YSL03
(maximum biomass concentration of 1.04 � 0.09 with a 51% lipid content). Our results
suggest that Chla. pitschmannii YSL03 is appropriate for producing biodiesel based on its
high lipid content and oleic acid proportion.
ª 2011 Elsevier Ltd. All rights reserved.
1. Introduction Biomass is one of the better sources of energy to mitigate
The basic resources currently exploited to obtain energy are
petroleum, natural gas, coal, hydropower, and nuclear power.
Continued use of petroleum-based fuels is now widely
recognized as unsustainable because of limited supplies and
the contribution of these fuels to atmospheric pollution. Fossil
fuel combustion is also a major source of greenhouse gases
responsible for global warming. Renewable, carbon-neutral,
economically viable alternatives to fossil fuels are urgently
needed to avert the impending oil crisis and the dramatic
consequences of climate change [1].
6; fax: þ82 33 760 2571.. Jeon).
ier Ltd. All rights reserved
greenhouse gas emissions and to function as a substitute for
fossil fuels [2]. Large-scale introductionof biomass energy could
contribute to sustainable development on environmental,
social, and economic fronts. Biodiesel (monoalkyl esters) is one
such alternative fuel, obtained by the transesterification of
triglyceride oil with monohydric alcohols. Commercial bio-
diesel has been obtained successfully from rapeseed, soybean,
sunflower, corn, palm, and waste cooking oil, as well as from
animal fat [3].However, large-scaleproductionofbiodiesel from
those resources cannot realistically satisfy the existing demand
for transport fuels [1]. Biodiesel has received considerable
.

b i om a s s an d b i o e n e r g y 3 5 ( 2 0 1 1 ) 3 0 7 9e3 0 8 53080
attention in recent years, as it is a biodegradable, renewable,
andnon-toxic fuel. It contributesnonet carbondioxideor sulfur
to the atmosphere and emits less gaseous pollutants than does
normal diesel [3]. While biodiesel is a desirable product, the
significant economic and environmental impacts of using
agricultural crops, especially food crops, as a feedstock for bio-
fuels has raised crucial sustainability issues. For example, the
transformation of primary food resources into bio-fuels has led
to a clash of interests, as using food crops to derived biodiesel
has resulted in a reduced supply to poor countries and an
increase in food costs [4]. All of these factors have stimulated
the search for other sources of biodiesel production that are
both sustainable and economical.
Microalgae are microscopic heterotrophiceautotrophic
photosynthesizingorganismsthatareabletousesolarenergyto
combine water with carbon dioxide to create biomass. Micro-
algae are present in all existing earth ecosystems, both aquatic
and terrestrial, and can flourish under a wide range of envi-
ronmental conditions, including freshwater, brackish water,
seawater, and even wastewater [5]. Microalgae have been sug-
gested as good candidates for fuel production because of their
higher photosynthetic efficiency, higher biomass production
and faster growth compared to those of other energy crops [6].
Microalgae systems also use far less water than do traditional
oilseed crops. For these reasons, microalgae are capable of
producingmore oil per unit area of land compared to terrestrial
oilseed crops [7]. According to some estimates, the yield (per
acre) of oil from algae is over 200 times the yield from the best-
performing plant/vegetable oils [8].
Hundreds of microalgal strains capable of producing large
quantities of lipids have been screened, and their lipid
production metabolisms characterized and reported. Most of
these organisms are marine microalgae [8]. Some species of
algae produce large quantities of oil as a storage product,
regularly achieving 50%e60% of their dry weights as lipid [7].
Biosynthesis of fatty acids can vary significantly according to
the external environmental conditions [9] with temperature,
composition in the culture medium and bubbling gas concen-
tration being important in microalgal growth.
Microalgae with high contents of fatty acids, neutral lipids,
and polar lipids as well as a high growth rate in the natural
environment have yet to be exploited for biodiesel production,
and the isolation and characterization of microalgae with the
potential for more efficient oil production remain the focus of
continuing research [10]. In this study, the growth rate, algal
biomass, and lipid content of some environmental microalgal
isolates were determined. Furthermore, these naturally iso-
lated microalgal species were subjected to fatty acid profile
analysis.Moreover, the large subunit (LSU) ribosomal DNA
(D1-D2)-encoding gene of the isolates was sequenced to
confirm the identities of the microalgal species.
2. Materials and methods
2.1. Isolation, purification and identification ofmicroalgae
Water samples used to isolate microalgae were collected
aseptically from sites that appeared to contain algal growth in
a freshwater lake at Yonsei University, Wonju, South Korea.
Bold basal medium (BBM) was used in this study [11]. The
media was autoclaved at 1.2 atm for 15 min before use. Ten
milliliters of water sample was inoculated into 200 mL media
in a 500 mL conical flask, and then incubated on a rotary
shaker at 27 �C and 150 rpm under continuous illumination
with white fluorescent light for three weeks.
Every two days, the flasks were examined for algal growth
using an optical microscope, and serial dilutions were
prepared in BBM from flasks showing growth. Subcultures
were made by inoculating 50 mL onto petri plates containing
BBM solidified with 1.5% (w/v) of bacteriological agar.
Further, 50 mL aliquots of the same dilution were placed into
wells of a 96-well microtiter plate containing 200 mL BBM.
These procedures were repeated for each of the original
flasks.
Both the Petri andmicrotiter plates were incubated at 27 �Cunder continuous illumination using white fluorescent light
for two weeks. The purities of the culture were ensured by
repeated plating and regular observation under a microscope.
Microscopic identification was performed [12] and was
confirmed using molecular markers.
2.2. DNA extraction, PCR amplification, sequencing, andphylogenetic analysis
An aliquot of cultured cells (1mL) was harvested in themid- to
late exponential growth phase (10e14 days) by centrifugation
(13,000� g for 3 min at 4 �C) in a sterile microcentrifuge tube.
Genomic DNA was extracted using a Plant Genomic DNA
extraction kit (SolGent, Daejeon, S. Korea) according to the
manufacturer’s instructions and protocols. The DNA
concentration of the extracted DNA was measured at 260 nm
using a spectrophotometer (HACH�, DR/4000v, USA). To
amplify the D1-D2 (LSU) coding region of the rDNA, amplifi-
cation reactions were performed on a T-Gradient thermo-
cycler (Biometra GmbH, Gottingen, Germany) using the
universal eukaryotic primers 50-AGCGGAGGAAAAGAAACTA-’3 as forward and 50-TACTAGAAGGTTCGATTAGTC-’3 as
reverse, according to the PCR protocol described by Sonnen-
berg et al. [13]. Aliquots (10 mL) of the reaction mixtures were
analyzed by 1% horizontal agarose gel electrophoresis to
confirm the presence of product. The PCR products were
purified using the Gel PCR Clean-Up System (Applied Bio-
systems, Foster, CA). Sequencing reactions were performed
using a Dye Deoxy Terminator Cycle Sequencing Ready
Reaction Kit (Applied Biosystems, Foster City, CA), and
sequencing fragments were analyzed on a ABI Prism 377 DNA
Sequencer.
Ribosomal RNA gene sequences from the isolates were
searched against GenBank using BLAST [14]. Aligned
sequences were checked manually and were edited with
Genedoc [15]. Sequences containing fewer than 200 nucleo-
tides or in excess of 1000 nucleotides were removed, and
sequences not belonging to greenmicroalgal specieswere also
discarded from our study. A phylogenetic tree was con-
structed using the neighbor-joining (NJ) algorithm using
Kimura’s two-parameter model of sequence evolution, as
implemented in the MEGA4 program package [16].

b i om a s s a n d b i o e n e r g y 3 5 ( 2 0 1 1 ) 3 0 7 9e3 0 8 5 3081
2.3. Microalgae cultivation and biomass
One hundredmilliliters BBM in a 250mL Erlenmeyer flask was
inoculated with the cells (OD680 0.05) and incubated at 27 �Cwith shaking at 150 rpm under continuous illumination for
three weeks. Algal growth was monitored by measuring daily
changes in optical density at 680 nm with a spectrophotom-
eter. If the optical density of the undiluted sample was greater
than 1.0, the sample was diluted to give an absorbance in the
range of 0.1e1.0.
Microalgae dry weight per liter (g L�1) was measured
according to a method previously reported [17]. Microalgal
cells were harvested by centrifugation and washed twice
with deionized water. Microalgal pellets were dried overnight
at 105 �C for dry weight measurement [18]. Experiments
were carried out in triplicate, and data are expressed as
mean � SD.
2.4. Extraction of total lipids
Total lipids were extracted from fresh microalgal biomass
using a slightly modified method of Bligh and Dyer [19]. The
lipids were extracted with chloroform-methanol (2:1, v/v)
and then separated into chloroform and aqueous methanol
layers by the addition of methanol and water to give a final
solvent ratio of chloroform: methanol: water of 1:1:0.9. The
chloroform layer was washed with 20 mL of a 5% NaCl
solution, and evaporated to dryness. Thereafter, the weight
of the crude lipid obtained from each sample was measured
gravimetrically. Experiments were carried out in triplicate,
and data are expressed as mean � SD.
2.5. Fatty acid composition analysis
The fatty acids were analyzed using the modified method of
Lepage and Roy [20]. The crude lipid (w10 mg) was dissolved
using 2 mL of a freshly prepared chloroform-methanol
mixture (2: 1, v/v) and transferred into capped test tube. One
mL of chloroform containing nonadecanoic acid (500 mg L�1) as
internal standard, 1 mL methanol, and 300 mL of sulfuric acid
as transmethylation reagents were added to the tube, mixed
for 5 min and then incubated at 100 �C for 10 min. The fatty
acid-containing phase was separated by adding 1 ml distilled
water and was then recovered. The organic phase was filtered
using a hypodermic 0.22 mm PVDF syringe filter (Millex-GV,
Millipore, USA). Methyl esters of fatty acids were analyzed
Fig. 1 e Light microscope (40x) picture
using a gas chromatograph (GC-7890, Agilent, USA) equipped
with a flame ionization detector anda HP-INNO wax capillary
column (Agilent Technologies, USA). The temperatures of
injector and detector were set at 250 �C and 275 �C, respec-tively. Oven temperature conditions were maintained at 50 �Cfor 1min, 200 �C for 12min, and 250 �C for 2min. Mix RM3,Mix
RM5, GLC50, and GLC70 (Supelco Co., USA), and a-linolenic
acid (Sigma Chemical Co. USA) were used as standard mate-
rials. All reagents were of analytical grade. The components
were identified by comparing their retention times and frag-
mentation patterns with those of the standards [21].
3. Results and discussion
3.1. Isolation and identification of microalgae
A total of 45 algal cultures were isolated from a freshwater
lake at Yonsei University, Wonju, S. Korea. Out of 45
cultures, five green microalgal isolates (YSL02, YSL03,
YSL05, YSL04 and YSL07) were selected based on their
morphologies (i.e., cell shape and size) and because they
could be successfully cultivated in pure form under our test
conditions. Light microscopic images of the new species
isolated in this study are shown in Fig. 1. Microscopic
observation of algal isolates revealed their colonial exis-
tences and purities. Microscopic analysis of the samples
allowed preliminary identification of isolates YSL02, YSL03,
YSL04, YSL05, and YSL07 as genus Scenedesmus, Chlamydo-
monas, Chlorella, Scenedesmus, and Chlamydomonas, respec-
tively. Komarek and Marvan [22] proposed the existence of
at least 13 species of Botryococcus on the basis of morpho-
logical differences by omitting the chemical analyses.
Metzger and Largeau [23] reported that for algae, within
each chemical race and for the same strain, morphology
could vary in relation to age and culture conditions. The
morphological heterogeneity of algae makes microscopic
identification difficult. Therefore, we isolated total DNA and
PCR-amplified rRNA (LSU) to confirm our morphology-based
species identifications.
3.1.1. LSU-rRNA (D1-D2) coding region amplification andsequencingPCR amplification of the genomic DNA of the algal isolates
with the universal forward and reverse primers revealed
efficient amplification. A single band of amplified LSU rDNA
s of the tested microalgal isolates.

b i om a s s an d b i o e n e r g y 3 5 ( 2 0 1 1 ) 3 0 7 9e3 0 8 53082
(D1-D2) product with a size of w 850 bp was recorded for all
isolates. The LSU rRNA gene has a higher evolutionary rate
than does the SSU rRNA gene [13] and should therefore allow
for better discrimination between closely-related species
using short diagnostic sequences. Based on the LSU rDNA (D1-
D2) sequences, we concluded that microalgal isolates YSL02,
YSL03, YSL04, YSL05 and YSL07 were closely related to Scene-
desmus obliquus, Chlamydomonas pitschmannii, Chlorella vulgaris,
S. obliquus and Chlamydomonas mexicana, based on 97%, 99%,
97%, 97% and 99% sequence similarities, respectively. The
DNA sequences were published in the NCBI databases
(accession numbers are provided in Table 1). The lengths of
the LSU rDNA (D1-D2) regions of the five species ofmicroalgae,
their specific accession numbers and the nearest identifiable
match present in the GenBank nucleotide database are shown
in Table 1. Identification of the fivemicroalgal strains was also
supported by the results from the phylogenetic analysis of the
LSU rDNA D1-D2 sequence. In the phylogram (Fig. 2), YSL02
and YSL05 which were identified as S. obliquus clearly group
with the microalgal strain S. obliquus AF183482 (Table 1). The
LSU rDNA sequences of isolates YSL03, YSL04, and YSL07
confirmed their identification as Chla. pitschmannii, C. vulgaris,
and Chla. mexicana; they had sequence similarities of 99%,
97%, and 99% to Chla. pitschmannii AF183462, C. vulgaris
AB237642, and Chla. mexicana AF395501, respectively (Fig. 2
and Table 1).
Fig. 2 e Phylogenetic tree showing the relationshipsamong LSU rDNA D1-D2 sequences of isolates YSL02,
YSL03, YSL04, YSL05, and YSL07 and the most similar
sequences retrieved from the NCBI nucleotide database.
3.2. Growth rates of the microalgal strains
Under suitable conditions and sufficient nutrients, microalgae
can grow profusely. Their biomass usually doubles within
3.5e24 h during the exponential growth phase [1]. The net
growth rates differed among the examined species under
similar environmental conditions (Fig. 3). The average growth
rates of C. vulgaris YSL04, Chla. mexicana YSL07, S. obliquus
YSL05, S. obliquus YSL02, and Chla. pitschmannii YSL03, were
1.83 � 0.07, 1.65 � 0.04, 1.24 � 0.07, 1.07 � 0.17, and 0.61 � 0.11
days�1, respectively. The growth rate of C. vulgaris YSL04 after
20 days of incubation was 3.29� 0.20 compared with an initial
reading of 0.69 � 0.19 at OD 680 nm. This result indicates that
C. vulgaris YSL04 is suitable for high-density culture. Algal
growth is directly affected by the availability of nutrients,
light, the stability of pH, temperature and the initial inoculum
density [24]. An increase in the initial inoculum density leads
to better algal growth and increases the nutrient removal
efficiency [25].
Table 1 e The accession numbers, lengths in base pairs, similasequences for five strains of environmentally isolated microal
Microalgal strain Accession number Length
S. obliquus YSL02 GU732415 864
Chla. pitschmannii YSL03 GU732416 874
C. vulgarisYSL04 GU732417 881
S. obliquus YSL05 GU732418 864
Chla. mexicana YSL07 GU732420 868
S ¼ Scenedesmus, Chla ¼ Chlamydomonas, C ¼ Chlorella and (nt) ¼ nucleotid
3.3. Biomass, lipid content, and lipid productivity
The five microalgal species were tested for lipid production by
evaluating biomass productivity and lipid content in 250 mL
flask laboratory cultures under the same conditions after 21
days of incubation (Table 2). Biomass productivities (g dwt L�1)
of 1.84� 0.30; 1.71� 0.53; 1.65� 0.07, and 1.53� 0.30were found
for S. obliquus YSL02; S. obliquus YSL05; C. vulgaris YSL04 and
Chla.mexicanaYSL07, respectively. S. obliquusYSL02 showed the
highest biomassproductivity at 1.84� 0.30 gdwt L�1,whileChla.
pitschmannii YSL03 had the lowest biomass productivity at
1.04 � 0.09 g dwt L�1.
Under our experimental growth conditions, the total lipid
contents of the microalgae cultured in this study ranged from
rities between amplified sequences, and the closest relativegae.
(nt) Closest relative andGenBank accession number
% Similarity
S. obliquus AF183482 97
Chla. pitschmannii AF183462 99
C. vulgarisAB237642 97
S. obliquus AF183482 97
Chla. mexicana AF395501 99
es.
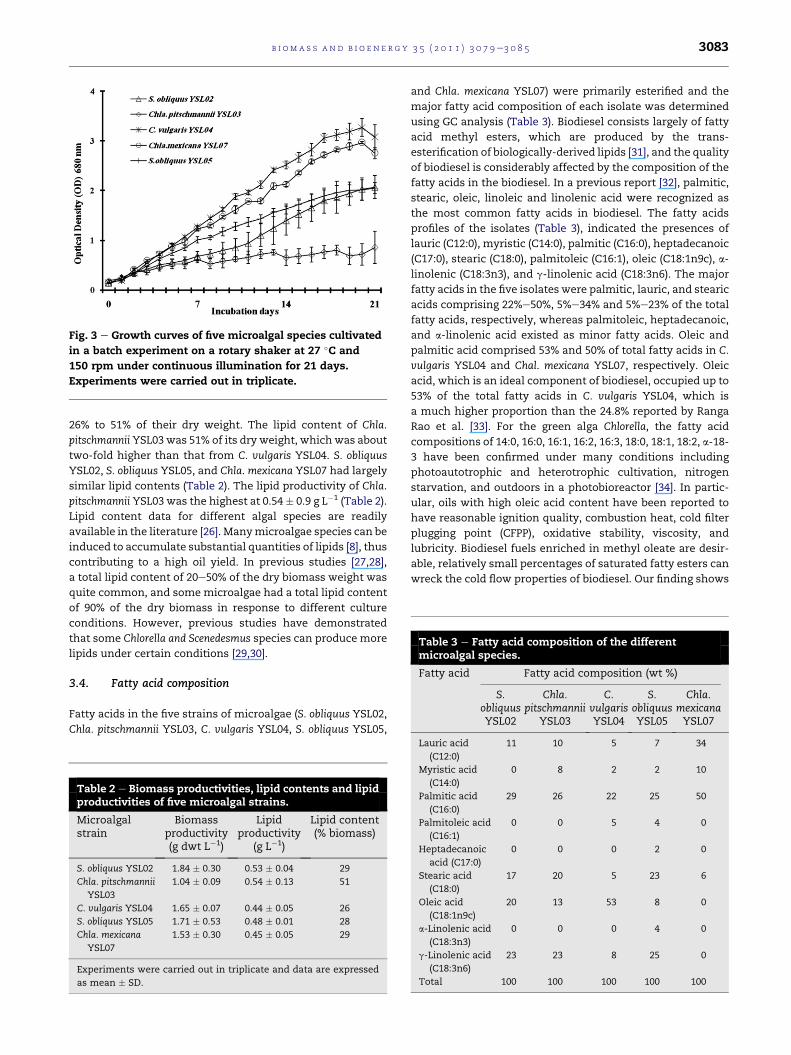
Fig. 3 e Growth curves of five microalgal species cultivated
in a batch experiment on a rotary shaker at 27 �C and
150 rpm under continuous illumination for 21 days.
Experiments were carried out in triplicate.
Table 3 e Fatty acid composition of the differentmicroalgal species.
Fatty acid Fatty acid composition (wt %)
S.obliquusYSL02
Chla.pitschmannii
YSL03
C.vulgarisYSL04
S.obliquusYSL05
Chla.mexicanaYSL07
b i om a s s a n d b i o e n e r g y 3 5 ( 2 0 1 1 ) 3 0 7 9e3 0 8 5 3083
26% to 51% of their dry weight. The lipid content of Chla.
pitschmannii YSL03 was 51% of its dry weight, which was about
two-fold higher than that from C. vulgaris YSL04. S. obliquus
YSL02, S. obliquus YSL05, and Chla. mexicana YSL07 had largely
similar lipid contents (Table 2). The lipid productivity of Chla.
pitschmannii YSL03 was the highest at 0.54 � 0.9 g L�1 (Table 2).
Lipid content data for different algal species are readily
available in the literature [26]. Manymicroalgae species can be
induced to accumulate substantial quantities of lipids [8], thus
contributing to a high oil yield. In previous studies [27,28],
a total lipid content of 20e50% of the dry biomass weight was
quite common, and somemicroalgae had a total lipid content
of 90% of the dry biomass in response to different culture
conditions. However, previous studies have demonstrated
that some Chlorella and Scenedesmus species can produce more
lipids under certain conditions [29,30].
3.4. Fatty acid composition
Fatty acids in the five strains of microalgae (S. obliquus YSL02,
Chla. pitschmannii YSL03, C. vulgaris YSL04, S. obliquus YSL05,
Table 2 e Biomass productivities, lipid contents and lipidproductivities of five microalgal strains.
Microalgalstrain
Biomassproductivity(g dwt L�1)
Lipidproductivity
(g L�1)
Lipid content(% biomass)
S. obliquus YSL02 1.84 � 0.30 0.53 � 0.04 29
Chla. pitschmannii
YSL03
1.04 � 0.09 0.54 � 0.13 51
C. vulgaris YSL04 1.65 � 0.07 0.44 � 0.05 26
S. obliquus YSL05 1.71 � 0.53 0.48 � 0.01 28
Chla. mexicana
YSL07
1.53 � 0.30 0.45 � 0.05 29
Experiments were carried out in triplicate and data are expressed
as mean � SD.
and Chla. mexicana YSL07) were primarily esterified and the
major fatty acid composition of each isolate was determined
using GC analysis (Table 3). Biodiesel consists largely of fatty
acid methyl esters, which are produced by the trans-
esterification of biologically-derived lipids [31], and the quality
of biodiesel is considerably affected by the composition of the
fatty acids in the biodiesel. In a previous report [32], palmitic,
stearic, oleic, linoleic and linolenic acid were recognized as
the most common fatty acids in biodiesel. The fatty acids
profiles of the isolates (Table 3), indicated the presences of
lauric (C12:0), myristic (C14:0), palmitic (C16:0), heptadecanoic
(C17:0), stearic (C18:0), palmitoleic (C16:1), oleic (C18:1n9c), a-
linolenic (C18:3n3), and g-linolenic acid (C18:3n6). The major
fatty acids in the five isolates were palmitic, lauric, and stearic
acids comprising 22%e50%, 5%e34% and 5%e23% of the total
fatty acids, respectively, whereas palmitoleic, heptadecanoic,
and a-linolenic acid existed as minor fatty acids. Oleic and
palmitic acid comprised 53% and 50% of total fatty acids in C.
vulgaris YSL04 and Chal. mexicana YSL07, respectively. Oleic
acid, which is an ideal component of biodiesel, occupied up to
53% of the total fatty acids in C. vulgaris YSL04, which is
a much higher proportion than the 24.8% reported by Ranga
Rao et al. [33]. For the green alga Chlorella, the fatty acid
compositions of 14:0, 16:0, 16:1, 16:2, 16:3, 18:0, 18:1, 18:2, a-18-
3 have been confirmed under many conditions including
photoautotrophic and heterotrophic cultivation, nitrogen
starvation, and outdoors in a photobioreactor [34]. In partic-
ular, oils with high oleic acid content have been reported to
have reasonable ignition quality, combustion heat, cold filter
plugging point (CFPP), oxidative stability, viscosity, and
lubricity. Biodiesel fuels enriched in methyl oleate are desir-
able, relatively small percentages of saturated fatty esters can
wreck the cold flow properties of biodiesel. Our finding shows
Lauric acid
(C12:0)
11 10 5 7 34
Myristic acid
(C14:0)
0 8 2 2 10
Palmitic acid
(C16:0)
29 26 22 25 50
Palmitoleic acid
(C16:1)
0 0 5 4 0
Heptadecanoic
acid (C17:0)
0 0 0 2 0
Stearic acid
(C18:0)
17 20 5 23 6
Oleic acid
(C18:1n9c)
20 13 53 8 0
a-Linolenic acid
(C18:3n3)
0 0 0 4 0
g-Linolenic acid
(C18:3n6)
23 23 8 25 0
Total 100 100 100 100 100

b i om a s s an d b i o e n e r g y 3 5 ( 2 0 1 1 ) 3 0 7 9e3 0 8 53084
that most of these strains contain w 20% of saturated fatty
acids (C16 and C18). Certainly this amount would negatively
impact cold flow properties such as CFPP. Higher oleic acid
content increases the oxidative stability of fuel, enabling
longer storage [32], and decreases the CFPP of the fuel,
allowing it to be used in cold regions [35]. Among the tested
microalgal species, C. vulgaris RAI04 showed the highest oleic
acid content, making it the most suitable isolate for the
production of good quality biodiesel.
4. Conclusions
Biomass productivity, lipid cell content, and overall lipid
productivity are some of the key parameters that determine
the economic feasibility of algal oil for biodiesel production.
To select microalgae with a high biomass and lipid produc-
tivity, five microalgal cultures were selected based on their
ease of cultivation and were identified as S. obliquus YSL02,
Chla. pitschmannii YSL03, C. vulgaris YSL04, S. obliquus YSL05,
and Chla. mexicana YSL07. Under similar environmental
conditions, the average growth rates (OD680 nm) of C. vulgaris
YSL04, Chla. mexicana YSL07, S. obliquus YSL05, S. obliquus
YSL02, and Chla. pitschmannii YSL03 were 1.83 � 0.071,
1.65 � 0.04, 1.24 � 0.07, 1.07 � 0.17 and 0.61 � 0.11 days�1,
respectively. These results indicated that the C. vulgaris YSL04
strainwas themost suitable of the five strains for high-density
culture. The total lipid contents of the algae were 51%, 29%,
29%, 28%, and 26% for Chla. pitschmannii YSL03, Chla. mexicana
YSL07, S. obliquus YSL02, S. obliquus YSL05, and C. vulgaris
YSL04, respectively. The composition of fatty acids in the
studied species was mainly C12:0, C16:0, C18:0, C18:1n9c,
C18:3n6, C16:1, and C14:0. The results of this study indicate
that the naturally isolated microalgal strain Chla. pitschmannii
YSL03 may be a valuable candidate for biodiesel production.
Acknowledgements
This work was supported by the Students’ Association of the
Graduate School of Yonsei University and was funded by the
Graduate School of Yonsei University, and Yonsei University
research fund of 2009, 21st Frontier research project
(Sustainable Water Resources Research Center 3-4-3), Global
Research Laboratory project (Korea Institute of Geosciences
and Mineral Resources NP2008-019) and the Brain Korea-21
(BK-21) program of the Ministry of Education, Korea.
r e f e r e n c e s
[1] Chisti Y. Biodiesel from microalgae. Biotechnol Adv 2007;25:294e306.
[2] Widjaja A, Chao-Chang Chien, Yi-Hsu Ju. Study of increasinglipid production from fresh water microalgae Chlorellavulgaris. J Taiwan Ins Chem Engin 2009;40:13e20.
[3] Lang X, Dalai AK, Bakhshi NN, Reaney MJ, Hertz PB.Preparation and characterization of biodiesels from variousBio-Oils. Biores Technol 2002;80:53e62.
[4] Puppan D. Environmental evaluation of biofuels. PeriodicaPolytechnic Ser Soc Man Sci 2002;10:95e116.
[5] Richmond A. Handbook of microalgal culture: Biotechnologyand applied phycology. Blackwell Science Ltd; 2004.
[6] Becker K. Measurement of algal growth. In: Microalgaebiotechnology and microbiology. Cambridge UniversityPress; 1994. p. 56e62.
[7] Chisti Y. Biodiesel from microalgae beats bioethanol. TrendsBiotechnol 2008;26:126e31.
[8] Sheehan J, Dunahay T, Benemann J, Roessler P. A look backat the U.S. Department of Energy’s aquatic species program:biodiesel from algae. Close-Out report. Golden, Colorado, U.S.A: National Renewable Energy Lab, Department of Energy;1998. Report number NREL/TP-580e24190.
[9] Los DA, Murata N. Membrane fluidity and its roles in theperception of environmental signals. Biochim Biophys Acta-Biomembranes 2004;1666:142e57.
[10] Qiang H, Sommerfeld M, Jarvis E, Ghirardi M, Posewitz M,Seibert M, et al. Microalgal triacylglycerols as feedstocks forbiofuel production: perspectives and advances. Plant J 2008;54:621e39.
[11] Kanz T, Bold HC. Publication No. In: Physiological studies,morphological and taxonomical investigation of nostoc andanabaenainculture.Austin,TX:UniversityofTexas;1969.p.6924.
[12] John DM, Whitton BA, Brook AJ. The freshwater algal flora ofthe British Isles an identification guide to freshwater andterrestrial algae. Cambridge: Cambridge University press;2003. 39e43.
[13] Sonnenberg R, Nolte AW, Tautz D. An evaluation of LSUrDNA D1-D2 sequences for their use in species identification.Front Zool 2007;4:1e12.
[14] Altschul SF, Thomas LM, Alejandro AS, Jinghui Z, Zheng Z,Webb M, et al. Gapped BLAST and PSI-BLAST: a newgeneration of protein database search programs. NucleicAcids Res 1997;25:3389e402.
[15] Nicholas KB, Nicholas HB. Alignment editor and shadingutility. 2.6.001 Ed, http://www.PSC.edu/biomed/genedoc; 1997.
[16] Tamura K, Dudley J. MECA4:molecular evolutionary geneticsanalysis (MEGA) softwareversion4.0.MolBiol Evol 2007;24:1596e9.
[17] American Public Health Association. Methods for biomassproduction. In: Standard methods for the examination ofwater and wastewater. Baltimore, MD, USA: American PublicHealth Association; 1998.
[18] Takagi M, Karseno S, Yoshida T. Effect of salt concentration onintracellularaccumulationof lipidsandtriacylglycerideinmarinemicroalgae Dunaliella cells. J Biosci Bioeng 2006;101:223e6.
[19] Bligh EG, Dyer WJ. A rapid method of total lipid extractionand purification. Can J Biochem Physiol 1959;37:911e7.
[20] Lepage G, Roy CC. Improved recovery of fatty acid throughdirect transesterification without prior extraction orpurification. J Lipid Res 1984;25:1391e6.
[21] Xu N, Zhang X, Fan X, Han L, Zeng C. Effects of nitrogensource and concentration on growth rate and fatty acidcomposition of Ellipsoidion sp. (Eustigmatophyta). J ApplPhycol 2001;13:463e9.
[22] Komarek J, Marvan P. Morphological differences in naturalpopulations of the genus Botryococcus (chlorophyceae).Archiv Fur Protistenkunde 1992;141:65e100.
[23] Metzger P, Largeau C. Botryococcus braunii: a rich source forhydrocarbons and related ether lipids. Appl MicrobiolBiotechnol 2005;66:486e96.
[24] Wang L, Yecong L, Chen P, Min M, Chen Y, Zhu J, et al.Anaerobic digested dairy manure as a nutrient supplementfor cultivation of oil-rich green microalgae Chlorella sp. BioresTechnol 2010;101:2623e8.
[25] Lau PS, Tam NFY, Wong YS. Effect of algal density onnutrient removal from primary settled wastewater. EnvironPollut 1995;89:59e66.

b i om a s s a n d b i o e n e r g y 3 5 ( 2 0 1 1 ) 3 0 7 9e3 0 8 5 3085
[26] Griffiths MJ, Harrison STL. Lipid productivity as a keycharacteristic for choosing algal species for biodieselproduction. J Appl Phycol 2009;21:493e507.
[27] SpolaoreP, Joannis-CassanC,DuranE, IsambertA.Commercialapplications of microalgae. J Biosci Bioeng 2006;101:87e96.
[28] Li Y, Horsman M, Wu N, Lan CQ, Dubois-Calero N. Biofuelsfrom microalgae. Biotechnol Prog 2008;24:815e20.
[29] Illman AM, Scragg AH, Shales SW. Increase in Chlorellastrains calorific values when grown in low nitrogen medium.Enz Microb Technol 2000;27:631e5.
[30] Rodolfi L, Zittelli GC, Bassi N, Padovani G, Biondi N, Bonin G,et al. Microalgae for oil: strain selection, induction of lipidsynthesis and outdoor mass cultivation in a low-costphotobioreactor. Biotechnol Bioeng 2009;102:100e12.
[31] Wackett LP. Biomass to fuels via microbial transformations.Curr Opin Chem Biol 2008;12:187e93.
[32] Knothe G. “Designer” biodiesel: optimizing fatty estercomposition to improve fuel properties. Energy Fuels 2008;22:1358e64.
[33] Ranga Rao A, Sarada TR, Ravishankar GA. Influence ofCO2 on growth and hydrocarbon production inBotryococcus braunii. J Microbiol Biotechnol 2007;17:414e9.
[34] Petkov G, Garcia G. Which are fatty acids of the green algaeChlorella. Biochem Syst Ecol 2007;35:281e5.
[35] Stournas S, Lois E, Serdari A. Effects of fatty acid derivativeson the ignition quality and cold flow of diesel fuel. J Am OilChem Soc 1995;72:433e7.






























