Urban forests as hubs for novel zoonosis: blood meal analysis, seasonal variation in Culicoides (Diptera: Ceratopogonidae) vectors, and avian haemosporidians DIEGO SANTIAGO-ALARCON 1,2 *, PETER HAVELKA 3 , EDUARDO PINEDA 2 , GERNOT SEGELBACHER 4 and H. MARTIN SCHAEFER 1 1 Department of Ecology and Evolutionary Biology, University of Freiburg, Hauptstr. 1, 79104 Freiburg, Baden-Württemberg, Germany 2 Biología y Conservación de Vertebrados, Instituto de Ecología A.C. Carretera Antigua a Coatepec 351, El Haya, C.P. 91070 Xalapa, Veracruz, México 3 Department of Zoology, Staatliches Museum für Naturkunde, Erbprinzenstraße 13, 76133 Karlsruhe, Baden-Württemberg, Germany 4 Department of Wildlife Ecology and Management, University of Freiburg, Tennenbacher Straße 4, 79106 Freiburg, Baden-Württemberg, Germany (Received 3 May 2013; revised 6 June 2013; accepted 9 June 2013) SUMMARY Culicoides vectors can transmit a diverse array of parasites and are globally distributed. We studied feeding preferences and seasonal variation of Culicoides (Diptera: Ceratopogonidae) vectors in an urban forest of Germany to determine whether humans living nearby are readily exposed to vector-borne parasites from wild animals. We used a fragment of the mtDNA COI gene to identify hosts from blood meals. We amplified a fragment of the mtDNA cyt b to detect haemosporidian infections in Culicoides abdomens and thoraxes. We detected a total of 22 Culicoides species. Fifty-eight blood meals (84%) were from humans, 10 from birds, and one from livestock. We found Culicoides kibunensis (considered ornithophilic) with 29 human blood meals. Host generalist Culicoides festivipennis and Culicoides obsoletus had 14 human blood meals. Culicoides clastrieri and Culicoides semimaculatus fed on birds; previously humans were their only known host. Six thoraxes and three abdomens were infected with either Haemoproteus pallidulus or Haemoproteus parabelopolskyi. There were changes in Culicoides community structure across months. Culicoides pictipennis was the dominant species during spring, C. kibunensis and C. clastrieri were dominant during summer, and C. obsoletus was dominant by early autumn. All dominant species were generalists feeding on birds, livestock and humans. Our results indicate that humans can serve as a blood source for dominant Culicoides species instead of the normal wild animal hosts in urban areas. Key words: Culicoides, Haemoproteus, Sylvia atricapilla, blood meal analysis, emergent disease, conservation medicine, host–parasite interaction. INTRODUCTION Haemosporidian parasites are responsible for millions of infections and thousands of deaths in humans, as well as domestic and wild animals each year (Valkiūnas, 2005; Enayati and Hemingway, 2010). Infections may spread further owing to current environmental changes because climate warming enhances the geographical expansion of both parasites and vectors into regions from which they were previously absent (Jones et al. 2008). Despite their relevance for human and animal health, insect vectors feeding on different vertebrate species and transmitting haemosporidians are poorly known (Santiago-Alarcon et al. 2012a). This creates two challenges. First, as the insect vectors of most of the haemosporidian parasites described to date are unknown, we do not know how and with whom these vectors directly and indirectly interact. Second, if vectors suck blood from humans and other vertebrates (e.g. Mayne, 1928; Coggeshall, 1940; Lefèvre et al. 2009), humans will be exposed to a wide range of parasites existing in the animal community surrounding them (Santiago-Alarcon et al. 2012b). Hence, it is important to know what blood parasites make it into the bloodstream of humans, even if they initially do not develop into an infection, because this is the first step towards new emerging diseases (Arinaminpathy and McLean, 2009). Haemosporidians are mainly transmitted by five families of Diptera: Culicidae, Hippoboscidae, Simu- liidae, Ceratopogonidae and Psychodidae (Valkiūnas, 2005; Santiago-Alarcon et al. 2012a). Until recently it was believed that most vector species had special- ized feeding habits, in general feeding on different vertebrate classes, but recent molecular data have found some vector species with generalized feeding habits (e.g. Santiago-Alarcon et al. 2012b). Studies * Corresponding author: Biología y Conservación de Vertebrados, Instituto de Ecología A.C. Carretera Antigua a Coatepec 351, El Haya, C.P. 91070 Xalapa, Veracruz, México. E-mail: [email protected] 1 Parasitology, Page 1 of 12. © Cambridge University Press 2013 doi:10.1017/S0031182013001285

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Urban forests as hubs for novel zoonosis: blood meal analysis,seasonal variation in Culicoides (Diptera: Ceratopogonidae)vectors, and avian haemosporidians
DIEGO SANTIAGO-ALARCON1,2*, PETER HAVELKA3, EDUARDO PINEDA2,GERNOT SEGELBACHER4 and H. MARTIN SCHAEFER1
1Department of Ecology and Evolutionary Biology, University of Freiburg, Hauptstr. 1, 79104 Freiburg,Baden-Württemberg, Germany2Biología y Conservación de Vertebrados, Instituto de Ecología A.C. Carretera Antigua a Coatepec 351, El Haya,C.P. 91070 Xalapa, Veracruz, México3Department of Zoology, Staatliches Museum für Naturkunde, Erbprinzenstraße 13, 76133 Karlsruhe, Baden-Württemberg,Germany4Department of Wildlife Ecology and Management, University of Freiburg, Tennenbacher Straße 4, 79106 Freiburg,Baden-Württemberg, Germany
(Received 3 May 2013; revised 6 June 2013; accepted 9 June 2013)
SUMMARY
Culicoides vectors can transmit a diverse array of parasites and are globally distributed. We studied feeding preferences andseasonal variation of Culicoides (Diptera: Ceratopogonidae) vectors in an urban forest of Germany to determine whetherhumans living nearby are readily exposed to vector-borne parasites from wild animals. We used a fragment of the mtDNACOI gene to identify hosts from blood meals. We amplified a fragment of the mtDNA cyt b to detect haemosporidianinfections in Culicoides abdomens and thoraxes. We detected a total of 22 Culicoides species. Fifty-eight blood meals (84%)were from humans, 10 from birds, and one from livestock. We found Culicoides kibunensis (considered ornithophilic) with29 human bloodmeals. Host generalistCulicoides festivipennis andCulicoides obsoletus had 14 human bloodmeals.Culicoidesclastrieri and Culicoides semimaculatus fed on birds; previously humans were their only known host. Six thoraxes and threeabdomens were infected with either Haemoproteus pallidulus or Haemoproteus parabelopolskyi. There were changes inCulicoides community structure across months. Culicoides pictipennis was the dominant species during spring, C. kibunensisandC. clastrieriwere dominant during summer, andC. obsoletuswas dominant by early autumn. All dominant species weregeneralists feeding on birds, livestock and humans. Our results indicate that humans can serve as a blood source fordominant Culicoides species instead of the normal wild animal hosts in urban areas.
Key words: Culicoides, Haemoproteus, Sylvia atricapilla, blood meal analysis, emergent disease, conservation medicine,host–parasite interaction.
INTRODUCTION
Haemosporidian parasites are responsible formillions of infections and thousands of deaths inhumans, as well as domestic and wild animals eachyear (Valkiūnas, 2005; Enayati and Hemingway,2010). Infections may spread further owing tocurrent environmental changes because climatewarming enhances the geographical expansion ofboth parasites and vectors into regions from whichthey were previously absent (Jones et al. 2008).Despite their relevance for human and animal health,insect vectors feeding on different vertebrate speciesand transmitting haemosporidians are poorly known(Santiago-Alarcon et al. 2012a). This creates twochallenges. First, as the insect vectors of most ofthe haemosporidian parasites described to date are
unknown, we do not know how and with whomthese vectors directly and indirectly interact. Second,if vectors suck blood from humans and othervertebrates (e.g. Mayne, 1928; Coggeshall, 1940;Lefèvre et al. 2009), humans will be exposed to awiderange of parasites existing in the animal communitysurrounding them (Santiago-Alarcon et al. 2012b).Hence, it is important to know what blood parasitesmake it into the bloodstream of humans, even if theyinitially do not develop into an infection, because thisis the first step towards new emerging diseases(Arinaminpathy and McLean, 2009).Haemosporidians are mainly transmitted by five
families ofDiptera:Culicidae,Hippoboscidae, Simu-liidae, Ceratopogonidae and Psychodidae (Valkiūnas,2005; Santiago-Alarcon et al. 2012a). Until recentlyit was believed that most vector species had special-ized feeding habits, in general feeding on differentvertebrate classes, but recent molecular data havefound some vector species with generalized feedinghabits (e.g. Santiago-Alarcon et al. 2012b). Studies
* Corresponding author: Biología y Conservación deVertebrados, Instituto de Ecología A.C. CarreteraAntigua a Coatepec 351, El Haya, C.P. 91070 Xalapa,Veracruz, México. E-mail: [email protected]
1
Parasitology, Page 1 of 12. © Cambridge University Press 2013doi:10.1017/S0031182013001285

on host feeding preferences have demonstrated thatmany species of Culicoides (i.e. biting midges) arespecialized on either birds (e.g. Culicoides kibunensis)or mammals (e.g. Culicoides chiopterus, Culicoidesdeltus), but that some species are host generalists (e.g.Culicoides festivipennis, Culicoides obsoletus) (Bennett,1960; Burýlova, 1975; Santiago-Alarcon et al. 2012b).Host generalists are of special interest because theyare capable of feeding on different vertebrate groups,and thus can facilitate the emergence of new diseasesby spreading parasites to new hosts. Furthermore,under unfavourable or altered ecological conditions,vectors specialized on a specific vertebrate group canbe found to feed on suboptimal hosts (Lefèvre et al.2009). A recent study has shown that C. kibunensisreadily takes blood from humans in an urban forestof southwestern Germany, and that some of thesemidges are infected with avian blood parasites(Santiago-Alarcon et al. 2012b). These results,together with the knowledge that Culicoides vectorsare competent for a diverse array of parasites (e.g.viruses, trypanosomes, microfilarids, haemospori-dians: Borkent, 2005), suggest that we do not have aclear understanding of the parasites to which humansare exposed. Moreover, the above-mentioned studiesimply that parasite–vector–host interactions areless constant than typically acknowledged. Thismeans that, despite the fact that humans have beenhistorically exposed to a diverse array of parasites (i.e.people have not always lived in cities or towns),new parasite–human interactions may become estab-lished. Hence, novel health problems for humansmay arise under a scenario of geographical rangeexpansion of both insect vectors and the diseases theytransmit due to habitat alteration and climate change(e.g. Jones et al. 2008).
Even in the best-studied regions, we lack knowl-edge on the diversity of competent vectors transmit-ting haemosporidians. This limitation contrasts withthe better-known diversity of haemosporidian para-sites, particularly for birds in temperate areas(Santiago-Alarcon et al. 2012a). Some studies havecharacterized the seasonal variation in the occurrenceof avian haemosporidians in different bird species(e.g. Herman, 1938; Clark, 1964; Bennett andCameron, 1974; Super and van Riper, 1995; Benschand Åkesson, 2003; Santiago-Alarcon et al. 2011).However, few have compared those seasonal changesin parasite abundance with the abundance of putativevectors (e.g. Janovy, 1966; Greiner, 1975; Klei andDeGiusti, 1975; Atkinson, 1988; Garvin andGreiner, 2003). A time lag separates the emergenceof competent vectors and the onset of parasites in theperipheral blood of birds in temperate areas (Janovy,1966; Klei and DeGiusti, 1975; Atkinson et al. 1988;Sol et al. 2000), and so determining the temporalavailability of competent vectors is fundamental tounderstanding and predicting disease dynamics(Keeling and Rohani, 2008). Most previous studies
were conducted prior to the development of mol-ecular methods to identify vectors, parasites andhost blood meals, and so new opportunities nowexist to characterize potential pathways of diseasetransmission.
Here, we studied feeding preferences and seasonalvariation of Culicoides (Diptera: Ceratopogonidae)vectors in an urban forest (Mooswald) in the city ofFreiburg in southwestern Germany, and comparedthe observed seasonality with variation in haemo-sporidian prevalence in blackcaps between early(spring) and late breeding season (summer/autumn).Our study site is embedded in a matrix of buildingsand farms, and humans frequently use it for recrea-tion. This raises the possibility that humans may beexposed to pathogens of wildlife bitten by potentialvectors (e.g. ornithophilic Culicoides feeding onhumans). In this study, we determine (1) thevertebrate species from which Culicoides vectorsobtain blood meals, (2) the haemosporidian parasitesthat infect Culicoides vectors, (3) changes inCulicoides species and abundances across seasons,and (4) seasonal variation in haemosporidian preva-lence in blackcaps and its relationship to variation inCulicoides populations. This information will help usto understand exposure of urban human populationsto Culicoides vector species and vector-borne para-sites from wild animals.
MATERIALS AND METHODS
Bird species model and bird sampling
We used the blackcap (Sylvia atricapilla) as a systemto analyse associations between the temporal patternsof haemosporidian infections and the temporalavailability of haemosporidians putative Culicoidesvectors. The blackcap is a migratory passerine birdthat is abundant and widely distributed acrossEurope (Pérez-Tris et al. 2004) and well-studiedacross its geographical range (Pérez-Tris andTelleria, 2002; Telleria and Pérez-Tris, 2003;Bearhop et al. 2005; Rolshausen et al. 2009, 2010).Haemosporidian parasites infecting this bird havebeen sampled across Western and Central Europe(Pérez-Tris et al. 2007; Santiago-Alarcon et al. 2011),with some samples from Sweden and the Baltic Searegion (Križanauskienė et al. 2010). It is common tofind high prevalence (>70%) ofHaemoproteus spp. inblackcaps across their distributional range (Pérez-Tris and Bensch, 2005a,b; Pérez-Tris et al. 2007;Križanauskienė et al. 2010; Santiago-Alarcon et al.2011). However, these parasites seem to have nosignificant effect on the condition of this bird speciesother than a short-term weight loss (Valkiūnas et al.2006). Haemoproteus parabelopolskyi, a parasite ofblackcaps, is transmitted by the midge Culicoidesimpunctatus in Eastern Europe (Curonian Spit,Russia), which is considered to be an important
2Diego Santiago-Alarcon and others

vector of Haemoproteus species across Europe(Valkiūnas et al. 2002; Valkiūnas and Iezhova,2004a,b). In addition, preliminary data indicate thatH. parabelopolskyi can be transmitted by C. kibu-nensis in southern Germany, where C. impunctatusis rare (Santiago-Alarcon et al. 2012b).We captured blackcaps in southwestern Germany,
in Freiburg (48°00′N07°51′E) from 2007 to 2011 andin Radolfzell (47°45′N, 08°59′E) in 2006, using mistnets aided by tape recordings of blackcap song. Weanalysed haemosporidian infection of 166 blackcapscaught during spring and autumn 2006, 169 caughtduring spring and summer 2007, 120 caught duringautumn 2009 (see Santiago-Alarcon et al. (2011) fordetailed sample sizes), and 18 during summer 2011.Each individual bird was marked with a standardaluminium ring, sexed, and aged based on plumageand skull pneumatization (Shirihai et al. 2001). Wetook *75 μL of blood from the brachial vein of eachcaptured bird using heparinized capillary tubes andstored the samples at −20 °C in the laboratory. Eachbird was released after sampling.
Vector sampling
We collectedCulicoidesmidges during 2010 and 2011using BG Sentinel® traps (Regensburg, Germany)with UV light; we added dry ice (CO2 source) as anadditional attractor in 2011. Collections were madein an urban forest of Freiburg in SouthwesternGermany (Mooswald Forest, 48°00N, 07°51E). Thisforest is a mixed deciduous forest with water availablein small streams and ponds whereDiptera vectors candevelop. The forest contains many walking trailsincluding some forest roads. Traps were operatedovernight starting at 18:00 h, and picked up nextmorning at 08:00 h. We sampled from late Marchuntil the beginning of October in 2010 conducting atotal of 32 trapping nights using two traps, whichyielded 64 samples; we detected no Culicoides inMarch and we did not sample in August. From Juneto August 2011, we conducted 14 trapping nightsusing two traps for a total of 28 samples. Insectscollected in 2010 were used to study the phenology ofthe Culicoides community.Samples were subsequently processed in the lab
with a stereomicroscope; each Culicoides sample wasdivided into engorged (i.e. insects with a full orpartial blood meal) and unfed midges. Engorgedmidges from 2011 were used for blood meal analysis;bloodmeal results for midges from 2010 can be foundin Santiago-Alarcon et al. (2012b). We dissected theabdomen and thorax of each individual midge;dissecting tools were sterilized with alcohol andflame for every insect. We extracted the DNA fromthe blood contained in the abdomen and conductedPCR (see below). Midges that previously had a bloodmeal (i.e. parous) might have been infected with
haemosporidian parasites, in which case differentdevelopmental stages could be present in the insect’smid gut wall (i.e. oocysts) or in the salivary glands(i.e. sporozoites), which would imply that thesevectors could inject haemosporidian sporozoitesduring subsequent feeding bouts. The thorax ofeach dissectedmidge collected in 2011 was also storedin 70% alcohol for subsequent laboratory work (DNAextraction and PCR to detect parasites). The head,genitals, legs, and wings of dissected midges weremounted on microscope slides in Euparal forsubsequent morphological identification. Unfedmidges were kept in 70% alcohol until they wereprocessed for identification. Determination wasconducted using taxonomic keys (Kremer, 1965;Havelka, 1976; Országh, 1976; Szadziewski, 1984;Delécolle, 1985). Representative Culicoides speci-mens were mounted on slides and they are kept inthe insect collection of Havelka and Aguilar at theStaatliches Museum für Naturkunde in Karlsruhe,Germany.
Molecular work
We extracted DNA from blood meals, abdomens andthoraxes of theCulicoidesmidges and frombird bloodsamples using the DNeasy Blood and Tissue® kit(QIAGEN, Hilden). DNA quality was checked on a1·2% agarose gel. We conducted a nested PCR toamplify the bar code (COImitochondrial DNA gene)of each blood meal using the protocol described inAlcaide et al. (2009), which amplifies *758 bp. Wealso used parasite genus-specific primers in a nestedPCR protocol that amplifies a fragment of *524 bpof the cytochrome b mitochondrial DNA geneof haemosporidian parasites (Hellgren et al. 2004)to verify whether avian blood samples and vectorabdomens and thoraxes were infected. All PCRswere run with positive controls (blackcap haemo-sporidian-infected blood sample, and blood samplesfrom another five bird species (blue tit (Cyanistescaeruleus), great tit (Parus major), robin (Erithacusrubecula), blackbird (Turdus merula) and songthrush(Turdus philomelos) as positives for the blood mealanalysis) and a negative control (ddH2O). Sub-sequently, we cleaned the PCR products with theMinElute kit (QUIAGEN,Hilden), placed them in a96-well plate and sent them out for sequencing atMicrosynth AG, Switzerland. Sequences were editedwith 4Peaks v.1.7.2 (mekentosj.com). We used theidentification engine of the BOLD (Barcode of LifeData Systems) systems v2.5 (Ratnasingham andHebert, 2007) to identify the vertebrate species foreach COI sequence obtained from the blood meals.Cyt b sequences obtained from haemosporidian-infected blood meals, abdomens and thoraxes werecompared against DNA sequences available inGenBank™ by using the BLAST algorithm of the
3Culicoides blood meal analysis in urban forest

NCBI (National Center for Biotechnology Infor-mation) database. Sequences were deposited inGenBank™ (accession numbers for mtDNA cyt b:KF192995-KF193002, and for mtDNA COI:KF193003-KF193056).
Statistical analyses
To analyse changes in Culicoides species throughoutthe study period, we used a χ2 test to compare theabundances of Culicoides recorded in each month,assuming as expected values the average monthlyabundance. Subsequently, to detect changes inassemblage structure, we contrasted the monthlyrank-abundance curves (Feinsinger, 2001) and thenwe calculated the Exponential Shannon Index usingthe SPADE program (Chao and Shen, 2010), whichreflects the number of equivalent species with respectto their relative abundance (Jost, 2006, 2010).Finally, we evaluated changes in assemblage struc-ture between months through a randomization test,assigning 10000 random partitions (Solow, 1993,two-sided test). The test was performed in theSpecies Diversity and Richness v.3.02 program
(Henderson and Seaby, 2002). Compositional simi-larity between contiguous months was analysed usingthe Jaccard Index based on incidence, as well as theJaccard Index based on abundance (including thecalculation of standard error by means of a boot-strapping procedure), both indices were calculatedusing the SPADE program (Chao and Shen, 2010).
We used the programQuantitative Parasitology 3.0(Rózsa et al. 2000) to calculate unbiased haemo-sporidian prevalence estimates. We estimated 95%confidence intervals by Sterne’s exact method(Reiczigel, 2003).
RESULTS
Blood meal analysis
We trapped 98 engorged (i.e. insects with full orpartial blood meals) Culicoides midges of 13 differentspecies (Table 1). We obtained good-quality COIsequences from 69 blood meals; most likely the DNAin the other 29 blood meals was degraded due todigestion in the vector’s mid gut. Fifty-eight bloodmeals (84%) were derived from humans, 10 frombirds (E. rubecula, T. philomelos, Tadorna ferruginea),
Table 1. Blood meals from Culicoides vector species found in the Mooswald forest of Freiburg duringsummer 2011
Vector species
Numberof bloodmeals
Vertebrate hostdetected inblood mealsa Previously known vertebrate hostsb
Culicoides clastrieri 22 Homo sapiens (9)Tadorna ferruginea (1)Turdus philomelos (1)
Man15
C. chiopterus 0 Cows, man8, 11, 12, 13, 15, 18, birds9
C. deltus 2 Homo sapiens (1) Horses, cows, man15, 18
C. dewulfi 5 Homo sapiens (3) Man15
C. duddingstoni 0 NIC. festivipennis 17 Homo sapiens (13) Cows, man, birds5, 7, 11, 15, 18
C. heliophilus 0 Man, cows, sheep, dogs6, 8, 13–15, 18
C. kibunensis 40 Erithacus rubecula (6)Homo sapiens (29)
Birds3, 11, 18 man15
C. newsteadi 1 Homo sapiens (1) Man, poultry2, 18
C. obsoletus 3 Homo sapiens (1) Birds, man, livestock1, 3, 4, 12, 14, 15, 18
C. pallidicornis 2 Cow (1) Sheep, cows, birds, man4, 6, 10, 13–15, 18
C. pictipennis 1 Erithacus rubecula (1) Birds, man3, 4, 11, 14, 15, 18
C. poperinghensis 0 Man15
C. pseudoheliophilus 0 NIC. pulicaris 1 UN Cows, sheep, horses, buffaloes, man4, 6, 8, 12–15, 18
C. punctatus 1 UN Man, cows11, 12, 18, birds9
C. scoticus 2 Homo sapiens (1) Cows, horses, sheep, man15, 18, birds9
C. semimaculatus 1 Erithacus rubecula (1) Man15
C. stigma 0 Cows, man6, 12, 18
C. vexans 0 Horses, goats, man, birds8, 10, 13, 18
C. impunctatus 0 Birds16, 17
NI = no information available; UN= unidentified host (PCR failed to amplify or sequence was of low quality).a Number of times a vertebrate host was identified from a blood meal is in parenthesis.b References: (1) Bennett (1960); (2) Braverman et al. (1977); (3) Burýlova (1975); (4) Butenko (1967); (5) Fritz (1982);(6) Havelka (1976); (7) Kitaoka et al. (1965); (8) Kremer (1965); (9) Lassen et al. (2011); (10) Minár (1965); (11) Nielsen(1964); (12) Nielsen (1971); (13) Országh (1976); (14) Paclt et al. (1970); (15) Santiago-Alarcon et al. (2012a,b);(16) Valkiūnas and Iezhova (2004a); (17) Valkiūnas and Iezhova (2004b); (18) http://www.bluetonguevirus.org/culicoides
4Diego Santiago-Alarcon and others

and one from livestock (cow). We identified eightCulicoides species with blood meals derived fromhumans (Table 1). For a second time, we foundC. kibunensis, previously known to feed only on birds,with 29 human blood meals; we also detected twohost generalist (i.e. bird- and mammal-feeding)Culicoides species (C. festivipennis, C. obsoletus) with14 human blood meals. We present for the first timebird records for Culicoides clastrieri and C. semima-culatus, which makes them generalists consideringthat we previously found them feeding on humans(Table 1). We corroborate our previous finding onCulicoides dewulfi feeding exclusively on humans(Table 1).We found haemosporidian parasites in the thoraxes
of six midges and the abdomens of three additionalmidges (Table 2). Haemoproteus pallidulus (lineagesSYAT03 and SYAT35) infected four specimensof C. kibunensis and one of Culicoides pictipennis.One C. kibunensis infected with H. pallidulus fedon E. rubecula, which represents a new bird hostrecord for this parasite.Haemoproteus parabelopolskyi(lineages SYAT01 and SYAT02) infected two indi-viduals of C. kibunensis and one of Culicoides scoticus.
Culicoides diversity and seasonality
We identified 22 Culicoides species in the Mooswaldforest. Vector sampling in 2011 added Culicoidesfurcillatus (only one specimen was collected) to the 21species identified during 2010 (see Santiago-Alarconet al. 2012b). During 2010, species richness permonth varied between 15 species (in June) and fourspecies (in October; Table 3). Altogether, we caught851 Culicoides individuals, with notable differencesbetween months (χ2 = 167·2, P<0·001, D.F. = 5):the highest abundance (216 individuals) occurredin September, while the lowest (56 individuals)
occurred in both May and October (Table 3).Culicoides obsoletus and C. pictipennis were the mostabundant species in the study, with 252 and 249individuals respectively, which together represents59% of all species recorded. In contrast, eight specieswere recorded just once or twice: C. chiopterus,C. deltus, Culicoides newsteadi, Culicoides punctatus,C. semimaculatus, Culicoides stigma, C. impunctatusand Culicoides vexans. Concerning temporal distri-bution, three species were recorded in all months ofthe study (C. dewulfi, C. obsoletus and C. scoticus),while six species were recorded in just 1 month (seeTable 3). The remaining species were detected in 2 to5 months.The structure of the species assemblage changed
over the study period. Besides variation in speciesnumber, there were also changes in relative abun-dance and hence in the hierarchical position of species(Fig. 1). The slopes of range-abundance curves weresteeper in the first and last months of the study,while during the intermediate months, in particularJune and July, the slopes were shallower, indicatinghigher evenness in the assemblage. As a consequence,the Exponential Shannon Index values were higher(see Fig. 1). Assemblage structure did not differbetween June and July, but these months differedfrom the assemblage structure of the other months.Also, assemblage structure in October differedfrom all other sampled months, while no differenceswere found between April, May and September (seeSupplementary Table S1 – in Online version onlyfor delta values and P-values). We detected a changein the dominant species throughout the study period.Culicoides pictipennis was the dominant species dur-ing April–June, but in July its hierarchical positiondecreased due to a notable decrease in abundance,and only one individual was recorded in August.No individuals of C. pictipennis were detected during
Table 2. Haemosporidian parasites found in blood meals (abdomens) and thoraxes from Culicoides speciescollected in the Mooswald forest of Freiburg during summer 2011
Vector species
Vertebrate host identifiedfrom haemosporidian-infectedblood meal
Body part whereparasite was detected Haemosporidian parasite
C. kibunensis Erithacus rubecula (1)a,UN (1), NBP (2)
Abdomen (1); Thorax (3) Haemoproteus pallidulus (4)[SYAT03 and SYAT35]
C. kibunensis NBP (3) Abdomen (1); Thorax (2) Haemoproteus parabelopolskyi (2)[SYAT01 and SYAT02];Haemoproteus sp. (1)[several lineages matched the sequence,unable to identify species]
C. pictipennis NBP Thorax (1) Haemoproteus pallidulus (1)[SYAT03 and SYAT35]
C. scoticus NBP Abdomen (1) Haemoproteus parabelopolskyi (1)[SYAT01 and SYAT02]
UN= unidentified host (PCR failed to amplify or sequence was of low quality); NBP = no blood, parous specimen.Parasite lineages are indicated in brackets and the number of times the interaction was observed is indicated in parentheses.a New bird host record for H. pallidulus (Sylvia atricapilla was the only known host, see Križanauskienė et al. 2010).
5Culicoides blood meal analysis in urban forest
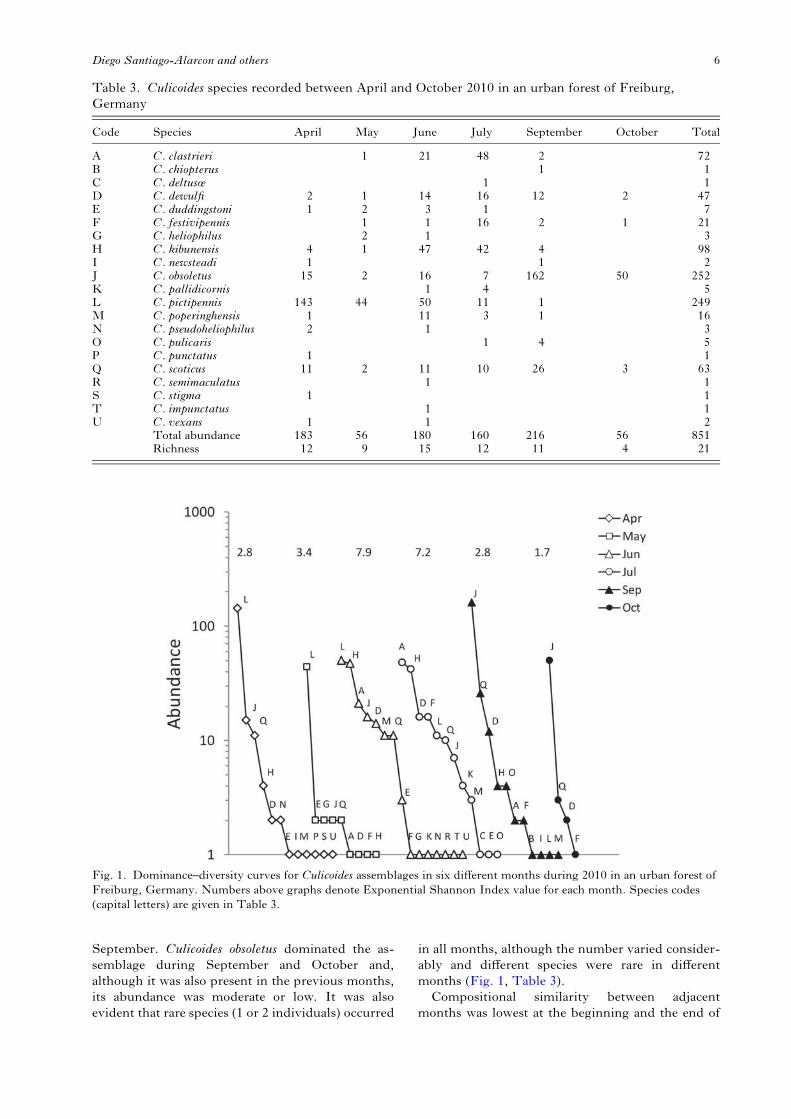
September. Culicoides obsoletus dominated the as-semblage during September and October and,although it was also present in the previous months,its abundance was moderate or low. It was alsoevident that rare species (1 or 2 individuals) occurred
in all months, although the number varied consider-ably and different species were rare in differentmonths (Fig. 1, Table 3).
Compositional similarity between adjacentmonths was lowest at the beginning and the end of
Table 3. Culicoides species recorded between April and October 2010 in an urban forest of Freiburg,Germany
Code Species April May June July September October Total
A C. clastrieri 1 21 48 2 72B C. chiopterus 1 1C C. deltusœ 1 1D C. dewulfi 2 1 14 16 12 2 47E C. duddingstoni 1 2 3 1 7F C. festivipennis 1 1 16 2 1 21G C. heliophilus 2 1 3H C. kibunensis 4 1 47 42 4 98I C. newsteadi 1 1 2J C. obsoletus 15 2 16 7 162 50 252K C. pallidicornis 1 4 5L C. pictipennis 143 44 50 11 1 249M C. poperinghensis 1 11 3 1 16N C. pseudoheliophilus 2 1 3O C. pulicaris 1 4 5P C. punctatus 1 1Q C. scoticus 11 2 11 10 26 3 63R C. semimaculatus 1 1S C. stigma 1 1T C. impunctatus 1 1U C. vexans 1 1 2
Total abundance 183 56 180 160 216 56 851Richness 12 9 15 12 11 4 21
Fig. 1. Dominance–diversity curves for Culicoides assemblages in six different months during 2010 in an urban forest ofFreiburg, Germany. Numbers above graphs denote Exponential Shannon Index value for each month. Species codes(capital letters) are given in Table 3.
6Diego Santiago-Alarcon and others
the sampling season (Jaccard Index based on inci-dences, JIin = 0·40 and 0·36, respectively), whilein intermediate adjacent months (i.e. summer)compositional similarity was higher (JIin = 0·59and 0·64; Fig. 2a). However, when compositionalsimilarity was calculated using abundances (JaccardIndex based on abundances, JIab) the observedpattern was different. The lowest compositionalsimilarity was detected at the beginning of thesampling season (JIab = 0·89), then increased untilreaching a maximum value between June and July(JIab = 0·96) and subsequently slightly decreasedtowards the end of the study season (JIab = 0·94;Fig. 2b). Values of JIab agree with those of theExponential Shannon Index (Fig. 1), which indicateshigher evenness (i.e. more similar abundances amongspecies) for the summer months.
Parasite prevalence
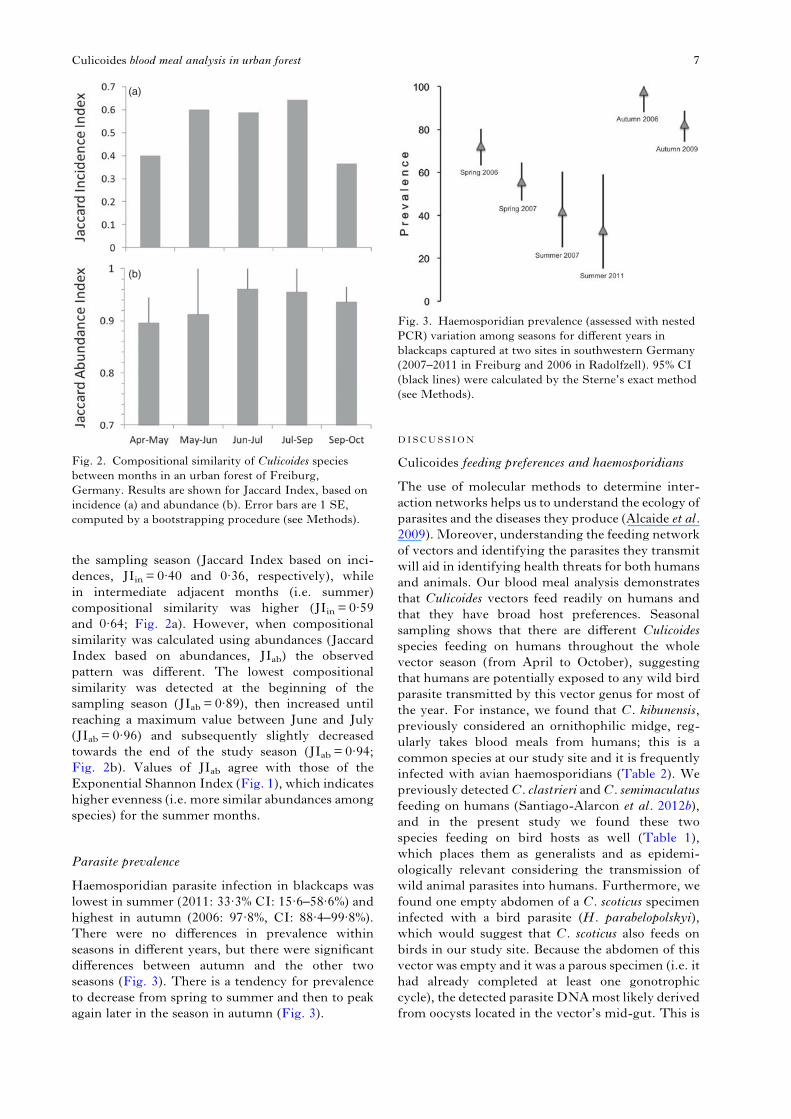
Haemosporidian parasite infection in blackcaps waslowest in summer (2011: 33·3% CI: 15·6–58·6%) andhighest in autumn (2006: 97·8%, CI: 88·4–99·8%).There were no differences in prevalence withinseasons in different years, but there were significantdifferences between autumn and the other twoseasons (Fig. 3). There is a tendency for prevalenceto decrease from spring to summer and then to peakagain later in the season in autumn (Fig. 3).
DISCUSSION
Culicoides feeding preferences and haemosporidians
The use of molecular methods to determine inter-action networks helps us to understand the ecology ofparasites and the diseases they produce (Alcaide et al.2009). Moreover, understanding the feeding networkof vectors and identifying the parasites they transmitwill aid in identifying health threats for both humansand animals. Our blood meal analysis demonstratesthat Culicoides vectors feed readily on humans andthat they have broad host preferences. Seasonalsampling shows that there are different Culicoidesspecies feeding on humans throughout the wholevector season (from April to October), suggestingthat humans are potentially exposed to any wild birdparasite transmitted by this vector genus for most ofthe year. For instance, we found that C. kibunensis,previously considered an ornithophilic midge, reg-ularly takes blood meals from humans; this is acommon species at our study site and it is frequentlyinfected with avian haemosporidians (Table 2). Wepreviously detectedC. clastrieri andC. semimaculatusfeeding on humans (Santiago-Alarcon et al. 2012b),and in the present study we found these twospecies feeding on bird hosts as well (Table 1),which places them as generalists and as epidemi-ologically relevant considering the transmission ofwild animal parasites into humans. Furthermore, wefound one empty abdomen of a C. scoticus specimeninfected with a bird parasite (H. parabelopolskyi),which would suggest that C. scoticus also feeds onbirds in our study site. Because the abdomen of thisvector was empty and it was a parous specimen (i.e. ithad already completed at least one gonotrophiccycle), the detected parasite DNAmost likely derivedfrom oocysts located in the vector’s mid-gut. This is
(a)
(b)
Fig. 2. Compositional similarity of Culicoides speciesbetween months in an urban forest of Freiburg,Germany. Results are shown for Jaccard Index, based onincidence (a) and abundance (b). Error bars are 1 SE,computed by a bootstrapping procedure (see Methods).
Fig. 3. Haemosporidian prevalence (assessed with nestedPCR) variation among seasons for different years inblackcaps captured at two sites in southwestern Germany(2007–2011 in Freiburg and 2006 in Radolfzell). 95% CI(black lines) were calculated by the Sterne’s exact method(see Methods).
7Culicoides blood meal analysis in urban forest

the second time C. scoticus is registered feedingon birds (see Lassen et al. 2011); previously thisspecies was known to feed only on non-humanmammals, but our results indicate that this speciescommonly feeds on humans, as well (Table 1, seealso Santiago-Alarcon et al. 2012b). Additionally,our analysis has also detected new parasite–birdassociations. Haemoproteus pallidulus is a recentlydescribed species that is known to infect onlyblackcaps (Križanauskienė et al. 2010). Here, wedetected H. pallidulus infecting the abdomen ofa C. kibunensis vector that fed on a robin, whichimplicates the robin (E. rubecula) as a new bird hostfor this parasite. Cautionmust be practiced, however,before a definitive statement on this new record canbe made because we have only found genetic lineagesof Plasmodium relictum, Haemoproteus balmorali andLeucocytozoon majoris infecting robins at our studysite (Santiago-Alarcon et al. unpublished results)and because the vector was parous, meaning thatinfection with H. pallidulus could have derived froma previous feeding bout on a different bird species(e.g. blackcap). Finally, C. impunctatus, which is thecompetent vector for H. parabelopolskyi in EasternEurope, is rare in surveys conducted in Freiburg,which seems to indicate that the competent vector inEastern Europe for H. parabelopolskyi is replaced insouthwest Germany by C. kibunensis, and to a lesserdegree, based on their abundances, also byC. scoticusand Culicoides poperinghensis (Santiago-Alarcon et al.2012b; Tables 1 and 2). Thus, our results indicatethat many unknown haemosporidian–host associ-ations that are potentially relevant for human andanimal health remain to be discovered.
Experimental studies have demonstrated that anavian parasite, Plasmodium lophurae, can adapt to andbe viable in mice after just four rounds of infectiousinoculations (McGhee, 1951). Furthermore, erythro-cytes of different mammal species have been shownto be susceptible to invasion by bird Plasmodiumparasites (McGhee, 1957). These studies suggest thathaemosporidian parasites can adapt to new distanthosts rather quickly. Our results show that manyCulicoides species of medical and veterinary impor-tance are feeding across a diverse array of vertebratehosts (Table 1), which posit them as ideal vectors totransmit parasites across large taxonomic distances.Many of those generalist vector species are suscep-tible to avian haemosporidian parasites (Plasmodiumsp. and Haemoproteus sp., Santiago-Alarcon et al.2012b; Table 2). This is important because thepotential of parasites to rapidly adapt to new hostswill be one of the greatest challenges to human andanimal health, as well as of increasing importance inthe study of zoonotic diseases. Although Culicoidesvectors infected with avian haemosporidians couldtransmit these parasites to humans, it is difficult topredict if a disease will eventually develop becauseparasites not adapted to the immune system of novel
hosts can be rapidly detected and cleared (Ewald,1994).
Plasticity of feeding preferences could vary fordifferent vector families in an urban setting. Forexample, Kim et al. (2009) studied feeding prefer-ences of Culex sasai (Diptera: Culicidae) in an urbanpark in Tokyo, Japan; different from our Culicoidesresults, this mosquito species seems to rarely usehumans as part of its diet despite its generalist feedinghabit (it fed on both birds and livestock). This resultis for a singleCulex species, however, and it might notbe representative of the feeding habits of Culicidaein general. For instance, Lefèvre et al. (2009) hasfound a plastic feeding response ofAnopheles gambiaemosquitoes in a rural community of Burkina Faso,where preferences from humans have changedtowards livestock due to year-round use of mosquitonets. Moreover, Culex quinquefasciatus alternatedtheir feeding preferences between humans and live-stock depending on the time of the year (Lefèvre et al.2009). Hence, a more thorough analysis of Culicidaeand other Diptera vector families feeding preferencesis needed before generalizations are made.
In reference to viral diseases transmitted byCulicoides, bluetongue virus (BTV, Reoviridae:Orbivirus) was detected in central Europe in 2006(Mehlhorn et al. 2008). This is a disease of domesticlivestock and wild ruminants that can create sub-stantial economic problems (Mehlhorn et al. 2007,2008). Culicoides obsoletus, Culicoides pulicaris andC. scoticus are considered the main vectors of BTV inGermany and obtain their blood meals only fromlivestock (Mehlhorn et al. 2007; Bartsch et al. 2009).In our previous (Santiago-Alarcon et al. 2012b) andpresent study we found C. obsoletus, C. scoticus andC. pulicaris feeding on humans (Table 1). Given thatBTV is already established in Germany, BTV couldpotentially also invade human blood via infectiousCulicoides bites, though BTV and other Orbivirus(Arbovirus: Reoviridae) apparently do not produceacute illness in humans (Mellor et al. 2000; Quanet al. 2008; Gethmann et al. 2010; but see Calisher,1994).
In areas with some degree of habitat modification(e.g. suburban and urban forests), infectious vectorscan bite hosts that can turn to be suboptimal for boththe parasite and the vector (i.e. host blood factors canbe detrimental for vectors). In the case of urbanforests or other areas with high densities andabundance of humans, humans can serve as a bloodsource that is a bigger and more accessible target thanthe wild animal source. Thus, humans can interceptvector infectious bites and can be acting either as adead-end host or as a highly mobile reservoir capableof carrying viable wild animal parasites to otherregions inhabited by naïve animal communities (e.g.islands). Moreover, continuous exposure of humansto novel parasites poses potential health problemscreated by emergent diseases. It is, therefore, a
8Diego Santiago-Alarcon and others

priority to study the biodiversity of haemosporidianparasites in wild vertebrates and of the vectorstransmitting them since we must first know what ispresent to be prepared for any epidemiologicalemergency. Thus, future studies should focus onurban and suburban forests, where the transmissionof putative non-human parasites into humans is morelikely compared with sparsely populated rural areas(e.g. Bradley and Altizer, 2007).
Culicoides diversity and seasonality
We found a total of 22 species at one location inFreiburg (Mooswald Forest, 48°00N, 07°51E).Fifty-six species of Culicoides have been registeredfor Germany (Havelka and Aguilar, 1999). Twenty-five Culicoides species (removing Culicoides puncti-collis from their list, which is a synonym ofC. nubeculosus) have been recorded at 20 locations innorthwest Germany (Kiel et al. 2009), 10 species at19 locations in the state of Nordrhein-Westfalen(Mehlhorn et al. 2009), and 22 species further southat 12 locations in the states of Saarland andRheinland-Pfalz (Vorsprach et al. 2009). A surveyof Culicoides species in the state of Baden-Wuertemberg, where our sampling was conducted,have only determined species groups (i.e. C. obsoletuss.l., and C. pulicaris s.l.) and no further details onspecies identities were provided (Hörbrand andGeier, 2009). Twenty-nine species of Culicoideshave been identified for the state of Baden-Wuertemberg (Havelka unpublished information).We observed a peak of species richness in April(n = 12) and then another in June (n = 15); speciesrichness rapidly declined in October (n = 4). Thispattern of species richness is different to the oneobserved in more northern regions of Germany,where the highest number of species was recordedduring September and October (Vorsprach et al.2009). It is remarkable that we found at a singlelocation about the same number of Culicoides speciesas the above-mentioned studies, which sampledbroadly and throughout the whole year. This canpertain to different climatic conditions that are foundin Freiburg and the southern parts of Germany, moreMediterranean-like weather in late spring andthroughout the summer, which would certainlyinfluence faunal diversity. Hence, we expect to finda much larger species count and possibly differentspecies composition in this region of Germany whena more intense and complete sampling is performed.Assemblages were more similar during summer
months compared with early spring and earlyautumn, where Culicoides communities were lesssimilar in both abundance and species composition(Fig. 1). There was a change in species compositionacross the study period; of particular importance arethe changes in dominant species at the beginning,
middle and end of the study period. Culicoidespictipennis was the dominant species during springand was gradually declining as the season progressed,and completely disappeared by the end of thesummer. Culicoides kibunensis and C. clastrieri weredominant during summer months, disappearing bythe end of the summer. Culicoides obsoletus was thedominant species by the end of the summer and earlyautumn, which is in agreement with other surveysconducted in Germany (Hörbrand and Geier, 2009;Mehlhorn et al. 2009; Vorsprach et al. 2009).Culicoides dewulfi, C. obsoletus and C. scoticus werepresent during the whole season, of these C. scoticusmaintained similar abundances among the differentsampling months (Table 3). All these Culicoidesspecies are generalists, feeding on humans, livestockand birds (Santiago-Alarcon et al. 2012b, Table 1),except C. dewulfi, which is known to feed only onhumans (Santiago-Alarcon et al. 2012b, Table 1).The relevant issue is that these Culicoides species(including C. festivipennis) are abundant and dom-inate the vector community throughout the aviancommunity’s breeding season, which makes thempotential vectors for parasite transmission acrossvertebrate groups. For instance, C. obsoletus,C. scoticus and C. pulicaris are known vectors ofviruses that are lethal to livestock (Mehlhorn et al.2007; Bartsch et al. 2009, see above), and thesespecies readily feed on humans (Santiago-Alarconet al. 2012b, Table 1). Furthermore, we have de-tected abdomens and thoraxes of C. kibunensis andC. pictipennis specimens infected with DNA of avianHaemoproteus and Plasmodium parasites (Santiago-Alarcon et al. 2012b, Table 2). Considering thatCulicoides are able to transmit a large array ofparasites (e.g. viruses: Mellor et al. 2000; trypano-somes, microfilarids, haemosporidians: Borkent,2005), that most abundant and generalist midgesare present throughout the whole season (despitesignificant changes in assemblage structure), and thatthey readily feed on humans, it is a priority to studythe feeding preferences of dipteran vectors and theparasites they are susceptible to in an urban/suburbansetting, where human populations are denser andchances of novel zoonosis are higher. We propose,under a conservation medicine framework, to designstudies focused on a gradient going from urban tosuburban forests and to more isolated well-conservedforests, in order to identify threats to human healthas feeding preferences and composition of parasiteand vector communities change along the habitatperturbation gradient (see also Bradley and Altizer,2007).
Seasonal haemosporidian prevalence
Blackcap (S. atricapilla) haemosporidian prevalencewas >50% during spring and it decreased duringsummer (30–40%), peaking later on during early
9Culicoides blood meal analysis in urban forest

autumn (Fig. 3). The way we interpret this patternis as follows: (1) because many blackcaps arrive inmid-March when no detectableCulicoides vectors arepresent at our study site, we assume that there is alarge fraction of blackcaps returning from winteringquarters that are already infected (see Santiago-Alarcon et al. 2011), and a few others should acquireinfection in breeding grounds during April and earlyMay, when midges are already available for parasitetransmission (i.e. infection probability increases laterduring the breeding season, see Santiago-Alarconet al. 2013); (2) parasite prevalence decreases towardsthe summer because new susceptible birds are addedto the blackcap population, ‘diluting’ the proportionof birds already infected; and (3) because during thebreeding period and throughout the summer there isa well-developed Culicoides vector community, mostchicks/fledglings could get infected. Parasites inperipheral blood will not be detected until latesummer or early autumn, however, when parasiteshave completed merogony in internal organs ofbirds. It is particularly relevant that C. kibunensis,one of the most common midges throughout thesummer, has been detected feeding on blackcapsat our study population (Santiago-Alarcon et al.2012b); moreover, we detected C. kibunensis abdo-mens and thoraxes infected with H. parabelopolskyi(see Table 2), a common parasite of blackcaps(Santiago-Alarcon et al. 2011). Thus, the observedpattern is in agreement with that found in otherstudies conducted in temperate regions, where thereis a time lag between the availability of competentvectors and the onset of haemosporidians in theperipheral blood of birds (Janovy, 1966; Klei andDeGiusti, 1975; Atkinson et al. 1988).
ACKNOWLEDGEMENTS
We thank R. Mettler, C. Hermes, M. Melchior andM. Hoffman for assistance during fieldwork. R. E.Ricklefs and two anonymous reviewers provided commentsthat improved the quality of the paper.
FINANCIAL SUPPORT
This work was supported by the Deutsche Forschungsge-meinschaft (H. M. S., grant number 1008/6-1); theAlexander von Humboldt Foundation (D.S.-A., post-doctoral grant); and by the Wissenschaftliche GesellschaftFreiburg (H. M. S. and G. S.).
DATA ACCESSIBILITY
Because we have no newmtDNACOI or cyt b sequences toreport to GenBank™, we have decided instead to providetext files containing all sequences used for our analyses inFASTA format. We are providing two files, one for eachgene, which are named: SupplementaryData S1 – inOnlineversion only (contains: mtDNA-COI blood meal sequencedata) and Supplementary Data S2 – in Online version only(contains: mtDNA-Cyt b haemosporidian sequence data).We provide an Excel file (Supplementary Data S3 – in
Online version only) containing Culicoides data used fordiversity analyses collected in 2010, the same file alsoincludes numbers of Culicoides species collected duringsummer 2011 and the complete list of 2011 Culicoidesthat were dissected for subsequent molecular analyses(blood meal and haemosporidian parasite screening ofboth thoraxes and abdomens). Finally, we provide an Excelfile (Supplementary Data S4 – in Online version only) withthe haemosporidian infection data for blackcaps for thedifferent years reported in the study. Data are archived inDryad doi: 10.5061/dryad.5nt04.
AUTHOR CONTRIBUTIONS
DS-A wrote the paper; DS-A and HMS designed thestudy; DS-A and PH mounted Culicoides specimens andPH made all Culicoides taxonomic identifications; EPconducted all diversity analyses; DS-A and GS did allmolecular work; HMS provided valuable insights through-out the different versions of the paper.
SUPPLEMENTARY MATERIAL
To view supplementary material for this article, please visithttp://dx.doi.org/10.1017/S0031182013001285.
REFERENCES
Alcaide, M., Rico, C., Ruíz, S., Soriguer, R., Muñoz, J. andFiguerola, J. (2009). Disentangling vector-borne transmission networks:a universal DNA barcoding method to identify vertebrate hosts fromarthropod blood meals. PLoS ONE 4, e7092.Arinaminpathy,N. andMcLean, A. R. (2009). Evolution and emergenceof novel human infections. Proceedings of the Royal Society of London, B 276,3937–3943.Atkinson, C. T. (1988). Epizootiology of Haemoproteus meleagridis(Protozoa: Haemosporina) in Florida: potential vectors and prevalence innaturally infectedCulicoides (Diptera: Ceratopogonidae). Journal ofMedicalEntomology 25, 39–44.Atkinson, C. T., Forrester, D. J. andGreiner, E. C. (1988). Epizootiologyof Haemoproteus meleagridis (Protozoa: Haemosporina) in Florida: seasonaltransmission and vector abundance. Journal of Medical Entomology 25,45–51.Bartsch, S., Bauer, B., Wiemann, A., Clausen, P.-H. and Steuber, S.(2009). Feeding patterns of biting midges of the Culicoides obsoletus andCulicoides pulicaris groups on selected farms in Brandenburg, Germany.Parasitology Research 105, 373–380.Bearhop, S., Fiedler, W., Furness, R.W., Votier, S. C., Waldron, S.,Newton, J., Bowen, G. J., Berthold, P. and Farnsworth, K. (2005).Assortativemating as amechanism for rapid evolution of a migratory divide.Science 310, 502–504.Bennett, G. F. (1960). On some ornithophilic blood-sucking dipterain the Algonquin Park, Ontario, Canada. Canadian Journal of Zoology 38,377–389.Bennett, G. F. and Cameron, M. (1974). Seasonal prevalence of avianhematozoa in passeriform birds of Atlantic Canada. Canadian Journal ofZoology 52, 1259–1264.Bensch, S. and Åkesson, S. (2003). Temporal and spatial variation ofhematozoans in Scandinavian Willow warblers. Journal of Parasitology 89,388–391.Borkent, A. (2005). The biting midges, the Ceratopogonidae (Diptera). InBiology of Disease Vectors, 2nd Edn (ed. Marquardt, C.), pp. 113–126.Elsevier Academic Press, San Diego, USA.Bradley, C. A. and Altizer, S. (2007). Urbanization and the ecology ofwildlife diseases. Trends in Ecology and Evolution 22, 95–102.Braverman, Y., Boreham, P. F. L., Galun, R. and Ziv, M. (1977). Theorigin of blood meals of biting midges (Diptera: Ceratopogonidae) andmosquitoes (Diptera: Culicidae) trapped in turkey runs in Israel. RhodesianJournal of Agricultural Research 15, 101–104.Burýlova, A.M. (1975). On bloodsucking midges attacking the birds in theforests of Prikamje. Parazitologiya 9, 197–200.
10Diego Santiago-Alarcon and others

Butenko, O.M. (1967). Some data on the feeding of blood-sucking insects(gnats) on birds. Bjulleten Moskovskogo obsiestva spytatelej prirody, otdelbiologiceskij 72, 132–136.Calisher, C. H. (1994). Medically important arboviruses of the UnitedStates and Canada. Clinical Microbiology Reviews 7, 89–116.Chao, A. and Shen, T. J. (2010). Program SPADE (Species Prediction AndDiversity Estimation). http://chao.stat.nthu.edu.tw.Clark, G.W. (1964). Frequency of infection and seasonal variation ofLeucocytozoon berestneffi in the yellow-billed magpie, Pica nuttalli. Journalof Protozoology 11, 481–484.Coggeshall, L. T. (1940). The infection of Anopheles quadrimaculatuswith a monkey malaria parasite, Plasmodium cynomolgi, and with an avianparasite, Plasmodium lophurae. Journal of Parasitology 26, S44–S45.Delécolle, J. C. (1985). Nouvelle contribution à l’étude systématiqueet iconographieque des espèces du genre Culicoides (Diptera: Ceratopogo-nidae) du Nord –Est de la France. Thèse. Université Louis Pasteur deStrasbourg, pp. 1–238.Enayati, A. and Hemingway, J. (2010). Malaria management: past,present, and future. Annual Review of Entomology 55, 569–591.Ewald, P.W. (1994). Evolution of Infectious Disease. Oxford UniversityPress, New York, USA.Feinsinger, P. (2001).Designing Field Studies for Biodiversity Conservation.The Nature Conservancy, Island Press, Washington, USA.Fritz, H. G. (1982). Dissertation –Ökologische und systematische Unter-suchungen an Diptera/Nematocera (Insecta) in Überschwemmungsge-bieten des Nördlichen Oberrheins –Ein Beitrag zur Ökologie großerFlußauen –Fachbereich 10: Biologie der Technischen HochschuleDarmstadt. Dissertation, 296 pp. Information from p. 108.Garvin, M. C. and Greiner, E. C. (2003). Ecology of Culicoides (Diptera:Ceratopogonidae) in Southcentral Florida and experimental Culicoidesvectors of the avian hematozoanHaemoproteus danilewskyiKruse. Journal ofWildlife Diseases 39, 170–178.Gethmann, J., Hoffmann, B., Probst, C., Beer, M., Conraths, F. J.and Mettenleiter, T. C. (2010). A survey of three years with bluetonguedisease serotype 8 in Germany (in German). Tierärztliche Umschau 64,4–12.Greiner, E. C. (1975). Prevalence and potential vectors of Haemoproteus inNebraska mourning doves. Journal of Wildlife Diseases 11, 150–156.Havelka, P. (1976). Limnologische und systematische Studien anCeratopogoniden (Diptera: Nematocera). Beiträge zur Entomologie Berlin26, 211–305.Havelka, P. and Aguilar, M. (1999). Ceratopogonidae. In Checkliste derDipteren Deutschlands: Studia Dipterologica (ed. Schumann, H.,Bährmann, R. and Stark, A.), pp. S33–S38. Suppl. 2. Ampyx-Verlag,Halle (Saale), Germany.Hellgren, O., Waldenström, J. and Bensch, S. (2004). A new PCR assayfor simultaneous studies of Leucocytozoon, Plasmodium, and Haemoproteusfrom avian blood. Journal of Parasitology 90, 797–802.Henderson, P. A. and Seaby, R.M.H. (2002). Species Diversity andRichness v.3.02. PISCES Conservation Ltd., Pennington, UK.Herman, C.M. (1938). Mosquito transmission of avian malaria parasites(Plasmodium circumflexum andP. cathemerium).American Journal ofHygiene27, 345–350.Hörbrand, T. and Geier, M. (2009). Monitoring of Culicoides at ninelocations in southern Germany (2007–2008). Parasitology Research 105,387–392.Institute for Animal Health (IAH), Vector-borne diseases – culicoides.net –UKCulicoides reference laboratory. http://www.bluetonguevirus.org/culicoides.Janovy, J., Jr. (1966). Epidemiology ofPlasmodium hexameriumHuff, 1935,in meadowlarks and starlings of the Cheyenne Bottoms, Barton County,Kansas. Journal of Parasitology 52, 573–578.Jones, K. E., Pater, N. G., Levy, M. A., Storeygard, A., Balk, D.,Gittleman, J. L. and Daszak, P. (2008). Global trends in emerginginfectious diseases. Nature 451, 990–993.Jost, L. (2006). Entropy and diversity. Oikos 113, 363–375.Jost, L. (2010). The relation between evenness and diversity. Diversity 2,207–232.Keeling, M. J. and Rohani, P. (2008). Modeling Infectious Diseases inHumans and Animals. Princeton University Press, Princeton, NJ, USA.Kiel, E., Liebisch, G., Focke, R. and Liebisch, A. (2009). Monitoring ofCulicoides at 20 locations in northwest Germany. Parasitology Research 105,351–357.Kim, K. S., Tsuda, Y., Sasaki, T., Kobayashi, M. and Hirota, Y. (2009).Mosquito blood-meal analysis for avian malaria study in wild birdcommunities: laboratory verification and application to Culex sasai(Diptera: Culicidae) collected in Tokyo, Japan. Parasitology Research 105,1351–1357.
Kitaoka, S., Morii, T. and Kosuge, M. (1965). Field experiments on therepellents to chicken bitingmidges. Japanese Journal of Sanitary Zoology 16,244–248.Klei, T. R. and DeGiusti, D. L. (1975). Seasonal occurrence ofHaemoproteus columbae Kruse and its vector Pseudolynchia canariensisBequaert. Journal of Wildlife Diseases 11, 130–135.Kremer, M. (1965). Contribution a l’étude du genre Culicoides Latreille,particulièrement en France. Editions P. Lechevalier Paris. EncyclopédieEntomologique Série, A 39, 3–299.Križanauskienė, A., Pérez-Tris, J., Palinauskas, V., Hellgren, O.,Bensch, S. and Valkiūnas, G. (2010). Molecular phylogenetic andmorphological analysis of haemosporidian parasites (Haemosporida)in a naturally infected European songbird, the Blackcap Sylvia atricapilla,with description of Haemoproteus pallidulus sp. nov. Parasitology 137,217–227.Lassen, S. B., Nielsen, S. A., Skovgård, H. and Kristensen, M. (2011).Molecular identification of blood meals from biting midges (Diptera:Ceratopogonidae: Culicoides Latreille) in Denmark. Parasitology Research108, 823–829.Lefèvre, T., Gouagna, L.-C., Dabiré, K. R., Elguero, E., Fontenille, D.,Renaud, F., Costantini, C. and Thomas, F. (2009). Beyond nature andnurture: phenotypic plasticity in blood-feeding behavior of Anophelesgambiae s.s. when humans are not readily accessible. American Journal ofTropical Medicine and Hygiene 81, 1023–1029.Mayne, B. (1928). An anopheline mosquito as host for the parasites of birdmalaria. Indian Journal of Medical Research 16, 557–558.McGhee, R. B. (1951). The adaptation of the avian malaria parasitePlasmodium lophurae to a continuous existence in infant mice. Journal ofInfectious Diseases 88, 86–97.McGhee, R. B. (1957). Comparative susceptibility of various erythrocytesto four species of avian Plasmodia. Journal of Infectious Diseases 100, 92–96.Mehlhorn, H., Walldorf, V., Klimpel, S., Jahn, B., Jaeger, F.,Eschweiler, J., Hoffmann, B. and Beer, M. (2007). First occurrence ofCulicoides obsoletus-transmitted bluetongue virus epidemic in centralEurope. Parasitology Research 101, 219–228.Mehlhorn, H.,Walldorf, V., Klimpel, S., Schmahl, G. (2008). Outbreakof bluetongue disease (BTD) in Germany and the danger for Europe.Parasitology Research 103(Suppl.), S79–S86.Mehlhorn, H., Walldorf, V., Klimpel, S., Schmahl, G., Al-Quraishy,S., Walldorf, U., Mehlhorn, B. and Bätza, H.-J. (2009). Entomologicalsurvey on vectors of bluetongue virus in Northrhine-Westfalia (Germany)during 2007 and 2008. Parasitology Research 105, 321–329.Mellor, P. S., Boorman, J. and Baylis, M. (2000). Culicoides bitingmidges: their role as arbovirus vectors. Annual Review of Entomology 45,307–340.Minár, J. (1965). Tabanidae, Ceratopogonidae, Simuliidae from the LipnoWater –Reservoir. Československá Parazitologie 12, 197–206.Nielsen, O. B. (1964). Studies on the Danish biting midges, CulicoidesLatreille (Dipt., Ceratopogonidae). Entomlogiske Meddelelser 32, 261–276.Nielsen, O. B. (1971). Some observations on biting midges (Diptera:Ceratopogonidae) attacking grazing cattle in Denmark. EntomologicaScandinavica 2, 94–98.Országh, I. (1976). Die Gattung Culicoides Latr. 1809 (Diptera,Ceratopogonidae) in Slowakei. Acta Facultatis Rerum NaturaliumUniversitatis Comenianae –Zoologia 21, 1–88.Paclt, J., Callot, J. and Kremer, M. (1970). Cératopogonidés piqueurshabitant les tourbières et les biotopes non-tourbeux de la partie supérieured’orava (Diptera Nematocera). Biológia (Bratislava) 25, 751–759.Pérez-Tris, J. and Bensch, S. (2005a). Diagnosing genetically diverseavian malarial infections using mixed-sequence analysis and TA-cloning.Parasitology 131, 15–23.Pérez-Tris, J. and Bensch, S. (2005b). Dispersal increases localtransmission of avian malaria parasites. Ecology Letters 8, 838–845.Pérez-Tris, J. and Tellería, J. L. (2002). Migratory and sedentaryblackcaps in sympatric non-breeding grounds: implications for theevolution of avian migration. Journal of Animal Ecology 71, 211–224.Pérez-Tris, J., Bensch, S., Carbonell, R., Helbig, A. J. andTellería, J. L. (2004). Historical diversification of migration patterns in apasserine bird. Evolution 58, 1819–1832.Pérez-Tris, J., Hellgren, O., Križanauskienė, A., Waldenström, J.,Secondi, J., Bonneaud, C., Fjeldså, J., Hasselquist, D. and Bensch, S.(2007). Within-host speciation of malaria parasites. PLoS ONE 2, e235.Quan, M., Van Vuuren, M., Howel, P. G., Groenewald, D. andGuthrie, A. J. (2008). Molecular epidemiology of the African horse sicknessvirus S10 gene. Journal of General Virology 89, 1159–1168.Ratnasingham, S. and Hebert, P. D. N. (2007). BOLD: the barcodeof life data system (www.barcodinglife.org). Molecular Ecology Notes 7,355–364.
11Culicoides blood meal analysis in urban forest

Reiczigel, J. (2003). Confidence intervals for the binomial parameter: somenew considerations. Statistics in Medicine 22, 611–621.Rolshausen, G., Segelbacher, G., Hobson, K. A. and Schaefer, H.M.(2009). Contemporary evolution of reproductive isolation and pheno-typic divergence in sympatry along a migratory divide. Current Biology19, 2097–2101.Rolshausen, G., Hobson, K. A. and Schaefer, H.M. (2010). Springarrival along a migratory divide of sympatric blackcaps (Sylvia atricapilla).Oecologia 162, 175–183.Rózsa, L., Reiczigel, J. andMajoros, G. (2000). Quantifying parasites insamples of hosts. Journal of Parasitology 86, 228–232.Santiago-Alarcon, D., Bloch, R., Rolshausen, G., Schaefer, H.M. andSegelbacher, G. (2011). Prevalence, diversity, and interaction patterns ofavian haemosporidians in a four-year study of blackcaps in a migratorydivide. Parasitology 138, 824–835.Santiago-Alarcon, D., Palinauskas, V. and Schaefer, H.M.(2012a). Diptera vectors of avian haemosporidian parasites:untangling parasite life cycles and their taxonomy. Biological Reviews 87,928–964.Santiago-Alarcon, D., Havelka, P., Schaefer, H.M. andSegelbacher, G. (2012b). Blood meal analysis reveals avian Plasmodiuminfections and broad host preferences of Culicoides (Diptera:Ceratopogonidae) vectors. PLoS ONE 7, e31098.Santiago-Alarcon, D., Mettler, R., Segelbacher, G. andSchaefer, H.M. (2013). Haemosporidian parasitism in the blackcap(Sylvia atricapilla) in relation to spring arrival and body condition.Journal of Avian Biology (in press) doi: 10.1111/j.1600-048X.2013.00181.x.Shirihai, H., Gargallo, G. and Helbig, A. J. (2001). Sylvia Warblers.Princeton University Press, Princeton, NJ, USA.Sol, D., Jovani, R. and Torres, J. (2000). Geographical variationin blood parasites in feral pigeons: the role of vectors. Ecography 23,307–314.
Solow, A. R. (1993). A simple test for change in community structure.Journal of Animal Ecology 62, 191–193.Super, P. E. and van Riper, C., III (1995). A comparison of avianhematozoan epizootiology in two California coastal scrub communities.Journal of Wildlife Diseases 31, 447–461.Szadziewski, R. (1984). On synonymy and morphology of someCulicoides species (Diptera, Ceratopogonidae). Polskie Pismo Entomolgiczne53, 559–556.Tellería, J. L. and Pérez-Tris, J. (2003). Seasonal distribution of amigratory bird: effects of local and regional resource tracking. Journal ofBiogeography 30, 1583–1591.Valkiūnas, G. (2005). Avian Malaria Parasites and other Haemosporidia.CRC Press, Boca Raton, FL, USA.Valkiūnas, G. and Iezhova, T. A. (2004a). Detrimental effects ofHaemoproteus infections on the survival of biting midge Culicoidesimpunctatus (Diptera: Ceratopogonidae). Journal of Parasitology 90,194–196.Valkiūnas, G. and Iezhova, T. A. (2004b). The transmission ofHaemoproteus belopolskyi (Haemosporida: Haemoproteidae) of blackcap byCulicoides impunctatus (Diptera: Ceratopogonidae). Journal of Parasitology90, 196–198.Valkiūnas, G., Liutkevičius, G. and Iezhova, T. A. (2002). Completedevelopment of three species of Haemoproteus (Haemosporida,Haemoproteidae) in the biting midge Culicoides impunctatus (Diptera,Ceratopogonidae). Journal of Parasitology 88, 864–868.Valkiūnas, G., Žičkus, T., Shapoval, A. P. and Iezhova, T. A. (2006).Effect of Haemoproteus belopolskyi (Haemosporida: Haemoproteidae) onbody mass of the blackcap Sylvia atricapilla. Journal of Parasitology 92,1123–1125.Vorsprach, B., Meiser, C. K., Werner, D., Balczun, C. andSchaub, G. A. (2009). Monitoring of Ceratopogonidae in southwestGermany. Parasitology Research 105, 337–344.
12Diego Santiago-Alarcon and others
Related Documents











