White matter and cognitive function in schizophrenia Andrew J. Dwork 1,2,3 , Branislav Mancevski 1,2 and Gorazd Rosoklija 1,2,4 1 Department of Neuroscience, New York State Psychiatric Institute, New York, NY, USA Departments of 2 Psychiatry and 3 Pathology, College of Physicians and Surgeons of Columbia University, New York, NY, USA 4 Macedonian Academy of Sciences and Arts, Skopje, Republic of Macedonia Abstract Abnormalities of cerebral white matter, oligodendrocytes, and myelin have been observed in schizo- phrenia with in-vivo imaging and post-mortem biochemistry. White-matter abnormalities are also fre- quently associated with cognitive impairment in both healthy and diseased individuals, and cognitive dysfunction is an important component of schizophrenia. While many studies have documented these associations, only a handful have examined the role of white matter in cognitive function in schizophrenia. In this paper, we explore what is known about white-matter deficits in relation to schizophrenia, cognitive deficits, or both together, in order to generate a theoretical model for the role that compromise of white matter might play in producing cognitive impairment in schizophrenia. Received 30 August 2006; Reviewed 12 October 2006; Revised 15 January 2006; Accepted 18 January 2006; First published online 21 February 2007 Key words : Cognition, myelin, neuropathology, oligodendrocyte, schizophrenia. Introduction Abnormalities of cerebral white matter, myelin, and oligodendrocytes have been reported in schizophrenia (reviewed in Davis et al., 2003; Sullivan and Pfefferbaum, 2003 ; Walterfang et al., 2006). White- matter abnormalities are also a common feature of dementia, particularly, but not exclusively, when of vascular origin (Bronge, 2002 ; Chin and Goldman, 1996 ; Hachinski, 1990). Cognitive impairment is now recognized as a core feature of schizophrenia. In young individuals with schizophrenia, cognitive defi- cits are often detectable on psychological testing (Goldberg and Weinberger, 1988 ; Green et al., 2004). In the majority of elderly individuals with schizophrenia, cognitive impairments are of a severity that would be characterized as dementia in individuals without schizophrenia (Arnold et al., 1995 ; Davidson et al., 1995; Dwork et al., 1998). The neuropathological substrates of cognitive im- pairment in schizophrenia are unknown. In elderly individuals with schizophrenia, there is no increase in Alzheimer-type changes (Arnold et al., 1998 ; Dwork et al., 1998; Purohit et al., 1998), and the cholinergic deficits found in Alzheimer’s disease are not present (Haroutunian et al., 1994). Although diminished cog- nitive reserve in schizophrenia may increase suscep- tibility to the cognitive effects of mild Alzheimer-type changes that commonly occur with age, many elderly individuals with schizophrenia and severe cognitive impairment lack such changes (Dwork et al., 1998 ; Ortakov et al., 1999). Two neuropathological processes, however, are common to dementia and schizophrenia. The first is a loss of presynaptic and post-synaptic elements in cerebral cortex (Bigio et al., 2001; Browning et al., 1993; Davidsson et al., 1999; Dawson and Hallenbeck, 1996; DeKosky and Scheff, 1990; Eastwood et al., 1995 ; Eastwood and Harrison, 1995, 1999; Gabriel et al., 1997; Garey et al., 1998; Glantz and Lewis, 1997, 2000 ; Goto and Hirano, 1990 ; Hamos et al., 1989; Harrison, 1999; Honer et al., 1992, 1997; Honer and Young, 2004 ; Karson et al., 1999 ; Law et al., 2004; Masliah et al., 1989, 1991; Perdahl et al., 1984; Perrone-Bizzozero et al., 1996; Regeur et al., 1994; Rosoklija et al., 2000 ; Tcherepanov and Sokolov, 1997; Terry et al., 1991; Vawter et al., 1999). The second, which is the focus of this study, is an alter- ation of cerebral white matter. Although white-matter changes appear to be less severe in schizophrenia than in many forms of dementia, they could nonetheless be an intrinsic com- ponent of schizophrenia that contributes eventually Address for correspondence : Dr A. J. Dwork, 1051 Riverside Dr., Unit 62, New York, NY 10032, USA. Tel. : 212-543-5563 Fax : 212-543-6017 E-mail : [email protected] International Journal of Neuropsychopharmacology (2007), 10, 513–536. Copyright f 2007 CINP doi:10.1017/S1461145707007638 SPECIAL SECTION CINP

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

White matter and cognitive functionin schizophrenia
Andrew J. Dwork1,2,3, Branislav Mancevski1,2 and Gorazd Rosoklija1,2,41 Department of Neuroscience, New York State Psychiatric Institute, New York, NY, USA
Departments of 2 Psychiatry and 3 Pathology, College of Physicians and Surgeons of Columbia University, New York, NY, USA4 Macedonian Academy of Sciences and Arts, Skopje, Republic of Macedonia
Abstract
Abnormalities of cerebral white matter, oligodendrocytes, and myelin have been observed in schizo-
phrenia with in-vivo imaging and post-mortem biochemistry. White-matter abnormalities are also fre-
quently associated with cognitive impairment in both healthy and diseased individuals, and cognitive
dysfunction is an important component of schizophrenia. While many studies have documented these
associations, only a handful have examined the role of white matter in cognitive function in schizophrenia.
In this paper, we explore what is known about white-matter deficits in relation to schizophrenia, cognitive
deficits, or both together, in order to generate a theoretical model for the role that compromise of white
matter might play in producing cognitive impairment in schizophrenia.
Received 30 August 2006; Reviewed 12 October 2006; Revised 15 January 2006; Accepted 18 January 2006;
First published online 21 February 2007
Key words : Cognition, myelin, neuropathology, oligodendrocyte, schizophrenia.
Introduction
Abnormalities of cerebral white matter, myelin, and
oligodendrocytes have been reported in schizophrenia
(reviewed in Davis et al., 2003; Sullivan and
Pfefferbaum, 2003; Walterfang et al., 2006). White-
matter abnormalities are also a common feature of
dementia, particularly, but not exclusively, when of
vascular origin (Bronge, 2002; Chin and Goldman,
1996; Hachinski, 1990). Cognitive impairment is now
recognized as a core feature of schizophrenia. In
young individuals with schizophrenia, cognitive defi-
cits are often detectable on psychological testing
(Goldberg andWeinberger, 1988; Green et al., 2004). In
the majority of elderly individuals with schizophrenia,
cognitive impairments are of a severity that would be
characterized as dementia in individuals without
schizophrenia (Arnold et al., 1995; Davidson et al.,
1995; Dwork et al., 1998).
The neuropathological substrates of cognitive im-
pairment in schizophrenia are unknown. In elderly
individuals with schizophrenia, there is no increase in
Alzheimer-type changes (Arnold et al., 1998; Dwork
et al., 1998; Purohit et al., 1998), and the cholinergic
deficits found in Alzheimer’s disease are not present
(Haroutunian et al., 1994). Although diminished cog-
nitive reserve in schizophrenia may increase suscep-
tibility to the cognitive effects of mild Alzheimer-type
changes that commonly occur with age, many elderly
individuals with schizophrenia and severe cognitive
impairment lack such changes (Dwork et al., 1998;
Ortakov et al., 1999). Two neuropathological processes,
however, are common to dementia and schizophrenia.
The first is a loss of presynaptic and post-synaptic
elements in cerebral cortex (Bigio et al., 2001;
Browning et al., 1993; Davidsson et al., 1999; Dawson
and Hallenbeck, 1996; DeKosky and Scheff, 1990;
Eastwood et al., 1995; Eastwood and Harrison, 1995,
1999; Gabriel et al., 1997; Garey et al., 1998; Glantz
and Lewis, 1997, 2000; Goto and Hirano, 1990; Hamos
et al., 1989; Harrison, 1999; Honer et al., 1992, 1997;
Honer and Young, 2004; Karson et al., 1999; Law et al.,
2004; Masliah et al., 1989, 1991; Perdahl et al., 1984;
Perrone-Bizzozero et al., 1996; Regeur et al., 1994;
Rosoklija et al., 2000; Tcherepanov and Sokolov,
1997; Terry et al., 1991; Vawter et al., 1999). The
second, which is the focus of this study, is an alter-
ation of cerebral white matter.
Although white-matter changes appear to be less
severe in schizophrenia than in many forms of
dementia, they could nonetheless be an intrinsic com-
ponent of schizophrenia that contributes eventually
Address for correspondence: Dr A. J. Dwork, 1051 Riverside Dr.,
Unit 62, New York, NY 10032, USA.
Tel. : 212-543-5563 Fax : 212-543-6017
E-mail : [email protected]
International Journal of Neuropsychopharmacology (2007), 10, 513–536. Copyright f 2007 CINPdoi:10.1017/S1461145707007638
SPECIAL SECTION
CINP

to dementia. In addition, normal or pathological vari-
ation in white matter that is not related to schizo-
phrenia could have cognitive consequences that are
exaggerated in individuals with schizophrenia. One
such consequence might be a lessening of cognitive
reserve, or the capacity to resist the cognitive manifes-
tations of normal ageing processes or age-associated
degenerative changes (Stern, 2006). The concept of
cognitive reserve comprises both anatomical capacity
(e.g. size of brain or neurons) and the ability to recruit
neuronal activity. Both types of reserve are probably
impaired in schizophrenia. Pre-existing impairment
may predispose to the development of schizophrenia
and may worsen its prognosis, and schizophrenia may
progressively impair cognitive reserve (Barnett et al.,
2006). Myelin integrity correlates with cognitive pro-
cessing speed (Liston et al., 2006), and might thus
contribute to cognitive reserve. On the other hand,
factors conventionally associated with cognitive
reserve, such as education or physical activity, ap-
pear to protect against loss of processing speed in
conditions associated with myelin loss, e.g. ageing
(Dik et al., 2003), infection with human immuno-
deficiency virus (Stern et al., 1996), or white-matter
hyperintensities on magnetic resonance imaging
(MRI) (Nebes et al., 2006). It thus seems likely that
early-life myelination is relatively resistant to ageing
and contributes to cognitive reserve, while the various
components of cognitive reserve impart resistance to
the cognitive effects of pathological or age-related loss
of myelin.
White-matter abnormalities in schizophrenia
The majority of studies implicating white-matter
alterations in schizophrenia employ in-vivo imaging.
T2-weighted signal intensity is largely determined
by water content, and therefore serves as a sensitive
indicator of demyelination, oedema, or inflammation.
Early studies, looking for regions of T2 hyperintensity,
gave mixed results, but white-matter hyperintensities
were certainly not consistently present (Brown et al.,
1992, 1995; Hulshoff Pol et al., 2000; Keshavan et al.,
1996; Lane et al., 1996; Rivkin et al., 2000; Symonds
et al., 1997). Brown et al. (1995) concluded that T2 hy-
perintensities in schizophrenia were associated with
stroke or hypertension and were no more common
than in non-psychiatric subjects. Thus, the abnormali-
ties uncovered by more sensitive MRI techniques must
be considered relatively subtle, which is consistent
with the failure of conventional neuropathological
studies of schizophrenia to note any excess of white-
matter lesions.
Volumetric studies of white matter (reviewed in
Walterfang et al., 2006) give conflicting results, par-
ticularly with regard to the size and shape of the cor-
pus callosum. The more consistent findings include
reduction in volume of the left uncinate fasciculus and
the anterior limb of the internal capsule. These struc-
tures contain fibres connecting the thalamus with
frontal and cingulate cortices, and frontal cortex with
rostral temporal cortex, respectively. In general, simi-
lar decrements in schizophrenia are reported for grey-
matter volumes and white-matter volumes, suggest-
ing that the latter may simply reflect the former.
However, this interpretation should be viewed with
caution; with the possible exception of the medial
dorsal nucleus of the thalamus (Byne et al., 2002;
Dorph-Petersen et al., 2004; Pakkenberg, 1992; Young
et al., 2000) there is little replicated evidence for neu-
ronal loss in schizophrenia (reviewed in Dwork, 1997;
Harrison, 1999; Heckers, 1997). Therefore, white-
matter deficits probably cannot be simply explained
by deficits in neurons giving rise to axons.
Magnetic resonance spectroscopy (MRS) exploits
the magnetic resonance spectra of specific organic
compounds to localize and quantify these compounds
in vivo. Commonly analysed compounds include
N-acetylaspartate (NAA), which is believed to be un-
ique to neurons and is generally accepted as a measure
of density or viability of neurons and axons. NAA is
quantified either in absolute terms or relative to cre-
atine, a relatively stable component of neurons and
glia (Hammen et al., 2003). MRS has demonstrated
decreased NAA, increased creatine or a decreased
NAA/creatine ratio in right prefrontal white matter
(Choe et al., 1994), white matter in general (Lim et al.,
1998), and parietal white matter (Auer et al., 2001) in
schizophrenia. Bartha et al. (1999), finding no change
in NAA signal in a medial temporal region of interest
designed to include predominantly grey matter, con-
cluded that previously reported decreases in NAA
probably reflected a loss in white matter. Reductions
of NAA in prefrontal white matter reported by Steel
et al. (2001) were not statistically significant.
The studies pointing most frequently to abnormali-
ties of white matter in schizophrenia employed dif-
fusion tensor imaging (DTI). DTI applies several
magnetic gradients in various orientations in order to
obtain a set of diffusion-weighted images, from which
is derived a tensor corresponding to the apparent rate
of diffusion of water in three orthogonal dimensions
within each voxel (typically several mm in each di-
mension). For each voxel, several indices are obtained
from this procedure, including (a) trace or mean dif-
fusivity (D=trace/3), measures of average diffusivity,
514 A. J. Dwork et al.

independent of direction, (b) fractional anisotropy
(FA) or relative anisotropy (RA), measures of the de-
gree of directionality of apparent diffusion (these em-
ploy different formulas, but both range theoretically
from 0 with equal diffusion in all directions to 1 with
diffusion confined to one direction), (c) lk, or axial
diffusivity, the apparent diffusion constant in the di-
rection of maximal apparent diffusivity, and (d) l?, or
radial diffusivity, the mean rate of diffusion in the
perpendicular dimensions (Basser and Jones, 2002;
Basser and Pierpaoli, 1996; Kingsley and Monahan,
2005). Comparison of diffusion tensors in adjacent
voxels is employed for fibre tract tracing (diffusion
tractography, e.g. Pierpaoli et al., 2001) who point out
the danger of artefactual construction of non-existent
tracts) or measures of intervoxel coherence, the aver-
age angle between the vector of maximum diffusivity
in a given voxel and those of its neighbours (Pierpaoli
and Basser, 1996).
In complex systems, such as cerebral white matter,
the arrangement of fibres must obviously contribute to
anisotropy (see, e.g. Pierpaoli et al., 2001). In tissues
such as peripheral or optic nerve, where most of the
fibres are oriented in parallel, lk is the apparent dif-
fusion constant in the direction of fibre orientation;
however, even in isolated preparations of such tissue,
the anatomical factors contributing to the tensor are
complex (reviewed in Beaulieu, 2002). In healthy
nerve, lk is nearly equal to the diffusion of free water,
while l? is several times smaller. Myelin probably
contributes to anisotropy by reducing l?, but it is not
required for anisotropic diffusion, which is readily
observed in normally unmyelinated nerves or in gen-
etically myelin-deficient rat spinal cords. The latter
show increased lk and somewhat greater increases in
l?. With Wallerian degeneration, in which axons and
myelin disintegrate distal to an injury of peripheral
nerve, lk decreases and l? increases ; anisotropy
eventually returns to normal with regeneration, al-
though regenerated peripheral nerves typically have
thinner myelin sheaths than previously. While atten-
tion is frequently focused on diffusion of water within
axons, diffusion in the extracellular space should also
have important effects on anisotropy.
In pathological conditions of the nervous system,
axonal damage, myelin loss, and reactive processes
tend to occur in unison. Thus, it is very difficult to
determine, even with histological examination and
DTI of the same piece of tissue, how an individual
component of tissue damage affects various indices of
diffusion. One exception is the shiverer mouse, in
which a mutation of the gene for myelin basic protein
(MBP) results in myelin sheaths that are thin, loosely
wrapped, or absent, without evidence of axonal injury
or tissue reaction. In major cerebral white-matter tracts
of live shiverer mice, Song et al. (2002) found decreases
in RA (y20%), with lk unchanged, but l? increased
by y20%. Thus, a relatively pure deficit in myelin re-
sulted in a modest increase in radial diffusivity, with
no effect on axial diffusivity and preservation of a
considerable degree of anisotropy. This is essentially
consistent with the conclusion of Beaulieu (2002) that
the axolemma forms the principal boundary to radial
diffusion, with the myelin sheath playing a mod-
ulatory role. It is also notable that, while axonal pack-
ing density was unchanged in the shiverer mice of
Song et al. (2002), the illustrations show an increase in
water-accessible space between adjacent myelin
sheaths and between layers of individual myelin
sheaths. One difficulty in interpreting such studies is
that tissue can change during fixation and histological
preparation. In white matter of fixed shiverer brains,
soaked in the contrast-enhancing agent gadoteridol, lkand l? were both increased, with a 10–20% decrease
in FA and a slightly larger increase trace, compared
with wild-type mice (Tyszka et al., 2006).
In more complex pathological models [retinal
ischaemia (Song et al., 2003; Sun et al., 2006a),
b-amyloid overexpression (Song et al., 2004; Sun et al.,
2005), experimental allergic encephalomyelitis (Kim
et al., 2006) cuprizone toxicity (Song et al., 2005; Sun
et al., 2006b)], increases in l? and decreases in lk,
were interpreted as damage to myelin and axons,
respectively, but as noted by the authors, other inter-
pretations are plausible.
Another refinement of MRI is the measurement of
magnetization transfer (MT), simplistically, the sup-
pression of the T2 signal of water by the transfer of
proton spin from adjacent, immobilized macro-
molecules. Presumably, loss of macromolecules, such
as myelin lipids, results in decreased MT, which is
distinguishable from a simple increase in water con-
tent (Grossman et al., 1994).
Of at least 39 DTI or MT studies of schizophrenia
(Table 1), only four were entirely negative (Begre et al.,
2003; Foong et al., 2002; Price et al., 2005; Wang et al.,
2003) ; only one of these four (Foong et al., 2002) sur-
veyed all regions of cerebral white matter, and that
study imposed stringent criteria for statistical signifi-
cance. The remaining studies, while all showing de-
creased FA somewhere in the white matter, are not
entirely consistent about where such loss is located. In
prefrontal white matter, the results suggest a pre-
dominance of ventral abnormalities (Ardekani et al.,
2003; Buchsbaum et al., 1998, 2006a,b; Hao et al., 2003;
Kumra et al., 2004). Some studies found generalized
White matter and cognition 515
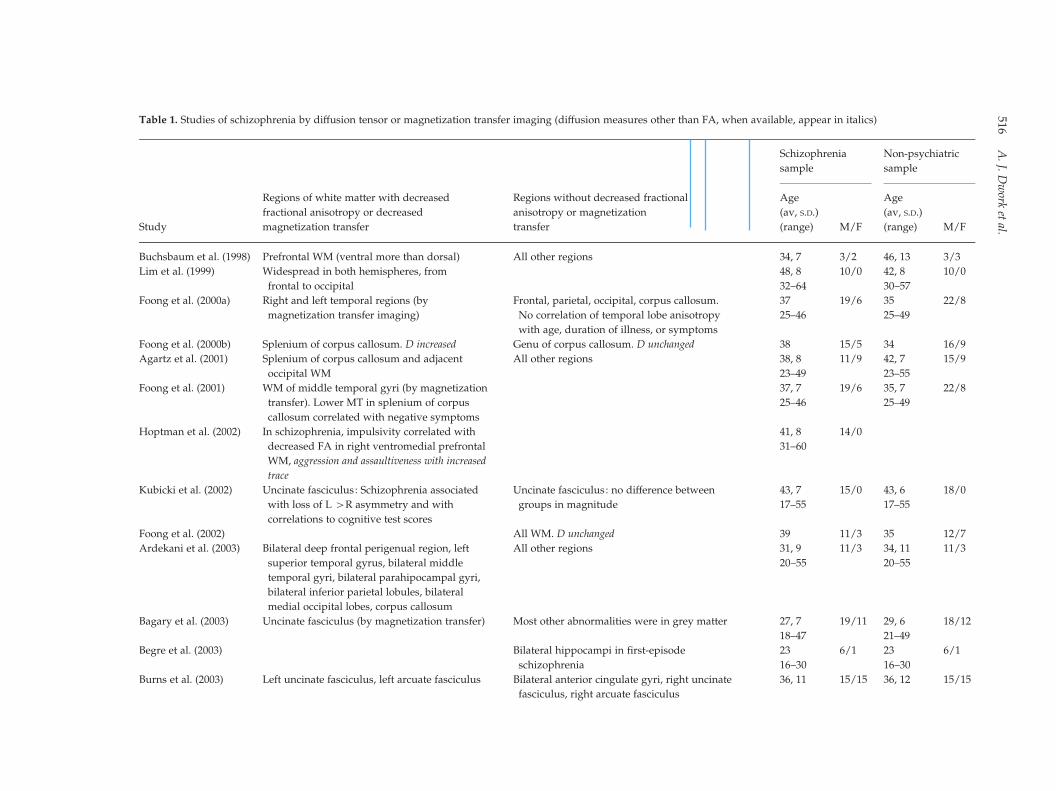
Table 1. Studies of schizophrenia by diffusion tensor or magnetization transfer imaging (diffusion measures other than FA, when available, appear in italics)
Study
Regions of white matter with decreased
fractional anisotropy or decreased
magnetization transfer
Regions without decreased fractional
anisotropy or magnetization
transfer
Schizophrenia
sample
Non-psychiatric
sample
Age
(av, S.D.)
(range) M/F
Age
(av, S.D.)
(range) M/F
Buchsbaum et al. (1998) Prefrontal WM (ventral more than dorsal) All other regions 34, 7 3/2 46, 13 3/3
Lim et al. (1999) Widespread in both hemispheres, from
frontal to occipital
48, 8 10/0 42, 8 10/0
32–64 30–57
Foong et al. (2000a) Right and left temporal regions (by
magnetization transfer imaging)
Frontal, parietal, occipital, corpus callosum. 37 19/6 35 22/8
25–46 25–49No correlation of temporal lobe anisotropy
with age, duration of illness, or symptoms
Foong et al. (2000b) Splenium of corpus callosum. D increased Genu of corpus callosum. D unchanged 38 15/5 34 16/9
Agartz et al. (2001) Splenium of corpus callosum and adjacent
occipital WM
All other regions 38, 8 11/9 42, 7 15/9
23–49 23–55
Foong et al. (2001) WM of middle temporal gyri (by magnetization
transfer). Lower MT in splenium of corpus
callosum correlated with negative symptoms
37, 7 19/6 35, 7 22/8
25–46 25–49
Hoptman et al. (2002) In schizophrenia, impulsivity correlated with
decreased FA in right ventromedial prefrontal
WM, aggression and assaultiveness with increased
trace
41, 8 14/0
31–60
Kubicki et al. (2002) Uncinate fasciculus: Schizophrenia associated
with loss of L >R asymmetry and with
correlations to cognitive test scores
Uncinate fasciculus: no difference between
groups in magnitude
43, 7 15/0 43, 6 18/0
17–55 17–55
Foong et al. (2002) All WM. D unchanged 39 11/3 35 12/7
Ardekani et al. (2003) Bilateral deep frontal perigenual region, left
superior temporal gyrus, bilateral middle
temporal gyri, bilateral parahipocampal gyri,
bilateral inferior parietal lobules, bilateral
medial occipital lobes, corpus callosum
All other regions 31, 9 11/3 34, 11 11/3
20–55 20–55
Bagary et al. (2003) Uncinate fasciculus (by magnetization transfer) Most other abnormalities were in grey matter 27, 7 19/11 29, 6 18/12
18–47 21–49
Begre et al. (2003) Bilateral hippocampi in first-episode
schizophrenia
23 6/1 23 6/1
16–30 16–30
Burns et al. (2003) Left uncinate fasciculus, left arcuate fasciculus Bilateral anterior cingulate gyri, right uncinate
fasciculus, right arcuate fasciculus
36, 11 15/15 36, 12 15/15
516A.J.
Dwork
etal.
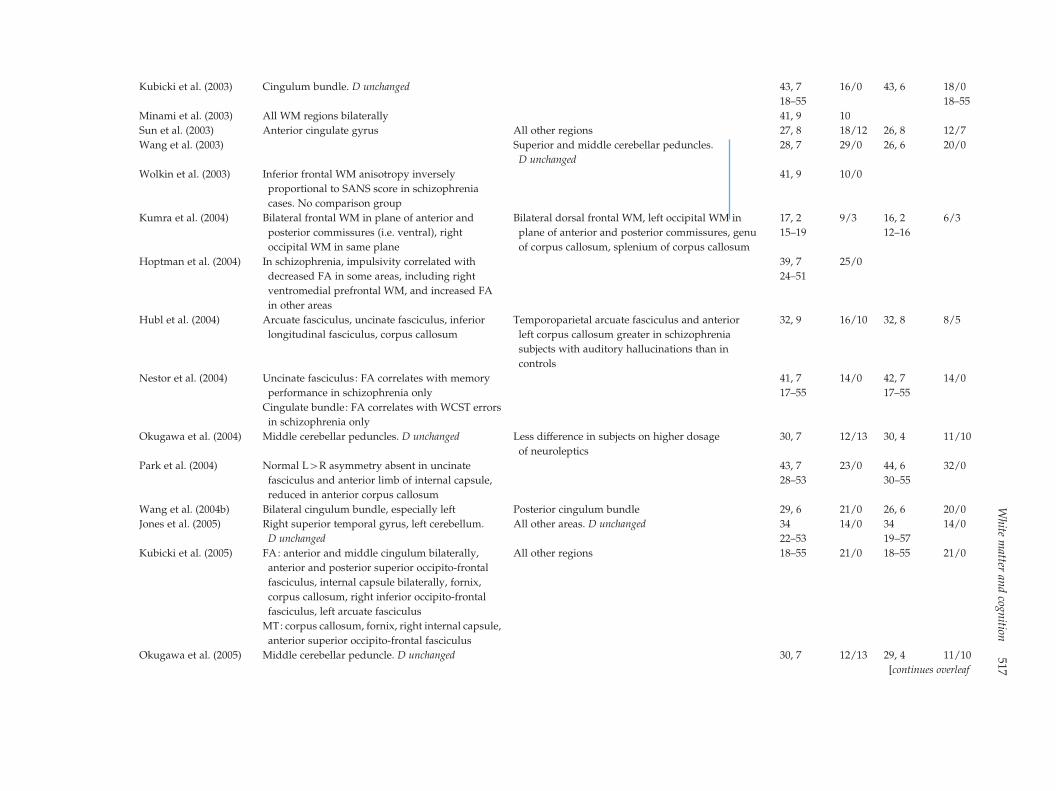
Kubicki et al. (2003) Cingulum bundle. D unchanged 43, 7 16/0 43, 6 18/0
18–55 18–55
Minami et al. (2003) All WM regions bilaterally 41, 9 10
Sun et al. (2003) Anterior cingulate gyrus All other regions 27, 8 18/12 26, 8 12/7
Wang et al. (2003) Superior and middle cerebellar peduncles.
D unchanged
28, 7 29/0 26, 6 20/0
Wolkin et al. (2003) Inferior frontal WM anisotropy inversely
proportional to SANS score in schizophrenia
cases. No comparison group
41, 9 10/0
Kumra et al. (2004) Bilateral frontal WM in plane of anterior and
posterior commissures (i.e. ventral), right
occipital WM in same plane
Bilateral dorsal frontal WM, left occipital WM in
plane of anterior and posterior commissures, genu
of corpus callosum, splenium of corpus callosum
17, 2 9/3 16, 2 6/3
15–19 12–16
Hoptman et al. (2004) In schizophrenia, impulsivity correlated with
decreased FA in some areas, including right
ventromedial prefrontal WM, and increased FA
in other areas
39, 7 25/0
24–51
Hubl et al. (2004) Arcuate fasciculus, uncinate fasciculus, inferior
longitudinal fasciculus, corpus callosum
Temporoparietal arcuate fasciculus and anterior
left corpus callosum greater in schizophrenia
subjects with auditory hallucinations than in
controls
32, 9 16/10 32, 8 8/5
Nestor et al. (2004) Uncinate fasciculus: FA correlates with memory
performance in schizophrenia only
41, 7 14/0 42, 7 14/0
17–55 17–55
Cingulate bundle: FA correlates with WCST errors
in schizophrenia only
Okugawa et al. (2004) Middle cerebellar peduncles. D unchanged Less difference in subjects on higher dosage
of neuroleptics
30, 7 12/13 30, 4 11/10
Park et al. (2004) Normal L>R asymmetry absent in uncinate
fasciculus and anterior limb of internal capsule,
reduced in anterior corpus callosum
43, 7 23/0 44, 6 32/0
28–53 30–55
Wang et al. (2004b) Bilateral cingulum bundle, especially left Posterior cingulum bundle 29, 6 21/0 26, 6 20/0
Jones et al. (2005) Right superior temporal gyrus, left cerebellum.
D unchanged
All other areas. D unchanged 34 14/0 34 14/0
22–53 19–57
Kubicki et al. (2005) FA: anterior and middle cingulum bilaterally,
anterior and posterior superior occipito-frontal
fasciculus, internal capsule bilaterally, fornix,
corpus callosum, right inferior occipito-frontal
fasciculus, left arcuate fasciculus
All other regions 21/0 21/018–55 18–55
MT: corpus callosum, fornix, right internal capsule,
anterior superior occipito-frontal fasciculus
Okugawa et al. (2005) Middle cerebellar peduncle. D unchanged 30, 7 12/13 29, 4 11/10
[continues overleaf
White
matter
andcogn
ition517
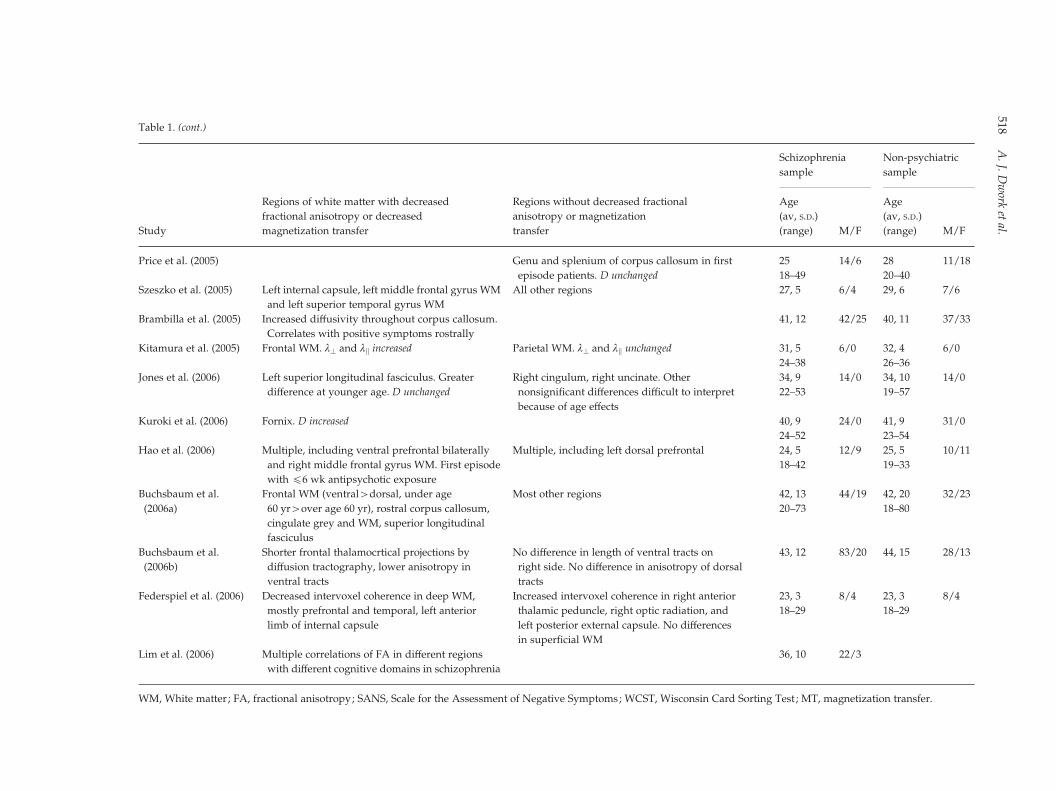
Table 1. (cont.)
Study
Regions of white matter with decreased
fractional anisotropy or decreased
magnetization transfer
Regions without decreased fractional
anisotropy or magnetization
transfer
Schizophrenia
sample
Non-psychiatric
sample
Age
(av, S.D.)
(range) M/F
Age
(av, S.D.)
(range) M/F
Price et al. (2005) Genu and splenium of corpus callosum in first
episode patients. D unchanged
25 14/6 28 11/18
18–49 20–40
Szeszko et al. (2005) Left internal capsule, left middle frontal gyrus WM
and left superior temporal gyrus WM
All other regions 27, 5 6/4 29, 6 7/6
Brambilla et al. (2005) Increased diffusivity throughout corpus callosum. 41, 12 42/25 40, 11 37/33
Correlates with positive symptoms rostrally
Kitamura et al. (2005) Frontal WM. l? and lk increased Parietal WM. l? and lk unchanged 31, 5 6/0 32, 4 6/0
24–38 26–36
Jones et al. (2006) Left superior longitudinal fasciculus. Greater
difference at younger age. D unchanged
Right cingulum, right uncinate. Other
nonsignificant differences difficult to interpret
because of age effects
34, 9 14/0 34, 10 14/0
22–53 19–57
Kuroki et al. (2006) Fornix. D increased 40, 9 24/0 41, 9 31/0
24–52 23–54
Hao et al. (2006) Multiple, including ventral prefrontal bilaterally
and right middle frontal gyrus WM. First episode
with f6 wk antipsychotic exposure
Multiple, including left dorsal prefrontal 24, 5 12/9 25, 5 10/11
18–42 19–33
Buchsbaum et al.
(2006a)
Frontal WM (ventral>dorsal, under age
60 yr>over age 60 yr), rostral corpus callosum,
cingulate grey and WM, superior longitudinal
fasciculus
Most other regions 42, 13 44/19 42, 20 32/23
20–73 18–80
Buchsbaum et al.
(2006b)
Shorter frontal thalamocrtical projections by
diffusion tractography, lower anisotropy in
ventral tracts
No difference in length of ventral tracts on
right side. No difference in anisotropy of dorsal
tracts
43, 12 83/20 44, 15 28/13
Federspiel et al. (2006) Decreased intervoxel coherence in deep WM,
mostly prefrontal and temporal, left anterior
limb of internal capsule
Increased intervoxel coherence in right anterior
thalamic peduncle, right optic radiation, and
left posterior external capsule. No differences
in superficial WM
23, 3 8/4 23, 3 8/4
18–29 18–29
Lim et al. (2006) Multiple correlations of FA in different regions
with different cognitive domains in schizophrenia
36, 10 22/3
WM, White matter; FA, fractional anisotropy; SANS, Scale for the Assessment of Negative Symptoms; WCST, Wisconsin Card Sorting Test ; MT, magnetization transfer.
518A.J.
Dwork
etal.

decreases that included dorsal prefrontal white matter
(Federspiel et al., 2006; Lim et al., 1999; Minami et al.,
2003), underlying a cortical region implicated in func-
tional imaging studies of cognition in schizophrenia
(Callicott et al., 2003; Weinberger et al., 1986). None of
the DTI studies of schizophrenia involved elderly
subjects, except for the large study by Buchsbaum
et al. (2006a), who reported that decreases in frontal
anisotropy were enhanced when subjects aged>60 yr
were removed from the sample. According to
Jones et al. (2006), by middle age, differences in FA
between schizophrenia and comparison subjects start
to become less evident with age.
As noted above, decreases in FA may be the result
of alterations in myelin. A subtle axonopathy, e.g. as
part of a slow dying-back process, might be consistent
with the reported loss of anisotropy and with the loss
of NAA signal in white matter, with relative preser-
vation of normal histology (see below). For example, it
appears that in amyotrophic lateral sclerosis, DTI may
be more sensitive than histological examination for
detecting abnormalities in the suprabulbar portions of
the corticospinal tracts (Toosy et al., 2003), which
usually appear normal on myelin stains.
Although a number of studies describe decreased
anisotropy in the uncinate fasciculus, or in ventral
frontal white matter, where it terminates, Highley et al.
(2002) report normal size and axon number in the un-
cinate fasciculus at autopsy. The methodology did not
allow myelin changes to be ruled out, but the discre-
pancy between normal axonal number and decreased
anisotropy could also be explained by disorganization
of the fasciculus. This possibility is supported by
tractography evidence of shorter frontal fibre bundles,
which could indicate dispersion of fibre tracts before
they reach their targets (Buchsbaum et al., 2006b).
Resolution of these issues will require stereological
studies of axonal orientation and, in plastic-embedded
tissue, of axonal diameter and thickness of myelin
sheaths.
Few of the DTI studies report diffusion measures
other than FA (Table 1). In four studies, decreased FA
was associated with increased D ; in five studies it was
not. Only one study reports l? and lk, which were
both increased where FA was decreased, leading the
authors to attribute the changes to expansion of the
extracellular space in frontal white matter (Kitamura
et al., 2005).
Only a few post-mortem studies have specifically
evaluated white matter in schizophrenia (Table 2).
Hof et al. (2002, 2003), in small groups of aged,
chronically institutionalized subjects, found, using
Cresyl Violet stains, a 15–20% decrease in density of
oligodendrocytes in the white matter underlying
Brodmann’s area (BA) 9, and a 28% decrease when
oligodendrocytes were identified by immunoreac-
tivity for 2k,3k-cyclic nucleotide-3k-phosphodiesterase(CNPase). However, in the younger sample from the
Stanley Consortium, Uranova et al. (2004) found an
effect of schizophrenia on density of oligodendroglia
in this region only in the cortex, not in the white
matter. Marner and Pakkenberg (2003), comparing
eight schizophrenia subjects and nine controls with a
mean age of 60 yr, found no difference in the total
length of myelinated axons in the prefrontal white
matter, nor in the diameters of these axons. In a large
series of elderly, chronically institutionalized schizo-
phrenia subjects, we performed detailed histological
evaluations of Verhoeff myelin stains in the white
matter of the dorsal prefrontal region and found no
difference from non-psychiatric subjects (Dwork et al.,
2005). Falkai et al. (1999) found no evidence for astro-
cytosis in the white matter of the premotor cortex or
the subventricular zone of the third ventricle, which
suggests that, at least in these regions, a demyelinating
process is unlikely.
Highley et al. (2002) reported normal size and axon
number in an autopsy study of the uncinate fasciculus
in elderly subjects with schizophrenia. In the same
sample, the anterior commissure was of normal size,
but there was a 31% decrease in fibre density in the
female schizophrenia subjects compared with con-
trols, while there was no effect of diagnosis among the
males (Highley et al., 1999b). In the corpus callosum,
fibre density was decreased among females with
schizophrenia but not among males (Highley et al.,
1999a). In the fornix, there was a 32% increase in fibre
density on the left side in males with schizophrenia
(Chance et al., 1999). These studies found a loss of
fibres with age in most subregions of the corpus cal-
losum, but not in the uncinate fasciculus, anterior
commissure, or fornix. In other studies of the corpus
callosum of middle-aged (Casanova et al., 1989) and
elderly (Nasrallah et al., 1983) subjects, schizophrenia
was not associated with a loss of fibre density.
Ultrastructural studies of white matter in schizo-
phrenia are lacking, but there are several reports on
oligodendroglial and myelin morphology in frontal
cortical area BA 10. An electron microscopic study
of BA 10 biopsies found lipofuscin deposits in oligo-
dendroglial cytoplasm and increased numbers of
electron-dense granules in neuronal cytoplasm, axon–
oligodendrocyte interfaces, and myelin sheaths
(Miyakawa et al., 1972). An electron microscopic study
of autopsy material (Uranova et al., 2001) purported to
show subtle abnormalities in oligodendroglia and
White matter and cognition 519
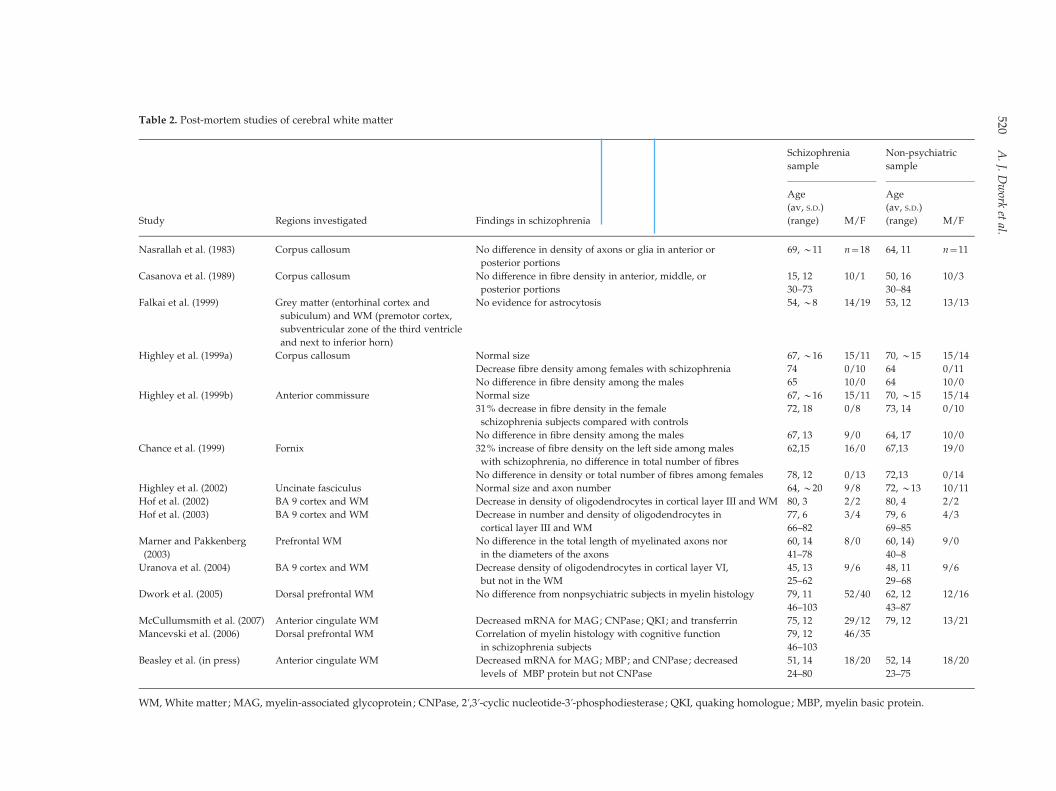
Table 2. Post-mortem studies of cerebral white matter
Study Regions investigated Findings in schizophrenia
Schizophrenia
sample
Non-psychiatric
sample
Age
(av, S.D.)
(range) M/F
Age
(av, S.D.)
(range) M/F
Nasrallah et al. (1983) Corpus callosum No difference in density of axons or glia in anterior or
posterior portions
69, y11 n=18 64, 11 n=11
Casanova et al. (1989) Corpus callosum No difference in fibre density in anterior, middle, or
posterior portions
15, 12 10/1 50, 16 10/3
30–73 30–84
Falkai et al. (1999) Grey matter (entorhinal cortex and
subiculum) and WM (premotor cortex,
subventricular zone of the third ventricle
and next to inferior horn)
No evidence for astrocytosis 54, y8 14/19 53, 12 13/13
Highley et al. (1999a) Corpus callosum Normal size 67, y16 15/11 70, y15 15/14
Decrease fibre density among females with schizophrenia 74 0/10 64 0/11
No difference in fibre density among the males 65 10/0 64 10/0
Highley et al. (1999b) Anterior commissure Normal size 67, y16 15/11 70, y15 15/14
31% decrease in fibre density in the female
schizophrenia subjects compared with controls
72, 18 0/8 73, 14 0/10
No difference in fibre density among the males 67, 13 9/0 64, 17 10/0
Chance et al. (1999) Fornix 32% increase of fibre density on the left side among males
with schizophrenia, no difference in total number of fibres
62,15 16/0 67,13 19/0
No difference in density or total number of fibres among females 78, 12 0/13 72,13 0/14
Highley et al. (2002) Uncinate fasciculus Normal size and axon number 64, y20 9/8 72, y13 10/11
Hof et al. (2002) BA 9 cortex and WM Decrease in density of oligodendrocytes in cortical layer III and WM 80, 3 2/2 80, 4 2/2
Hof et al. (2003) BA 9 cortex and WM Decrease in number and density of oligodendrocytes in
cortical layer III and WM
77, 6 3/4 79, 6 4/3
66–82 69–85
Marner and Pakkenberg
(2003)
Prefrontal WM No difference in the total length of myelinated axons nor
in the diameters of the axons
60, 14 8/0 60, 14) 9/0
41–78 40–8
Uranova et al. (2004) BA 9 cortex and WM Decrease density of oligodendrocytes in cortical layer VI,
but not in the WM
45, 13 9/6 48, 11 9/6
25–62 29–68
Dwork et al. (2005) Dorsal prefrontal WM No difference from nonpsychiatric subjects in myelin histology 79, 11 52/40 62, 12 12/16
46–103 43–87
McCullumsmith et al. (2007) Anterior cingulate WM Decreased mRNA for MAG; CNPase; QKI; and transferrin 75, 12 29/12 79, 12 13/21
Mancevski et al. (2006) Dorsal prefrontal WM Correlation of myelin histology with cognitive function
in schizophrenia subjects
79, 12 46/35
46–103
Beasley et al. (in press) Anterior cingulate WM Decreased mRNA for MAG; MBP; and CNPase; decreased
levels of MBP protein but not CNPase
51, 14 18/20 52, 14 18/20
24–80 23–75
WM, White matter; MAG, myelin-associated glycoprotein; CNPase, 2k,3k-cyclic nucleotide-3k-phosphodiesterase; QKI, quaking homologue; MBP, myelin basic protein.
520A.J.
Dwork
etal.

myelin sheaths of cortical layer VI of BA 10 and the
caudate nucleus in schizophrenia. These included de-
creased cross-sectional areas of nuclei and decreased
fraction of cytoplasmic volume occupied by mito-
chondria, but not the electron-dense granules or lipo-
fuscin described in the biopsies (Miyakawa et al.,
1972). The non-specific findings of these studies are
somewhat compromised by a failure to note whether
the examiners were masked to diagnosis. None of
these studies described evidence of demyelination,
and none measured the thicknesses of myelin sheaths,
which could be reduced in the case of remyelination or
altered by aberrant regulation of myelination (e.g. as
hypothesized in the case of an abnormality or insuf-
ficiency of neuregulin 1; see below).
In the only reported study of mRNA in white matter
(McCullumsmith et al., 2007), expression of myelin-
associated glycoprotein [MAG; transmembrane glyco-
protein of oligodendrocytes, largely localized to the
periaxonal collar and believed to be involved in signal
transduction between the axon and myelin sheath
(Schachner and Bartsch, 2000)], CNPase (cytoplasmic
protein of oligodendrocytes, with unknown function),
quaking homologue [QKI, regulates alternative spli-
cing of several myelin-related genes (Aberg et al.,
2006a; Wu et al., 2002)], and transferrin (iron transport
protein expressed by oligodendrocytes) were down-
regulated in the anterior cingulate white matter of 41
elderly, medicated and unmedicated schizophrenia
subjects, compared with 34 elderly non-psychiatric
subjects. Similarly, in a small, middle-aged sample, we
found decreased mRNA for MAG, MBP, and CNPase,
and decreased levels of MBP protein, but not CNPase,
in anterior cingulate white matter (Beasley et al., in
press).
Studies of myelin- and oligodendrocyte-related
proteins and mRNA levels in grey matter are more
numerous (Table 3). Flynn et al. (2003) found sub-
stantial decreases in immunoreactivity for CNPase
and MAG in prefrontal cortex. Hakak et al. (2001)
demonstrated down-regulation of mRNA for CNPase,
MAG, myelin and lymphocyte protein, gelsolin, and
transferrin, all of which are expressed predominantly
in oligodendrocytes, and for ErbB3, a receptor for
neuregulin (see below) in BA 46 of elderly schizo-
phrenia subjects. Similar results were obtained in BA 9
in a younger sample (Tkachev et al., 2003). Aberg et al.
(2006a) demonstrated down-regulation of mRNA for
proteolipid protein 1, MAG, and transferrin in frontal
cortex, and proposed that this was regulated by ex-
pression of the QKI 7-kb splice variant, which is also
down-regulated (Aberg et al., 2006b). Dracheva et al.
(2006) found, in an elderly sample, that mRNA for
MAG, CNPase, SRY (sex determining region Y)-box 10
(SOX10; oligodendroglial transcription regulator),
claudin11 (CLDN11, essential for the formation of
myelin tight junctions), and peripheral myelin protein
22 (PMP22) was reduced in the hippocampus and an-
terior cingulate cortex but not in the putamen of pa-
tients with schizophrenia, while transcripts for MBP
and myelin-associated oligodendrocyte basic protein
(MOBP), both involved in compaction of the myelin
sheath, were unaffected. Expression of CNPase, the
only protein examined in this study, was decreased in
the hippocampus but not in the putamen.
The aforementioned abnormalities of myelin-
related proteins and mRNA, although studied in grey
matter, have undoubtedly contributed to assumptions
or speculation that the decreases in FA and MT, ob-
served in MRI studies, are the result of abnormalities
in myelin. This line of thought is further supported by
genetic studies.Myelination-related geneswhose poly-
morphisms have been associated with schizophrenia
include Nrg1 (Harrison and Weinberger, 2005), which
is involved (at least in peripheral nerve) in regulating
the extent of wrapping of the myelin sheath around
the axon (Michailov et al., 2004). Although previous
reports involved non-coding regions, association of
psychosis with a missense mutation of the Nrg1 gene
was recently discovered (Walss-Bass et al., 2006).
Studies of white matter in these individuals have not
yet been reported. A genetic association of schizo-
phrenia was found with an exonic polymorphism of
CNPase, an allele that is less expressed than its
counterpart allele in the cerebral cortex of normal
heterozygous individuals (Peirce et al., 2006). Schizo-
phrenia has also been associated with a specific hap-
lotype in the region of the QKI gene and, in a different
population of schizophrenia patients, with altered pat-
terns of expression of QKI splice variants. Expression
of all QKI splice variants was elevated in subjects
treated with typical neuroleptics (Aberg et al., 2006b).
While the biology of oligodendrocytes in grey mat-
ter appears to be similar to that of oligodendrocytes in
white matter, there may be differences that are poorly
understood; for example, Graeber et al. (2002) refer to
the satellite oligodendrocytes that surround neurons
as an ‘enigma’ (p. 150). Thus, while white-matter
myelin may be abnormal in schizophrenia, direct evi-
dence for this is very limited, and such an abnormality
may or may not underlie the loss of anisotropy seen
with DTI. It should also be remembered that axonal
abnormalities, such as the slow dying-back of amyo-
trophic lateral sclerosis (motor neuron disease)
(Coleman and Ribchester, 2004), are associated with
loss of myelin, and demyelinating diseases, such as
White matter and cognition 521
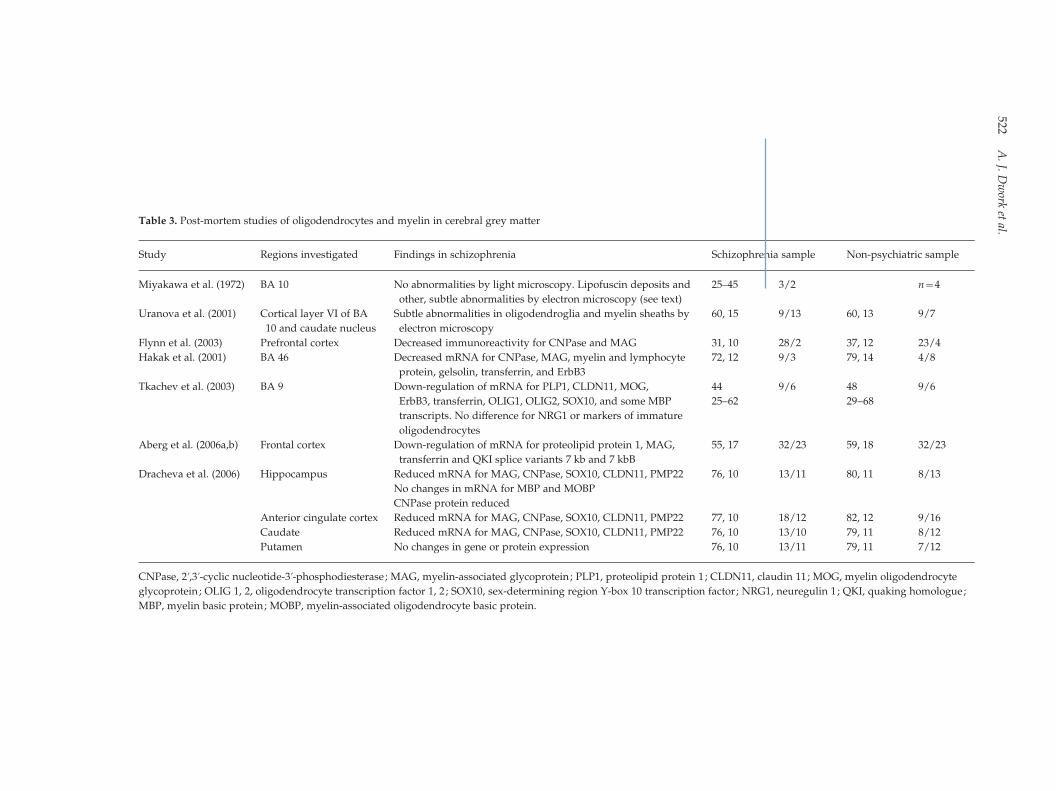
Table 3. Post-mortem studies of oligodendrocytes and myelin in cerebral grey matter
Study Regions investigated Findings in schizophrenia Schizophrenia sample Non-psychiatric sample
Miyakawa et al. (1972) BA 10 No abnormalities by light microscopy. Lipofuscin deposits and
other, subtle abnormalities by electron microscopy (see text)
25–45 3/2 n=4
Uranova et al. (2001) Cortical layer VI of BA
10 and caudate nucleus
Subtle abnormalities in oligodendroglia and myelin sheaths by
electron microscopy
60, 15 9/13 60, 13 9/7
Flynn et al. (2003) Prefrontal cortex Decreased immunoreactivity for CNPase and MAG 31, 10 28/2 37, 12 23/4
Hakak et al. (2001) BA 46 Decreased mRNA for CNPase, MAG, myelin and lymphocyte
protein, gelsolin, transferrin, and ErbB3
72, 12 9/3 79, 14 4/8
Tkachev et al. (2003) BA 9 Down-regulation of mRNA for PLP1, CLDN11, MOG,
ErbB3, transferrin, OLIG1, OLIG2, SOX10, and some MBP
transcripts. No difference for NRG1 or markers of immature
oligodendrocytes
44 9/6 48 9/6
25–62 29–68
Aberg et al. (2006a,b) Frontal cortex Down-regulation of mRNA for proteolipid protein 1, MAG,
transferrin and QKI splice variants 7 kb and 7 kbB
55, 17 32/23 59, 18 32/23
Dracheva et al. (2006) Hippocampus Reduced mRNA for MAG, CNPase, SOX10, CLDN11, PMP22 76, 10 13/11 80, 11 8/13
No changes in mRNA for MBP and MOBP
CNPase protein reduced
Anterior cingulate cortex Reduced mRNA for MAG, CNPase, SOX10, CLDN11, PMP22 77, 10 18/12 82, 12 9/16
Caudate Reduced mRNA for MAG, CNPase, SOX10, CLDN11, PMP22 76, 10 13/10 79, 11 8/12
Putamen No changes in gene or protein expression 76, 10 13/11 79, 11 7/12
CNPase, 2k,3k-cyclic nucleotide-3k-phosphodiesterase; MAG, myelin-associated glycoprotein; PLP1, proteolipid protein 1; CLDN11, claudin 11; MOG, myelin oligodendrocyte
glycoprotein; OLIG 1, 2, oligodendrocyte transcription factor 1, 2; SOX10, sex-determining region Y-box 10 transcription factor; NRG1, neuregulin 1; QKI, quaking homologue;
MBP, myelin basic protein; MOBP, myelin-associated oligodendrocyte basic protein.
522A.J.
Dwork
etal.

multiple sclerosis, result in axonal damage and loss
(Craner et al., 2004). It may ultimately prove diffi-
cult to determine whether myelin abnormalities in
schizophrenia are primary or secondary to neuronal
abnormalities. Furthermore, as we discuss below, re-
myelination can result in increased density of oligo-
dendrocytes and related mRNA, so lower levels of
mRNA for myelin-related proteins in schizophrenia
could represent either an abnormal increase in myelin
stability, or absence of repair of normally occurring
(e.g. age-related) demyelination.
Ageing, myelin, and ApoE genotype
Myelination of the human brain continues into adult-
hood (Yakovlev and Lecours, 1967), and by late middle
age there begins a process of demyelination (Marner
et al., 2003; Tang et al., 1997). In rhesus monkeys, age-
related demyelination is accompanied by evidence
of remyelination, at least in cerebral cortex, where
numerical densities of oligodendrocytes (Peters et al.,
1991; Peters and Sethares, 2004), grouping of oligo-
dendrocytes (Peters, 1996), and frequency of paranodal
profiles, short internodes, and thinly myelinated inter-
nodes (Peters and Sethares, 2003) all increase with age.
Some of these changes, as well as a loss of axons, are
also observed in the anterior commissure (Sandell and
Peters, 2003), corpus callosum (Peters and Sethares,
2002), and optic nerve (Sandell and Peters, 2002) of
elderly rhesus monkeys. Sloane et al. (2003) found in-
creasing calpain activation with age in rhesus white
matter, accompanied by increased levels of intact and
degraded CNPase but decreased levels of MAG [poss-
ibly explaining observations by Peters and colleagues
of impaired adherence of myelin sheaths to axons, a
process in partmediated byMAG (Kursula, 2001)] ; this
pattern of protein expression may be common to early
demyelination of various aetiologies (Aboul-Enein
et al., 2003). An interesting question is why these
monkeys, like humans (Marner et al., 2003; Tang et al.,
1997), undergo a loss of axons in white matter, while
axonal numbers are maintained in the simian cortex.
Sandell and Peters (2003) suggest a dying-back process
that affects distal axons in white matter but not proxi-
mal axons in cortex. This explanation is quite plausible ;
equally plausible is the possibility that axonal damage
is secondary to primary demyelination. White-matter
oligodendrocytes could be more vulnerable than cor-
tical ones to age-related stressors, or distal axons could
be more vulnerable than proximal ones to damage
secondary to demyelination.
In humans, imaging (reviewed in Bartzokis, 2004;
Bartzokis et al., 2006) and post-mortem studies (Ansari
and Loch, 1975; Berlet and Volk, 1980; Chia et al.,
1983; Marner et al., 2003; Meier-Ruge et al., 1992;
Miller et al., 1980; Svennerholm et al., 1991, 1997; Tang
et al., 1997; Wender et al., 1991; Wiggins et al., 1988;
Yakovlev and Lecours, 1967) indicate a loss of white
matter and its myelin content with ageing. Lintl and
Braak (1983) demonstrated an age-associated increase
in intra-cortical myelin (in the stria of Gennari) in the
first three decades, followed by a linear decrease with
age over the next seven decades, consistent with the
predictions of Yakovlev and Lecours (1967) that the
end-point of cortical myelination would be pathologi-
cal or senile demyelination. Kemper (1994), reviewing
data from the atlas of Kaes (1907), found pronounced
senile loss of myelin in association cortices, but not
in primary sensory or motor cortices. Presumably, re-
myelination also increases with age, as in rhesus
monkeys, but this remains to be proven histologically.
Preliminary data from the anterior nucleus of the
thalamus indicate an increase in oligodendrocytes
with normal ageing that is not seen in schizophrenia
(W. Byne, personal communication), suggesting that a
normal process of demyelination and remyelination
may be disrupted by schizophrenia.
It is frequently stated (e.g. Bartzokis, 2004) that the
latest axons to be myelinated are the first to be de-
myelinated. The evidence is consistent with this as-
sumption, but it is not conclusive. The most specific
statement that can be made about the effect of ageing
on white matter, based on quantitative histology, is
that there is a significant loss of myelinated fibres,
preferentially affecting those of small calibre (Marner
et al., 2003; Pakkenberg et al., 2003; Tang et al., 1997).
According to Yakovlev and Lecours (1967) and their
citations of Flechsig (1920), the latest white-matter
fibres to be myelinated are the connections between
thalamus and association cortex, long cortical associ-
ation fibres, and commissural fibres, with myelination
of the corpus callosum gradually extending from the
splenium rostrally. Kemper (1994) was of the opinion
that the material in Yakovlev’s collection showed age-
related pallor confined to corticocortical fibres. Among
the commissures of the rhesus monkey, the greatest
proportion of small-diameter fibres is in the genu of
the corpus callosum, connecting frontal association
areas (Lamantia and Rakic, 1990). DTI shows that
in normal human ageing, reduced anisotropy in the
corpus callosum is primarily in the genu (Head et al.,
2004), or at least spares the splenium (Ota et al., 2006).
Salat et al. (2005) found age-related decreases in ani-
sotropy in frontal white matter and the genu of the
corpus callosum, but also in the posterior limb of the
internal capsule [with differences between young and
White matter and cognition 523

middle-aged adults ; similarly Schneiderman et al. (in
press) found a decrease in anisotropy of the posterior
limb between adolescence and adulthood]. The pos-
terior limb of the internal capsule, containing mostly
corticospinal fibres and general somatic projections
from thalamus to postcentral gyrus, completes myeli-
nation in the first few years of life (Yakovlev and
Lecours, 1967), and reaches a plateau of anisotropy at
5–18 months of age (Morriss et al., 1999), with no fur-
ther increase between childhood and early adulthood
(Suzuki et al., 2003).
Myelination, demyelination, and remyelination are
probably all affected by genotype for apolipoprotein E
(apoE). In young adults, the e4 allele is associated with
an abnormal pattern of cortical activation during a
recall task, without deficits in performance of the task
(Scarmeas et al., 2005). Some studies find impaired
cognitive function in middle-aged and older adults
possessing an e4 allele, while others do not, and there
is controversy over whether such abnormalities rep-
resent early Alzheimer’s disease (AD) (reviewed in
Savitz et al., 2006). Presence of an e4 allele is associated
with increased risk and earlier onset of AD. In a sam-
ple of 29 cognitively normal adults with a mean age
of 65 yr, the presence of an e4 allele was associated
with a localized decrease in white-matter anisotropy
in the ventral parahippocampal region, but not 5 mm
or 10 mm dorsal to this (Nierenberg et al., 2005). The
localized effect in an age group at risk for AD suggests
that this white-matter change may reflect senile de-
generation in the entorhinal and transentorhinal cor-
tex [Braak stages I–II (Braak and Braak, 1991)], rather
than a generalized effect of the e4 allele on myelin.
Kalus et al. (2006) found localized loss of intervoxel
coherence in this region in subjects with mild cogni-
tive impairment (mean age 76 yr), presumably inde-
pendent of genotype. On the other hand, using T2
relaxation time as a measure of myelin integrity,
Bartzokis et al. (2006) found that the apoE e4 allele was
associated with decreased integrity and an exagger-
ated correlation of decreased integrity with age in
frontal white matter and genu of corpus callosum (re-
gions without apparent vulnerability in early AD)
among normal adults aged 55–75 yr, while an e2 allele
was protective.
Homozygosity for e4 is associated with AD-related
white-matter abnormalities on MRI (Bronge, 2002),
and with a mean age of onset for schizophrenia that
is 10 yr earlier than with other genotypes (Kampman
et al., 2004). As noted by Bartzokis et al. (2004), the
essential role of apoE in the recycling of lipids and
cholesterol could be crucial to myelin repair, and the
influence of genotype on this process could explain its
influence on the clinical course of a variety of brain
insults.
Onset of schizophrenia and the natural course of
myelination
Benes and colleagues have noted that the molecular
layer of the subiculum and pre- and parasubiculum is
myelinated in the second and third decades of life
(Benes, 1989; Benes et al., 1994), and they suggest that
maturation of the connections subserved by these
fibres is required for the onset of schizophrenia (Benes,
1989, 2004; Benes et al., 1994). In a slightly different
interpretation of this phenomenon, Bartzokis (2002)
suggests that schizophrenia may result from inter-
ference withmyelination at this time. In schizophrenia,
subicular apical dendrites, which extend into the
molecular layer, have decreased densities of dendritic
spines (Rosoklija et al., 2000) and decreased im-
munoreactivity for microtubule-associated proteins
2 and 5 (Arnold et al., 1991; Rosoklija et al., 2005).
mRNA for spinophilin is reduced in these neurons
(Law et al., 2004). These findings suggest that the
afferent axons may also be abnormal. However, the
question is unresolved, with only one relevant study
published. Chambers and Perrone-Bizzozero (2004)
compared optical density of MBP immunoreactivity in
16 schizophrenia subjects (10 males, mean age 52 yr;
6 females, mean age 70 yr) and 14 non-psychiatric
subjects (9 males, mean age 51 yr; 5 females, mean
age 70 yr). Among the females, optical density was
significantly lower in the schizophrenia group, while
among the males, there was no significant difference
between groups. Unfortunately, this study cannot
provide a definitive conclusion about the state of
myelination of the subicular/presubicular molecular
layer, since the number of subjects is small, and optical
density of MBP immunoreactivity is not a sensitive
method for evaluating the status of myelin [which
perhaps explains the small variance that these in-
vestigators report, while Benes et al. (1994) illustrate
considerable variability among control subjects, even
within a given age group].
DTI reveals widespread areas of white-matter
change between adolescence and adulthood
(Schneiderman et al., in press), any of which could be
a candidate for a developmental abnormality of
myelination in schizophrenia. Indeed, extension of
these studies to schizophrenia has revealed complex
patterns of differences in effects of age on FA
(Schneiderman et al., 2006, and personal communi-
cation). These important studies are somewhat limited
by cross-sectional design and modest sample size;
524 A. J. Dwork et al.

longitudinal follow-up to demonstrate maturational
changes in individuals would be invaluable.
The various presentations of metachromatic leuko-
dystrophy, an autosomal recessive defect in aryl sul-
fatase A that results in widespread demyelination,
suggests that brain functions in the process of devel-
opment may be particularly vulnerable to disruption
by demyelination. The late infantile form of meta-
chromatic leukodystrophy presents in the first 2 yr of
life with motor difficulties. The juvenile form presents
between ages 4 yr and 12 yr with behavioural and
speech difficulties, while the adult form presents after
puberty with psychiatric symptoms that are fre-
quently mistaken initially for schizophrenia. The
psychotic presentation of the adult-onset form is fre-
quently cited as evidence for a role of myelin pathol-
ogy in the aetiology of schizophrenia (e.g. Bartzokis,
2002; Hyde et al., 1992). However, we would caution
against over-interpretation of this clinical phenom-
enon. The adult onset form of globoid cell leukody-
strophy, a sphingolipidosis with similar gross
pathology (due to a defect in the next step of sphin-
golipid metabolism), presents with motor difficulties.
Both diseases, while characterized by loss of myelin,
are caused by enzymatic deficiencies that also have
other effects.
Myelin integrity and cognitive function in healthy
humans and monkeys
The relationship between cognitive function and
myelin integrity has been studied histologically in the
rhesus monkey. Demyelination and axonal degener-
ation in the anterior commissure increase with
age. Both processes presumably lead to a loss of mye-
linated axons, and the number of myelinated axons
was significantly correlated with cognitive function
between ages 5 and 20 yr. In older monkeys, aged
25–35 yr, the number of myelinated fibres was uni-
formly decreased to the minimum level found in the
younger group, and there was no correlation with
cognitive function, which was well-preserved in some
of the older animals (Sandell and Peters, 2003). How-
ever, degenerative changes of cortical myelin in frontal
area 46 (Peters and Sethares, 2002) and primary visual
cortex (Peters et al., 2000) were correlated with cogni-
tive impairment at all ages, although at least in the
visual cortex, there was no loss of fibres (Nielsen and
Peters, 2000).
In a study of 47 children, aged 5–18 yr, there was
a significant correlation between IQ and FA in
several brain areas, including frontal, parietal,
occipito-temporo-parietal, and corticospinal tracts
(Schmithorst et al., 2005). In a series of 23 children,
working memory was significantly correlated with FA
in dorsal left fronto-parietal, ventral left frontal, and
left temporo-occipital white matter and in rostral cor-
pus callosum, while reading correlated with FA in left
temporal white matter (Nagy et al., 2004). In both
studies, anisotropy in these regions also correlated
with age, suggesting that deficits were the result of
failed or delayed development. Young and middle-
aged adults with dyslexia also show decreased FA in
left temporal white matter (Klingberg et al., 2000),
suggesting persistence of a developmental deficit in-
volving temporal white matter that can still be de-
tected years later by DTI. An even more striking
example of developmental variability comes from two
studies of healthy Scottish subjects (Deary et al., 2006;
Shenkin et al., 2003) who had received IQ tests (Moray
House Test) in 1932, at age 11 yr. A total of 69 healthy
subjects were examined at ages 80 or 83 yr. Current FA
in the centrum semiovale was significantly correlated
with IQ at age 11 yr, as well as with current measures
of cognitive function and reading ability. (The cen-
trum semiovale is a large region, dorsal to the entire
body of the lateral ventricle. More detailed localization
would be of great interest.) The remarkable feature
of these studies is that normal correlates of cognitive
functional abilities are detectable by DTI years after
these abilities are established, and indeed, years after
they were measured. Long-term longitudinal studies
of DTI are not yet available. While the studies cited
would suggest that early variations persist, especially
in the case of those related to reading ability (generally
believed to be among the most stable of cognitive
functions), we would point out that this is not necess-
arily the case, and that the histological basis of a deficit
of FA might change. Theoretically, for instance, neu-
ronal pathology could first give rise to a functional
deficit without any structural abnormality in white
matter. Subsequently, this abnormality could lead to
decreased FA due to structural changes in the distal
axon, as part of a dying-back axonopathy that
eventually might result in loss of myelin.
Significant correlations have been reported between
cognitive decline in normal ageing and decreased FA
in all levels of centrum semiovale (Charlton et al.,
2006) and in rostral corpus callosum (Persson et al.,
2006b). However, Shenkin et al. (2005), in a study of
105 non-demented individuals aged 76–82 yr, found
no significant correlations of FA in frontal white
matter or centrum semiovale with any cognitive
measure, including reading ability. Studies that did
not include measures of cognitive decline found a
significant, negative correlation between age and FA
White matter and cognition 525

in frontal white matter or rostral corpus callosum, but
not in posterior regions (Head et al., 2004; Ota et al.,
2006; Pfefferbaum et al., 2005; Salat et al., 2005)
whereas posterior deficits in FA have been associated
with early-stage dementia (Head et al., 2004), and with
an apoE e4 allele in a sample of non-demented in-
dividuals aged 50–79 yr (Persson et al., 2006a).
[Presumably, these posterior deficits in white matter
are related to the posterior cortical deficits in resting
metabolism and activation that have long been re-
cognized in Alzheimer’s disease (Ingvar et al., 1975).]
Since cognitive function and frontal and rostral callo-
sal FA are all correlated with ageing, it is difficult
to determine whether age-related decline in anterior
FA is independently related to cognitive decline. After
Charlton et al. (2006) controlled for age and reading
ability, the only significant correlation that remained
was between working memory and FA in middle and
posterior centrum semiovale. It seems plausible that
a developmental component of frontal FA contributes
to a static component of cognitive function, while age-
related loss of frontal FA has little relationship with
cognitive function.
In a small autopsy study of elderly subjects with
no cognitive impairment, mild cognitive impairment,
or AD (Wang et al., 2004a), there was a significant
negative correlation between cognitive impairment
and immunoreactivity for MBP in white-matter homo-
genates from the middle frontal gyrus, and a negative
correlation of MBP with age among the unimpaired
subjects.
Myelin integrity and cognitive function in
schizophrenia
A few studies have examined the relationship between
white-matter integrity and cognitive function or
symptomatology and schizophrenia. In an autopsy
study of elderly, chronically institutionalized schizo-
phrenia subjects, dorsal prefrontal myelin ratings
(from left or right hemisphere, selected randomly)
were associated with severity of cognitive dysfunction
[evaluated by review of medical records (Ortakov
et al., 1999)] at the onset of schizophrenia (typically
y50 yr before death) and during the last years of
life, but not with the intervening change (Mancevski
et al., 2006). Age at death and neuropathological evi-
dence for vasculopathy, infarction or AD were all as-
sociated with lower myelin ratings, greater cognitive
impairment at the end of life, and greater progression
of cognitive impairment over the course of illness,
but not with severity of cognitive impairment at the
onset of illness. Thus, as suggested above for healthy
subjects, normal variability early in life may contri-
bute to a deficit of white-matter integrity and a static
component of cognitive deficit, or decreased cognitive
reserve. Whereas in healthy individuals this may
correspond only to a few points of IQ, in schizo-
phrenia it is manifested by mild but clinically evident
deficits on most items of the adapted Clinical
Dementia Rating scale (Ortakov et al., 1999).
Several DTI studies of younger (see Table 1 for ages)
schizophrenia subjects found correlations between
anisotropy and cognitive measures. Hoptman et al.
(2002) found, in a group of male schizophrenia sub-
jects, that in right ventral frontal white matter, but
not right dorsal or left, impulsiveness correlated with
decreased FA. In a subsequent voxelwise study, the
correlation with impulsiveness was confirmed, and
other regions were also involved (Hoptman et al.,
2004). These two studies did not include healthy sub-
jects, but the result is generally consistent with normal
involvement of ventral prefrontal cortex in impul-
siveness. Kubicki et al. (2002) found that in patients,
but not in comparison subjects, FA of the right uncin-
ate fasciculus at the level of the anterior commissure
correlated inversely with measures of attention and
verbal abstraction, and FA of the left with immediate
recall, but neither side was correlated with perform-
ance on the Wisconsin Card Sorting Test (WCST).
Nestor et al. (2004) found that in patients but not
comparison subjects, WCST performance was in-
versely correlated with FA in the left cingulum
bundle, while FA in the uncinate fasciculi was related
to memory functions and general intelligence. Lim
et al. (2006), in a whole-brain voxelwise study, found
significant correlations (r2o0.41), between three cog-
nitive dimensions and FA in various regions of white
matter. Verbal declarative memory was positively
correlated with FA in both hippocampi. Attention was
positively correlated with FA bilaterally in anterior
cingulate and prefrontal regions and white matter
adjacent to the caudate nucleus. Executive function
was positively correlated with FA in the anterior
cingulate region bilaterally, the corpus callosum, and
widespread areas in the left hemisphere. This sum-
mary of the findings is not exhaustive. Correlations
were found in the expected regions, as well as others,
possibly reflecting correlations among performances
in a number of cognitive domains. On all of the tests
in all of these studies there were significant positive
correlations with frontal FA, except for the WCST.
However, in the studies that employed the WCST,
the frontal region of interest was limited to the uncin-
ate fasciculus, which is situated far ventrally, while
functional imaging demonstrates activation of the
526 A. J. Dwork et al.

dorsolateral prefrontal cortex during this task
(Callicott et al., 2003; Weinberger et al., 1986).
Although correlations of cognitive performance
with FA in the uncinate fasciculus and cingulum
bundle could be explained by the neuroanatomical lo-
calization of the specific tasks in healthy people, these
correlations were found only in the schizophrenia
subjects, in whom mean values of FA in these regions
were depressed. This is apparently not a measurement
issue; performance was impaired in schizophrenia,
but variance in performance was very similar among
schizophrenia and healthy comparison subjects
(Nestor et al., 2004). This raises the possibility that,
rather than a static, developmental element of dimin-
ished cognitive reserve, the components of FA in these
two regions that correlate with specific cognitive tasks
are caused by processes related to schizophrenia itself.
A theoretical model
Characterization of white-matter abnormalities in
schizophrenia is a formidable undertaking that is still
in its early stages. While many imaging studies indi-
cate the presence of abnormalities, only a handful have
examined correlations with cognitive function. There
are no studies of the longitudinal course of FA, and
there is none of elderly subjects with schizophrenia.
There are only a few autopsy studies, and most are
small. Thus, the findings that must be explained by a
model will undoubtedly change over the next few
years.
For now, we believe that a model should explain the
following phenomena:
(1) There are deficits of white-matter integrity that are
specifically related to schizophrenia.
(2) Schizophrenia-specific white-matter deficits be-
come less prominent with advancing age.
(3) Schizophrenia-specific white-matter deficits are
related to schizophrenia-specific cognitive deficits
that are permanent and progressive, often result-
ing in dementia.
(4) There is normal developmental variability in
white-matter integrity that contributes a static
component of cognitive variability both to in-
dividuals who remain healthy and to those who
develop schizophrenia.
(5) Myelin integrity normally declines with age
through a process of demyelination and re-
myelination.
(6) Oligodendrocyte density in some areas of grey
matter increases with age in healthy humans and
monkeys.
We propose a model (Figure 1) in which a deficit of
myelin in schizophrenia arises from a partial or com-
plete failure of myelination in late adolescence and
early adulthood. The concept of ‘ last myelinated, first
demyelinated’ would predict that the fibres that fail to
myelinate in schizophrenia would be the first to de-
myelinate with age in healthy individuals, while
schizophrenia subjects would have less of this myelin
to lose at the same age. Healthy ageing individuals,
undergoing more demyelination than ageing in-
dividuals with schizophrenia, would respond with
more remyelination; hence, shorter internodes re-
quiring more oligodendrocytes. (Remyelination could
also be pathologically impaired in schizophrenia,
but in our proposed model, the efficacy of normal re-
myelination or its decrement in schizophrenia would
be insufficient to amount to a significant contribution
Age
Myelin
~18 ~25 (onset of schizophrenia) ~60
Schizophrenia
Normal
Lowdevelopmentalendowment
Highdevelopmentalendowment
Devel
opm
enta
l mye
linde
ficit
Schizophrenia-relatedmyelin deficit
Figure 1. Hypothetical model of myelination in development
and schizophrenia. In this model, normal myelination
(red lines) progresses into adulthood, and then regresses.
Differences occurring early in life (solid red line vs. dashed
red line) remain constant (shaded blue area). Myelination in
individuals who will develop schizophrenia (blue lines)
starts off normally, with similar variations among individuals
(dashed blue line vs. dotted blue line). Some time during late
adolescence, several years before the onset of schizophrenia,
myelination is interrupted, depriving affected individuals of
much (shown here as all) of the myelin normally acquired in
adulthood. For simplicity, the schizophrenia-related deficit
is indicated, by diagonal shading, only for individuals with
initially high levels of myelination. In middle age, healthy
individuals begin to lose myelin acquired in adulthood, while
individuals with schizophrenia have less (shown here as
none) such myelin to lose, so levels of myelination in healthy
individuals decline to the level seen in schizophrenia. At
older ages still (shown here as y60 yr), there may be a
loss of myelin acquired prior to late adolescence, whose
initial acquisition and senescent loss are independent of
schizophrenia.
White matter and cognition 527

to the myelin deficit.) The adverse consequences of
failed myelination in schizophrenia could be myriad,
including greater vulnerability of the axon to injury,
greater metabolic requirements of the neuron, im-
paired transport of neurotrophic factors, and trans-
synaptic effects on axonal targets. These effects could
already be manifest at the onset of clinical disease,
which may follow several years of failing myelination,
and they would become more severe over time, but
they would not necessarily entail further deficits of
myelin. A normal, earlier-developing, stable varia-
bility in white-matter integrity, corresponding to a
component of variability among individuals in natural
cognitive endowment, would be superimposed on this
process.
Acknowledgements
This work was supported by Supported by MH60877,
MH64168, the National Alliance for Research on
Schizophrenia and Depression, and the Lieber Center
for Schizophrenia Research at the Department of
Psychiatry, College of Physicians and Surgeons of
Columbia University. We thank Dr Jasmine Chen and
Dr Cheryl Corcoran for thoughtful review of the
manuscript, and Dr Ana Sotrel for helpful discussions.
Statement of Interest
None.
References
Aberg K, Saetre P, Jareborg N, Jazin E (2006a). Human QKI,
a potential regulator of mRNA expression of human
oligodendrocyte-related genes involved in schizophrenia.
Proceedings of the National Academy of Sciences USA 103,
7482–7487.
Aberg K, Saetre P, Lindholm E, Ekholm B, Pettersson U,
Adolfsson R, Jazin E (2006b). Human QKI, a new
candidate gene for schizophrenia involved in myelination.
American Journal of Medical Genetics, B: Neuropsychiatric
Genetics 141, 84–90.
Aboul-Enein F, Rauschka H, Kornek B, Stadelmann C,
Stefferl A, Bruck W, Lucchinetti C, Schmidbauer M,
Jellinger K, Lassmann H (2003). Preferential loss of
myelin-associated glycoprotein reflects hypoxia-like
white matter damage in stroke and inflammatory brain
diseases. Journal of Neuropathology and Experimental
Neurology 62, 25–33.
Agartz I, Andersson JL, Skare S (2001). Abnormal brain
white matter in schizophrenia: a diffusion tensor imaging
study. Neuroreport 12, 2251–2254.
Ansari KA, Loch J (1975). Decreased myelin basic protein
content of the aged human brain. Neurology 25, 1045–1050.
Ardekani BA, Nierenberg J, Hoptman MJ, Javitt DC,
Lim KO (2003). MRI study of white matter diffusion
anisotropy in schizophrenia. Neuroreport 14, 2025–2029.
Arnold SE, Gur RE, Shapiro RM, Fisher KR, Moberg PJ,
Gibney MR, Gur RC, Blackwell P, Trojanowski JQ
(1995). Prospective clinicopathologic studies of
schizophrenia: accrual and assessment of patients.
American Journal of Psychiatry 152, 731–737.
Arnold SE, Lee VM, Gur RE, Trojanowski JQ (1991).
Abnormal expression of two microtubule-associated
proteins (MAP2 and MAP5) in specific subfields of the
hippocampal formation in schizophrenia. Proceedings of
the National Academy of Sciences USA 88, 10850–10854.
Arnold SE, Trojanowski JQ, Gur RE, Blackwell P, Han LY,
Choi C (1998). Absence of neurodegeneration and neural
injury in the cerebral cortex in a sample of elderly patients
with schizophrenia. Archives of General Psychiatry 55,
225–232.
Auer DP, WilkeM, Grabner A, Heidenreich JO, Bronisch T,
Wetter TC (2001). Reduced NAA in the thalamus and
altered membrane and glial metabolism in schizophrenic
patients detected by 1H-MRS and tissue segmentation.
Schizophrenia Research 52, 87–99.
BagaryMS, SymmsMR, Barker GJ, Mutsatsa SH, Joyce EM,
Ron MA (2003). Gray and white matter brain
abnormalities in first-episode schizophrenia inferred
from magnetization transfer imaging. Archives of General
Psychiatry 60, 779–788.
Barnett JH, Salmond CH, Jones PB, Sahakian BJ (2006).
Cognitive reserve in neuropsychiatry. Psychological
Medicine 36, 1053–1064.
Bartha R, al-Semaan YM, Williamson PC, Drost DJ,
Malla AK, Carr TJ, DensmoreM, CanaranG, Neufeld RW
(1999). A short echo proton magnetic resonance
spectroscopy study of the left mesial-temporal lobe in
first-onset schizophrenic patients. Biological Psychiatry 45,
1403–1411.
Bartzokis G (2002). Schizophrenia: breakdown in the
well-regulated lifelong process of brain development
and maturation [see comment]. Neuropsychopharmacology
27, 672–683.
Bartzokis G (2004). Age-related myelin breakdown: a
developmental model of cognitive decline and Alzheimer’s
disease. Neurobiology of Aging 25, 5–18 (author reply,
49–62).
Bartzokis G, Lu PH, Geschwind DH, Edwards N, Mintz J,
Cummings JL (2006). Apolipoprotein E genotype and
age-related myelin breakdown in healthy individuals:
implications for cognitive decline and dementia.
Archives of General Psychiatry 63, 63–72.
Bartzokis G, Sultzer D, Lu PH, Nuechterlein KH, Mintz J,
Cummings JL (2004). Heterogeneous age-related
breakdown of white matter structural integrity:
implications for cortical ‘disconnection’ in aging and
Alzheimer’s disease. Neurobiology of Aging 25, 843–851.
Basser PJ, Jones DK (2002). Diffusion-tensor MRI: theory,
experimental design and data analysis – a technical
review. NMR in Biomedicine 15, 456–467.
528 A. J. Dwork et al.

Basser PJ, Pierpaoli C (1996). Microstructural and
physiological features of tissues elucidated by
quantitative-diffusion-tensor MRI. Journal of Magnetic
Resonance, Series B 111, 209–219.
Beasley C, Dracheva S, Rosoklija G, Serafimova T,
Mancevski B, Duma A, JJ M, WG H, Haroutunian V,
Dwork AJ (in press). Myelin-associated proteins and
mRNA in the cingulum bundle. International Congress
on Schizophrenia Research, Colorado Springs, CO,
28 March–1 April, 2007.
Beaulieu C (2002). The basis of anisotropic water diffusion
in the nervous system – a technical review. NMR in
Biomedicine 15, 435–455.
Begre S, Federspiel A, Kiefer C, Schroth G, Dierks T,
Strik WK (2003). Reduced hippocampal anisotropy
related to anteriorization of alpha EEG in schizophrenia.
Neuroreport 14, 739–742.
Benes FM (1989). Myelination of cortical-hippocampal relays
during late adolescence. Schizophrenia Bulletin 15, 585–593.
Benes FM (2004). A disturbance of late myelination as a
trigger for Alzheimer’s disease. Neurobiology of Aging 25,
41–43.
Benes FM, Turtle M, Khan Y, Farol P (1994). Myelination
of a key relay zone in the hippocampal formation occurs
in the human brain during childhood, adolescence, and
adulthood. Archives of General Psychiatry 51, 477–484.
Berlet HH, Volk B (1980). Studies of human myelin proteins
during old age. Mechanisms of Ageing and Development 14,
211–222.
Bigio EH, Vono MB, Satumtira S, Adamson J, Sontag E,
Hynan LS, White III CL, Baker M, Hutton M (2001).
Cortical synapse loss in progressive supranuclear palsy.
Journal of Neuropathology and Experimental Neurology 60,
403–410.
Braak H, Braak E (1991). Neuropathological stageing of
Alzheimer-related changes. Acta Neuropathologica 82,
239–259.
Brambilla P, Cerini R, Gasparini A, Versace A, Andreone N,
Vittorini E, Barbui C, Pelizza L, Nose M, Barlocco L, et al.
(2005). Investigation of corpus callosum in schizophrenia
with diffusion imaging. Schizophrenia Research 79, 201–210.
Bronge L (2002). Magnetic resonance imaging in dementia. A
study of brain white matter changes. Acta Radiologica
428 (Suppl.), 1–32.
Brown FW, Lewine RJ, Hudgins PA, Risch SC (1992). White
matter hyperintensity signals in psychiatric and
nonpsychiatric subjects. American Journal of Psychiatry 149,
620–625.
Brown FW, Lewine RR, Hudgins PA (1995). White matter
hyperintensity signals associated with vascular risk factors
in schizophrenia. Progress in Neuro-Psychopharmacology and
Biological Psychiatry 19, 39–45.
BrowningMD, Dudek EM, Rapier JL, Leonard S, Freedman
R (1993). Significant reductions in synapsin but not
synaptophysin specific activity in the brains of some
schizophrenics. Biological Psychiatry 34, 529–535.
Buchsbaum MS, Friedman J, Buchsbaum BR, Chu KW,
Hazlett EA, Newmark R, Schneiderman JS, Torosjan Y,
Tang C, Hof PR, et al. (2006a). Diffusion tensor imaging in
schizophrenia. Biological Psychiatry 60, 1181–1187.
Buchsbaum MS, Schoenknecht P, Torosjan Y, Newmark R,
Chu KW, Mitelman S, Brickman AM, Shihabuddin L,
Haznedar MM, Hazlett EA, et al. (2006b). Diffusion tensor
imaging of frontal lobe white matter tracts in
schizophrenia. Annals of General Psychiatry 5, 19.
Buchsbaum MS, Tang CY, Peled S, Gudbjartsson H, Lu D,
Hazlett EA, Downhill J, Haznedar M, Fallon JH,
Atlas SW (1998). MRI white matter diffusion anisotropy
and PET metabolic rate in schizophrenia. Neuroreport 9,
425–430.
Burns J, Job D, Bastin ME, Whalley H, Macgillivray T,
Johnstone EC, Lawrie SM (2003). Structural
disconnectivity in schizophrenia: a diffusion tensor
magnetic resonance imaging study. British Journal of
Psychiatry 182, 439–443.
Byne W, Buchsbaum MS, Mattiace LA, Hazlett EA,
Kemether E, Elhakem SL, Purohit DP, Haroutunian V,
Jones L (2002). Postmortem assessment of thalamic nuclear
volumes in subjects with schizophrenia. American Journal of
Psychiatry 159, 59–65.
Callicott JH, Egan MF, Mattay VS, Bertolino A, Bone AD,
Verchinksi B, Weinberger DR (2003). Abnormal fMRI
response of the dorsolateral prefrontal cortex in cognitively
intact siblings of patients with schizophrenia. American
Journal of Psychiatry 160, 709–719.
Casanova MF, Zito M, Bigelow LB, Berthot B, Sanders RD,
Kleinman JE (1989). Axonal counts of the corpus callosum
of schizophrenic patients. Journal of Neuropsychiatry and
Clinical Neurosciences 1, 391–393.
Chambers JS, Perrone-Bizzozero NI (2004). Altered
myelination of the hippocampal formation in subjects with
schizophrenia and bipolar disorder. Neurochemical Research
29, 2293–2302.
Chance SA, Highley JR, Esiri MM, Crow TJ (1999). Fiber
content of the fornix in schizophrenia: lack of evidence for
a primary limbic encephalopathy. American Journal of
Psychiatry 156, 1720–1724.
Charlton RA, Barrick TR, McIntyre DJ, Shen Y,
O’Sullivan M, Howe FA, Clark CA, Morris RG,
Markus HS (2006). White matter damage on diffusion
tensor imaging correlates with age-related cognitive
decline. Neurology 66, 217–222.
Chia LS, Thompson JE, Moscarello MA (1983). Changes in
lipid phase behaviour in human myelin during maturation
and aging. Involvement of lipid peroxidation. FEBS Letters
157, 155–158.
Chin SS, Goldman JE (1996). Glial inclusions in CNS
degenerative diseases. Journal of Neuropathology and
Experimental Neurology 55, 499–508.
Choe BY, Kim KT, Suh TS, Lee C, Paik IH, Bahk YW,
Shinn KS, Lenkinski RE (1994). 1H magnetic resonance
spectroscopy characterization of neuronal dysfunction in
drug-naive, chronic schizophrenia. Academic Radiology 1,
211–216.
Coleman MP, Ribchester RR (2004). Programmed axon
death, synaptic dysfunction and the ubiquitin proteasome
White matter and cognition 529

system. Current Drug Targets – CNS & Neurological
Disorders 3, 227–238.
Craner MJ, Newcombe J, Black JA, Hartle C, Cuzner ML,
Waxman SG (2004). Molecular changes in neurons in
multiple sclerosis: altered axonal expression of Nav1.2
and Nav1.6 sodium channels and Na+/Ca2+ exchanger.
Proceedings of the National Academy of Sciences USA 101,
8168–8173.
Davidson M, Harvey PD, Powchik P, Parrella M, White L,
Knobler HY, Losonczy MF, Keefe RS, Katz S, Frecska E
(1995). Severity of symptoms in chronically
institutionalized geriatric schizophrenic patients.
American Journal of Psychiatry 152, 197–207.
Davidsson P, Gottfries J, Bogdanovic N, Ekman R,
Karlsson I, Gottfries CG, Blennow K (1999). The
synaptic-vesicle-specific proteins rab3a and synaptophysin
are reduced in thalamus and related cortical brain
regions in schizophrenic brains. Schizophrenia Research 40,
23–29.
Davis KL, Stewart DG, Friedman JI, Buchsbaum M,
Harvey PD, Hof PR, Buxbaum J, Haroutunian V (2003).
White matter changes in schizophrenia: evidence for
myelin-related dysfunction. Archives of General Psychiatry
60, 443–456.
Dawson DA, Hallenbeck JM (1996). Acute focal ischemia-
induced alterations in MAP2 immunostaining: description
of temporal changes and utilization as a marker for
volumetric assessment of acute brain injury. Journal of
Cerebral Blood Flow and Metabolism 16, 170–174.
Deary IJ, Bastin ME, Pattie A, Clayden JD, Whalley LJ,
Starr JM, Wardlaw JM (2006). White matter integrity
and cognition in childhood and old age [see comment].
Neurology 66, 505–512.
DeKosky ST, Scheff SW (1990). Synapse loss in frontal cortex
biopsies in Alzheimer’s disease: correlation with cognitive
severity. Annals of Neurology 27, 457–464.
Dik M, Deeg DJ, Visser M, Jonker C (2003). Early life
physical activity and cognition at old age. Journal of Clinical
and Experimental Neuropsychology 25, 643–653.
Dorph-Petersen KA, Pierri JN, Sun Z, Sampson AR,
Lewis DA (2004). Stereological analysis of the mediodorsal
thalamic nucleus in schizophrenia: volume, neuron
number, and cell types. Journal of Comparative Neurology
472, 449–462.
Dracheva S, Davis KL, Chin B, Woo DA, Schmeidler J,
Haroutunian V (2006). Myelin-associated mRNA and
protein expression deficits in the anterior cingulate cortex
and hippocampus in elderly schizophrenia patients.
Neurobiology of Disease 21, 531–540.
Dwork A, Ilievski B, Mancevski B, Kurzon M, Serafimova
T, Trencevska I, Rosoklija G, Keilp J (2005). Histological
evaluation of prefrontal white matter in schizophrenia.
Society for Neuroscience Abstracts. Abstract no. 474.478.
Dwork AJ (1997). Postmortem studies of the hippocampal
formation in schizophrenia. Schizophrenia Bulletin 23,
385–402.
Dwork AJ, Susser ES, Keilp J, Waniek C, Liu D, Kaufman
M, Zemishlany Z, Prohovnik I (1998). Senile degeneration
and cognitive impairment in chronic schizophrenia.
American Journal of Psychiatry 155, 1536–1543.
Eastwood SL, Burnet PW, Harrison PJ (1995). Altered
synaptophysin expression as a marker of synaptic
pathology in schizophrenia. Neuroscience 66, 309–319.
Eastwood SL, Harrison PJ (1995). Decreased synaptophysin
in the medial temporal lobe in schizophrenia
demonstrated using immunoautoradiography.
Neuroscience 69, 339–343.
Eastwood SL, Harrison PJ (1999). Detection and
quantification of hippocampal synaptophysin messenger
RNA in schizophrenia using autoclaved, formalin-fixed,
paraffin wax-embedded sections. Neuroscience 93, 99–106.
Falkai P, Honer WG, David S, Bogerts B, Majtenyi C, Bayer
TA (1999). No evidence for astrogliosis in brains of
schizophrenic patients. A post-mortem study.
Neuropathology and Applied Neurobiology 25, 48–53.
Federspiel A, Begre S, Kiefer C, Schroth G, Strik WK,
Dierks T (2006). Alterations of white matter connectivity
in first episode schizophrenia. Neurobiology of Disease 22,
702–709.
Flechsig P (1920). Anatomie des menschlichen Gehirns und
Ruckenmarks auf myelogenetischer Grundlage. Leipzig:
G. Thieme.
Flynn SW, Lang DJ, Mackay AL, Goghari V, Vavasour IM,
Whittall KP, Smith GN, Arango V, Mann JJ, Dwork AJ,
et al. (2003). Abnormalities of myelination in
schizophrenia detected in vivo with MRI, and post-
mortem with analysis of oligodendrocyte proteins.
Molecular Psychiatry 8, 811–820.
Foong J, Maier M, Barker GJ, Brocklehurst S, Miller DH,
Ron MA (2000a). In vivo investigation of white matter
pathology in schizophrenia with magnetisation transfer
imaging. Journal of Neurology, Neurosurgery and Psychiatry
68, 70–74.
Foong J, MaierM, Clark CA, Barker GJ, Miller DH, RonMA
(2000b). Neuropathological abnormalities of the corpus
callosum in schizophrenia: a diffusion tensor imaging
study. Journal of Neurology, Neurosurgery and Psychiatry 68,
242–244.
Foong J, Symms MR, Barker GJ, Maier M, Miller DH,
Ron MA (2002). Investigating regional white matter in
schizophrenia using diffusion tensor imaging. Neuroreport
13, 333–336.
Foong J, Symms MR, Barker GJ, Maier M, Woermann FG,
Miller DH, Ron MA (2001). Neuropathological
abnormalities in schizophrenia: evidence from
magnetization transfer imaging. Brain 124, 882–892.
Gabriel SM, Haroutunian V, Powchik P, Honer WG,
Davidson M, Davies P (1997). Increased concentrations of
presynaptic proteins in the cingulate cortex of subjects
with schizophrenia. Archives of General Psychiatry 54,
559–566.
Garey LJ, Ong WY, Patel TS, Kanani M, Davis A, Mortimer
AM, Barnes TR, Hirsch SR (1998). Reduced dendritic
spine density on cerebral cortical pyramidal neurons in
schizophrenia. Journal of Neurology, Neurosurgery and
Psychiatry 65, 446–453.
530 A. J. Dwork et al.

Glantz LA, Lewis DA (1997). Reduction of synaptophysin
immunoreactivity in the prefrontal cortex of subjects with
schizophrenia. Regional and diagnostic specificity.
Archives of General Psychiatry 54, 943–952.
Glantz LA, Lewis DA (2000). Decreased dendritic spine
density on prefrontal cortical pyramidal neurons in
schizophrenia. Archives of General Psychiatry 57, 65–73.
Goldberg TE, Weinberger DR (1988). Probing prefrontal
function in schizophrenia with neuropsychological
paradigms. Schizophrenia Bulletin 14, 179–183.
Goto S, Hirano A (1990). Neuronal inputs to hippocampal
formation in Alzheimer’s disease and in parkinsonism-
dementia complex on Guam. Acta Neuropathologica 79,
545–550.
Graeber M, Blakemore W, Kreutzberg G (2002). Celllular
pathology of the central nervous system. In: Graham D,
Lantos P (Eds.), Greenfield’s Neuropathology (pp. 123–191).
New York: Arnold.
Green MF, Nuechterlein KH, Gold JM, Barch DM, Cohen J,
Essock S, Fenton WS, Frese F, Goldberg TE, Heaton RK,
et al. (2004). Approaching a consensus cognitive battery
for clinical trials in schizophrenia: the NIMH-MATRICS
conference to select cognitive domains and test criteria.
Biological Psychiatry 56, 301–307.
Grossman RI, Gomori JM, Ramer KN, Lexa FJ, Schnall MD
(1994). Magnetization transfer: theory and clinical
applications in neuroradiology. Radiographics 14, 279–290.
Hachinski VC (1990). The decline and resurgence of vascular
dementia. Canadian Medical Association Journal 142,
107–111.
Hakak Y, Walker JR, Li C, Wong WH, Davis KL,
Buxbaum JD, Haroutunian V, Fienberg AA (2001).
Genome-wide expression analysis reveals dysregulation of
myelination-related genes in chronic schizophrenia.
Proceedings of the National Academy of Sciences USA 98,
4746–4751.
Hammen T, Stefan H, Eberhardt KE, W-Huk BH,
Tomandl BF (2003). Clinical applications of 1H-MR
spectroscopy in the evaluation of epilepsies – what do
pathological spectra stand for with regard to current
results and what answers do they give to common clinical
questions concerning the treatment of epilepsies?
Acta Neurologica Scandinavica 108, 223–238.
Hamos JE, DeGennaro LJ, Drachman DA (1989). Synaptic
loss in Alzheimer’s disease and other dementias.Neurology
39, 355–361.
Hao J, JanssenWG, Tang Y, Roberts JA, McKay H, Lasley B,
Allen PB, Greengard P, Rapp PR, Kordower JH, et al.
(2003). Estrogen increases the number of spinophilin-
immunoreactive spines in the hippocampus of young and
aged female rhesus monkeys. Journal of Comparative
Neurology 465, 540–550.
Hao Y, Liu Z, Jiang T, Gong G, Liu H, Tan L, Kuang F, Xu L,
Yi Y, Zhang Z (2006). White matter integrity of the whole
brain is disrupted in first-episode schizophrenia.
Neuroreport 17, 23–26.
Haroutunian V, Davidson M, Kanof PD, Perl DP, Powchik
P, Losonczy M, McCrystal J, Purohit DP, Bierer LM,
Davis KL (1994). Cortical cholinergic markers in
schizophrenia. Schizophrenia Research 12, 137–144.
Harrison PJ (1999). The neuropathology of schizophrenia.
A critical review of the data and their interpretation.
Brain 122, 593–624.
Harrison PJ, Weinberger DR (2005). Schizophrenia genes,
gene expression, and neuropathology: on the matter
of their convergence [Erratum appears in Molecular
Psychiatry (2005) 10, 420]. Molecular Psychiatry 10, 40–68
(image 45).
Head D, Buckner RL, Shimony JS, Williams LE, Akbudak
E, Conturo TE, McAvoy M, Morris JC, Snyder AZ (2004).
Differential vulnerability of anterior white matter in
nondemented aging with minimal acceleration in
dementia of the Alzheimer type: evidence from diffusion
tensor imaging. Cerebral Cortex 14, 410–423.
Heckers S (1997). Neuropathology of schizophrenia: cortex,
thalamus, basal ganglia, and neurotransmitter-specific
projection systems. Schizophrenia Bulletin 23, 403–421.
Highley JR, Esiri MM, McDonald B, Cortina-Borja M,
Herron BM, Crow TJ (1999a). The size and fibre
composition of the corpus callosum with respect to gender
and schizophrenia: a post-mortem study. Brain 122,
99–110.
Highley JR, Esiri MM, McDonald B, Roberts HC, Walker
MA, Crow TJ (1999b). The size and fiber composition of
the anterior commissure with respect to gender and
schizophrenia. Biological Psychiatry 45, 1120–1127.
Highley JR, Walker MA, Esiri MM, Crow TJ, Harrison PJ
(2002). Asymmetry of the uncinate fasciculus: a post-
mortem study of normal subjects and patients with
schizophrenia. Cerebral Cortex 12, 1218–1224.
Hof PR, Haroutunian V, Copland C, Davis KL, Buxbaum JD
(2002). Molecular and cellular evidence for an
oligodendrocyte abnormality in schizophrenia.
Neurochemical Research 27, 1193–1200.
Hof PR, Haroutunian V, Friedrich Jr. VL, Byne W, Buitron
C, Perl DP, Davis KL (2003). Loss and altered spatial
distribution of oligodendrocytes in the superior frontal
gyrus in schizophrenia. Biological Psychiatry 53, 1075–1085.
Honer WG, Dickson DW, Gleeson J, Davies P (1992).
Regional synaptic pathology in Alzheimer’s disease.
Neurobiology of Aging 13, 375–382.
Honer WG, Falkai P, Young C, Wang T, Xie J, Bonner J,
Hu L, BoulianneGL, Luo Z, TrimbleWS (1997). Cingulate
cortex synaptic terminal proteins and neural cell adhesion
molecule in schizophrenia. Neuroscience 78, 99–110.
Honer WG, Young CE (2004). Presynaptic proteins and
schizophrenia. International Review of Neurobiology 59,
175–199.
Hoptman MJ, Ardekani BA, Butler PD, Nierenberg J, Javitt
DC, Lim KO (2004). DTI and impulsivity in schizophrenia:
a first voxelwise correlational analysis. Neuroreport 15,
2467–2470.
Hoptman MJ, Volavka J, Johnson G, Weiss E, Bilder RM,
Lim KO (2002). Frontal white matter microstructure,
aggression, and impulsivity in men with schizophrenia: a
preliminary study. Biological Psychiatry 52, 9–14.
White matter and cognition 531

Hubl D, Koenig T, StrikW, Federspiel A, Kreis R, Boesch C,
Maier SE, Schroth G, Lovblad K, Dierks T (2004).
Pathways that make voices: white matter changes in
auditory hallucinations. Archives of General Psychiatry 61,
658–668.
Hulshoff Pol HE, Hoek HW, Susser E, Brown AS,
Dingemans A, Schnack HG, van Haren NE, Pereira
Ramos LM, Gispen-de Wied CC, Kahn RS (2000).
Prenatal exposure to famine and brain morphology in
schizophrenia. American Journal of Psychiatry 157,
1170–1172.
Hyde TM, Ziegler JC, Weinberger DR (1992). Psychiatric
disturbances in metachromatic leukodystrophy. Insights
into the neurobiology of psychosis [see comment].
Archives of Neurology 49, 401–406.
Ingvar DH, Risberg J, Schwartz MS (1975). Evidence of
subnormal function of association cortex in presenile
dementia. Neurology 25, 964–974.
Jones DK, Catani M, Pierpaoli C, Reeves SJ, Shergill SS,
O’SullivanM, Golesworthy P, McGuire P, HorsfieldMA,
Simmons A, et al. (2006). Age effects on diffusion tensor
magnetic resonance imaging tractography measures of
frontal cortex connections in schizophrenia. Human Brain
Mapping 27, 230–238.
Jones DK, Symms MR, Cercignani M, Howard RJ (2005).
The effect of filter size on VBM analyses of DT-MRI data.
Neuroimage 26, 546–554.
Kaes T (1907). Die Grosshirnrinde des Menschen in ihern Massen
und in ihrem Fasergehalt. Jena: Gustav Fisher.
Kalus P, Slotboom J, Gallinat J, Mahlberg R, Cattapan-
Ludewig K, Wiest R, Nyffeler T, Buri C, Federspiel A,
Kunz D, et al. (2006). Examining the gateway to the limbic
system with diffusion tensor imaging: the perforant
pathway in dementia. Neuroimage 30, 713–720.
Kampman O, Anttila S, Illi A, Mattila KM, Rontu R,
Leinonen E, Lehtimaki T (2004). Apolipoprotein E
polymorphism is associated with age of onset in
schizophrenia. Journal of Human Genetics 49, 355–359.
Karson CN, Mrak RE, Schluterman KO, Sturner WQ,
Sheng JG, Griffin WS (1999). Alterations in synaptic
proteins and their encoding mRNAs in prefrontal cortex
in schizophrenia: a possible neurochemical basis for
‘hypofrontality’. Molecular Psychiatry 4, 39–45.
Kemper T (1994). Neuroanatomical and neuropathological
changes during aging and dementia. In: Albert M, Knoefel
J (Eds.), Clinical Neurology of Aging (pp. 3–67). New York:
Oxford University Press.
Keshavan MS, Mulsant BH, Sweet RA, Pasternak R,
Zubenko GS, Krishnan RR (1996). MRI changes in
schizophrenia in late life: a preliminary controlled study.
Psychiatry Research 60, 117–123.
Kim JH, Budde MD, Liang HF, Klein RS, Russell JH, Cross
AH, Song SK (2006). Detecting axon damage in spinal cord
from a mouse model of multiple sclerosis. Neurobiology of
Disease 21, 626–632.
Kingsley PB, Monahan WG (2005). Contrast-to-noise ratios
of diffusion anisotropy indices. Magnetic Resonance in
Medicine 53, 911–918.
Kitamura H, Matsuzawa H, Shioiri T, Someya T, Kwee IL,
Nakada T (2005). Diffusion tensor analysis in chronic
schizophrenia. A preliminary study on a high-field (3.0T)
system. European Archives of Psychiatry and Clinical
Neuroscience 255, 313–318.
Klingberg T, Hedehus M, Temple E, Salz T, Gabrieli JD,
Moseley ME, Poldrack RA (2000). Microstructure of
temporo-parietal white matter as a basis for reading
ability: evidence from diffusion tensor magnetic resonance
imaging. Neuron 25, 493–500.
Kubicki M, Park H, Westin CF, Nestor PG, Mulkern RV,
Maier SE, Niznikiewicz M, Connor EE, Levitt JJ,
Frumin M, et al. (2005). DTI and MTR abnormalities in
schizophrenia: analysis of white matter integrity.
Neuroimage 26, 1109–1118.
Kubicki M, Westin CF, Maier SE, Frumin M, Nestor PG,
Salisbury DF, Kikinis R, Jolesz FA, McCarley RW,
Shenton ME (2002). Uncinate fasciculus findings in
schizophrenia: a magnetic resonance diffusion tensor
imaging study. American Journal of Psychiatry 159, 813–820.
Kubicki M, Westin CF, Nestor PG, Wible CG, Frumin M,
Maier SE, Kikinis R, Jolesz FA, McCarley RW, Shenton
ME (2003). Cingulate fasciculus integrity disruption in
schizophrenia: a magnetic resonance diffusion tensor
imaging study. Biological Psychiatry 54, 1171–1180.
Kumra S, Ashtari M, McMeniman M, Vogel J, Augustin R,
Becker DE, Nakayama E, Gyato K, Kane JM, Lim K,
Szeszko P (2004). Reduced frontal white matter integrity
in early-onset schizophrenia: a preliminary study.
Biological Psychiatry 55, 1138–1145.
Kuroki N, Kubicki M, Nestor PG, Salisbury DF, Park HJ,
Levitt JJ, Woolston S, Frumin M, Niznikiewicz M,
Westin CF, et al. (2006). Fornix integrity and hippocampal
volume in male schizophrenic patients. Biological
Psychiatry 60, 22–31.
Kursula P (2001). The current status of structural studies on
proteins of the myelin sheath [Review]. International Journal
of Molecular Medicine 8, 475–479.
Lamantia AS, Rakic P (1990). Cytological and quantitative
characteristics of four cerebral commissures in the
rhesus monkey. Journal of Comparative Neurology 291,
520–537.
Lane B, Sullivan EV, Lim KO, Beal DM, Harvey Jr. RL,
Meyers T, Faustman WO, Pfefferbaum A (1996). White
matter MR hyperintensities in adult patients with
congenital rubella. American Journal of Neuroradiology 17,
99–103.
Law AJ, Weickert CS, Hyde TM, Kleinman JE, Harrison PJ
(2004). Reduced spinophilin but not microtubule-
associated protein 2 expression in the hippocampal
formation in schizophrenia and mood disorders:
molecular evidence for a pathology of dendritic spines.
American Journal of Psychiatry 161, 1848–1855.
Lim KO, Adalsteinsson E, Spielman D, Sullivan EV,
Rosenbloom MJ, Pfefferbaum A (1998). Proton magnetic
resonance spectroscopic imaging of cortical gray and
white matter in schizophrenia. Archives of General
Psychiatry 55, 346–352.
532 A. J. Dwork et al.

Lim KO, Ardekani BA, Nierenberg J, Butler PD, Javitt DC,
Hoptman MJ (2006). Voxelwise correlational analyses of
white matter integrity in multiple cognitive domains in
schizophrenia. American Journal of Psychiatry 163,
2008–2010.
Lim KO, HedehusM, Moseley M, de Crespigny A, Sullivan
EV, Pfefferbaum A (1999). Compromised white matter
tract integrity in schizophrenia inferred from diffusion
tensor imaging. Archives of General Psychiatry 56, 367–374.
Lintl P, Braak H (1983). Loss of intracortical myelinated
fibers: a distinctive age-related alteration in the human
striate area. Acta Neuropathologica 61, 178–182.
Liston C, Watts R, Tottenham N, Davidson MC, Niogi S,
Ulug AM, Casey BJ (2006). Frontostriatal microstructure
modulates efficient recruitment of cognitive control.
Cerebral Cortex 16, 553–560.
Mancevski B, Ilievski B, Trencevska I, Serafimova T,
Rosoklija G, Keilp J, Dwork A (2006). Myelin and
cognition in schizophrenia. Society for Neuroscience
Abstracts. Abstract no. 20.26.
Marner L, Nyengaard JR, Tang Y, Pakkenberg B (2003).
Marked loss of myelinated nerve fibers in the human brain
with age. Journal of Comparative Neurology 462, 144–152.
Marner L, Pakkenberg B (2003). Total length of nerve fibers
in prefrontal and global white matter of chronic
schizophrenics. Journal of Psychiatric Research 37, 539–547.
Masliah E, Terry RD, Alford M, DeTeresa R, Hansen LA
(1991). Cortical and subcortical patterns of
synaptophysinlike immunoreactivity in Alzheimer’s
disease. American Journal of Pathology 138, 235–246.
Masliah E, Terry RD, DeTeresa RM, Hansen LA (1989).
Immunohistochemical quantification of the synapse-
related protein synaptophysin in Alzheimer disease.
Neuroscience Letters 103, 234–239.
McCullumsmith R, Gupta D, Beneyto M, Kreger E,
Haroutunian V, Davis K, Meador-Woodruff J (2007).
Expression of transcripts for myelin-associated genes
in the anterior cingulate cortex in schizophrenia.
Schizophrenia Research 90, 15–27.
Meier-Ruge W, Ulrich J, Bruhlmann M, Meier E (1992).
Age-related white matter atrophy in the human brain.
Annals of the New York Academy of Sciences 673, 260–269.
Michailov GV, Sereda MW, Brinkmann BG, Fischer TM,
Haug B, Birchmeier C, Role L, Lai C, Schwab MH, Nave
KA (2004). Axonal neuregulin-1 regulates myelin sheath
thickness [see comment]. Science 304, 700–703.
Miller AK, Alston RL, Corsellis JA (1980). Variation with
age in the volumes of grey and white matter in the cerebral
hemispheres of man: measurements with an image
analyser. Neuropathology and Applied Neurobiology 6,
119–132.
Minami T, Nobuhara K, Okugawa G, Takase K, Yoshida T,
Sawada S, Ha-Kawa S, Ikeda K, Kinoshita T (2003).
Diffusion tensor magnetic resonance imaging of disruption
of regional white matter in schizophrenia.
Neuropsychobiology 47, 141–145.
Miyakawa T, Sumiyoshi S, Deshimaru M, Suzuki T,
Tomonari H (1972). Electron microscopic study on
schizophrenia. Mechanism of pathological changes.
Acta Neuropathologica 20, 67–77.
Morriss MC, Zimmerman RA, Bilaniuk LT, Hunter JV,
Haselgrove JC (1999). Changes in brain water diffusion
during childhood. Neuroradiology 41, 929–934.
Nagy Z, Westerberg H, Klingberg T (2004). Maturation of
white matter is associated with the development of
cognitive functions during childhood. Journal of Cognitive
Neuroscience 16, 1227–1233.
Nasrallah HA,McCalley-WhittersM, Bigelow LB, Rauscher
FP (1983). A histological study of the corpus callosum in
chronic schizophrenia. Psychiatry Research 8, 251–260.
Nebes RD, Meltzer CC, Whyte EM, Scanlon JM, Halligan
EM, Saxton JA, Houck PR, Boada FE, Dekosky ST (2006).
The relation of white matter hyperintensities to cognitive
performance in the normal old: education matters.
Neuropsychology, Development, and Cognition. Section B,
Aging, Neuropsychology and Cognition 13, 326–340.
Nestor PG, Kubicki M, Gurrera RJ, Niznikiewicz M,
Frumin M, McCarley RW, Shenton ME (2004).
Neuropsychological correlates of diffusion tensor imaging
in schizophrenia. Neuropsychology 18, 629–637.
Nielsen K, Peters A (2000). The effects of aging on the
frequency of nerve fibers in rhesus monkey striate cortex.
Neurobiology of Aging 21, 621–628.
Nierenberg J, Pomara N, Hoptman MJ, Sidtis JJ,
Ardekani BA, Lim KO (2005). Abnormal white matter
integrity in healthy apolipoprotein E epsilon4 carriers.
Neuroreport 16, 1369–1372.
Okugawa G, Nobuhara K, Minami T, Tamagaki C,
Takase K, Sugimoto T, Sawada S, Kinoshita T (2004).
Subtle disruption of the middle cerebellar peduncles in
patients with schizophrenia. Neuropsychobiology 50,
119–123.
Okugawa G, Nobuhara K, Sugimoto T, Kinoshita T (2005).
Diffusion tensor imaging study of the middle cerebellar
peduncles in patients with schizophrenia. Cerebellum 4,
123–127.
Ortakov V, Mancevski B, Keilp J, Oppenheim S, Dwork AJ
(1999). Application of cognitive scales to medical records of
schizophrenia inpatients. Schizophrenia Research 35,
131–140.
Ota M, Obata T, Akine Y, Ito H, Ikehira H, Asada T, Suhara
T (2006). Age-related degeneration of corpus callosum
measured with diffusion tensor imaging. Neuroimage 31,
1445–1452.
Pakkenberg B (1992). Stereological quantitation of human
brains from normal and schizophrenic individuals. Acta
Neurologica Scandinavica 137 (Suppl.), 20–33.
Pakkenberg B, Pelvig D, Marner L, Bundgaard MJ,
Gundersen HJ, Nyengaard JR, Regeur L (2003). Aging
and the human neocortex. Experimental Gerontology 38,
95–99.
Park HJ, Westin CF, Kubicki M, Maier SE, Niznikiewicz M,
Baer A, Frumin M, Kikinis R, Jolesz FA, McCarley RW,
Shenton ME (2004). White matter hemisphere
asymmetries in healthy subjects and in schizophrenia: a
diffusion tensor MRI study. Neuroimage 23, 213–223.
White matter and cognition 533

Peirce TR, Bray NJ, Williams NM, Norton N, Moskvina V,
Preece A, Haroutunian V, Buxbaum JD, Owen MJ,
O’Donovan MC (2006). Convergent evidence for
2k,3k-cyclic nucleotide 3k-phosphodiesterase as a possible
susceptibility gene for schizophrenia. Archives of General
Psychiatry 63, 18–24.
Perdahl E, Adolfsson R, Alafuzoff I, Albert KA, Nestler EJ,
Greengard P, Winblad B (1984). Synapsin I (protein I) in
different brain regions in senile dementia of Alzheimer
type and in multi-infarct dementia. Journal of Neural
Transmission 60, 133–141.
Perrone-Bizzozero NI, Sower AC, Bird ED, Benowitz LI,
Ivins KJ, Neve RL (1996). Levels of the growth-associated
protein GAP-43 are selectively increased in association
cortices in schizophrenia. Proceedings of the National
Academy of Sciences USA 93, 14182–14187.
Persson J, Lind J, Larsson A, Ingvar M, Cruts M, Van
Broeckhoven C, Adolfsson R, Nilsson LG, Nyberg L
(2006a). Altered brain white matter integrity in healthy
carriers of the APOE epsilon4 allele : a risk for AD?
Neurology 66, 1029–1033.
Persson J, Nyberg L, Lind J, Larsson A, Nilsson LG,
Ingvar M, Buckner RL (2006b). Structure-function
correlates of cognitive decline in aging. Cerebral Cortex 16,
907–915.
Peters A (1996). Age-related changes in oligodendrocytes in
monkey cerebral cortex. Journal of Comparative Neurology
371, 153–163.
Peters A, Josephson K, Vincent SL (1991). Effects of aging on
the neuroglial cells and pericytes within area 17 of the
rhesus monkey cerebral cortex. Anatomical Record 229,
384–398.
Peters A, Moss MB, Sethares C (2000). Effects of aging on
myelinated nerve fibers in monkey primary visual cortex.
Journal of Comparative Neurology 419, 364–376.
Peters A, Sethares C (2002). Aging and the myelinated fibers
in prefrontal cortex and corpus callosum of the monkey.
Journal of Comparative Neurology 442, 277–291.
Peters A, Sethares C (2003). Is there remyelination during
aging of the primate central nervous system? Journal of
Comparative Neurology 460, 238–254.
Peters A, Sethares C (2004). Oligodendrocytes, their
progenitors and other neuroglial cells in the aging primate
cerebral cortex. Cerebral Cortex 14, 995–1007.
PfefferbaumA, Adalsteinsson E, Sullivan EV (2005). Frontal
circuitry degradation marks healthy adult aging: evidence
from diffusion tensor imaging. Neuroimage 26, 891–899.
Pierpaoli C, Barnett A, Pajevic S, Chen R, Penix LR, Virta A,
Basser P (2001). Water diffusion changes in Wallerian
degeneration and their dependence on white matter
architecture. Neuroimage 13, 1174–1185.
Pierpaoli C, Basser PJ (1996). Toward a quantitative
assessment of diffusion anisotropy. Magnetic Resonance in
Medicine 36, 893–906.
Price G, Bagary MS, Cercignani M, Altmann DR, Ron MA
(2005). The corpus callosum in first episode schizophrenia:
a diffusion tensor imaging study. Journal of Neurology,
Neurosurgery and Psychiatry 76, 585–587.
Purohit DP, Perl DP, Haroutunian V, Powchik P, Davidson
M, Davis KL (1998). Alzheimer disease and related
neurodegenerative diseases in elderly patients with
schizophrenia: a postmortem neuropathologic study of
100 cases. Archives of General Psychiatry 55, 205–211.
Regeur L, Jensen GB, Pakkenberg H, Evans SM,
Pakkenberg B (1994). No global neocortical nerve cell
loss in brains from patients with senile dementia of
Alzheimer’s type. Neurobiology of Aging 15, 347–352.
Rivkin P, Kraut M, Barta P, Anthony J, Arria AM, Pearlson
G (2000). White matter hyperintensity volume in late-onset
and early-onset schizophrenia. International Journal of
Geriatric Psychiatry 15, 1085–1089.
Rosoklija G, Keilp JG, Toomayan G, Mancevski B,
Haroutunian V, Liu D, Malespina D, Hays AP, Sadiq S,
Latov N, Dwork AJ (2005). Altered Subicular MAP2
Immunoreactivity in Schizophrenia. Prilozi 26, 13–34.
Rosoklija G, Toomayan G, Ellis SP, Keilp J, Mann JJ,
Latov N, Hays AP, Dwork AJ (2000). Structural
abnormalities of subicular dendrites in subjects with
schizophrenia and mood disorders: preliminary findings.
Archives of General Psychiatry 57, 349–356.
Salat DH, Tuch DS, Greve DN, van der Kouwe AJ,
Hevelone ND, Zaleta AK, Rosen BR, Fischl B, Corkin S,
Rosas HD, Dale AM (2005). Age-related alterations in
white matter microstructure measured by diffusion tensor
imaging. Neurobiology of Aging 26, 1215–1227.
Sandell JH, Peters A (2002). Effects of age on the glial cells in
the rhesus monkey optic nerve. Journal of Comparative
Neurology 445, 13–28.
Sandell JH, Peters A (2003). Disrupted myelin and axon loss
in the anterior commissure of the aged rhesus monkey.
Journal of Comparative Neurology 466, 14–30.
Savitz J, Solms M, Ramesar R (2006). Apolipoprotein E
variants and cognition in healthy individuals: a
critical opinion. Brain Research. Brain Research Reviews 51,
125–135.
Scarmeas N, Habeck CG, Hilton J, Anderson KE, Flynn J,
Park A, Stern Y (2005). APOE related alterations in
cerebral activation even at college age. Journal of Neurology,
Neurosurgery and Psychiatry 76, 1440–1444.
Schachner M, Bartsch U (2000). Multiple functions of the
myelin-associated glycoprotein MAG (siglec-4a) in
formation and maintenance of myelin. Glia 29, 154–165.
Schmithorst VJ, Wilke M, Dardzinski BJ, Holland SK
(2005). Cognitive functions correlate with white matter
architecture in a normal pediatric population: a diffusion
tensor MRI study. Human Brain Mapping 26, 139–147.
Schneiderman J, Buchsbaum M, Haznedar M, Hazlett E,
Brickman A, Shihabuddin L, Brand J, Torosjan Y,
Newmark R, Tang C, et al. (in press). Diffusion tensor
anisotropy in adolescents and adults. Neurospsychobiology.
Schneiderman JS, Buchsbaum M, Haznedar M, Hazlett EA,
Shihabuddin L, Brand J, Brickman AM, Torosjan Y,
Newmark RE, Tang C, Aranowitz J, Paul-Odougard R
(2006). Age and diffusion tensor anisotropy in adolescent
and adult patients with schizophrenia. Society for
Neuroscience Abstracts. Abstract no. 188.186.
534 A. J. Dwork et al.

Shenkin SD, BastinME,Macgillivray TJ, Deary IJ, Starr JM,
Rivers CS, Wardlaw JM (2005). Cognitive correlates of
cerebral white matter lesions and water diffusion tensor
parameters in community-dwelling older people.
Cerebrovascular Diseases 20, 310–318.
Shenkin SD, Bastin ME, MacGillivray TJ, Deary IJ,
Starr JM, Wardlaw JM (2003). Childhood and current
cognitive function in healthy 80-year-olds: a DT-MRI
study. Neuroreport 14, 345–349.
Sloane JA, Hinman JD, Lubonia M, Hollander W,
Abraham CR (2003). Age-dependent myelin degeneration
and proteolysis of oligodendrocyte proteins is associated
with the activation of calpain-1 in the rhesus monkey.
Journal of Neurochemistry 84, 157–168.
Song SK, Kim JH, Lin SJ, Brendza RP, Holtzman DM (2004).
Diffusion tensor imaging detects age-dependent white
matter changes in a transgenic mouse model with amyloid
deposition. Neurobiology of Disease 15, 640–647.
Song SK, Sun SW, Ju WK, Lin SJ, Cross AH, Neufeld AH
(2003). Diffusion tensor imaging detects and differentiates
axon and myelin degeneration in mouse optic nerve after
retinal ischemia. Neuroimage 20, 1714–1722.
Song SK, Sun SW, Ramsbottom MJ, Chang C, Russell J,
Cross AH (2002). Dysmyelination revealed throughMRI as
increased radial (but unchanged axial) diffusion of water.
Neuroimage 17, 1429–1436.
Song SK, Yoshino J, Le TQ, Lin SJ, Sun SW, Cross AH,
Armstrong RC (2005). Demyelination increases radial
diffusivity in corpus callosum of mouse brain. Neuroimage
26, 132–140.
Steel RM, Bastin ME, McConnell S, Marshall I,
Cunningham-Owens DG, Lawrie SM, Johnstone EC,
Best JJ (2001). Diffusion tensor imaging (DTI) and proton
magnetic resonance spectroscopy (1H MRS) in
schizophrenic subjects and normal controls. Psychiatry
Research 106, 161–170.
Stern RA, Silva SG, Chaisson N, Evans DL (1996). Influence
of cognitive reserve on neuropsychological functioning in
asymptomatic human immunodeficiency virus-1 infection.
Archives of Neurology 53, 148–153.
Stern Y (2006). Cognitive reserve and Alzheimer
disease. Alzheimer Disease and Associated Disorders 20,
112–117.
Sullivan EV, PfefferbaumA (2003). Diffusion tensor imaging
in normal aging and neuropsychiatric disorders. European
Journal of Radiology 45, 244–255.
Sun SW, Liang HF, Le TQ, Armstrong RC, Cross AH,
Song SK (2006a). Differential sensitivity of in vivo and
ex vivo diffusion tensor imaging to evolving optic nerve
injury in mice with retinal ischemia. Neuroimage 32,
1195–1204.
Sun SW, Liang HF, Trinkaus K, Cross AH, Armstrong RC,
Song SK (2006b). Noninvasive detection of cuprizone
induced axonal damage and demyelination in the mouse
corpus callosum. Magnetic Resonance in Medicine 55,
302–308.
Sun SW, Song SK, Harms MP, Lin SJ, Holtzman DM,
Merchant KM, Kotyk JJ (2005). Detection of
age-dependent brain injury in a mouse model of brain
amyloidosis associated with Alzheimer’s disease using
magnetic resonance diffusion tensor imaging. Experimental
Neurology 191, 77–85.
Sun Z, Wang F, Cui L, Breeze J, Du X, Wang X, Cong Z,
Zhang H, Li B, Hong N, Zhang D (2003). Abnormal
anterior cingulum in patients with schizophrenia: a
diffusion tensor imaging study. Neuroreport 14, 1833–1836.
Suzuki Y, Matsuzawa H, Kwee IL, Nakada T (2003).
Absolute eigenvalue diffusion tensor analysis for
human brain maturation. NMR in Biomedicine 16,
257–260.
Svennerholm L, Bostrom K, Helander CG, Jungbjer B
(1991). Membrane lipids in the aging human brain.
Journal of Neurochemistry 56, 2051–2059.
Svennerholm L, Bostrom K, Jungbjer B (1997). Changes in
weight and compositions of major membrane components
of human brain during the span of adult human life of
Swedes. Acta Neuropathologica 94, 345–352.
Symonds LL, Olichney JM, Jernigan TL, Corey-Bloom J,
Healy JF, Jeste DV (1997). Lack of clinically significant
gross structural abnormalities in MRIs of older patients
with schizophrenia and related psychoses. Journal of
Neuropsychiatry and Clinical Neurosciences 9, 251–258.
Szeszko PR, Ardekani BA, Ashtari M, Kumra S, Robinson
DG, Sevy S, Gunduz-Bruce H, Malhotra AK, Kane JM,
Bilder RM, Lim KO (2005). White matter abnormalities in
first-episode schizophrenia or schizoaffective disorder: a
diffusion tensor imaging study. American Journal of
Psychiatry 162, 602–605.
Tang Y, Nyengaard JR, Pakkenberg B, Gundersen HJ (1997).
Age-induced white matter changes in the human brain: a
stereological investigation. Neurobiology of Aging 18,
609–615.
Tcherepanov AA, Sokolov BP (1997). Age-related
abnormalities in expression of mRNAs encoding synapsin
1A, synapsin 1B, and synaptophysin in the temporal cortex
of schizophrenics. Journal of Neuroscience Research 49,
639–644.
Terry RD, Masliah E, Salmon DP, Butters N, DeTeresa R,
Hill R, Hansen LA, Katzman R (1991). Physical basis of
cognitive alterations in Alzheimer’s disease: synapse loss
is the major correlate of cognitive impairment. Annals of
Neurology 30, 572–580.
Tkachev D, Mimmack ML, Ryan MM, Wayland M,
FreemanT, Jones PB, StarkeyM,WebsterMJ, YolkenRH,
Bahn S (2003). Oligodendrocyte dysfunction in
schizophrenia and bipolar disorder. Lancet 362, 798–805.
Toosy AT, Werring DJ, Orrell RW, Howard RS, King MD,
Barker GJ, Miller DH, Thompson AJ (2003). Diffusion
tensor imaging detects corticospinal tract involvement
at multiple levels in amyotrophic lateral sclerosis.
Journal of Neurology, Neurosurgery and Psychiatry 74,
1250–1257.
Tyszka JM, Readhead C, Bearer EL, Pautler RG, Jacobs RE
(2006). Statistical diffusion tensor histology reveals
regional dysmyelination effects in the shiverer mouse
mutant. Neuroimage 29, 1058–1065.
White matter and cognition 535

Uranova N, Orlovskaya D, Vikhreva O, Zimina I,
Kolomeets N, Vostrikov V, Rachmanova V (2001).
Electron microscopy of oligodendroglia in severe mental
illness. Brain Research Bulletin 55, 597–610.
Uranova NA, Vostrikov VM, Orlovskaya DD, Rachmanova
VI (2004). Oligodendroglial density in the prefrontal cortex
in schizophrenia and mood disorders: a study from the
Stanley Neuropathology Consortium. Schizophrenia
Research 67, 269–275.
Vawter MP, Howard AL, Hyde TM, Kleinman JE, Freed WJ
(1999). Alterations of hippocampal secreted N-CAM in
bipolar disorder and synaptophysin in schizophrenia.
Molecular Psychiatry 4, 467–475.
Walss-Bass C, Liu W, Lew DF, Villegas R, Montero P,
Dassori A, Leach RJ, Almasy L, Escamilla M,
Raventos H (2006). A novel missense mutation in the
transmembrane domain of Neuregulin 1 is associated
with schizophrenia. Biological Psychiatry 60, 548–553.
Walterfang M, Wood SJ, Velakoulis D, Pantelis C
(2006). Neuropathological, neurogenetic and
neuroimaging evidence for white matter pathology in
schizophrenia. Neuroscience and Biobehavioral Reviews 30,
918–948.
Wang DS, Bennett DA, Mufson EJ, Mattila P, Cochran E,
Dickson DW (2004a). Contribution of changes in ubiquitin
and myelin basic protein to age-related cognitive decline.
Neuroscience Research 48, 93–100.
Wang F, Sun Z, Cui L, Du X, Wang X, Zhang H, Cong Z,
Hong N, Zhang D (2004b). Anterior cingulum
abnormalities in male patients with schizophrenia
determined through diffusion tensor imaging. American
Journal of Psychiatry 161, 573–575.
Wang F, Sun Z, Du X, Wang X, Cong Z, Zhang H, Zhang D,
Hong N (2003). A diffusion tensor imaging study of
middle and superior cerebellar peduncle in male patients
with schizophrenia. Neuroscience Letters 348, 135–138.
Weinberger DR, Berman KF, Zec RF (1986). Physiologic
dysfunction of dorsolateral prefrontal cortex in
schizophrenia. I. Regional cerebral blood flow evidence.
Archives of General Psychiatry 43, 114–124.
Wender M, Adamczewska-Goncerzewicz Z, Dorszewska J
(1991). Myelin proteins in aging human brain. Molecular
and Chemical Neuropathology 14, 1–10.
Wiggins RC, Gorman A, Rolsten C, Samorajski T,
Ballinger Jr. WE, Freund G (1988). Effects of aging and
alcohol on the biochemical composition of histologically
normal human brain. Metabolic Brain Disease 3, 67–80.
Wolkin A, Choi SJ, Szilagyi S, Sanfilipo M, Rotrosen JP,
Lim KO (2003). Inferior frontal white matter anisotropy
and negative symptoms of schizophrenia: a diffusion
tensor imaging study. American Journal of Psychiatry 160,
572–574.
Wu JI, Reed RB, Grabowski PJ, Artzt K (2002). Function of
quaking in myelination: regulation of alternative splicing.
Proceedings of the National Academy of Sciences USA 99,
4233–4238.
Yakovlev P, Lecours A (1967). The myelogenetic cycles of
regional maturation of the brain. In: Minowski, A (Ed.),
Regional Development of the Brain in Early Life (pp. 3–70).
Philadelphia: F.A. Davis.
Young KA, Manaye KF, Liang C, Hicks PB, German DC
(2000). Reduced number of mediodorsal and anterior
thalamic neurons in schizophrenia. Biological Psychiatry 47,
944–953.
536 A. J. Dwork et al.
Related Documents











