VP7 gene polymorphism of serotype G9 rotavirus strains and its impact on G genotype determination by PCR Norma Santos a, *, Eduardo M. Volota ˜o a , Caroline C. Soares a , Maria Carolina M. Albuquerque a , Fabiano M. da Silva a , Vladimir Chizhikov b , Yasutaka Hoshino c a Departamento de Virologia, Instituto de Microbiologia, Universidade Federal do Rio de Janeiro, Cidade Universita ´ria, CCS-Bl. I, Ilha do Funda ˜o, Rio de Janeiro, RJ 21.941-590, Brazil b Center for Biologics Evaluation and Research, Food and Drug Administration, Kensington, MD 20895, USA c Laboratory of Infectious Diseases, National Institutes of Health, Bethesda, MD 20892, USA Received 24 January 2002; received in revised form 24 May 2002; accepted 24 May 2002 Abstract Rotaviruses are the single most important etiologic agents of severe diarrhea of infants and young children worldwide. Surveillance of rotavirus serotypes/genotypes (both VP7[G] and VP4[P]) is in progress globally in which polymerase chain reaction (PCR) has been the assay of choice. We investigated polymorphism of the VP7 gene of serotype G9 rotavirus strains and its impact on the determination of VP7 gene genotype by PCR assay. By VP7 gene sequence analysis, we and others have previously shown that the G9 rotavirus strains belong to one of three VP7 gene lineages. By PCR assay using three different sets of commonly used primers specific for G1-4, 8 and 9, 23 Brazilian G9 strains and 5 well-characterized prototype G9 strains which collectively represented all three VP7 gene lineages were typed as: (i) G3; (ii) G4; (iii) G9; (iv) G3 and G9; or (v) G9 and G4 depending on a primer pool employed. This phenomenon appeared to be due to: (i) a VP7 gene lineage-specific polymorphism, more specifically mutation(s) in the primer binding region of the VP7 gene of G9 strain; and (ii) the magnitude of difference in nucleotide homology at respective primer binding site between homotypic (G9) and heterotypic (G3 or G4) primers present in a primer pool employed. # 2002 Elsevier Science B.V. All rights reserved. Keywords: Rotavirus; PCR G genotyping; VP7 gene polymorphism; Gastroenteritis 1. Introduction It has been well established that virtually every child becomes infected with a rotavirus at least once by 3 years of age (Bern and Glass, 1994; Kapikian et al., 2001). The consequences of rotavirus infection, how- ever, are markedly different depending on the economic status of each country. In the U.S., for example, rotavirus gastroenteritis is estimated to be responsible for more than 500,000 physician visits, 50,000 hospita- lizations, approximately 20 deaths and over $1 billion of societal expenses each year (Parashar et al., 1998a,b; Tucker et al., 1998). In developing countries, estimates indicate that rotavirus diarrhea kill 1600 /2400 children/ day, or 600,000 /870,000 children/year (Glass et al., 1999). Thus, the development and universal implemen- tation of a safe and effective rotavirus vaccine is a global public health goal (Bresee et al., 1999; Kapikian et al., 2001). Rotavirus is a member of the Reoviridae family which includes seven groups (rotavirus A /G). A complete virus particle possesses 11 double-stranded (ds) RNA segments surrounded by three concentric protein layers. The outermost layer is composed of two proteins, VP7 (which defines G serotypes) and VP4 (which defines P serotypes) that independently evoke neutralizing anti- bodies. There are 14 G serotypes, however, types G1 / G4 are the most common globally and, therefore, are the targets for current vaccine development strategies (Hoshino and Kapikian, 2000). More recently, various epidemiologic studies have shown the emergence/re- PII of original article: S 0 1 6 8 - 1 7 0 2 ( 0 2 ) 0 0 1 0 6 - 5 * Corresponding author. Tel.: /55-21-260-9311; fax: /55-21-560- 8344. E-mail address: [email protected] (N. Santos). Virus Research 93 (2003) 127 /138 www.elsevier.com/locate/virusres 0168-1702/02/$ - see front matter # 2002 Elsevier Science B.V. All rights reserved. doi:10.1016/S0168-1702(02)00318-0

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

VP7 gene polymorphism of serotype G9 rotavirus strains and itsimpact on G genotype determination by PCR�
Norma Santos a,*, Eduardo M. Volotao a, Caroline C. Soares a, Maria CarolinaM. Albuquerque a, Fabiano M. da Silva a, Vladimir Chizhikov b, Yasutaka Hoshino c
a Departamento de Virologia, Instituto de Microbiologia, Universidade Federal do Rio de Janeiro, Cidade Universitaria, CCS-Bl. I, Ilha do Fundao, Rio
de Janeiro, RJ 21.941-590, Brazilb Center for Biologics Evaluation and Research, Food and Drug Administration, Kensington, MD 20895, USA
c Laboratory of Infectious Diseases, National Institutes of Health, Bethesda, MD 20892, USA
Received 24 January 2002; received in revised form 24 May 2002; accepted 24 May 2002
Abstract
Rotaviruses are the single most important etiologic agents of severe diarrhea of infants and young children worldwide.
Surveillance of rotavirus serotypes/genotypes (both VP7[G] and VP4[P]) is in progress globally in which polymerase chain reaction
(PCR) has been the assay of choice. We investigated polymorphism of the VP7 gene of serotype G9 rotavirus strains and its impact
on the determination of VP7 gene genotype by PCR assay. By VP7 gene sequence analysis, we and others have previously shown
that the G9 rotavirus strains belong to one of three VP7 gene lineages. By PCR assay using three different sets of commonly used
primers specific for G1-4, 8 and 9, 23 Brazilian G9 strains and 5 well-characterized prototype G9 strains which collectively
represented all three VP7 gene lineages were typed as: (i) G3; (ii) G4; (iii) G9; (iv) G3 and G9; or (v) G9 and G4 depending on a
primer pool employed. This phenomenon appeared to be due to: (i) a VP7 gene lineage-specific polymorphism, more specifically
mutation(s) in the primer binding region of the VP7 gene of G9 strain; and (ii) the magnitude of difference in nucleotide homology at
respective primer binding site between homotypic (G9) and heterotypic (G3 or G4) primers present in a primer pool employed.
# 2002 Elsevier Science B.V. All rights reserved.
Keywords: Rotavirus; PCR G genotyping; VP7 gene polymorphism; Gastroenteritis
1. Introduction
It has been well established that virtually every child
becomes infected with a rotavirus at least once by 3
years of age (Bern and Glass, 1994; Kapikian et al.,
2001). The consequences of rotavirus infection, how-
ever, are markedly different depending on the economic
status of each country. In the U.S., for example,
rotavirus gastroenteritis is estimated to be responsible
for more than 500,000 physician visits, 50,000 hospita-
lizations, approximately 20 deaths and over $1 billion of
societal expenses each year (Parashar et al., 1998a,b;
Tucker et al., 1998). In developing countries, estimates
indicate that rotavirus diarrhea kill 1600�/2400 children/
day, or 600,000�/870,000 children/year (Glass et al.,
1999). Thus, the development and universal implemen-
tation of a safe and effective rotavirus vaccine is a global
public health goal (Bresee et al., 1999; Kapikian et al.,
2001).
Rotavirus is a member of the Reoviridae family which
includes seven groups (rotavirus A�/G). A complete
virus particle possesses 11 double-stranded (ds) RNA
segments surrounded by three concentric protein layers.
The outermost layer is composed of two proteins, VP7
(which defines G serotypes) and VP4 (which defines P
serotypes) that independently evoke neutralizing anti-
bodies. There are 14 G serotypes, however, types G1�/
G4 are the most common globally and, therefore, are
the targets for current vaccine development strategies
(Hoshino and Kapikian, 2000). More recently, various
epidemiologic studies have shown the emergence/re-
�PII of original article: S 0 1 6 8 - 1 7 0 2 ( 0 2 ) 0 0 1 0 6 - 5
* Corresponding author. Tel.: �/55-21-260-9311; fax: �/55-21-560-
8344.
E-mail address: [email protected] (N. Santos).
Virus Research 93 (2003) 127�/138
www.elsevier.com/locate/virusres
0168-1702/02/$ - see front matter # 2002 Elsevier Science B.V. All rights reserved.
doi:10.1016/S0168-1702(02)00318-0

emergence of rotavirus serotype G9 as an important
human pathogen worldwide and, it is likely to become
the fifth globally important strain (Urasawa et al., 1992;
Ramachandran et al., 1996, 1998; Cunliffe et al., 1999;Unicomb et al., 1999; Bon et al., 2000; Cubitt et al.,
2000; Griffin et al., 2000; Maneekarn and Ushijima,
2000; O’Halloran et al., 2000; Oka et al., 2000; Palombo
et al., 2000; Widdowson et al., 2000; Adah et al., 2001;
Araujo et al., 2001; Santos et al., 2001; Van Man et al.,
2001; Zhou et al., 2001; Bok et al., 2001). In addition,
sequence analysis of the VP7 gene of various G9
rotavirus isolates carried out by us and others haveshown the existence of at least three phylogenetic VP7
gene lineages (I�/III) (Ramachandran et al., 2000; Oka et
al., 2000; Bok et al., 2001; Santos et al., 2001; Zhou et
al., 2001).
Polymerase chain reaction (PCR) methodologies for
rotavirus genotyping were developed over a decade ago.
Because available VP7 gene sequences were rather
limited then, selection of type-specific primers for Ggenotyping was also limited. Moreover, since rotaviruses
were demonstrated to undergo constant genetic varia-
tion via sequential point mutations or ‘antigenic drift’,
genetic reassortment or ‘antigenic shift’, genomic re-
arrangement or intragenic recombination (Gouvea and
Brantly, 1995; Desselberger, 1996; Iturriza-Gomara et
al., 2001; Suzuki et al., 1998), the emergence of strains
that could not be easily identified by this method wasnot completely unexpected. Since neutralizing antibo-
dies and their type specificities appear to play an
important role in protection against rotavirus disease
and/or infection (Hoshino and Kapikian, 2000), epide-
miologic surveillance of rotavirus serotypes/genotypes is
underway throughout the world. However, a potential
impact of rotavirus genome polymorphism on G
genotype identification by PCR has just begun to befully explored (Nakagomi et al., 1991; Adah et al., 1997;
Iturriza-Gomara et al., 2000; Cunliffe et al., 2001a).
Here we describe: (i) a genomic variation of the VP7
gene of rotavirus serotype G9 strains isolated in Brazil
in comparison with that of G9 strains isolated in other
parts of the world which collectively represent all three
phylogenetic VP7 gene lineages; and (ii) the impact of
such variation on G genotype identification by PCR.
2. Materials and methods
2.1. Viruses
Twenty-three G9 strains were derived from children
under 5 years of age with acute diarrhea detected during
epidemiologic studies in Brazil from 1997 to 1999 aspreviously reported (Santos et al., 2001). The prototype
G9 strains WI61 (Clark et al., 1987), F45 (Akatani and
Ikegami, 1987), AU32 (Nakagomi et al., 1990), 116E
(Das et al., 1994) and US1205 (Ramachandran et al.,
1998) belonged to the rotavirus collection of the
Laboratory of Infectious Diseases, National Institutes
of Health, Bethesda, USA.
2.2. RNA extraction and RT-PCR
Rotavirus dsRNA was extracted from stool samples
or infected cell culture lysates by use of guanidine
isothiocyanate method (Santos and Gouvea, 1994) or
TRIzol method (Life Technologies Inc., Grand Island,
NY) and precipitated with ethanol. Reverse transcrip-tion (RT)-PCR for determination of rotavirus G types
was performed by using primers specific for genotypes
G1 to G6, G8 to G10 and G11 (Gouvea et al., 1990;
Taniguchi et al., 1992; Isegawa et al., 1993; Das et al.,
1994; Gouvea et al., 1994). Four different sets of G
typing primers were used (Fig. 1): (1) H1 pool which
contained the original set of primers described by
Gouvea et al. (1990), specific for G1 to G4, G8 andG9 genotypes; (2) H2 pool in which Gouvea’s primers
aET3, aDT4 and aAT8 (G3-, G4-, and G8-specific,
respectively) were replaced by S3, S4, and S8 primers
described by Taniguchi et al. (1992); (3) C pool reported
by Das et al. (1994), specific for G1 to G4 and G9
genotypes; and (4) A pool described by Gouvea et al.
(1994) which contained primers specific for G5, G6, G8,
and G11 genotypes in which the ET10 primer (G10-specific) was replaced by G10 primer described by
Isegawa et al. (1993). The dsRNA samples were
subjected to one cycle of reverse transcription (42 8C,
45 min) followed by 30 cycles of PCR. Each PCR cycle
for G or P genotyping contained steps of 1 min at
94 8C, 2 min at 50 8C and 3 min at 72 8C.
2.3. Sequence analysis
The VP7 gene of six Brazilian G9 strains (R44, R136,
R143, R146, R160 and R172) was sequenced and then
analyzed by using the DNASTAR program. The VP7 gene
sequence of selected G3, G4 and other G9 rotavirus
strains was obtained from GenBank.
2.4. Neutralization assay
Antigenic characterization of selected G9 isolates was
performed by a 60% plaque reduction neutralization
(PRN) assay using guinea pig hyperimmune antiserum
raised against each of G1 to G14 rotavirus prototype
strain as described previously (Wyatt et al., 1982).
2.5. Analysis of rotavirus dsRNA by polyacrylamide gel
electrophoresis
Rotavirus genomic dsRNA was extracted using
standard phenol�/chloroform methods and precipitated
Erratum128

with ethanol. Each RNA sample was analyzed on a
standard discontinuous 10% acrylamide slab gel with a
3.5% acrylamide stacking gel. After electrophoresis,dsRNAs were visualized by staining gel with silver
nitrate (Herring et al., 1982).
3. Results
3.1. G genotype variability displayed by G9 rotavirus
strains against three different primer sets
When analyzed by RT-PCR, each of 23 Brazilian G9
strains displayed different reactivity pattern against
primer pool H1, H2 or C and fell into one of three
distinct reactivity groups (Table 1). One group consist-
ing of 17 strains (represented by R44 strain) was typed
as G3 versus (vs) H1 pool, G4 vs H2 pool and G9 vs C
pool. Second group (strain R179) was typed as G3 vs H1
pool, G4 and 9 vs H2 pool and G9 vs C pool. Thirdgroup of five strains (represented by R143 strain) were
typed as G3 and 9 vs H1 pool, G9 and 4 vs H2 pool and
G9 vs C pool. None of the samples yielded positive
reactivity with A pool (data not shown) and only the C
pool provided a reliable detection of the G9 strains.
A panel of five prototype G9 rotavirus strains (the US
strains US1205 and WI61, the Japanese strains F45 and
AU32, and the Indian strain 116E) was also analyzed byRT-PCR against the same four sets of G typing primers
(Table 1). Strains WI61, F45 and AU32 were typed as
G9 vs H1, H2 or C pool. Strains US1205 and 116E
showed reactivity pattern similar to that of R143 strain
against three primer sets (H1, H2 and C). The A pool
did not produce any reactivity against any of the fiveprototype G9 strains (data not shown).
3.2. Reactivity of selected G9 rotavirus strains vs G3-,
G4- or G9-specific primer in H1 pool
Six Brazilian strains (R44, R136, R172, R143, R146
and R160) and one US strain (US1205), belonging to
the VP7 gene lineage III, had identical five nucleotide
mismatches (76.2% nucleotide [nt] homology) at theaET3 primer binding site (nt 689�/709) and all of them
were amplified (Table 2). Strain 116E belonging to
lineage II which had six nucleotide mismatches (71.4%
nucleotide homology) was amplified weakly with the
primer. The primer binding site of strain WI61, F45 or
AU32 belonging to lineage I had six identical nucleotide
mismatches (71.4% homology) different from those
found in lineage II virus and was not primed with theaET primer. All 11 G9 rotavirus strains analyzed had 9�/
12 nucleotide mismatches (36.8�/52.6% homology) at the
aDT4 primer binding site and none were amplified
(Table 2).
At the aFT9-specific binding site (nt 757�/776) strains
R143, R146, R160 and US1205, which presented three
or four mismatches (85.0 and 80.0% nucleotide homol-
ogy, respectively), were weakly amplified (Table 2). TheBrazilian strain R44, R136 or R172 belonging to lineage
III which presented four identical mismatches (80.0%
nucleotide homology) was not amplified by primer
Fig. 1. The position and directions of primers in H-1, H-2, C or A primer pool used in this study in relation to the schematic VP7-encoding gene.
Variable regions A�/F on the gene are denoted by black boxes. 1, Primers described by Gouvea et al. (1990); 2, Primers described by Gouvea et al.
(1990) and Taniguchi et al. (1992); 3, Primers described by Das et al. (1994); 4, Primers described by Gouvea et al. (1994) and Isegawa et al. (1993).
Erratum 129
aFT9. The Indian strain 116E belonging to lineage II
that had a set of four mismatches different from those
found in lineage III viruses was amplified. Each of three
prototype strains WI61, AU32, and F45 belonging to
lineage I showed perfect matches (100.0% nucleotide
homology) at the primer binding site.
3.3. Reactivity of selected G9 strains vs G3-, G4- or G9-
specific primer in H2 pool
With primer S3 (nt 481�/498), G3-specific, each of 11
G9 strains analyzed presented eight or nine mismatches
(55.6 and 50.0% nucleotide homology, respectively) and
none were amplified with this primer (Table 3). All seven
G9 strains belonging to lineage III which had identical
five nucleotide mismatches (75% homology) at the S4
primer binding site (nt 669�/688) were amplified (Table3). Although strain WI61, F45 or AU32 (lineage I) had
only four or five nucleotide mismatches (75�/80%
homology), none were amplified with the primer. Strain
116E (lineage II) which had four nucleotide mismatches
identical to those of F45 was weakly amplified.
Strains R44, R136 and R172 (lineage III) had four
identical nucleotide mismatches (80% homology) at the
aFT9 primer binding site and none were amplified.
Strains R143, R146, R160 and US1205 (lineage III) had
three nucleotide mismatches (85% homology) and wereamplified. Strains WI61, F45 and AU32 (lineage I)
which presented a 100% nucleotide homology at the
primer binding site were amplified, as expected. Strain
116E (lineage II) which had four nucleotide mismatches
(80% homology) different from those of lineage III
viruses and was still amplified.
3.4. Reactivity of selected G9 strains vs G3-, G4- or G9-
specific primer in C pool
All 11 G9 strains analyzed presented 7�/9 nucleotide
mismatches (50.0�/61.1% homology) with primer 9T-3P
(nt 484�/503) and none were amplified (Table 4). Strains
Table 1
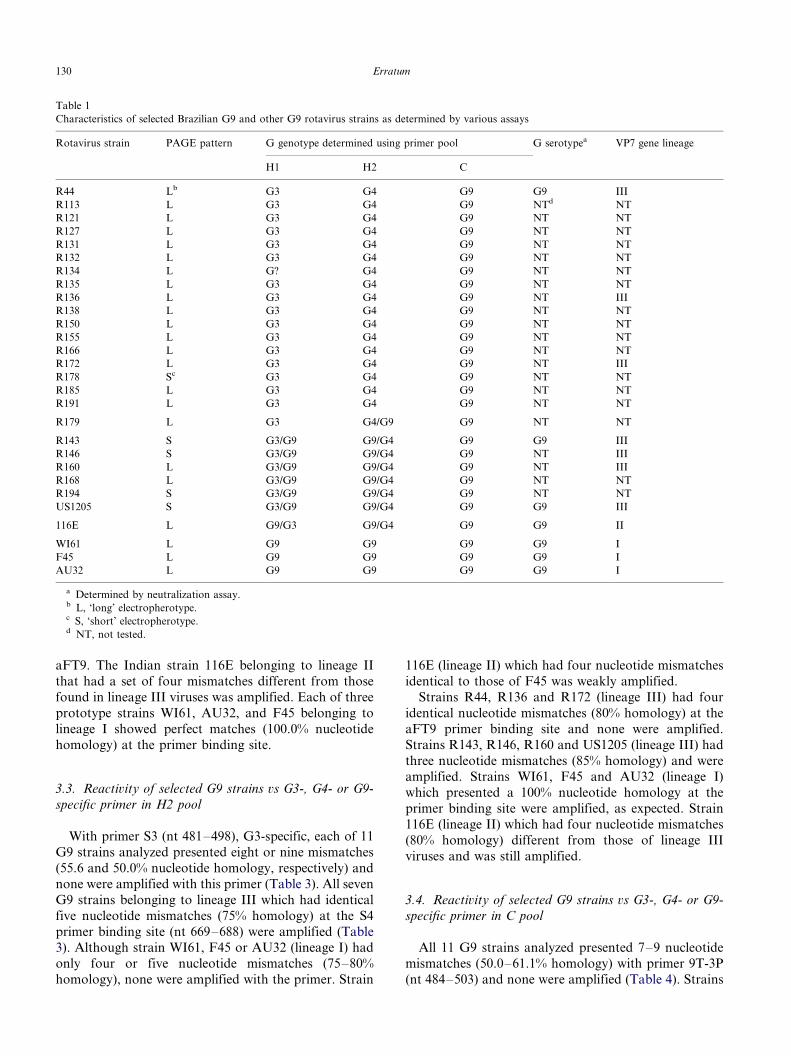
Characteristics of selected Brazilian G9 and other G9 rotavirus strains as determined by various assays
Rotavirus strain PAGE pattern G genotype determined using primer pool G serotypea VP7 gene lineage
H1 H2 C
R44 Lb G3 G4 G9 G9 III
R113 L G3 G4 G9 NTd NT
R121 L G3 G4 G9 NT NT
R127 L G3 G4 G9 NT NT
R131 L G3 G4 G9 NT NT
R132 L G3 G4 G9 NT NT
R134 L G? G4 G9 NT NT
R135 L G3 G4 G9 NT NT
R136 L G3 G4 G9 NT III
R138 L G3 G4 G9 NT NT
R150 L G3 G4 G9 NT NT
R155 L G3 G4 G9 NT NT
R166 L G3 G4 G9 NT NT
R172 L G3 G4 G9 NT III
R178 Sc G3 G4 G9 NT NT
R185 L G3 G4 G9 NT NT
R191 L G3 G4 G9 NT NT
R179 L G3 G4/G9 G9 NT NT
R143 S G3/G9 G9/G4 G9 G9 III
R146 S G3/G9 G9/G4 G9 NT III
R160 L G3/G9 G9/G4 G9 NT III
R168 L G3/G9 G9/G4 G9 NT NT
R194 S G3/G9 G9/G4 G9 NT NT
US1205 S G3/G9 G9/G4 G9 G9 III
116E L G9/G3 G9/G4 G9 G9 II
WI61 L G9 G9 G9 G9 I
F45 L G9 G9 G9 G9 I
AU32 L G9 G9 G9 G9 I
a Determined by neutralization assay.b L, ‘long’ electropherotype.c S, ‘short’ electropherotype.d NT, not tested.
Erratum130
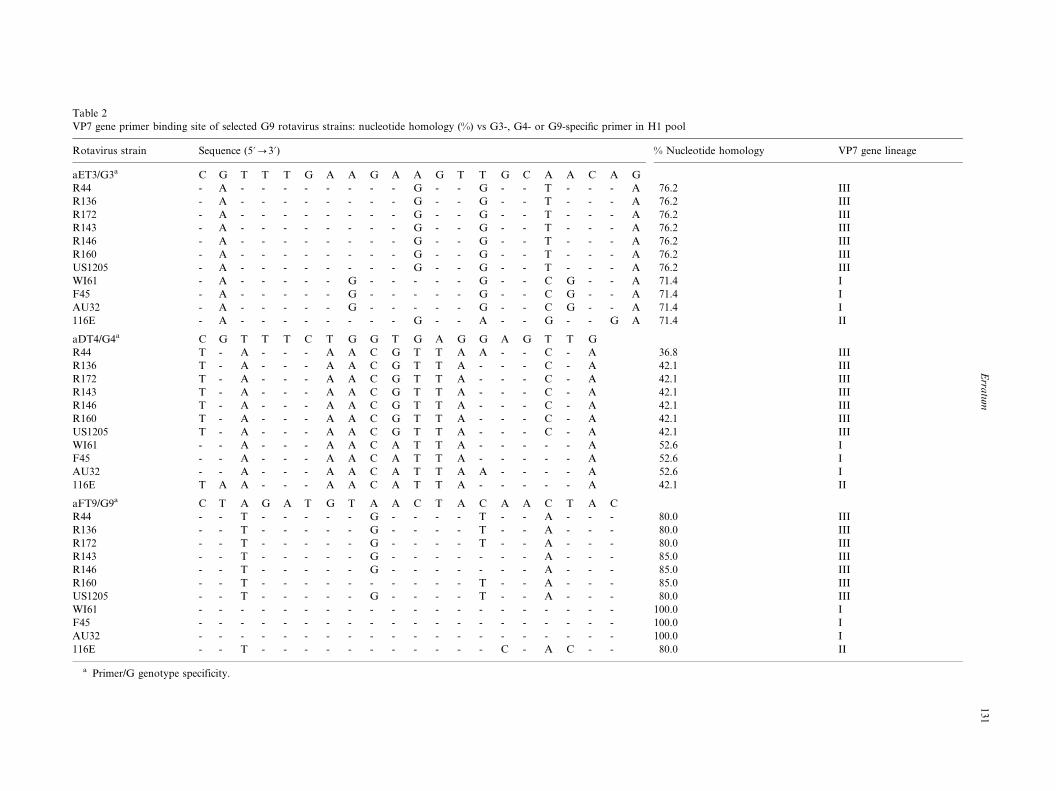
Table 2
VP7 gene primer binding site of selected G9 rotavirus strains: nucleotide homology (%) vs G3-, G4- or G9-specific primer in H1 pool
Rotavirus strain Sequence (5?0/3?) % Nucleotide homology VP7 gene lineage
aET3/G3a C G T T T G A A G A A G T T G C A A C A G
R44 - A - - - - - - - - G - - G - - T - - - A 76.2 III
R136 - A - - - - - - - - G - - G - - T - - - A 76.2 III
R172 - A - - - - - - - - G - - G - - T - - - A 76.2 III
R143 - A - - - - - - - - G - - G - - T - - - A 76.2 III
R146 - A - - - - - - - - G - - G - - T - - - A 76.2 III
R160 - A - - - - - - - - G - - G - - T - - - A 76.2 III
US1205 - A - - - - - - - - G - - G - - T - - - A 76.2 III
WI61 - A - - - - - G - - - - - G - - C G - - A 71.4 I
F45 - A - - - - - G - - - - - G - - C G - - A 71.4 I
AU32 - A - - - - - G - - - - - G - - C G - - A 71.4 I
116E - A - - - - - - - - G - - A - - G - - G A 71.4 II
aDT4/G4a C G T T T C T G G T G A G G A G T T G
R44 T - A - - - A A C G T T A A - - C - A 36.8 III
R136 T - A - - - A A C G T T A - - - C - A 42.1 III
R172 T - A - - - A A C G T T A - - - C - A 42.1 III
R143 T - A - - - A A C G T T A - - - C - A 42.1 III
R146 T - A - - - A A C G T T A - - - C - A 42.1 III
R160 T - A - - - A A C G T T A - - - C - A 42.1 III
US1205 T - A - - - A A C G T T A - - - C - A 42.1 III
WI61 - - A - - - A A C A T T A - - - - - A 52.6 I
F45 - - A - - - A A C A T T A - - - - - A 52.6 I
AU32 - - A - - - A A C A T T A A - - - - A 52.6 I
116E T A A - - - A A C A T T A - - - - - A 42.1 II
aFT9/G9a C T A G A T G T A A C T A C A A C T A C
R44 - - T - - - - - G - - - - T - - A - - - 80.0 III
R136 - - T - - - - - G - - - - T - - A - - - 80.0 III
R172 - - T - - - - - G - - - - T - - A - - - 80.0 III
R143 - - T - - - - - G - - - - - - - A - - - 85.0 III
R146 - - T - - - - - G - - - - - - - A - - - 85.0 III
R160 - - T - - - - - - - - - - T - - A - - - 85.0 III
US1205 - - T - - - - - G - - - - T - - A - - - 80.0 III
WI61 - - - - - - - - - - - - - - - - - - - - 100.0 I
F45 - - - - - - - - - - - - - - - - - - - - 100.0 I
AU32 - - - - - - - - - - - - - - - - - - - - 100.0 I
116E - - T - - - - - - - - - - - C - A C - - 80.0 II
a Primer/G genotype specificity.
Erra
tum
13
1
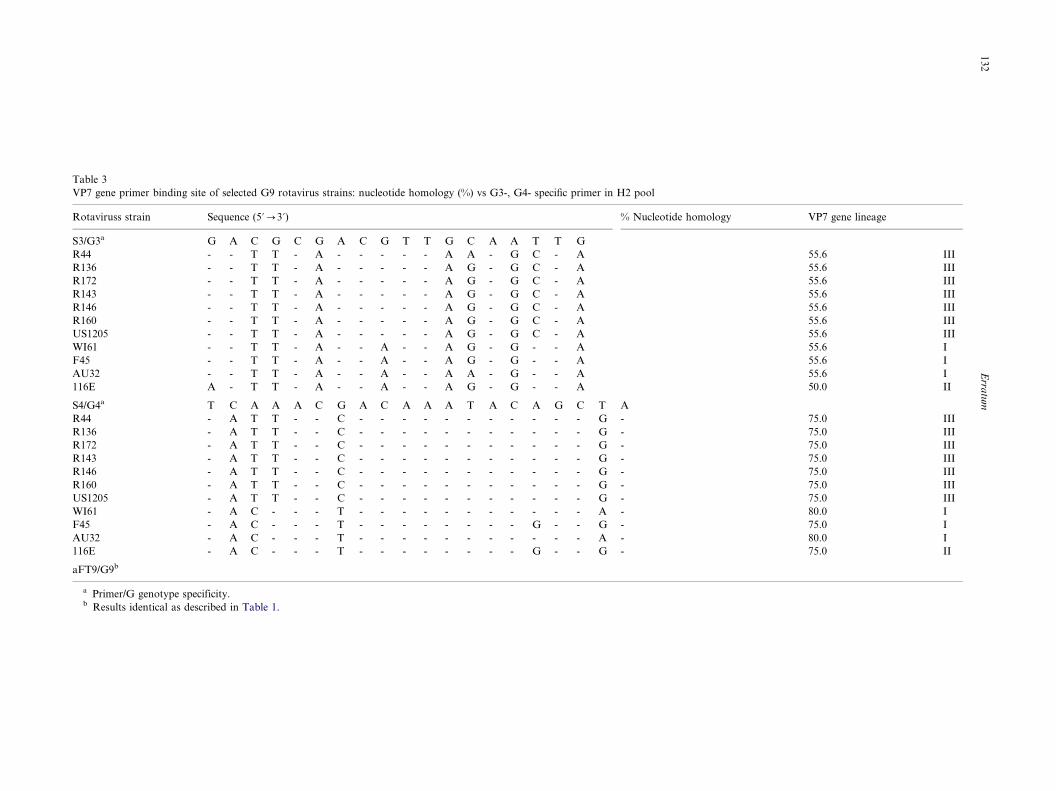
Table 3
VP7 gene primer binding site of selected G9 rotavirus strains: nucleotide homology (%) vs G3-, G4- specific primer in H2 pool
Rotaviruss strain Sequence (5?0/3?) % Nucleotide homology VP7 gene lineage
S3/G3a G A C G C G A C G T T G C A A T T G
R44 - - T T - A - - - - - A A - G C - A 55.6 III
R136 - - T T - A - - - - - A G - G C - A 55.6 III
R172 - - T T - A - - - - - A G - G C - A 55.6 III
R143 - - T T - A - - - - - A G - G C - A 55.6 III
R146 - - T T - A - - - - - A G - G C - A 55.6 III
R160 - - T T - A - - - - - A G - G C - A 55.6 III
US1205 - - T T - A - - - - - A G - G C - A 55.6 III
WI61 - - T T - A - - A - - A G - G - - A 55.6 I
F45 - - T T - A - - A - - A G - G - - A 55.6 I
AU32 - - T T - A - - A - - A A - G - - A 55.6 I
116E A - T T - A - - A - - A G - G - - A 50.0 II
S4/G4a T C A A A C G A C A A A T A C A G C T A
R44 - A T T - - C - - - - - - - - - - - G - 75.0 III
R136 - A T T - - C - - - - - - - - - - - G - 75.0 III
R172 - A T T - - C - - - - - - - - - - - G - 75.0 III
R143 - A T T - - C - - - - - - - - - - - G - 75.0 III
R146 - A T T - - C - - - - - - - - - - - G - 75.0 III
R160 - A T T - - C - - - - - - - - - - - G - 75.0 III
US1205 - A T T - - C - - - - - - - - - - - G - 75.0 III
WI61 - A C - - - T - - - - - - - - - - - A - 80.0 I
F45 - A C - - - T - - - - - - - - G - - G - 75.0 I
AU32 - A C - - - T - - - - - - - - - - - A - 80.0 I
116E - A C - - - T - - - - - - - - G - - G - 75.0 II
aFT9/G9b
a Primer/G genotype specificity.b Results identical as described in Table 1.
Erra
tum
13
2
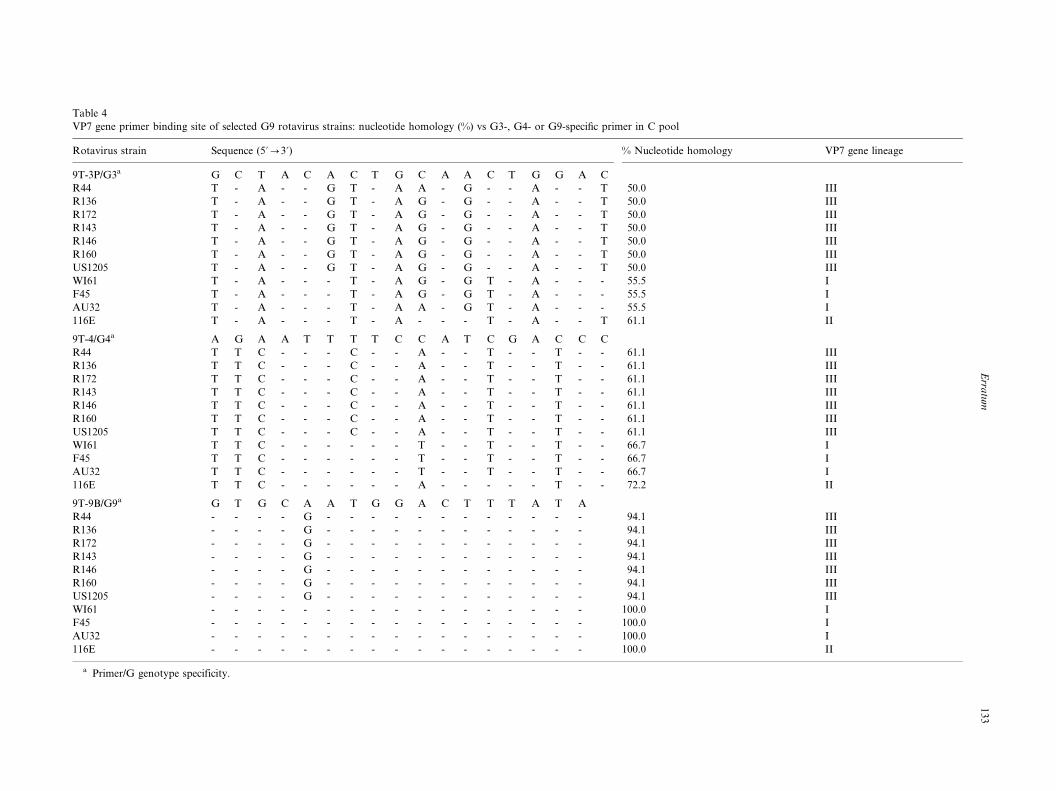
Table 4
VP7 gene primer binding site of selected G9 rotavirus strains: nucleotide homology (%) vs G3-, G4- or G9-specific primer in C pool
Rotavirus strain Sequence (5?0/3?) % Nucleotide homology VP7 gene lineage
9T-3P/G3a G C T A C A C T G C A A C T G G A C
R44 T - A - - G T - A A - G - - A - - T 50.0 III
R136 T - A - - G T - A G - G - - A - - T 50.0 III
R172 T - A - - G T - A G - G - - A - - T 50.0 III
R143 T - A - - G T - A G - G - - A - - T 50.0 III
R146 T - A - - G T - A G - G - - A - - T 50.0 III
R160 T - A - - G T - A G - G - - A - - T 50.0 III
US1205 T - A - - G T - A G - G - - A - - T 50.0 III
WI61 T - A - - - T - A G - G T - A - - - 55.5 I
F45 T - A - - - T - A G - G T - A - - - 55.5 I
AU32 T - A - - - T - A A - G T - A - - - 55.5 I
116E T - A - - - T - A - - - T - A - - T 61.1 II
9T-4/G4a A G A A T T T T C C A T C G A C C C
R44 T T C - - - C - - A - - T - - T - - 61.1 III
R136 T T C - - - C - - A - - T - - T - - 61.1 III
R172 T T C - - - C - - A - - T - - T - - 61.1 III
R143 T T C - - - C - - A - - T - - T - - 61.1 III
R146 T T C - - - C - - A - - T - - T - - 61.1 III
R160 T T C - - - C - - A - - T - - T - - 61.1 III
US1205 T T C - - - C - - A - - T - - T - - 61.1 III
WI61 T T C - - - - - - T - - T - - T - - 66.7 I
F45 T T C - - - - - - T - - T - - T - - 66.7 I
AU32 T T C - - - - - - T - - T - - T - - 66.7 I
116E T T C - - - - - - A - - - - - T - - 72.2 II
9T-9B/G9a G T G C A A T G G A C T T T A T A
R44 - - - - G - - - - - - - - - - - - 94.1 III
R136 - - - - G - - - - - - - - - - - - 94.1 III
R172 - - - - G - - - - - - - - - - - - 94.1 III
R143 - - - - G - - - - - - - - - - - - 94.1 III
R146 - - - - G - - - - - - - - - - - - 94.1 III
R160 - - - - G - - - - - - - - - - - - 94.1 III
US1205 - - - - G - - - - - - - - - - - - 94.1 III
WI61 - - - - - - - - - - - - - - - - - 100.0 I
F45 - - - - - - - - - - - - - - - - - 100.0 I
AU32 - - - - - - - - - - - - - - - - - 100.0 I
116E - - - - - - - - - - - - - - - - - 100.0 II
a Primer/G genotype specificity.
Erra
tum
13
3

belonging to lineage I or III presented six or seven
nucleotide mismatches (61.1 or 66.7% homology) at the
primer 9T-4 binding site (nt 423�/440) and none were
amplified. Although strain 116E (lineage II) presentedonly five nucleotide mismatches (72.2% homology), it
was not amplified with the primer.
Strains belonging to lineage III which presented one
identical nucleotide mismatch (94.1% homology) at the
9T-9B primer binding site (nt 131�/147) were amplified.
No mismatches were observed at the primer binding site
of strains belonging to lineage I or II.
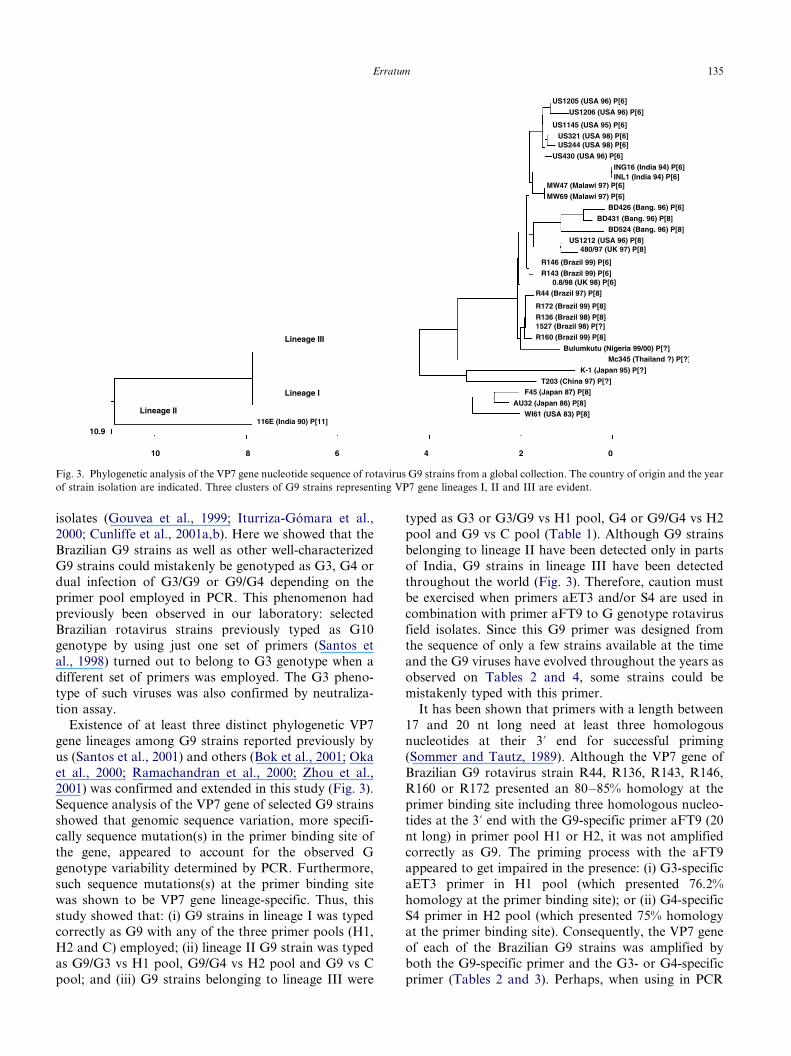
3.5. Phylogenetic analysis of VP7 gene of Brazilian and
other G9 rotavirus strains
Phylogenetic comparison of the VP7 encoding gene of
selected strains belonging to G3, G4 or G9 showed that
the 12 G9 strains that were analyzed in this study
clustered together in G9 genotype which was distinct
from G3 or G4 genotype phylogenetically (Fig. 2), even
though such G9 strains showed a high degree of cross-
reactivity with G3- or G4-specific primer in PCR
amplification (Tables 2 and 3).The VP7 gene sequence analysis of the Brazilian as
well as other G9 strains from a global collection showed
the existence of at least three VP7 gene lineages among
them (Fig. 3): lineage I represented by strains WI61,
F45, and AU32; lineage II formed by the Indian strain
116E; and lineage III consisted of strains from Brazil,
U.S., India, Malawi, Bangladesh, Nigeria, UK, China
and Japan.
3.6. Characterization by neutralization of selected
Brazilian G9 rotavirus strains
Brazilian G9 strains R44 and R143 were cell culture-
adapted and characterized by PRN assay. Each rota-
virus strain was tested against guinea pig hyperimmune
antiserum raised to each of G1 to G14 rotavirus strains.
Both strains R44 and R143 were neutralized strongly
only by hyperimmune antiserum to WI61 or F45
prototype G9 strain.
3.7. Characterization by PAGE of Brazilian and other
G9 rotavirus strains
Four of 23 Brazilian G9 rotavirus strains exhibited so-called ‘short’ (S) pattern and the remaining 19 strains
had a ‘long’ (L) pattern when analyzed by polyacryla-
mide gel electrophoresis (PAGE) (Table 1). These four
strains with S pattern may be naturally-occurring
reassortants. Four prototype G9 strains (WI61, F45,
AU32 and 116E) displayed in PAGE gel an L pattern
whereas strain US1205 exhibited a S pattern as de-
scribed previously.
4. Discussion
Sequence analysis of selected rotavirus genes has
provided information leading to substantial advances
in our understanding of: (i) the phylogenetic diversity of
this virus; and (ii) genetic and antigenic diversity of
outer capsid protective antigens VP7 and VP4, the
targets of current vaccine development strategies. Suchstudies have also provided new insight into genetic and
antigenic intratypic VP7 variation within a given human
rotavirus G serotype including G1 (Xin et al., 1993; Jin
et al., 1996; Maunula and von Bonsdorff, 1998; Diwa-
karla and Palombo, 1999), G2 (Wen et al., 1995;
Coulson et al., 1996; Zao et al., 1999), G3 (Wen et al.,
1997), G4 (Coulson et al., 1996; Palombo et al., 1997),
and G9 (Coulson et al., 1999; Ramachandran et al.,2000). Furthermore, for rotavirus serotypes G1, G2, G4
and G8, such VP7 gene variations were shown to be
associated with a failure in correct G type identification
by PCR (Nakagomi et al., 1991; Adah et al., 1997;
Cunliffe et al., 2001a,b). Genetic variation has also been
described for the most common human rotavirus VP4
serotype P1A[8], leading to a failure to P genotype such
Fig. 2. Phylogenetic tree of VP7 gene nucleotide sequences of rotavirus serotypes G3, G4 and G9 strains. Branch length is proportional to the
percentage of nucleotide substitution indicated in the scale.
Erratum134
isolates (Gouvea et al., 1999; Iturriza-Gomara et al.,
2000; Cunliffe et al., 2001a,b). Here we showed that the
Brazilian G9 strains as well as other well-characterized
G9 strains could mistakenly be genotyped as G3, G4 or
dual infection of G3/G9 or G9/G4 depending on the
primer pool employed in PCR. This phenomenon had
previously been observed in our laboratory: selected
Brazilian rotavirus strains previously typed as G10genotype by using just one set of primers (Santos et
al., 1998) turned out to belong to G3 genotype when a
different set of primers was employed. The G3 pheno-
type of such viruses was also confirmed by neutraliza-
tion assay.
Existence of at least three distinct phylogenetic VP7
gene lineages among G9 strains reported previously by
us (Santos et al., 2001) and others (Bok et al., 2001; Oka
et al., 2000; Ramachandran et al., 2000; Zhou et al.,2001) was confirmed and extended in this study (Fig. 3).
Sequence analysis of the VP7 gene of selected G9 strains
showed that genomic sequence variation, more specifi-
cally sequence mutation(s) in the primer binding site of
the gene, appeared to account for the observed G
genotype variability determined by PCR. Furthermore,
such sequence mutations(s) at the primer binding site
was shown to be VP7 gene lineage-specific. Thus, this
study showed that: (i) G9 strains in lineage I was typedcorrectly as G9 with any of the three primer pools (H1,
H2 and C) employed; (ii) lineage II G9 strain was typed
as G9/G3 vs H1 pool, G9/G4 vs H2 pool and G9 vs C
pool; and (iii) G9 strains belonging to lineage III were
typed as G3 or G3/G9 vs H1 pool, G4 or G9/G4 vs H2
pool and G9 vs C pool (Table 1). Although G9 strains
belonging to lineage II have been detected only in parts
of India, G9 strains in lineage III have been detected
throughout the world (Fig. 3). Therefore, caution must
be exercised when primers aET3 and/or S4 are used in
combination with primer aFT9 to G genotype rotavirus
field isolates. Since this G9 primer was designed fromthe sequence of only a few strains available at the time
and the G9 viruses have evolved throughout the years as
observed on Tables 2 and 4, some strains could be
mistakenly typed with this primer.
It has been shown that primers with a length between
17 and 20 nt long need at least three homologous
nucleotides at their 3? end for successful priming
(Sommer and Tautz, 1989). Although the VP7 gene of
Brazilian G9 rotavirus strain R44, R136, R143, R146,R160 or R172 presented an 80�/85% homology at the
primer binding site including three homologous nucleo-
tides at the 3? end with the G9-specific primer aFT9 (20
nt long) in primer pool H1 or H2, it was not amplified
correctly as G9. The priming process with the aFT9
appeared to get impaired in the presence: (i) G3-specific
aET3 primer in H1 pool (which presented 76.2%
homology at the primer binding site); or (ii) G4-specific
S4 primer in H2 pool (which presented 75% homologyat the primer binding site). Consequently, the VP7 gene
of each of the Brazilian G9 strains was amplified by
both the G9-specific primer and the G3- or G4-specific
primer (Tables 2 and 3). Perhaps, when using in PCR
Fig. 3. Phylogenetic analysis of the VP7 gene nucleotide sequence of rotavirus G9 strains from a global collection. The country of origin and the year
of strain isolation are indicated. Three clusters of G9 strains representing VP7 gene lineages I, II and III are evident.
Erratum 135

amplification a pool of two or more primers which have
a close nucleotide homology at each respective primer
binding site, the priming process may favor an annealing
of one primer over others regardless of the number and
position of nucleotide mismatches at the primer binding
site. Further studies need to be explored.
Of interest was the behavior in PCR of three Brazilian
strains (R143, R146 and R160) against G9-specific aFT9
primer (which showed an 85% homology at primer
binding site) present in H1 or in H2 pool (Tables 2 and
3). Data obtained in this study appear to suggest that at
an annealing temperature of 50 8C in PCR, if the VP7
gene nucleotide homology difference between homoty-
pic and heterotypic primers at each respective primer
binding site in a pooled primer set is: (i) higher than
20%, only the homotypic priming may take place; (ii)
lower than 5%, only the heterotypic priming may occur;
and (iii) between 5 and 10%, both homotypic and
heterotypic priming may be seen. More detailed studies
need to be performed.
Epidemiologic surveillance of rotavirus serotypes/
genotypes (both VP7[G] and VP4[P]) is underway
worldwide, because neutralizing antibodies appear to
play an important protective role against rotavirus
disease in a type-specific manner (Hoshino and Kapi-
kian, 2000). The advent of PCR-typing methodology
has greatly enhanced an efficiency of such epidemiologic
surveillance of rotavirus genotypes. Nonetheless, the
results described here reinforce the need for improve-
ment on such methodology. For example, the use of
more than one set of type-specific primer pools may be
necessary for an accurate and efficient genotype deter-
mination of rotavirus field isolates. In addition, since an
‘antigenic drift’ of the rotavirus VP7 gene has been
shown to occur as reported by us (this study) and others
(Palombo et al., 1997), continuous efforts need to be
made to design a type-specific primer which recognizes
as many rotavirus isolates as possible within a given G
genotype. Furthermore, since errors in PCR genotyping
due to cross-priming of multiplexed primers could
occasionally occur, selective confirmation of PCR
results by other assays will be needed including sequen-
cing, probe hybridization or serotyping with type-
specific antibodies.
The primer aFT9 which was designed from the VP7
gene sequence of lineage I G9 strains WI61, AU32 and
F45 isolated in the 1980s was shown to fail to prime
correctly more recent G9 isolates belonging to lineage II
or III. Whereas the primer 9T-9B designed from the VP7
gene sequence of more recent 116E strain (lineage II)
genotyped correctly all the G9 isolates analyzed. If in
the present study only H1 or H2 primer pool had been
used, Brazilian and Indian G9 strains would have been
typed wrongly. Actually, it is possible that the real
frequency of rotavirus G9 genotype in Brazil, and other
countries as well, could be underestimated due to the
phenomenon that we described in this study.
5. Nucleotide sequence accession numbers
The sequences were deposited in the GenBank under
the accession numbers AF438227, AF438228,
AF274969, AF274970 and AF274971. Nucleotide se-
quences of the VP7 gene from strains R172 and R136
are identical.
Acknowledgements
We thank Ronald Jones, Jerri Ross, and Mariam
Wagner for their expert technical assistance; and Albert
Kapikian for encouragement throughout the study. This
work was partially supported by CNPq, FINEP,
FAPERJ, FUJB, and CAPES Brazil, and TWAS, Italy.
References
Adah, M.I., Rohwedder, A., Olaleyly, O.D., Werchau, H., 1997.
Nigerian rotavirus serotype G8 could not be typed by PCR due to
nucleotide mutation at the 3? end of the primer binding site. Arch.
Virol. 142, 1881�/1887.
Adah, M.I., Wade, A., Taniguchi, K., 2001. Molecular epidemiology
of rotaviruses in Nigeria: detection of unusual strains with G2P[6]
and G8P[1] specificities. J. Clin. Microbiol. 39, 3969�/3975.
Akatani, K., Ikegami, N., 1987. Typing of fecal rotavirus specimens by
an enzyme-linked immunosorbent assay using monoclonal anti-
bodies. Clin. Virol. 15, 61�/68 (in Japanese).
Araujo, I.T., Ferreira, M.S.R., Fialho, A.M., Assis, R.M., Cruz, C.M.,
Rocha, M., Leite, J.P.G., 2001. Rotavirus genotypes P[4]G9,
P[6]G9, and P[8]G9 in hospitalized children with acute gastro-
enteritis in Rio de Janeiro, Brazil. J. Clin. Microbiol. 39, 1999�/
2001.
Bern, C., Glass, R.I., 1994. Impact of diarrheal diseases worldwide. In:
Kapikian, A.Z. (Ed.), Viral Infections of the Gastrointestinal
Tract, 2nd ed.. Marcel Dekker, New York, pp. 1�/6.
Bok, K., Palacios, G., Sijvarger, K., Matson, D., Gomez, J., 2001.
Emergence of G9P[6] human rotaviruses in Argentina: phyloge-
netic relationships among G9 strains. J. Clin. Microbiol. 39, 4020�/
4025.
The Azay Group, Bon, F., Fromantin, C., Aho, S., Pothier, P., Kohli,
E., 2000. G and P genotyping of rotavirus strains circulating in
France over a 3-year period: detection of G9 rotavirus and P[6]
strains at low frequencies. J. Clin. Microbiol. 38, 1681�/1683.
Bresee, J.S., Glass, R.I., Ivanoff, B., Gentsch, J.R., 1999. Current
status and future priorities for rotavirus vaccine development,
evaluation and implementation in developing countries. Vaccine
17, 2207�/2222.
Clark, H.F., Hoshino, Y., Bell, L.M., Groff, J., Hess, G., Bachman, P.,
Offit, P.A., 1987. Rotavirus isolate WI61 representing a presump-
tive new human serotype. J. Clin. Microbiol. 25, 1757�/1762.
Coulson, B.S., Kirkwood, C.D., Masendyez, P.J., Bishop, R.F.,
Gerna, G., 1996. Amino acid involved in distinguishing between
monotypes of rotavirus G serotypes 2 and 4. J. Gen. Virol. 77,
239�/245.
Erratum136

Coulson, B.S., Gentsch, J.R., Das, K., Bhan, M.K., Glass, R.I., 1999.
Comparison of enzyme immunoassay and reverse transcriptase
PCR for identification of serotype G9 rotaviruses. J. Clin.
Microbiol. 37, 3187�/3193.
Cubitt, W.D., Steele, A.D., Iturriza, M., 2000. Characterization of
rotavirus from children treated at a London hospital during 1996:
emergence of strains G9P2A[6] and G3P2A[6]. J. Med. Virol. 61,
150�/154.
Cunliffe, N.A., Gondwe, J.S., Broadhead, R.L., Molyneux, M.E.,
Woods, P.A., Bresee, J.S., Glass, R.I., Gentsch, J.R., Hart, C.A.,
1999. Rotavirus G and P types in children with acute diarrhea in
Blantyre, Malawi, from 1997 to 1998: predominance of a novel
P[6]G8 strains. J. Med. Virol. 57, 308�/312.
Cunliffe, N.A., Gondwe, J.S., Graham, S.M., Thindwa, B.D.M.,
Dove, W., Broadhead, R.L., Molyneux, M.E., Hart, C.A., 2001a.
Rotavirus strains diversity in Blantyre, Malawi, from 1997 to 1999.
J. Clin. Microbiol. 39, 836�/843.
Cunliffe, N.A., Dove, W., Bunn, J.E.G., Ramadam, M.B., Nyangao,
J.W.O., Riveron, R.L., Cuevas, L.E., Hart, C.A., 2001b. Expand-
ing global distribution of rotavirus serotype G9: detection in Libya,
Kenya, and Cuba. Emerg. Infect. Dis. 7, 890�/892.
Das, B.K., Gentsch, J.R., Cicirelo, H.G., Woods, P.A., Gupta, A.,
Ramachandran, M., Kumar, R., Bhan, M.K., Glass, R.G., 1994.
Characterization of rotavirus strains from newborns in New Delhi,
India. J. Clin. Microbiol. 32, 1820�/1822.
Desselberger, U., 1996. Genome rearrangements of rotavirus. Adv.
Virus Res. 46, 69�/95.
Diwakarla, C.S., Palombo, E.A., 1999. Genetic and antigenic variation
of capsid protein VP7 of serotype G1 human rotavirus isolates. J.
Gen. Virol. 80, 341�/344.
Glass, R.I., Bresee, J.S., Parashar, U.D., Gentsch, J.R., 1999. First
rotavirus vaccine licensed: is there really a need? Acta Paediatr.
Suppl. 426, 2�/8.
Gouvea, V., Glass, R.I., Woods, P.A., Taniguchi, K., Clark, H.F.,
Forrester, B., Fang, Z.Y., 1990. Polymerase chain reaction
amplification and typing of rotavirus nucleic acid from stool
specimens. J. Clin. Microbiol. 28, 276�/282.
Gouvea, V., Santos, N., Timenetsky, M., 1994. Identification of bovine
and porcine G types by PCR. J. Clin. Microbiol. 32, 1338�/1340.
Gouvea, V., Brantly, M., 1995. Is rotavirus a population of reassor-
tants? Trends Microbiol. 3, 159�/162.
Gouvea, V., Lima, R.C.C., Linhares, R.E., Clark, H.F., Nozawa,
C.M., Santos, N., 1999. Identification of two lineages (WA-like and
F45-like) whitin the major rotavirus genotype P[8]). Virus Res. 59,
141�/147.
The National Rotavirus Strain Surveillance System Collaborating
Laboratories, Griffin, D.D., Kirkwood, C.D., Parashar, U.D.,
Woods, P.A., Bresee, J.S., Glass, R.I., Gentsch, J.R., 2000.
Surveillance of rotavirus strains in the United States: identification
of unusual strains. J. Clin. Microbiol. 38, 2784�/2787.
Herring, A.J., Inglis, N.F., Ojhe, C.K., Snodgrass, D.R., Menzies,
J.D., 1982. Rapid diagnosis of rotavirus infection by direct
detection of a viral nucleic acid in silver-stained polyacrylamide
gels. J. Clin. Microbiol. 16, 473�/477.
Hoshino, Y., Kapikian, A.Z., 2000. Rotavirus serotypes: classification
and importance in rotavirus epidemiology, immunity and vaccine
development. J. Health Popl. Nutr. 18, 5�/14.
Isegawa, Y., Nakagomi, O., Nakagomi, T., Uesugi, S., Ueda, S., 1993.
Determination of bovine rotavirus G and P serotypes by poly-
merase chain reaction. Mol. Cell. Prob. 7, 277�/284.
Iturriza-Gomara, M., Green, J., Brown, D.W.G., Desselberger, U.,
Gray, J.J., 2000. Diversity whitin the VP4 gene of rotavirus P[8]
genotyping. J. Clin. Microbiol. 38, 898�/901.
Iturriza-Gomara, M., Isherwood, B., Desselberger, U., Gray, J., 2001.
Reassortment in vivo: driving force for diversity of human
rotavirus strains isolated in the United Kingdom between 1995
and 1999. J. Virol. 75, 3696�/3705.
Jin, Q., Ward, R.L., Knowlton, D.R., Gabbay, Y.B., Linhares, A.C.,
Rappaport, R., Woods, P.A., Glass, R.I., Gentsch, J.R., 1996.
Divergence of VP7 genes of G1 rotaviruses isolated from infants
vaccinated with reassortant rhesus rotaviruses. Arch. Virol. 141,
2057�/2076.
Kapikian, A.Z., Hoshino, Y., Chanock, R.M., 2001. Rotaviruses. In:
Knipe, D.M., Howley, R.M. (Eds.), Fields Virology, 4th ed..
Lippincott, Williams and Wilkins, Philadelphia, Pennsylvania, pp.
1787�/1825.
Maneekarn, N., Ushijima, H., 2000. Epidemiology of rotavirus
infection in Thailand. Pediatr. Int. 42, 415�/521.
Maunula, L., von Bonsdorff, C.H., 1998. Short sequences define
genetic lineages: phylogenetic analysis of group A rotaviruses based
on partial sequences of genome segments 4 and 9. J. Gen. Virol. 79,
321�/332.
Nakagomi, O., Oyamada, H., Nakagomi, T., 1991. Experience with
serotyping rotavirus strains by reverse transcription and two-step
polymerase chain reaction with generic and type-specific primers.
Mol. Cell. Prob. 5, 285�/289.
Nakagomi, T., Oshima, A., Akatani, K., Ikegami, N., Katsushima, N.,
Nakagomi, O., 1990. Isolation and molecular characterization of a
serotype 9 human rotavirus strain. Microbiol. Immunol. 34, 77�/82.
O’Halloran, F., Lynch, M., Cryan, B., O’Shea, H., Fanning, 2000.
Molecular characterization of rotavirus in Ireland: detection of
novel strains circulating in the population. J. Clin. Microbiol. 38,
3370�/3374.
Oka, T., Nakagomi, T., Nakagomi, O., 2000. Apparent re-emergence
of serotype G9 in 1995 among rotaviruses recovered from Japanese
children hospitalized with acute gastroenteritis. Microbiol. Immu-
nol. 44, 957�/961.
Palombo, E.A., Bugg, H.C., Masendycz, P.J., Bishop, R.F., 1997.
Sequence of the VP7 gene of an atypical human rotavirus: evidence
for genetic and antigenic drift. DNA Seq. 7, 307�/311.
Palombo, E., Masendycz, P.J., Bugg, H.C., Bogdanovic-Sakran, N.,
Barnes, G.L., Bishop, R.F., 2000. Emergence of serotype G9
human rotavirus in Australia. J. Clin. Microbiol. 38, 1305�/1306.
Parashar, U.D., Holman, R.C., Clarke, M.J., Bresee, J.S., Glass, R.I.,
1998a. Hospitalizations associated with rotavirus diarrhea in the
United Sates, 1993 through 1995: surveillance based on the new
ICD-9-CM rotavirus specific diagnostic code. J. Infect. Dis. 177,
7�/13.
Parashar, U.D., Bresee, J.S., Gentsch, J.R., Glass, R.I., 1998b.
Rotavirus. Emerg. Infect. Dis. 4, 561�/570.
Ramachandran, M., Das, B.K., Vij, A., Kumar, R., Bhambal, S.S.,
Kesari, N., Rawat, H., Bahl, L., Thakur, S., Woods, P.A., Glass,
R.I., Bhan, M.K., Gentsch, J.R., 1996. Unusual diversity of human
rotavirus G and P genotypes in India. J. Clin. Microbiol. 34, 436�/
439.
The National Rotavirus Strain Surveillance System Collaborating
Laboratories, Ramachandran, M., Gentsch, J.R., Parashar, U.D.,
Jin, S., Woods, P.A., Holmes, J.L., Kirkwood, C.D., Bishop, R.F.,
Greenberg, H.B., Urasawa, S., Gerna, G., Coulson, B.S., Tani-
guchi, K., Bresee, J.S., Glass, R.I., 1998. Detection and character-
ization of novel rotavirus strains in the United States. J. Clin.
Microbiol. 36, 3223�/3229.
Ramachandran, M., Kirkwood, C.D., Unicomb, L., Cunliffe, N.A.,
Ward, R.L., Bhan, R.L., Clark, H.F., Glass, R.I., Gentsch, J.R.,
2000. Molecular characterization of serotype G9 rotavirus strains
from a global collection. Virology 278, 436�/444.
Santos, N., Gouvea, V., 1994. Improved method for purification of
viral RNA from fecal specimens for rotavirus detection. J. Virol.
Methods 46, 11�/21.
Santos, N., Lima, R.C.C., Pereira, C.F.A., Gouvea, V., 1998.
Detection of rotavirus types G8 and G10 among Brazilian children
with diarrhea. J. Clin. Microbiol. 36, 2727�/2729.
Santos, N., Volotao, E.M., Soares, C.C., Albuquerque, M.C.M., da
Silva, F.M., de Carvalho, T.R.B., Pereira, C.F.A., Chizhikov, V.,
Erratum 137

Hoshino, Y., 2001. Rotavirus strains bearing genotype G9 or P[9]
recovered from Brazilian children with diarrhea from 1997 to 1999.
J. Clin. Microbiol. 39, 1157�/1160.
Sommer, R., Tautz, D., 1989. Minimal homology requirements for
PCR primers. Nucl. Acids Res. 17, 6749.
Suzuki, Y., Gojobori, T., Nakagomi, O., 1998. Intragenic recombina-
tion in rotaviruses. FEBS 427, 183�/187.
Taniguchi, K., Wakasugi, F., Ponsuwanna, Y., Urasawa, T., Ukae, S.,
Chiba, S., Urasawa, S., 1992. Identification of human and bovine
rotavirus serotypes by polymerase chain reaction. Epidemiol.
Infect. 109, 303�/312.
Tucker, A.W., Haddix, A.C., Bresee, J.S., Holman, R.C., Parashar,
U.D., Glass, R.I., 1998. Cost-effectiveness analysis of a rotavirus
immunization program for the United States. J. Am. Med. Assoc.
297, 1371�/1376.
Unicomb, L.E., Podder, G., Gentsch, J.R., Woods, P.A., Hasan, K.Z.,
Faruque, A.S.G., Albert, M.J., Glass, R.I., 1999. Evidence of high-
frequency genomic reassortment of group A rotavirus strains in
Bangladesh: emergence of type G9 in 1995. J. Clin. Microbiol. 37,
1885�/1891.
Urasawa, S., Hasegawa, A., Urasawa, T., Taniguchi, K., Wakasugi,
F., Suzuki, H., Inouye, S., Pongprot, B., Supawadee, J., Suprasert,
S., Rangsiyamond, P., Tonusin, S., Yamazi, Y., 1992. Antigenic
and genetic analyses of human rotaviruses in Chiang Mai, Thai-
land: evidence for a close relationship between human and animal
rotavirus. J. Infect. Dis. 166, 227�/234.
The Vietnam Rotavirus Surveillance Network, Van Man, N., Van
Trang, N., Lien, H.P., Trach, D.D., Thanh, N.T.H., Tu, P.V., Long,
N.T., Luan, I.T., Ivanoff, B., Gentsch, J.R., Glass, R.I., 2001. The
epidemiology and disease burden of rotavirus in Vietnam: sentinel
surveillance at 6 hospitals. J. Infect. Dis. 183, 1707�/1712.
Wen, L., Ushijima, H., Kakizawa, J., Fang, Z.Y., Nishio, O.,
Morikawa, S., Motohiro, T., 1995. Genetic variation in VP7 gene
of human rotavirus serotype 2 (G2 type) isolated in Japan, China,
and Pakistan. Microbiol. Immunol. 39, 911�/915.
Wen, L., Nakayama, M., Yamanishi, Y., Nishio, O., Fang, Z.-Y.,
Nakagomi, O., Araki, K., Nishimura, S., Hasegawa, A., Muller,
W.E.G., Ushijima, H., 1997. Genetic variation in the VP7 gene of
human rotavirus serotype 3 (G3 type) isolated in China and Japan.
Arch. Virol. 142, 1481�/1489.
Widdowson, M.A., van Doornum, G.J., van der Poel, W.H., de Boer,
A.S., Mahdi, U., Koopmans, M., 2000. Emerging group-A
rotavirus and a nosocomial outbreak of diarrhoea. Lancet 356,
1161�/1162.
Wyatt, R.G., Greenberg, H.B., James, H.D., Jr., Pittman, A.L.,
Kalica, A.R., Flores, J., Chanock, R.M., Kapikian, A.Z., 1982.
Definition of human rotavirus serotypes by plaque reduction assay.
Infect. Immun. 37, 110�/115.
Xin, K.-Q., Morikawa, S., Fang, Z.-Y., Mukoyama, A., Okuda, K.,
Ushima, H., 1993. Genetic variation in VP7 gene of human
rotavirus serotype 1 (G1 type) isolated in Japan and China.
Virology 197, 813�/816.
Zao, C.-L., Yu, W.-N., Kao, C.-L., Taniguchi, K., Lee, C.-Y., Lee, C.-
N., 1999. Sequence analysis of VP1 and VP7 genes suggests
occurrence of a reassortant of G2 rotavirus responsible for an
epidemic of gastroenteritis. J. Gen. Virol. 80, 1407�/1415.
Zhou, Y., Supawadee, J., Khamwan, C., Tonusin, S., Peerakome, S.,
Kim, B., Kaneshi, K., Ueda, Y., Nakaya, S., Akatani, K.,
Maneekarn, N., Ushijima, H., 2001. Characterization of human
rotavirus serotype G9 isolated in Japan and Thailand from 1995 to
1997. J. Med. Virol. 65, 619�/628.
Erratum138
Related Documents










![Rotavirus Vaccines: an Overview - myCMEmedia.mycme.com/documents/23/rotavirus_expert_reviews...rotavirus strains bearing VP7 G serotypes G1 to G4 and G9 and VP4 P genotypes P1B[4],](https://static.cupdf.com/doc/110x72/5f8e21d6883e9848ab510511/rotavirus-vaccines-an-overview-rotavirus-strains-bearing-vp7-g-serotypes.jpg)
