Journal of Alzheimer’s Disease 45 (2015) 127–138 DOI 10.3233/JAD-141008 IOS Press 127 Type 2 Diabetes Aggravates Alzheimer’s Disease-Associated Vascular Alterations of the Aorta in Mice Cristina M. Sena a,b,∗ , Ana M. Pereira a,b , Cristina Carvalho d , Rosa Fernandes b , Raquel M. Seic ¸a a,b , Catarina R. Oliveira c,d and Paula I. Moreira a,d,∗ a Institute of Physiology, Faculty of Medicine, University of Coimbra, Coimbra, Portgual b IBILI - Institute of Biomedical Imaging and Life Sciences, Faculty of Medicine, University of Coimbra, Coimbra, Portugal c Institute of Biochemistry, Faculty of Medicine, University of Coimbra, Coimbra, Portugal d CNC - Center for Neuroscience and Cell Biology, University of Coimbra, Coimbra, Portugal Accepted 12 November 2014 Abstract. Vascular risk factors are associated with a higher incidence of dementia. In fact, diabetes mellitus is considered a main risk factor for Alzheimer’s disease (AD) and both diseases are characterized by vascular dysfunction. However, the underlying mechanisms remain largely unknown. Here, the effects of high-sucrose-induced type 2 diabetes (T2D) in the aorta of wild type (WT) and triple-transgenic AD (3xTg-AD) mice were investigated. 3xTg-AD mice showed a significant decrease in body weight and an increase in postprandial glycemia, glycated hemoglobin (HbA1c), and vascular nitrotyrosine, superoxide anion (O 2 • – ), receptor for the advanced glycation end products (RAGE) protein, and monocyte chemoattractant protein-1 (MCP-1) levels when compared to WT mice. High-sucrose intake caused a significant increase in body weight, postprandial glycemia, HbA1c, triglycerides, plasma vascular cell adhesion molecule 1 (VCAM-1), and vascular nitrotyrosine, O 2 • – , RAGE, and MCP-1 levels in both WT and 3xTg-AD mice when compared to the respective control group. Also, a significant decrease in nitric oxide- dependent vasorelaxation was observed in 3xTg-AD and sucrose-treated WT mice. In conclusion, AD and T2D promote similar vascular dysfunction of the aorta, this effect being associated with elevated oxidative and nitrosative stress and inflammation. Also, AD-associated vascular alterations are potentiated by T2D. These findings support the idea that metabolic alterations predispose to the onset and progression of dementia. Keywords: Alzheimer’s disease, aorta, endothelium, inflammation, oxidative stress, type 2 diabetes INTRODUCTION Alzheimer’s disease (AD) is a complex neurode- generative disorder and is the most common form of dementia affecting millions of people worldwide. ∗ Correspondence to: Cristina M. Sena, Institute of Physiology, Faculty of Medicine, University of Coimbra, 3000-548 Coimbra, Portugal. E-mail: [email protected] and Paula I. Moreira, Institute of Physiology, Faculty of Medicine, University of Coimbra & Center for Neuroscience and Cell Biology, University of Coimbra, 3004-504 Coimbra, Portugal. E-mail: [email protected]/[email protected]. Several vascular risk factors (e.g., diabetes, hyper- lipidemia, obesity, atherosclerosis, hypertension, and metabolic syndrome) have been found to be associ- ated with AD [1, 2]. It has been consistently shown that cerebral vascular insufficiency plays an impor- tant role in neurodegenerative diseases ranging from vascular cognitive impairment to AD [3]. Also, heart disease and the resultant vascular insufficiency have the potential to impair function in other organs, includ- ing the brain [4]. In fact, reduced cerebral blood flow due to heart disease of any kind worsens the vascular ISSN 1387-2877/15/$35.00 © 2015 – IOS Press and the authors. All rights reserved

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Alzheimer’s Disease 45 (2015) 127–138DOI 10.3233/JAD-141008IOS Press
127
Type 2 Diabetes Aggravates Alzheimer’sDisease-Associated Vascular Alterationsof the Aorta in Mice
Cristina M. Senaa,b,∗, Ana M. Pereiraa,b, Cristina Carvalhod, Rosa Fernandesb, Raquel M. Seicaa,b,Catarina R. Oliveirac,d and Paula I. Moreiraa,d,∗aInstitute of Physiology, Faculty of Medicine, University of Coimbra, Coimbra, PortgualbIBILI - Institute of Biomedical Imaging and Life Sciences, Faculty of Medicine, University of Coimbra, Coimbra,PortugalcInstitute of Biochemistry, Faculty of Medicine, University of Coimbra, Coimbra, PortugaldCNC - Center for Neuroscience and Cell Biology, University of Coimbra, Coimbra, Portugal
Accepted 12 November 2014
Abstract. Vascular risk factors are associated with a higher incidence of dementia. In fact, diabetes mellitus is considered a mainrisk factor for Alzheimer’s disease (AD) and both diseases are characterized by vascular dysfunction. However, the underlyingmechanisms remain largely unknown. Here, the effects of high-sucrose-induced type 2 diabetes (T2D) in the aorta of wild type(WT) and triple-transgenic AD (3xTg-AD) mice were investigated. 3xTg-AD mice showed a significant decrease in body weightand an increase in postprandial glycemia, glycated hemoglobin (HbA1c), and vascular nitrotyrosine, superoxide anion (O2
•–),receptor for the advanced glycation end products (RAGE) protein, and monocyte chemoattractant protein-1 (MCP-1) levelswhen compared to WT mice. High-sucrose intake caused a significant increase in body weight, postprandial glycemia, HbA1c,triglycerides, plasma vascular cell adhesion molecule 1 (VCAM-1), and vascular nitrotyrosine, O2
•–, RAGE, and MCP-1 levelsin both WT and 3xTg-AD mice when compared to the respective control group. Also, a significant decrease in nitric oxide-dependent vasorelaxation was observed in 3xTg-AD and sucrose-treated WT mice. In conclusion, AD and T2D promote similarvascular dysfunction of the aorta, this effect being associated with elevated oxidative and nitrosative stress and inflammation.Also, AD-associated vascular alterations are potentiated by T2D. These findings support the idea that metabolic alterationspredispose to the onset and progression of dementia.
Keywords: Alzheimer’s disease, aorta, endothelium, inflammation, oxidative stress, type 2 diabetes
INTRODUCTION
Alzheimer’s disease (AD) is a complex neurode-generative disorder and is the most common formof dementia affecting millions of people worldwide.
∗Correspondence to: Cristina M. Sena, Institute of Physiology,Faculty of Medicine, University of Coimbra, 3000-548 Coimbra,Portugal. E-mail: [email protected] and Paula I. Moreira, Institute ofPhysiology, Faculty of Medicine, University of Coimbra & Centerfor Neuroscience and Cell Biology, University of Coimbra, 3004-504Coimbra, Portugal. E-mail: [email protected]/[email protected].
Several vascular risk factors (e.g., diabetes, hyper-lipidemia, obesity, atherosclerosis, hypertension, andmetabolic syndrome) have been found to be associ-ated with AD [1, 2]. It has been consistently shownthat cerebral vascular insufficiency plays an impor-tant role in neurodegenerative diseases ranging fromvascular cognitive impairment to AD [3]. Also, heartdisease and the resultant vascular insufficiency havethe potential to impair function in other organs, includ-ing the brain [4]. In fact, reduced cerebral blood flowdue to heart disease of any kind worsens the vascular
ISSN 1387-2877/15/$35.00 © 2015 – IOS Press and the authors. All rights reserved

128 C.M. Sena et al. / Vascular Risk Factors and Alzheimer’s Disease
homeostasis of the brain, and magnifies any cognitiveproblems caused by the buildup of tau and amyloid-�(A�) proteins, key hallmarks of AD [5].
The endothelium is a common target for vascularrisk factors, and functional impairment of the endothe-lium in response to damage occurs long before thedevelopment of overt disease [6, 7]. Vascular endothe-lium is a major target of oxidative stress therebyplaying a major role in the pathophysiology of sev-eral diseases including AD and type 2 diabetes (T2D)[8]. A growing body of evidence supports a vascular-neuronal axis in AD involving vascular mechanismsin the development and/or progression of the disease[9, 10]. Thus, the vascular endothelium is a key tar-get in AD, particularly important in conditions ofhyperglycemia and metabolic syndrome, where thelevels of oxidative stress are substantially increased[8]. However, the mechanisms underlying T2D and/orAD-associated vascular and endothelial anomalies arepoorly explored. Thus, we aimed to investigate andcompare the effects of high-sucrose-induced T2D andAD in aortic vascular wall and endothelial function.For this purpose, four groups of experimental ani-mals were used: 1) wild type (WT) control mice;2) sucrose-treated WT mice; 3) triple transgenic AD(3xTg-AD) mice, and 4) sucrose-treated 3xTg-ADmice. Mice were subjected to a sucrose solution giventhat excessive consumption of sugars plays a keyrole in the epidemic of obesity and T2D, two riskfactors for AD [11, 12]. Several parameters were eval-uated in aortas isolated from the above-mentionedmice: vascular constriction and relaxation, histolog-ical alterations, oxidative and nitrosative stress andinflammation markers, and the levels of A� oligomers,considered the most neurotoxic species in AD.
MATERIALS AND METHODS
Drugs
Phenylephrine, acetylcholine, and NG-nitro-L-arginine-methyl ester (L-NAME) were obtained fromSigma (St. Louis, MO, USA). Anti-RAGE and anti-nitrotyrosine were obtained from Abcam (Cambridge,UK) and Upstate Biotechnology (Lake Placid, NY,USA), respectively. Anti-MCP-1 and anti-�-actin wereobtained from Chemicon International Inc. (Temec-ula, CA, USA). Anti-amyloid beta clone 6E10 wasobtained from Signet Laboratories (Princeton, NJ,USA). Dihydroethidium (DHE) was obtained fromInvitrogen (Barcelona, Spain). All other chemicals andreagents used in the study were of high grade.
Animals
Male WT and 3xTg-AD mice (4 months old)were housed in our animal facility (Faculty ofMedicine/Center for Neuroscience and Cell Biol-ogy, University of Coimbra). The 3xTg-AD mice arehomozygous for three mutant alleles: presenilin (PS) 1(M146V), amyloid-� protein precursor (A�PP) (Swe),and tau (P301L). WT and 3xTg-AD mice were ran-domly divided into two groups: 1) control group and2) sucrose-treated animals with free access to 20%sucrose solution during 7 months. Mice were main-tained under controlled light (12-h day/night cycle)and humidity with free access (except in the fastingperiod) to water (WT and 3xTg-AD mice at basalconditions) or 20% sucrose solution (sucrose-treatedWT and sucrose-treated 3xTg-AD mice) and pow-dered rodent chow (URF1; Charles River). Adheringto procedures approved by the Federation of Labo-ratory Animal Science Associations (FELASA), theanimals (11 months old) were sacrificed at the endof the treatment period by cervical displacement anddecapitation.
Determination of metabolic parameters
The levels of blood glucose, glycated hemoglobin(HbA1c), triglycerides, cholesterol, and VCAM-1 weredetermined by standard procedures.
Isometric tension studies
Aorta were rapidly excised and freed of connec-tive tissue. The aorta was divided into two segments(4 mm width, Fig. 1). Ring segments were mountedbetween stainless steel triangles into individual organchambers filled with oxygenated (95% O2, 5% CO2)Krebs-Henseleit buffer (37◦C, pH 7.4) (composition inmmol l−1: NaCl 119; KCl 4.7; CaCl2 1.6; MgSO4 1.2;NaHCO3 25; KH2PO4 1.2; glucose 11.0). Aortic ringswere subject to a resting tension of 1 g. After equili-bration for 90 min, all vessels were preconstricted withphenylephrine. Ligand stimulated receptor-mediatednitric oxide (NO) bioavailability was assessed bya dose-dependent relaxation to acetylcholine (ACh,10−9 to 10−2 M), whereas sodium nitroprusside (SNP,10−9 to 10−2 M) was used as an endothelium-independent agonist. Relaxation responses to ACh andSNP were expressed as percentage of relaxation froma submaximal phenylephrine-induced constriction andconcentration-response curves were obtained as previ-ously described [13, 14].

C.M. Sena et al. / Vascular Risk Factors and Alzheimer’s Disease 129
Fig. 1. Mice aortic preparation. Mouse aorta was gently removed from the animal after sacrifice. Fat and connective tissues were cleaned off andcut transversely into rings of 3- to 4-mm in length. Extreme care was taken to not damage the endothelium in the rings. The rings were mountedvertically between two stainless steel wire hooks tied with thread and suspended in 10 ml organ baths containing Krebs-Henseleit buffer (pH7.4). The pH of the buffer was maintained with 95% O2 and 5% CO2 at 37◦C. Aortic rings were equilibrated for 90 min with a resting force of1 g before the start of the experiment.
Detection of superoxide anion levels
Unfixed frozen, 30 �m-thick sections of proximalaorta were incubated with DHE (2 × 10−6 mol l−1)in PBS for 30 min at 37◦C in a humidified cham-ber protected from light. DHE is oxidized on reactionwith O2
• – to ethidium bromide (EtBr), which binds toDNA in the nucleus and fluoresces red [15]. Polyethy-lene glycol-superoxide dismutase (PEG-SOD, 500 Uml−1) abolished EtBr fluorescence, confirming speci-ficity of the fluorescent signal for O2
• – (data notshown). For EtBr detection, images were obtained witha fluorescence microscope (Leica DMIRE200, Wet-zlar, Germany) and fluorescence was detected with a568 nm filter. Tissues obtained from the four groupsof animals were processed and imaged in parallelwith identical settings. Microscope and camera settingswere kept constant for all preparations. Fluorescencewas quantified using ImageJ (1.40 g, NIH).
Histology
Mice were anesthetized and perfused transcardia-cally with ice-cold saline for 2 min. The arterialsystem was perfusion-fixed for 15 min with phosphate-buffered 4% paraformaldehyde (pH 7.4). The aortaswere dissected and further fixed overnight in 4%paraformaldehyde for subsequent immunohistochem-
ical analysis. Tissue sections (4 mm) were stainedwith Hematoxylin-Eosin (H&E) or with Periodic Acid-Schiff (PAS) in a standard manner.
Assessment of aortic immunofluorescence
Sections (6 �m) of abdominal aorta were washedwith PBS and fixed in ice cold acetone, for 10 min.Sections were then permeabilized for 10 min in 1%Triton X-100 in PBS, pH 7.4, and blocked with10% goat serum for 30 min. Primary antibodies werediluted in PBS containing 0.02% BSA (PBS/BSA).The primary antibodies were added and the sectionswere incubated overnight at 4◦C. After incubation,the sections were extensively washed with PBS/BSAsolution. After, sections were incubated with the sec-ondary antibodies, diluted in PBS/BSA for 1 h. Thecoverslips were washed before mounting with Glyc-ergel Dako mounting medium (Dako, Carpinteria, CA,USA). Immunostained aortic sections were visualizedwith a Leica DMIRE200 fluorescence microscope.Immunostained aortic sections were counterstainedwith DAPI and examined, photographed and quantifiedas described above for DHE fluorescence.
Western blot analysis
Segments of endothelium-intact aortas were washedwith cold PBS and chilled in buffer containing

130 C.M. Sena et al. / Vascular Risk Factors and Alzheimer’s Disease
(mmol l−1): Tris–HCl 50, NaCl 150, ethylenedi-amine tetraacetic acid (EDTA) 1 and ethylene glycoltetraacetic acid (EGTA) 0.1 with NP-40 0.1%, SDS0.1% and deoxycholate 0.5%. Phenylmethylsulfonylfluoride (1 mmol l−1), aprotonin (10 �g ml−1), leu-peptin (10 �g ml−1), and pepstatin (10 �g ml−1) allfrom Sigma Chemicals (St. Louis, MO, USA) wereadded as protease inhibitors. Tissues were homoge-nized in a standard manner, followed by centrifugationat 14,000× g for 20 min at 4◦C. The supernatantswere collected and total protein concentration wasdetermined using a Bio-Rad protein assay kit. Sam-ples containing 10 �g of protein were loaded on to12% sodium dodecyl sulfate-polyacrylamide gel elec-trophoresis (SDS-PAGE) gel, run and electroblottedonto polyvinylidene difluoride (PVDF) membrane.Prestained molecular weight marker proteins wereused as standards for the SDS-PAGE. A ponceaustaining was performed to verify the quality of thetransfer and to ensure equal protein loading. Blotswere blocked in 5% skimmed nonfat milk in PBSfor 1 h, treated overnight with primary antibodies andthen incubated with alkaline phosphatase secondaryantibodies for 1 h. Immunoblots were developed withan ECF western blotting detection system (AmershamBiosciences).
Statistical analysis
All data were analyzed by standard computerprograms (GraphPad Prism PC Software version 3.0,ANOVA) and are expressed as mean ± SE/SEM.Significant differences were evaluated using eitherthe t-test or ANOVA. p < 0.05 was consideredsignificant. Dose-response curves were fitted bynonlinear regression with simplex algorithm.Relaxation responses were given as the percent-age of phenylephrine-mediated preconstriction.
Comparisons of dose–response curves were evaluatedby two-way ANOVA for repeated measures.
RESULTS
Characterization of experimental animals
Compared to WT mice, 3xTg-AD animals pre-sented a significant decrease in body weight and asignificant increase in HbA1c and postprandial glucoselevels. Sucrose intake caused a significant increase inbody weight, blood glucose, HbA1c, triglycerides, andVCAM-1 levels in both WT and 3xTg-AD mice whencompared to the respective control groups at basal con-ditions (Table 1). The metabolic alterations induced bysucrose intake are typical of type 2 diabetes.
T2D and AD cause endothelial dysfunction
Compared to age-matched WT mice, endothelium-mediated vascular relaxation of phenylephrine-precontracted aortic rings in response to ACh wasimpaired in 3xTg-AD mice (Fig. 2A). Noteworthy, aor-tic rings from sucrose-treated WT mice presented arelaxation in response to ACh similar to that observedin 3xTg-AD mice (Fig. 2A, no significant differ-ences were observed in maximal relaxation), although3xTg-AD mice show impaired endothelial-dependentvasodilation at lower ACh concentrations (the impair-ment was observed in the presence of 0.1 �M AChin 3xTg-AD mice and only at 10 �M ACh in WT+Smice). Strikingly, sucrose did not aggravate endothelialdysfunction in 3xTg-AD mice (Fig. 2A, no signifi-cant differences were observed between 3xTg-AD and3xTg-AD+S at all ACh concentrations). Preincuba-tion of the arterial rings with the nitric oxide synthase(NOS) inhibitor L-NAME and the cyclooxygenaseinhibitor indomethacin almost completely abolished
Table 1Mice characterization
WT WT+S 3xTg-AD 3xTg-AD+S
Body weight (g) 35.20 ± 1.09 42.86 ± 1.27∗∗∗ 31.14 ± 0.75∗∗ 35.21 ± 0.84φ
Postprandial glycemia (mg/dl) 100.7 ± 2.19 116.6 ± 3.72∗ 121.3 ± 8.53∗ 136.9 ± 7.6∗∗∗Insulin levels (�g/l) 1.42 ± 0.25 4.25 ± 0.86∗∗ 0.88 ± 0.17 0.76 ± 0.07HbA1c (%) (mmol/mol) 3.36 ± 0.07 3.77 ± 0.08∗∗ 3.99 ± 0.11∗∗∗ 4.67 ± 0.18∗∗∗φφ
≈ 13 ≈ 18 ≈ 20 ≈ 28Triglyceride levels (mg/dl) 159.8 ± 19.16 277.4 ± 30.60∗∗ 168.2 ± 17.80 280.4 ± 23.05∗∗φφ
Cholesterol levels (mg/dl) 161.3 ± 2.81 160.1 ± 1.91 160.6 ± 2.48 163.7 ± 2.85VCAM levels (ng/ml) 554.7 ± 24.23 733.2 ± 45.40∗∗ 626.7 ± 27.38 817.5 ± 54.53φφ
Data shown represent mean ± SEM from 10–12 animals. Statistical significance: ∗p < 0.05; ∗∗p < 0.01; ∗∗∗p < 0.001 when compared with WTcontrol animals; φp < 0.05; φφp < 0.01 when compared with 3xTg-AD control animals.
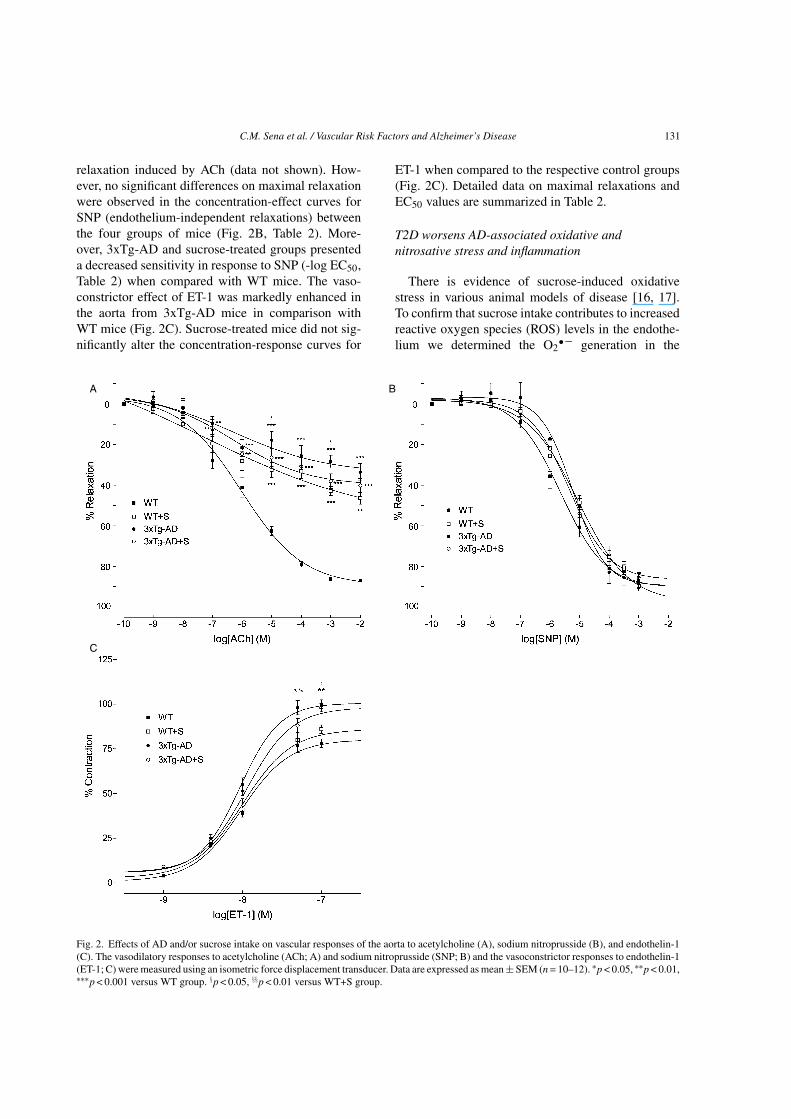
C.M. Sena et al. / Vascular Risk Factors and Alzheimer’s Disease 131
relaxation induced by ACh (data not shown). How-ever, no significant differences on maximal relaxationwere observed in the concentration-effect curves forSNP (endothelium-independent relaxations) betweenthe four groups of mice (Fig. 2B, Table 2). More-over, 3xTg-AD and sucrose-treated groups presenteda decreased sensitivity in response to SNP (-log EC50,Table 2) when compared with WT mice. The vaso-constrictor effect of ET-1 was markedly enhanced inthe aorta from 3xTg-AD mice in comparison withWT mice (Fig. 2C). Sucrose-treated mice did not sig-nificantly alter the concentration-response curves for
ET-1 when compared to the respective control groups(Fig. 2C). Detailed data on maximal relaxations andEC50 values are summarized in Table 2.
T2D worsens AD-associated oxidative andnitrosative stress and inflammation
There is evidence of sucrose-induced oxidativestress in various animal models of disease [16, 17].To confirm that sucrose intake contributes to increasedreactive oxygen species (ROS) levels in the endothe-lium we determined the O2
• – generation in the
Fig. 2. Effects of AD and/or sucrose intake on vascular responses of the aorta to acetylcholine (A), sodium nitroprusside (B), and endothelin-1(C). The vasodilatory responses to acetylcholine (ACh; A) and sodium nitroprusside (SNP; B) and the vasoconstrictor responses to endothelin-1(ET-1; C) were measured using an isometric force displacement transducer. Data are expressed as mean ± SEM (n = 10–12). ∗p < 0.05, ∗∗p < 0.01,∗∗∗p < 0.001 versus WT group. §p < 0.05, §§p < 0.01 versus WT+S group.

132 C.M. Sena et al. / Vascular Risk Factors and Alzheimer’s Disease
Table 2Maximal relaxations and EC50 values of the aortic rings
WT WT+S 3xTg-AD 3xTg-AD+S
ACh
% Max. relaxation 87.01 ± 2.53 46.36 ± 2.96∗∗∗ 33.60 ± 4.2∗∗∗ 40.17 ± 3.54∗∗∗-logEC50 6.02 ± 0.15 7.27 ± 0.08∗ 6.32 ± 0.23 6.3 ± 0.4SNP
% Max. relaxation 88.2 ± 2.86 89.57 ± 2.56 86.9 ± 3.5 85.57 ± 2.56-logEC50 5.64 ± 0.09 5.05 ± 0.10∗∗∗ 5.22 ± 0.08∗∗ 5.34 ± 0.06∗
ET − 1% Max. contraction 78 ± 2.4 86 ± 3.2 100 ± 3.7∗ 98 ± 3.4∗∗-logEC50 8.02 ± 0.05 7.99 ± 0.06 8.03 ± 0.03 7.95 ± 0.04
Data shown represent mean ± SEM from 10–12 animals. Statistical significance: ∗p < 0.05; ∗∗p < 0.01;∗∗∗p < 0.001 when compared with WT control animals.
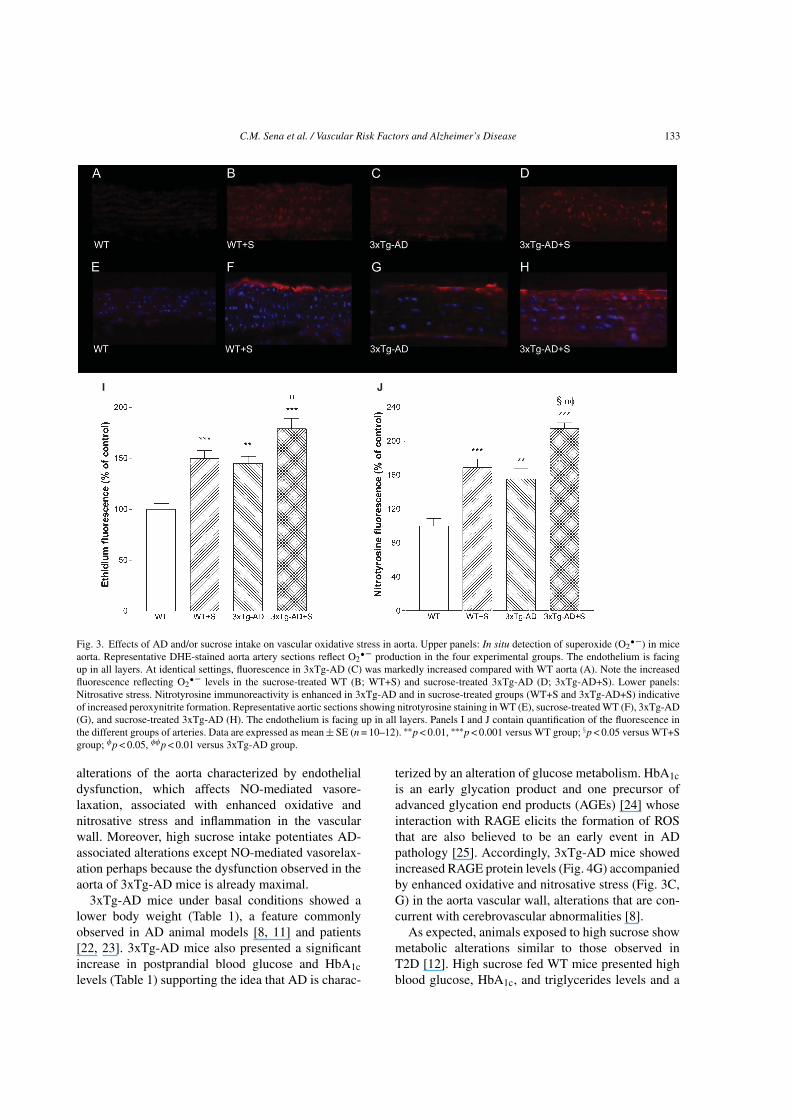
vasculature by immunofluorescence using the fluores-cent dye DHE. As expected, sucrose induced a 1.5-foldincrease in O2
• – production in WT aorta (p < 0.001;Fig. 3B, I). Similarly, the fluorescence of DHE wassignificantly higher in the aorta of 3xTg-AD mice com-pared to WT mice (p < 0.01; Fig. 3C, I). Additionally,sucrose significantly increased DHE fluorescence in3xTg-AD mice (p < 0.05; Fig. 3D, I).
Accordingly, we sought to investigate whetherthe enhanced O2
• – production in aortic tissue ofsucrose-treated mice was associated with peroxynitriteformation and the nitration of tyrosine residues. 3xTg-AD mice had increased immunoreactive nitrotyrosinelevels in their aortas (p < 0.01; Fig. 3G; J), suggest-ing elevated peroxynitrite-mediated protein oxidation.Sucrose significantly increased nitrotyrosine stainingin both WT and 3xTg-AD mice (Fig. 3F, H, and J) indi-cating an increment in oxidative stress in the vascularwall.
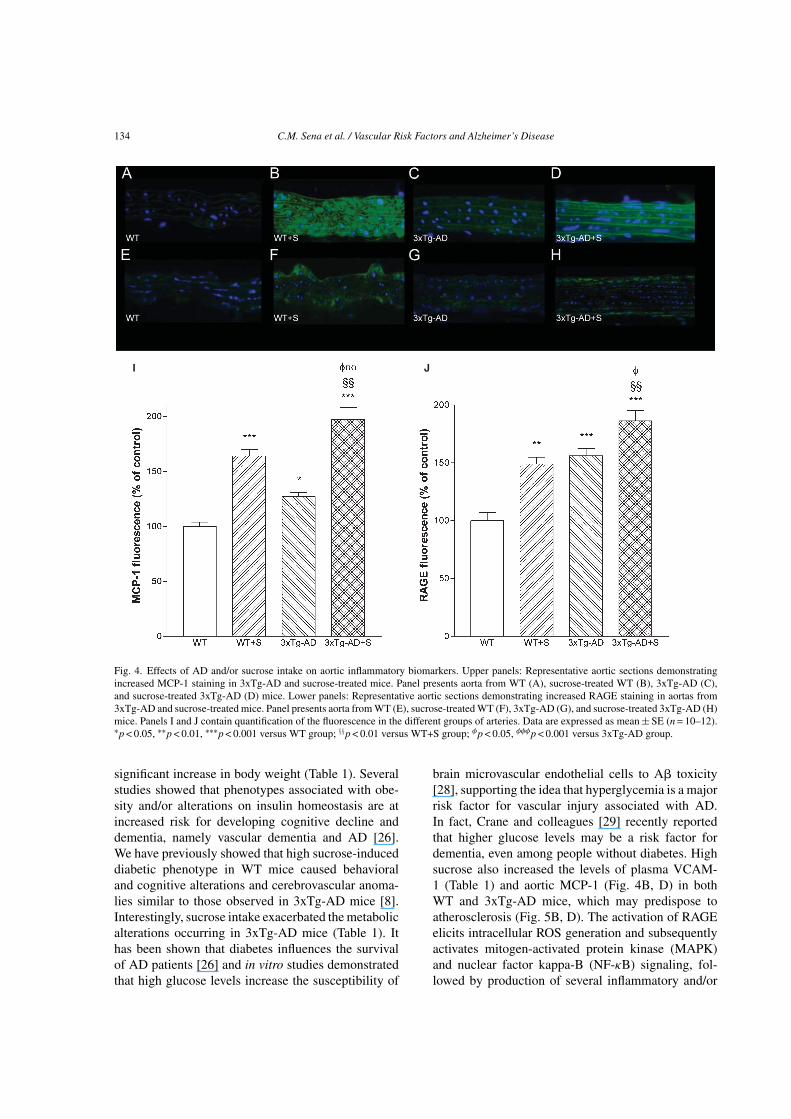
To investigate the underlying mechanism ofendothelial dysfunction, we examined an indicator ofinflammation, the aortic MCP-1 considered one of theearliest molecular markers of vascular inflammationin atherogenesis [18]. MCP-1 levels were significantlyincreased (27%) in the aortas of 3xTg-AD mice com-pared to WT mice (Fig. 4C, I). Additionally, sucroseinduced a 64% increment in MCP-1 levels in WT mice(Fig. 4B, I). Sucrose intake also increased (70%) MCP-1 levels in 3xTg-AD aorta when compared to 3xTg-ADat basal conditions (Fig. 4D, I).
An additional oxidative stress marker that hasbeen implicated in the complications of diabetesand AD is RAGE. Activation of RAGE causes aself-perpetuating, pro-oxidant, pro-inflammatory, andpro-adhesive milieu, which drives tissue damage [19].RAGE levels in aortas of 3xTg-AD mice were signif-icantly increased (56%) when compared to WT mice(Fig. 4G, J). Aorta from sucrose-treated WT and 3xTg-
AD mice showed a significant increase (49% and 30%,respectively) in immunoreactive RAGE levels in thevascular wall when compared to WT and 3xTg-ADcontrol groups (Fig. 4F, H, and J).
T2D worsens AD-associated aorta wallsthickening
For each group of mice, we have also performedH&E and PAS staining. The different groups of ani-mals presented no major changes in aorta morphology(data not shown). Accumulation of PAS positive mate-rial has been described in diabetic arteries independentof the presence of atherosclerotic plaques [20]. Anincrement in PAS positive material in the intima ofsucrose-treated WT and 3xTg-AD mice (Fig. 5B and5D, respectively) was observed.
T2D does not affect Aβ oligomers levels
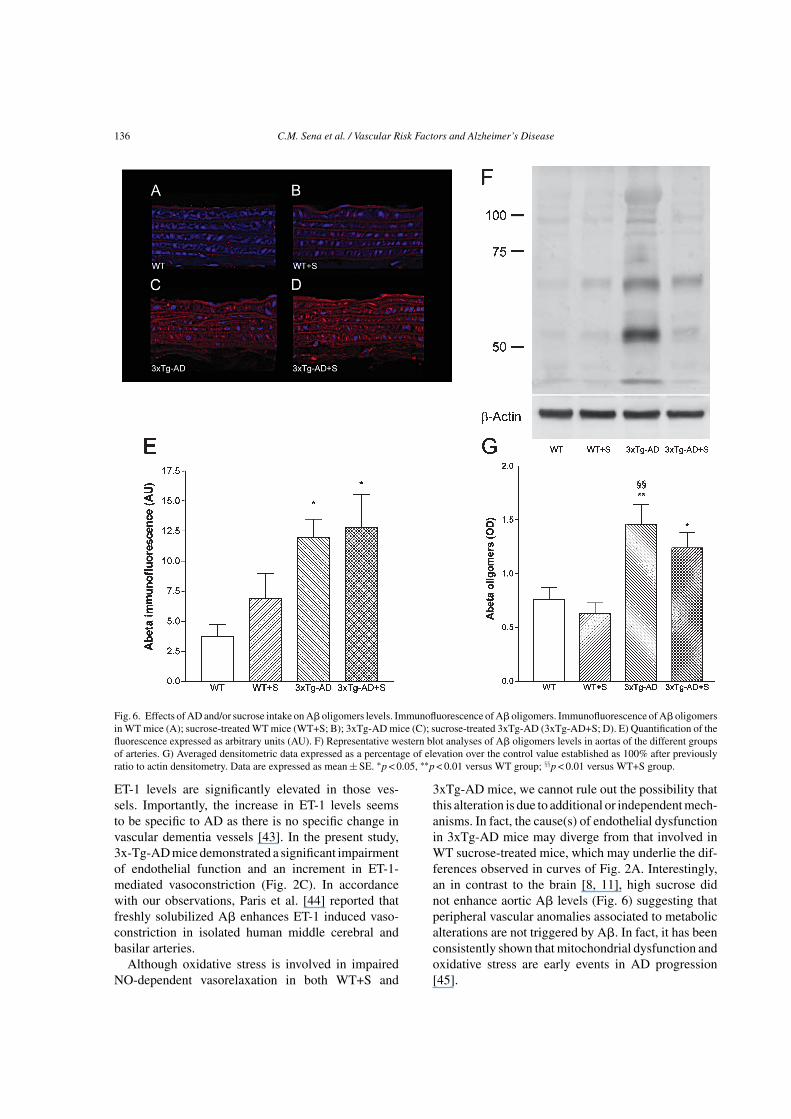
A� oligomers are considered the most neurotoxicspecies in AD brains [21]. A� oligomers levels wereassessed by western blot and by immunofluorescencewith confocal microscopy in aorta arteries. 3xTg-AD mice presented significantly higher levels of A�oligomers at the vascular wall (Fig. 6). Interestingly,high sucrose did not promote any significant alterationsin the levels of these neurotoxic species (Fig. 6). Impor-tantly, no differences on A�PP levels were observedin the groups studied (Supplementary Figure 1). Addi-tionally, the ratio of phosphorylated tau/total tau didnot significantly changed between the groups (Supple-mentary Figure 1).
DISCUSSION
The present study shows for the first time that T2Dinduced by high-sucrose intake and AD cause similar
C.M. Sena et al. / Vascular Risk Factors and Alzheimer’s Disease 133
I J
Fig. 3. Effects of AD and/or sucrose intake on vascular oxidative stress in aorta. Upper panels: In situ detection of superoxide (O2• –) in mice
aorta. Representative DHE-stained aorta artery sections reflect O2• – production in the four experimental groups. The endothelium is facing
up in all layers. At identical settings, fluorescence in 3xTg-AD (C) was markedly increased compared with WT aorta (A). Note the increasedfluorescence reflecting O2
• – levels in the sucrose-treated WT (B; WT+S) and sucrose-treated 3xTg-AD (D; 3xTg-AD+S). Lower panels:Nitrosative stress. Nitrotyrosine immunoreactivity is enhanced in 3xTg-AD and in sucrose-treated groups (WT+S and 3xTg-AD+S) indicativeof increased peroxynitrite formation. Representative aortic sections showing nitrotyrosine staining in WT (E), sucrose-treated WT (F), 3xTg-AD(G), and sucrose-treated 3xTg-AD (H). The endothelium is facing up in all layers. Panels I and J contain quantification of the fluorescence inthe different groups of arteries. Data are expressed as mean ± SE (n = 10–12). ∗∗p < 0.01, ∗∗∗p < 0.001 versus WT group; §p < 0.05 versus WT+Sgroup; φp < 0.05, φφp < 0.01 versus 3xTg-AD group.
alterations of the aorta characterized by endothelialdysfunction, which affects NO-mediated vasore-laxation, associated with enhanced oxidative andnitrosative stress and inflammation in the vascularwall. Moreover, high sucrose intake potentiates AD-associated alterations except NO-mediated vasorelax-ation perhaps because the dysfunction observed in theaorta of 3xTg-AD mice is already maximal.
3xTg-AD mice under basal conditions showed alower body weight (Table 1), a feature commonlyobserved in AD animal models [8, 11] and patients[22, 23]. 3xTg-AD mice also presented a significantincrease in postprandial blood glucose and HbA1clevels (Table 1) supporting the idea that AD is charac-
terized by an alteration of glucose metabolism. HbA1cis an early glycation product and one precursor ofadvanced glycation end products (AGEs) [24] whoseinteraction with RAGE elicits the formation of ROSthat are also believed to be an early event in ADpathology [25]. Accordingly, 3xTg-AD mice showedincreased RAGE protein levels (Fig. 4G) accompaniedby enhanced oxidative and nitrosative stress (Fig. 3C,G) in the aorta vascular wall, alterations that are con-current with cerebrovascular abnormalities [8].
As expected, animals exposed to high sucrose showmetabolic alterations similar to those observed inT2D [12]. High sucrose fed WT mice presented highblood glucose, HbA1c, and triglycerides levels and a
134 C.M. Sena et al. / Vascular Risk Factors and Alzheimer’s Disease
I J
Fig. 4. Effects of AD and/or sucrose intake on aortic inflammatory biomarkers. Upper panels: Representative aortic sections demonstratingincreased MCP-1 staining in 3xTg-AD and sucrose-treated mice. Panel presents aorta from WT (A), sucrose-treated WT (B), 3xTg-AD (C),and sucrose-treated 3xTg-AD (D) mice. Lower panels: Representative aortic sections demonstrating increased RAGE staining in aortas from3xTg-AD and sucrose-treated mice. Panel presents aorta from WT (E), sucrose-treated WT (F), 3xTg-AD (G), and sucrose-treated 3xTg-AD (H)mice. Panels I and J contain quantification of the fluorescence in the different groups of arteries. Data are expressed as mean ± SE (n = 10–12).∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001 versus WT group; §§p < 0.01 versus WT+S group; φp < 0.05, φφφp < 0.001 versus 3xTg-AD group.
significant increase in body weight (Table 1). Severalstudies showed that phenotypes associated with obe-sity and/or alterations on insulin homeostasis are atincreased risk for developing cognitive decline anddementia, namely vascular dementia and AD [26].We have previously showed that high sucrose-induceddiabetic phenotype in WT mice caused behavioraland cognitive alterations and cerebrovascular anoma-lies similar to those observed in 3xTg-AD mice [8].Interestingly, sucrose intake exacerbated the metabolicalterations occurring in 3xTg-AD mice (Table 1). Ithas been shown that diabetes influences the survivalof AD patients [26] and in vitro studies demonstratedthat high glucose levels increase the susceptibility of
brain microvascular endothelial cells to A� toxicity[28], supporting the idea that hyperglycemia is a majorrisk factor for vascular injury associated with AD.In fact, Crane and colleagues [29] recently reportedthat higher glucose levels may be a risk factor fordementia, even among people without diabetes. Highsucrose also increased the levels of plasma VCAM-1 (Table 1) and aortic MCP-1 (Fig. 4B, D) in bothWT and 3xTg-AD mice, which may predispose toatherosclerosis (Fig. 5B, D). The activation of RAGEelicits intracellular ROS generation and subsequentlyactivates mitogen-activated protein kinase (MAPK)and nuclear factor kappa-B (NF-κB) signaling, fol-lowed by production of several inflammatory and/or

C.M. Sena et al. / Vascular Risk Factors and Alzheimer’s Disease 135
WT
A B
C DWT+S
3×Tg-AD 3×Tg-AD+S
Fig. 5. Effects of AD and/or sucrose intake on aortic PAS staining. Representative slides of PAS staining of aortic sections (400x). WT mice(A); sucrose-treated WT mice (WT+S; B); 3xTg-AD mice (C); sucrose-treated 3xTg-AD (3xTg-AD+S; D).
profibrotic factors such as VCAM-1, intercellularadhesion molecule-1 (ICAM-1), plasminogen activa-tor inhibitor-1 (PAI-1), and MCP-1, thereby beinginvolved in the progression of atherosclerosis [30].
AD patients also exhibit impaired brachial arteryflow-mediated dilation, a measure of endothelialfunction and vascular disease, in comparison withnon-demented controls [31]. Consistent with thesefindings, others reported that peripheral pulse pressureand aortic pulse wave velocity, measures of arterialstiffness and subclinical atherosclerosis, are associ-ated with declines in cognitive domains known to beaffected in early AD [32, 33]. Accordingly, 3xTg-ADmice presented impaired NO-dependent vasorelax-ation (Fig. 2A). Thus, in addition to cerebrovascularanomalies, the extent of vascular disease to the periph-ery is another hallmark of AD. Cumulative evidencesuggests a possible link between A� and vascu-lar disease. Interestingly, vascular changes includingendothelial damage appear to parallel A� deposits[34, 35] and increased amounts of soluble A� havebeen shown in isolated vascular tissues from AD sub-jects [34]. Interestingly, we observed an increase inA� oligomers levels in 3xTg-AD mice aorta (Fig. 6).The 3xTg-AD line was originally generated by co-microinjection of human A�PP (K670M/N671L) andtau (P301L) transgenes under the control of the Thy 1.2
promoter into mutant PS-1 (M146V) knock-in mice[36]. The Thy 1.2 promoter can be responsible for theincreased levels of A� oligomers found in 3xTg-ADmice aorta since it has been recently described thatCD90/Thy-1 is expressed on lymphatic endothelialcells [37].
The importance of an interaction between A� andthe vascular endothelium was further described bythe presence of A�-binding to cell surface receptorson vascular endothelial cells and the upregulation ofthese receptors in AD [38], RAGE being one of thesereceptors. Several AD animal models have shown adecrement in vasodilatory capacity largely attributedto A�-induced oxidative stress [39–41]. In fact, O2
• –
triggers the synthesis of other ROS and the sequestra-tion of NO, thus impairing resting vascular tone andNO-dependent dilations. Accordingly, an increment inaortic O2
• – (Fig. 3B) and A� oligomers (Fig. 6) levelsand impaired NO-dependent vasorelaxation (Fig. 2A)were observed in 3xTg-AD. A� also contributes tocerebral smooth muscle cell contraction directly ormay induce neuronal production and release of ET-1, apotent vasoconstrictor, and upregulation of endothelin-converting enzyme 2 (ECE-2), the enzyme that cleavesET-1 from its inactive precursor [42]. Palmer and col-laborators [43] studied isolated leptomeningeal bloodvessels from postmortem AD brains and found that
136 C.M. Sena et al. / Vascular Risk Factors and Alzheimer’s Disease
Fig. 6. Effects of AD and/or sucrose intake on A� oligomers levels. Immunofluorescence of A� oligomers. Immunofluorescence of A� oligomersin WT mice (A); sucrose-treated WT mice (WT+S; B); 3xTg-AD mice (C); sucrose-treated 3xTg-AD (3xTg-AD+S; D). E) Quantification of thefluorescence expressed as arbitrary units (AU). F) Representative western blot analyses of A� oligomers levels in aortas of the different groupsof arteries. G) Averaged densitometric data expressed as a percentage of elevation over the control value established as 100% after previouslyratio to actin densitometry. Data are expressed as mean ± SE. ∗p < 0.05, ∗∗p < 0.01 versus WT group; §§p < 0.01 versus WT+S group.
ET-1 levels are significantly elevated in those ves-sels. Importantly, the increase in ET-1 levels seemsto be specific to AD as there is no specific change invascular dementia vessels [43]. In the present study,3x-Tg-AD mice demonstrated a significant impairmentof endothelial function and an increment in ET-1-mediated vasoconstriction (Fig. 2C). In accordancewith our observations, Paris et al. [44] reported thatfreshly solubilized A� enhances ET-1 induced vaso-constriction in isolated human middle cerebral andbasilar arteries.
Although oxidative stress is involved in impairedNO-dependent vasorelaxation in both WT+S and
3xTg-AD mice, we cannot rule out the possibility thatthis alteration is due to additional or independent mech-anisms. In fact, the cause(s) of endothelial dysfunctionin 3xTg-AD mice may diverge from that involved inWT sucrose-treated mice, which may underlie the dif-ferences observed in curves of Fig. 2A. Interestingly,an in contrast to the brain [8, 11], high sucrose didnot enhance aortic A� levels (Fig. 6) suggesting thatperipheral vascular anomalies associated to metabolicalterations are not triggered by A�. In fact, it has beenconsistently shown that mitochondrial dysfunction andoxidative stress are early events in AD progression[45].

C.M. Sena et al. / Vascular Risk Factors and Alzheimer’s Disease 137
In summary, these results demonstrate that AD isalso characterized by peripheral vascular alterations,a situation that can be triggered and potentiated bymetabolic alterations associated to T2D corroboratingthe idea that a close connection does exist between bothpathologies.
ACKNOWLEDGMENTS
The authors’ work is supported by Fundacaopara a Ciencia e a Tecnologia (FCT). StrategicProject (PEst-C/SAU/LA0001/2013 and PEst-C/SAU/UI3282/2011), Programa Operacional Factoresde Competitividade (COMPETE), and Quadro deReferencia Estrategico Nacional (Projecto n◦ 13853QREN DO-IT).
Authors’ disclosures available online (http://j-alz.com/manuscript-disclosures/14-1008r2).
SUPPLEMENTARY MATERIAL
The supplementary material is available in theelectronic version of this article: http://dx.doi.org/10.3233/JAD-141008.
REFERENCES
[1] Humpel C (2011) Chronic mild cerebrovascular dysfunctionas a cause for Alzheimer’s disease. Exp Gerontol 46, 225-232.
[2] Exalto LG, Whitmer RA, Kappele LJ, Biessels GJ (2012) Anupdate on type 2 diabetes, vascular dementia and Alzheimer’sdisease. Exp Gerontol 47, 858-864.
[3] Rosenberg PB, Tan CO (2013) Cocoa, neurovascular cou-pling, and neurodegeneration: The good, the bad, and the ugly.Neurology 81, 863-864.
[4] Wolozin B, Bednar MM (2006) Interventions for heart diseaseand their effects on Alzheimer’s disease. Neurol Res 28, 630-636.
[5] Jefferson AL (2010) Cardiac output as a potential risk factorfor abnormal brain aging. J Alzheimers Dis 20, 813-621.
[6] Libby P, Ridker PM, Hansson GK, Leducq TransatlanticNetwork on Atherothrombosis (2009) Inflammation inatherosclerosis: From pathophysiology to practice. J Am CollCardiol 54, 2129-2138.
[7] Sena CM, Pereira AM, Seica R (2013) Endothelial dysfunc-tion - A major mediator of diabetic vascular disease. BiochimBiophys Acta 1832, 2216-2231.
[8] Carvalho C, Machado N, Mota PC, Correia SC, Cardoso S,Santos RX, Santos MS, Oliveira CR, Moreira PI (2013) Type 2diabetic and Alzheimer’s disease mice present similar behav-ioral, cognitive, and vascular anomalies. J Alzheimers Dis 35,623-635.
[9] Grammas P (2011) Neurovascular dysfunction, inflammationand endothelial activation: Implications for the pathogenesisof Alzheimer’s disease. J Neuroinflammation 8, 26-38.
[10] Kalaria RN, Akinyemi R, Ihara M (2012) Does vascularpathology contribute to Alzheimer changes? J Neurol Sci 322,141-147.
[11] Carvalho C, Cardoso S, Correia SC, Santos RX, Santos MS,Baldeiras I, Oliveira CR, Moreira PI (2012) Metabolic alter-ations induced by sucrose intake and Alzheimer’s diseasepromote similar brain mitochondrial abnormalities. Diabetes61, 1234-1242.
[12] Moreira PI (2013) High-sugar diets, type 2 diabetes andAlzheimer’s disease. Curr Opin Clin Nutr Metab Care 16,440-445.
[13] Sena CM, Nunes E, Louro T, Proenca T, Fernandes R, BoarderMR, Seica RM (2008) Effects of alpha-lipoic acid on endothe-lial function in aged diabetic and high-fat fed rats. Br JPharmacol 153, 894-906.
[14] Sena CM, Louro T, Matafome P, Nunes E, Monteiro P, SeicaR (2009) Antioxidant and vascular effects of gliclazide in type2 diabetic rats fed high-fat diet. Physiol Res 58, 203-209.
[15] Miller FJ Jr, Gutterman DD, Rios CD, Heistad DD, DavidsonBL (1998) Superoxide production in vascular smooth mus-cle contributes to oxidative stress and impaired relaxation inatherosclerosis. Circ Res 82, 1298-1305.
[16] Blouet C, Mariotti F, Azzout-Marniche D, Mathe V,Mikogami T, Tome D, Huneau JF (2007) Dietary cysteine alle-viates sucrose-induced oxidative stress and insulin resistance.Free Radic Biol Med 42, 1089-1097.
[17] Busserolles J, Rock E, Gueux E, Mazur A, Grolier P,Rayssiguier Y (2002) Short-term consumption of a high-sucrose diet has a pro-oxidant effect in rats. Br J Nutr 87,337-342.
[18] Piga R, Naito Y, Kokura S, Handa O, Yoshikawa T(2007) Short-term high glucose exposure induces monocyte-endothelial cells adhesion and transmigration by increasingVCAM-1 and MCP-1 expression in human aortic endothelialcells. Atherosclerosis 193, 328-334.
[19] Yan SF, Yan SD, Ramasamy R, Schmidt AM (2009)Tempering the wrath of RAGE: An emerging therapeutic strat-egy against diabetic complications, neurodegeneration, andinflammation. Ann Med 41, 408-422.
[20] Ledet T, Heickendorff L, Rasmussen LM (1988) Pathologyof macrovascular disease. Baillieres Clin Endocrinol Metab2, 391-405.
[21] Lambert MP, Barlow AK, Chromy BA, Edwards C, Freed R,Liosatos M, Morgan TE, Rozovsky I, Trommer B, Viola KL,Wals P, Zhang C, Finch CE, Krafft GA, Klein WL (1998)Diffusible, nonfibrillar ligands derived from Abeta1-42 arepotent central nervous system neurotoxins. Proc Natl AcadSci U S A 95, 6448-6453.
[22] Guyonnet S, Nourhashemi F, Andrieu S (1998) A prospectivestudy in the nutritional status of Alzheimer’s patients. ArchGerontol Geriat 6, 255-262.
[23] Power DA, Noel J, Collins R, O’Neill D (2001) Circulatingleptin levels and weight loss in Alzheimer’s disease patients.Dement. Geriatr Cogn Disord 12, 167-170.
[24] Makita Z, Vlassara H, Rayfield E, Cartwright K, FriedmanE, Rodby R, Cerami A, Bucala R (1992) Hemoglobin-AGE:A circulating marker of advanced glycosylation. Science 258,651-653.
[25] Carvalho C, Correia SC, Santos RX, Cardoso S, MoreiraPI, Clark TA, Zhu X, Smith MA, Perry G (2009)Role of mitochondrial-mediated signaling pathways inAlzheimer disease and hypoxia. J Bioenerg Biomembr 41,433-440.
[26] Luchsinger JA, Tang MX, Shea S, Mayeux R (2004) Hyper-insulinemia and risk of Alzheimer disease. Neurology 63,1187-1192.
[27] Magierski R, Kłoszewska I, Sobow TM (2010) The influenceof vascular risk factors on the survival rate of patients with

138 C.M. Sena et al. / Vascular Risk Factors and Alzheimer’s Disease
dementia with Lewy bodies and Alzheimer disease. NeurolNeurochir Pol 44, 139-147.
[28] Carvalho C, Katz PS, Dutta S, Katakam PV, Moreira PI, BusijaDW (2014) Increased susceptibility to amyloid-� toxicity inrat brain microvascular endothelial cells under hyperglycemicconditions. J Alzheimers Dis 38, 75-83.
[29] Crane PK, Walker R, Hubbard RA, Li G, Nathan DM, ZhengH, Haneuse S, Craft S, Montine TJ, Kahn SE, McCormick W,McCurry SM, Bowen JD, Larson EB (2013) Glucose levelsand risk of dementia. N Engl J Med 369, 540-548.
[30] Fukami K, Yamagishi SI, Okuda S (2014) Role of AGEs-RAGE system in cardiovascular disease. Curr Pharm Des 20,2395-2402.
[31] Dede DS, Yavuz B, Yavuz BB, Cankurtaran M, Halil M, UlgerZ, Cankurtaran ES, Aytemir K, Kabakci G, Ariogul S (2007)Assessment of endothelial function in Alzheimer’s disease: IsAlzheimer’s disease a vascular disease? J Am Geriatr Soc 55,1613-1617.
[32] Waldstein SR, Rice SC, Thayer JF, Najjar SS, Scuteri A, Zon-derman AB (2008) Pulse pressure and pulse wave velocityare related to cognitive decline in the Baltimore LongitudinalStudy of Aging. Hypertension 51, 99-104.
[33] Nation DA, Wierenga CE, Delano-Wood L, Jak AJ, DelisDC, Salmon DP, Bondi MW (2010) Elevated pulse pressureis associated with age-related decline in language ability. JInt Neuropsychol Soc 16, 933-938.
[34] Kalaria RN, Premkumar DR, Pax AB, Cohen DL, LieberburgI (1996) Production and increased detection of amyloid betaprotein and amyloidogenic fragments in brain microvessels,meningeal vessels and choroid plexus in Alzheimer’s disease.Brain Res Mol Brain Res 35, 58-68.
[35] Ellis RJ, Olichney JM, Thal LJ, Mirra SS, Morris JC, BeeklyD, Heyman A (1996) Cerebral amyloid angiopathy in thebrains of patients with Alzheimer’s disease: The CERADexperience, Part XV. Neurology 46, 1592-1596.
[36] Oddo S, Caccamo A, Shepherd JD, Murphy MP, Golde TE,Kayed R, Metherate R, Mattson MP, Akbari Y, LaFerla FM(2003) Triple-transgenic model of Alzheimer’s disease with
plaques and tangles: Intracellular Abeta and synaptic dysfunc-tion. Neuron 39, 409-421.
[37] Kretschmer S, Dethlefsen I, Hagner-Benes S, Marsh LM,Garn H, Konig P (2013) Visualization of intrapulmonarylymph vessels in healthy and inflamed murine lung usingCD90/Thy-1 as a marker. PLoS One 8, e55201.
[38] Stamler JS (1996) Alzheimer’s disease. A radical vascularconnection. Nature 380, 108-111.
[39] Iadecola C, Zhang F, Niwa K, Eckman C, Turner SK,Fischer E, Younkin S, Borchelt DR, Hsiao KK, Carlson GA(1999) SOD1 rescues cerebral endothelial dysfunction in miceoverexpressing amyloid precursor protein. Nat Neurosci 2,157-161.
[40] Tong XK, Nicolakakis N, Kocharyan A, Hamel E (2005)Vascular remodeling versus amyloid beta-induced oxidativestress in the cerebrovascular dysfunctions associated withAlzheimer’s disease. J Neurosci 25, 11165-11174.
[41] Nicolakakis N, Aboulkassim T, Ongali B, Lecrux C,Fernandes P, Rosa-Neto P, Tong XK, Hamel E (2008) Com-plete rescue of cerebrovascular function in aged Alzheimer’sdisease transgenic mice by antioxidants and pioglitazone, aperoxisome proliferator-activated receptor gamma agonist. JNeurosci 28, 9287-9296.
[42] Palmer J, Love S (2011) Endothelin receptor antago-nists: Potential in Alzheimer’s disease. Pharmacol Res 63,525-531.
[43] Palmer BW, Ryan KA, Kim HM, Karlawish JH, AppelbaumPS, Kim SY (2013) Neuropsychological correlates of capacitydeterminations in Alzheimer disease: Implications for assess-ment. Am J Geriatr Psychiatry 21, 373-381.
[44] Paris D, Humphrey J, Quadros A, Patel N, Crescentini R,Crawford F, Mullan M (2003) Vasoactive effects of Abetain isolated human cerebrovessels and in a transgenic mousemodel of Alzheimer’s disease: Role of inflammation. NeurolRes 25, 642-651.
[45] Moreira PI, Carvalho C, Zhu X, Smith MA, Perry G (2010)Mitochondrial dysfunction is a trigger of Alzheimer’s diseasepathophysiology. Biochim Biophys Acta 1802, 2-10.
Related Documents











