Two NHX-type transporters from Helianthus tuberosus improve the tolerance of rice to salinity and nutrient deficiency stress Yang Zeng 1,2 , Qing Li 2 , Haiya Wang 1,2 , Jianliang Zhang 1,2 , Jia Du 1,2 , Huimin Feng 1,2 , Eduardo Blumwald 3 , Ling Yu 1,2, * and Guohua Xu 1,2, * 1 State Key Laboratory of Crop Genetics and Germplasm Enhancement, Nanjing Agricultural University, Nanjing, China 2 Key Laboratory of Plant Nutrition and Fertilization in Low-Middle Reaches of the Yangtze River, Ministry of Agriculture, Nanjing Agricultural University, Nanjing, China 3 Department of Plant Sciences, University of California, Davis, CA, USA Received 15 April 2017; revised 30 May 2017; accepted 8 June 2017. *Correspondence (Tel +0086-25-84395624; fax +0086-25-84395210; email [email protected] (L.Y.)) and (Tel +0086-25-84396246; fax +0086-25-84396246; email [email protected] (G.X.)) Keywords: Helianthus tuberosus, NHX-type transporters, nutrient deficiency, potassium, rice, salt stress. Summary The NHX-type cation/H + transporters in plants have been shown to mediate Na + (K + )/H + exchange for salinity tolerance and K + homoeostasis. In this study, we identified and characterized two NHX homologues, HtNHX1 and HtNHX2 from an infertile and salinity tolerant species Helianthus tuberosus (cv. Nanyu No. 1). HtNHX1 and HtNHX2 share identical 5 0 - and 3 0 - UTR and coding regions, except for a 342-bp segment encoding 114 amino acids (L 272 to Q 385 ) which is absent in HtNHX2. Both hydroponics and soil culture experiments showed that the expression of HtNHX1 or HtNHX2 improved the rice tolerance to salinity. Expression of HtNHX2, but not HtNHX1, increased rice grain yield, harvest index, total nutrient uptake under K + -limited salt-stress or general nutrient deficiency conditions. The results provide a novel insight into NHX function in plant mineral nutrition. Introduction Naturally occurred and fertilization-induced soil salinity, and low nutrient use efficiency are significant constraints in modern agriculture production (Heuer et al., 2016; Horie et al., 2009; Munns and Gilliham, 2015; Munns and Tester, 2008; Xu et al., 2012). At the cellular level, the extrusion of Na + ions at the cell plasma membrane and the compartmentation of Na + into vacuoles are efficient mechanisms to avoid excessive cytosolic Na + concentration and maintain an adequate cytosolic K + /Na + ratio (Julkowska and Testerink, 2015; Munns and Tester, 2008). The plasma membrane electrical potential difference of root cells is usually maintained around 120 mV, while the tonoplast (vacuole membrane) potential is positive and around 20–40 mV (Shabala et al., 2016). These potentials allow the root to acquire sufficient K + via high-affinity transporter systems (HATS) in K + - limited soils or via low-affinity transporter systems (LATS) at normal external K + supplies. The presence of high external Na + concentrations suppresses the K + conductance through LATS (Qi and Spalding, 2004) and competes with K + uptake through HATS (Alem an et al., 2011; Leidi et al., 2010), causing a decrease in intracellular K + with a concomitant [K + ]/[Na + ] imbalance. The plant vacuolar Na + /H + antiporters (NHXs) were shown originally to mediate the electroneutral Na + /H + exchange, driving the excess cytosolic Na + into the vacuole (Apse et al., 1999; Blumwald and Poole, 1985; Gaxiola et al., 1999). The NHX proteins belong to the large mono-valent cation/proton trans- porters (CPA) family, showing three distinct functional clades (Chanroj et al., 2012). In Arabidopsis, in addition to the plasma membrane-located NHX7 and NHX8, also known as SOS (salt- over-sensitive), intracellular NHXs sharing high sequence similarity are further divided into type-I and type-II, based on their subcellular location (Chanroj et al., 2012). Type-I NHXs (AtNHX1-AtNHX4) are vacuolar-located, while type-II NHXs (AtNHX5 and AtNHX6) are found at endosome, Trans-Golgi Network (TGN)/Golgi and prevacuolar compartments (Andr es et al., 2014; Barrag an et al., 2012; Bassil et al., 2011a,b; McCubbin et al., 2014). In the rice genome, there are at least five NHX members with OsNHX1-OsNHX4 belonging to the type-I and OsNHX5 to the type-II (Fukuda et al., 2011). The operation of the NHXs also affects the pH gradients across the different luminal compartments (Bassil et al., 2012). Both type-I and type-II NHXs have different subcellular localiza- tion; they may have different mechanisms in salt tolerance, and other not yet defined functions. Large efforts have been paid to increasing the plant salt tolerance by overexpressing NHX ortho- logue genes from different species. In rice, overexpression of the NHX1 homologue genes from Oryza sativa, Chenopodium glau- cum and Atriplex dimorphostegia resulted in the enhanced tolerance to salt stress (Li et al., 2008). However, these NHX proteins actually mediate both Na + /H + and K + /H + exchange and their functions cannot be solely explained by accumulating Na + into vacuole (Andr es et al., 2014; Barrag an et al., 2012; Bassil et al., 2011b; Jiang et al., 2010; Leidi et al., 2010; Reguera et al., 2014). The type-II NHXs have also been shown to be involved in salt tolerance. AtNHX5 and AtNHX6 are the only two type-II NHXs in Please cite this article as: Zeng, Y., Li, Q., Wang, H., Zhang, J., Du, J., Feng, H., Blumwald, E., Yu, L. and Xu, G. (2017) Two NHX-type transporters from Helianthus tuberosus improve the tolerance of rice to salinity and nutrient deficiency stress. Plant Biotechnol. J., https://doi.org/10.1111/pbi.12773 ª 2017 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd. This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited. 1 Plant Biotechnology Journal (2017), pp. 1–12 doi: 10.1111/pbi.12773

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Two NHX-type transporters from Helianthus tuberosusimprove the tolerance of rice to salinity and nutrientdeficiency stressYang Zeng1,2, Qing Li2, Haiya Wang1,2, Jianliang Zhang1,2, Jia Du1,2, Huimin Feng1,2, Eduardo Blumwald3,Ling Yu1,2,* and Guohua Xu1,2,*
1State Key Laboratory of Crop Genetics and Germplasm Enhancement, Nanjing Agricultural University, Nanjing, China2Key Laboratory of Plant Nutrition and Fertilization in Low-Middle Reaches of the Yangtze River, Ministry of Agriculture, Nanjing Agricultural University, Nanjing, China3Department of Plant Sciences, University of California, Davis, CA, USA
Received 15 April 2017;
revised 30 May 2017;
accepted 8 June 2017.
*Correspondence (Tel +0086-25-84395624;
fax +0086-25-84395210; email
[email protected] (L.Y.)) and
(Tel +0086-25-84396246;
fax +0086-25-84396246;
email [email protected] (G.X.))
Keywords: Helianthus tuberosus,
NHX-type transporters, nutrient
deficiency, potassium, rice, salt stress.
SummaryThe NHX-type cation/H+ transporters in plants have been shown to mediate Na+(K+)/H+
exchange for salinity tolerance and K+ homoeostasis. In this study, we identified and
characterized two NHX homologues, HtNHX1 and HtNHX2 from an infertile and salinity tolerant
species Helianthus tuberosus (cv. Nanyu No. 1). HtNHX1 and HtNHX2 share identical 50- and 30-UTR and coding regions, except for a 342-bp segment encoding 114 amino acids (L272 to Q385)
which is absent in HtNHX2. Both hydroponics and soil culture experiments showed that the
expression of HtNHX1 or HtNHX2 improved the rice tolerance to salinity. Expression of HtNHX2,
but not HtNHX1, increased rice grain yield, harvest index, total nutrient uptake under K+-limited
salt-stress or general nutrient deficiency conditions. The results provide a novel insight into NHX
function in plant mineral nutrition.
Introduction
Naturally occurred and fertilization-induced soil salinity, and low
nutrient use efficiency are significant constraints in modern
agriculture production (Heuer et al., 2016; Horie et al., 2009;
Munns and Gilliham, 2015; Munns and Tester, 2008; Xu et al.,
2012). At the cellular level, the extrusion of Na+ ions at the cell
plasma membrane and the compartmentation of Na+ into
vacuoles are efficient mechanisms to avoid excessive cytosolic
Na+ concentration and maintain an adequate cytosolic K+/Na+
ratio (Julkowska and Testerink, 2015; Munns and Tester, 2008).
The plasma membrane electrical potential difference of root cells
is usually maintained around �120 mV, while the tonoplast
(vacuole membrane) potential is positive and around 20–40 mV
(Shabala et al., 2016). These potentials allow the root to acquire
sufficient K+ via high-affinity transporter systems (HATS) in K+-
limited soils or via low-affinity transporter systems (LATS) at
normal external K+ supplies. The presence of high external Na+
concentrations suppresses the K+ conductance through LATS (Qi
and Spalding, 2004) and competes with K+ uptake through HATS
(Alem�an et al., 2011; Leidi et al., 2010), causing a decrease in
intracellular K+ with a concomitant [K+]/[Na+] imbalance.
The plant vacuolar Na+/H+ antiporters (NHXs) were shown
originally to mediate the electroneutral Na+/H+ exchange, driving
the excess cytosolic Na+ into the vacuole (Apse et al., 1999;
Blumwald and Poole, 1985; Gaxiola et al., 1999). The NHX
proteins belong to the large mono-valent cation/proton trans-
porters (CPA) family, showing three distinct functional clades
(Chanroj et al., 2012). In Arabidopsis, in addition to the plasma
membrane-located NHX7 and NHX8, also known as SOS (salt-
over-sensitive), intracellular NHXs sharing high sequence similarity
are further divided into type-I and type-II, based on their
subcellular location (Chanroj et al., 2012). Type-I NHXs
(AtNHX1-AtNHX4) are vacuolar-located, while type-II NHXs
(AtNHX5 and AtNHX6) are found at endosome, Trans-Golgi
Network (TGN)/Golgi and prevacuolar compartments (Andr�es
et al., 2014; Barrag�an et al., 2012; Bassil et al., 2011a,b;
McCubbin et al., 2014). In the rice genome, there are at least
five NHX members with OsNHX1-OsNHX4 belonging to the type-I
and OsNHX5 to the type-II (Fukuda et al., 2011). The operation of
the NHXs also affects the pH gradients across the different
luminal compartments (Bassil et al., 2012).
Both type-I and type-II NHXs have different subcellular localiza-
tion; they may have different mechanisms in salt tolerance, and
other not yet defined functions. Large efforts have been paid to
increasing the plant salt tolerance by overexpressing NHX ortho-
logue genes from different species. In rice, overexpression of the
NHX1 homologue genes from Oryza sativa, Chenopodium glau-
cum and Atriplex dimorphostegia resulted in the enhanced
tolerance to salt stress (Li et al., 2008). However, these NHX
proteins actually mediate both Na+/H+ and K+/H+ exchange and
their functions cannot be solely explained by accumulating Na+ into
vacuole (Andr�es et al., 2014; Barrag�an et al., 2012; Bassil et al.,
2011b; Jiang et al., 2010; Leidi et al., 2010; Reguera et al., 2014).
The type-II NHXs have also been shown to be involved in salt
tolerance. AtNHX5 and AtNHX6 are the only two type-II NHXs in
Please cite this article as: Zeng, Y., Li, Q., Wang, H., Zhang, J., Du, J., Feng, H., Blumwald, E., Yu, L. and Xu, G. (2017) Two NHX-type transporters from Helianthus
tuberosus improve the tolerance of rice to salinity and nutrient deficiency stress. Plant Biotechnol. J., https://doi.org/10.1111/pbi.12773
ª 2017 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd.This is an open access article under the terms of the Creative Commons Attribution License, which permits use,distribution and reproduction in any medium, provided the original work is properly cited.
1
Plant Biotechnology Journal (2017), pp. 1–12 doi: 10.1111/pbi.12773
Arabidopsis. Although their function appears to be redundant,
the double knockout Atnhx5nhx6 displayed high sensitivity to salt
stress (Bassil et al., 2011a). In tomato, the overexpression of
LeNHX2, a type-II NHX located in prevacuolar compartments and
Golgi, enhanced salt tolerance at high external K+ levels (Venema
et al., 2003).
Plants use a number of strategies to deal with high salinity
(Munns and Tester, 2008). Halophytes rely heavily on the
homoeostasis of three major inorganic ions (Na+, Cl�, and K+)
to maintain their osmotic and turgor pressure under saline
conditions, while glycophytes predominantly increase the synthe-
sis of compatible solutes (Deinlein et al., 2014). For salt tolerant
plants grown in high external saline conditions, the efficient
compartmentation of Na+ into vacuole and other cell organs via
intracellular NHXs is of importance, together with the extrusion of
Na+ at the roots, via plasma membrane-bound antiporters such as
SOS (Deinlein et al., 2014; Munns and Tester, 2008).
Jerusalem artichoke (Helianthus tuberosus L.) belonging to the
same Asteraceae family as sunflower (Helianthus annuus L.) is an
herbaceous perennial plant that have potential as a biorefinery
crop (Johansson et al., 2015). H. tuberosus is highly tolerant to
infertile, drought and saline stresses and its variety Nanyu No. 1
(NY-1) could fully emerge in the coastal region containing 1% salt
at soil surface layer (0–15 cm) and grow well in soil containing
0.5%–0.6% salt or under irrigation with 50%–75% sea water
(Long et al., 2008). In this study, we isolated two putative NHX
genes, HtNHX1 and HtNHX2, from H. tuberosus cv. NY-1 and
examined their roles in enhancing the tolerance of salt stress and
nutrient deficiency in rice, Arabidopsis and yeast. Our results
indicate that HtNHX2 could function in enhancing the plant
tolerance to salinity stress and improving plant nutrient efficiency.
Results
Cloning of salt-stress up-regulated HtNHX1 and HtNHX2genes from Helianthus tuberosus
To clone NHX genes in H. tuberosus, we designed the primers
(Table S1) based on the conserved regions of known NHXs in
plants (Figure S1) for PCR amplification of cDNA isolated from the
salt-treated cv. NY-1. The two different fragments were obtained,
and their full lengths were further isolated by 50 and 30 RACE-PCR.The two putative NHX genes with 2148- and 1806-bp encoding
regions were named HtNHX1 and HtNHX2, respectively.
Interestingly, HtNHX1 and HtNHX2 have identical 269 bp of 50
UTR and 501 bp of 30UTR. Furthermore, their coding sequences
are also identical except a segment of 342 bp in HtNHX1
encoding 114 amino acids (L272 to Q385) which is absent in
HtNHX2 (Figure S1A). By alignment of the amino acids sequences
of HtNHX1 and HtNHX2 with more than forty NHX members
from different plant species, both HtNHX1 and HtNHX2 are
classified as IC type-I NHXs (Figure S1B). Transmembrane topol-
ogy predicted by TMpred (http://www.ch.embnet.org/software/
TMPRED_form.html) or TMHMM2.0 (http://www.cbs.dtu.dk/se
rvices/TMHMM/) indicated that HtNHX1 comprised 10 transmem-
brane domains, while HtNHX2 has seven to eight transmembrane
domains (Figure S1CD).
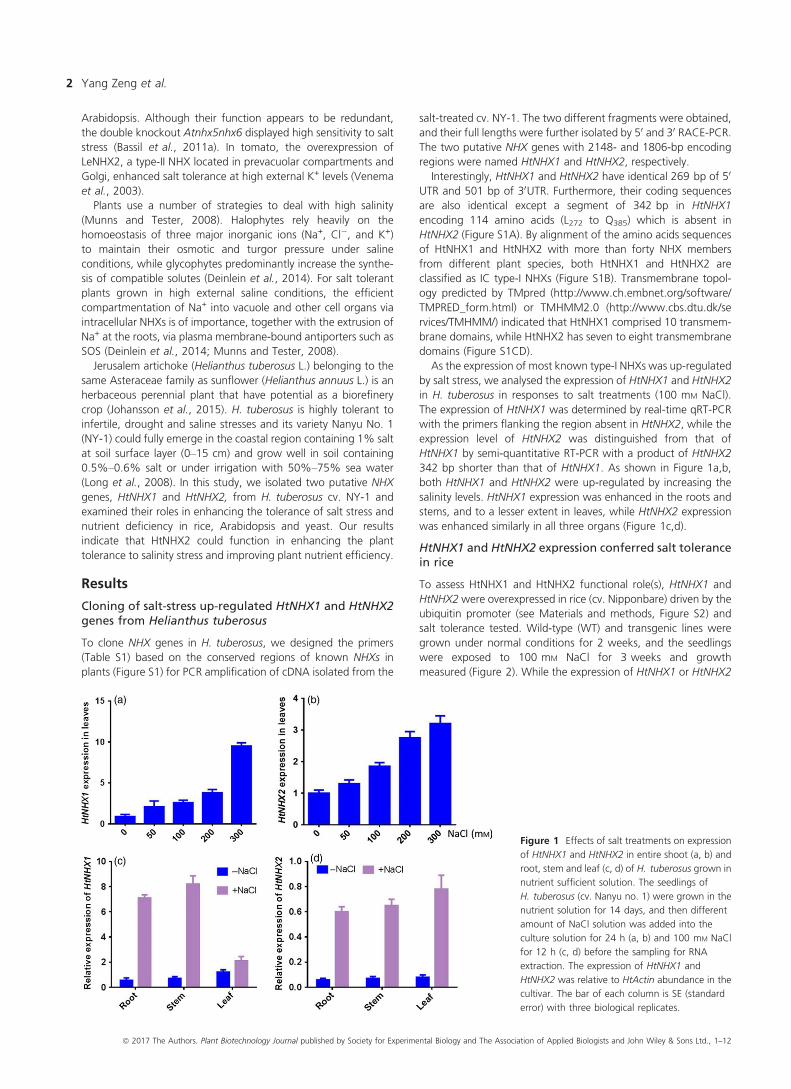
As the expression of most known type-I NHXs was up-regulated
by salt stress, we analysed the expression of HtNHX1 and HtNHX2
in H. tuberosus in responses to salt treatments (100 mM NaCl).
The expression of HtNHX1 was determined by real-time qRT-PCR
with the primers flanking the region absent in HtNHX2, while the
expression level of HtNHX2 was distinguished from that of
HtNHX1 by semi-quantitative RT-PCR with a product of HtNHX2
342 bp shorter than that of HtNHX1. As shown in Figure 1a,b,
both HtNHX1 and HtNHX2 were up-regulated by increasing the
salinity levels. HtNHX1 expression was enhanced in the roots and
stems, and to a lesser extent in leaves, while HtNHX2 expression
was enhanced similarly in all three organs (Figure 1c,d).
HtNHX1 and HtNHX2 expression conferred salt tolerancein rice
To assess HtNHX1 and HtNHX2 functional role(s), HtNHX1 and
HtNHX2 were overexpressed in rice (cv. Nipponbare) driven by the
ubiquitin promoter (see Materials and methods, Figure S2) and
salt tolerance tested. Wild-type (WT) and transgenic lines were
grown under normal conditions for 2 weeks, and the seedlings
were exposed to 100 mM NaCl for 3 weeks and growth
measured (Figure 2). While the expression of HtNHX1 or HtNHX2
Figure 1 Effects of salt treatments on expression
of HtNHX1 and HtNHX2 in entire shoot (a, b) and
root, stem and leaf (c, d) of H. tuberosus grown in
nutrient sufficient solution. The seedlings of
H. tuberosus (cv. Nanyu no. 1) were grown in the
nutrient solution for 14 days, and then different
amount of NaCl solution was added into the
culture solution for 24 h (a, b) and 100 mM NaCl
for 12 h (c, d) before the sampling for RNA
extraction. The expression of HtNHX1 and
HtNHX2 was relative to HtActin abundance in the
cultivar. The bar of each column is SE (standard
error) with three biological replicates.
ª 2017 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd., 1–12
Yang Zeng et al.2
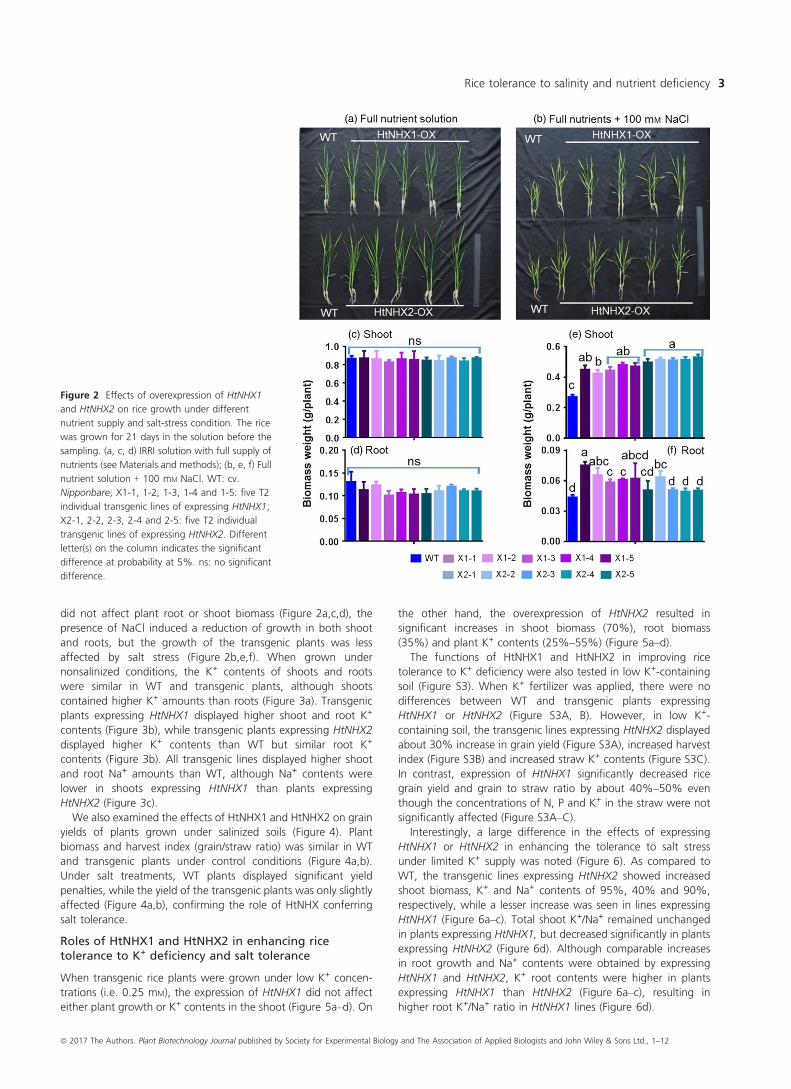
did not affect plant root or shoot biomass (Figure 2a,c,d), the
presence of NaCl induced a reduction of growth in both shoot
and roots, but the growth of the transgenic plants was less
affected by salt stress (Figure 2b,e,f). When grown under
nonsalinized conditions, the K+ contents of shoots and roots
were similar in WT and transgenic plants, although shoots
contained higher K+ amounts than roots (Figure 3a). Transgenic
plants expressing HtNHX1 displayed higher shoot and root K+
contents (Figure 3b), while transgenic plants expressing HtNHX2
displayed higher K+ contents than WT but similar root K+
contents (Figure 3b). All transgenic lines displayed higher shoot
and root Na+ amounts than WT, although Na+ contents were
lower in shoots expressing HtNHX1 than plants expressing
HtNHX2 (Figure 3c).
We also examined the effects of HtNHX1 and HtNHX2 on grain
yields of plants grown under salinized soils (Figure 4). Plant
biomass and harvest index (grain/straw ratio) was similar in WT
and transgenic plants under control conditions (Figure 4a,b).
Under salt treatments, WT plants displayed significant yield
penalties, while the yield of the transgenic plants was only slightly
affected (Figure 4a,b), confirming the role of HtNHX conferring
salt tolerance.
Roles of HtNHX1 and HtNHX2 in enhancing ricetolerance to K+ deficiency and salt tolerance
When transgenic rice plants were grown under low K+ concen-
trations (i.e. 0.25 mM), the expression of HtNHX1 did not affect
either plant growth or K+ contents in the shoot (Figure 5a–d). On
the other hand, the overexpression of HtNHX2 resulted in
significant increases in shoot biomass (70%), root biomass
(35%) and plant K+ contents (25%–55%) (Figure 5a–d).The functions of HtNHX1 and HtNHX2 in improving rice
tolerance to K+ deficiency were also tested in low K+-containing
soil (Figure S3). When K+ fertilizer was applied, there were no
differences between WT and transgenic plants expressing
HtNHX1 or HtNHX2 (Figure S3A, B). However, in low K+-
containing soil, the transgenic lines expressing HtNHX2 displayed
about 30% increase in grain yield (Figure S3A), increased harvest
index (Figure S3B) and increased straw K+ contents (Figure S3C).
In contrast, expression of HtNHX1 significantly decreased rice
grain yield and grain to straw ratio by about 40%–50% even
though the concentrations of N, P and K+ in the straw were not
significantly affected (Figure S3A–C).Interestingly, a large difference in the effects of expressing
HtNHX1 or HtNHX2 in enhancing the tolerance to salt stress
under limited K+ supply was noted (Figure 6). As compared to
WT, the transgenic lines expressing HtNHX2 showed increased
shoot biomass, K+ and Na+ contents of 95%, 40% and 90%,
respectively, while a lesser increase was seen in lines expressing
HtNHX1 (Figure 6a–c). Total shoot K+/Na+ remained unchanged
in plants expressing HtNHX1, but decreased significantly in plants
expressing HtNHX2 (Figure 6d). Although comparable increases
in root growth and Na+ contents were obtained by expressing
HtNHX1 and HtNHX2, K+ root contents were higher in plants
expressing HtNHX1 than HtNHX2 (Figure 6a–c), resulting in
higher root K+/Na+ ratio in HtNHX1 lines (Figure 6d).
Figure 2 Effects of overexpression of HtNHX1
and HtNHX2 on rice growth under different
nutrient supply and salt-stress condition. The rice
was grown for 21 days in the solution before the
sampling. (a, c, d) IRRI solution with full supply of
nutrients (see Materials and methods); (b, e, f) Full
nutrient solution + 100 mM NaCl. WT: cv.
Nipponbare; X1-1, 1-2, 1-3, 1-4 and 1-5: five T2
individual transgenic lines of expressing HtNHX1;
X2-1, 2-2, 2-3, 2-4 and 2-5: five T2 individual
transgenic lines of expressing HtNHX2. Different
letter(s) on the column indicates the significant
difference at probability at 5%. ns: no significant
difference.
ª 2017 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd., 1–12
Rice tolerance to salinity and nutrient deficiency 3

HtNHX2, but not HtNHX1, enhanced rice tolerance tolimited supply of major nutrients
While the ectopic expression of HtNHX1 or HtNHX2 did not
induce phenotypical differences in rice grown under control
conditions, transgenic rice plants expressing HtNHX2 displayed
remarkable higher growth than WT or transgenic plants express-
ing HtNHX1, when grown under low K supply (Figure 5).
Considering the essential roles of K+ in balancing uptake and
distribution of anions, particularly, nitrate and phosphate (Drew
and Saker, 1984; Drew et al., 1990; Engels and Marschner, 1992)
and improving plant growth, we characterized the effects of
expressing HtNHX1 and HtNHX2 on enhancing the tolerance of
rice to nutrient deficiency. The reduction in nutrient supply to 1/4
of its full strength decreased the growth and biomass of WT and
HtNHX1-expressing rice equally (Figure 7a,b) while did not affect
the growth of HtNHX2-expressing rice plants (Figure 7a,b). The
HtNHX2 lines showed about 35%, 25% and 45% increase in
total biomass, N and P contents, respectively, in comparison with
WT (Figure 7b–d), indicating that HtNHX2 expression could
remarkably enhance the root acquisition of N and P, thus
improving plant growth under limited nutrient supply conditions.
In contrast, N and P contents in the HtNHX1 lines were lower than
those in WT. 15N quantification analysis of rice grown in low N
and K conditions showed that HtNHX2 expression did not alter
root N contents, but significantly increased shoot N contents
(Figure S4), suggesting the enhanced N translocation from shoot
to root, in addition to the increase in total N uptake.
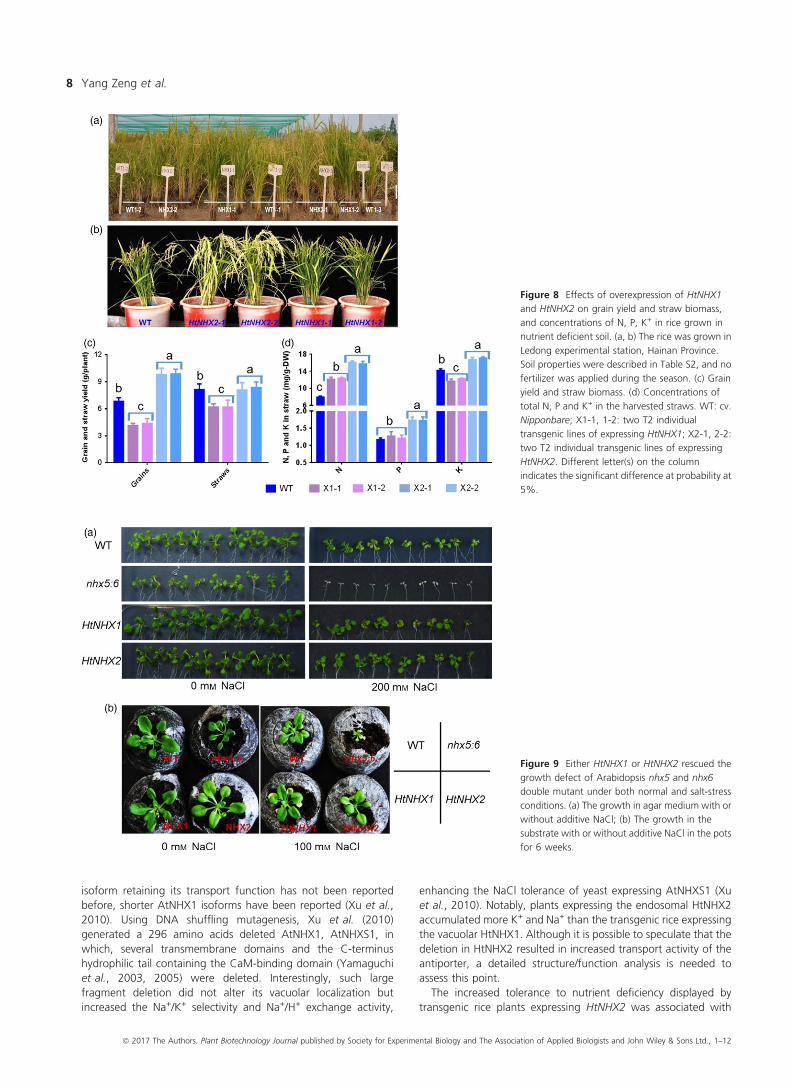
The notion of the role(s) of HtNHX2, but not HtNHX1, in
enhancing K+ uptake and mediating N and P accumulation was
further supported by the growth of rice plants in infertile paddy
fields (low N and K contents) (see Table S2). Compared to WT,
HtNHX1 lines showed 20% lower straw weight (biomass) and
40% lower grain yield, resulting in significant lower grain harvest
index due to a larger portion of unfilled grains (Figure 8a–c). TheHtNHX1 lines contained significant higher total N, similar P and
lower K+ in the straw at mature stage (Figure 8d). Remarkably,
the HtNHX2 lines showed 45% increase of total grain yield and
90%, 40% and 13% higher N, P and K+ contents in the straw
(Figure 8c,d).
Both HtNHX1 and HtNHX2 could rescue the growthimpairment of nhx5 and nhx6 double mutant ofArabidopsis
In Arabidopsis, AtNHX5 and AtNHX6 are located at TGN, Golgi
and prevacuolar compartments and the double knockout
Atnhx5nhx6 displayed growth impairment under normal growth
condition and high sensitivity to salinity (Bassil et al., 2011a). We
found that either HtNHX1 or HtNHX2 expression could rescue the
growth defects and salt sensitivity of the Atnhx5nhx6 mutant
similarly (Figure 9a,b), suggesting a functional similarity between
HtNHX1, HtNHX2 and AtNHX5, AtNHX6. Interestingly, only
HtNHX1, but not HtNHX2, could enhance the tolerance of nhx1
mutant yeast (AXT3 strains) to hygromycin (Figure S5). In
addition, the transformation of the Atnhx5nhx6 double mutant
with HtNHX1 or HtNHX2 resulted in increased hygromycin
resistance, although HtNHX1-expressing plants displayed much
stronger hygromycin tolerance (Figure S5).
HtNHX1 and HtNHX2 showed different subcellularlocalization
The differential response displayed by transgenic rice plants
expressing HtNHX1 or HtNHX2 would suggest their dissimilar
cellular localization. Therefore, we used rice protoplasts express-
ing HtNHX1-GFP or HtNHX2-GFP encoding chimeras and several
Arabidopsis markers to detect their localization. HtNHX1-GFP, but
not HtNHX2-GFP fluorescence colocalized with the RFP signal of
the tonoplast marker AtTPK1 (Figure 10a). Either HtNHX1-GFP or
HtNHX2-GFP fluorescence was largely separated from RFP signal
of the endoplasmic reticulum (ER) marker AtHDEL (Figure 10b).
Figure 3 Effects of overexpression of HtNHX1 and HtNHX2 on uptake of
K+ and Na+ under full nutrient supply and salt-stress condition. (a, b) K+
concentration in shoot and root under full nutrient supply and
nonsalinized condition (1 mM K+ + 0 mM Na+) (a) and under salinized
condition (1 mM K+ + 100 mM Na+) (b); (c) Na+ concentration in shoot and
root under salinized condition (1 mM K+ + 100 mM NaCl). WT: cv.
Nipponbare; X1-1, 1-2, 1-3, 1-4 and 1-5: five T2 individual transgenic lines
of expressing HtNHX1; X2-1, 2-2, 2-3, 2-4 and 2-5: five T2 individual
transgenic lines of expressing HtNHX2. Different letter(s) on the column
indicates the significant difference at probability at 5%. ns: no significant
difference.
ª 2017 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd., 1–12
Yang Zeng et al.4

It has been addressed that the type-I NHX members including
AtNHX1-AtNHX4 are located mainly at the vacuolar membranes
while the type-II NHX members including AtNHX5, AtNHX6,
LeNHX2, HvNHX4 and OsNHX5 are at endosome membranes
(Bassil et al., 2012).We aligned sequences of HtNHX1 andHtNHX2
with those of known plant NHXs and found a patch of conserved
eight amino acids (S/T,P/F,G,X,S/T,Ø,X,V) in type-I NHXs ismissed in
the sequences of type-II NHXs (Figure S1C). Interestingly, this eight
amino acids motif is also missed in HtNHX2 (Figure S1C). To gain
insight on the effect of these eight amino acids on the cellular
localization of type-I NHXmembers, we generated the HtNHX1del-
GFP encoding chimera and expressed it in rice protoplasts.
Interestingly, the mutated HtNHX1 was separated from AtTPK1
location (Figure 10a), but largely overlapped with the ER marker
(AtHDEL) (Figure 10b). The results suggested that the eight amino
acids patch was important for targeting HtNHX1 to the vacuolar
membrane.
In the rice protoplasts, HtNHX2 localized similar to AtNHX5 and
partially overlapped with AtSYP61 and AtScamp (Figure 10c),
indicating that HtNHX2 was largely distributed at endosomal
membranes.
Discussion
Plant NHX-type antiporters play important roles in salt tolerance
and the maintenance of cellular K+ and Na+ homoeostasis and
the regulation of intracellular pH. In this study, we cloned two
highly identical salt-stress up-regulated NHX orthologues genes,
HtNHX1 and HtNHX2 from H. tuberosus (Figures 1 and S1).
HtNHX1 and HtNHX2 share identical 50-UTR, 30-UTR and coding
sequences, except for a fragment of consecutive 342 bp in
HtNHX1 which is absent in HtNHX2. Whether the two genes
were alternatively spliced or evolutionary independently gener-
ated is not clear. According to their sequences, both genes are
predicted to belong to the type-I family and to locate at the
vacuole (Figure S1B). The ability of HtNHX1 but not HtNHX2 to
confer resistance to hygromycin in yeast and Arabidopsis
(Figure S5) and different cellular localization in rice (Figure 10)
would suggest functional differences between the two isoforms.
Notably, the role of HtNHX2 in conferring tolerance to nutrient
deficiency, in addition to conferring tolerance to salinity, provides
a novel insight on NHX functions (Figures 5, 6, 7, 8, S3 and S4).
The contribution of HtNHX1 and HtNHX2 to salttolerance in rice depends on the K+-supply levels
We observed that HtNHX1 or HtNHX2 improved rice tolerance to
salt stress (comparison of Figures 2b,e,f and 3b with Figure 2a,c,
d; Figure 4a,c), but they did not have significant influence on
growth and uptake of K+ and Na+ at normal growth conditions
(Figures 2a,c,d, 3a and 4a,c). At salinized conditions, HtNHX1
increased rice K+ and Na+ accumulation, that is keeping the same
ratio of K+/Na+ as that in WT (Figure 3d). This indicated that
HtNHX1 indistinguishably transported K+ and Na+, which is
similar to that of AtNHX1 and AtNHX2 in Arabidopsis (Barrag�an
et al., 2012; Bassil et al., 2011b). In addition, the K+ and Na+
contents of plant expressing HtNHX1 were higher in the roots
(about 40%–50%) and the shoots (about 10%–25%) than in WT
plants (Figure 3b,c), supporting the role of HtNHX1 as type-I
NHXs (Figure 10a) in sequestering Na+(K+) into the vacuoles
(Barrag�an et al., 2012; Bassil et al., 2011b). Interestingly, the
accumulation and distribution of K+ and Na+ in plants expressing
HtNHX2 differed from that seen in HtNHX1 plants, as HtNHX2
plants accumulated relatively more Na+ and K+ (Figure 3c). These
results were in agreement with the localization of HtNHX2 to
intracellular compartments other than the vacuole (Figure 10a).
Endosomal trafficking and the vesicle fusion to the vacuole are
important components of the response of plant cells to salinity.
Transporters playing roles in cation sequestration, such as NHX1
(Apse et al., 1999) and H+-pumps (Gaxiola et al., 2001), depend
on vesicular trafficking for their delivery to the tonoplast. Thus,
the expression of endosomal NHXs may influence protein
trafficking from the Golgi/TGN to the vacuoles (Bassil et al.,
Figure 4 Effects of overexpression of HtNHX1 and HtNHX2 on rice growth, harvest index grown in the soil with different amount of additive salts
(NaCl). Two-week-old seedlings were acclimated to the soil containing 0.1% (1-g/kg soil) NaCl for 1 week before being transplanted to the pot
filled with 40 kg of soil which had been fertilized and mixture with indicated extra amount of salts for 2 weeks. Control: no additive NaCl was
provided into the fertile soil (see Table S2); 0.13% and 0.18% salts: adding 1.3 and 1.8 g NaCl per kg soil in the pot. WT: cv. Nipponbare; X1-1, 1-2: two
T2 individual transgenic lines of expressing HtNHX1; X2-1, 2-2: two T2 individual transgenic lines of expressing HtNHX2. Different letter(s) on
the column indicates the significant difference at probability at 5%. ns: no significant difference.
ª 2017 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd., 1–12
Rice tolerance to salinity and nutrient deficiency 5
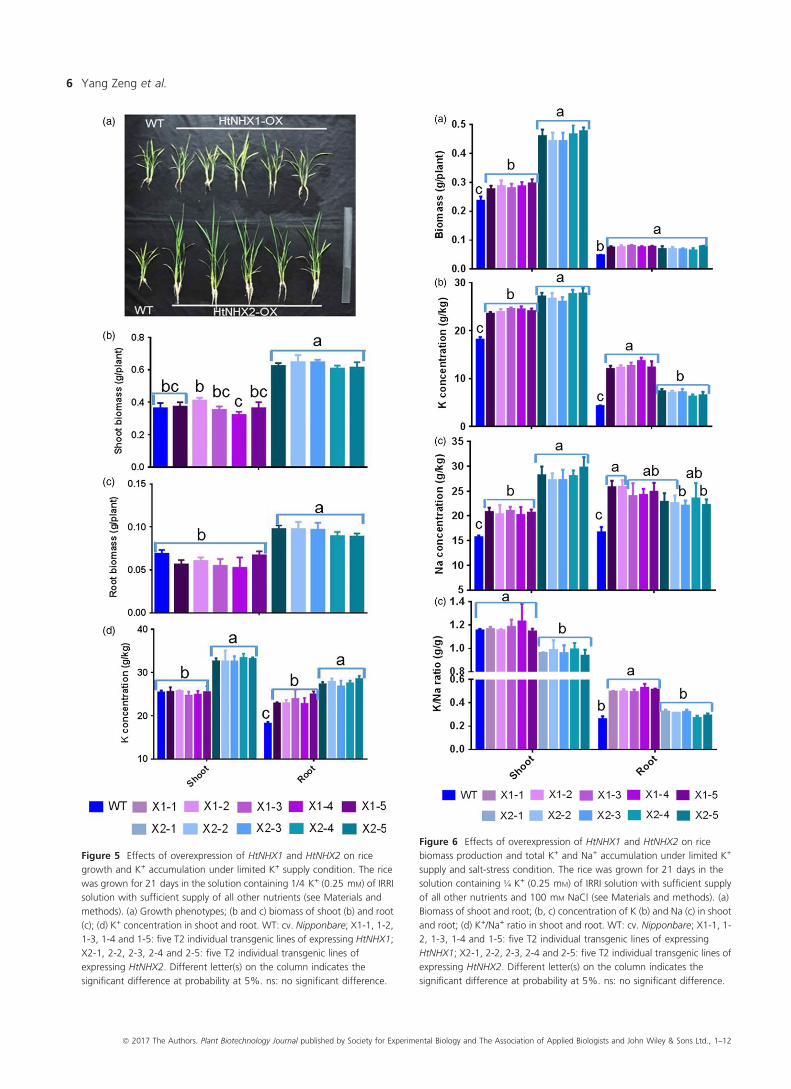
Figure 5 Effects of overexpression of HtNHX1 and HtNHX2 on rice
growth and K+ accumulation under limited K+ supply condition. The rice
was grown for 21 days in the solution containing 1/4 K+ (0.25 mM) of IRRI
solution with sufficient supply of all other nutrients (see Materials and
methods). (a) Growth phenotypes; (b and c) biomass of shoot (b) and root
(c); (d) K+ concentration in shoot and root. WT: cv. Nipponbare; X1-1, 1-2,
1-3, 1-4 and 1-5: five T2 individual transgenic lines of expressing HtNHX1;
X2-1, 2-2, 2-3, 2-4 and 2-5: five T2 individual transgenic lines of
expressing HtNHX2. Different letter(s) on the column indicates the
significant difference at probability at 5%. ns: no significant difference.
Figure 6 Effects of overexpression of HtNHX1 and HtNHX2 on rice
biomass production and total K+ and Na+ accumulation under limited K+
supply and salt-stress condition. The rice was grown for 21 days in the
solution containing ¼ K+ (0.25 mM) of IRRI solution with sufficient supply
of all other nutrients and 100 mM NaCl (see Materials and methods). (a)
Biomass of shoot and root; (b, c) concentration of K (b) and Na (c) in shoot
and root; (d) K+/Na+ ratio in shoot and root. WT: cv. Nipponbare; X1-1, 1-
2, 1-3, 1-4 and 1-5: five T2 individual transgenic lines of expressing
HtNHX1; X2-1, 2-2, 2-3, 2-4 and 2-5: five T2 individual transgenic lines of
expressing HtNHX2. Different letter(s) on the column indicates the
significant difference at probability at 5%. ns: no significant difference.
ª 2017 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd., 1–12
Yang Zeng et al.6

2011), necessary for the response to high salinity. The up-
regulation of endosomal NHXs, such as AtNHX5 in response to
salt stress, but not to osmotic shock, supported the role of
endosomal AtNHXs (Bassil et al., 2011a; Yokoi et al., 2002). In
addition, salinity induced bulk endocytosis, promoted the rapid
increase in vacuolar volume and the accumulation of sodium into
the vacuole (Baral et al., 2015; Hamaji et al., 2009; Leshem et al.,
2006; Mimura et al., 2003). These results suggest that the
overexpression of endosomal NHXs, such as HtNHX2, contributed
to salt tolerance through several mechanisms: increasing vesicle
fusion to the vacuole, contributing to the accumulation of Na+
and by increasing the trafficking of transporters that could explain
the improved nutrient uptake in the transgenic plants.
HtNHX1 and HtNHX2 rescued the salt sensitivity of the
Arabidopsis Atnhx5 nhx6 double mutant at the same extent
(Figure 9a,b), but HtNHX1 was more effective than HtNHX2 in
conferring tolerance of Arabidopsis to external hygromycin
(Figure S5). It has been shown that AtNHX5 and AtNHX6 located
at endosomes, TGN and prevacuolar compartments and nhx5
nhx6 double knockouts displayed abnormal vesicular trafficking
and sensitivity to salinity (Bassil et al., 2011a). In rice, HtNHX2
showed similar cellular localization as AtNHX5 (Figure 10c); thus,
it is plausible that HtNHX2 may play a similar role as AtNHX5 in
the regulation of endosomal ion and pH homoeostasis. Never-
theless, these results should be taken with some caution as both
HtNHX1 and HtNHX2 were expressed under the control of a
constitutive promoter (ubiquitin) and abnormal localization due to
unregulated expression cannot be ruled out.
The expression of HtNHX2, but not HtNHX1, improvedrice growth and grain yield
Potassium is a plant essential nutrient, and vacuolar K+ plays roles in
the regulation of cellular volume and tissue expansion. Although
plants grown under different K+ supply can change their vacuolar
K+ contents, cytosolic K+ ismaintained at a relative constant level of
around 60–100 mM (Cuin et al., 2003; Walker et al., 1996). The
activity ofNHXsplays a significant role in the transport of K+ into the
vacuole (Bassil et al., 2012). The overexpression of AtNHX1 in
tomato led to increased vacuolar K+ accumulation (Zhang and
Blumwald, 2001) and higher tissue K+ contents. In the knockout
Atnhx1, Apse et al. (2003) reported a reduction in K+/H+ and Na+/
H+ exchange and reduced cell expansion. Bassil et al. (2011b)
observed lower vacuole pH and K+ concentration in the nhx1 nhx2
mutant, further supporting the role of AtNHX1 and AtNHX2 in
driving the uptake of K+ into the vacuole.
The expression of HtNHX2 improved plant growth and grain
yield at low K+ concentrations (Figures 5, S3), and the tissue K+
contents of the transgenic plants were higher than the WT and
HtNHX1-expressing plants. Notably, HtNHX2 localization
appeared to be similar to that of AtNHX5 (Figure 10), that is
intracellular vesicles, TGN. (Bassil et al., 2011a), thus associated
with vesicular trafficking and storage protein sorting (Reguera
et al., 2014). In yeast, nhx1 mutants have been shown to have
impaired vacuolar biogenesis and protein sorting (Qiu and Fratti,
2010). In tomato, LeNHX2 colocalized with prevacuolar and Golgi
markers and appeared to be more selective to K+ than Na+
(Venema et al., 2003). Interestingly, HtNHX2 lacks a continuous
114 amino acids stretch at predicted six to eight transmembrane
domains of HtNHX1 (Figure S1D). This stretch also comprises a
segment of eight amino acids that is absent in type-II NHXs
(Figure S1C). Although the occurrence of a shorter HtNHX2
Figure 7 Effects of overexpression of HtNHX1 and HtNHX2 on rice growth
and N and P concentrations under supply of¼ concentrations of all the
essential nutrients in IRRI culture solution. (a) The rice was grown in IRRI
solutionwith¼ concentrationsof the full supply; (b)biomassof shootand root;
(c, d) concentration of N (c) and P (d) in shoot and root. WT: cv. Nipponbare;
X1-1, 1-2: two T2 individual transgenic lines of expressingHtNHX1; X2-1, 2-2:
twoT2 individual transgenic lines ofexpressingHtNHX2. Different letteron the
column indicates the significant difference at probability at 5%.
ª 2017 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd., 1–12
Rice tolerance to salinity and nutrient deficiency 7
isoform retaining its transport function has not been reported
before, shorter AtNHX1 isoforms have been reported (Xu et al.,
2010). Using DNA shuffling mutagenesis, Xu et al. (2010)
generated a 296 amino acids deleted AtNHX1, AtNHXS1, in
which, several transmembrane domains and the C-terminus
hydrophilic tail containing the CaM-binding domain (Yamaguchi
et al., 2003, 2005) were deleted. Interestingly, such large
fragment deletion did not alter its vacuolar localization but
increased the Na+/K+ selectivity and Na+/H+ exchange activity,
enhancing the NaCl tolerance of yeast expressing AtNHXS1 (Xu
et al., 2010). Notably, plants expressing the endosomal HtNHX2
accumulated more K+ and Na+ than the transgenic rice expressing
the vacuolar HtNHX1. Although it is possible to speculate that the
deletion in HtNHX2 resulted in increased transport activity of the
antiporter, a detailed structure/function analysis is needed to
assess this point.
The increased tolerance to nutrient deficiency displayed by
transgenic rice plants expressing HtNHX2 was associated with
Figure 8 Effects of overexpression of HtNHX1
and HtNHX2 on grain yield and straw biomass,
and concentrations of N, P, K+ in rice grown in
nutrient deficient soil. (a, b) The rice was grown in
Ledong experimental station, Hainan Province.
Soil properties were described in Table S2, and no
fertilizer was applied during the season. (c) Grain
yield and straw biomass. (d) Concentrations of
total N, P and K+ in the harvested straws. WT: cv.
Nipponbare; X1-1, 1-2: two T2 individual
transgenic lines of expressing HtNHX1; X2-1, 2-2:
two T2 individual transgenic lines of expressing
HtNHX2. Different letter(s) on the column
indicates the significant difference at probability at
5%.
Figure 9 Either HtNHX1 or HtNHX2 rescued the
growth defect of Arabidopsis nhx5 and nhx6
double mutant under both normal and salt-stress
conditions. (a) The growth in agar medium with or
without additive NaCl; (b) The growth in the
substrate with or without additive NaCl in the pots
for 6 weeks.
ª 2017 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd., 1–12
Yang Zeng et al.8

increased K+ contents and enhanced N assimilation as seen by the
increase in 15N translocation from root to shoot (Figure S4) and
the amounts of N, P and K+ in the straw of plants grown in
infertile soil (Figure 8a,b). K+ is the major accompanying ion for
the translocation of NO3� and sucrose in plants (Drew and Saker,
1984; Drew et al., 1990). About 40%–90% of root acquired K+
could be retranslocated from the shoot via the phloem and
recycled through the roots (Lu et al., 2005; Peuke, 2010), and K+
recycling in plants can act as an important signal for feedback
control of nutrient uptake (Drew et al., 1990; Engels and
Marschner, 1992). It could be speculated that the expression of
HtNHX2 improved the synthesis/delivery of the transporters of N,
P and K+ to plasma membranes by improving vesicular trafficking
and/or protein targeting (Reguera et al., 2014).
The functions of NHX-type transporters have been extensively
studied (Bassil et al., 2012 and references therein), and the
effects of the overexpression of NHXs on salt tolerance have been
shown in different crop species (Bassil and Blumwald, 2014).
Here, we show that although the expression of HtNHX1
conferred salt tolerance, only the expression of HtNHX2, a
shorter isoform ot the known type-II NHXs, conferred salt
tolerance under nutrient stress conditions.
Experimental procedures
Plant materials and growth conditions
The tubers of H. tuberosus genotype NY-1 after dormancy were
germinated on moist sand in an incubator. Uniformly germinated
slices of the buds were selected, sown in sands and then
transplanted into 1/2 Hoagland nutrient solution in the green-
house as described previously (Li et al., 2014). After emergence
of the fourth leaf, the seedlings were treated with 50, 100, 200
and 300 mM NaCl for 24 h or with 100 mM NaCl for 12 h before
the sampling for analysis of HtNHX1 and HtNHX2 expression.
Each treatment was replicated three times.
Arabidopsis thaliana (ecotype Columbia (Col-0) wild-type (WT)
and nhx5 nhx6 double knockout mutants (Bassil et al., 2011b)
were provided by Professor Eduardo Blumwald at University of
California (Davis, California). For the salt and hygromycin toler-
ance analysis in agar medium, the seeds were germinated and
grown by following the method described by Bassil et al. (2011b).
For the salt tolerance analysis in the pot filled with peat culture
medium, the seedlings of uniform size after 2 weeks growth in a
plate were selected and transplanted to the nutrient-enriched
medium in a growth chamber at 23 °C with a 14 : 10 h L : D
photoperiod. For salt treatment, NaCl was added to final
concentrations as indicated in the figure legends.
The rice hypotonic culture and soil pot culture were basically
followed the protocols previously described by Li et al. (2014) and
Yang et al. (2014). Briefly, for hypotonic culture, 2-week-old
seedlings of similar size were transferred to IRRI nutrient solution
with specific treatment (indicated in figure legends) for additional
3 weeks. The nutrient concentrations (mM) of IRRI solution are as
follows: N 2.5, P 0.3, K 1, Ca 1, Mg 1, Si 0.5, Fe 0.02, B 0.02, Mn
0.009, Zn 0.00077, Cu 0.00032, Mo 0.00039. For salt-stressed
pot culture, 2-week-old seedlings were acclimated to the soil
containing 0.1% (1-g/kg soil) NaCl for 1 week before being
transplanted to the pot filled with 40 kg of soil which had been
fertilized and mixture with indicated extra amount of salts (0, 1.3
and 1.8-g/kg soil, respectively) for 2 weeks. For either K+ deficient
or N + P deficient soil culture, 4-week-old seedlings were
transplanted to the respective isolated paddy field and no extra
fertilizer was added during the growth season. The soil properties
used for the plant tolerance to salt stress, K+ deficiency or N+Pdeficiency were described in Table S2.
Cloning of HtNHX1 and HtNHX2 sequences
Both cDNAs of HtNHX1 and HtNHX2 were reversely transcribed
from the RNA extracted from the leaves of NY-1. Degenerate
primers (P1, P2) designed for cloning the Na+/H+ antiporters
(Table S1) were based on alignment of the sequences of yeast,
microbes, moss and plant Na+/H+ antiporters. The amplified
fragments were ligated into the pEasy-Blunt vector (Transgene,
Figure 10 Subcellular localization of HtNHX1 and HtNHX2 in rice
protoplast. Expression of HtNHX1, HtNHX2, HtNHX1del fused with GFP,
andAtTPK1 (a), AtHDEL (b)AtNHX5, AtSYP61, AtScamp (c) fusedwith RFP in
rice protoplast cells.. HtNHX1del: expression of mutated HtNHX1 by
deletion of conserved eight amino acids in NHX1 group from different plant
species (see Figure S1). AtTPK1, AtHDEL, AtNHX5, AtSYP61 and AtScamp
are the localization markers on tonoplast, endoplasmic reticulum, and
endosome or trans-Golgi network/Golgi, respectively, in Arabidopsis.
ª 2017 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd., 1–12
Rice tolerance to salinity and nutrient deficiency 9

China). Based on the sequences of this RT-PCR fragment, rapid-
amplification of cDNA ends (RACE) PCR was carried out using
SMARTer RACE cDNA amplification kit (Clontech, China) to
obtain the full-length cDNA sequences according to the instruc-
tion. First strand of 50- and 30-RACE-ready cDNA was synthesized
according to the kit manual with total RNA. The 30 and 50 RACE-PCRs were carried out with each pair of gene-specific and
universal primers using the first strand cDNA as the template. The
30 nested PCR was performed with the nested gene-specific
primer and nested universal primer A (provided with the kit) using
diluted 30 RACE-PCR product as template because the level of
background in the 30-RACES reaction was too high. The primers
used in RACE-PCR were designed based on the partial DNA
sequence: P3: (for 50 RACE-PCR), P4: (for 50 nested RACE-PCR),
P5: (for 30 RACE-PCR), and P6: (for 30 nested RACE-PCR). The PCR
product was purified and cloned into pEasy-Blunt vector and
sequenced. By aligning the sequences of the 50-end 30-end PCR
products and the previous partial DNA sequence, the full-length
cDNA sequence of HtNHX1 and HtNHX2 was deduced and
obtained by RT-PCR with KOD-PLUS (TOYOBO, Japan) using the
specific primers: P7 and P8 (Table S1).
HtNHX1 and HtNHX2 transcripts analysis
Total RNA was extracted from root or shoot of H. tuberosus
genotype NY-1 subjected to different salt stresses. Real-time qRT-
PCR analysis of HtNHX1 was performed as described by Li et al.
(2014). Semi-quantitative RT-PCR analysis for HtNHX2 was
performed according to the protocol described previously
(Tsuchiya et al., 2004; Yan et al., 2011). The target genes and
actin standards in 1 : 10, 1 : 100 and 1 : 1000 dilutions were
always present in the experiments. All the primers are listed in
Table S1. All the PCR products were checked by electrophoresis
and sequenced to confirm their identity.
Generating the transgenic rice expressing either HtNHX1or HtNHX2
The open reading frames of HtNXH1 and HtNXH2 were amplified
by gene-specific primers (see Table S1). The fragment was treated
with restriction enzymes and inserted into pTCK303 expression
vectors under the control of the Ubi-promoter. The final
constructed vector was sequenced before transformation. The
gene transformation into rice (Oryza sativa L. cv. Nipponbare) was
performed by Agrobacterium-mediated cocultivation method
(Upadhyaya et al., 2010; Li et al., 2014). Total 13 lines of
HtNHX1 and 17 lines of HtNHX2 expressed homozygotes were
initially obtained from T1 generation. By Southern blot analysis,
each five lines of HtNHX1 or HtNHX2 expressing rice plants with
only one copy of the gene insertion (Figure S2A) were selected
and propagated for getting the seeds of T2-T4 generations. The
exact integration sites of HtNHX1 or NtNHX2 in the rice genome
for HtNHX1.1-HtNHX1.2 and HtNHX2.1-HtNHX2.2 lines were
sequenced (Figure S2B), and their T3 generation was used for
further experiments.
Expression of HtNH1 or HtNHX2 gene in Arabidopsisnhx5 nhx6 double mutant
The HtNHX1- or HtNHX2-coding sequence was amplified and
built into the BamHI/SacI sites of the pTCK303 vector driven by
the ubiquitin promoter. The resulting constructs were introduced
into nhx5-2 nhx6-2 for the functional complementation by
Agrobacterium tumefaciens strain EHA105 using the floral
dipping method (Clough and Bent, 1998).
Southern blot and TAIL-PCR analysis
The independent transgenic lines with HtNHX1 or HtNHX2
expression lines, NHX1-1, NHX1-2 and NHX2-1, NHX2-2, were
determined by Southern blot analysis following the procedures
described previously (Yan et al., 2011). A TAIL-PCR procedure
was performed as described previously (Liu et al., 1995). The PCR
products were purified and then cloned into pMD-19 T vector
(TaKaRa Biotechnology Co., Ltd., Dalian, China) for sequencing.
Sequence identity was determined by blasting against the NCBI
database. All of the primers used for TAIL-PCR and procedures are
listed in Table S1.
HtNHX1 and HtNHX2 subcellular localization assay
Rice protoplasts were isolated according to Yang et al. (2014), C-
terminal or N-terminal GFP fused HtNHX1 or HtNHX2 was
constructed in the vector and transiently expressed in the rice
protoplasts and detected by laser scanning microscope (LSM410;
Carl Zeiss) as described by Yang et al. (2014). The separation of
the membrane fractions of BY2 cells was conducted according to
the bio-protocol of Membrane Preparation, Sucrose Density
Gradients and Two-phase Separation Fractionation from
5-day-old Arabidopsis seedlings (http://www.bio-protocol.org/
e1014) by Yang et al. (2013).
Determination total Na+, K+, N and P content in rice
All the samples were dried at 105 °C for 30 min and then kept at
70 °C in an oven for 72 h before weighing. After grinding, the
samples were extracted by 2 M HCl for 2 days before measuring
K+ and Na+ by a flame emission spectrometry. For N and P,
samples were digested completely with 98% H2SO4–30% H2O2
and measured as described by Tang et al. (2012) and Ai et al.
(2009).
Statistical analysis
Data were analysed by ANOVA using the SPSS 10 program (SPSS).
Different letters above columns in the figures indicate statistical
differences (Probability at 0.05) between individual transgenic
lines and wild-type and/or between different treatments.
Acknowledgements
This work was supported by the National R&D Program for
Transgenic Crops (Grant no. 2016ZX08009003005-008), National
Natural Science Foundation of China (Grant no. 31272226), The
National Key Program for Research and Development (Grant no.
2016YFD0100700) and International Potash Institute (IPI). We
thank Prof. Huixia Shou and Dr. Yinhui Ying from Zhejiang
University, Prof. Yiqun Bao for providing the localization markers,
Dr. Francisco Javier Quintero from Consejo Superior de Investiga-
ciones Cientificas for some critical comments of this work.
Conflict of interest
The authors declare no conflict of interest.
References
Ai, P., Sun, S.B., Zhao, J.N., Fan, X.R., Xin, W.J., Guo, Q., Yu, L. et al. (2009)
Two rice phosphate transporters, ORYsa;Pht1;2 and ORYsa;Pht1;6, have
different functions and kinetic properties in uptake and translocation. Plant J.
57, 798–809.
ª 2017 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd., 1–12
Yang Zeng et al.10

Alem�an, F., Nieves-Cordones, M., Mart�ınez, V. and Rubio, F. (2011) Root K(+)
acquisition in plants: the Arabidopsis thaliana model. Plant Cell Physiol. 52,
1603–1612.
Andr�es, Z., P�erez-Hormaeche, J., Leidi, E.O., Schl€ucking, K., Steinhorst, L.,
McLachlan, D.H., Schumacher, K. et al. (2014) Control of vacuolar dynamics
and regulation of stomatal aperture by tonoplast potassium uptake. Proc.
Natl Acad. Sci. USA, 111, E1806–E1814.
Apse, M.P., Aharon, G.S., Snedden, W.A. and Blumwald, E. (1999) Salt
tolerance conferred by overexpression of a vacuolar Na+/H+ antiport in
Arabidopsis. Science, 285, 1256–1258.
Apse, M.P., Sottosanto, J.B. and Blumwald, E. (2003) Vacuolar cation/H+
exchange, ion homeostasis, and leaf development are altered in a T-DNA
insertional mutant of AtNHX1, the Arabidopsis vacuolar Na+/H+ antiporter.
Plant J. 36, 229–239.
Baral, A., Shruthi, K.S. and Mathew, M.K. (2015) Vesicular trafficking and
salinity responses in plants. IUBMB Life, 67, 677–686.
Barrag�an, V., Leidi, E.O., Andr�es, Z., Rubio, L., De Luca, A., Fern�andez, J.A.,
Cubero, B. et al. (2012) Ion exchangers NHX1 and NHX2 mediate active
potassium uptake into vacuoles to regulate cell turgor and stomatal function
in Arabidopsis. Plant Cell, 24, 1127–1142.
Bassil, E. and Blumwald, E. (2014) The ins and outs of intracellular ion
homeostasis: NHX-type cation/H+ antiporters. Curr. Opin. Plant Biol. 22, 1–6.
Bassil, E., Ohto, M.A., Esumi, T., Tajima, H., Zhu, Z., Cagnac, O., Belmonte, M.
et al. (2011a) The Arabidopsis intracellular Na+/H+ antiporters NHX5 and
NHX6 are endosome associated and necessary for plant growth and
development. Plant Cell, 23, 224–239.
Bassil, E., Tajima, H., Liang, Y., Ohto, M., Ushijima, K., Nakano, R., Esumi, T.
et al. (2011b) The Arabidopsis Na+/H+ antiporters NHX1 and NHX2 control
vacuolar pH and K+ homeostasis to regulate growth, flower development,
and reproduction. Plant Cell, 23, 3482–3497.
Bassil, E., Coku, A. and Blumwald, E. (2012) Cellular homeostasis: emerging
roles of intracellular NHX Na+/H+ antiporters in plant growth and
development. J. Exp. Bot. 3, 5727–5740.
Blumwald, E. and Poole, R.J. (1985) Na+/H+ antiport in isolated tonoplast
vesicles from storage tissue of Beta vulgaris. Plant Physiol., 78, 163–167.
Chanroj, S., Wang, G., Venema, K., Zhang, M.W., Delwiche, C.F. and Sze, H.
(2012) Conserved and diversified gene families of monovalent cation/H+
antiporters from algae to flowering plants. Front. Plant Sci. 3, 1–18.
Clough, S.J. and Bent, A.F. (1998) Floral dip: a simplified method for
Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J.
16, 735–743.
Cuin, T.A., Miller, A.J., Laurie, S.A. and Leigh, R.A. (2003) Potassium activities in
cell compartments of salt-grown barley leaves. J. Exp. Bot. 54, 657–661.
Deinlein, U., Stephan, A.B., Horie, T., Luo, W., Xu, G.H. and Schroeder, J.I.
(2014) Plant salt-tolerance mechanisms. Trends Plant Sci. 19, 371–379.
Drew, M.C. and Saker, L.R. (1984) Uptake and long-distance transport of
phosphate, potassium and chloride in relation to internal ion concentrations
in barley: evidence of non-allosteric regulation. Planta, 160, 500–507.
Drew, M.C., Webb, J. and Saker, L.R. (1990) Regulation of K+ uptake and
transport to the xylem in barley roots: K+ distribution determined by electron
probe X-ray microanalysis of frozen-hydrated cells. J. Exp. Bot. 41, 815–825.
Engels, C. and Marschner, H. (1992) Adaptation of potassium translocation into
the shoot of maize (Zea mays) to shoot demand: evidence for xylem loading
as a regulating step. Physiol. Plant. 86, 263–268.
Fukuda, A., Nakamura, A., Hara, N., Toki, S. and Tanaka, Y. (2011) Molecular
and functional analyses of rice NHX-type Na+/H+ antiporter genes. Planta,
233, 175–188.
Gaxiola, R.A., Rao, R., Sherman, A., Grisafi, P., Alper, S.L. and Fink, G.R. (1999)
The Arabidopsis thaliana transporters, AtNHX1 and Avp1, can function in
cation detoxification in yeast. Proc. Natl Acad. Sci. USA, 96, 1480–1485.
Gaxiola, R.A., Li, J., Undurraga, S., Dang, L.M., Allen, G.J., Alper, S.L. and Fink,
G.R. (2001) Drought- and salt-tolerant plants result from overexpression of
the AVP1 H1-pump. Proc. Natl Acad. Sci. USA, 98, 11444–11449.
Hamaji, K., Nagira, M., Yoshida, K., Ohnishi, M., Oda, Y., Uemura, T., Goh, T.
et al. (2009) Dynamic aspects of ion accumulation by vesicle traffic under salt
stress in Arabidopsis. Plant Cell Physiol. 50, 2023–2033.
Heuer, S., Gaxiola, R., Schilling, R., Herrera-Estrella, L., L�opez-Arredondo, D.,
Wissuwa, M., Delhaize, E. et al. (2016) Improving phosphorus use efficiency
—a complex trait with emerging opportunities. Plant J. 90, 868–885.
https://doi.org/10.1111/tpj.13423.
Horie, T., Hauser, F. and Schroeder, J.I. (2009) HKT transporter-mediated
salinity resistance mechanisms in Arabidopsis and monocot crop plants.
Trends Plant Sci. 14, 660–668.
Jiang, X., Leidi, E.O. and Pardo, J.M. (2010) How do vacuolar NHX exchangers
function in plant salt tolerance? Plant Signal Behav. 5, 792–795.
Johansson, E., Prade, T., Angelidaki, I., Svensson, S.E., Newson, W.R.,
Gunnarsson, I.B. and Hovmalm, H.P. (2015) Economically viable
components from Jerusalem artichoke (Helianthus tuberosus L.) in a
biorefinery concept. Int. J. Mol. Sci. 16, 8997–9016.
Julkowska, M.M. and Testerink, C. (2015) Tuning plant signaling and growth to
survive salt. Trends Plant Sci. 20, 586–594.
Leidi, E.O., Barragan, V., Rubio, L., El-Hamdaoui, A., Ruiz, M.T., Cubero, B.,
Fernandez, J.A. et al. (2010) The AtNHX1 exchanger mediates potassium
compartmentation in vacuoles of transgenic tomato. Plant J. 61, 495–
506.
Leshem, Y., Melamed-Book, N., Cagnac, O., Ronen, G., Nishri, Y., Solomon, M.,
Cohen, G. et al. (2006) Suppression of Arabidopsis vesicle-SNARE expression
inhibited fusion of H2O2-containing vesicles with tonoplast and increased salt
tolerance. Proc. Natl Acad. Sci. USA, 103, 18008–18013.
Li, J.Y., He, X.W., Xu, L., Zhou, J., Wu, P., Shou, H.X. and Zhang, F.C. (2008)
Molecular and functional comparisons of the vacuolar Na+/H+ exchangers
originated from glycophytic and halophytic species. J. Zhejiang Univ. Sci. B, 9,
132–140.
Li, Q., Tang, Z., Hu, Y., Yu, L., Liu, Z. and Xu, G.H. (2014) Functional analyses of
a putative plasma membrane Na+/H+ antiporter gene isolated from salt
tolerant Helianthus tuberosus. Mol. Biol. Rep. 41, 5097–5108.
Liu, Y.G., Mitsukawa, N., Oosumi, T. and Whittier, R.F. (1995) Efficient isolation
and mapping of Arabidopsis thaliana T-DNA insert junctions by thermal
asymmetric interlaced PCR. Plant J. 8, 457–463.
Long, X.H., Mehta, S. and Liu, Z.P. (2008) Effect of NO3–N enrichment on
seawater stress tolerance of Jerusalem artichoke (Helianthus tuberosus).
Pedosphere, 18, 113–123.
Lu, Y.X., Li, C.J. and Zhang, F.S. (2005) Transpiration, potassium uptake and
flow in tobacco as affected by nitrogen forms and nutrient levels. Ann. Bot.
95, 991–998.
McCubbin, T., Bassil, E., Zhang, S. and Blumwald, E. (2014) Vacuolar Na+/H+
antiporters are required for cellular K+ homeostasis, microtubule organization
and directional root growth. Plants, 3, 409–426.
Mimura, T., Kura-Hotta, M., Tsujimura, T., Ohnishi, M., Miura, M., Okazaki, Y.,
Mimura, M. et al. (2003) Rapid increase of vacuolar volume in response to
salt stress. Planta, 216, 397–402.
Munns, R. and Gilliham, M. (2015) Salinity tolerance of crops—what is the cost?
New Phytol. 208, 668–673.
Munns, R. and Tester, M. (2008) Mechanisms of salinity tolerance. Annu. Rev.
Plant Biol. 59, 651–681.
Peuke, A.D. (2010) Correlations in concentrations, xylem and phloem flows,
and partitioning of elements and ions in intact plants. A summary and
statistical re-evaluation of modelling experiments in Ricinus communis. J. Exp.
Bot. 61, 635–655.
Qi, Z. and Spalding, E.P. (2004) Protection of plasma membrane K+ transport by
the salt overly sensitive1 Na+-H+ antiporter during salinity stress. Plant
Physiol. 136, 2548–2555.
Qiu, Q.S. and Fratti, R.A. (2010) The Na+/H+ exchanger Nhx1p regulates the
initiation of Saccharomyces cerevisiae vacuole fusion. J. Cell Sci. 123, 3266–
3275.
Reguera, M., Bassil, E. and Blumwald, E. (2014) Intracellular NHX-type cation/
H+ antiporters in plants. Mol. Plant. 7, 261–263.
Shabala, S., Bose, J., Fuglsang, A.T. and Pottosin, I. (2016) On a quest for stress
tolerance genes: membrane transporters in sensing and adapting to hostile
soils. J. Exp. Bot. 67, 1015–1031.
Tang, Z., Fan, X.R., Li, Q., Feng, H.M., Miller, A.J., Shen, Q.R. and Xu, G.H.
(2012) Knock down of a rice stelar nitrate transporter alters long distance
translocation but not root influx. Plant Physiol. 160, 2052–2063.
Tsuchiya, T., Takesawa, T., Kanzaki, H. and Nakamura, I. (2004) Genomic
structure and differential expression of two tandem-arranged GSTZ genes in
rice. Gene, 335, 141–149.
ª 2017 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd., 1–12
Rice tolerance to salinity and nutrient deficiency 11

Upadhyaya, N.M., Surin, B., Ramm, K., Gaudron, J., Schunmann, P.H.D., Taylor,
W., Waterhouse, P.M., et al. (2000) Agrobacterium-mediated transformation
of Australian rice cultivars Jarrah and Amaroo using modified promoters and
selectable markers. Aust. J. Plant Physiol. 27, 201–210.
Venema, K., Belver, A., Marin-Manzano, M.C., Rodr�ıguez-Rosales, M.P. and
Donaire, J.P. (2003) A novel intracellular K+/H+ antiporter related to Na+/H+
antiporters is important for K+ ion homeostasis in plants. J. Biol. Chem. 278,
22453–22459.
Walker, D.J., Leigh, R.A. and Miller, A.J. (1996) Potassium homeostasis in
vacuolate plant cells. Proc. Natl Acad. Sci. USA, 93, 10510–10514.
Xu, K., Zhang, H., Blumwald, E. and Xia, T. (2010) A novel plant vacuolar Na+/
H+ antiporter gene evolved by DNA shuffling confers improved salt tolerance
in yeast. J. Biol. Chem. 285, 22999–23006.
Xu, G.H., Fan, X.R. and Miller, A.J. (2012) Plant nitrogen assimilation and use
efficiency. Annu. Rev. Plant Biol. 63, 153–182.
Yamaguchi, T., Apse,M.P., Shi, H. and Blumwald, E. (2003) Topological analysis of
a plant vacuolar Na+/H+ antiporter reveals a luminal C terminus that regulates
antiporter cation selectivity. Proc. Natl Acad. Sci. USA, 100, 12510–12515.
Yamaguchi, T., Aharon, G.S., Sottosanto, J.B. and Blumwald, E. (2005)
Vacuolar Na+/H+ antiporter cation selectivity is regulated by calmodulin
from within the vacuole in a Ca2 + - and pH-dependent manner. Proc. Natl
Acad. Sci. USA, 102, 16107–16112.
Yan, M., Fan, X.R., Feng, H.M., Miller, A.J., Shen, Q.R. and Xu, G.H. (2011) Rice
OsNAR2.1 interacts with OsNRT2.1, OsNRT2.2 and OsNRT2.3a nitrate
transporters to provide uptake over high and low concentration ranges.
Plant Cell Environ. 34, 1360–1372.
Yang, H., Richter, G.L., Wang, X., Mlodzinska, E., Carraro, N., Ma, G., Jenness,
M., et al. (2013) Sterols and sphingolipids differentially function in trafficking
of the Arabidopsis ABCB19 auxin transporter. Plant J. 74, 37–47.
Yang, T.Y., Zhang, S., Hu, Y.B., Hu, F.C., Hu, Q.D., Chen, G., Cai, J. et al.
(2014) The role of a potassium transporter OsHAK5 in potassium acquisition
and transport from roots to shoots in rice at low potassium supply levels.
Plant Physiol. 166, 945–959.
Yokoi, S., Quintero, F.J., Cubero, B., Ruiz, M.T., Bressan, R.A., Hasegawa, P.M.
and Pardo, J.M. (2002) Differential expression and function of Arabidopsis
thaliana NHX Na+/H+ antiporters in the salt stress response. Plant J. 30, 529–
539.
Zhang, H.X. and Blumwald, E. (2001) Transgenic salt-tolerant tomato plants
accumulate salt in foliage but not in fruit. Nat. Biotechnol. 19, 765–768.
Supporting information
Additional Supporting Information may be found online in the
supporting information tab for this article:
Figure S1 Bioinformatic prediction of HtNHX1 and HtNHX2
structures.
Figure S2 Isolation of single copy inserted rice lines of HtNHX1 or
HtNHX2 expression.
Figure S3 Effects of overexpression of HtNHX1 and HtNHX2 on
grain yield, harvest index, and NPK concentrations in the rice
grown in low K supplied soil.
Figure S4 Effects of HtNHX1 and HtNHX2 expression on
ammonium N acquisition and distribution under low N and K
supply condition.
Figure S5 HtNHX1, but not HtNHX2, functioned in enhancing
tolerance to external hygromycin in both nhx1 mutated yeast cells
(A) and Arabidopsis (B).
Table S1 The primers used in the experiments.
Table S2 The properties of the soil used in the experiments.
ª 2017 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd., 1–12
Yang Zeng et al.12
Related Documents











