��������� VOL. SPEC. - N° 10, 2005

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

���������
VOL. SPEC. - N° 10, 2005

ISSN 0253-6730
1 Department of Evolution, Systematics and Ecology, The Hebrew University, Jerusalem 91904, Israel2 CNRS, France (retired)
Revue de Paléobiologie, Genève (décembre 2005) Vol. spéc. 10 : 209-224
Counting Cattle :Trends in Neolithic Bos Frequencies from the Southern Levant
Liora Kolska HORWITZ1 & Pierre DUCOS1,2
AbstractThe marked increase in caprine frequencies observed by the mid-Pre-Pottery Neolithic, is generally accepted as heralding the onset of their domestication/or introduction as domesticates into the Southern Levant.In order to investigate whether cattle exploitation shows a similar diachronic patterning to that shown by caprines, the relative proportions of their remains in archaeological sites from the Southern Levant were examined by chronological phase and geographic region. Results show that the increase in remains of cattle is not synchronous throughout the region but varies between geographic zones. An initial increase in cattle frequencies was noted in the Mid-Pre-Pottery Neolithic B. It was followed by a major increase in numbers by the Pre-Pottery Neolithic C accompanied by reduction in body size, indicating a significant time lag in the introduction/or domestication of cattle relative to caprines. Key wordsAurochs, cattle, domestication, Neolithic, Southern Levant.
RésuméQuantifier les boeufs : leur importance relative au Levant sud durant le Néolithique.- La fréquence des Caprinés sʼaccroît de manière très évidente pendant le Néolithique Pré-poterie Moyen. Il est généralement admis que cet accroissement indique le commencement de leur domestication ou leur introduction à lʼétat domestique au Levant sud.Afin de savoir si lʼexploitation des Bovidés est similaire diachroniquement à celle observée pour les Caprinés, nous avons examiné, par phases chronologiques et régions géographiques, les fréquences de leurs restes osseux dans les sites du Levant sud. Les résultats montrent que lʼaccroissement des fréquences des Bovidés ne se produit pas synchroniquement, mais à des moments différents selon les zones géographiques. Une première augmentation des fréquences sʼobserve dans le PPNB moyen, se généralise au PPNC, accompagné alors dʼune diminution de la taille. Par rapport à la domestication/introduction des Caprinés, celles des Bovidés se fait avec un décalage chronologique significatif.
Mots-clésAurochs, boeuf, domestication, Néolithique, Levant sud.
1. INTRODUCTION
Several criteria have been advanced for assessing the domestic status of animals (MEADOW, 1989 ; VIGNE et al., 2005 and references cited therein). They include : the presence of a species outside its natural range, evidence for biometric and morphological changes, shifts in sex ratios and age distributions, the manner in which carcasses were processed and most recently, specific isotopic and DNA signatures. An additional diagnostic feature is a reduction in species diversity and an increased and sustained reliance on a few select species. This signifies dietary specialisation as a species is appropriated into the anthropogenic sphere. Resource specialisation is defined by ENLOE (1999 : 502) as, “when a particular resource offers a better solution to the problem of year-round, long-term food acquisition than does a general mix of other potential food resources”. Under these conditions, a
range of resources may continue to be utilized, but a key resource takes precedence over the others since it is able to fulfil the specific needs of the group, either in the short term or long term. Domestication of herd animals in the Near East is an excellent example of resource specialisation. It is clearly demonstrated by the sudden rise in goat frequencies in mid-Pre-Pottery Neolithic B (MPPNB) sites in the Southern Levantine, and is associated with a concomitant decrease in the exploitation of wild taxa, especially small-sized game and gazelle. All these features have been interpreted by researchers as heralding the onset of proto-domestication/domestication of caprines in the region (CLUTTON-BROCK, 1979, 1999 ; DAVIS, 1981, 1982 ; DUCOS, 1968, 1993, 1995 ; HORWITZ, 1993, 1996 ; HORWITZ & TCHERNOV, 1998 ; HORWITZ et al., 1999 ; MUNRO, 2004 ; TCHERNOV, 1993). The current paper focuses on the timing of cattle

domestication in the Southern Levant, the area encompassing Syria south of the Damascus Basin, Lebanon, Jordan, Israel and the Sinai Peninsula. It aims at investigating whether, like caprines, the onset of cattle domestication during the Neolithic, was marked by a significant increase in their numbers as well as the extent to which this was associated with changes in body size, age and sex ratios. These data will be used to evaluate the association between changes in the numbers of animals exploited and their domestic status. Gazelle, a wild species that was never domesticated, will serve as a standard for the extent of hunting practiced. Caprines will provide a standard for the extent of numerical changes expected in a taxon undergoing domestication as well as offer some indication as to the timing of this event. Since the Southern Levant is characterised by a mosaic of contrasting physical environments, the faunal assemblages have been analysed using a diachronic/geographic approach. This will facilitate examination of regional variation in contemporaneous modes of production for cattle, and determine whether they resemble those previously reported for caprines (HORWITZ, 1993, 2003).
MATERIAL & METHODS
The assemblages examined here derive from 40 Neolithic sites from the Southern Levant, spanning the Pre-Pottery Neolithic A (PPNA) to Pottery Neolithic (PN) periods (Fig. 1, Tables 1 & 2). The data were calculated from published archaeozoological reports as well as several unpublished data-sets of the authors. Since many of the published sources have been referenced in two other syntheses (HORWITZ, 1996 ; HORWITZ & TCHERNOV, 1998),
Fig. 1 : Map showing Neolithic sites in the Southern Levant. Due to the small scale of the map, several sites that are located close to each other are denoted by the same number.
(1) Hagoshrim, (2) Yiftahʼel, Kfar HaHoresh, (3) Munhatta, Tel Ali, (4) Gesher, (5) ʻAin Ghazal, (6) Jericho, (7) Abu Ghosh, (8) El Khiam, (9) Nahal Oren, Atlit Yam, Newe Yam, (10) Tel Teʼo (11) Ramad, (12) Aswad, (13) Beidha, (14) Basta, (15) Ayn Abu Nukhayla, (16) Jilat sites, (17) Dhuweila, (18) Azraq, Burqu, (19) Gilgal, Netiv Hagdud, (20) Ziqim, Ashkelon, (21) Qatif Y3, (22) Khirbet Hammam, (23) Faynan 16, Wadi Fidan A & C, (24) Labwe, (25) Jebel Abu Thawwab, Wadi Shuʼeib, (26) Nahal Zehora, (27) Shaʼar Hagolan, (28) Wadi Tbeik, (29) Ujret el-Mehed, (30) Hatoula.
Table 1 : Chronology of periods in the Southern Levant (uncalibrated B.P. dates after KUIJT & GORING-MORRIS, 2002).
Time Period Stratigraphic Units Culture/Phase Dates (uncalibrated B.P.)
Late Epipaleolithic Natufian 13,000-10,300Neolithic Pre-Pottery Neolthic A PPNA 10,300-9,600 Pre-Pottery Neolithic B EPPNB 9,600-9,300 MPPNB 9,300-8,500 LPPNB 8,500-8,000 Terminal PPNB or PPNC 8,000-7,500 Pottery Neolithic PN 7,500-7,000
Yarmukian & Wadi Rabah phases
210 L. K. HORWITZ & P. DUCOS

Table 2 : Southern Levantine sites used in this study. Sites are divided into five geographic zones : (1) Western Israel-Mediterranean coast, (2) Beqaʼa –Jordan Valley, (3) Mediterranean Zone, (4) Eastern Arid Zone, (5) Southern Arid Zone.
Sites whose full references appear in two previous studies (HORWITZ, 1996 ; HORWITZ & TCHERNOV, 1998) are denoted by a * and are not cited here.
PERIOD MAP NO. SITE ZONE REFERENCEPPNA 9 Nahal Oren Western NOY et al., 1973 30 Hatoula Mediterranean DAVIS et al., 1994 * 4 Gesher Beqaʼa –Jordan Valley HORWITZ & GARFINKEL, 1991 * 19 Netiv Hagdud Beqaʼa –Jordan Valley TCHERNOV, 1994 * 19 Gilgal I Beqaʼa –Jordan Valley HORWITZ et al., in press 6 Jericho Beqaʼa –Jordan Valley/Arid CLUTTON-BROCK, 1971, 1979 * 16 Jilat 7-1 Eastern Arid MARTIN, 1999 8 ʻAin Darat Southern Arid HORWITZ & SIMMONS, unpubl. 8 El Khiam Southern Arid DUCOS, 1997 23 Wadi Faynan 16 Southern Arid CARRUTHERS, 2002MPPNB 2 Kfar HaHoresh Western GORING-MORRIS et al., 1994-5 * 2 Yiftahʼel Western HORWITZ, 2003 9 Nahal Oren Western NOY et al., 1973 7 Abu Ghosh Mediterranean DUCOS & HORWITZ, 2003 5 ʻAin Ghazal Mediterranean ROLLEFSON & KOHLER-ROLLEFSON, 1993* 13 Beidha Mediterranean/Southern Arid HECKER, 1975 12 Aswad Beqaʼa –Jordan Valley DUCOS, 1993a,b 12 Ghoraife Beqaʼa –Jordan Valley DUCOS, 1993a,b 3 Munhatta Beqaʼa –Jordan Valley DUCOS, 1968 Beisamoun Beqaʼa –Jordan Valley DAVIS, 1982*
6 Jericho Beqaʼa –Jordan Valley/Southern Arid CLUTTON-BROCK, 1971, 1979* 16 Jilat 7-2,3,4 Eastern Arid MARTIN, 1999 8 El Khiam Southern Arid DUCOS, 1997 15 Ayn Abu Nukhayla Southern Arid HENRY et al., 2003
28 Wadi Tbeik Southern Arid TCHERNOV & BAR-YOSEF, 1982 * 29 Ujret el-Mehed Southern Arid DAYAN et al., 1986*LPPNB 5 ʻAin Ghazal Mediterranean ROLLEFSON & KÖHLER-ROLLEFSON, 1993 25 Wadi Shuʼeib Mediterranean SIMMONS et al., 2001
14 Basta Mediterranean/Southern Arid BECKER, 1991, 1997, 1998
11 Ramad 1 Beqaʼa –Jordan Valley DUCOS, 1993a,b 12 Ghoraife 1 Beqaʼa –Jordan Valley DUCOS, 1993a,b 17 Dhuweila 1 Eastern Arid MARTIN, 1999 22 Khirbet al Hammam Southern Arid PETERSON, 2004 23 Wadi Fidan A Southern Arid RICHARDSON, 1997PPNC 2 Yiftahʼel Western HORWITZ, 2003 9 Atlit Yam Western GALILI et al., 1993* 20 Ashkelon Western DUCOS, 1968 5 ʻAin Ghazal Mediterranean ROLLEFSON & KÖHLER-ROLLEFSON, 1993* 25 Wadi Shuʼeib Mediterranean SIMMONS et al., 2001 1 Hagoshrim 6 Beqaʼa –Jordan Valley HABER, 2001 10 Tel Teʼo Beqaʼa –Jordan Valley HORWITZ, 2001 3 Tel Ali Beqaʼa –Jordan Valley LEV-TOV, 2000
Counting Cattle : Trends in Neolithic Bos Frequencies from the Southern Levant 211

they are not cited here and only new references are listed (Table 2). Aside from ̒ Ain Ghazal, few Neolithic sites were occupied for more than one period or phase. Consequently, in order to investigate diachronic trends for the Southern Levant as a whole, coeval sites located in the same geographic area were grouped into five general zones. Three zones fall in the Mediterranean phytogeographic region : 1. Western Israel and the Mediterranean coastal plain. 2. The Mediterranean zone comprising northern Israel
and northern Jordan including the Jordan highlands as far south as Petra.
3. The Beqaʼa-Jordan Valley which constitutes ʻthe Levantine Corridor ̓(BAR-YOSEF, 2000).
Two zones encompass the desert margins :4. The eastern desert of Jordan including the Black and
Basalt deserts.5. The southern deserts – the Judean and Negev deserts
of Israel, southern Jordan and the Sinai Peninsula.The five areas listed above offer a more precise breakdown of the three climatic-topographic zones, that also differed in their modes of production, which were recognised by HORWITZ (2003) during the MPPNB : a. the Beqaʼa-Jordan Valley – characterised by very high
caprine frequencies – indicative of the onset of their incipient domestication, with little hunting of gazelle but cultivation of cereals and legumes ;
b. western Israel and the Mediterranean coastal plain – with an inverse pattern, indicating a focus on
gazelle hunting but with low caprine frequencies and evidence for agriculture especially of legumes ;
c. the eastern and southern desert regions – where hunting of gazelle and ibex continued as in the preceding Epi-Palaeolithic, associated with gathering of wild plants.
Several methodological issues pertaining to the current study need to be raised. For this research, the number of individual specimens (NISP counts) was preferred over estimates of the minimum number of individuals (MNIʼs). The latter data are seldom provided in Near Eastern Neolithic faunal reports. Moreover, given the variable methods and problems associated with the calculation of MNIʼs (GRAYSON, 1984), such estimates are clearly problematic. Related to this issue is the fact that NISP counts of a species are not necessarily the best indicator of the dietary importance of a taxon. This is especially true given the great disparity in meat yield between cattle and smaller sized gazelle or goats. In the absence of a standardised method for calculation of MNIʼs for all Neolithic sites that would enable a simple calculation of meat weights, these may be calculated from the numbers of bones following the method outlined in GRIGSON (1995). Consequently, the abundance of identified bones of each taxon has been used as an indicator of their relative importance in the diet. A further complication is that the relative frequencies of taxa are reported in the literature in a variety of fashions ;
PERIOD MAP NO. SITE ZONE REFERENCE 23 Wadi Fidan C Southern Arid RICHARDSON, 1997 16 Jilat 25 Eastern Arid MARTIN, 1999 16 Jilat 13 Eastern Arid MARTIN, 1999 18 Azraq 31 Eastern Arid MARTIN, 1999PN 9 Newe Yam Western HORWITZ, GALILI & LERNAU, in press 20 Ziqim Western GARFINKEL et al., 2002 21 Qatif Y3 Western/Southern Arid GRIGSON, 1983, 1995 26 Nahal Zehora Mediterranean DAVIS, in press 25 Jebel Abu Thawwab Mediterranean KAFAFI, 1988 25 Wadi Shuʼeib Mediterranean SIMMONS et al., 2001 5 ʻAin Ghazal Mediterranean ROLLEFSON & KÖHLER-ROLLEFSON, 1993* 7 Abu Ghosh Mediterranean HORWITZ, 2003 24 Labwe Beqaʼa –Jordan Valley BOKONYI, 1978 10 Tel Teʼo Beqaʼa –Jordan Valley HORWITZ, 2001 1 Hagoshrim Beqaʼa –Jordan Valley HABER, 2001 27 Shaʼar Hagolan Beqaʼa –Jordan Valley HESSE, 2002 3 Munhatta 2 Beqaʼa –Jordan Valley DUCOS, 1968 17 Dhuweila 1 Eastern Arid MARTIN, 1999 18 Burqu 27-2 Eastern Arid MARTIN, 1999 6 Jericho Beqaʼa –Jordan Valley/Southern Arid CLUTTON-BROCK, 1971, 1979*
212 L. K. HORWITZ & P. DUCOS

some reports exclude birds, fish and small mammals from the total identified fauna, while others include them as well as non-dietary items such as molluscs used for ornaments. To counter these inconsistencies, the relative frequencies for each taxon used in this paper have been calculated for each site and period from the sum of identified bones (NISP counts) for only three taxa (cattle, goat and gazelle). The archaeozoological samples from the PPNA through Late Pre-Pottery Neolithic B (LPPNB) periods have all been sieved, using 5 mm or smaller mesh sizes. However, most Pre-Pottery Neolithic C (PPNC) and PN samples were hand-collected or selectively sieved, usually with a large-sized mesh (5 cm or larger). Hence it is possible that in the PPNC and PN samples, cattle bones are over-represented, their larger, more easily identified bones, having been preferentially retrieved in the field.A taphonomic problem that is encountered in many sites, but which could not be accounted for in this study, is that the large-sized cattle bones have suffered a greater degree of fragmentation than smaller sized goat or gazelle bones (noted also by von den DRIESCH & WODTKE, 1997 : 530). Hence, fewer Bos bones were identified with certainty and a higher proportion of these remains are represented by unidentified shaft fragments. This is well illustrated in the LPPNB site of Basta where BECKER (2002) reported that cattle bones comprised only 4 % of all identified material. However, this frequency increased to 19 % when bone weights were used that included large sized fragments and damaged bones. Moreover due to breakage, few cattle bones can be measured, sexed or aged, limiting the type of information available on these issues.Finally, shifts in the exploitation pattern of a taxon may reflect changes in the environment such as climate change or decimation of the environment. Increased Bos frequencies may then indicate improved pasture conditions rather than the onset of domestication, while decreased frequencies may reflect the limited availability of pasture and degradation of the environment (DUCOS & HORWITZ, 1998). Consequently, examination of the climatic record, as well as charting coeval changes in gazelle and goat frequencies, may offer insights into the cause of the observed shift in cattle representation.
FINDINGS
Several published studies have addressed the issue of Bos exploitation in the prehistoric record of the Southern Levant. As shown by DAVIS (1982), prior to the PPNB cattle were infrequently exploited. According to TCHERNOV (1993), in the Kebaran they constituted 1-5 % of identified ungulate remains (mean = 4.7 %) ; in the Natufian, 0-6 % of all ungulate remains (mean = 3.0 %) and in the PPNA only 0-9.5 % of all identified ungulates (mean = 5.0 %). This frequency rose in the PPNB, attaining a mean of 11 % (range 0-43.0 %). However, in his study TCHERNOV
(1993) treated the PPNB as a single entity. In the absence of data for the sub-phases now recognised within this period (Table 1), it is not possible to pin-point the exact timing of this increase nor elucidate whether it was a localised or a region-wide occurrence. Examination of cattle remains from the LPPNB site of Basta (BECKER, 2002), revealed that in the uppermost levels cattle frequencies were double those of the lower levels (6 % compared to 3 %), illustrating the increased importance of this taxon over time. VON DEN DRIESCH & WODTKE (1997) also reported a slight increase in the frequency of cattle at ʻAin Ghazal, from 3.3 % of all identified remains in the Late MPPNB, to 3.8 % in the LPPNB peaking at 4.9 % in the PN (Yarmoukian phase). However, data for both Basta and ʻAin Ghazal are based on the sum of all identified remains and as such include relative frequencies of a wide range of taxa. Consequently, in all Neolithic phases at these sites, cattle actually constitute a minor component of the assemblage – less than 5 % of identified remains. The magnitude of changes in cattle exploitation patterns are thus masked by data on other taxa. By focusing solely on cattle, caprines and gazelle – these being the three major ungulates exploited in the Neolithic of the Southern Levant – as has been done in the present study, greater resolution is obtained in examination of diachronic trends.
REGIONAL VARIATION
Although some clear patterns emerge from comparison of cattle frequencies for the five different regions (Figs 2, 3 & 4), variation among contemporaneous sites within each zone is clearly evident. This may be due to differences in micro-environment of each site, recovery techniques, sample size and/or bone preservation. Moreover, periods and zones vary as to the number of sites they comprise such that sample sizes examined are not equal.
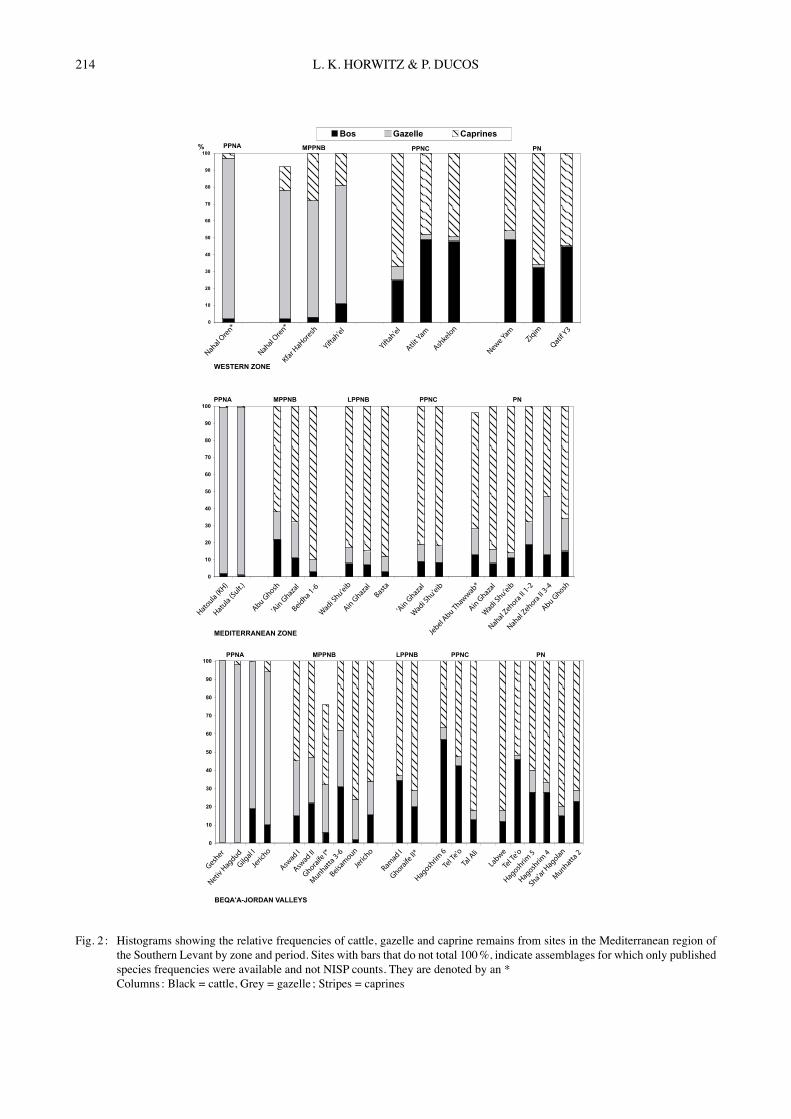
Western Israel-Coastal ZoneAs illustrated in Fig. 2a, for the zone encompassing Western Israel and the Mediterranean coastal plain, both PPNA and MPPNB sites are dominated by gazelle, with low frequencies of caprines and cattle (the latter comprising <5 % to 10 %). Only in the PPNC do caprines dominate the ungulate sample. Cattle frequencies are even lower than those of caprines in the PPNA (mean = 2.1 %) but show an increase in the MPPNB (mean = 9.6 %). Their numbers continue to rise, reaching frequencies ranging from 20 % to 50 % (mean = 47.1 %) in the PPNC (Figs 2a & 4). This high level is retained into the PN (mean = 45.7). PPNC and PN gazelle frequencies decrease even more as a result of the increased abundance of Bos. Unfortunately, no LPPNB assemblages are available from this region such that it is not possible to evaluate whether cattle numbers begin to increase in this period.
Counting Cattle : Trends in Neolithic Bos Frequencies from the Southern Levant 213
0
10
20
30
40
50
60
70
80
90
100
BEQA'A-JORDAN VALLEYS
PPNA MPPNB LPPNB PPNC PN
0
10
20
30
40
50
60
70
80
90
100
Bos Gazelle Caprines
WESTERN ZONE
PPNA MPPNB PPNC PN%
0
10
20
30
40
50
60
70
80
90
100
MEDITERRANEAN ZONE
PPNA MPPNB LPPNB PPNC PN
���������
��
���������
�����
��������
��
���������
���������
�������
��
�������
�
��������
�����
�������
�
����������
������
���������
�
���������
����
������
�����
�������
�����������
�����
�����
�����
����
������
��
�����������
�������
��������������
������
��
�����������
������������
�������
������������
�������
���������
������
�������
���������
������
�����
�������
��������
��������
���
��������
���������
����
�����
��
�������
��������
����������
�����
�����
���
�����
��
�����������
�����
�����
���
���������
���
���������
��������������
Fig. 2 : Histograms showing the relative frequencies of cattle, gazelle and caprine remains from sites in the Mediterranean region of the Southern Levant by zone and period. Sites with bars that do not total 100 %, indicate assemblages for which only published species frequencies were available and not NISP counts. They are denoted by an *
Columns : Black = cattle, Grey = gazelle ; Stripes = caprines
214 L. K. HORWITZ & P. DUCOS
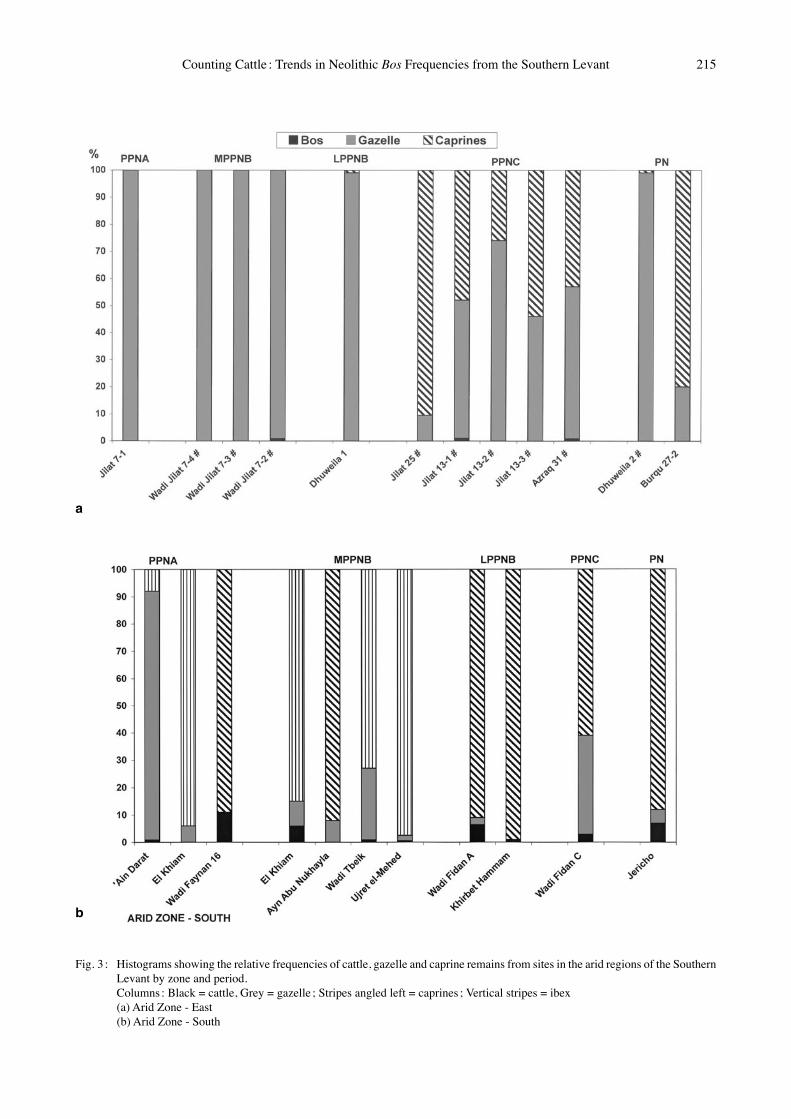
Fig. 3 : Histograms showing the relative frequencies of cattle, gazelle and caprine remains from sites in the arid regions of the Southern Levant by zone and period.
Columns : Black = cattle, Grey = gazelle ; Stripes angled left = caprines ; Vertical stripes = ibex (a) Arid Zone - East (b) Arid Zone - South
a
b
Counting Cattle : Trends in Neolithic Bos Frequencies from the Southern Levant 215
Mediterranean Zone PPNA assemblages in the Mediterranean zone are clearly dominated by gazelle with extremely low numbers of caprines (5 % or less) (Fig. 2b). In contrast, all MPPNB assemblages have extremely high frequencies of caprines, comprising over 50 % of ungulates exploited. This peak is retained into the LPPNB, PPNC and PN periods. A concomitant dramatic decrease is observed in gazelle frequencies over the same period of time. This shift has been associated in the Southern Levant with the onset of proto-domestication of goats or their introduction as domesticates and the near cessation of gazelle hunting (DAVIS, 1982 ; HORWITZ, 2003). In the Mediterranean zone, cattle frequencies are low in the PPNA (mean = 1.1 %), but increase noticeably in the MPPNB (mean = 12.2 %) (Fig. 4). However, between the MPPNB sites frequencies vary markedly with Abu Ghosh containing 20 % cattle while the coeval site of Beidha yielded <5 % cattle remains. In none of the subsequent
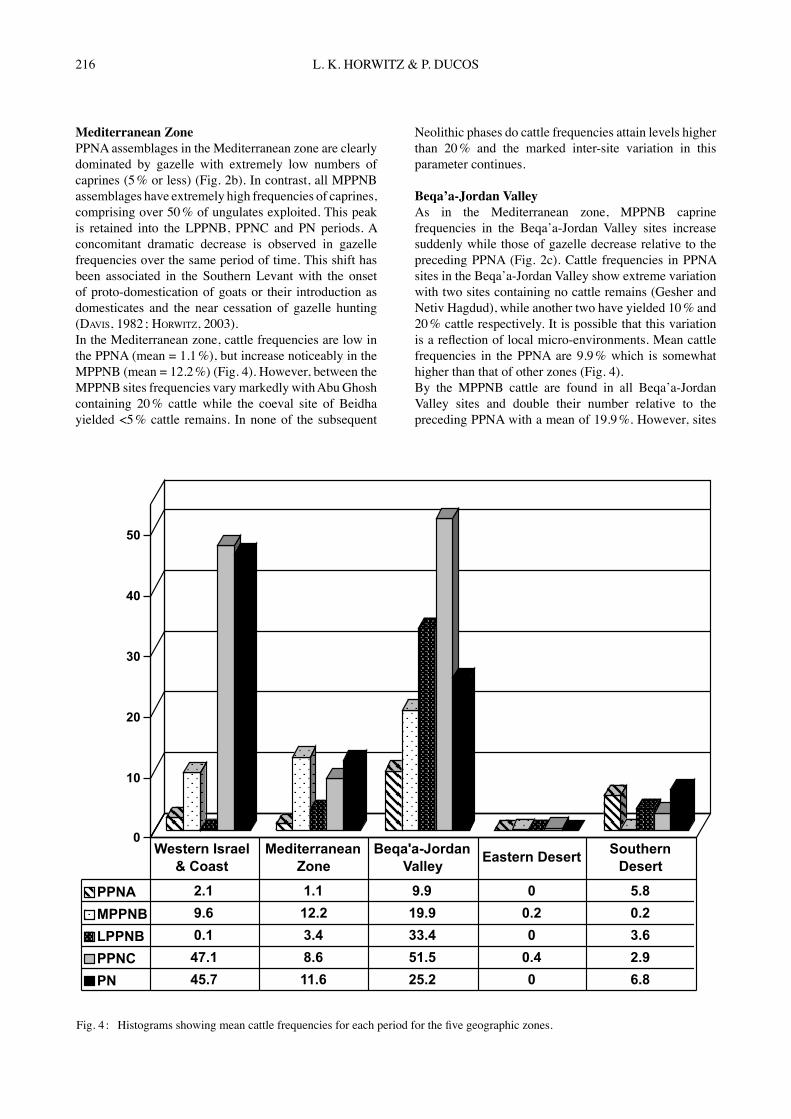
Neolithic phases do cattle frequencies attain levels higher than 20 % and the marked inter-site variation in this parameter continues. Beqaʼa-Jordan ValleyAs in the Mediterranean zone, MPPNB caprine frequencies in the Beqaʼa-Jordan Valley sites increase suddenly while those of gazelle decrease relative to the preceding PPNA (Fig. 2c). Cattle frequencies in PPNA sites in the Beqaʼa-Jordan Valley show extreme variation with two sites containing no cattle remains (Gesher and Netiv Hagdud), while another two have yielded 10 % and 20 % cattle respectively. It is possible that this variation is a reflection of local micro-environments. Mean cattle frequencies in the PPNA are 9.9 % which is somewhat higher than that of other zones (Fig. 4).By the MPPNB cattle are found in all Beqaʼa-Jordan Valley sites and double their number relative to the preceding PPNA with a mean of 19.9 %. However, sites
Fig. 4 : Histograms showing mean cattle frequencies for each period for the five geographic zones.
216 L. K. HORWITZ & P. DUCOS

continue to vary greatly, even those located in close proximity to each other. For example Beisamoun has <5 % cattle while ca. 30 % are found at Munhatta 3-6 (Figs 1 & 2c). During the LPPNB cattle frequencies continue to increase and achieve a mean value of 33.4 % for the period (Fig. 4). Examination of sites from a circumscribed geographic area within the valley offers good resolution of this diachronic trend. In the Damascus Basin a significant increase in cattle frequencies is found between the MPPNB sites (Aswad I, II Ghoraife I) and those of the LPPNB (Ramad, Ghoraife II) (Fig. 2c). For the Beqaʼa-Jordan Valley region as a whole, frequencies in most PPNC sites are extremely high with a mean of 51.5 % (Figure 4). However, inter-site variation is evident both here and in the subsequent PN, where for example 50 % cattle are found at Tel Teʼo but only some 10 % in Labwe and Shaʼar Hagolan. Overall, cattle frequencies in the PN decrease compared to the earlier MPPNB and PPNC (PN mean = 25.2 %). This is observed even in the same site e.g. Munhatta and Tel Teʼo.
Arid Zone – EastMARTIN (1999) has described the faunal trends for sites in the eastern desert region of Jordan. Until the LPPNB, gazelle is almost the sole ungulate exploited – exceptions are isolated bones of Bos in Wadi Jilat 7-2, and of Capra in Dhuweila (Fig. 3a). A shift of great magnitude is observed in this region in the PPNC, when caprines begin to comprise a significant portion of the fauna. Negligible numbers of cattle remains are found in all period with means ranging from 0 to 0.4 % (Fig. 4). In PPNC Jilat 13-1 and Azraq 31 the presence of cattle has been explained by MARTIN (1999) as representing the introduction of domestic animals into the region. No cattle remains are found in the PN samples. However, the two assemblages attributed to this period are both located in an extremely arid region that may not support Bos (Figs 1 & 3a).
Arid Zone - South With the exception of the PPNA site of ʻAin Darat, where gazelle are the dominant species, irrespective of period, goats are the dominant taxon in this zone. As shown in Fig. 3b, the arid dwelling ibex (Capra ibex) has been identified in four of the sites from this climatic-topographic zone (ʻAin Darat, El Khiam, Wadi Tbeik, Ujret el-Mehed). As demonstrated by HECKER (1975) for Beidha, and suggested by CLUTTON-BROCK (1979) with reference to Jericho, it is possible that both wild bezoar goat (Capra aegagrus) and ibex co-occur in sites in the desert margins, but that in most instances their remains have not have been separated. In the southern desert margins cattle comprise a small component of the ungulate fauna. In the PPNA the high mean value of 5.8 % is due to the abundance of Bos in the Wadi Fidan sites – ca. 10 % of the assemblages in
both the PPNA and LPPNB (Fig. 3b). However, in the MPPNB to PPNC the means remain low, ranging from 0.2 % to 2.9 % respectively. A substantial increase in the number of cattle occurs by the PN, where the mean value peaks at 6.8 %.
DIACHRONIC TRENDS
PPNA sites, with no osteological evidence for domesticated animals provided base-line data of the relative frequency of game animals hunted prior to the advent of domestication. As illustrated in Figures 2, 3 and 4, all PPNA sites are dominated by hunted taxa ; those in the Mediterranean zone by high frequencies of gazelle and the two arid zone sites by goat – Capra ibex at El Khiam and probably Capra aegagrus at Faynan 16. The frequency of cattle varies according to the geographic location of the sites, from <5 % in Hatoula (Mediterranean zone) to 20 % in Gilgal (Beqaʼa-Jordan Valley). Indeed, the highest Bos frequencies for PPNA sites occur in assemblages located in the Beqaʼa-Jordan Valleys and its southern-most extension south of the Dead Sea (Gilgal I, Jericho and Wadi Faynan 16). However, even at their numerical peak, cattle remains in all sites in this period are far fewer than those of gazelle (ratio of 1 :4).Data for the Early PPNB is not shown here since this period occurs only in three sites in the Southern Levant ; Horvat Galil situated close to the Lebanese border in the north-west of Israel, Motza located on the outskirts of the city of Jerusalem and Jilat in the eastern desert of Jordan. No Bos remains were found in Horvat Galil (HORWITZ, unpublished data) or in Jilat (MARTIN, 1999), while at Motza they constituted only 1.4 % of the identified sample (SAPIR, 2005). As such the EPPNB closely resembles the preceding PPNA in patterns of cattle exploitation.The MPPNB is characterised by a rise in the proportion of cattle in the Mediterranean zones in general, in Western Israel and the coastal plain, as well as in the Beqaʼa-Jordan Valley. The magnitude of this increase is circa. 10 % (Fig. 4).However, a high degree of inter-site variation is observed in each zone. Sites in the arid regions contain low numbers of cattle remains (Fig. 3a, b). It is interesting to note that the geographic patterning in frequencies observed for cattle, corresponds to that previously reported for caprines in the MPPNB. This is clearly illustrated in Fig. 2b. The highest numbers of caprines are found in sites in the Jordan-Beqaʼa Valley (termed the Levantine Corridor – BAR-YOSEF, 2000). These sites also contain the lowest gazelle numbers, while an inverse pattern is found in sites located in western Israel. The desert sites in eastern Jordan are dominated by gazelle, while those in the southern deserts and Sinai Peninsula have high frequencies of goats – ibex and bezoar goat mixed, or only ibex (HECKER, 1975 ; HORWITZ, 1993, 2003). The augmented frequencies of
Counting Cattle : Trends in Neolithic Bos Frequencies from the Southern Levant 217

MPPNB caprines in sites in the Levantine Corridor has been interpreted as either representing (i) the zone where authochtonous domestication of goats first took place following diffusion of the concept southwards from the Northern Levant (HORWITZ, 2003), or else (b) the southward dispersion of domestic stock from the north (BAR-YOSEF, 2000). As shown in Fig. 2b, the high frequencies of both Bos and caprines in sites in the Levantine Corridor are undoubtedly related to the high carrying capacity of this zone which offers plentiful water resources and pasture (TCHERNOV & HORWITZ, 1990). This zone would then have offered the most amenable physical conditions for both wild animals as well as for their domestication. If however animals were physically introduced into the Southern Levant from southeastern Anatolia, then the Beqaʼa-Jordan Valley represents a natural conduit, having served since the Late Epipaleolithic as a trade route (BAR-YOSEF, 2000). Notably, both caprine and cattle frequencies in the Valley sites increase in the MPPNB relative to earlier periods, but Bos shows less change. Gazelle frequencies drop, indicating that their importance decreased.Data for cattle in the LPPNB are uneven due to the small number of sites available. This may account for the drop in mean values in the Mediterranean zone and absence of Bos in sites in the eastern desert margin. However, in the Beqaʼa-Jordan Valley mean values increase from 19.9 % to 33.4 % and in the Southern desert margin they rise from 0.2 % to 3.6 %. Inter-site variation is clearly evident (Fig. 2a-e) with the highest Bos frequencies (20 %-30 %) in the Levantine Corridor sites, especially those located furthest to the north (Aswad, Ramad). A second, major increase in the frequency of cattle remains is documented for all zones in the PPNC, where frequencies attain unprecedented high levels of 40 % to 50 % (Figs 2 & 3). This trend is clearly evident in sites situated in Western Israel (mean = 47.1 %), the Mediterranean zone (mean = 8.6 %) and Beqaʼa-Jordan Valley (mean =51.5 %) (Fig. 4). The PPNC also marks the re-appearance of Bos in the eastern deserts of Jordan (Jilat 13-1, Azraq 31), suggested by MARTIN (1999) to represent domestic animals, while those in the preceding MPPNB were aurochs. In the southern desert margin, mean values decrease slightly (from 3.6 % to 2.9 %), although in some southern sites on the desert margin such as Wadi Fidan, frequencies remain almost unchanged relative to the MPPNB.By the PN, cattle comprise over 20 % of herd animals exploited in at least half the sites investigated in this study (Figs 2 & 3). In all zones, mean values for cattle plateau or decrease with the exception of the Mediterranean zone where the mean value for Bos increases from 8.6 % to 11.6 % (Fig. 4). Due to poor bone preservation, it is unclear whether cattle had dispersed into the southern arid interior by the PN. The high frequencies of their remains at Qatif Y3, located in the Gaza Strip (Fig. 1), indicates that they were present at least on the southern coast.
Based on the data presented here it is clear that a gradual increase in the proportion of Bos occurred in the Southern Levant especially evident in the three Mediterranean zones – western Israel, northern Israel and Jordan and the Beqaʼa-Jordan Valley. Evidence for this trend may be observed in the Damascus Basin sites (Ghoraife, Aswad, Ramad) where an increase in cattle frequencies is found between the MPPNB and LPPNB. However, there is no indication that these animals had undergone changes in size, sex ratios or mortality profiles (DUCOS, 1993, 1995). By the PPNC a major shift in Bos exploitation patterns had occurred. In this period cattle comprised a significantly higher frequency of ungulates than exploited before and their remains are found in sites throughout the Southern Levant including the arid margins. This suggests a gradual dispersion of domestic cattle, either as a local innovation or due to the physical movement of domestic stock from the core Mediterranean area outwards.Caprines follow a similar diachronic trend to that of cattle. In the MPPNB, caprine frequencies are significantly higher than in previous periods. This is especially evident in sites in the Beqaʼa-Jordan Valley sites. By the LPPNB this trend is widespread throughout the region, although it is only by the PPNC that caprines are commonly found in sites even in the desert zones (HORWITZ, 1993, 2003 ; MARTIN, 1999). Gazelle frequencies show an inverse linkage to those of caprines and cattle, decreasing as the others numbers increased. According to HOROWITZ (1992) and SANLAVILLE (1996), at the onset of the PPNA climatic conditions were warm and dry improving by the latter part of the PPNB which experienced warm, pluvial conditions with summer rainfall. Recently, KISLEV et al. (2004) reported that cooler more humid conditions characterised the PPNC. Thus, it is possible that the trend of increasing Bos frequencies observed here reflects the greater availability of suitable pasture and water resources under conditions of climatic amelioration relative rather than domestication. The main argument to counter this is that there is a patent decline in gazelle numbers, as well as those of small game, over the same time period (HORWITZ, 1996 ; HORWITZ & TCHERNOV, 1998 ; MUNRO, 2004). This offers unequivocal proof that hunting of wild animals decreased in importance in Neolithic societies. Consequently, the increased Bos frequencies observed here can only reflect a focus on this taxon with the advent of domestic cattle rather than more intensive hunting of aurochs.
DISCUSSION
Over the past decade, DNA analyses have demonstrated that domestic cattle had a dual, if not tripartite origin (BRADLEY et al., 1996, 1998 ; HANOTTE et al., 2002). Domestic cattle (Bos taurus) of the Near East and Europe arose from a common Near Eastern ancestor, the aurochs,
218 L. K. HORWITZ & P. DUCOS

Bos primigenius primigenius (TROY et al., 2001). This species appears to have enjoyed a broad Palaearctic distribution until its extinction in the 17th century. The presence of aurochs in the arid regions of the Southern and Eastern Levant has led researchers to suggest that it could tolerate fairly dry conditions and inhabited forests as well as open scrub, probably being adapted to both grazing and browsing (CLUTTON-BROCK, 1999 ; UERPMANN, 1987). However like cattle, aurochs probably required frequent access to water a factor that would have severely restricted their geographic range, probably more so than the available vegetation cover (UERPMANN, 1987). These physiological characteristics find some corroboration in recent genetic data which demonstrate the limited genetic diversity of the ancestral Near Eastern aurochs. This suggests that they originate from a single, geographically restricted nuclear zone, possibly constrained in its range due to climate (TROY et al., 2001). Current archaeozoological research points to the Northern Levant as the earliest centre of cattle domestication (HELMER et al., 2005 ; PETERS et al., 1999). Despite the importance of cattle in the iconography of the PPNA and PPNB periods in the Northern Levant (HELMER et al., 2004 ; PETERS & SCHMIDT, 2004), the frequency of skeletal remains of Bos in these periods is consistently low, ranging from 5 % to 20 % in the PPNA and EPPNB (HELMER et al., 2005). An exception is the EPPNB level IVA at Mureybet where cattle frequencies attain a high 30 % to 40 % and have been interpreted as representing selective hunting or “protoélevage” (DUCOS, 1978). Corroboration for this hypothesis may be found in the EPPNB site of Djaʼde. It has been proposed that at this site, cattle may have been undergoing proto-domestication as reflected by their intermediate position in sex ratio, body size and extent of sexual dimorphism relative to wild and domestic cattle from other sites in the region (HELMER et al., 2005). Only in the MPPNB site of Tell Halula are cattle remains encountered that clearly exhibit the morphological and metrical changes associated with domestication, a reduced degree of sexual dimorphism as well as increased frequencies (HELMER et al., 2005 ; SAÑA SEGUI, 1999). However, even here the frequency of Bos remains continues to be low (less than 20 % of all identified species), and does not show a distinct increase in the LPPNB (SAÑA SEGUI, 1999). According to HELMER and colleagues, this may reflect their exploitation primarily for milk products rather than for meat (HELMER et al., 2005). Cattle are first found on Cyprus in EPPNB contexts. Since Bos was not part of the Pleistocene fauna of this island, its presence in Neolithic sites clearly indicates that it was introduced (VIGNE et al., 2003). Many researchers accept this as proof of their domestic status despite the fact that on the mainland no conclusive evidence for morphologically domestic Bos has been found in EPPNB contexts. Moreover, the age profiles, morphology and biometry of the Cypriot cattle do not
offer equivocal support for their domestic status. An alternative explanation has been offered by HORWITZ et al. (2004), namely that the cattle represent wild animals released on the island as a food source. This is especially plausible since Bos became extinct on Cyprus by the end of PPNB and was reintroduced as a domesticate only in the Bronze Age (CROFT, 1991).In the Southern Levant, cattle do not feature as an important element in the iconography of the PPNA period. However, they comprise a significant ritual and artistic element in the MPPNB and LPPNB (GARFINKEL, 1995 ; GORING-MORRIS & HORWITZ, in press and references therein). Despite the clear shift in the symbolic importance of cattle, the archaeozoological evidence for their domestication has been ambiguous but as we show here, their importance as a source of food increased markedly during this period.The earliest osteological evidence for domestic cattle in the Southern Levant is from the LPPNB sites of Basta, Baʼja and ʻAin Ghazal (BECKER, 2002 ; VON DEN DRIESCH & WODTKE, 1997 ; HORWITZ et al., 1999). For LPPNB Basta, BECKER (2002) has presented a cogent argument in favour of the domestic status of cattle at this site. There are a majority of females in the assemblage, but sample sizes are small. Likewise, the small dental sample used for ageing suggests a predominance of young cattle but data on post-cranial bone fusion show no coherent patterning. The small increase documented in cattle frequencies, as noted above, may also demonstrate the increased importance of this taxon over time. Most importantly, the Basta cattle display a broad range of size variation that has been interpreted as indicating a mixed domestic cattle-aurochs component. On the basis of the small size of cattle at Baʼja, it has been suggested that they may represent domesticates, but the sample size of measurable bones is small (HORWITZ et al., 1999). The high frequency of juvenile animals in the PPNB at ʻAin Ghazal has been interpreted as evidence of an attempt to domesticate local aurochs, an endeavour which then failed since body size increased between the PPNB and PN (von den DRIESCH & WODTKE, 1997).The size index of cattle bones from Basta shows them to be smaller than Northern Levantine PPNA and EPPNB specimens, but of similar size to both MPPNB animals from Tell Halula and a combined sample of LPPNB/PPNC cattle from ʻAin Ghazal. However, the influence of geographic clines (BERGMANʼs Rule) as well as varying sex ratios in the different sites need to be accounted for before these results can be used to argue in favour of the domestic status of the Basta animals, while the use of a mixed LPPNB/PPNC sample from ʻAin Ghazal may mask inter-period size variation at this site. Data on sex ratios and age profiles in Bos from Neolithic site in the Southern Levant are limited and vary greatly between sites. Although both BECKER (2002) and VON DEN DRIESCH & WODTKE (1997) report high frequencies of females and juveniles in MPPNB assemblages, at other
Counting Cattle : Trends in Neolithic Bos Frequencies from the Southern Levant 219

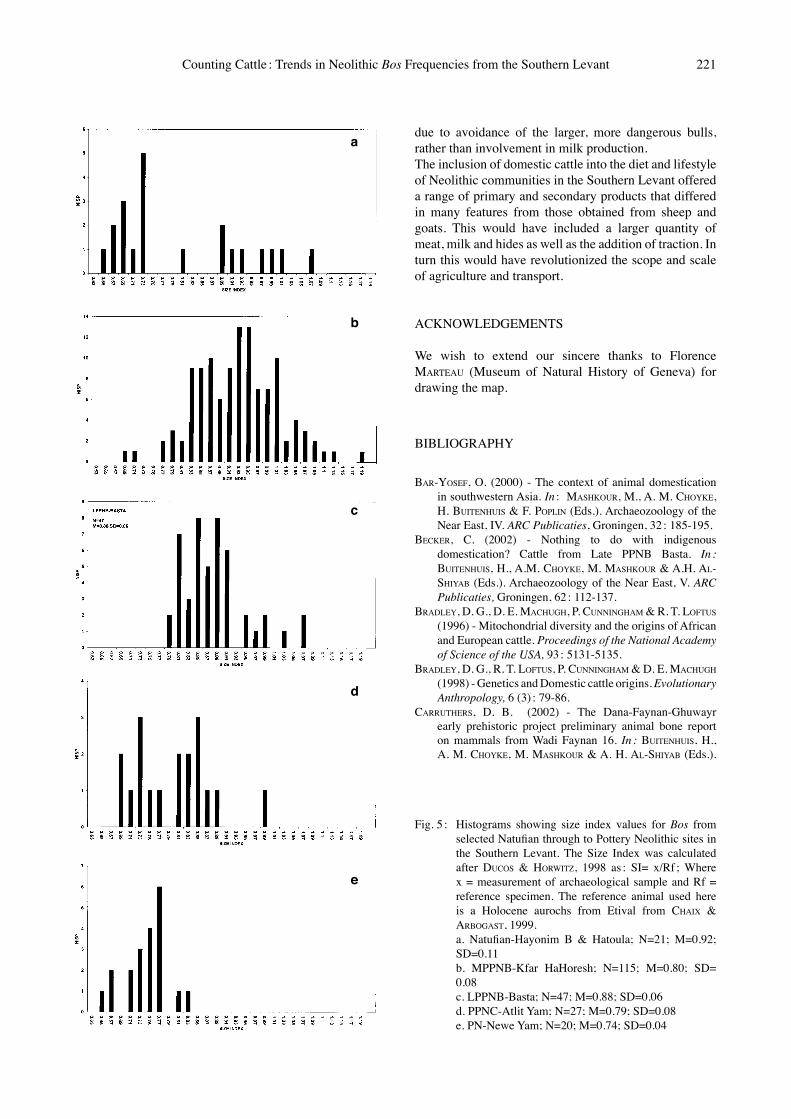
sites a predominance of adults was found (e.g. HORWITZ, 2003). A similar dominance of adult animals is noted for PPNC sites (e.g. GARFINKEL et al., in press ; HABER, 2001). When the size indices for Bos are examined (Fig. 5) it is evident that none of the samples exhibit clear bimodal peaks representing males and females, suggesting that they contain unequal sex ratios. This is a point that requires further investigation, although many faunal assemblages are limited by the small numbers of bones that may be aged or sexed (e.g. CARRUTHERS, 2002 ; DUCOS, 1995 ; DUCOS & HORWITZ, 2003 ; HORWITZ, 2003). In a diachronic study of body size of several taxa from the Southern Levant, DAVIS (1981) reported a significant size diminution in Bos following the MPPNB which he linked to their domestication. However, the data set used by DAVIS lacked samples from all sub-phases of the PPNB, especially the critical LPPNB and PPNC. Examination of a broader Near Eastern data-base by GRIGSON (1989), illustrated a significant reduction in cattle body size in Southern Levantine sites beginning in the 6th millennium B.C. i.e. the Terminal PPNB/PPNC, and resulting in a definite diminution by the 5th millennium B.C. This size reduction was associated with an increase in the proportion of females. In a more recent study, DUCOS & HORWITZ (1998) reported a diminution in Bos in the PPNC. The magnitude of size reduction observed in these cattle, which is evident in both the mean and the range of the size index, was interpreted as probably signifying their domestic status. However, the sample sizes used in this study are small and no LPPNB assemblage was included. Moreover, the PPNC animals show little change in mortality profiles or skeletal morphology that would support their identification as domestic (HORWITZ et al., 1999 ; GARFINKEL et al., in press). At ʻAin Ghazal the smallest animals were found in the PPNC, but size increased in the PN (VON DEN DRIESCH & WODTKE, 1997), while at Jericho no size change was reported for cattle between the Pre-Pottery Neolithic and PN levels at the site (CLUTTON-BROCK, 1971).Fig. 5 presents a graphic illustration of cattle size indices from a series of Southern Levantine sites spanning the Natufian through Pottery Neolithic periods. Again sample sizes are small, but it is evident that the Natufian sample, comprising aurochs from Hayonim Cave (Level B) and Hatoula, presents the full size range of Bos primigenius in the region. The animals recovered from a single pit at the MPPNB site of Kfar HaHoresh fall within the Natufian range, as do those from LPPNB Basta. In this respect, cattle follow the same pattern as observed for Natufian through MPPNB gazelle, but differ from caprines (DUCOS & HORWITZ, 1998) suggesting that like gazelle, MPPNB cattle still represented hunted elements or were in the early stages of proto-domestication. Most Natufian to LPPNB Bos were significantly larger than any represented in the PPNC from Atlit Yam or PN from Newe Yam (Fig. 5). The apparent diminution in body size (both in the range of size as well as the mean) in these site
undoubtedly indicates the presence of domestic cattle at this time, post-dating diminution observed in caprines.
CONCLUSIONS
Based on trends in the relative frequency of the three main ungulate taxa, it is evident that a diachronic increase in the numbers of cattle exploited began in the Southern Levant in the MPPNB i.e. at the same time as caprine frequencies show a major increase. This is especially evident in sites located in the Mediterranean region of the Southern Levant. This trend cannot be explained by climate change and probably represents the onset of local, proto-domestication of cattle i.e. focussed hunting or corralling of incipient domestic stock or alternately, the introduction of domestic animals. The highest cattle frequencies are found in sites in the Beqaʼa-Jordan Valley (i.e the Levantine Corridor), indicating that local communities were influenced by those in the Middle Euphrates where cattle domestication was already advanced by the MPPNB (SAÑA SEGUI, 1999). However, it is only some 1000 years later, by the PPNC, that there is substantial evidence for the widespread occurrence of domestic cattle throughout the Southern Levant, manifest in augmented bone frequencies and a marked diminution in their size. This indicates a significant lag period between the first appearance of morphologically distinct domestic cattle in the Northern Levant and those in the south. Unfortunately data on mortality profiles and sex ratios are lacking for most Southern Levantine sites, but there are some hints that by the LPPNB selection of juveniles and females was practised. The large inter-site variation and small sample sizes available however dictate that these data be treated with some caution.At this point in time the available data do not appear to be sufficient or rigorous enough to provide a definitive answer as to whether cattle were domesticated locally in the Southern Levant or imported. BECKER (2002) considered that domestic Bos were introduced into the Petra region, but did not negate the possibility of authochtonous domestication in the Southern Levant at sites such as ̒ Ain Ghazal. As noted by her, local communities would have had extensive experience of aurochs from hunting since this taxon inhabited the region. Moreover, they would have amassed knowledge of domestication from herding caprines which predated the advent of domestic cattle by ca. 500 years (HORWITZ, 1993, 2003 ; HORWITZ & DUCOS, 1998 ; HORWITZ et al., 1999 ; MARTIN, 1999). There is no evidence to support or contradict the idea that cattle were primarily exploited for milk in the initial stages of their domestication, as suggested by HELMER et al. (2005) for the Northern Levant. Cattle frequencies in the Southern Levant increase substantially, such that they would have constituted a significant meat resource. Despite the paucity of data on age and sex profiles, preferential selection of young and/or female animals may have been
220 L. K. HORWITZ & P. DUCOS
due to avoidance of the larger, more dangerous bulls, rather than involvement in milk production. The inclusion of domestic cattle into the diet and lifestyle of Neolithic communities in the Southern Levant offered a range of primary and secondary products that differed in many features from those obtained from sheep and goats. This would have included a larger quantity of meat, milk and hides as well as the addition of traction. In turn this would have revolutionized the scope and scale of agriculture and transport.
ACKNOWLEDGEMENTS
We wish to extend our sincere thanks to Florence MARTEAU (Museum of Natural History of Geneva) for drawing the map.
BIBLIOGRAPHY
BAR-YOSEF, O. (2000) - The context of animal domestication in southwestern Asia. In : MASHKOUR, M., A. M. CHOYKE, H. BUITENHUIS & F. POPLIN (Eds.). Archaeozoology of the Near East, IV. ARC Publicaties, Groningen, 32 : 185-195.
BECKER, C. (2002) - Nothing to do with indigenous domestication? Cattle from Late PPNB Basta. In : BUITENHUIS, H., A.M. CHOYKE, M. MASHKOUR & A.H. AL-SHIYAB (Eds.). Archaeozoology of the Near East, V. ARC Publicaties, Groningen, 62 : 112-137.
BRADLEY, D. G., D. E. MACHUGH, P. CUNNINGHAM & R. T. LOFTUS (1996) - Mitochondrial diversity and the origins of African and European cattle. Proceedings of the National Academy of Science of the USA, 93 : 5131-5135.
BRADLEY, D. G., R. T. LOFTUS, P. CUNNINGHAM & D. E. MACHUGH (1998) - Genetics and Domestic cattle origins. Evolutionary Anthropology, 6 (3) : 79-86.
CARRUTHERS, D. B. (2002) - The Dana-Faynan-Ghuwayr early prehistoric project preliminary animal bone report on mammals from Wadi Faynan 16. In : BUITENHUIS, H., A. M. CHOYKE, M. MASHKOUR & A. H. AL-SHIYAB (Eds.).
Fig. 5 : Histograms showing size index values for Bos from selected Natufian through to Pottery Neolithic sites in the Southern Levant. The Size Index was calculated after DUCOS & HORWITZ, 1998 as : SI= x/Rf ; Where x = measurement of archaeological sample and Rf = reference specimen. The reference animal used here is a Holocene aurochs from Etival from CHAIX & ARBOGAST, 1999.
a. Natufian-Hayonim B & Hatoula; N=21; M=0.92; SD=0.11
b. MPPNB-Kfar HaHoresh; N=115; M=0.80; SD= 0.08
c. LPPNB-Basta; N=47; M=0.88; SD=0.06 d. PPNC-Atlit Yam; N=27; M=0.79; SD=0.08 e. PN-Newe Yam; N=20; M=0.74; SD=0.04
a
b
c
d
e
221Counting Cattle : Trends in Neolithic Bos Frequencies from the Southern Levant

Archaeozoology of the Near East, V. ARC Publicaties, Groningen, 62 : 93-97.
CHAIX, L. & R.-M. ARBOGAST (1999) - Holocene Aurochs from western Europe : Osteometrical data. In : WENIGER, G.-C. (Ed.). Archaeologie und Biologie des Aurochsen. Wissenschaftliche Schriften des Neanderthal Museums, 1 : 35-48.
CLUTTON-BROCK, J. (1979) - The mammalian remains from the Jericho Tell. Proceedings of the Prehistoric Society, 45 : 135-157.
CLUTTON-BROCK, J. (1999) - Natural History of Domesticated Mammals, 2nd edition. Cambridge University Press, Cambridge.
CROFT, P. (1991) - Man and beast in Chalcolithic Cyprus. Bulletin of the American School of Oriental Research, 282-283 : 63-79.
DAVIS, S. J. M. (1981) - The effects of temperature change and domestication on body size of Late Pleistocene to Holocene mammals of Israel. Paleobiology, 7 : 101-114.
DAVIS, S. J. M. (1982) - Climatic change and the advent of domestication : the succession of ruminant artiodactyls in the late Pleistocene-Holocene in the Israel region. Paléorient, 8 : 5-15.
DAVIS, S. J. M. (in press) - The animal bones from Nahal Zehora I and II. In : GOPHER, A. (Ed.). Archaeological Investigations at Nahal Zehora : Villages of the Pottery Neolithic in the Menashe Hills, Israel. Institute of Archaeology, Tel Aviv University.
DRIESCH, A. VON DEN & U. WODTKE (1997) - The fauna of ʻAin Ghazal, a major PPN and early PN settlement in central Jordan. In : GEBEL, H. G. K., Z. KAFAFI & G. O. ROLLEFSON (Eds.). The Prehistory of Jordan II. Perspectives from 1997. Studies in Early Near Eastern Production, Subsistence and Environment, Berlin, 4 : 511-556.
DUCOS, P. (1968) - LʼOrigine des Animaux Domestiques en Palestine. Publications de lʼInstitut de Préhistoire de lʼUniversité de Bordeux, Memoire 6.
DUCOS, P. (1978) - Tell-Mureybet (Syrie, IXe-VIIe millénaires). Etude archeozoologique et précis dʼécologie humaine. CNRS Editions, Lyon.
DUCOS, P. (1993) - Proto-élevage au Levant sud au VIIe millenaire BC. Les données de la Damascene. Paléorient, 19 : 153-174.
DUCOS, P. (1995) - Note préliminaire sur les faunes dʼAswad et Ghoraife. In : DE CONTENSON, H. (Ed.). Aswad et Ghoraife sites Neolithiques en Damascene (Syrie) aux IXeme et VIIIeme millénaires avant lʼère Chrétienne. Bibliothèque Archéologique et Historique T CXXXVII. Institut Francais dʼArchéologie du Proche-Orient, Beyrouth : 339-349.
DUCOS, P. (1997) - A re-evaluation of the fauna from the Neolithic levels of El-Khiam. Journal of the Israel Prehistoric Society, 27 : 75-81.
DUCOS, P. & L.R.K. HORWITZ (1998) - The influence of climate on artiodactyl size during the late Pleistocene-early Holocene of the southern Levant. Paléorient, 23 (2) : 229-247.
DUCOS, P. & L.K. HORWITZ (2003) - The Pre-Pottery Neolithic B fauna from the Lechevalier excavations at Abu Ghosh. In : KHALAILY, H. & O. MARDER (Eds.). The Neolithic Site of Abu Ghosh. The 1995 Excavations. Israel Antiquities Authority Reports, Jerusalem, 19 : 103-119.
ENLOE, J. G. (1999) - Hunting specialization : Single-species locus and human adaptation. In : BRUGAL, J.-P., F. DAVID, J. G. ENLOE & J. JAUBERT (Eds). Le Bison : Gibier et moyen de
subsistence des hommes du Paléolithique aux paléoindiens des grandes plaines. Editions APDCA, Antibes : 501-509.
GARFINKEL, Y. (1995) - Human and Animal Figurines of Munhatta (Israel). Les Cahiers de Missions Archéologiques Français en Israël, 8, Paléorient. Paris.
GARFINKEL, Y., D. DAG, L. K. HORWITZ, O. LERNAU & H. K. MIENIS (2002) - Ziqim, A Pottery Neolithic site in the southern coastal plain of Israel. A final report. Journal of the Israel Prehistoric Society, 32 : 73-145.
GARFINKEL, Y., D. DAG, B. HESSE, P. WAPNISH, D. ROOKIS, G. HARTMAN, D.E. BAR-YOSEF MAYER & O. LERNAU (2005) - Neolithic Ashkelon : meat processing and early pastoralism on the Mediterranean coast. Eurasian Prehistory, 3 (1) : 43-72.
GORING-MORRIS, N. & L. K. HORWITZ (in press) - Funerals and feasts during the Pre-Pottery Neolithic B of the Near East. Antiquity.
GRAYSON, D. K. (1984) - Quantitative Zooarchaeology. Topics in the Analysis of Archaeological Faunas. Academic Press, London.
GRIGSON, C. (1983) - Preliminary report on the mammal bones from Neolithic Qatif, site Y3 on the Sinai coastal plain (excavations of 1979, 1980 and 1983). Unpublished manuscript.
GRIGSON, C. (1989) - Size and sex : evidence for the domestication of cattle in the Near East. In : MILLES, A. D., D. WILLIAMS & N. GARDNER (Eds). The Beginnings of Agriculture. BAR International Series, Oxford, 496 : 77-109.
GRIGSON, C. (1995) - Plough and Pasture in the early economy of the southern Levant. In : LEVY, T. E. (Ed.). The Archaeology of Society in the Holy Land. Leicester University Press, London : 245-268.
HABER, A. (2001) - The Faunal Analysis of Hagoshrim : Biological and Economical Aspects of Prehistoric Agricultural Societies and the process of Domestication. Unpublished M.Sc Thesis, Tel-Aviv University (in Hebrew).
HANOTTE, O., D.G. BRADLEY, J.W. OCHIENG, Y. VERJEE, E.W. HILL & J.E. REGE (2002) - African pastoralism : genetic imprints of origins and migrations. Science, 296 : 336-339.
HECKER, H. (1975) - The Faunal Analysis of The Primary Food Animals from Pre-Pottery Neolithic Beidha (Jordan). University Microfilms International, Ann Arbor : Michigan.
HELMER, D., L. GOURICHON & D. STORDEUR (2004) - A lʼaube de la domestication animale. Imaginaire et symbolisme animal dans les premières sociétés néolithiques du nord du Proche-Orient. Anthropozoologia, 39 (1) : 143-163.
HELMER, D., L. GOURICHON, H. MONCHOT, J. PETERS & M. SANA SEGUI (2005) - Identifying early domestic cattle from Pre-Pottery Neolithic sites on the Middle Euphrates using sexual dimorphism. In : VIGNE J.-D., D. HELMER & J. PETERS (Eds.). The First Steps of Animal Domestication. Oxbow Books, Oxford : 86-95.
HENRY, D. O., C. CARDOVA, J. J. WHITE, R. M. DEAN, J. E. BEAVER, H. EKSTROM, S. KADOWAKI, J. MCCORRISTON, J. NOWELL & L. SCOTT-CUMMINGS (2003) - The early Neolithic site of Ayn Abu Nukhayla, Southern Jordan. Bulletin of the American School of Oriental Research, 330 : 1-30.
HESSE, B. (2002) - Between the revolutions : Animal use at Shaʼar Hagolan during the Yarmukian. In : GARFINKEL, Y. & M.A. MILLER (Eds.). Shaʼar Hagolan, I. Neolithic Art in Context. Oxbow Books, Oxford : 247-255.
222 L. K. HORWITZ & P. DUCOS

HOROWITZ, A. (1992) - Palynology of Arid Lands. Elsevier : Amsterdam.
HORWITZ, L. K. (1993) - The development of ovicaprine domestication during the PPNB of the Southern Levant. In : BUITENHUIS, H. & A. T. CLASON (Eds). Archaeozoology of the Near East, I. Universal Book Services, Leiden : 27-36.
HORWITZ, L. K. (1996) - The impact of animal domestication on species richness : A pilot study from the Neolithic of the southern Levant. ArchaeoZoologia, 8 (1-2) : 53-70.
HORWITZ, L. K. (2001) - The mammalian fauna. In : EISENBERG, E., R. GREENBERG & A. GOPHER (Eds). Tel Teʼo, A Neolithic, Chalcolithic, and Early Bronze Age site in the Hula Valley. Israel Antiquities Reports, Jerusalem, 13 : 171-194.
HORWITZ, L. K. (2003) - Temporal and spatial variation in Neolithic caprine exploitation strategies : A case study of fauna from the site of Yiftahʼel, (Israel). Paléorient, 21 : 19-58.
HORWITZ, L. K. & E. TCHERNOV (1998) - Diachronic and synchronic changes in patterns of animal exploitation during the Neolithic of the southern Levant. In : ANREITER, P., L. BARTOSIEWICZ, E. JEREM & W. MEID (Eds). Man and The Animal World. Archaeolingua, Budapest : 307-318.
HORWITZ, L.K., E. TCHERNOV, P. DUCOS, C. BECKER, A. VON DEN DRIESCH, L. MARTIN & A. GARRARD (1999) - Animal Domestication in the Southern Levant. Paléorient, 25 : 63-80.
HORWITZ, L.K., E. TCHERNOV & H. HONGO (2004) - The domestic status of the early Neolithic fauna of Cyprus : A view from the mainland. In : PELTENBURG, E. & A. WASSE (Eds). Neolithic Revolution. New Perspectives on Southwest Asia in Light of Recent Discoveries on Cyprus. Levant Supplementary Series 1, Oxbow Books, Oxford : 35-48.
HORWITZ, L. K., E. GALILI & O. LERNAU (in press) - Fauna from the Pottery Neolithic site of Newe Yam. Journal of the Israel Prehistoric Society.
HORWITZ, L. K., T. SIMMONS, O. LERNAU & E. TCHERNOV (in press) - Fauna from the sites of Gilgal I-III. In : BAR-YOSEF, O., A. GOPHER & N. GORING-MORRIS (Eds). The Epipaleolithic and Early Holocene Site Complex of Gilgal, Israel. Peabody Museum Publications, Brill.
KAFAFI, Z. (1988) - Jebel Abu Thawwab : a Pottery Neolithic village in north Jordan. In : GARRARD, A. & H-G. GEBEL (Eds). The Prehistory of Jordan. The State of Research in 1986. BAR International Series, Oxford, 396 : 451-471.
KISLEV, M.E., A. HARTMANN & E. GALILI (2004) - Archaeobotanical and archaeoentomological evidence from a well at Atlit Yam indicates colder, more humid cli-mate on the Israeli coast during the PPNC period. Journal of Archaeological Science, 31 : 1301-1310.
KUIJT, I. & A. GORING-MORRIS (2002) - Foraging, farming and social complexity in the Pre-Pottery Neolithic of the Southern Levant : A review and synthesis. Journal of World Prehistory, 16 : 361-440.
LEV-TOV, J. (2000) - Late prehistoric faunal remains from new excavations at Tel Ali (Northern Israel). In : MASHKOUR, M., A. M. CHOYKE, BUITENHUIS, H. & F. POPLIN (Eds). Archaeozoology of the Near East, IV. ARC Publicaties, Groningen, 32 : 208-217.
MARTIN, L. (1999) - Mammal remains from the eastern Jordanian Neolithic, and the nature of caprine herding in the steppe. Paléorient, 25(2) : 87-104.
MEADOW, R.H. (1989) - Osteological evidence for the process
of animal domestication. In : CLUTTON-BROCK, J. (Ed.). The Walking Larder. Unwin Hyman, London : 80-90.
MUNRO, N.D. (2004) - Small game indicators of human foraging efficiency and early herd management at the transition to agriculture in south-west Asia. In : BRUGAL, J-P. & J. DESSE (Eds). Petits animaux sociétés humaines. Du complément alimentaire aux ressources utilitaires. Editions APDCA, Antibes : 515-531.
NOY, T., A.J. LEGGE & E.S. HIGGS (1973) - Recent excavations at Nahal Oren, Israel. Proceedings of the Prehistoric Society, 39 : 75-99.
PETERS, J. & K. SCHMIDT (2004) - Animals in the symbolic world of Pre-Pottery Neolithic Göbekli Tepe, south-eastern Turkey : A Preliminary assessment. Anthropozoologica, 39 (1) : 179-218.
PETERS, J., D. HELMER, A. VON DEN DRIESCH & M. SANA SEGUI (1999) - Early animal husbandry in the northern Levant. Paléorient, 25 : 27-48
PETERSON, J. (2004) - Khirbet Hammam (WHS 149) : A late Pre-Pottery Neolithic B settlement in the Wadi el-Hasa, Jordan. Bulletin of the American School of Oriental Research, 334 : 1-17.
RICHARDSON, J.E. (1997) - An analysis of the faunal assemblages from two Pre-Pottery Neolithic sites in the Wadi Fidan, Jordan. In : GEBEL, H.G.K., Z. KAFAFI & G.O. ROLLEFSON (Eds). The Prehistory of Jordan II. Perspectives from 1997. Studies in Early Near Eastern Production, Subsistence and Environment, Berlin, 4 : 497-510.
SANA SEGUI, M. (1999) - Arqueología de la domesticacíon animal. La gestión de los recursos animales en Tell Halula (Valle del Éufrates, Siria) de 8.800 al 7.000 BP. Treballs dʼArqueologia del Pròxim Orient 1. Barcelone.
SANLAVILLE, P. (1996) - Changements climatiques dans la région Levantine à la fin du Pleistocène supérieur et au début de lʼHolocène. Leurs relation avec lʼévolution des sociétés humaines. Paléorient, 22 (1) : 7-30.
SAPIR, L. (2005) - The Faunal Remains from Motza : Paleoeconomy and Paleoecology of the Last Hunters. Unpublished MSc Thesis, Tel-Aviv University (in Hebrew).
SIMMONS, A.H., M. AL-NAHAR, G.O. ROLLEFSON, J. COOPER, Z. KAFAFI, I. KOHLER-ROLLEFSON, R. D. MADEL & K. R. DURAND (2001) - Wadi Shuʼeib, a large Neolithic community in Central Jordan : Final report of test investigations. Bulletin of the American School of Oriental Research, 321 : 1-39.
TCHERNOV, E. (1993) - From sedentism to domestication- A preliminary review of the soutern Levant. In : CLASON, A. T., S. PAYNE & H.-P. UERPMANN (Eds). Skeletons in Her Cupboard. Oxbow Monographs, Oxford, 34 : 189-234.
TCHERNOV, E. & L.K. HORWITZ (1990) - Herd management in the past and its impact on the landscape of the southern Levant. In : BOTTEMA, S., G. ENTJES-NIEBORG & W. VAN ZEIST (Eds). Man s̓ Role in the Shaping of the Eastern Mediterranean Landscape. Balkema Publishers, Rotterdam : 207-215.
TROY, C. S., D. E. MACHUGH, J. F. BALLEY, D. A. MAGEE, R. T. LOFTUS, P. CUNNINGHAM, A. T. CHAMBERLAIN, B. C. SYKES & D. G. BRADLEY (2001) - Genetic evidence for Near-Eastern origins of European cattle. Nature, 410 : 1086-1091.
UERPMANN, H.-P. (1987) - The Ancient Distribution of Ungulate Mammals in the Middle East. Beihefte Zum Tubinger Atlas Des Vorderen Orients. Reihe A, Nr. 27. L. Reichert, Wiesbaden.
VIGNE, J.-D., I. CARRERE & J. GUILAINE (2003) - Unstable status
Counting Cattle : Trends in Neolithic Bos Frequencies from the Southern Levant 223

of early domestic ungulates in the Near East : The example of Shillourokambos (Cyprus, IX-VIIIth millennia cal. B.C.). In : GUILAINE, J.& A. LE BRUN (Eds). Le Néolithique de Chypre. Actes du Colloque International, Nicosia : 239-251.
VIGNE, J.-D., D. HELMER & J. PETERS (2005) - New archaeozoological approaches to trace the first steps of animal domestication : general presentation, reflections and proposals. In : VIGNE, J.-D., D. HELMER & J. PETERS (Eds). The First Steps of Animal Domestication. Oxbow Books, Oxford : 1-16.
224 L. K. HORWITZ & P. DUCOS
Related Documents











