by David M. Knott; extracted from: South Carolina Department of Natural Resources and National Oceanic and Atmospheric Administration, Coastal Services Center. 2000. Characterization of the Ashepoo-Combahee- Edisto (ACE) Basin, South Carolina. CD-ROM. SC Marine Resources Center Special Scientific Report Number 17. NOAA/CSC/20010-CD. Charleston, SC: NOAA Coastal Services Center. Definition Organisms that live in aquatic environments face certain challenges that their terrestrial counterparts do not. One of the obvious differences is the motion of the fluid medium, which presents opportunities and drawbacks that are unique to animals and plants that live suspended in the water column. Among the benefits this lifestyle offers are enhanced dispersal of the population, which may be achieved at a relatively low energy cost, the resultant high gene flow among dispersed populations, and the ability to readily expand into new habitats. Aquatic organisms with limited swimming ability relative to the strength of ambient currents are said to be planktonic. The term plankton is derived from the Greek word planktos, which means wandering or drifting. Organisms such as these, whose distributions are closely tied to the movement of the water mass in which they reside, are at risk of being transported away from conditions that are necessary for their survival. Classification Biologists typically classify plankton into three general categories based on their phylogeny:

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

by David M. Knott; extracted from:
South Carolina Department of Natural Resources and National Oceanic and Atmospheric Administration, Coastal Services Center. 2000. Characterization of the Ashepoo-Combahee-Edisto (ACE) Basin, South Carolina. CD-ROM. SC Marine Resources Center Special Scientific Report Number 17. NOAA/CSC/20010-CD. Charleston, SC: NOAA Coastal Services Center.
Definition
Organisms that live in aquatic
environments face certain challenges that
their terrestrial counterparts do not. One
of the obvious differences is the motion
of the fluid medium, which presents
opportunities and drawbacks that are
unique to animals and plants that live
suspended in the water column. Among
the benefits this lifestyle offers are
enhanced dispersal of the population,
which may be achieved at a relatively low
energy cost, the resultant high gene flow
among dispersed populations, and the
ability to readily expand into new
habitats.
Aquatic organisms with limited
swimming ability relative to the strength
of ambient currents are said to be
planktonic. The term plankton is derived
from the Greek word planktos, which
means wandering or drifting. Organisms
such as these, whose distributions are
closely tied to the movement of the water
mass in which they reside, are at risk of
being transported away from conditions
that are necessary for their survival.
Classification
Biologists typically classify plankton into three general categories based on their phylogeny:
phytoplankton are microscopic algae and other photosynthetic organisms; zooplankton are
animals, mainly invertebrates; and ichthyoplankton comprise the larval fish component of the
plankton.
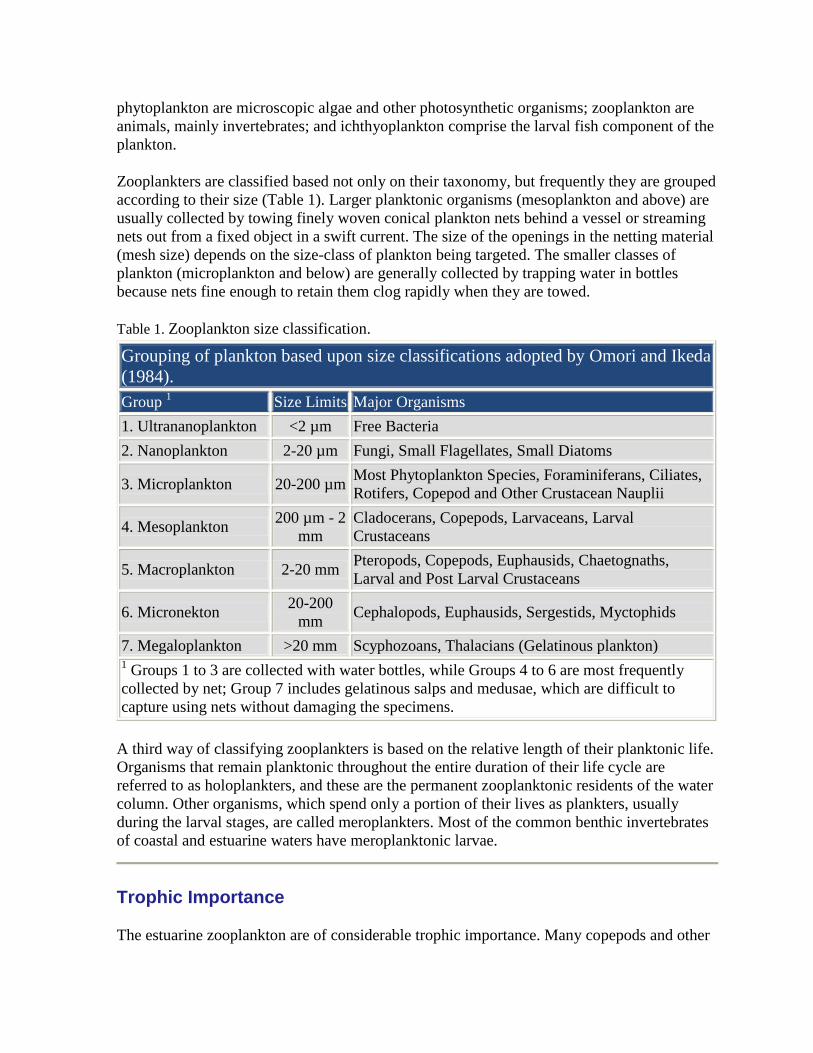
Zooplankters are classified based not only on their taxonomy, but frequently they are grouped
according to their size (Table 1). Larger planktonic organisms (mesoplankton and above) are
usually collected by towing finely woven conical plankton nets behind a vessel or streaming
nets out from a fixed object in a swift current. The size of the openings in the netting material
(mesh size) depends on the size-class of plankton being targeted. The smaller classes of
plankton (microplankton and below) are generally collected by trapping water in bottles
because nets fine enough to retain them clog rapidly when they are towed.
Table 1. Zooplankton size classification.
Grouping of plankton based upon size classifications adopted by Omori and Ikeda
(1984).
Group 1
Size Limits Major Organisms
1. Ultrananoplankton <2 µm Free Bacteria
2. Nanoplankton 2-20 µm Fungi, Small Flagellates, Small Diatoms
3. Microplankton 20-200 µm Most Phytoplankton Species, Foraminiferans, Ciliates,
Rotifers, Copepod and Other Crustacean Nauplii
4. Mesoplankton 200 µm - 2
mm
Cladocerans, Copepods, Larvaceans, Larval
Crustaceans
5. Macroplankton 2-20 mm Pteropods, Copepods, Euphausids, Chaetognaths,
Larval and Post Larval Crustaceans
6. Micronekton 20-200
mm Cephalopods, Euphausids, Sergestids, Myctophids
7. Megaloplankton >20 mm Scyphozoans, Thalacians (Gelatinous plankton) 1 Groups 1 to 3 are collected with water bottles, while Groups 4 to 6 are most frequently
collected by net; Group 7 includes gelatinous salps and medusae, which are difficult to
capture using nets without damaging the specimens.
A third way of classifying zooplankters is based on the relative length of their planktonic life.
Organisms that remain planktonic throughout the entire duration of their life cycle are
referred to as holoplankters, and these are the permanent zooplanktonic residents of the water
column. Other organisms, which spend only a portion of their lives as plankters, usually
during the larval stages, are called meroplankters. Most of the common benthic invertebrates
of coastal and estuarine waters have meroplanktonic larvae.
Trophic Importance
The estuarine zooplankton are of considerable trophic importance. Many copepods and other

zooplankters, especially estuarine species, are omnivores that derive the majority of their
nutrition by feeding on heterotrophic protists such as ciliates and dinoflagellates, although
under some circumstances they may rely more heavily on microphytoplankton (Kleppel et al.
1998). In localities where macrophytes are abundant, such as salt marshes or seagrass beds,
zooplankton standing stocks may obtain much of their nutrition by feeding on detritus
(Roman et al. 1983). In estuaries, heterotrophic protists are an important component of the
microzooplankton, since they provide a link between bacterial production and higher trophic
levels (Heip et al. 1995). Their importance in the diets of many marine and freshwater
zooplankton species was emphasized by Sanders and Wickham (1993), who noted that
protists serve as a necessary link in the transfer of bacterial biomass to larger organisms.
Zooplankton density and volume specific biomass are usually greater in estuaries than in
other aquatic habitats, reflecting the generally higher productivity of an estuarine
environment. The species of fish and shellfish responsible for over 85 percent (by weight) of
the commercial fisheries landings of the southeastern Atlantic states are estuarine or
estuarine-dependent at some life stage (Burrell 1975a). For many of these species that depend
on estuaries as spawning or nursery grounds (e.g., Atlantic croaker, Atlantic menhaden,
seatrout, drum, blue crab, and white shrimp), an abundant zooplanktonic population is
necessary. Recently, Allen et al. (1995) described how competition for zooplankton as food in
a high salinity South Carolina estuary may be minimized by vertical and lateral partitioning
and temporal shifts in dietary selectivity. Similar partitioning of zooplankton food sources,
based upon prey size, has been documented for freshwater fish species such as the threadfin
shad and blueback herring introduced to the Jocassee Reservoir in the 1970s (Davis and Foltz
1991).
Certain mesoplankters, particularly copepods and cladocerans, are essential as food for early
fish larvae and for larger predacious zooplankters, which in turn are fed upon by late larval
and postlarval fish and other organisms. In estuaries, macroplankters such as mysid shrimp
and gammarid amphipods may be the most important food chain link in habitats bounded by
extensive salt and brackish marshes, which themselves often are important fish nursery
grounds (Ragotzkie 1959; Van Engel and Joseph 1968; University of Georgia Marine
Institute 1971). In fresh water, most larval fish are zooplanktivores, frequently selecting
small-bodied organisms like rotifers and copepods. Cladocerans, which are generally larger,
are preferentially selected by older fish.
Zooplankton Behavior
Although the diverse assemblages of zooplankton in marine, estuarine and freshwater habitats
are all subjected to the vagaries of the water in which they reside, they do not all respond
similarly to the forces that cause the water to move. By using selective behavior in response
to various physical cues, even planktonic organisms can exert some influence on the ultimate
outcome of their transport (Epifanio 1988). Thus, by responding to salinity cues, some
planktonic species may be distributed only within restricted zones in coastal waters, such as
the low-salinity regions of estuaries, while others with may reside only in coastal waters and
the high-salinity reaches near the estuary mouth.
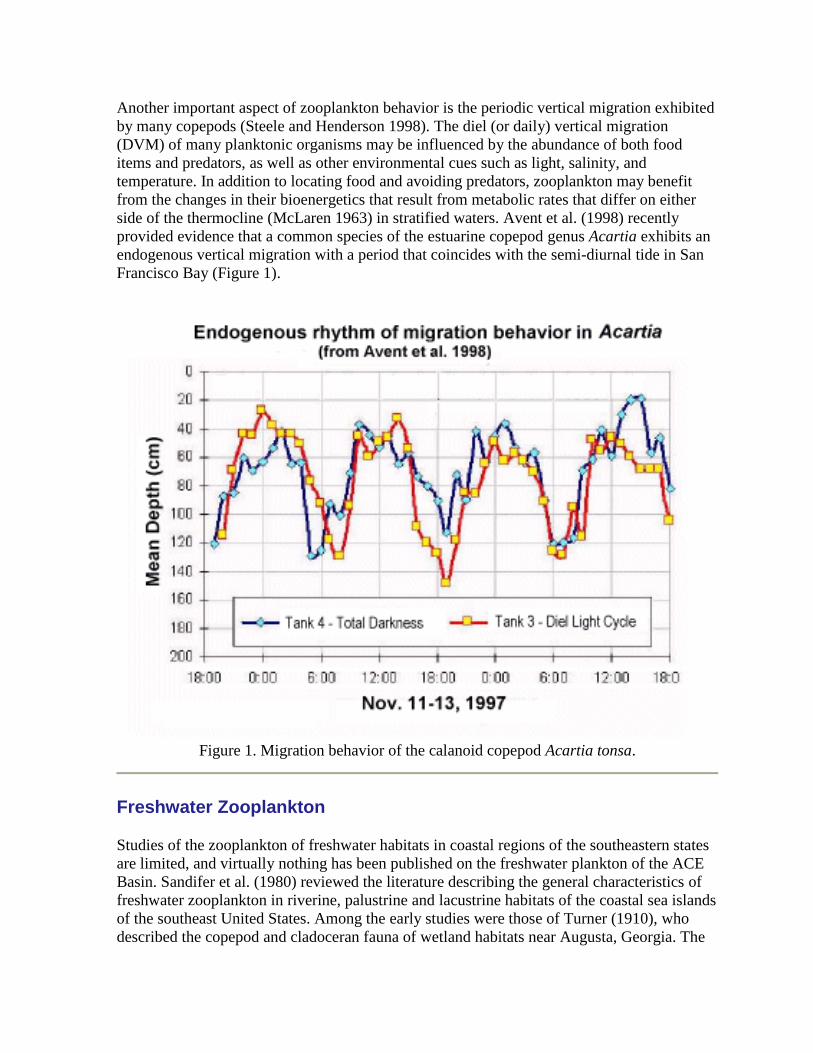
Another important aspect of zooplankton behavior is the periodic vertical migration exhibited
by many copepods (Steele and Henderson 1998). The diel (or daily) vertical migration
(DVM) of many planktonic organisms may be influenced by the abundance of both food
items and predators, as well as other environmental cues such as light, salinity, and
temperature. In addition to locating food and avoiding predators, zooplankton may benefit
from the changes in their bioenergetics that result from metabolic rates that differ on either
side of the thermocline (McLaren 1963) in stratified waters. Avent et al. (1998) recently
provided evidence that a common species of the estuarine copepod genus Acartia exhibits an
endogenous vertical migration with a period that coincides with the semi-diurnal tide in San
Francisco Bay (Figure 1).
Figure 1. Migration behavior of the calanoid copepod Acartia tonsa.
Freshwater Zooplankton
Studies of the zooplankton of freshwater habitats in coastal regions of the southeastern states
are limited, and virtually nothing has been published on the freshwater plankton of the ACE
Basin. Sandifer et al. (1980) reviewed the literature describing the general characteristics of
freshwater zooplankton in riverine, palustrine and lacustrine habitats of the coastal sea islands
of the southeast United States. Among the early studies were those of Turner (1910), who
described the copepod and cladoceran fauna of wetland habitats near Augusta, Georgia. The

zooplankton of the temporary and permanent ponds and ditches sampled by Turner (1910)
included 4 species of calanoid copepods, 10 cyclopoids, 1 harpacticoid, and 24 species of
cladocerans. The copepod Cyclops serulatus and the cladoceran Simocephalus serrulatus
were the most widely distributed taxa.
More recent research has described the species richness and population dynamics of
zooplankton in another type of palustrine habitat, the Carolina bays of the Savannah River
site of the U.S. Department of Energy. These geological features are shallow, poorly drained
elliptical or oval depressions that number in the hundreds of thousands throughout the
Atlantic coastal plain from New Jersey to Florida. Their distribution and ecological status in
South Carolina was addressed by Bennett and Nelson (1991), who noted that 20 of these
features are located in Colleton County; however their precise locations, and consequently
their inclusion within the ACE Basin characterization area, was not described. Sharitz and
Gibbons (1982) discussed the ecology of southeastern Carolina bays, but made no mention of
their zooplankton.
Mahoney et al. (1990) reported that Carolina bays on the upper South Carolina coastal plain
support exceptionally rich zooplankton communities, compared with temporary ponds
elsewhere. These communities are generally dominated early in the wet season by crustacean
taxa with long generation times, such as anostracans, conchostracans and calanoid copepods.
In the 23 bays studied, seven species of the calanoid genus Diaptomus were common; none of
which are typically found in nearby permanent waters. Another group of crustaceans, the
cladocerans, were represented by 26 genera and at least 44 species, many of which showed
considerable overlap between the fauna of the temporary bay ponds and permanent reservoir
waters. Other major invertebrate taxa collected in the Carolina bays were cyclopoid and
harpacticoid copepods; the crustacean orders Amphipoda, Isopoda, and Ostracoda; the insect
orders Ephemeroptera, Odonata, Coleoptera, Trichoptera; the Dipteran families
Ceratopogonidae, Chaoboridae, Chironomidae, and Culicidae; and oligochaetes, nematodes
and aquatic mites.
The population dynamics of zooplankton in Rainbow Bay, one of the 23 Carolina bays
mentioned above, were studied by Taylor and Mahoney (1990). They observed a temporal
pattern in that Carolina bay pond that was typical of many others. The community was
initially dominated by the copepods Diaptomus stagnalis and either Acanthocyclops vernalis
or Diacyclops haueri, but later in the hydroperiod by cyclopoid copepods and cladocerans,
including Daphnia laevis and Simocephalus spp. Experiments conducted on sediments from
the dry pond bed suggested that the time of emergence from resting stages was a determinant
of the initial succession of species in this temporary aquatic habitat. Predation by amphibian
larvae (primarily salamanders) was not sufficient in this pond to limit the abundance of the
predominant zooplankters; thus, population growth was limited for extended periods by
insufficient food.
The zooplankton of lakes and rivers is generally dominated by the free-living non-
photosynthetic protists, rotifers and microcrustaceans; however, the species composition of
these groups may be quite different in lacustrine habitats than in riverine ones (Sandifer et al.
1980). Hudson (1975) described the zooplankton of Keowee Reservoir, a man-made lake in

the South Carolina piedmont region. Of the 53 species of copepods and cladocerans identified
from the reservoir, only about 15 were common in the plankton, while the remainder were
littoral or benthic species. Diaptomus mississippiensis, Mesocyclops edax, and Tropocyclops
prasinus were the most abundant copepods, while Diaphanosoma branchyurum, Holopedium
amazonicum, Daphnia ambigua, and two species of Bosmina were the most abundant
planktonic cladocerans. More recent research on the zooplankton of reservoirs in South
Carolina focused on the spatial heterogeneity of the plankton communities (Betsill and Van
den Avyle 1994) and the effects of thermal stresses caused by nuclear reactor cooling
effluents (Taylor et al. 1993).
Riverine zooplankton of coastal South Carolina has not been intensively studied. Herlong and
Mallin (1985) noted that the zooplankton below an impoundment on Black Creek, South
Carolina, was augmented by the impoundment outfall, making it much denser than that
upstream from the impoundment. Dames and Moore Associates (1975) sampled freshwater
creeks and portions of the Cooper River, collecting 12 taxa of rotifers, 4 taxa of copepods,
and 2 taxa of cladocerans. Rotifers and copepods together comprised nearly 90 percent or
more of the total number of zooplankters at all six sample sites. The most abundant rotifers
were Polyarthra sp. and Keratella cochlearis, while the only genus of copepod identified was
Diaptomus. The cladocerans Bosmia longirostris and Alonella sp. were dominant within that
taxon.
Estuarine Zooplankton
Estuarine Mesozooplankton The abundance of mesozooplankton
in the North Edisto River at Bluff
Point (Figure 2), near the boundary
of the ACE Basin Characterization
Area, was described by Knott
(1980). Surface samples collected
weekly from May 1975 to May
1976 in the river and two adjacent
upland saltwater impoundments
yielded more than 146 unique taxa
(Table 2).
Figure 2. Site sampled for estuarine mesozooplankton by
Knott (1980).
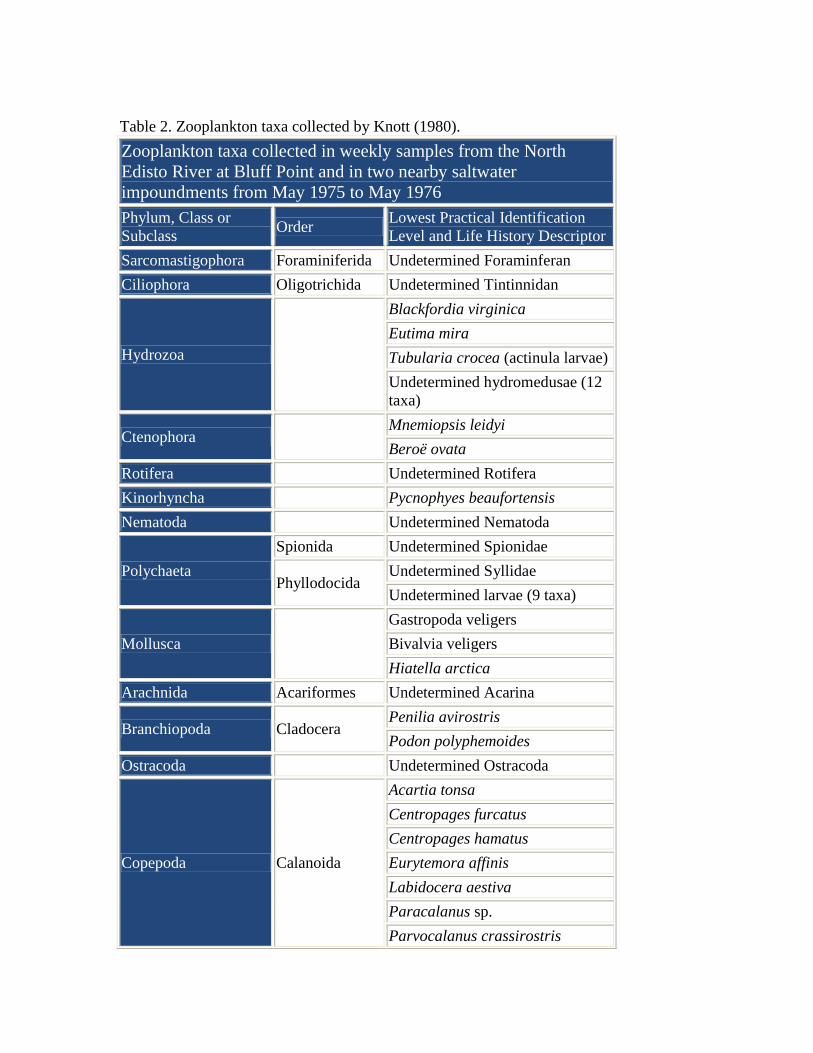
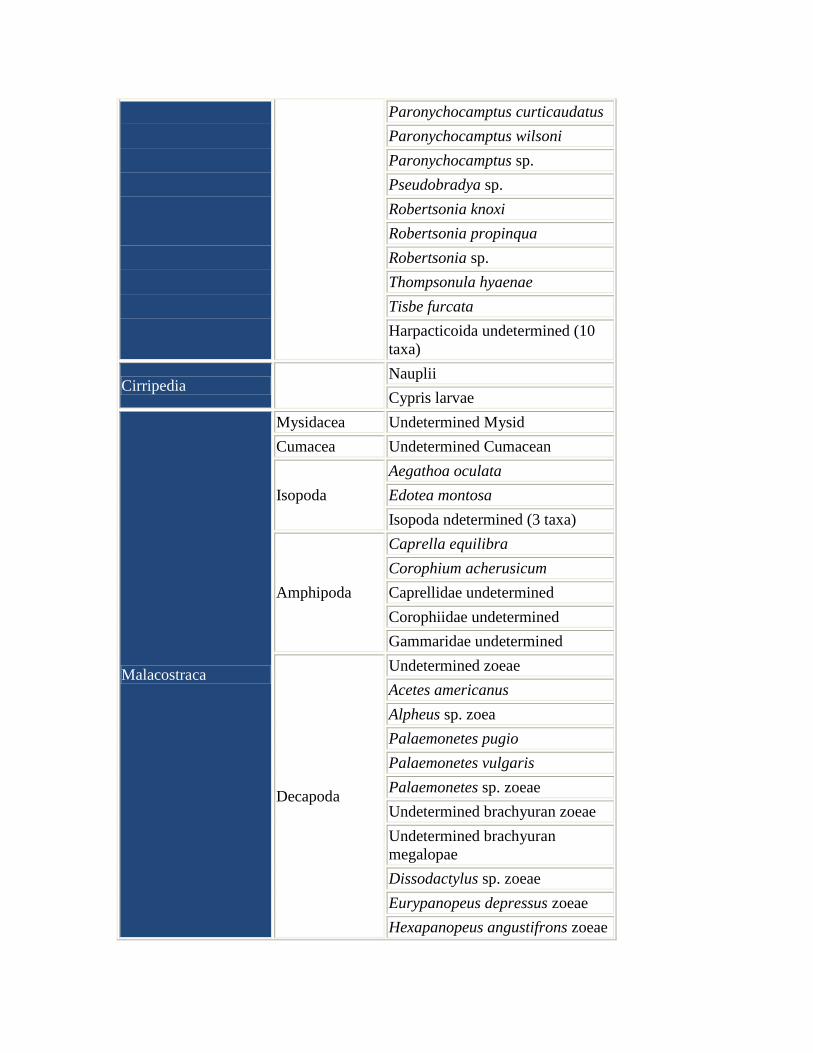
Table 2. Zooplankton taxa collected by Knott (1980).
Zooplankton taxa collected in weekly samples from the North
Edisto River at Bluff Point and in two nearby saltwater
impoundments from May 1975 to May 1976
Phylum, Class or
Subclass Order
Lowest Practical Identification
Level and Life History Descriptor
Sarcomastigophora Foraminiferida Undetermined Foraminferan
Ciliophora Oligotrichida Undetermined Tintinnidan
Hydrozoa
Blackfordia virginica
Eutima mira
Tubularia crocea (actinula larvae)
Undetermined hydromedusae (12
taxa)
Ctenophora Mnemiopsis leidyi
Beroë ovata
Rotifera Undetermined Rotifera
Kinorhyncha Pycnophyes beaufortensis
Nematoda Undetermined Nematoda
Polychaeta
Spionida Undetermined Spionidae
Phyllodocida Undetermined Syllidae
Undetermined larvae (9 taxa)
Mollusca
Gastropoda veligers
Bivalvia veligers
Hiatella arctica
Arachnida Acariformes Undetermined Acarina
Branchiopoda Cladocera Penilia avirostris
Podon polyphemoides
Ostracoda Undetermined Ostracoda
Copepoda Calanoida
Acartia tonsa
Centropages furcatus
Centropages hamatus
Eurytemora affinis
Labidocera aestiva
Paracalanus sp.
Parvocalanus crassirostris
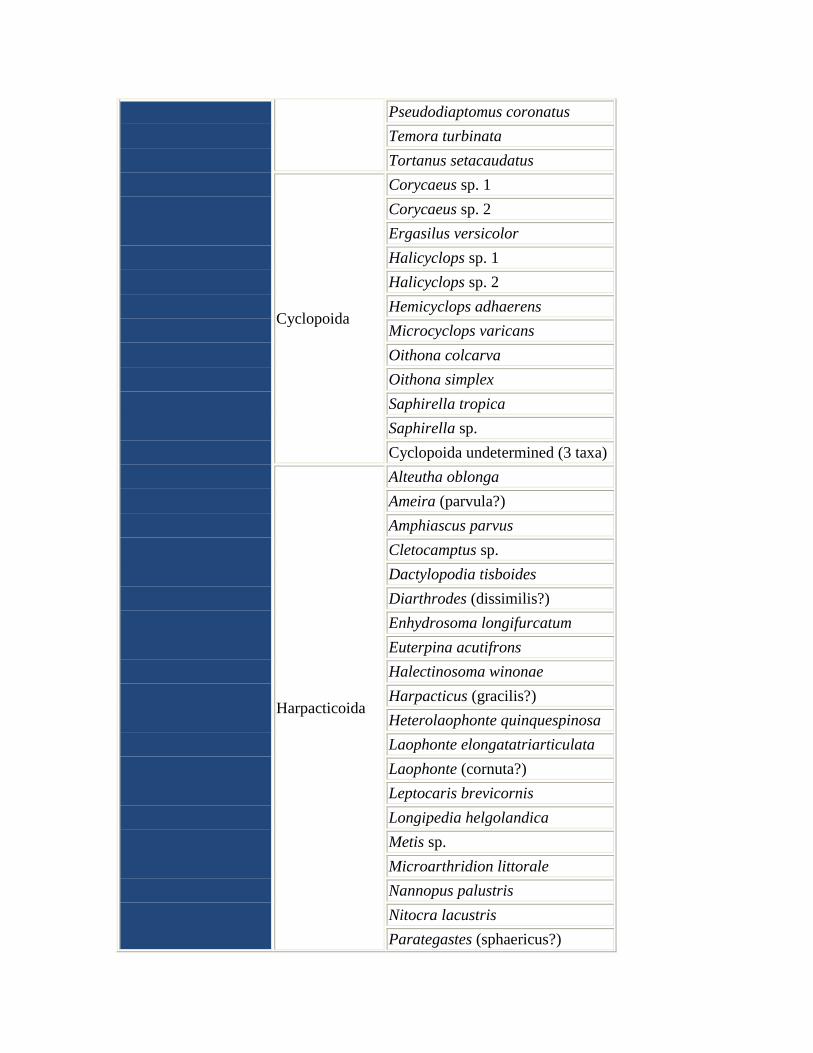
Pseudodiaptomus coronatus
Temora turbinata
Tortanus setacaudatus
Cyclopoida
Corycaeus sp. 1
Corycaeus sp. 2
Ergasilus versicolor
Halicyclops sp. 1
Halicyclops sp. 2
Hemicyclops adhaerens
Microcyclops varicans
Oithona colcarva
Oithona simplex
Saphirella tropica
Saphirella sp.
Cyclopoida undetermined (3 taxa)
Harpacticoida
Alteutha oblonga
Ameira (parvula?)
Amphiascus parvus
Cletocamptus sp.
Dactylopodia tisboides
Diarthrodes (dissimilis?)
Enhydrosoma longifurcatum
Euterpina acutifrons
Halectinosoma winonae
Harpacticus (gracilis?)
Heterolaophonte quinquespinosa
Laophonte elongatatriarticulata
Laophonte (cornuta?)
Leptocaris brevicornis
Longipedia helgolandica
Metis sp.
Microarthridion littorale
Nannopus palustris
Nitocra lacustris
Parategastes (sphaericus?)
Paronychocamptus curticaudatus
Paronychocamptus wilsoni
Paronychocamptus sp.
Pseudobradya sp.
Robertsonia knoxi
Robertsonia propinqua
Robertsonia sp.
Thompsonula hyaenae
Tisbe furcata
Harpacticoida undetermined (10
taxa)
Cirripedia Nauplii
Cypris larvae
Malacostraca
Mysidacea Undetermined Mysid
Cumacea Undetermined Cumacean
Isopoda
Aegathoa oculata
Edotea montosa
Isopoda ndetermined (3 taxa)
Amphipoda
Caprella equilibra
Corophium acherusicum
Caprellidae undetermined
Corophiidae undetermined
Gammaridae undetermined
Decapoda
Undetermined zoeae
Acetes americanus
Alpheus sp. zoea
Palaemonetes pugio
Palaemonetes vulgaris
Palaemonetes sp. zoeae
Undetermined brachyuran zoeae
Undetermined brachyuran
megalopae
Dissodactylus sp. zoeae
Eurypanopeus depressus zoeae
Hexapanopeus angustifrons zoeae
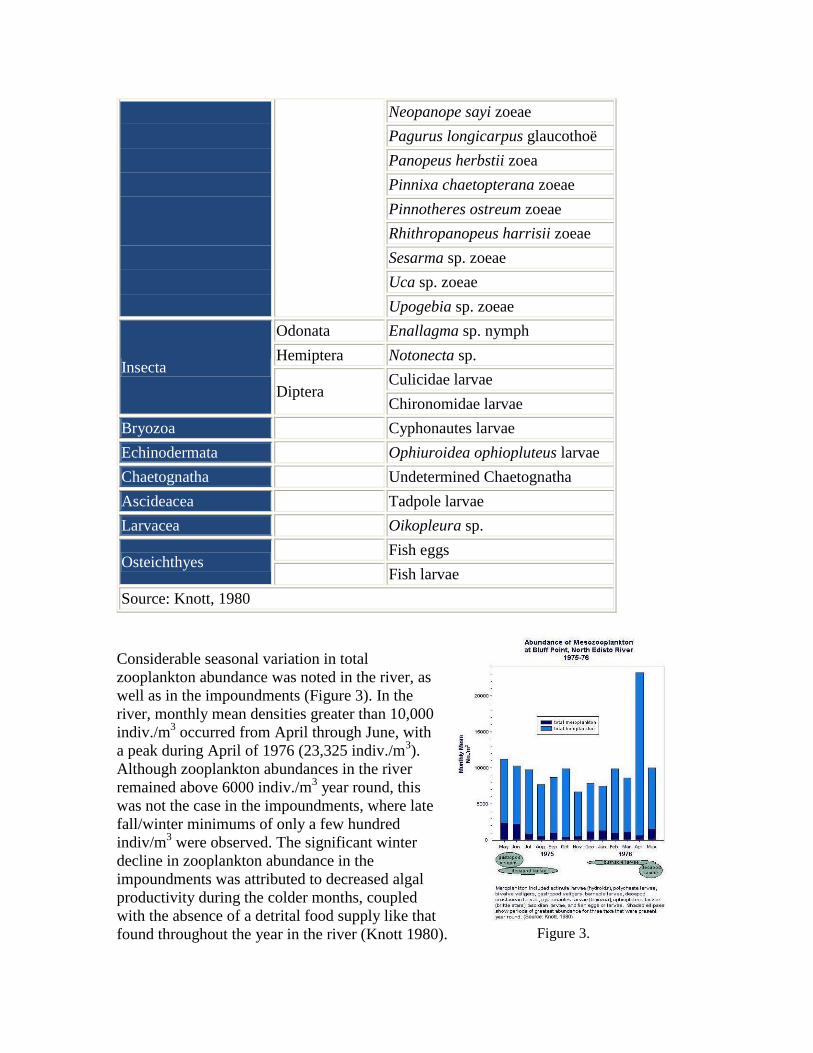
Neopanope sayi zoeae
Pagurus longicarpus glaucothoë
Panopeus herbstii zoea
Pinnixa chaetopterana zoeae
Pinnotheres ostreum zoeae
Rhithropanopeus harrisii zoeae
Sesarma sp. zoeae
Uca sp. zoeae
Upogebia sp. zoeae
Insecta
Odonata Enallagma sp. nymph
Hemiptera Notonecta sp.
Diptera Culicidae larvae
Chironomidae larvae
Bryozoa Cyphonautes larvae
Echinodermata Ophiuroidea ophiopluteus larvae
Chaetognatha Undetermined Chaetognatha
Ascideacea Tadpole larvae
Larvacea Oikopleura sp.
Osteichthyes Fish eggs
Fish larvae
Source: Knott, 1980
Considerable seasonal variation in total
zooplankton abundance was noted in the river, as
well as in the impoundments (Figure 3). In the
river, monthly mean densities greater than 10,000
indiv./m3 occurred from April through June, with
a peak during April of 1976 (23,325 indiv./m3).
Although zooplankton abundances in the river
remained above 6000 indiv./m3 year round, this
was not the case in the impoundments, where late
fall/winter minimums of only a few hundred
indiv/m3 were observed. The significant winter
decline in zooplankton abundance in the
impoundments was attributed to decreased algal
productivity during the colder months, coupled
with the absence of a detrital food supply like that
found throughout the year in the river (Knott 1980). Figure 3.
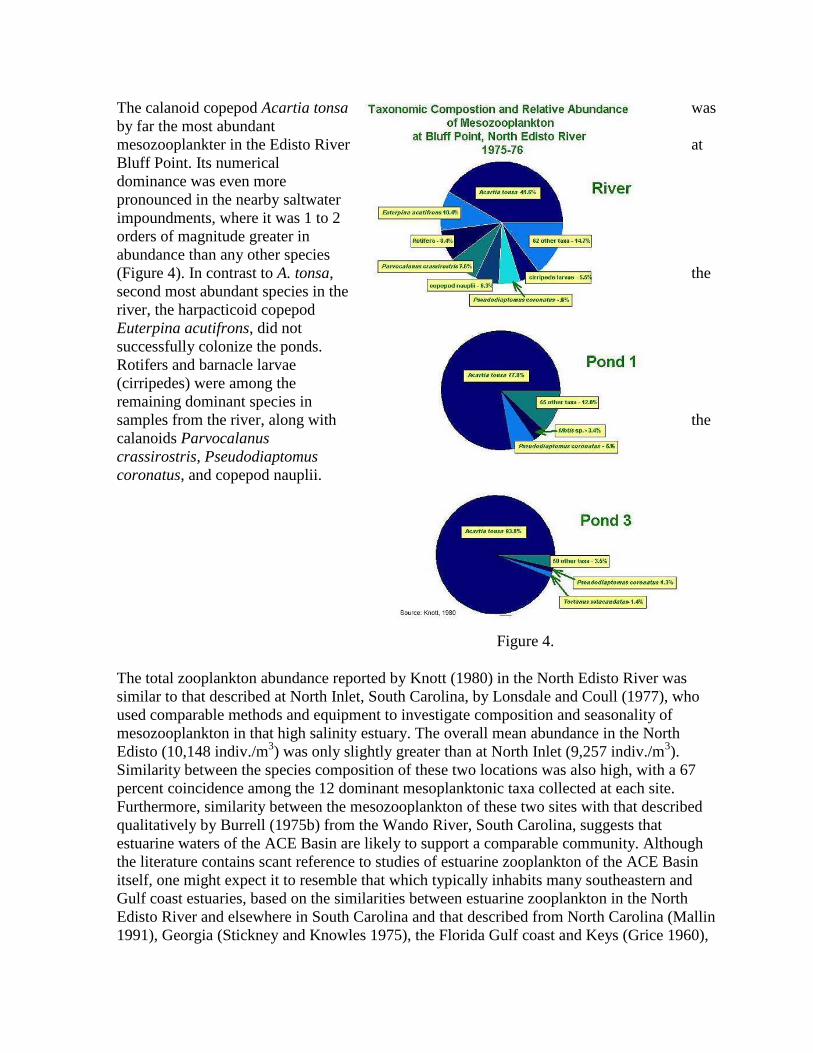
The calanoid copepod Acartia tonsa was
by far the most abundant
mesozooplankter in the Edisto River at
Bluff Point. Its numerical
dominance was even more
pronounced in the nearby saltwater
impoundments, where it was 1 to 2
orders of magnitude greater in
abundance than any other species
(Figure 4). In contrast to A. tonsa, the
second most abundant species in the
river, the harpacticoid copepod
Euterpina acutifrons, did not
successfully colonize the ponds.
Rotifers and barnacle larvae
(cirripedes) were among the
remaining dominant species in
samples from the river, along with the
calanoids Parvocalanus
crassirostris, Pseudodiaptomus
coronatus, and copepod nauplii.
Figure 4.
The total zooplankton abundance reported by Knott (1980) in the North Edisto River was
similar to that described at North Inlet, South Carolina, by Lonsdale and Coull (1977), who
used comparable methods and equipment to investigate composition and seasonality of
mesozooplankton in that high salinity estuary. The overall mean abundance in the North
Edisto (10,148 indiv./m3) was only slightly greater than at North Inlet (9,257 indiv./m
3).
Similarity between the species composition of these two locations was also high, with a 67
percent coincidence among the 12 dominant mesoplanktonic taxa collected at each site.
Furthermore, similarity between the mesozooplankton of these two sites with that described
qualitatively by Burrell (1975b) from the Wando River, South Carolina, suggests that
estuarine waters of the ACE Basin are likely to support a comparable community. Although
the literature contains scant reference to studies of estuarine zooplankton of the ACE Basin
itself, one might expect it to resemble that which typically inhabits many southeastern and
Gulf coast estuaries, based on the similarities between estuarine zooplankton in the North
Edisto River and elsewhere in South Carolina and that described from North Carolina (Mallin
1991), Georgia (Stickney and Knowles 1975), the Florida Gulf coast and Keys (Grice 1960),

and the Gulf of Mexico (Buskey 1993).
The relative contribution of meroplanktonic organisms to total zooplankton abundance was
uniformly low (3 to 21 percent) in the North Edisto River. The predominant meroplanktonic
taxa were gastropod veligers (which peaked in spring), barnacle larvae (which peaked in
winter/early spring), and decapod crustacean larvae (which peaked in spring and summer).
Although a variety of decapod crustacean larvae were collected (at least 20 species ), they
contributed relatively few numbers to the total mesoplankton community (Table 2). Many of
the planktonic larvae of decapods are macroplanktonic, and they may not have been
efficiently captured by the 30 centimeter (12 inch) diameter net with 147 mm mesh deployed
by Knott (1980). Samples collected in North Inlet, South Carolina, by Lonsdale and Coull
(1977), using a nearly identical plankton net, also contained relatively few meroplanktonic
larvae (25 percent by number), suggesting a sampling bias against some of the larger
crustacean larvae that might be able to avoid capture by these small fine-meshed nets.
Copepods were predominant among the mesozooplankton of the North Edisto River, both in
terms of species richness (63 different species) and abundance (78 percent in the river; 95 to
98 percent in the ponds)(Knott 1980). The copepods comprised 24 truly planktonic species in
the orders Calanoida and Cyclopoida and a rich representation of the Harpacticoida (39
species), all but three of which were typically benthic organisms that were suspended at the
shallow river station by tidal turbulence.
Estuarine Macrozooplankton Early studies of macrozooplankton in South Carolina targeted the larval stages of
commercially important crustaceans. Fisheries researchers conducted periodic plankton
sampling in the Wando, Cooper and Ashley Rivers near Charleston, South Carolina, and in
the Santee River to the north, using nets designed to capture macroplankton (Bears Bluff
Laboratories, Inc. 1964). In addition to larval crustaceans, two taxa were among the
numerically dominant organisms at most stations: copepods (which were not quantitatively
represented because of the coarse mesh nets) and the medusa stage (jellyfish) of
undifferentiated species of coelenterates. Burrell (1975b) also found coelenterate medusae to
be seasonally abundant in the Wando River, including Blackfordia virginica and Nemopsis
bachei, along with the comb jelly Mnemiopsis. Hester (1976) and Calder and Hester (1978)
described a rich planktonic coelenterate fauna in South Carolina estuaries.
SCDNR Ingress Studies
Blue crab and white shrimp are two decapod crustaceans with significant commercial value in
South Carolina. Both of these species have a life history that includes offshore larval
development and an estuarine nursery habitat (Figures 5 and 6).
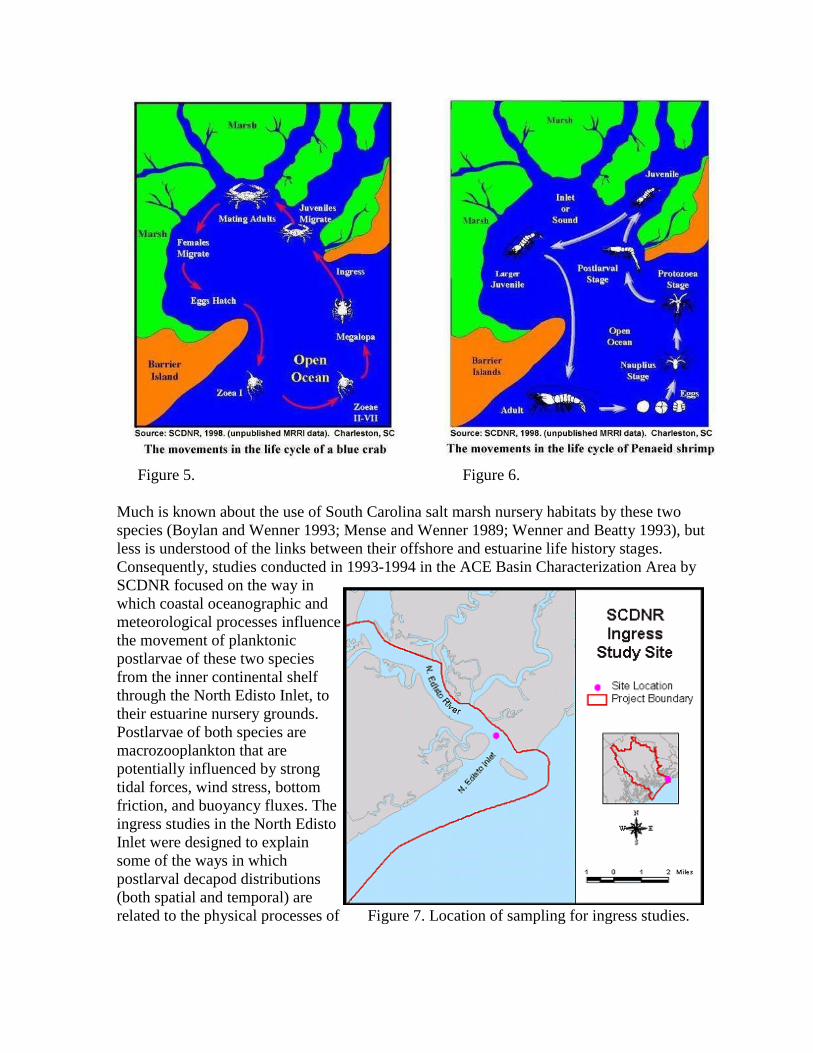
Figure 5. Figure 6.
Much is known about the use of South Carolina salt marsh nursery habitats by these two
species (Boylan and Wenner 1993; Mense and Wenner 1989; Wenner and Beatty 1993), but
less is understood of the links between their offshore and estuarine life history stages.
Consequently, studies conducted in 1993-1994 in the ACE Basin Characterization Area by
SCDNR focused on the way in
which coastal oceanographic and
meteorological processes influence
the movement of planktonic
postlarvae of these two species
from the inner continental shelf
through the North Edisto Inlet, to
their estuarine nursery grounds.
Postlarvae of both species are
macrozooplankton that are
potentially influenced by strong
tidal forces, wind stress, bottom
friction, and buoyancy fluxes. The
ingress studies in the North Edisto
Inlet were designed to explain
some of the ways in which
postlarval decapod distributions
(both spatial and temporal) are
related to the physical processes of Figure 7. Location of sampling for ingress studies.

transport. These specifically address ways in which periodic phenomena such as tides and
daily or lunar cycles, and less predictable ones such as wind-generated currents, interact to
influence the transport of planktonic larvae of these two species through the inlet and into the
estuary.
Blue crabs spawn in the lower estuary, and
their planktonic larvae are exported to the
coastal ocean where they develop to the
postlarval stage known as the megalopa
(Figure 5). Subsequent ingress of
megalopae through coastal inlets precedes
their settlement and recruitment to the salt
marsh nursery habitat. In South Carolina,
ingress and settlement peak between
August and November.
Megalopal ingress was investigated during
the spring and fall of 1993 and 1994 by
Knott and others (unpublished data), who
examined plankton collections from the
North Edisto River Inlet. Plankton samples
were classified based on wind stress
conditions at the time of their collection.
With the exception of the spring 1993
cruise, when very few megalopae were
collected, ingress was significantly greater when the wind blew up the coast and slightly
onshore. A quantitative vector-scalar correlation (the correlation is between wind stress
vectors and the corresponding scalar values of megalopal density, thereby accounting for the
magnitude as well as the precise direction of wind stress) also showed a significant
association between ingress and upcoast winds with an onshore component during the fall
1993 cruise. Although a dissimilar result was obtained for the fall 1994 cruise, when
downcoast winds were significantly correlated with ingress, these apparently contrasting
results were both consistent with 3-D model simulations of the study area, which predict
significant ingress of passive particles released at the surface during both upwelling and
downwelling conditions (Blanton et al. 1998).
The transport of blue crab megalopae may differ from that described for postlarval white
shrimp largely due to differences in the behavior and vertical distribution of the two species
in near shore coastal waters. Penaeus setiferus broadcasts its eggs on the seafloor of the
shallow continental shelf over several months in the spring and early summer. After the
hatchlings pass through several larval stages, they reach the postlarval stage, which ingresses
through inlets to the salt marsh nursery grounds (Figure 7). In South Carolina, the period of
peak ingress usually occurs in June (DeLancey and others 1994).
Wenner et al. (1998) described the importance of wind stress on the transport of white shrimp

postlarvae through the North Edisto River inlet
during 1993 and 1994. Their study demonstrated
that ingress of white shrimp is enhanced by easterly
to northeasterly winds, which push surface waters
toward the coast. This forcing may move many
shrimp postlarvae into the shallow area near the
inlet mouth, where they can be easily transported
through the inlet into the estuary. The authors noted
that variability in winds and other physical
processes may explain much of the inter-annual
variability in transport of larvae to their nursery
grounds. white shrimp postlarva
Studies by other researchers working in the North Inlet estuary of South Carolina provide
additional insight into the composition and dynamics of a macroplanktonic community that is
likely to closely resemble that in the ACE Basin. Tidal, day-night, and day-to-day patterns of
macrozooplanktonic abundance were described by Houser and Allen (1996), who observed
large pulses of crab and shrimp larvae originating from nocturnal hatching events in the upper
reaches of a tidal creek. The most abundant organisms in their 6-month series of daily
samples were fish larvae (primarily the goby Gobiosoma), larval and postlarval decapod
crustaceans (including the snapping shrimps Alpheus spp., the fiddler crabs Uca spp., the
grass shrimps Palaemonetes spp., and the commercially valuable shrimps Penaeus spp.),
juvenile bivalves, the holoplanktonic chaetognaths (arrow worms), gammarid amphipods and
hydromedusae. Further seaward in Town Creek, near the inlet of the same estuary, Moore and
Reis (1983) observed a similar macrozooplanktonic community dominated by the mysid
crustacean Neomysis americanus. At that locality they also noted greater numbers of the
holoplanktonic decapod crustaceans Acetes americanus and Lucifer faxoni, which are more
typical residents of shallow coastal oceanic environments and high salinity inlets. Further
documentation of such tidal incursions of coastal macrozooplankton into this estuary was
provided by Costello and Stancyk (1983), who described the mechanism by which the
macroplanktonic appendicularian Oikopleura dioica enters the North Inlet from the ocean.
References
Allen, D. M., W. S. Johnson, and V. Ogburn-Matthews. 1995. Trophic relationships and seasonal utilization of
salt-marsh creeks by zooplanktivorous fishes. Environmental Biology of Fishes 42(1):37-50.
Avent, S. R., S. M. Bollens, and S. P. Troia. 1998. Diel vertical migration in zooplankton: experimental
investigations using video-microscopy and plankton mini-towers. In American Geophysical Union (ed).
1998 Ocean Sciences Meeting, 9-13 February 1998, San Diego, CA. American Geophysical Union,
Washington, DC.
Bears Bluff Laboratories, Inc. 1964. Biological studies of Charleston Harbor, S.C. and the Santee River.
Submitted to the USFWS, contract no. 14-16-0004-1024.
Bennett, S. H. and J. B. Nelson. 1991. Distribution and status of Carolina bays in South Carolina. Nongame and
Heritage Trust Section, South Carolina Wildlife and Marine Resources Department.
Betsill, R. K. and M. J. Van den Avyle. 1994. Spatial heterogeneity of reservoir zooplankton: a matter of
timing? Hydrobiologia 277(1):63-70.
Blanton, J. O., F. E. Werner, A. Kapolnai, B. O. Blanton, D. Knott, and E. L. Wenner. 1998. Wind-generated

transport of ficticious passive larvae into shallow tidal estuaries. Fisheries Oceanography 8 (suppl. 2):210-
223.
Bowman, T. E. 1971. The distribution of calanoid copepods off the southeastern United States between Cape
Hatteras and southern Florida. Smithsonian Contributions to Zoology 96.
Boylan, J. M. and E. L. Wenner. 1993. Settlement of brachyuran megalopae in a South Carolina estuary. Marine
Ecology Progress Series 97:237-246.
Brownlee, D. C. and F. Jacobs. 1987. Mesozooplankton and microzooplankton in the Chesapeake Bay. In S.K.
Majumdar, L.W. Hall Jr., and H.M. Austin (eds). Contaminant Problems and Management of Living
Chesapeake Bay Resources. Pennsylvania Academy of Science, Easton, PA.
Burrell, V. G., Jr. 1975a. The relationship of proposed offshore nuclear power plants to marine fisheries of the
South Atlantic region of the United States. In Institute of Electrical and Electronic Engineers (eds).
Proceedings of the Ocean '75 Conference, San Diego, CA. Institute of Electrical and Electronic Engineers,
New York, NY.
Burrell, V. G., Jr. 1975b. A preliminary list of the most common species of zooplankton found in the Wando
River, August 1972-August 1974. Unpublished data from the South Carolina Department of Natural
Resources, Charleston, SC. Summarized in Sandifer et al. 1980.
Buskey, E. J. 1993. Annual pattern of micro- and mesozooplankton abundance and biomass in a subtropical
estuary. Journal of Plankton Research 15(8):907-924.
Calder, D. R. and B. S. Hester. 1978. Phylum Cnidaria In An annotated checklist of the biota of the coastal zone
of South Carolina, Zingmark,R. G. (ed.) University of South Carolina Press, Columbia, SC.
Costello, J. and S. E. Stancyk. 1983. Tidal influence upon appendicularian abundance in North Inlet estuary,
South Carolina. Journal of Plankton Research 5(2):263-277.
Daborn, G. R. and M. Brylinsky. 1981. Zooplankton diversity and species associations in the inner Bay of
Fundy. Estuaries 4(3):253.
Dames and Moore Associates. 1975. Environmental assessment report on a proposed chemical plant, Berkeley
County, South Carolina. Prepared for Amoco Chemicals Corporation. Park Ridge, IL.
Davis, B. M. and J. W. Foltz. 1991. Food of blueback herring and threadfin shad in Jocassee Reservoir, South
Carolina. Transactions of the American Fisheries Society 120(5):605-613.
DeLancey, L. B., J. E. Jenkins, and J. D. Whitaker. 1994. Results of long-term, seasonal sampling for Penaeus
postlarvae at Breach Inlet, South Carolina. Fishery Bulletin 92:633-640.
Durbin, A. G. and E. G. Durbin. 1981. Standing stock and estimated production rates of phytoplankton and
zooplankton in Narragansett Bay, Rhode Island. Estuaries 4(1):24-41.
Epifanio, C. E. 1988. Transport of invertebrate larvae between estuaries and the continental shelf. American
Fisheries Society Symposium 3:104-114.
Fulton, R. S. III. 1983. Interactive effects of temperature and predation on an estuarine zooplankton community.
Journal of Experimental Marine Biology and Ecology 72:67-81.
Fulton, R. S. III. 1984. Predation, production and the organization of an estuarine copepod community. Journal
of Plankton Research 6(3):399-415.
Grice, G. D. 1960. Calanoid and cyclopoid copepods collected from the Florida Gulf coast and Florida Keys in
1954 and 1955. Bulletin of Marine Science 10:217-226.
Grice, G. D. and N. H. Marcus. 1981. Dormant eggs of marine copepods. Oceanography and Marine Biology:
An Annual Review 19:125-140.
Heip, C. H. R., N. K. Goosen, P. M. J. Herman, J. Kromkamp, J. J. Middelburg, and K. Soetaert. 1995.
Production and consumption of biological particles in temperate tidal estuaries. Oceanography and Marine
Biology: An Annual Review 33:1-149.
Herlong, D. D. and M. A. Mallin. 1985. The benthos-plankton relationship upstream and downstream of a
blackwater impoundment. Journal of Freshwater Ecology 3(1):47-59.
Hester, B. S. 1976. Distribution and seasonality of hydromedusae in South Carolina estuaries. Masters of
Science Thesis, College of Charleston, Charleston, SC.
Hopkins, T. L. 1977. Zooplankton distribution in surface waters of Tampa Bay, Florida. Bulletin of Marine
Science 27(3):467-478.
Houser, D. S. and D. M. Allen. 1996. Zooplankton dynamics in an intertidal salt-marsh basin. Estuaries
19(3):659-673.
Hudson, P. L. 1975. 1975 annual report: southeastern reservoir investigation. United States Fish and Wildlife
Service. Clemson, SC.

Kleppel, G. S., C. A. Burkart, L. Houchin, and C. Tomas. 1998. Egg production in the copepod Acartia tonsa in
Florida Bay during summer. The roles of food environment and diet. Estuaries 21(2):328-339.
Knott, D. M. 1980. The zooplankton of the North Edisto River and two artificial saltwater impoundments.
Masters of Science Thesis, College of Charleston, Charleston, SC.
Knott, D., J. Amft, C. Barans, J. Blanton, B. Stender, P. Verity, E. Wenner, and F. Werner. (in preparation,
1998). The influence of wind forcing on the ingress of blue crab megalopae into a South Carolina coastal
inlet.
Lonsdale, D. J. and B. C. Coull. 1977. Composition and seasonality of zooplankton of North Inlet, South
Carolina. Chesapeake Science 18(3):272-283.
Mahoney, D. L., M. A. Mort, and B. E. Taylor. 1990. Species richness of calanoid copepods, cladocerans and
other branchiopods in Carolina Bay temporary ponds. American Midlands Naturalist 123(2):244-258.
Mallin, M. A. 1991. Zooplankton abundance and community structure in a mesohaline North Carolina estuary.
Estuaries 14(4):481-488.
McLaren, I. A. 1963. Effects of temperature on growth of zooplankton and the adaptive value of vertical
migration. Journal of the Fisheries Research Board of Canada 26:199-220.
Mense, D. J. and E. L. Wenner. 1989. Distribution and abundance of early life history stages of the blue crab,
Callinectes sapidus, in tidal marsh creeks near Charleston, SC. Estuaries 12(3):157-168.
Moore, R. H. and R. R. Reis. 1983. Analysis of spatial and temporal variations in biomass and community
structure of motile organisms in Town Creek, a South Carolina tidal pass. Contributions in Marine Science
26:111-125.
Omori, M. and T. Ikeda. 1984. Methods in Marine Zooplankton Ecology. John Wiley & Sons. New York,NY.
Ragotzkie, R. A. 1959. Plankton productivity in estuarine waters of Georgia. Publications of the Institute of
Marine Science, University of Texas 6:146-158.
Roman, M. R., M. R. Reeve, and J. L. Froggatt. 1983. Carbon production and export from Biscayne Bay,
Florida. Vol. I: Temporal patterns in primary production, seston and zooplankton. Estuarine, Coastal and
Shelf Science 17:45-59.
Sanders, R. W. and S. A. Wickham. 1993. Planktonic protozoa and metazoa: predation, food quality and
population control. Marine Microbial Food Webs 7(2):197-223.
Sandifer, P. A., J. V. Miglarese, D. R. Calder, J. J. Manzi, and L. A. Barclay. 1980. Ecological characterization
of the Sea Island coastal region of South Carolina and Georgia. Vol. III: Biological features of the
characterization area. U.S. Fish and Wildlife Service, Office of Biological Services, Washington, D.C.
FWS/OBS-79/42.
Sharitz, R. R. and J. W. Gibbons. 1982. The ecology of southeastern shrub bogs (Pocosins) and Carolina bays: a
community profile. United States Fish and Wildlife Service, Division of Biological Services, Washington,
DC. FWS/OBS-82/04.
Steele, J. H. and E. W. Henderson. 1998. Vertical migration of copepods. Journal of Plankton Research
20(4):787-799.
Stickney, R. R. and S. C. Knowles. 1975. Summer zooplankton distribution in a Georgia estuary. Marine
Biology 33:147-154.
Taylor, B. E. and D. L. Mahoney. 1990. Zooplankton in Rainbow Bay, a Carolina bay pond: population
dynamics in a temporary habitat. Freshwater Biology 24:597-612.
Taylor, B. E., A. E. DeBiase, and D. L. Mahoney. 1993. Development of the zooplankton assemblage in a new
cooling reservoir. Archiv fur Hydrobiologie 128(2):129-148.
Turner, C. H. 1910. Ecological notes on the cladocera and copepoda of Augusta, Georgia, with description of
new or little known species. Transactions of the Academy of Sciences of St. Louis 19(10):151-176.
University of Georgia Marine Institute. 1971. Notes on the natural history and invertebrate fauna of the upper
north Newport River In An ecological survey of the north and south Newport Rivers and adjacent waters
with respect to possible effects of treated kraft mill effluent. Final Report to the Georgia Water Quality
Control Board. UGA #D2422-122.
Van Engel, W. A. and E. B. Joseph. 1968. The characterization of coastal and estuarine fish nursery grounds as
natural communities. U. S. Department of the Interior, Fish and Wildlife Service, Bureau of Commercial
Fisheries.
Wenner, E. L. and H. R. Beatty. 1993. Utilization of shallow estuarine habitats in South Carolina, U.S.A., by
postlarval and juvenile stages of Penaeus spp. (Decapoda: Penaeidae). Journal of Crustacean Biology
13(2):280-295.

Wenner, E., D. Knott, J. Blanton, C. Barans, and J. Amft. 1998. Roles of tidal and wind- generated currents in
transporting white shrimp (Penaeus setiferus) postlarvae through a South Carolina (USA) inlet. Journal of
Plankton Research 20(12):2333-2356.
Related Documents











