1648 • JID 2005:191 (15 May) • Brown et al. MAJOR ARTICLE Treatment of Schistosoma mansoni Infection Increases Helminth-Specific Type 2 Cytokine Responses and HIV-1 Loads in Coinfected Ugandan Adults Michael Brown, 1,2 Patrice A. Mawa, 1 Sarah Joseph, 3 Joseph Bukusuba, 1 Christine Watera, 1 James A. G. Whitworth, 1,2 David W. Dunne, 3 and Alison M. Elliott 1,2 1 Uganda Virus Research Institute, Entebbe, Uganda; 2 London School of Hygiene and Tropical Medicine, London, and 3 Department of Pathology, University of Cambridge, Cambridge, United Kingdom Background. Studies showing that helminths stimulate type 2 cytokine responses and influence responses to unrelated antigens suggest that helminths may accelerate human immunodeficiency virus type 1 (HIV-1) disease progression in coinfected individuals and that antihelminthic therapy may be beneficial. By the same logic, however, the increase in type 2 cytokines occurring immediately after antischistosomal treatment might increase viral replication and be detrimental. Methods. To assess the effect of antischistosomal therapy on immune responses and HIV-1 replication, a cohort of 163 Ugandans coinfected with Schistosoma mansoni and HIV-1 was treated with praziquantel. CD4 + T lymphocyte counts, eosinophil counts, and plasma HIV-1 RNA concentrations were measured before treatment and 1 month and 5 months after treatment. Schistosoma mansoni– and Mycobacterium tuberculosis–specific cytokine responses and serum interleukin (IL)–10 concentrations were analyzed. Results. Transient increases in viral load and sustained decreases in CD4 + T lymphocyte count were observed, especially in subjects with higher-intensity infections. Despite enhanced posttreatment S. mansoni–specific type 2 responses, no increase in eosinophils or in M. tuberculosis–specific type 2 responses nor any decline in M. tuber- culosis–specific interferon (IFN)–g responses were seen. A significant decline in circulating IL-10 concentrations was observed. Conclusion. Although the mechanisms underlying the increase in viral load after treatment with praziquantel are unclear, these results do not support the hypothesis that treating schistosomiasis is beneficial in the management of HIV-1 disease in Africa. Concurrent infection with helminths and HIV-1 is com- mon in sub-Saharan Africa. The type 1/type 2 model of immune responses to infection would suggest a detri- mental effect of helminth infection, since the balance Received 29 August 2004; accepted 20 December 2004; electronically published 5 April 2005. Presented in part: 52nd Annual Meeting of the American Society of Hygiene and Tropical Medicine, Philadelphia, Pennsylvania, 3–7 December 2003 (abstract 777). Financial support: Wellcome Trust (Research Training Fellowship 060116/Z/99/ Z to M.B.); UK Medical Research Council and Wellcome Trust program grants (to D.W.D. and S.J.). Reprints or correspondence: Dr. Michael Brown, Dept. of Infectious and Tropical Diseases, London School of Hygiene and Tropical Medicine, Keppel St., London WC1E 7HT, United Kingdom ([email protected]). The Journal of Infectious Diseases 2005; 191:1648–57 2005 by the Infectious Diseases Society of America. All rights reserved. 0022-1899/2005/19110-0010$15.00 in favor of type 2 cytokines at the expense of type 1 cytokines might encourage HIV-1 disease progression; immune control over HIV-1 replication appears to be dominated by type 1–mediated mechanisms, and the increased susceptibility of Th0 or Th2 cells to HIV-1 infection is particularly relevant to the course of HIV- 1 disease in the context of chronic helminth infection, in which type 2 cells are in abundance [1–3]. For this reason, as well as on the basis of evidence of chronic immune activation in coinfected individuals, it has been suggested that antihelminthic therapy is an appropriate intervention as part of the package of care for HIV-1– infected people in places where antiretroviral drugs are not widely available [4, 5]. Few studies have addressed this issue in epidemiologically meaningful cohorts [5]. We recently demonstrated that helminth infection, including Schistosoma mansoni infection, in HIV-1–in- by guest on August 16, 2016 http://jid.oxfordjournals.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1648 • JID 2005:191 (15 May) • Brown et al.
M A J O R A R T I C L E
Treatment of Schistosoma mansoni InfectionIncreases Helminth-Specific Type 2 CytokineResponses and HIV-1 Loads in CoinfectedUgandan Adults
Michael Brown,1,2 Patrice A. Mawa,1 Sarah Joseph,3 Joseph Bukusuba,1 Christine Watera,1
James A. G. Whitworth,1,2 David W. Dunne,3 and Alison M. Elliott1,2
1Uganda Virus Research Institute, Entebbe, Uganda; 2London School of Hygiene and Tropical Medicine, London, and 3Department of Pathology,University of Cambridge, Cambridge, United Kingdom
Background. Studies showing that helminths stimulate type 2 cytokine responses and influence responses tounrelated antigens suggest that helminths may accelerate human immunodeficiency virus type 1 (HIV-1) diseaseprogression in coinfected individuals and that antihelminthic therapy may be beneficial. By the same logic, however,the increase in type 2 cytokines occurring immediately after antischistosomal treatment might increase viralreplication and be detrimental.
Methods. To assess the effect of antischistosomal therapy on immune responses and HIV-1 replication, acohort of 163 Ugandans coinfected with Schistosoma mansoni and HIV-1 was treated with praziquantel. CD4+ Tlymphocyte counts, eosinophil counts, and plasma HIV-1 RNA concentrations were measured before treatmentand 1 month and 5 months after treatment. Schistosoma mansoni– and Mycobacterium tuberculosis–specific cytokineresponses and serum interleukin (IL)–10 concentrations were analyzed.
Results. Transient increases in viral load and sustained decreases in CD4+ T lymphocyte count were observed,especially in subjects with higher-intensity infections. Despite enhanced posttreatment S. mansoni–specific type 2responses, no increase in eosinophils or in M. tuberculosis–specific type 2 responses nor any decline in M. tuber-culosis–specific interferon (IFN)–g responses were seen. A significant decline in circulating IL-10 concentrationswas observed.
Conclusion. Although the mechanisms underlying the increase in viral load after treatment with praziquantelare unclear, these results do not support the hypothesis that treating schistosomiasis is beneficial in the managementof HIV-1 disease in Africa.
Concurrent infection with helminths and HIV-1 is com-
mon in sub-Saharan Africa. The type 1/type 2 model of
immune responses to infection would suggest a detri-
mental effect of helminth infection, since the balance
Received 29 August 2004; accepted 20 December 2004; electronically published5 April 2005.
Presented in part: 52nd Annual Meeting of the American Society of Hygieneand Tropical Medicine, Philadelphia, Pennsylvania, 3–7 December 2003 (abstract777).
Financial support: Wellcome Trust (Research Training Fellowship 060116/Z/99/Z to M.B.); UK Medical Research Council and Wellcome Trust program grants (toD.W.D. and S.J.).
Reprints or correspondence: Dr. Michael Brown, Dept. of Infectious and TropicalDiseases, London School of Hygiene and Tropical Medicine, Keppel St., LondonWC1E 7HT, United Kingdom ([email protected]).
The Journal of Infectious Diseases 2005; 191:1648–57� 2005 by the Infectious Diseases Society of America. All rights reserved.0022-1899/2005/19110-0010$15.00
in favor of type 2 cytokines at the expense of type 1
cytokines might encourage HIV-1 disease progression;
immune control over HIV-1 replication appears to be
dominated by type 1–mediated mechanisms, and the
increased susceptibility of Th0 or Th2 cells to HIV-1
infection is particularly relevant to the course of HIV-
1 disease in the context of chronic helminth infection,
in which type 2 cells are in abundance [1–3]. For this
reason, as well as on the basis of evidence of chronic
immune activation in coinfected individuals, it has been
suggested that antihelminthic therapy is an appropriate
intervention as part of the package of care for HIV-1–
infected people in places where antiretroviral drugs are
not widely available [4, 5]. Few studies have addressed
this issue in epidemiologically meaningful cohorts [5].
We recently demonstrated that helminth infection,
including Schistosoma mansoni infection, in HIV-1–in-
by guest on August 16, 2016
http://jid.oxfordjournals.org/D
ownloaded from

S. mansoni Treatment in HIV-1 Coinfection • JID 2005:191 (15 May) • 1649
Figure 1. Flow diagram of the study design, for subjects infected with Schistosoma mansoni at enrollment. The effects specific to praziquanteltreatment (PZQ) were analyzed by treating S. mansoni–infected subjects 1 month after empirical albendazole treatment (ABZ) had been given forintestinal nematodes. IL-10, interleukin-10.
fected adults in a semiurban cohort in Uganda was not associated
with higher viral load or lower CD4+ T lymphocyte count and
that, unexpectedly, effective antischistosomal treatment was as-
sociated with a CD4+ T lymphocyte decline greater than that
seen in subjects with persistent schistosomiasis or reinfection 5
months after treatment [6]. These results, as well as those from
a study conducted in Kenya that failed to demonstrate a decline
in viral load after successful treatment of S. mansoni in coinfected
adults [7], do not support the hypothesis that antischistosomal
treatment is a useful strategy for coinfected subjects.
The antischistosome immune response after praziquantel
therapy has been characterized in animal models and in studies
of humans. Damage to the worm tegument exposes antigens
that induce strong type 2 immune responses, characterized by
increased production of schistosome-specific interleukin (IL)–
4, IL-5, and IgG; total IgE concentration; peripheral blood eo-
sinophil counts; and histamine concentrations [8–14]. Such an
increase in type 2 responses might be expected to affect the
wider immune response, with suppressed type 1 cytokine re-
sponses and increased type 2 cytokine responses to unrelated
antigens [8–14]. Thus, rather than improving immune function
in coinfected adults, praziquantel treatment might be associ-
ated, at least transiently, with a loss of useful HIV-1–specific
immune responses and acceleration of viral replication, leading
to increased HIV-1 disease progression.
We studied the effect of praziquantel on viral load, CD4+ T
lymphocyte counts, peripheral blood eosinophil counts, cyto-
kine responses to S. mansoni and Mycobacterium tuberculosis
antigens in a whole-blood assay, and circulating IL-10 concen-
trations, within a larger study of interactions between helminths
and HIV-1 in Uganda.
SUBJECTS, MATERIALS, AND METHODS
Study population. Study subjects, who attend either the AIDS
Support Organisation clinic or the Uganda Virus Research In-
stitute clinic in Entebbe, were members of the Entebbe Cohort
(EC). This prospective cohort of HIV-1–infected adults was
established by the UK Medical Research Council in 1995 [15],
and recruitment into the cohort continues. Informed consent
was obtained from all patients; ethical approval for the study
was sought from and provided by the Uganda National Coun-
cil for Science and Technology, the ethics committee for the
Uganda Virus Research Institute, and the ethics committee of
the London School of Hygiene and Tropical Medicine. Details
of ethical approval and recruitment into the study of helminth–
HIV-1 interactions have been described elsewhere [6]. Briefly,
as illustrated in figure 1, enrollment into this study took place
during participants’ routine (every 6 months) cohort visits,
when a questionnaire was completed, stool and blood samples
were collected, and presumptive treatment with 400 mg of al-
bendazole was provided. Subjects identified on the basis of their
enrollment specimens as having S. mansoni infection returned
after 1 month, when repeat stool and blood samples were taken;
subjects were treated with 40 mg/kg praziquantel and returned
after 1 month, when blood was collected again. All participants
with S. mansoni were treated, because it was not considered
ethical to leave these infections untreated. Subjects were seen
at their subsequent routine EC appointment—that is, 5 months
after praziquantel treatment—when blood samples were col-
lected. Subjects who did not attend appointments were traced
at their homes by fieldworkers and were encouraged to attend
by guest on August 16, 2016
http://jid.oxfordjournals.org/D
ownloaded from

1650 • JID 2005:191 (15 May) • Brown et al.
the clinic. At the time of this study, antiretroviral therapy was
not available within the cohort.
Diagnostic parasitologic assessment. Duplicate 41.7-mg
Kato-Katz smears were performed on all fecal specimens. In-
fection intensity was estimated by averaging the egg counts in
the 2 stool samples collected—the first at enrollment into the
study and the second at the pre–praziquantel treatment visit 1
month later.
ELISA for circulating anodic antigen (CAA) of S. mansoni
[16] was performed on serum samples, to increase the diag-
nostic yield and to avoid loss of sensitivity resulting from the
reported effect of immunosuppression reducing S. mansoni fe-
cal egg excretion [17]. This assay was performed on all subjects
at enrollment. Subjects with a positive CAA ELISA result but
no eggs detectable in Kato-Katz smears were classified as having
0 eggs/g of feces.
In addition to infection assessment by the Kato-Katz method,
diagnosis of other helminth infections was performed by as-
sessment of formol-ether concentration and charcoal culture of
stool samples and by use of a modified Knott’s concentration
method for detection of circulating microfilaria [18]. These tech-
niques are described in detail elsewhere [6, 19].
Eosinophil, CD4+ T lymphocyte, and viral load quanti-
fication. Leukocyte differential counts were obtained by use
of an ACT Diff II analyzer (Beckman Coulter). Eosinophil
counts were read microscopically, by 1 of 5 technicians, from
a thin film stained with Leishman’s stain. Differential eosinophil
counts were determined by counting 100 consecutive leuko-
cytes. CD4+ T lymphocytes were enumerated in EDTA blood
by use of FACScount (Becton Dickinson). Plasma was stored
at �80�C for HIV-1 RNA quantification, which was performed
by use of a Bayer Versant branched DNA assay (version 3.0;
Bayer). The volume of each plasma sample assayed was 250
mL, giving a detectable range of 200–2,000,000 copies/mL. Sam-
ples from 1 subject were below the detectable range and were
classified as 100 copies/mL (2 log10) for the purposes of analysis.
Samples from 5 subjects were above the detectable range and,
because there was an insufficient amount of specimen for fur-
ther dilution, were classified as 2,000,000 copies/mL (6.3 log10)
for the purposes of analysis.
Whole-blood cytokine assay. Cytokine responses were ex-
amined using a whole-blood assay, as described elsewhere [20].
Briefly, unseparated blood collected in sodium heparin was
diluted to a final concentration of 1:4 with serum-free medium
(RPMI supplemented with penicillin and streptomycin and 2
mmol/L glutamine), and 200 mL/well was added to 96-well,
round-bottom plates. Cells were stimulated with antigen or
mitogen (phytohemagglutinin [PHA]; Sigma) at a final con-
centration of 10 mg/mL or were left unstimulated. Cultures were
incubated at 37�C with 5% CO2. Supernatants were harvested
on day 1 for IL-4, on day 4 for IL-13, and on day 6 for interfer-
on (IFN)–g and IL-5. Viral inactivation of supernatants with
0.03% tributyl phosphate (Sigma T4908) and 1% Tween 80
(Sigma P1754) was performed before freezing at �80�C. Crude
culture filtrate proteins (CFPs) of M. tuberculosis H37Rv were
provided by J. T. Belisle (Colorado State University, Fort Col-
lins), courtesy of the National Institutes of Health (National
Institute of Allergy and Infectious Diseases grant NO1 AI-
75320). This antigen is composed primarily of antigen 85, a
secreted protein complex that is involved in the synthesis of
the cell wall and is immunogenic in humans [20–23]. S. man-
soni adult worm antigen (SWA) and S. mansoni egg antigen (SEA)
were prepared as described elsewhere [24].
Supernatants from all time points were analyzed on the same
ELISA plate, to minimize the effects of interplate variability on
the comparison of results between time points. Plates were
deemed unacceptable and the ELISA was repeated if positive
control results were 12 SDs from the mean control result, if
the correlation coefficient for the standard curve was !0.96, or
if the lowest detectable standard was 122.2 pg/mL. Cytokine
responses were defined as positive for any result 17 pg/mL (the
lowest point on the standard curve in the majority of assays).
Low-level production of cytokines in unstimulated wells was
subtracted from the concentration produced in response to
stimulation.
Serum IL-10 ELISA method. ELISAs for IL-10 were per-
formed on sera (separated and stored at or below �20�C within
24 h of sample collection) from all subjects at all time points.
ELISAs were performed using an OPTEIA kit (Becton Dick-
inson) in accordance with the manufacturer’s protocol. Stan-
dard series of dilutions of recombinant IL-10 (1800 to 2.4 pg/
mL) were used. The lowest standard (2.4 pg/mL) was defined
as the cutoff and was subtracted from all values to give the
response above the limit of detection.
Statistical analysis. Study data were entered into Foxpro
for Windows (version 2.6; Microsoft) and analyzed with STATA
(version 7.0; STATA Corporation). Comparisons of median
CD4+ T lymphocyte counts and median absolute eosinophil
counts were performed by Wilcoxon signed rank tests, and
comparisons of mean log10 viral load were performed by Stu-
dent’s paired t test, where appropriate. Analyses of association
were performed using standard tables.2 � 2
The observed distribution of serum IL-10 responses was skewed,
requiring log10 transformation or nonparametric statistical tech-
niques; for most whole-blood assay cytokine responses, !50%
of subjects displayed concentrations above background values,
so most analyses compared proportions of subjects with de-
tectable cytokine responses; median values were used only after
restricting analyses to subjects with a detectable response. Mc-
Nemar’s x2 test was used for paired longitudinal analyses.
by guest on August 16, 2016
http://jid.oxfordjournals.org/D
ownloaded from

S. mansoni Treatment in HIV-1 Coinfection • JID 2005:191 (15 May) • 1651
Table 1. Baseline characteristics of 163 Schistosoma mansoni–infected HIV-1–positiveadults in Uganda.
Characteristic Value at baseline
Age, median (range), years 32 (19–51)Male sex 67 (41.1)World Health Organization clinical stage 3 or 4 61 (38)CD4+ T lymphocyte count, median (interquartile range), cells/mm3 298 (151–477)Log10 viral load, mean (SD), log10 copies/mL ( )n p 138 4.80 (0.82)S. mansoni infection intensity 1100 eggs/g 26 (16.0)Detectable mitogen-induced interferon-g response ( )n p 90 67 (74.4)Infected with other helminths 71 (43.6)Eosinophil count, median (interquartile range), cells�109/L ( )n p 139 0.29 (0.13–0.52)
NOTE. Data are no. (%) of subjects, unless otherwise indicated.
RESULTS
Baseline characteristics. Of 663 subjects enrolled in the study,
116 were excluded because of incomplete parasitological data
or other reasons, as discussed elsewhere [6]. Of the remaining
547 subjects, 173 were infected with S. mansoni. Of these, 9
failed to attend follow-up visits for praziquantel treatment.
CD4+ T lymphocyte count was unavailable for 1 subject at this
visit. The remaining 163 S. mansoni–infected subjects were in-
cluded in subsequent analyses. Baseline characteristics of these
subjects are displayed in table 1.
S. mansoni infection was diagnosed by Kato-Katz smear in 91
subjects and was diagnosed by CAA ELISA in an additional 72
subjects. Estimates of infection intensity in subjects with positive
Kato-Katz smears correlated well with CAA concentrations (Spear-
man rank correlation coefficient ; ). Therer p 0.63 P ! .0001
was no association between infection intensity and CD4+ T
lymphocyte count (CAA concentration vs. baseline CD4+ T
lymphocyte count, ; ) or viral load (CAA con-r p 0.01 P p .86
centration vs. baseline viral load, ; ).r p 0.04 P p .62
CD4+ T lymphocyte count and viral load after treatment with
praziquantel. During the follow-up period, 3 subjects de-
faulted, 3 subjects were too sick to attend follow-up visits, and
4 subjects died. Measurement of CD4+ T lymphocyte count failed
for 1 subject at the 5 month follow-up, leaving 152 subjects with
follow-up data. Because of insufficient plasma sample volumes,
viral load data were not available at all time points for 33 subjects.
There were no differences in CD4+ T lymphocyte count or S.
mansoni infection intensity between subjects with and subjects
without complete viral load data (data not shown).
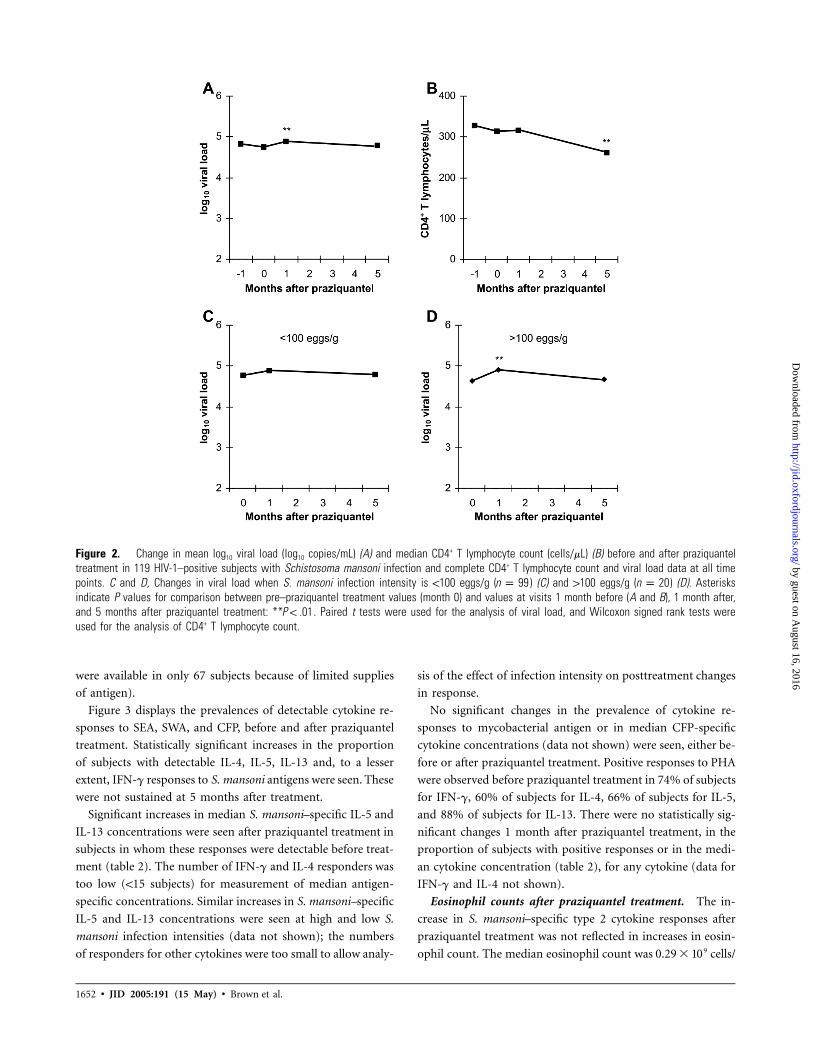
Figure 2A and 2B shows mean log10 viral load and median
CD4+ T lymphocyte count before and after praziquantel treat-
ment for 119 subjects with complete CD4+ T lymphocyte count
and viral load data. A statistically significant increase in viral
load 1 month after treatment, from 4.76 to 4.89 log10 copies/
mL ( , paired t test), was not sustained at 5 monthsP p .001
after treatment. There was a concomitant but not statistically
significant decline in CD4+ T lymphocyte count 1 month after
praziquantel treatment in these subjects (as displayed in figure
2B, median CD4+ T lymphocyte count did not fall, but rank
values declined; , Wilcoxon signed rank test); however,P p .10
a significant decline in median CD4+ T lymphocyte count after
praziquantel treatment (from 306 to 261 cells/mL at 1 month;
, Wilcoxon signed rank test) was observed when all 152P p .03
subjects with CD4+ T lymphocyte count data at all time points
were analyzed.
The increase in viral load seen after praziquantel treatment
was greater in subjects with higher S. mansoni infection inten-
sities (figure 2C and 2D). A weak correlation between initial
CAA concentration and viral load increase was observed (r
p0.21, ). CD4+ T lymphocyte decline was 22 cells/mLP p .02
in subjects with 1100 S. mansoni eggs/g and 17 cells/mL in sub-
jects with !100 eggs/g ( ).P p .84
No statistically significant changes in viral load ( ,P p .28
paired t test) or CD4+ T lymphocyte count ( , WilcoxonP p .09
signed rank test) occurred in the month before praziquantel
treatment (figure 2). When the analysis was restricted to subjects
free of other helminths, mean viral load was 4.84 log10 copies/
mL at enrollment and 4.83 log10 copies/mL at the pre–prazi-
quantel treatment visit ( ; , paired t test); for then p 63 P p .91
same subjects, the median CD4+ T lymphocyte count was 263
cells/mL at enrollment and 286 cells/mL at the pre–praziquantel
treatment visit ( , Wilcoxon signed rank test).P p .24
Antigen-specific cytokine responses. Complete data on an-
tigen-specific and mitogen-induced cytokine responses were
unavailable for 62 subjects, because of plate contamination,
insufficient sample, or ELISAs that failed despite the repeated
assay of initially unacceptable plates. There were no significant
differences in CD4+ T lymphocyte count, log10 viral load, or S.
mansoni infection intensity between subjects with and subjects
without complete cytokine data (data not shown). Complete
cytokine data were available at all time points in 90 subjects
(with the exception of data on SEA-specific responses, which
by guest on August 16, 2016
http://jid.oxfordjournals.org/D
ownloaded from
1652 • JID 2005:191 (15 May) • Brown et al.
Figure 2. Change in mean log10 viral load (log10 copies/mL) (A) and median CD4+ T lymphocyte count (cells/mL) (B) before and after praziquanteltreatment in 119 HIV-1–positive subjects with Schistosoma mansoni infection and complete CD4+ T lymphocyte count and viral load data at all timepoints. C and D, Changes in viral load when S. mansoni infection intensity is !100 eggs/g ( ) (C) and 1100 eggs/g ( ) (D). Asterisksn p 99 n p 20indicate P values for comparison between pre–praziquantel treatment values (month 0) and values at visits 1 month before (A and B), 1 month after,and 5 months after praziquantel treatment: ** . Paired t tests were used for the analysis of viral load, and Wilcoxon signed rank tests wereP ! .01used for the analysis of CD4+ T lymphocyte count.
were available in only 67 subjects because of limited supplies
of antigen).
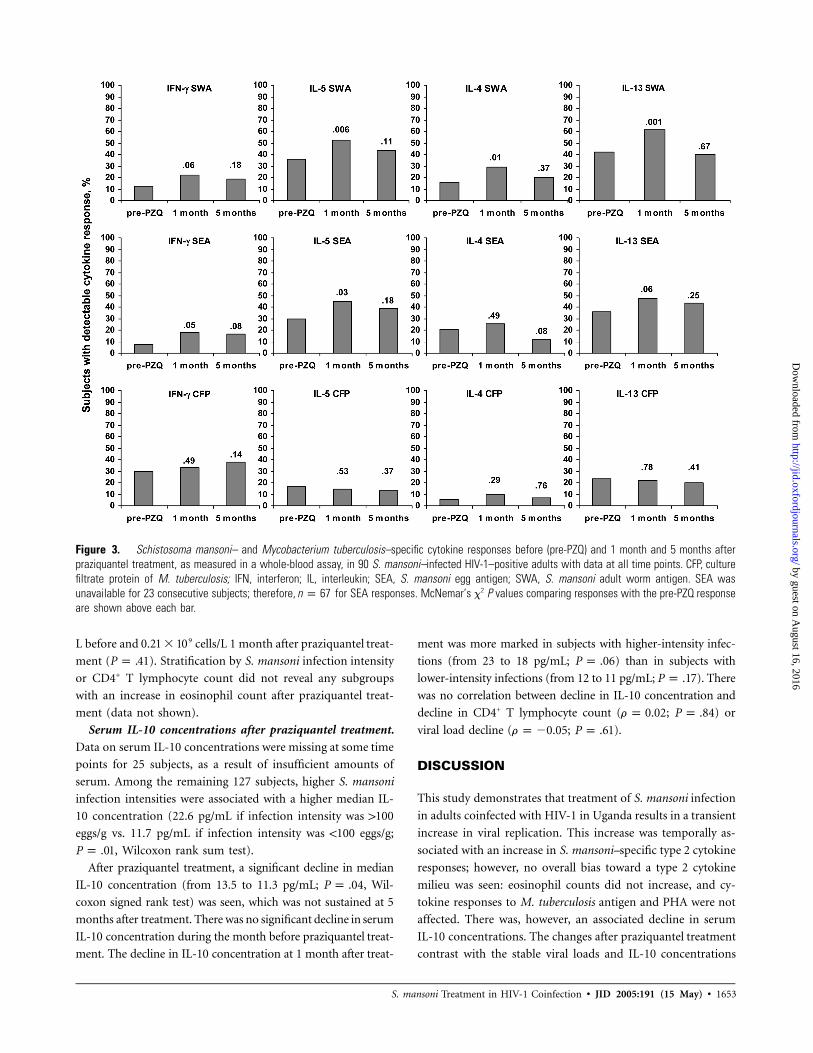
Figure 3 displays the prevalences of detectable cytokine re-
sponses to SEA, SWA, and CFP, before and after praziquantel
treatment. Statistically significant increases in the proportion
of subjects with detectable IL-4, IL-5, IL-13 and, to a lesser
extent, IFN-g responses to S. mansoni antigens were seen. These
were not sustained at 5 months after treatment.
Significant increases in median S. mansoni–specific IL-5 and
IL-13 concentrations were seen after praziquantel treatment in
subjects in whom these responses were detectable before treat-
ment (table 2). The number of IFN-g and IL-4 responders was
too low (!15 subjects) for measurement of median antigen-
specific concentrations. Similar increases in S. mansoni–specific
IL-5 and IL-13 concentrations were seen at high and low S.
mansoni infection intensities (data not shown); the numbers
of responders for other cytokines were too small to allow analy-
sis of the effect of infection intensity on posttreatment changes
in response.
No significant changes in the prevalence of cytokine re-
sponses to mycobacterial antigen or in median CFP-specific
cytokine concentrations (data not shown) were seen, either be-
fore or after praziquantel treatment. Positive responses to PHA
were observed before praziquantel treatment in 74% of subjects
for IFN-g, 60% of subjects for IL-4, 66% of subjects for IL-5,
and 88% of subjects for IL-13. There were no statistically sig-
nificant changes 1 month after praziquantel treatment, in the
proportion of subjects with positive responses or in the medi-
an cytokine concentration (table 2), for any cytokine (data for
IFN-g and IL-4 not shown).
Eosinophil counts after praziquantel treatment. The in-
crease in S. mansoni–specific type 2 cytokine responses after
praziquantel treatment was not reflected in increases in eosin-
ophil count. The median eosinophil count was cells/90.29 � 10
by guest on August 16, 2016
http://jid.oxfordjournals.org/D
ownloaded from
S. mansoni Treatment in HIV-1 Coinfection • JID 2005:191 (15 May) • 1653
Figure 3. Schistosoma mansoni– and Mycobacterium tuberculosis–specific cytokine responses before (pre-PZQ) and 1 month and 5 months afterpraziquantel treatment, as measured in a whole-blood assay, in 90 S. mansoni–infected HIV-1–positive adults with data at all time points. CFP, culturefiltrate protein of M. tuberculosis; IFN, interferon; IL, interleukin; SEA, S. mansoni egg antigen; SWA, S. mansoni adult worm antigen. SEA wasunavailable for 23 consecutive subjects; therefore, for SEA responses. McNemar’s x2 P values comparing responses with the pre-PZQ responsen p 67are shown above each bar.
L before and cells/L 1 month after praziquantel treat-90.21 � 10
ment ( ). Stratification by S. mansoni infection intensityP p .41
or CD4+ T lymphocyte count did not reveal any subgroups
with an increase in eosinophil count after praziquantel treat-
ment (data not shown).
Serum IL-10 concentrations after praziquantel treatment.
Data on serum IL-10 concentrations were missing at some time
points for 25 subjects, as a result of insufficient amounts of
serum. Among the remaining 127 subjects, higher S. mansoni
infection intensities were associated with a higher median IL-
10 concentration (22.6 pg/mL if infection intensity was 1100
eggs/g vs. 11.7 pg/mL if infection intensity was !100 eggs/g;
, Wilcoxon rank sum test).P p .01
After praziquantel treatment, a significant decline in median
IL-10 concentration (from 13.5 to 11.3 pg/mL; , Wil-P p .04
coxon signed rank test) was seen, which was not sustained at 5
months after treatment. There was no significant decline in serum
IL-10 concentration during the month before praziquantel treat-
ment. The decline in IL-10 concentration at 1 month after treat-
ment was more marked in subjects with higher-intensity infec-
tions (from 23 to 18 pg/mL; ) than in subjects withP p .06
lower-intensity infections (from 12 to 11 pg/mL; ). ThereP p .17
was no correlation between decline in IL-10 concentration and
decline in CD4+ T lymphocyte count ( ; ) orr p 0.02 P p .84
viral load decline ( ; ).r p �0.05 P p .61
DISCUSSION
This study demonstrates that treatment of S. mansoni infection
in adults coinfected with HIV-1 in Uganda results in a transient
increase in viral replication. This increase was temporally as-
sociated with an increase in S. mansoni–specific type 2 cytokine
responses; however, no overall bias toward a type 2 cytokine
milieu was seen: eosinophil counts did not increase, and cy-
tokine responses to M. tuberculosis antigen and PHA were not
affected. There was, however, an associated decline in serum
IL-10 concentrations. The changes after praziquantel treatment
contrast with the stable viral loads and IL-10 concentrations
by guest on August 16, 2016
http://jid.oxfordjournals.org/D
ownloaded from

Tabl
e2.
Med
ian
Schi
stos
oma
man
soni
–spe
cific
inte
rleu
kin
(IL)–
5an
dIL
-13
conc
entr
atio
nsbe
fore
and
1m
onth
and
5m
onth
saf
ter
praz
iqua
ntel
(PZQ
)tr
eatm
ent
ofS.
man
soni
-in
fect
edH
IV-1
–pos
itive
adul
tsw
ithde
tect
able
base
line
S.m
anso
ni–s
peci
ficre
spon
ses.
Cyt
okin
e,m
easu
rem
ent
SW
A-s
peci
ficre
spon
sea
SE
A-s
peci
ficre
spon
seb
PH
A-in
duce
dre
spon
sec
Pre
-PZQ
1m
onth
5m
onth
sP
re-P
ZQ1
mon
th5
mon
ths
Pre
-PZQ
1m
onth
5m
onth
s
IL-5 C
once
ntra
tion,
med
ian
(inte
rqua
rtile
rang
e),p
g/m
L13
1(5
6–45
8)61
5(2
22–1
534)
289
(8–1
223)
133
(20–
335)
445
(3–1
420)
160
(0–1
022)
124
(0–1
063)
125
(0–1
274)
162
(0–1
209)
Wilc
oxon
sign
edra
nkP
valu
ed.0
001
.005
.01
.50
.23
.05
IL-1
3
Con
cent
ratio
n,m
edia
n(in
terq
uart
ilera
nge)
,pg/
mL
26(5
–159
)13
8(1
2–51
3)79
(0–4
81)
23(8
–191
)11
4(7
–492
)48
(0–2
93)
177
(22–
841)
207
(17–
819)
327
(21–
940)
Wilc
oxon
sign
edra
nkP
valu
ed.0
01.0
2.0
7.9
2.1
4.0
3
NO
TE
.P
HA
,ph
ytoh
emag
glut
inin
;SE
A,
S.
man
soni
egg
antig
en;S
WA
,S
.m
anso
niad
ult
wor
man
tigen
.a
Sam
ple
size
s:IL
-5,
;IL
-13,
.n
p32
np
38b
Sam
ple
size
s:IL
-5,
;IL
-13,
.n
p20
np
24c
Sam
ple
incl
udes
all9
0su
bjec
tsw
ithco
mpl
ete
cyto
kine
data
atal
ltim
epo
ints
.d
Pva
lues
com
pare
follo
w-u
pco
ncen
trat
ions
with
pre–
PZQ
trea
tmen
tco
ncen
trat
ions
.
by guest on August 16, 2016
http://jid.oxfordjournals.org/D
ownloaded from

S. mansoni Treatment in HIV-1 Coinfection • JID 2005:191 (15 May) • 1655
observed during the preceding month. These results do not
support the hypothesis that treatment of S. mansoni infection
has a beneficial effect on HIV-1 disease progression in people
coinfected with HIV-1.
There is evidence that HIV-1 infection may suppress fecal
egg excretion and result in unreliable egg counts [17]; however,
we were unable to replicate this finding in the present study.
We found good correlations between CAA concentrations and
egg counts and found no association between infection intensity
and CD4+ T lymphocyte count.
A preliminary study of this cohort demonstrated a similar
increase in viral load 1 month after antihelminthic therapy in
39 helminth-infected subjects, 28 of whom were infected with
S. mansoni and received praziquantel [21]. In the present study,
an increase in viral load was seen only after praziquantel treat-
ment and not after albendazole treatment; a slight decline in
viral load in the month before praziquantel treatment was seen
in the subjects coinfected with other helminths. Furthermore,
greater increases in viral load were seen in subjects with higher-
intensity S. mansoni infections. Similarly, greater declines in
CD4+ T lymphocyte count were seen after praziquantel treat-
ment than after albendazole treatment, and slightly greater de-
clines were seen in subjects with higher-intensity S. mansoni
infections. The transient nature of the increase in viral load
and the absence of an increase during the preceding month
suggest an effect of praziquantel rather than merely the natural
progression of HIV-1 disease. CD4+ T lymphocyte counts de-
clined at 5 months after treatment, as expected with HIV-1
progression and as seen among helminth-free subjects (from
265 to 244 cells/mL; ), but the rate of decline appearedP ! .0001
to be greater during the first month after treatment; further-
more, we have already reported greater declines in CD4+ T
lymphocyte count in those whose S. mansoni infections had
cleared at follow-up than in those with persistent infection [6].
Reinfection with S. mansoni or other helminths during the
follow-up period, the high level of comorbidity in subjects with
advanced immunosuppression, or other factors may have over-
whelmed the praziquantel treatment–induced effects and, thus,
may explain the transient nature of the changes we observed.
Increases in S. mansoni–specific type 2 cytokine responses
after praziquantel treatment have been described. It is believed
that such treatment-induced responses may be useful in gen-
erating protective antischistosomal immunity [9, 25]. Studies
have demonstrated that HIV-1 infection suppresses posttreat-
ment cytokine responses, which may partly explain increased
reinfection rates among HIV-1–infected adults [14, 26, 27]. In
this study, concentrations of SWA-specific IL-5 and IL-13 and
SEA-specific IL-5 increased significantly after treatment, as did
the proportion of subjects with detectable SWA-specific IL-4,
IL-5, and IL-13 and SEA-specific IL-5 responses; the absence
of significant increases in posttreatment SEA-specific IL-4 and
IL-13 responses may partly reflect smaller numbers of subjects
but does accord with findings in HIV-1–negative subjects [9].
The cohort in our study comprised subjects with varying de-
grees of HIV-1–induced immunosuppression, and the expected
increase in posttreatment responses was observed even in sub-
jects with lower CD4+ T lymphocyte counts (data not shown).
We did not have an HIV-1–negative control group for assess-
ment of whether posttreatment responses were suppressed by
HIV-1 infection, and comparison of our study with previous
studies suggests that the magnitude of the responses may be
smaller in the present study. Nevertheless, these findings do
demonstrate that, despite immunological impairment, there is
still a type 2 cytokine increase after praziquantel treatment.
Despite the increase in type 2 cytokine responses to S. mansoni
antigens, no concomitant increase in eosinophil counts was ob-
served. This was despite preserved posttreatment responses of
IL-5, a cytokine implicated in eosinophil production [28]. This
may be a matter of timing: previous data have shown an increase
in eosinophil counts at 3 weeks after treatment [11]. Alternatively,
it may be a real effect of HIV-1 immunosuppression. Data show-
ing failure to augment antibody responses in HIV-1–infected
subjects suggest that some posttreatment type 2 responses are
impaired [14]; presumably, mechanisms of eosinophil prolifer-
ation independent of—or requiring higher levels of—type 2 cy-
tokines explain this impaired eosinophil production.
Some studies have demonstrated enhanced type 2 and im-
paired type 1 cytokine responses to unrelated antigens in hel-
minth-infected subjects [29–31]. Assessment of the effect of
helminth infection on HIV-1–specific immune responses is ham-
pered by the low numbers of HIV-1–specific CD4+ T lympho-
cytes in subjects with chronic HIV-1 infection. Studies have
more successfully explored the impact of helminth infection
and treatment on response to mycobacterial antigens. Many
have failed to demonstrate any type 2 bias in the immune
response to mycobacterial antigens but have shown impaired
type 1 responses to these antigens [32–36]. M. tuberculosis–
specific responses in our study were infrequent (a response to
any cytokine assessed was detectable in only 45% of subjects),
which reflects the level of immunodeficiency in the cohort. No
evidence of increased M. tuberculosis–specific type 2 cytokine
or reduced IFN-g production was seen after praziquantel treat-
ment, despite the increase in schistosome-specific type 2 cy-
tokine responses.
How might praziquantel treatment of HIV-1–positive people
with schistosomiasis lead to an increase in viral load? The lack
of evidence of suppression of responses to mycobacterial an-
tigens suggests that suppression of beneficial, type 1 responses
to HIV-1 may be an unlikely explanation. However, an increase
in viral load concomitant with the increase in schistosome type
2 cytokine responses after praziquantel treatment could be due
to increased HIV-1 replication in activated Th2 lymphocytes
by guest on August 16, 2016
http://jid.oxfordjournals.org/D
ownloaded from

1656 • JID 2005:191 (15 May) • Brown et al.
[3]. Alternatively, the demonstrated decline in serum IL-10
concentrations after treatment may be relevant: IL-10 is known
to suppress immune activation and type 2 responses and is
capable of inhibiting viral replication by binding the long ter-
minal repeat sequence of proviral HIV-1 DNA [37–39]. The
role of IL-10 in suppressing host immune responses in helminth
infections is the subject of increasing interest, with the discovery
of regulatory T cells expressing this cytokine [40–42]. A decline
in regulatory T cell activity after antischistosomal treatment
could also explain the IL-10 decline, but very little is known
about the role of these cells in HIV-1 disease pathogenesis.
Available evidence suggests that they suppress antiviral immune
responses and, therefore, might enhance viral replication [43].
Examination of the regulatory T cell axis should be the subject
of specific consideration in future studies of the impact of
helminth infection on HIV-1 disease progression.
Acknowledgments
We thank the staff and clients of the AIDS Support Organisation andUganda Virus Research Institute clinics, as well as Sam Rowland andJacqueline Kyosimiire, for viral load quantification.
References
1. Clerici M, Balotta C, Meroni L, et al. Type 1 cytokine production andlow prevalence of viral isolation correlate with long-term nonprogressionin HIV infection. AIDS Res Hum Retroviruses 1996; 12:1053–61.
2. Ostrowski MA, Gu JX, Kovacs C, Freedman J, Luscher MA, MacDonaldKS. Quantitative and qualitative assessment of human immunodefi-ciency virus type 1 (HIV-1)–specific CD4+ T cell immunity to gag inHIV-1–infected individuals with differential disease progression: recip-rocal interferon-g and interleukin-10 responses. J Infect Dis 2001; 184:1268–78.
3. Maggi E, Mazzetti M, Ravina A, et al. Ability of HIV to promote aTH1 to TH0 shift and to replicate preferentially in TH2 and TH0 cells.Science 1994; 265:244–8.
4. Bentwich Z, Maartens G, Torten D, Lal AA, Lal RB. Concurrent in-fections and HIV pathogenesis. AIDS 2000; 14:2071–81.
5. Fincham JE, Markus MB, Adams VJ. Could control of soil-transmittedhelminthic infection influence the HIV/AIDS pandemic. Acta Trop 2003;86:315–33.
6. Brown M, Kizza M, Watera C, et al. Helminth infection is not associatedwith faster progression of HIV disease in coinfected adults in Uganda.J Infect Dis 2004; 190:1869–79.
7. Lawn SD, Karanja DM, Mwinzia P, et al. The effect of treatment ofschistosomiasis on blood plasma HIV-1 RNA concentration in coin-fected individuals. AIDS 2000; 14:2437–43.
8. Fallon P, Fookes R, Wharton G. Temporal differences in praziquantel-and oxamniquine-induced tegumental damage to adult Schistosomamansoni: implications for drug-antibody synergy. Parasitology 1996;112:47–58.
9. Joseph S, Jones FM, Walter K, et al. Increases in human T helper 2cytokine responses to Schistosoma mansoni worm and worm tegumentantigens are specifically induced by treatment with praziquantel. J In-fect Dis 2004; 190:835–42.
10. Grogan JL, Kremsner PG, Deelder AM, Yazdanbakhsh M. Elevatedproliferation and interleukin-4 release from CD4+ cells after chemo-therapy in human Schistosoma haematobium infection. Eur J Immunol1996; 26:1365–70.
11. Kimani G, Chunge CN, Butterworth AE, et al. Eosinophilia and eo-sinophil helminthotoxicity in patients treated for Schistosoma mansoniinfections. Trans R Soc Trop Med Hyg 1991; 85:489–92.
12. Butterworth AE, Capron M, Cordingley JS, et al. Immunity after treat-ment of human schistosomiasis mansoni. II. Identification of resistantindividuals, and analysis of their immune responses. Trans R Soc TropMed Hyg 1985; 79:393–408.
13. Satti M, Cahen P, Skov P, et al. Changes in IgE- and antigen-dependenthistamine-release in peripheral blood of Schistosoma mansoni-infectedUgandan fishermen after treatment with praziquantel. BMC Immunol2004; 5:6.
14. Joseph S, Jones FM, Laidlaw ME, et al. Impairment of the Schistosomamansoni–specific immune responses elicited by treatment with prazi-quantel in Ugandans with HIV-1 co-infection. J Infect Dis 2004; 190:613–8.
15. French N, Nakiyingi J, Carpenter LM, et al. 23-valent pneumococcalpolysaccharide vaccine in HIV-1-infected Ugandan adults: double-blind,randomised and placebo controlled trial. Lancet 2000; 355:2106–11.
16. Deelder AM, De Jonge N, Boerman OC, et al. Sensitive determinationof circulating anodic antigen in Schistosoma mansoni infected individ-uals by an enzyme-linked immunosorbent assay using monoclonalantibodies. Am J Trop Med Hyg 1989; 40:268–72.
17. Karanja DM, Colley DG, Nahlen BL, Ouma JH, Secor WE. Studies onschistosomiasis in western Kenya: I. Evidence for immune-facilitatedexcretion of schistosome eggs from patients with Schistosoma mansoniand human immunodeficiency virus coinfections. Am J Trop Med Hyg1997; 56:515–21.
18. Melrose WD, Turner PF, Pisters P, Turner B. An improved Knott’sconcentration test for the detection of microfilariae. Trans R Soc TropMed Hyg 2000; 94:176.
19. Brown M, Bukusuba J, Hughes P, et al. Screening for intestinal helminthinfestation in a semi-urban cohort of HIV-infected people in Uganda:a combination of techniques may enhance diagnostic yield in the ab-sence of multiple stool samples. Trop Doct 2003; 33:72–6.
20. Elliott AM, Hurst TJ, Balyeku MN, et al. The immune response toMycobacterium tuberculosis in HIV-infected and uninfected adults inUganda: application of a whole blood cytokine assay in an epidemi-ological study. Int J Tuberc Lung Dis 1999; 3:239–47.
21. Elliott AM, Mawa PA, Joseph S, et al. Associations between helminthinfection and CD4+ T cell count, viral load and cytokine responses inHIV-1-infected Ugandan adults. Trans R Soc Trop Med Hyg 2003; 97:103–8.
22. Hodsdon WS, Luzze H, Hurst TJ, et al. HIV-1–related pleural tuber-culosis: elevated production of IFN-g, but failure of immunity to My-cobacterium tuberculosis. AIDS 2001; 15:467–75.
23. Huygen K, van Vooren JP, Turneer M, Bosmans R, Dierckx P, De BruynJ. Specific lymphoproliferation, gamma-interferon production and se-rum immunoglobulin G directed against a purified 32-kDa mycobac-terial antigen (P32) in patients with active tuberculosis. Scand J Im-munol 1988; 27:187–94.
24. Webster M, Libranda-Ramirez BD, Aligui GD, et al. The influence ofsex and age on antibody isotype responses to Schistosoma mansoni andSchistosoma japonicum in human populations in Kenya and the Phil-ippines. Parasitology 1997; 114:383–93.
25. Woolhouse ME, Hagan P. Seeking the ghost of worms past. Nat Med1999; 5:1225–7.
26. Mwinzi PN, Karanja DM, Colley DG, Orago AS, Secor WE. Cellularimmune responses of schistosomiasis patients are altered by human im-munodeficiency virus type 1 coinfection. J Infect Dis 2001; 184:488–96.
27. Karanja DM, Hightower AW, Colley DG, et al. Resistance to reinfectionwith Schistosoma mansoni in occupationally exposed adults and effectof HIV-1 co-infection on susceptibility to schistosomiasis: a longitu-dinal study. Lancet 2002; 360:592–6.
28. Sanderson C. Interleukin-5, eosinophils, and disease. Blood 1992; 79:3101–9.
29. Sabin EA, Araujo MI, Carvalho EM, Pearce EJ. Impairment of tetanus
by guest on August 16, 2016
http://jid.oxfordjournals.org/D
ownloaded from

S. mansoni Treatment in HIV-1 Coinfection • JID 2005:191 (15 May) • 1657
toxoid-specific Th1-like immune responses in humans infected withSchistosoma mansoni. J Infect Dis 1996; 173:269–72.
30. Kamal SM, Bianchi L, Al Tawil A, et al. Specific cellular immuneresponse and cytokine patterns in patients coinfected with hepatitis Cvirus and Schistosoma mansoni. J Infect Dis 2001; 184:972–82.
31. Satoh M, Tsukidate S, Fujita K, Yamamoto K. Strongyloidiasis influ-ences the elevation of adult T-cell leukemia-associated antigen antibodytiter. Int Arch Allergy Appl Immunol 1991; 96:95–6.
32. Cooper PJ, Guderian RH, Nutman TB, Taylor DW. Human infectionwith Onchocerca volvulus does not affect the T-helper cell phenotypeof the cellular immune response to mycobacterial antigen. Trans R SocTrop Med Hyg 1997; 91:350–2.
33. Malhotra I, Mungai P, Wamachi A, et al. Helminth- and Bacillus Cal-mette-Guerin-induced immunity in children sensitized in utero to fil-ariasis and schistosomiasis. J Immunol 1999; 162:6843–8.
34. Borkow G, Leng Q, Weisman Z, et al. Chronic immune activationassociated with intestinal helminth infections results in impaired signaltransduction and anergy. J Clin Invest 2000; 106:1053–60.
35. Elias D, Wolday D, Akuffo H, Petros B, Bronner U, Britton S. Effectof deworming on human T cell responses to mycobacterial antigensin helminth-exposed individuals before and after bacille Calmette-Guerin (BCG) vaccination. Clin Exp Immunol 2001; 123:219–25.
36. Sartono E, Kruize YC, Kurniawan A, et al. Elevated cellular immuneresponses and interferon-gamma release after long-term diethylcar-bamazine treatment of patients with human lymphatic filariasis. J InfectDis 1995; 171:1683–7.
37. Hoffmann KF, Cheever AW, Wynn TA. IL-10 and the dangers of im-mune polarization: excessive type 1 and type 2 cytokine responses in-
duce distinct forms of lethal immunopathology in murine schistoso-miasis. J Immunol 2000; 164:6406–16.
38. Schottelius AJ, Mayo MW, Sartor RB, Baldwin AS Jr. Interleukin-10signaling blocks inhibitor of kB kinase activity and nuclear factor kBDNA binding. J Biol Chem 1999; 274:31868–74.
39. Duh EJ, Maury WJ, Folks TM, Fauci AS, Rabson AB. Tumor necrosisfactor a activates human immunodeficiency virus type 1 through in-duction of nuclear factor binding to the NF-kB sites in the long terminalrepeat. Proc Natl Acad Sci USA 1989; 86:5974–8.
40. Doetze A, Satoguina J, Burchard G, et al. Antigen-specific cellularhyporesponsiveness in a chronic human helminth infection is mediatedby Th3/Tr1-type cytokines IL-10 and transforming growth factor-b butnot by a Th1 to Th2 shift. Int Immunol 2000; 12:623–30.
41. Groux H, O’Garra A, Bigler M, et al. A CD4+ T-cell subset inhibitsantigen-specific T-cell responses and prevents colitis. Nature 1997; 389:737–42.
42. O’Garra A, Vieira P. Regulatory T cells and mechanisms of immune
system control. Nat Med 2004; 10:801–5.
43. Aandahl EM, Michaelsson J, Moretto W, Hecht FM, Nixon DF. Human
CD4+ CD25+ regulatory T cells control T-cell responses to human
immunodeficiency virus and cytomegalovirus antigens. J Virol 2004;
78:2454–9.
by guest on August 16, 2016
http://jid.oxfordjournals.org/D
ownloaded from
Related Documents







![HELMINTH PARASITES IN MAMMALSparasite.org.au/para-site/text/helminth-checklist.pdf · HELMINTH PARASITES IN MAMMALS ... Subclass: EUTHERIA [placental mammals] ... NEM:Asc Ascaris](https://static.cupdf.com/doc/110x72/5ad4fa137f8b9a5d058c90e9/helminth-parasites-in-parasites-in-mammals-subclass-eutheria-placental-mammals.jpg)

