Two new cestode species of Neoskrjabinolepis Spasskii, 1947 (Cyclophyllidea: Hymenolepididae) from the tundra shrew Sorex tundrensis Merriam (Mammalia: Soricidae) in Alaska and Chukotka Svetlana A. Kornienko • Nikolaj E. Dokuchaev Received: 27 March 2012 / Accepted: 23 June 2012 Ó Springer Science+Business Media B.V. 2012 Abstract Descriptions, illustrations and differential diagnoses for two new species of Neoskrjabinolepis Spassky, 1947, i.e. N. (Neoskrjabinolepis) fertilis n. sp. and N. (Neoskrjabinolepidoides) hobergi n. sp., are given. Neoskrjabinolepis (Neoskrjabinolepis) fertilis was found in the shrew Sorex tundrensis Merriam on the Seward Peninsula (Alaska, USA) and from the mouth of the Anadyr River (Chukotka, Russia), and N. (Neoskrjabinolepidoides) hobergi was found in Sorex tundrensis on the Seward Peninsula (Alaska, USA). Neoskrjabinolepis fertilis n. sp. is characterised by having: rostellar hooks 38–42 lm long and provided with a small epiphyseal thickening of the handle; a long cirrus (85–100 lm), consisting of a basal region with claw-shaped spines and a parabasal region with small, thin needle-shaped spines; a cirrus- sac extending well into the median field; and 55–70 eggs per gravid uterus. Neoskrjabinolepis (Neoskrja- binolepidoides) hobergi n. sp. is characterised by having: rostellar hooks 63–65 lm long and provided with a large epiphyseal thickening of the handle; a short cirrus (45–50 lm), consisting of a basal region with small claw-shaped spines and a parabasal region with thin, needle-shaped spines; cirrus-sac reaching slightly into the median field; and 36–45 eggs per gravid uterus. Introduction The last checklist of the helminth parasites of shrews from North America by Kinsella & Tkach (2009) included 34 species of cestodes (including larvae). However, only 18 cestode species of 11 genera are currently known for shrews of the genus Sorex L. in Alaska (Rausch & Kuns, 1950; Voge & Rausch, 1955; Dunagan, 1956; Olsen, 1969; Gulyaev, 2003; Melnik- ova et al., 2003; Dokuchaev & Gulyaev, 2007; Gulyaev et al., 2007, 2009; Kinsella, 2007; Kinsella et al., 2008; Kinsella & Tkach, 2009). More recently, the cestode Spasskylepis rauschi Gulyaev, Dokuchaev & Lykova, 2010 has been found in Alaskan specimens of Sorex cinereus Kerr (Gulyaev et al., 2010). Thus, 10 of the 12 recorded genera are common for Eurasia and North America and only two, i.e. Lockerraushia Yamaguti, 1959 and Vogelepis Vaucher in Czaplinski & Vaucher, 1994, are North-American endemics. The latter is morphologically similar to Neoskrjabinol- epis Spassky, 1947, but Neoskrjabinolepis spp. have recently been considered to parasitise only Palaearctic shrews and were thought not to occur in the Nearctic S. A. Kornienko (&) Institute of Systematics and Ecology of Animals, Siberian Branch, Russian Academy of Sciences, 11 Frunze Street, Novosibirsk 630091, Russia e-mail: [email protected] N. E. Dokuchaev Institute of Biological Problems of the North, Far-East Branch, Russian Academy of Sciences, 18 Portovaja Street, Magadan 685000, Russia e-mail: [email protected] 123 Syst Parasitol (2012) 83:179–188 DOI 10.1007/s11230-012-9383-6

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Two new cestode species of Neoskrjabinolepis Spasskii, 1947(Cyclophyllidea: Hymenolepididae) from the tundra shrewSorex tundrensis Merriam (Mammalia: Soricidae) in Alaskaand Chukotka
Svetlana A. Kornienko • Nikolaj E. Dokuchaev
Received: 27 March 2012 / Accepted: 23 June 2012
� Springer Science+Business Media B.V. 2012
Abstract Descriptions, illustrations and differential
diagnoses for two new species of Neoskrjabinolepis
Spassky, 1947, i.e. N. (Neoskrjabinolepis) fertilis n.
sp. and N. (Neoskrjabinolepidoides) hobergi n. sp., are
given. Neoskrjabinolepis (Neoskrjabinolepis) fertilis
was found in the shrew Sorex tundrensis Merriam on
the Seward Peninsula (Alaska, USA) and from the
mouth of the Anadyr River (Chukotka, Russia), and
N. (Neoskrjabinolepidoides) hobergi was found in
Sorex tundrensis on the Seward Peninsula (Alaska,
USA). Neoskrjabinolepis fertilis n. sp. is characterised
by having: rostellar hooks 38–42 lm long and
provided with a small epiphyseal thickening of the
handle; a long cirrus (85–100 lm), consisting of a
basal region with claw-shaped spines and a parabasal
region with small, thin needle-shaped spines; a cirrus-
sac extending well into the median field; and 55–70
eggs per gravid uterus. Neoskrjabinolepis (Neoskrja-
binolepidoides) hobergi n. sp. is characterised by
having: rostellar hooks 63–65 lm long and provided
with a large epiphyseal thickening of the handle; a
short cirrus (45–50 lm), consisting of a basal region
with small claw-shaped spines and a parabasal region
with thin, needle-shaped spines; cirrus-sac reaching
slightly into the median field; and 36–45 eggs per
gravid uterus.
Introduction
The last checklist of the helminth parasites of shrews
from North America by Kinsella & Tkach (2009)
included 34 species of cestodes (including larvae).
However, only 18 cestode species of 11 genera are
currently known for shrews of the genus Sorex L. in
Alaska (Rausch & Kuns, 1950; Voge & Rausch, 1955;
Dunagan, 1956; Olsen, 1969; Gulyaev, 2003; Melnik-
ova et al., 2003; Dokuchaev & Gulyaev, 2007;
Gulyaev et al., 2007, 2009; Kinsella, 2007; Kinsella
et al., 2008; Kinsella & Tkach, 2009). More recently,
the cestode Spasskylepis rauschi Gulyaev, Dokuchaev
& Lykova, 2010 has been found in Alaskan specimens
of Sorex cinereus Kerr (Gulyaev et al., 2010). Thus, 10
of the 12 recorded genera are common for Eurasia and
North America and only two, i.e. Lockerraushia
Yamaguti, 1959 and Vogelepis Vaucher in Czaplinski
& Vaucher, 1994, are North-American endemics. The
latter is morphologically similar to Neoskrjabinol-
epis Spassky, 1947, but Neoskrjabinolepis spp. have
recently been considered to parasitise only Palaearctic
shrews and were thought not to occur in the Nearctic
S. A. Kornienko (&)
Institute of Systematics and Ecology of Animals, Siberian
Branch, Russian Academy of Sciences, 11 Frunze Street,
Novosibirsk 630091, Russia
e-mail: [email protected]
N. E. Dokuchaev
Institute of Biological Problems of the North,
Far-East Branch, Russian Academy of Sciences,
18 Portovaja Street, Magadan 685000, Russia
e-mail: [email protected]
123
Syst Parasitol (2012) 83:179–188
DOI 10.1007/s11230-012-9383-6

(Gulyaev et al., 2009). However, in helminthological
samples from shrews in Alaska, cestodes correspond-
ing morphologically to the genus Neoskrjabinolepis
have been found (Kornienko & Gulyaev, 2008).
For many years, only two species of Neoskrjabi-
nolepis were recognised: N. schaldybini Spassky, 1947
and N. singularis (Cholodkowsky, 1912) Spassky,
1954 (Spassky, 1947, 1954; Czaplinski & Vaucher
1994). Sometimes, N. schaldybini has been considered
a junior synonym of N. singularis (Kobuley, 1953;
Zarnowski, 1955; Kisielewska, 1958; Prokopic, 1956,
1958; Pojmanska, 1957; Rybicka, 1959), but its
validity was supported by Spassky (1954), Vaucher
(1971) and Genov (1984). Recent studies have been
undertaken on cestodes from Sorex spp. belonging to
Neoskrjabinolepis originating from various parts of the
Palaearctic Region. These studies have resulted in
redescriptions of the two known forms and the
description of a further 10 species (Kornienko et al.,
2006, 2007, 2008, 2010). Neoskrjabinolepis now
includes two subgenera whose species differ in the
pattern of their strobilar development: Neoskrjabinol-
epis (Neoskrjabinolepis) Spasskii, 1947, with gradual
strobilar maturation, and Neoskrjabinolepis (Neoskrja-
binolepidoides) Kornienko, Gulyaev & Mel’nikova,
2006, with strobila consisting of a series of proglottides
at the same developmental stage (Kornienko et al.,
2006). Both types of strobilar organisation are recorded
in cestodes found in Sorex tundrensis Merriam from
Alaska. The aim of the present article is to describe two
new species, one from each of these two subgenera.
Materials and methods
Cestodes were collected from the intestine of shrews
Sorex tundrensis caught on the Seward Peninsula
(Alaska, USA) and in the mouth of Anadyr River
(Chukotka, Russia) during the summers of 2001 and
2002. Host specimens were dissected immediately
after their death. Live cestodes were isolated, washed,
relaxed in water and fixed in 70% ethanol. They were
stained in Ehrlich’s haematoxylin, differentiated in a
3% aqueous solution of ferric ammonium-sulphate-
12-hydrate, dehydrated in an ethanol series, cleared in
clove oil and mounted in Canada balsam. Some
specimens were mounted in Berlese’s medium to
facilitate examination of the rostellar hooks and
copulatory apparatus. Type-specimens are deposited
in the collection of Zoology Museum of the Institute of
Systematics and Ecology of Animals, Novosibirsk,
Russia (ISEA) and in the Natural History Museum of
Geneva, Switzerland (MHNG).
In order to establish the stage of proglottid matu-
ration, the terminology of Mas-Coma & Galan-
Puchades (1991) is used. Measurements are given in
micrometres except where otherwise indicated. They
are presented as the range followed by the mean and
the number of the measurements (n) in parentheses.
Neoskrjabinolepis (Neoskrjabinolepis) fertilis n. sp.
Type-host: Sorex tundrensis Merriam (Mammalia:
Soricidae), the tundra shrew.
Type-locality: Seward Peninsula, Alaska, USA;
65�519.120N, 164�4145.360W.
Other locality: Vicinity of Dionisija Mountain, Chuk-
otka, Russia; 64�344.490N, 177�1848.120E; vicinity of
Shachterskii Village, Chukotka, Russia, 64�4845.420N,
177�3254.310E.
Type-material: Holotype: ISEA 18.11.6.1, collected on
10 August 2001. Paratypes: 14 slides, ISEA
18.11.6.2–18.11.6.5, 18.11.6.7–18.11.13, Seward Pen-
insula, Alaska, USA, collected on 10 August. 2001;
ISEA 18.11.6.14, vicinity of Dionisija Mountain,
Chukotka, Russia, collected on 29 July 2002; ISEA
18.11.6.15, vicinity of the Shachterskii Village, Chuk-
otka, Russia, 3 August 2002; MHNG INVE 82149,
Seward Peninsula, Alaska, USA, collected on 10
August 2001.
Etymology: The species name is derived from the
Latin fertilis (=fertile) and refers to the great number
of eggs in the gravid proglottides of this species.
Description (Figs. 1, 2)
Gravid specimens 8.0–10.0 mm (8.9 mm, n = 10)
long; strobila consists of 190–220 (200, n = 10)
proglottides. Strobilation gradual. Premature and
mature proglottides with developing uterus wider than
long; pregravid and gravid proglottides almost equal
in width and length. Internal formation of proglottides
distinct before external segmentation is apparent;
external segmentation appears at level of postmature
proglottides.
Scolex 250–290 wide (260, n = 7), distinctly wider
than neck (Fig. 1A). Suckers round, 100–130 100–110
180 Syst Parasitol (2012) 83:179–188
123
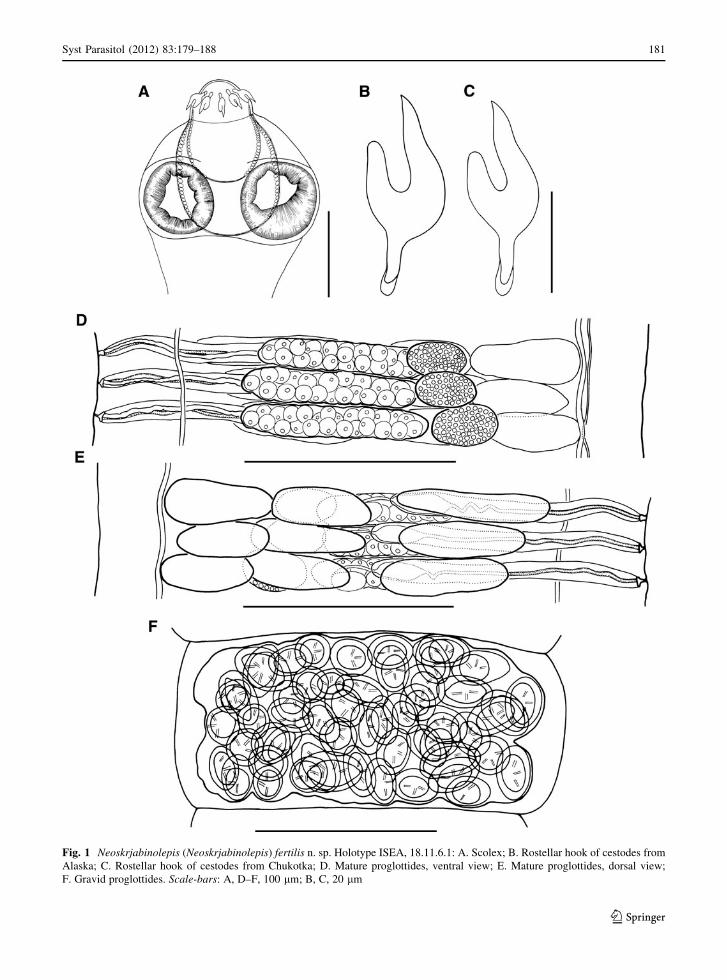
Fig. 1 Neoskrjabinolepis (Neoskrjabinolepis) fertilis n. sp. Holotype ISEA, 18.11.6.1: A. Scolex; B. Rostellar hook of cestodes from
Alaska; C. Rostellar hook of cestodes from Chukotka; D. Mature proglottides, ventral view; E. Mature proglottides, dorsal view;
F. Gravid proglottides. Scale-bars: A, D–F, 100 lm; B, C, 20 lm
Syst Parasitol (2012) 83:179–188 181
123
(95 9 90, n = 7), with well-developed musculature.
Rostellar apparatus complex. Rhynchus short, 62–65
long, 78–87 wide. Rostellum sac-like, 125–140 9
100–110 (121 9 72, n = 7); its walls consist of
external layer of longitudinal muscular fibres and
internal layer of circular muscle bundles. Rostellar
hooks 10 in number, arranged in single row, 38–42
(41, n = 15) long, with characteristic pincer-like
shape; claw-like blade longer than guard and axis of
blade almost parallel to axis of guard; blade with
curved middle region; handle provided with small
epiphyseal thickening (Fig. 1B, C). When rostellar
apparatus retracted, rostellar hooks with blades
directed anteriorly. Rostellar pouch voluminous,
140–145 9 110–140 (144 9 129, n = 7), reaches
beyond level of posterior margins of suckers; its wall
consists of longitudinal muscular fibres and circular
muscular bundles.
Proglottides acraspedote. Mature proglottides
15–17 9 230–260 (15 9 246, n = 10) [as external
segmentation appears only at level of postmature
proglottides, length of mature proglottides is measured
as distance between genital atria of adjacent proglot-
tides] (Fig. 1D), with lateral fields 37–38 wide; gonads
densely situated in median field, with Neoskrjabino-
lepidoides-type organisation (as defined by Gulyaev &
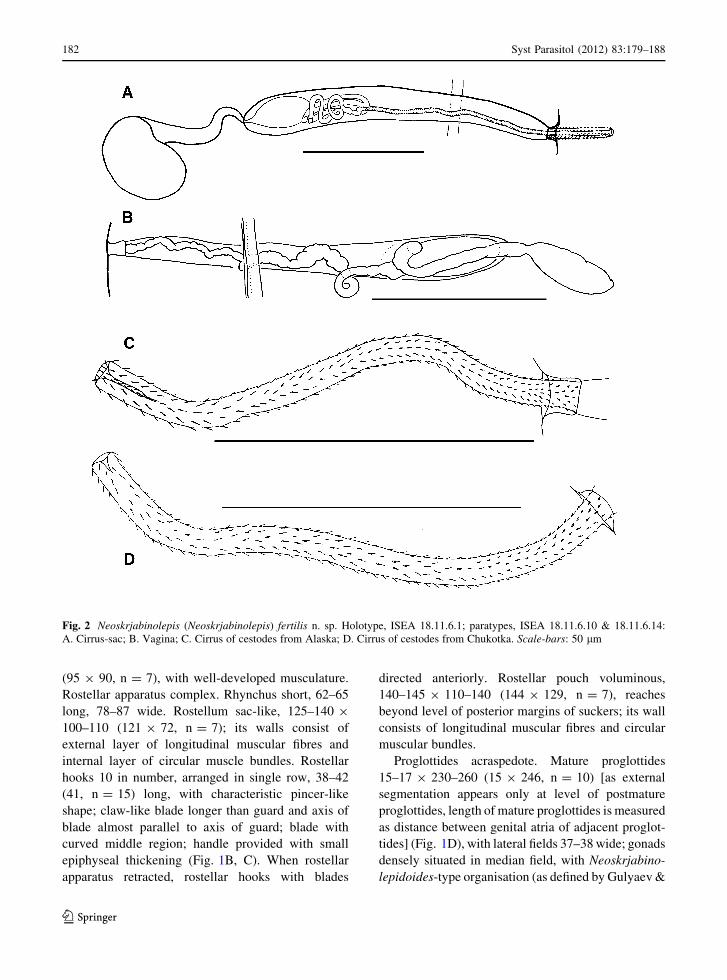
Fig. 2 Neoskrjabinolepis (Neoskrjabinolepis) fertilis n. sp. Holotype, ISEA 18.11.6.1; paratypes, ISEA 18.11.6.10 & 18.11.6.14:
A. Cirrus-sac; B. Vagina; C. Cirrus of cestodes from Alaska; D. Cirrus of cestodes from Chukotka. Scale-bars: 50 lm
182 Syst Parasitol (2012) 83:179–188
123

Kornienko, 2009). Gravid proglottides 180–210 9
280–300 (200 9 290, n = 10). Osmoregulatory canals
arranged in pairs, without transverse anastomoses;
ventral canals 4–6 wide; dorsal canals 1–2 wide.
Genital pores unilateral, dextral. Genital atrium sim-
ple, 5–6 deep, 5–6 in diameter.
Testes 3, of almost equal size, 15–23 9 34–50
(20 9 45, n = 12), situated in line; poral testis
separated from 2 antiporal testes by external seminal
vesicle (Fig. 1E). Diameter of testes greater than
proglottis length and, consequently, dense dorsal
testicular field formed in mature region of strobila.
Cirrus-sac cigar-shaped, 100–130 9 12–15 (120 9 13,
n = 12), slightly winding, passes deeply into median
field but does not reach median line of proglottis
(Fig. 2A). Cirrus long, 85–100 (93, n = 8), cylindri-
cal, armed with small, thin, needle-shaped spines
along its entire length; its basal region armed with
small, claw-shaped spines (Fig. 2C, D). Ductus ejac-
ulatorius forms several coils. Internal seminal vesicle
small, 25–35 9 10–12 (28 9 10, n = 12), occupies
less than quarter of cirrus-sac length even when filled.
External seminal vesicle 32–35 9 15–17 (33 9 17,
n = 10), connected to cirrus-sac by long, narrow
duct.
Vitellarium subspherical, 25–28 9 15–17 (26 9
17, n = 11), situated antiporally to ovary. Ovary
transversely elongate, compact, 10–15 9 90–110
(14 9 103, n = 11), situated in poral half of median
field, overlaps cirrus-sac, external seminal vesicle and
testes ventrally (Fig. 1D). Vagina thin-walled, passes
ventrally to cirrus-sac. Conductive part of vagina
gradually enlarges and passes into small, sac-like
seminal receptacle (Fig. 2B).
Uterus not extending into lateral fields. Uterine wall
thin, membranous throughout morphogenesis of uterus.
Number of eggs in uterus 55–70 (Fig. 1F).
Neoskrjabinolepis (Neoskrjabinolepidoides) hobergi
n. sp
Type-host: Sorex tundrensis Merriam (Mammalia:
Soricidae), the tundra shrew.
Type-locality: Seward Peninsula, Alaska, USA;
65�519.120N, 164�4145.360W.
Type-material: Holotype: ISEA 18.11.19.1, collected
on 7 August 2001. The holotype and another specimen
belonging to the same species are mounted on one
slide. The specimen marked N 1 was chosen as the
holotype. Paratypes 4: MHNG INVE 82150, ISEA
18.11.19.3–18.11.19.5; collected on 7 August 2001.
Etymology: This species is named for Dr Eric P.
Hoberg in recognition of his contributions to
helminthology.
Description (Figs. 3, 4)
Gravid specimens 8.0–10.0 mm (8.6 mm, n = 4)
long; strobila consists of 480–520 (510, n = 4)
proglottides. Strobilation serial; pregravid or gravid
strobila usually consisting of 4 series of proglottides,
each containing proglottides at same developmental
stage (first series of juvenile or premature proglottides;
second section of mature proglottides; third series of
postmature proglottides; fourth series of pregravid or
gravid proglottides); each series consists of c.110–130
proglottides. Strobilar regions containing juvenile,
premature or mature proglottides without external
segmentation; proglottides externally distinct at level
of postmature region of strobila.
Scolex 300–320 wide (316, n = 4), clearly wider
than neck (Fig. 3A). Suckers round, 100–110 9
100–110 (105 9 100, n = 4), with well-developed
musculature. Rostellar apparatus complex. Rhynchus
short, 65–66 long, 100–110 wide. Rostellum sac-like,
120–130 9 100–110 (121 9 72, n = 4); its walls
consist of external layer of longitudinal muscular fibres
and internal layer of circular muscular bundles.
Rostellar hooks 10 in number, arranged in single row,
63–66 (64, n = 5) long, with characteristic pincer-like
shape; blade twice length of handle and guard
(Fig. 3B), curved medially; handle provided with
large epiphyseal thickening. When rostellar appara-
tus retracted, blades of rostellar hooks are directed
anteriorly. Rostellar pouch voluminous, 180–190 9
170–180 (186 9 173, n = 5), reaches beyond level of
posterior margins of suckers; its wall consists of
longitudinal muscular fibres and circular muscular
bundles. Neck 160–180 (173, n = 4) wide.
Proglottides acraspedote, transversely elongate.
Mature proglottides 11–13 9 310–350 (11 9 336,
n = 4) (Fig. 3C) [as external segmentation appears
only at level of postmature proglottides, length of
mature proglottides is measured as distance between
genital atria of adjacent proglottides], with length/
width ratio 1:26–28; lateral fields 50–60 wide; gonads
densely situated in median field. Gravid proglottides
Syst Parasitol (2012) 83:179–188 183
123
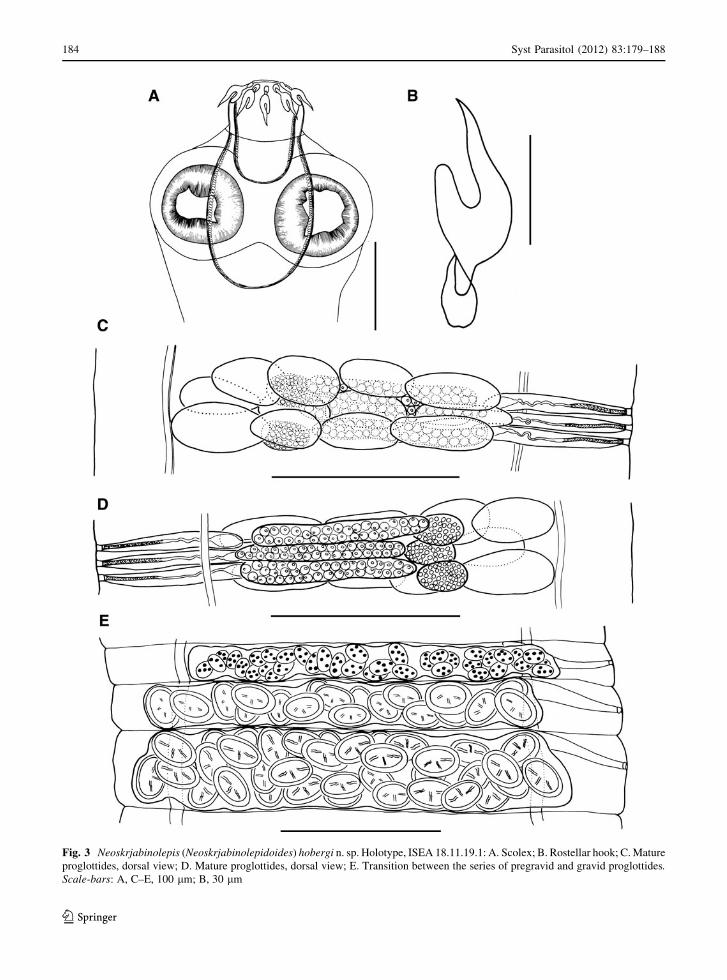
Fig. 3 Neoskrjabinolepis (Neoskrjabinolepidoides) hobergi n. sp. Holotype, ISEA 18.11.19.1: A. Scolex; B. Rostellar hook; C. Mature
proglottides, dorsal view; D. Mature proglottides, dorsal view; E. Transition between the series of pregravid and gravid proglottides.Scale-bars: A, C–E, 100 lm; B, 30 lm
184 Syst Parasitol (2012) 83:179–188
123

20–22 9 330–340. Osmoregulatory canals 2 pairs,
without transverse anastomoses; ventral canals 4–6
wide; dorsal canals 2–3 wide. Genital pores unilateral.
Genital atrium simple, 7–8 deep, 3–4 in diameter.
Testes 3, of almost equal size, 20–30 9 40–54
(20 9 35, n = 8), situated in line; poral testis sepa-
rated from 2 antiporal testes by external seminal
vesicle (Fig. 3C). Diameter of testes larger than
proglottis length and, consequently, dense dorsal
testicular field is formed in mature strobilar region.
Cirrus-sac cigar-shaped, 72–80 9 10–14 (76 9 11,
n = 4), thin-walled, crosses poral osmoregulatory
canals and reaches slightly into median field of
proglottis (Fig. 4A). Cirrus short, cylindrical, 45–50
(47, n = 4); its basal region armed with small, claw-
shaped spines; parabasal region provided with thin,
needle-shaped spines. Length of needle-shaped spines
decreases towards apex of cirrus (Fig. 4B). Ductus
ejaculatorius forms several coils. Internal seminal
vesicle small, 17–22 9 8–10 (21 9 8, n = 4), occu-
pies not more than quarter of cirrus-sac length even
when filled. External seminal vesicle 30–32 9 20–22
(30 9 20, n = 4), connected to cirrus-sac by narrow
duct.
Vitellarium, 16–19 9 8–11 (17 9 11, n = 4), sit-
uated antiporally to ovary. Ovary transversely elon-
gate, compact, 7–10 9 80–90 (9 9 84, n = 4),
medial or may be in poral half of median field; in
latter case, ovary overlaps cirrus-sac, external seminal
vesicle and testes ventrally (Fig. 3D). Vagina thin-
walled, passes ventrally to cirrus-sac (Fig. 4A).
Uterus extends into lateral fields of both postmature
and gravid proglottides. Uterine wall thin, membra-
nous throughout morphogenesis of uterus. Number of
eggs in uterus 36–45 (Fig. 3E).
Remarks
Currently, four species are included in the subgenus
N. (Neoskrjabinolepis), which is characterised by a
gradual maturation along the strobila. These are
N. schaldybini Spassky, 1947, N. longicirrosa Kor-
nienko, Gulyaev & Melnikova, 2006, N. pilosa
Kornienko, Gulyaev & Melnikova, 2007 and N. plagis
Kornienko, Gulyaev & Melnikova, 2007.
Neoskrjabinolepis fertilis n. sp. is morphologically
close to N. longicirrosa and N. pilosa. Despite the
similar lengths of the rostellar hooks (39–42, 41–45
Fig. 4 Neoskrjabinolepis (Neoskrjabinolepidoides) hobergi n. sp. Holotype, ISEA 18.11.19.1; paratype, ISEA 18.11.19.4: A. Cirrus-
sac and vagina; B. Cirrus. Scale-bars: 25 lm
Syst Parasitol (2012) 83:179–188 185
123

and 45–49 lm, respectively), the cirrus-sac (100–130,
150–165 and 120–140 lm, respectively) and the
cirrus (85–100, 122–125 and 100–110 lm, respec-
tively), there are significant differences in hook shape
and the number of eggs. The blade of the hooks of
N. fertilis has a curvature in the middle region, in
contrast to the almost straight blade of hooks of
N. longicirrosa and N. pilosa. The blade in N. pilosus
is twice the length of the guard, whereas in N. fertilis
this ratio is much less. In N. fertilis the rostellar pouch
reaches beyond level of the posterior margins of the
suckers, but in N. pilosa it extends almost to the neck.
Furthermore, there are differences in the number of
eggs per gravid uterus: this is 16–20 in N. longicirrosa,
35–47 in N. pilosa and 55–70 in N. fertilis. Further-
more, N. pilosa has not been found outside Western
Siberia (Kornienko 2011; Kornienko et al., 2008).
This new species differs from N. plagis in the
smaller size of its rostellar hooks (39–42 and
52–55 lm, respectively), the length of the cirrus
(85–100 and 45–50 lm, respectively) and by having
more than twice the number of eggs per gravid uterus.
The lengths of the hooks of N. fertilis are similar to
those of N. schaldybini (39–42 and 38–43 lm, respec-
tively). However, the new species has a longer cirrus-
sac (100–130 vs. 73–90 lm) and longer cirrus (85–100
vs. 40–42 lm). The two species also differ from one
another on the basis of the cirral armament. The
parabasal region of the cirrus of N. schaldybini is
armed with several large, claw-shaped spines, and the
middle and distal regions of the cirrus are armed with
sparsely distributed, sabre-shaped spines; whereas the
cirrus of N. fertilis is armed with small, thin, needle-
shaped spines along its entire length.
Currently, the subgenus N. (Neoskrjabinolepido-
ides), which is characterised by strobila with a series
of proglottides at the same stage of development,
contains six species: N. (Neoskrjabinolepidoides)
singularis (Cholodkowsky, 1912) Spasskii, 1954,
N. nadtochijae Kornienko, Gulyaev & Melnikova,
2006, N. corticirrosa Kornienko, Gulyaev & Melnik-
ova, 2007, N. kedrovensis Kornienko, Gulyaev &
Melnikova, 2007, N. nuda Kornienko, Gulyaev,
Melnikova & Georgiev, 2008 and N. merkushevae
Kornienko & Binkien _e, 2008.
Neoskrjabinolepis hobergi n. sp. is most similar to
N. singularis, a species with a Euro-Siberian geo-
graphical range, which does not occur in the Far East
(Kornienko 2011; Kornienko & Binkiene, 2008;
Kornienko & Gulyaev, 2008; Kornienko et al.,
2008). Although N. hobergi and N. singularis are
similar in terms of their rostellar hooks (63–66 and
56–65 lm in length, respectively), cirrus (45–50 and
40–47 lm in length, respectively) and the number of
eggs (35–45) in the gravid uterus, there are differences
in the number of proglottides in series and in the shape
of the gravid proglottides. Their shape in N. hobergi is
much wider than long, whereas in N. singularis it is
much longer than wide, and the number of proglottides
in N. singularis is about 350 but exceeds 520 in
N. hobergi. Furthermore, N. hobergi is unique in
Neoskrjabinolepis in that its uterus crosses the longi-
tudinal osmoregulatory canals and reaches into the
lateral fields of postmature, pregravid and gravid
proglottides.
Neoskrjabinolepis hobergi differs from all other
species in this subgenus by the size of the rostellar
hooks, cirrus-sac and cirrus, its cirral armament, the
number of proglottides and the number of eggs per
gravid uterus. Its rostellar hooks are larger (63–66 lm)
than those of N. corticirrosa (48–53 lm), N. nad-
tochijae and N. nuda (40–45 lm), N. kedrovensis
(36–38 lm) and N. merkushevae (35–37 lm). The
cirrus-sac of N. hobergi only slightly crosses the poral
excretory canals, whereas the long cirrus-sacs of
N. nuda, N. nadtochijae and N. kedrovensis extend
far into the median field of the proglottides. In terms of
the length of the cirrus-sac, that of N. hobergi
(72–80 lm) is similar to N. corticirrosa (90–95 lm)
and N. merkushevae (60–65 lm). However, the latter
species can be distinguished from each of these by
differences in armament of the cirri. The parabasal
region of each cirrus in N. corticirrosa is armed with
several small, claw-shaped spines and the remainder
with scarce, sabre-shaped spines. The middle region of
the cirrus of N. merkushevae is provided with sparse,
needle-shaped spines which decrease in number dis-
tally, and the parabasal region and distal end are
unarmed. The parabasal region of each cirrus of
N. hobergi is armed with thin, needle-shaped spines
which decrease in numbers towards the apex. More-
over, these species differ in the number of proglottides
and the number of eggs in gravid proglottides. More
than 520 proglottides are found in N. hobergi, c.420 in
N. corticirrosa and only c.200 in N. merkushevae. The
gravid uterus of N. hobergi contains 36–45 eggs,
186 Syst Parasitol (2012) 83:179–188
123

whereas that of N. corticirrosa and N. merkushevae are
characterised by 20 and 12–16 eggs per gravid uterus,
respectively.
On the basis of the above comparisons, we recognise
the two species described above as new to science.
Discussion
North-American shrews (Sorex spp.) have been
recorded as hosting 20 species of cestodes belonging
to 12 genera of the families Hymenolepididae and
Dilepididae (Melnikova et al., 2003; Dokuchaev &
Gulyaev, 2007; Gulyaev et al., 2007, 2009, 2010;
Kinsella, 2007; Kinsella et al., 2008; Kinsella &
Tkach, 2009). Taxonomic composition of tapeworm
fauna parasitising Nearctic shrews appears to be
similar to that of shrews in northern Asia. In North
American shrews, there are autochthonous species
belonging to 10 cestode genera which are also known
from the Palaearctic: Staphylocystis Villot, 1877,
Skrjabinacanthus Spassky & Morosov, 1959, Ecri-
nolepis Spassky & Karpenko, 1983, Mathevolepis
Spassky, 1948, Lineolepis Spassky, 1958, Staphylo-
cystoides Yamaguti, 1959, Soricinia Spassky &
Spasskaja, 1954, Spasskylepis Schaldybin, 1964,
Monocercus Villot, 1882 and Neoskrjabinolepis Spas-
sky, 1947. There are only two genera endemic to the
Nearctic, i.e. Vogelepis Vaucher in Czaplinski &
Vaucher, 1994 and Lockerraushia Yamaguti, 1959.
Our record of species of the genus Neoskrjabinolepis
in North-American shrews refutes the recent assump-
tion that in North America species of Neoskrjabinol-
epis are replaced by species of the morphologically
related but distinct genus Vogelepis (see Gulyaev,
2003).
Only one cestode species from shrews, Lineolepis
pribilofensis (Olsen, 1969) Dokuchaev & Gulyaev,
2007, has previously been recorded as occurring at
both sides of the Bering Strait (Dokuchaev & Gulyaev,
2007). The geographical distribution of N. fertilis n.
sp. in both Alaskan and Chukotkan shrews provides
further evidence for the presence of a close faunal
relationship between the Palaearctic and the Nearctic.
Acknowledgements This work was funded by the Russian
Fund for Fundamental Research (Project Nos. 11-04-49010-a,
11-04-00870-a, 11-04-00342) and by the National Science
Foundation ‘‘Beringian Coevolution Project’’ (NSF 0196095
and 0415668).
References
Czaplinski, B., & Vaucher, C. (1994). Family HymenolepididaeAriola, 1899. In L. F. Khalil, A. Jones, & R. A. Bray (Eds)
Keys to the cestode parasites of vertebrates. Wallingford,
UK: CAB International, pp. 595–663.
Dokuchaev, N. E., & Gulyaev, V. D. (2007). [Quaternary history
of Sorex shrews in North-East Asia in view of helmintho-
logical data.] The biology of Insectivora. Proceedings ofthe 3rd All-Russian Scientific Conference on the Biology ofInsectivora (15–20 September 2007, Novosibirsk). Novo-
sibirsk: Izdatelstvo’’ZERIS’’, pp. 38–40. (In Russian).
Dunagan, T. T. (1956). Hymenolepis sphenomorphus from
Sorex cinereus in Alaska. Journal of Parasitology, 42,
657–658.
Genov, T. (1984). [Helminths of insectivores and rodents inBulgaria.] Sofia: Izdatelstvo na Bulgarskata Akademiya na
Naukite, 348 pp. (In Bulgarian).
Gulyaev, V. D. (2003). [Species diversity and taxonomic
structure of hymenolepidid cestodes of shrews in Holarc-
tic.] The problems of modern parasitology. Proceedings ofthe International Conference and 3rd Congress of theRussian Society of Parasitologists–Russian Academy ofSciences. (Petrozavodsk, 6–12 October 2003). St Peters-
burg: Zoological Institute RAS, pp. 137–139. (In Russian).
Gulyaev, V. D., Dokuchaev, N. E., & Kornienko, S. A. (2007).
[The cestodes of genus Staphylocystoides Yamaguti, 1959
(Cestoda, Hymenolepididae) in shrews of Berengia].
Vestnik Severo-Vostochnogo Nauchnogo Tsentra DVORAN, 4, 75–84. (In Russian).
Gulyaev, V. D., Dokuchaev, N. E., & Kornienko, S. A. (2009).
[Hymenolepidid cestodes of shrews in Alaska]. Parasi-tology researches in Siberia and the Far East. Proceedingsof the 3rd Interregional Scientific Conference (Novosi-
birsk, 15–20 September 2009). Novosibirsk: ISEA SB RAS,pp. 71–72. (In Russian).
Gulyaev, V. D., Dokuchaev, N. E., & Lykova, K. A. (2010).
[Description of Spasskylepis rauschi sp. n. (Cestoda,
Hymenolepididae) from shrews in Alaska]. Vestnik Seve-ro-Vostochnogo Nauchnogo Tsentra DVO RAN, 2, 75–77.
(In Russian).
Gulyaev, V. D., & Kornienko, S. A. (2009). [On causes and
mechanisms of the appearance of mini polymerous
Hymenolepididae (Cyclophyllidea, Cestoda), parasites of
shrews]. Trudy Zoologicheskogo Instituta RAN, 313(3),
249–256. (In Russian).
Kinsella, J. (2007). Helminths of the vagrant shrew, Sorexvagrans, from western Montana, USA. Acta Parasitolog-ica, 52, 151–155.
Kinsella, J., & Tkach, V. (2009). Checklist of helminth parasites
of Soricomorpha (= Insectivora) of North America north of
Mexico. Zootaxa, 1969, 36–58.
Kinsella, J. M., Vandegrift, K. J., Grant, L. K., Bozick, B. A., &
Hudson, P. J. (2008). Gastrointestinal helminths of the
masked shrew, Sorex cinereus, from Pennsylvania. Com-parative Parasitology, 75, 141–144.
Kisielewska, K. (1958). Cysticercoid of the tapeworm Neoskr-jabinolepis singularis (Cholodkowsky 1912) Spassky,
1954 in a beetle of the family Catopidae. Bulletin del’Academie Polonaise de Sciences (Cl. 2), 6, 206–208.
Syst Parasitol (2012) 83:179–188 187
123

Kobuley, N. (1953). On the anatomy and systematics of poorly
known cestodes from shrews. Acta Veterinaria AcademiaeScientarum Hungaricae, 3, 431–438. (In Russian).
Kornienko, S. A. (2011). [The cestodes of the genus Neoskrjab-inolepis (Hymenolepididae, Cyclophyllidea, Cestoda)—
parasites of Palaearctic shrews (Soricidae)]. In: Modernproblems of evolutionary morphology of animals. Pro-ceedings 2nd All-Russian Scientific Conference with Inter-national Participation on the 105th Anniversary of the Birthof Academician A.V. Ivanov (17–19 October 2011, St
Petersburg). St Petersburg: Zoological Institute RAS,
pp. 196–199. (In Russian).
Kornienko, S. A., & Binkiene, R. (2008). Neoskrjabinolepismerkushevae sp. n. (Cyclophyllidea, Hymenolepididae), a
new cestode from shrews from Palaearctic. Folia Parasi-tologica, 55, 136–140.
Kornienko, S. A., & Gulyaev, V. D. (2008). [Species resources
of cestodes of the genus Neoskrjabinolepis of Palaearctic
shrews]. In: Galaktionov, K. V., & Dobrovolskij, A. A.
(Eds) ‘‘Parasitology of the 21st Century—problems,
methods, solutions’’. Proceedings of the 4th Congress ofthe Russian Society of Parasitologists – Russian Academyof Sciences. (20–25 October 2008, Zoological Institute
RAS, St Petersburg). St Petersburg: ‘‘Lema’’, 2, 72–75.
(In Russian).
Kornienko, S. A., Gulyaev, V. D., & Erzhanov, N. T. (2010). A
new cestode, Neoskrjabinolepis gvosdevi (Cyclophyllidea:
Hymenolepididae) from the shrew Sorex tundrensis Mer-
riam, 1900, in Kazakhstan. Zoosystematica Rossica, 19,
13–17.
Kornienko, S. A., Gulyaev, V. D., Mel’nikova, Y. A., & Geor-
giev, B. B. (2008). Neoskrjabinolepis nuda n. sp. from
shrews in Sakhalin Island, Russia, with a taxonomic review
of Neoskrjabinolepis Spasskii, 1947 (Cyclophyllidea,
Hymenolepididae). Systematic Parasitology, 70, 147–158.
Kornienko, S. A., Gulyaev, V. D., & Melnikova, Yu A. (2006).
[On the morphology and systematics of cestodes of the
genus Neoskrjabinolepis Spassky, 1947 (Cyclophyllidea,
Hymenolepididae)]. Zoologicheskiy Zhurnal, 85, 134–145.
(In Russian).
Kornienko, S. A., Gulyaev, V. D., & Mel’nikova, Yu. A. (2007).
[New species of cestodes of the genus NeoskrjabinolepisSpassky, 1947 (Cyclophyllidea, Hymenolepididae) from
shrews of Russia.] Zoologicheskiy Zhurnal, 86, 259–269.
(In Russian).
Mas-Coma, S., & Galan-Puchades, M. T. (1991). A methodol-
ogy for the morphoanatomic and systematic study of the
species of the family Hymenolepididae Railliet et Henry,
1909 (Cestoda: Cyclophyllidea). Research and Reviews inParasitology, 51, 139–173.
Melnikova, Yu. A., Gulyaev, V. D., & Dokuchaev, N. E. (2003).
[The structure of community of cestodes of shrews of West
Berengia and the Far East]. In: The problems of modernparasitology. Proceedings of the International Conferenceand 3rd Congress of the Russian Society of Parasitologists– Russian Academy of Sciences. (Petrozavodsk, 6–12
October 2003). St Petersburg: Zoological Institute, RAS,
pp. 18–20. (In Russian).
Olsen, O. W. (1969). Hymenolepis pribilofensis n. sp. (Cestoda:
Hymenolepididae) from the Pribilof shrew (Sorex pri-bilofensis Merriam) from the Pribilof Islands, Alaska.
Canadian Journal of Zoology, 47, 449–454.
Pojmanska, T. (1957). Pasozyty wewnetrzne (Cestoda, Trema-
toda) drobnych ssakowpolnych z okoliz Turwi kolo Pos-
nana. Acta Parasitologica Polonica, 5, 117–161.
Prokopic, J. (1956). Helmintofauna rejska obecneho (Sorexaraneus) v CSSR. Ceskoslovenska Parasitologie, 3,
109–131.
Prokopic, J. (1958). [On the helminth fauna of the genus Sorexin Czechoslovakia]. Zoologicheskiy Zhurnal, 38, 174–183.
(In Russian).
Rausch, R., & Kuns, M. L. (1950). Studies on some North
American shrew cestodes. Journal of Parasitology, 36,
433–438.
Rybicka, K. (1959). Tapeworms of forest micromammals
(Rodentia and Insectivora) from Kampinos Wilderness.
Acta Parasitologica Polonica, 7, 393–420.
Spassky, A. A. (1947). [The phenomenon of confluence of
proglottides and uteri in cestodes]. Doklady Akademii NaukSSSR, 58, 723–724. (In Russian).
Spassky, A. A. (1954). [Classification of hymenolepidids of
mammals]. Trudy Gel’mintologcheskoi Laboratorii, 7,
120–167. (In Russian).
Vaucher, C. (1971). Les cestodes parasites des Soricidae
d’Europe. Etude anatomique, revision taxonomique et
biologie. Revue Suisse de Zoologie, 78, 1–113.
Voge, M., & Rausch, R. (1955). Occurrence and distribution of
hymenolepidid cestodes in shrews. Journal of Parasitol-ogy, 41, 566–574.
Zarnowski, E. (1955). Robaki paso _zytnicze drobnych ssakow
lesnych (Rodentia i Insectivora) okolicy Pulaw(woj.
Lubelskie). I. Cestoda. Acta Parasitologica Polonica, 3,
279–368.
188 Syst Parasitol (2012) 83:179–188
123
Related Documents











