Use of light and inorganic carbon acquisition by two morphotypes of Zostera noltii Hornem Jesu ´s M. Mercado a, * , F.X. Niell b , Joa ˜o Silva c , Rui Santos c a Centro Oceanogra ´fico de Ma ´laga, Instituto Espan ˜ol de Oceanografı ´a, Puerto Pesquero s/n, Apdo. 285, 29640, Fuengirola, Ma ´laga, Spain b Departamento de Ecologı ´a, Universidad de Ma ´laga, Campus Universitario de Teatinos 29071, Ma ´laga, Spain c Centro de Cie ˆncias do Mar, CCMAR, Universidade do Algarve, Gambelas 8000, Faro, Portugal Received 1 January 2003; received in revised form 16 July 2003; accepted 23 July 2003 Abstract The affinity for dissolved inorganic carbon (DIC) and the mechanisms to use HCO 3 as a source of DIC for photosynthesis were investigated in two morphotypes of Zostera noltii Hornem. Both morphotypes were collected at Ria Formosa lagoon (Southern Portugal) at two different levels in the intertidal. Affinity for DIC at saturating photon fluence rate (PFR), estimated as photosynthetic conductance for CO 2 ( g p(CO 2 ) ), was reduced by 75% in the Z. noltii plants adapted to shade conditions (lower intertidal) in comparison to the sun morphotype (45 10 6 and 182 10 6 ms 1 , respectively), indicating that the plants acclimated to sun conditions (higher intertidal) had a higher capacity to use HCO 3 as DIC source for photosynthesis. Since external carbonic anhydrase activity was negligible and a large inhibitory effect was produced by Tris buffer addition, this HCO 3 use was attributed to the operation of H + ATPases creating low pH zones in periplasmic space. The photosynthetic CO 2 -flux supported for this mechanism was calculated to be 53 Amol O 2 m 2 s 1 in sun morphotype, about 80% out of maximum photosynthesis rate. In order to determine the possible photosynthetic energy cost of the HCO 3 use, the effect of decreasing light on photosynthetic rates and g p(CO 2 ) was estimated. Photosynthetic conductance decreased in both morphotypes at non- saturating PFR. This dependence of g p(CO 2 ) on PFR indicated the existence of a positive interactive effect between DIC and PFR which was more pronounced in the shade morphotype since the ascending slope of O 2 evolution vs. PFR curves at limiting PFRs was reduced from 7.2 to 2.3 mmol O 2 mol photon 1 at 4 and 0.5 mol m 3 of DIC, respectively. D 2003 Elsevier B.V. All rights reserved. Keywords: Carbonic anhydrase; Carbon acquisition; Eutrophication; Intertidal system; Photosynthesis; Seagrasses; Zostera 0022-0981/$ - see front matter D 2003 Elsevier B.V. All rights reserved. doi:10.1016/S0022-0981(03)00368-X * Corresponding author. Tel.: +34-952476955; fax: +34-952463808. E-mail address: [email protected] (J.M. Mercado). www.elsevier.com/locate/jembe Journal of Experimental Marine Biology and Ecology 297 (2003) 71 – 84

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.elsevier.com/locate/jembe
Journal of Experimental Marine Biology and Ecology
297 (2003) 71–84
Use of light and inorganic carbon acquisition by two
morphotypes of Zostera noltii Hornem
Jesus M. Mercadoa,*, F.X. Niellb, Joao Silvac, Rui Santosc
aCentro Oceanografico de Malaga, Instituto Espanol de Oceanografıa, Puerto Pesquero s/n, Apdo. 285,
29640, Fuengirola, Malaga, SpainbDepartamento de Ecologıa, Universidad de Malaga, Campus Universitario de Teatinos 29071, Malaga, Spain
cCentro de Ciencias do Mar, CCMAR, Universidade do Algarve, Gambelas 8000, Faro, Portugal
Received 1 January 2003; received in revised form 16 July 2003; accepted 23 July 2003
Abstract
The affinity for dissolved inorganic carbon (DIC) and the mechanisms to use HCO3� as a source of
DIC for photosynthesis were investigated in two morphotypes of Zostera noltii Hornem. Both
morphotypes were collected at Ria Formosa lagoon (Southern Portugal) at two different levels in the
intertidal. Affinity for DIC at saturating photon fluence rate (PFR), estimated as photosynthetic
conductance for CO2 ( gp(CO2)), was reduced by 75% in the Z. noltii plants adapted to shade conditions
(lower intertidal) in comparison to the sun morphotype (45� 10� 6 and 182� 10� 6 m s� 1,
respectively), indicating that the plants acclimated to sun conditions (higher intertidal) had a higher
capacity to use HCO3� as DIC source for photosynthesis. Since external carbonic anhydrase activity
was negligible and a large inhibitory effect was produced by Tris buffer addition, this HCO3� use
was attributed to the operation of H+ ATPases creating low pH zones in periplasmic space. The
photosynthetic CO2-flux supported for this mechanism was calculated to be 53 Amol O2 m� 2 s� 1 in
sun morphotype, about 80% out of maximum photosynthesis rate. In order to determine the possible
photosynthetic energy cost of the HCO3� use, the effect of decreasing light on photosynthetic rates
and gp(CO2)was estimated. Photosynthetic conductance decreased in both morphotypes at non-
saturating PFR. This dependence of gp(CO2)on PFR indicated the existence of a positive interactive
effect between DIC and PFR which was more pronounced in the shade morphotype since the
ascending slope of O2 evolution vs. PFR curves at limiting PFRs was reduced from 7.2 to 2.3 mmol
O2 mol photon� 1 at 4 and 0.5 mol m� 3 of DIC, respectively.
D 2003 Elsevier B.V. All rights reserved.
Keywords: Carbonic anhydrase; Carbon acquisition; Eutrophication; Intertidal system; Photosynthesis;
Seagrasses; Zostera
* Corresponding author. Tel.: +34-952476955; fax: +34-952463808.
0022-0981/$ - see front matter D 2003 Elsevier B.V. All rights reserved.
doi:10.1016/S0022-0981(03)00368-X
E-mail address: [email protected] (J.M. Mercado).

J.M. Mercado et al. / J. Exp. Mar. Biol. Ecol. 297 (2003) 71–8472
1. Introduction
Seagrasses are the dominant macrophytes of many near-shore soft-bottom ecosystems
where they constitute an important and highly productive component of the benthic
environment (McRoy and McMillan, 1977). A decline in seagrass-dominated areas has
been observed worldwide (Denninson et al., 1993). Seagrasses are being replaced by other
macrophytes, particularly green macroalgae (Hernandez et al., 1997; Clavero et al., 1999).
It has been proposed that this replacement is a consequence of the great sensitivity of
seagrass ecosystems to human-induced perturbations. The eutrophication of coastal areas
results in a higher growth of the phytoplankton. The subsequent light limitation of benthic
communities reduces their primary production (Bondsorff et al., 1997) and eventually
decreases seagrasses cover (Short and Neckles, 1999). Changes in riverine fluxes of
organic matter and nutrients or suspended matter due to human activities can produce
similar effects (Carreira et al., 1995).
According to Moore et al. (1996), the long-term survival of Zostera marina is affected
by water turbidity rather than changes in the nutrient concentration or salinity. This fact
suggests that the photosynthetic performance of seagrasses could contribute to their high
sensitivity to anthropogenic inputs. Previous reports demonstrate that the photosynthesis
rates are limited by the availability of DIC under natural conditions in many seagrasses
species (Madsen and Sand-Jensen, 1991; Durako, 1993; Beer and Koch, 1996; James and
Larkum, 1996; Invers et al., 1999). However, Hellblom et al. (2001) suggested recently
that this apparent low affinity for DIC have been largely an artefact of the use of buffers in
the incubation medium. Accordingly, Schwarz et al. (2000) demonstrated that the seawater
DIC concentration was enough to saturate in situ photosynthesis of two tropical seagrass
species of the genera Cymodocea and Halophila, which inhabit the upper part of the
intertidal system (i.e. fully exposed to solar radiation). By contrast, photosynthesis by
shade-adapted forms of these tropical species was limited at the seawater DIC concen-
tration (Schwarz et al., 2000). These results suggest that acclimation to decreasing light
intensity could affect the DIC acquisition mechanisms. The capacity to use HCO3� by
seagrasses has been demonstrated from kinetic measurements in species of the genera
Thalassia and Zostera (Sand-Jensen and Gordon, 1984; Durako, 1993; Beer and
Rehnberg, 1997; Touchette and Burkholder, 2000). At least three different mechanisms
of HCO3� use for photosynthesis have been described in seagrasses: (1) its transformation
into CO2 performed by an external CA (Millhouse and Stroher, 1986; Bjork et al., 1997),
(2) its direct uptake by means of specific transport systems located at plasma membrane
(Touchette and Burkholder, 2000), and (3) more recently, Hellblom et al. (2001) suggested
the use of HCO3� was dependent on low pH zones at periplasmic space created by H+
ATPases pumping protons to the outside of the plasma membrane. The ability of seagrass
species to modify the activity of these mechanisms in response to changes on light
intensity remains unexplored.
The goal of the experiments described in this paper was to determine the affinity to DIC
and the presence of different mechanisms for using HCO3� in two morphotypes of Z. noltii
which grow under different light conditions in the field. For this proposal, the effect on the
photosynthesis of carbonic anhydrase inhibitors and inhibitors for the proton pump
dependant, HCO3� use mechanism (i.e. Tris buffer) was determined (Axelsson et al.,

J.M. Mercado et al. / J. Exp. Mar. Biol. Ecol. 297 (2003) 71–84 73
2000). Additionally, photosynthesis vs. photon fluence rate (PFR) curves were performed
at different DIC concentrations in order to determine the dependence of affinity for DIC on
incident PFR.
2. Materials and methods
2.1. Sampling sites and plant material
Plants were collected in January 2000 from Ria Formosa lagoon. This is a shallow
mesotidal lagoon, extending for about 55 km along the coast of southern Portugal, with an
average depth of about 2 m. The tidal amplitude varies from an average of 2.80 m in spring
tides to 1.30 m in neap tides. The total area of the lagoon is 170 km2 with an exposed
intertidal area of 67 km2 during spring tides (Andrade, 1990).
Zostera noltii morphology in the Ria Formosa varies with vertical distribution of the
species in the intertidal. At the upper limit the leaves are thinner and shorter in contrast
with the lower limit where they are larger and longer. The two morphotypes are referred to
in this work as sun-type and shade-type, and respectively correspond to the small (SM)
and large (LM) morphotypes described by Peralta et al. (2001). Environmental conditions
change significantly along the vertical distribution of species, particularly light availability.
The lower limit of this bed is never completely emersed, while the upper limit has an
average emersion between 6 and 8 h every day, depending on tidal amplitude. As a
consequence, during January, the shade-type receives an average daily PAR dose of 4 mol
photons m� 2, whereas the sun-type receives around 15 mol photons m� 2. Other variables
as the pH also are different at each position. Thus, the pH values at the site where the sun
morphotype was collected increased from 7.4 at low tide up to 8.8 during high tide. In
contrast, at the shade-morphotype site, the pH varies from 7.9 to 8.3.
Plants were collected from Ria Formosa and kept cooled and wet in the dark while they
were transported to the laboratory. In the laboratory, plants were maintained in natural,
nutrient-poor seawater. About 10 g of plant were incubated into Plexiglas cylinder of 4 l in
capacity. They were aerated vigorously (about 3 l air min� 1) at 15 jC under white
fluorescent lamps (F20W/CW lamps), at a photoperiod of 12 h light per day and a photon
fluence rate (PFR) of 100 Amol photons m� 2 s� 1. The PFR was determined with a
quantum spherical PAR sensor (LiCor 193SB) connected to a radiometer (Li-1000).
The experiments were performed using pieces of leaves of 2 cm in length, cut
approximately 2 cm above the sheath. Prior the experiments, 10 mg of this plant material
were frozen at � 20 jC until the analysis of its photosynthetic pigment content. For this
purpose, chlorophyll a and b were extracted in N,N-dimethylformamide and their
concentrations were calculated according to Inskeep and Bloom (1985).
2.2. Oxygen evolution and effects of the buffer and CA inhibitors
Oxygen evolution was measured in small (8–9 ml) temperature-controlled (15–16 jC)seawater chambers containing oxygen electrodes. The suitable agitation of the medium in
the chambers was obtained by a magnetic stirrer. The 2 cm leaf fragments were transferred

J.M. Mercado et al. / J. Exp. Mar. Biol. Ecol. 297 (2003) 71–8474
to the oxygen-evolution chambers containing natural seawater. A PFR of 600 Amol
photons m� 2 s� 1 was provided by a Duro-Test lighting (model F13DTT/55K) lamp. The
effect of Tris buffer on photosynthesis was tested at pH 8.2 as described by Axelsson et al.
(2000), prior to testing the effect of the inhibitors of carbonic anhydrase (CA) on oxygen
evolution. Different Tris biological buffer (Sigma) concentrations ranging from 0 to 45
mol m� 3 were produced into the chambers by adding small amounts of a 2 kmol m� 3 Tris
solution. The pH electrode and meter (Crison 52-08, Alella, Spain) were calibrated in
buffer solutions referred to the NBS scale (Crison).
Since Tris produced a strong inhibitory effect on photosynthesis, unbuffered seawater
was used to test the effect of CA inhibitors, 6-ethoxyzolamide (EZ) and acetazolamide
(AZ; Sigma, Madrid, Spain). It was assumed that AZ cannot penetrate into the cell and
inhibits only the extracellular CA (Palmqvist et al., 1990; Haglund et al., 1992; Axelsson
et al., 1995) while EZ penetrates into the cell and inhibits the external and internal CA.
Carbon dioxide-free stock solutions of the inhibitors were prepared in 0.05 N NaOH to a
final concentration of 50 mmol m� 3. The assays of inhibitors were started by introducing
fragments of plant into the oxygen evolution chamber containing unbuffered seawater.
When a constant O2 evolution rate was obtained, AZ or EZ were added. A complete effect
was obtained within 2–3 min.
2.3. Oxygen evolution rate vs. PFR curves
The response of photosynthesis to increasing PFR was tested under six DIC concen-
trations (250, 500, 1000, 2000, 3000 and 4000 mmol m� 3). Pieces of plant were
introduced into the oxygen evolution chambers containing Ci free synthetic seawater
(450 mol m� 3, NaCl, 30 mol m� 3 MgSO4, 10 mol m� 3 KCl, 10 mol m� 3 CaCl2; Beer et
al., 1990). The pH was adjusted to 8.2 with Tris 2 mol m� 3 final concentration (it
produced an inhibition of the photosynthesis rates lower than 5%). This Tris concentration
was enough to maintain the pH within the range 8.2–8.3 during the experiments. After
zero net O2 exchange rate was attained, small amounts of a 200 mol m� 3 NaHCO3
solution were injected into the chambers in order to create the required DIC concentration.
Nine different PFRs (10, 15, 30, 70, 130, 210, 270, 360 and 600 Amol photons m� 2 s� 1)
were used to perform the P-PFR curves. They were obtained with glass neutral density
filters. A total of 600 Amol photons m� 2 s� 1 did not produce apparent photoinhibiton in
the two morphotypes. PFR values were measured inside the chamber with a spherical
sensor (20HM33CM12KG, Zemoko, Holland). Respiration was measured in darkness
before switching the light on. Oxygen evolution was recorded for 10–15 min at each PFR.
Maximal photosynthetic rates (Pmax) and ascending slope at limiting PFRs (a) were
obtained after the fitting of the curves with the rectangular hyperbole equation (Henley,
1993):
P ¼ Pmax½aPFR=ðPmax þ aPFRÞ� þ Rd
where P is the photosynthetic rate and Rd is the respiration rate in darkness. The light
saturation parameter (Ek) was estimated as Pmax/a. Three independent curves (i.e. startingfrom different samples) were constructed for each DIC concentration tested. The curves

J.M. Mercado et al. / J. Exp. Mar. Biol. Ecol. 297 (2003) 71–84 75
were analysed separately. Their goodness of fit to the model was tested by using least-
squares regression analysis. The average R2 value obtained from the least-squares
regression analysis was 0.99.
Oxygen evolution was expressed on a surface area basis. Fresh weight per unit area was
estimated by determining the surface area of samples that had previously been weighed.
Leaf surface area was determined after digitalisation using a computer video system as
described by Luning (1992).
2.4. Conductance for inorganic carbon ( gp)
The response of photosynthesis to increasing DIC concentration was determined at
different PFRs using the data of the P–PFRs curves. Photosynthesis vs. DIC concentration
curves were constructed at six selected PFRs (30, 70, 130, 270, 360 and 600 Amol photons
m� 2 s� 1). Conductance for DIC ( gp(DIC)) at each PFR was calculated as the initial slope
of these curves. Conductance for CO2 ( gp(CO2)) was obtained by multiplying gp(DIC) by the
CO2/DIC ratio estimated by using the apparent dissociation constants of carbonic acid in
seawater (Riley and Chester, 1977).
2.5. Assay of external CA activity
External CA activity was assayed using the potentiometric method. The time
required for a drop of 0.4 pH units within the pH range 8.4–7.4 was measured at
0–2 jC. About 50–200 mg FW were washed three times in a buffer (100 mol m� 3
Tris, 5 mol m� 3 EDTA-Na and 25 mol m� 3 ascorbic acid) before being placed in the
reaction vessel (Haglund et al., 1992). The reaction was started by rapidly introducing 1
ml of ice-cold CO2-saturated distilled water. One unit of enzyme activity (REA) was
defined as (to/tc)� 1 where to and tc are the times for pH change of the control (assay
buffer without sample) and sample, respectively. The activity was recalculated on a
fresh weight basis.
The external activity [termed CA (%)] was also determined by using the method
developed by Mercado et al. (1997). This method permits the relative importance of the
external CA for DIC acquisition to be assessed relative to direct uptake of HCO3�. 30 mg
of plant material was transferred to the O2 evolution chambers containing synthetic
seawater buffered at pH 8.7 with Tris 2 mol m� 3 final concentration. After zero net O2
exchange rate was attained, 200 Al of ice-cold CO2-saturated distilled water were added in
both the presence and absence of 50 mmol m� 3 AZ. CO2 and HCO3� dependent
photosynthetic rates were then calculated. Activity estimated according to Eq. (2) in
Mercado et al. (1997) was expressed in arbitrary units (ranging from 0 to 1, where a figure
of 1 represents the maximal detectable activity with this method).
2.6. Statistics
The results were expressed as the mean valuesF standard deviation (S.D.). Significant
differences among means were tested at p < 0.05 with a one-way ANOVA, followed by the
Fisher’s protected multirange test (Sokal and Rohlf, 1981).

J.M. Mercado et al. / J. Exp. Mar. Biol. Ecol. 297 (2003) 71–8476
3. Results
Differences between the two morphotypes were pronounced, especially for leaf width
which was higher in sun morphotype than in shade morphotype (see Peralta et al., 2001 for
a complete morphological description). Analysis of the morphometric characteristics of the
leaves utilized in our experiments (Table 1) revealed statistically significant differences in
fresh weight per unit area. The chlorophyll a and b contents expressed on a fresh weight
basis were similar in both morphotype although they expressed on an area surface basis
were higher in the shade morphotype. There were no significant differences in the Chl
b/Chl a ratio.
Photosynthetic oxygen production was strongly inhibited by the addition of Tris
without changing pH. The inhibition percentage was dependent on the buffer concentra-
tion (Fig. 1), reaching 80% inhibition at 45 mol m� 3 of Tris. Inhibition was lower than 5%
at 2 mol m� 3 of Tris, the concentration used in performing the P-PFR curves. This
concentration was shown to be adequate to maintain constant pH during the experiments
as tested by measuring pH after the incubations.
There was no agreement between the results of external CA activity obtained with the
two methods used. Thus, significant activity was detected by the potentiometric method
(Table 2), although it was very low in comparison with values reported for other
macrophytes (Mercado et al., 1998). In contrast, CA activity assayed using the method
described by Mercado et al. (1997) was not detected in three out of four samples, since the
criterion for determination of activity (i.e. a value of a/cV higher than 2 according to Eq.
(1) of Mercado et al., 1997) was not satisfied. It was due to the low inhibition of
photosynthesis obtained after acetazolamide (AZ) addition at pH 8.7 (the value used for
the assay). Since CA activity was detected potentiometrically, this low inhibition must
indicate that CO2 requirement of RUBISCO is fulfilled via mechanisms of HCO3� use
other that the external CA activity. Accordingly, the inhibitory effect of AZ at the normal
seawater pH was much less that it found after the addition of a high buffer concentration
(Fig. 2). In contrast to AZ, ethoxyzolamide inhibited photosynthesis by 80%. The
percentage inhibition of photosynthesis by AZ and EZ and the external CA activity were
similar in the two morphotypes.
Changes in the inorganic carbon concentration did not affect the respiration rates, which
were similar in the two morphotypes (9.73F 5.01 and 13.04F 4.86 Amol O2 m� 2 min� 1
for shade- and sun-morphotype, respectively). About 500 Amol photons m� 2 s� 1 were
Table 1
Fresh weight per area ratio (FW/area) and chlorophyll a and b contents in sun and shade morphotypes
Sun morphotype Shade morphotype
FW/area (g m� 2) 32.26F 9.12a (n= 10) 54.61F 20.83b (n= 11)
Chl a (mg m� 2) 63.55F 3.96a (n= 4) 85.74F 9.12b (n= 4)
Chl b (mg m� 2) 19.03F 1.07a (n= 4) 26.76F 1.94b (n= 4)
Chl b/a 0.31F 0.01a (n= 4) 0.30F 0.00a (n= 4)
Chl a/b ratio is also provided. The numbers are the meanF S.D. n indicates the numbers of independent samples
analysed.
Within each row, means with different superscripts are significantly different at p= 0.05.
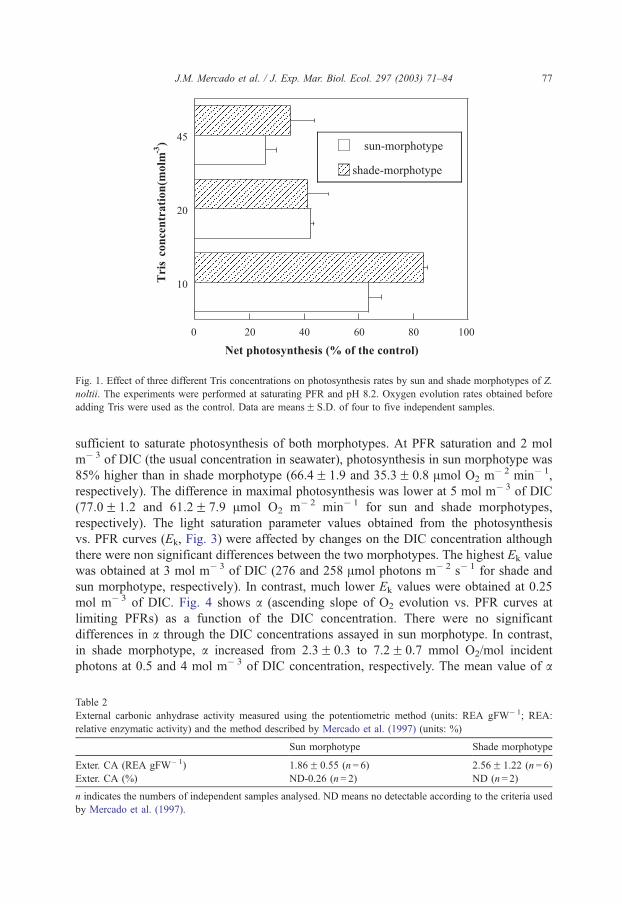
Fig. 1. Effect of three different Tris concentrations on photosynthesis rates by sun and shade morphotypes of Z.
noltii. The experiments were performed at saturating PFR and pH 8.2. Oxygen evolution rates obtained before
adding Tris were used as the control. Data are meansF S.D. of four to five independent samples.
J.M. Mercado et al. / J. Exp. Mar. Biol. Ecol. 297 (2003) 71–84 77
sufficient to saturate photosynthesis of both morphotypes. At PFR saturation and 2 mol
m� 3 of DIC (the usual concentration in seawater), photosynthesis in sun morphotype was
85% higher than in shade morphotype (66.4F 1.9 and 35.3F 0.8 Amol O2 m� 2 min� 1,
respectively). The difference in maximal photosynthesis was lower at 5 mol m� 3 of DIC
(77.0F 1.2 and 61.2F 7.9 Amol O2 m� 2 min� 1 for sun and shade morphotypes,
respectively). The light saturation parameter values obtained from the photosynthesis
vs. PFR curves (Ek, Fig. 3) were affected by changes on the DIC concentration although
there were non significant differences between the two morphotypes. The highest Ek value
was obtained at 3 mol m� 3 of DIC (276 and 258 Amol photons m� 2 s� 1 for shade and
sun morphotype, respectively). In contrast, much lower Ek values were obtained at 0.25
mol m� 3 of DIC. Fig. 4 shows a (ascending slope of O2 evolution vs. PFR curves at
limiting PFRs) as a function of the DIC concentration. There were no significant
differences in a through the DIC concentrations assayed in sun morphotype. In contrast,
in shade morphotype, a increased from 2.3F 0.3 to 7.2F 0.7 mmol O2/mol incident
photons at 0.5 and 4 mol m� 3 of DIC concentration, respectively. The mean value of a
Table 2
External carbonic anhydrase activity measured using the potentiometric method (units: REA gFW� 1; REA:
relative enzymatic activity) and the method described by Mercado et al. (1997) (units: %)
Sun morphotype Shade morphotype
Exter. CA (REA gFW� 1) 1.86F 0.55 (n= 6) 2.56F 1.22 (n= 6)
Exter. CA (%) ND-0.26 (n= 2) ND (n= 2)
n indicates the numbers of independent samples analysed. ND means no detectable according to the criteria used
by Mercado et al. (1997).
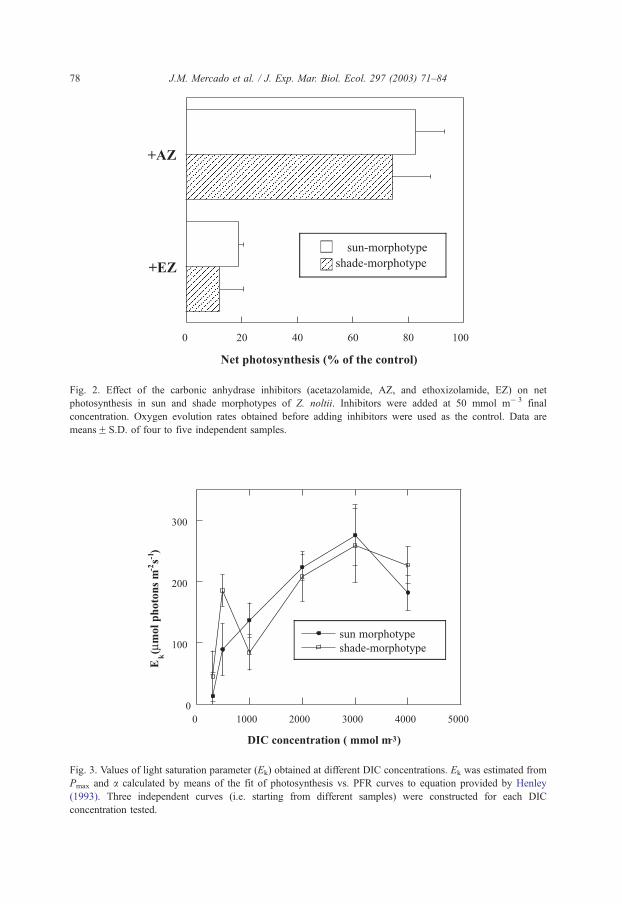
Fig. 3. Values of light saturation parameter (Ek) obtained at different DIC concentrations. Ek was estimated from
Pmax and a calculated by means of the fit of photosynthesis vs. PFR curves to equation provided by Henley
(1993). Three independent curves (i.e. starting from different samples) were constructed for each DIC
concentration tested.
Fig. 2. Effect of the carbonic anhydrase inhibitors (acetazolamide, AZ, and ethoxizolamide, EZ) on net
photosynthesis in sun and shade morphotypes of Z. noltii. Inhibitors were added at 50 mmol m� 3 final
concentration. Oxygen evolution rates obtained before adding inhibitors were used as the control. Data are
meansF S.D. of four to five independent samples.
J.M. Mercado et al. / J. Exp. Mar. Biol. Ecol. 297 (2003) 71–8478
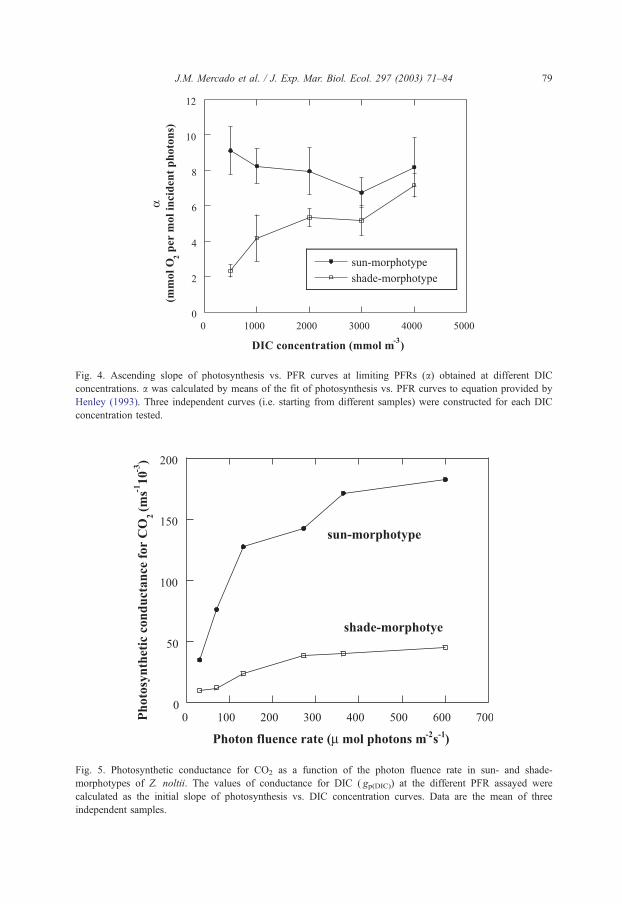
Fig. 4. Ascending slope of photosynthesis vs. PFR curves at limiting PFRs (a) obtained at different DIC
concentrations. a was calculated by means of the fit of photosynthesis vs. PFR curves to equation provided by
Henley (1993). Three independent curves (i.e. starting from different samples) were constructed for each DIC
concentration tested.
Fig. 5. Photosynthetic conductance for CO2 as a function of the photon fluence rate in sun- and shade-
morphotypes of Z. noltii. The values of conductance for DIC ( gp(DIC)) at the different PFR assayed were
calculated as the initial slope of photosynthesis vs. DIC concentration curves. Data are the mean of three
independent samples.
J.M. Mercado et al. / J. Exp. Mar. Biol. Ecol. 297 (2003) 71–84 79

J.M. Mercado et al. / J. Exp. Mar. Biol. Ecol. 297 (2003) 71–8480
was higher in sun morphotype (8.2F 0.7 mmol O2/mol photon) than in shade morphotype
(4.8F 1.8 mmol O2/mol photons).
The values of gp(CO2)showed a dependence on PFR (Fig. 5) with an asymptotic
relationship. The mean value of gp(CO2)increased by fivefold when PFR increased from 30
to 600 Amol photons m� 2 s� 1 for the two morphotypes. Higher gp(CO2)values in the sun
morphotype than in the shade morphotype were reached at each PFR used. In particular,
gp(CO2)at saturating PFR (600 Amol photons m� 2 s� 1) was fourfold higher in the sun
morphotype (182.3� 10� 6 m s� 1) than in the shade morphotype (45.1�10� 6 m s� 1).
4. Discussion
4.1. Inorganic carbon acquisition
The initial slope of photosynthesis rate vs. DIC curves (photosynthetic conductance for
CO2, gp(CO2)) at saturating irradiance has been proposed as an useful parameter for
determining the ability to extract Ci from seawater for photosynthesis (Johnston et al.,
1992). Thus, the different gp(CO2)values found for the two Zostera morphotypes at
saturating irradiance are indicative of a differential ability to use DIC because they were
fourfold lower in the shade morphotype when compared to the sun morphotype. The
possibility that gp(CO2)values are consistent with photosynthesis relying only on CO2
diffusive entry can be checked by calculating the minimum boundary layer thickness
which would permit these values (Johnston et al., 1992; Mercado et al., 1998). For this
proposal, it is assumed that no external transformation of HCO3� into CO2 is occurring.
The thickness of the diffusion boundary layer is calculated by multiplying CO2 diffusion
coefficient (1.3� 10� 9 m� 2 s� 1) to gp(CO2)(Johnston et al., 1992; Raven et al., 2002).
The calculated boundary layer thickness is 7 Am for sun morphotype and 29 Am for shade
morphotype. Different authors indicate that overall conductance for CO2 (including the
intracellular pathway) account for a diffusion boundary layer of 10–15 Am in macroalgae
without mechanisms for using HCO3� (Maberly et al., 1992; Raven et al., 2002). The value
obtained for shade morphotype is higher than it suggested by Maberly et al. (1992). Taking
into account the stirring conditions in the chamber where photosynthesis was measured, it
can be concluded that 29 Am is concordant with a use of DIC mainly based on CO2
diffusive entry. By contrast, the value of boundary layer thickness necessary to account for
measured gp(CO2)in sun morphotype is lower than the value calculated by Maberly et al.
(1992), therefore it indicates that use of HCO3� for photosynthesis is occurring in this
morphotype.
The reason for this difference in the ability to use HCO3� between the two morphotypes
could be the different activation state of the mechanisms involved in the HCO3� use.
Carbonic anhydrase-mediated indirect use of bicarbonate has been described for different
seagrasses species (James and Larkum, 1996; Invers et al., 1999), including two species of
Zostera, Z. muelleri (Millhouse and Stroher, 1986) and Z. marina (Beer and Rehnberg,
1997). In contrast, we have found that the role of the external CA on HCO3� indirect use in
Z. noltii must be very restricted since photosynthesis was only 20% inhibited with AZ and
no activity was detected by the method developed by Mercado et al. (1997). The findings

J.M. Mercado et al. / J. Exp. Mar. Biol. Ecol. 297 (2003) 71–84 81
suggest that more efficient use of HCO3�, not dependent on external CA must be
occurring, at least in the sun morphotype. An alternative mechanism to explain the use
of HCO3� independently of external CA activity, is the one recently described by Hellblom
et al. (2001) in Z. marina. According to these authors, HCO3� use in this species is
facilitated by proton transport towards the outside of the cell membrane. These protons are
then used to facilitate the inward transport of inorganic carbon through the cell membrane
(Axelsson et al., 2000). Hellblom et al. (2001) based their model on the high sensitivity of
photosynthesis to proton buffers. We demonstrate that photosynthesis was inhibited almost
80% by a high Tris concentration (i.e. 45 mol m� 3) suggesting that such a mechanism is
the main component of HCO3� use in Z. noltii sun morphotype. According to the values of
photosynthesis rate obtained in the presence of Tris, the DIC flux supported by this
mechanism could count for 53 Amol O2 m� 2 min� 1 in sun morphotype. If this DIC-flux is
compared to the one theoretically supported by the spontaneous dehydration of CO2
(which can be calculated from the photosynthetic rates obtained in the presence of AZ plus
Tris), it can be calculated that the HCO3� use mechanism sensitive to Tris produces a CO2
concentration increase by 5 fold in the periplasmic space. It has been noted that the shade
morphotype, with a comparatively lower capacity for use HCO3�, also exhibits a high
sensitivity to buffer. Therefore, restricted capacity for use of HCO3� cannot be ruled out.
4.2. Use of light by shade and sun morphotypes
The reason for a higher Pmax at saturating light and DIC in the sun morphotype could be
its higher capacity to use HCO3� and/or a higher Rubisco content, as both lead to higher
carbon dioxide fixation rates (Sukenik et al., 1987). The lower a found in the shade
morphotype could also be related to DIC acquisition, since it was increased by twofold in
this morphotype when DIC concentration was increased from 0.5 to 4 mol m� 3. It is worth
noting that no effect was found in sun morphotype. In our experiments, the CO2/O2 ratio
in the medium was modified by changing the DIC concentration (Raven, 1984, 1997;
Kerby and Raven, 1985; Beardall, 1989; Mercado and Niell, 1999). Since the shade
morphotype exhibited a very low capacity to use HCO3� it can be speculated that
photorespiration could contribute to the lower a values observed at low DIC concentration
in comparison to the sun morphotype. Our results contrast to the ones obtained by Kubler
and Raven (1995) who showed that light-harvesting efficiency was inversely related to the
effectiveness of DIC acquisition in Palmaria palmata. According to those authors, the
interactive effect is a consequence of the trade-off between photosynthetic efficiency and
bicarbonate use based on the fact that the use of HCO3� requires energy. In our
experiments, the reliance on CO2 found in the shade morphotype would indicate that
the photon cost of photorespiration is higher than the possible cost of HCO3� use (Raven,
1984).
The dependence of gp(CO2)on PFR has been also described by other authors (Maberly,
1990; Johnston et al., 1992; Mercado et al., 2000, 2001) and demonstrates that a positive
interactive effect exists between light use and DIC acquisition. In both morphotypes, an
increase of DIC concentration would be necessary to support an equivalent photosynthetic
rate when incident light is reduced. Apparently, the photosynthetic rate of the sun
morphotype depends more on light quantity than on DIC concentration since it exhibited

J.M. Mercado et al. / J. Exp. Mar. Biol. Ecol. 297 (2003) 71–8482
a relative high capacity for HCO3� use. In contrast, both factors (availability of light and
DIC) limit the photosynthetic rates of the shade morphotype, as photosynthetic efficiency
was dependent on DIC concentration. In any case, the reduction of incident light leads to a
lower affinity to DIC in both morphotype, which may increase the susceptibility of Z.
noltii to eutrophication.
Acknowledgements
This work was financed by the Projects REN2002-00340/MAR and REN2002-04022-
C02-01 (CICYT) and ECOFITO (Instituto Espanol de Oceanografıa). J.M. Mercado is
supported from Ramon y Cajal Programme of the Ministerio de Ciencia y Tecnologıa from
Spain. J. Silva is supported by PRAXIS XXI/BD/15782/98 grant. [RW]
References
Andrade, C.F., 1990. O ambiente de barreira da Ria Formosa Algarve-Portugal. PhD thesis, Universidade de
Lisboa, Portugal.
Axelsson, L., Ryberg, H., Beer, S., 1995. Two modes of bicarbonate utilization in the marine green macroalga
Ulva lactuca. Plant Cell Environ. 18, 439–445.
Axelsson, L., Mercado, J.M., Figueroa, F.L., 2000. Utilization of HCO3� at high pH by the brown macroalga
Laminaria saccharina. Eur. J. Phycol. 35, 53–59.
Beardall, J., 1989. Photosynthesis and photorespiration in marine phytoplankton. Aquat. Bot. 34, 105–130.
Beer, S., Koch, E., 1996. Photosynthesis of marine macroalgae and seagrasses in globally changing CO2 environ-
ments. Mar. Ecol. Prog. Ser. 141, 199–204.
Beer, S., Rehnberg, J., 1997. The acquisition of inorganic carbon by the seagrass Zostera marina. Aquat. Bot. 56,
277–283.
Beer, S., Israel, A., Drechsler, G.D., 1990. Photosynthesis in Ulva fasciata: V. Evidence for an inorganic carbon
concentrating system, and Rubisco CO2 kinetics. Plant Physiol. 94, 1542–1546.
Bjork, M., Weil, A., Semesi, S., Beer, S., 1997. The photosynthetic responses of eight seagrass species from
Zanzibar to increased inorganic carbon concentrations. Mar. Biol. 129, 363–366.
Bondsorff, E., Blompvist, E.M., Mattila, J., Norkko, A., 1997. Coastal eutrophication: causes, consequences
and perspectives in the Archipelago areas of the Northern Baltic Sea. Estuar. Coast. Shelf Sci. 44 (suppl. A),
63–72.
Carreira, J.A., Clavero, V., Galvez, J.A., Guevara, J.M., Hernandez, I., Lucena, J., Niell, F.X., Peralta, G., Perez-
Llorens, J.L., Vergara, J.J., 1995. Eutrophication progresses in the estuaries site at the end of small catchments
areas. In: Kamermans, P., Nienhuis, P.H. (Eds.), Eutrophication and Macrophytes. Netherland Institute of
Ecology, The Netherlands, pp. 113–116.
Clavero, V., Izquierdo, J.J., Palomo, L., Fernandez, J.A., Niell, F.X., 1999. Water management and climate
changes increase the phosphorous accumulation in the small shallow estuary of Palmones River (southern
Spain). Sci. Total Environ. 228, 193–202.
Denninson, W.C., Orth, R.J., Moore, K.A., Stevenson, J.C., Carter, V., Kollar, S., Bergstrom, P.W., Batiuk, R.A.,
1993. Assessing water quality with submerged aquatic vegetation. Bioscience 43, 86–94.
Durako, M.J., 1993. Photosynthetic utilization of CO2 and HCO3� in Thalassia testudintum (Hydrocharitaceae).
Mar. Biol. 115, 373–380.
Haglund, K., Bjork, M., Ramazanov, Z., Garcıa-Reina, G., Pedersen, M., 1992. Role of carbonic anhydrase in
photosynthesis and inorganic-carbon assimilation in the red alga Gracilaria tenuistipitata. Planta 187,
275–281.
Hellblom, F., Beer, S., Bjork, M., Axelsson, L., 2001. A buffer sensitive inorganic carbon utilisation system in
Zostera marina. Aquat. Bot. 69, 55–62.

J.M. Mercado et al. / J. Exp. Mar. Biol. Ecol. 297 (2003) 71–84 83
Henley, W.J., 1993. Measurement and interpretation of photosynthetic light-response curves in algae in the
context of photoinhibition and diel changes. J. Phycol. 29, 729–739.
Hernandez, I., Peralta, G., Perez-Llorens, J.A., Vergara, J.J., Niell, F.X., 1997. Biomass and dynamics of growth
of Ulva species in Palmones River Estuary. J. Appl. Phycol. 33, 764–772.
Inskeep, W., Bloom, P.R., 1985. Extinction coefficients of Chlorophyll a and b in N,N-dimethylformamide and
80% acetone. Plant Physiol. 77, 483–485.
Invers, O., Perez, M., Romero, J., 1999. Bicarbonate utilization in seagrass photosynthesis: role of carbonic
anhydrase in Posidonia oceanica (L.) Delile and Cymodocea nodosa (Ucria) Ascherson. J. Exp. Mar. Biol.
Ecol. 235, 125–133.
James, P.L., Larkum, A.W.D., 1996. Photosynthetic inorganic carbon acquisition of Posidonia australis. Aquat.
Bot. 55, 149–157.
Johnston, A.M., Maberly, S.C., Raven, J.A., 1992. The acquisition of inorganic carbon by four red macroalgae.
Oecologia 92, 317–326.
Kerby, N.W., Raven, J.A., 1985. Transport and fixation of inorganic carbon by marine algae. Adv. Bot. Res. 11,
71–123.
Kubler, J.E., Raven, J.A., 1995. The interaction between inorganic carbon acquisition and light supply in
Palmaria palmata (Rhodophyta). J. Phycol. 31, 369–375.
Luning, K., 1992. Day and night kinetics of growth rate in green, brown and red seaweed. J. Phycol. 28,
794–803.
Maberly, S.C., 1990. Exogenous sources of inorganic carbon for photosynthesis by marine macroalgae. J. Phycol.
26, 439–449.
Maberly, S.C., Raven, J.A., Johnston, A.M., 1992. Discrimination between 12C and 13C by marine plants.
Oecologia 91, 481–492.
Madsen, T.V., Sand-Jensen, K., 1991. Photosynthetic carbon assimilation in aquatic macrophytes. Aquat. Bot. 41,
5–40.
McRoy, C.P., McMillan, C., 1977. Production ecology and physiology of seagrasses. In: McRoy, C.P., Helferrich,
C. (Eds.), Production Ecology and Physiology of Seagrasses. Dekler, New York, USA, pp. 58–87.
Mercado, J.M., Niell, F.X., 1999. Carbonic anhydrase activity and use of HCO3� in Bostrychia scorpioides
(Ceramiales, Rhodophyceae). Eur. J. Phycol. 34, 13–19.
Mercado, J.M., Figueroa, F.L., Niell, F.X., Axelsson, L., 1997. A new method for estimating external carbonic
anhydrase in macroalgae. J. Phycol. 33, 999–1006.
Mercado, J.M., Gordillo, F.J.L., Figueroa, F.L., Niell, F.X., 1998. External carbonic anhydrase and affinity for
inorganic carbon in intertidal macroalgae. J. Exp. Mar. Biol. Ecol. 221, 209–220.
Mercado, J.M., Carmona, R., Niell, F.X., 2000. Affinity for inorganic carbon of Gracilaria tenuistipitata cultured
at low and high irradiance. Planta 210, 758–764.
Mercado, J.M., Niell, F.X., Gil-Rodrıguez, M.C., 2001. Photosynthesis might be limited by light, not inorganic
carbon availability, in three intertidal Gelidiales species. New Phytol. 149, 431–439.
Millhouse, J.V., Stroher, S., 1986. Salt-stimulated bicarbonate-dependent photosynthesis in the marine angio-
sperm Zostera muelleri. J. Exp. Bot. 37, 965–976.
Moore, K.A., Neckles, H.A., Orth, R.J., 1996. Zostera marina (eelgrass) growth and survival along a gradient of
nutrients and turbidity in the lower Chesapeake Bay. Mar. Ecol. Prog. Ser. 142, 247–259.
Palmqvist, K., Ramazanov, Z., Samuelsson, G., 1990. Role of extracellular carbonic anhydrase for accumulation
of inorganic carbon in the green alga Chlamydomonas reinhardtii. A comparison between wild-type and cell-
wall-less mutant cells. Physiol. Plant. 80, 267–276.
Peralta, G., Perez-Llorens, J.L., Hernandez, I., Brun, F., Vergara, J.J., Bartual, A., Galvez, J.A., Garcıa, C.M.,
2001. Morphological and physiological differences between two morphotypes of Zostera noltii Hornem.
From the south-western Iberian Peninsula. Helg. Mar. Res. 54, 80–86.
Raven, J.A., 1984. Energetic and Transport in Aquatic Plants. AR, Liss, New York.
Raven, J.A., 1997. Putting the C in phycology. Eur. J. Phycol. 32, 319–333.
Raven, J.A., Johnston, A.M., Kubler, J.E., Korb, R., McInroy, S.G., Handley, L.L., Scrimgeour, C.M., Walker,
D.I., Beardall, J., Vanderklift, M., Fredriksen, S., Dunton, K.H., 2002. Mechanistic interpretation of carbon
isotope discrimination by marine macroalgae and seagrasses. Funct. Plant Biol. 29, 355–378.
Riley, J.B., Chester, R., 1977. Introduction to Marine Chemistry. Academic Press, London.

J.M. Mercado et al. / J. Exp. Mar. Biol. Ecol. 297 (2003) 71–8484
Sand-Jensen, K., Gordon, D.M., 1984. Differential ability of marine and fresh water macrophytes to utilize
bicarbonate and carbon dioxide. Mar. Biol. 80, 247–254.
Schwarz, A.-M., Bjork, M., Buluda, T., Mtolera, M., Beer, S., 2000. Photosynthetic utilisation of carbon and light
by two tropical seagrass species as measured in situ. Mar. Biol. 137, 755–761.
Short, F.T., Neckles, H.A., 1999. The effects of global climate change on seagrasses. Aquat. Bot. 63, 169–196.
Sokal, P.R., Rohlf, F.J., 1981. Biometry, 2nd ed. WH Freeman, San Francisco.
Sukenik, A., Bennett, J., Falkowski, P., 1987. Light-saturated photosynthesis-limitation by electron transport or
carbon fixation? Biochem. Biophys. Acta 891, 205–215.
Touchette, B.W., Burkholder, J.M., 2000. Overview of the physiological ecology of carbon metabolism in
seagrasses. J. Exp. Mar. Biol. Ecol. 250, 169–205.
Related Documents











