1 Transthyretin blocks retinol uptake and cell signalling by the holo-retinol-binding protein 1 receptor STRA6 2 3 4 5 Daniel C. Berry 1,2 , Colleen M. Croniger 2 , Norbert B. Ghyselinck 3 , and Noa Noy 1,2* 6 7 1 Departments of Pharmacology and 2 Nutrition, Case Western Reserve University School of 8 Medicine, Cleveland, OH 44106, USA, 3 Institut de Génétique et de Biologie Moléculaire et 9 Cellulaire (IGBMC), Centre National de la Recherche Scientifique (CNRS UMR7104), Institut 10 National de la Santé et de la Recherche Médicale (INSERM) U964, Université de Strasbourg, 11 Illkirch, France 12 13 14 Running Title: TTR inhibits STRA6 activities 15 16 17 * Address correspondence to this author at Department of Pharmacology, Case Western Reserve 18 University School of Medicine, 10900 Euclid Ave. W333, Cleveland, OH 44106. Tel: 216-368- 19 0302, Fax: 216-368-1300, E. mail: [email protected] 20 21 22 Copyright © 2012, American Society for Microbiology. All Rights Reserved. Mol. Cell. Biol. doi:10.1128/MCB.00775-12 MCB Accepts, published online ahead of print on 23 July 2012 on June 13, 2016 by guest http://mcb.asm.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1
Transthyretin blocks retinol uptake and cell signalling by the holo-retinol-binding protein 1
receptor STRA6 2
3
4
5
Daniel C. Berry1,2, Colleen M. Croniger2, Norbert B. Ghyselinck3, and Noa Noy1,2* 6
7
1Departments of Pharmacology and 2Nutrition, Case Western Reserve University School of 8
Medicine, Cleveland, OH 44106, USA, 3Institut de Génétique et de Biologie Moléculaire et 9
Cellulaire (IGBMC), Centre National de la Recherche Scientifique (CNRS UMR7104), Institut 10
National de la Santé et de la Recherche Médicale (INSERM) U964, Université de Strasbourg, 11
Illkirch, France 12
13
14
Running Title: TTR inhibits STRA6 activities 15
16
17
*Address correspondence to this author at Department of Pharmacology, Case Western Reserve 18
University School of Medicine, 10900 Euclid Ave. W333, Cleveland, OH 44106. Tel: 216-368-19
0302, Fax: 216-368-1300, E. mail: [email protected] 20
21
22
Copyright © 2012, American Society for Microbiology. All Rights Reserved.Mol. Cell. Biol. doi:10.1128/MCB.00775-12 MCB Accepts, published online ahead of print on 23 July 2012
on June 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

2
Abstract 23
Vitamin A is secreted from cellular stores and circulates in blood bound to retinol binding 24
protein (RBP). In turn, holo-RBP associates in plasma with transthyretin (TTR) to form a ternary 25
RBP-retinol-TTR complex. It is believed that binding to TTR prevents the loss of RBP by 26
filtration in the kidney. At target cells, holo-RBP is recognized by STRA6, a plasma membrane 27
protein that serves a dual role: it mediates uptake of retinol from extracellular RBP into cells and 28
it functions as a cytokine receptor that, upon binding holo-RBP, triggers a JAK/STAT signalling 29
cascade. We previously showed that STRA6-mediated signalling underlies the ability of RBP to 30
induce insulin resistance. However, the role that TTR, the binding partner of holo-RBP in blood, 31
in STRA6-mediated activities remained unknown. Here we show that TTR blocks the ability of 32
holo-RBP to associate with STRA6 and thereby effectively suppresses both STRA6-mediated 33
retinol uptake and STRA6-initiated cell signalling. Consequently, TTR protects mice from RBP-34
induced insulin resistance reflected by reduced phosphorylation of insulin receptor and glucose 35
tolerance tests. The data indicate that STRA6 functions only under circumstances where plasma 36
RBP levels exceeds that of TTR and demonstrate that, in addition to preventing the loss of RBP, 37
TTR plays a central role in regulating holo-RBP/STRA6 signalling. 38
39
40
on June 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

3
Introduction 41
Vitamin A (retinol, ROH) plays critical roles both in the embryo and in the adult where it 42
regulates multiple cellular processes and is essential for embryonic development, reproduction, 43
immune function and vision (29, 32, 33). The vitamin exerts many of its biological activities by 44
giving rise to active metabolites: the visual chromophore 11-cis-retinaldehyde and retinoic acid 45
(RA), which regulates gene transcription by activating specific nuclear receptors (11, 27). ROH 46
is stored in various tissues, including white adipose tissue (WAT), lung, and retinal pigment 47
epithelium in the eye but its main storage site is the liver. ROH is secreted from storage into the 48
circulation bound to retinol-binding protein (RBP), a 21 KDa polypeptide that contains one 49
binding site for ROH. In most mammals, ROH-bound RBP (holo-RBP) does not circulate alone 50
but is associated with another protein called transthyretin (TTR), a 56 KDa homotetramer that, in 51
addition to associating with RBP, functions as a carrier for thyroid hormones (23, 24). ROH thus 52
reaches target tissues bound in a holo-RBP-TTR complex that, under normal circumstances, 53
displays a 1:1 molar stoichiometry. It is believed that binding of RBP to TTR serves to prevent 54
the loss of the smaller protein from blood by filtration in the glomeruli. The concentration of the 55
holo-RBP-TTR complex in plasma is kept constant at 1-2 µM except in extreme cases of vitamin 56
A deficiency or in disease states. Notably, RBP levels are markedly elevated in blood of obese 57
mice and humans and it was reported that, under these circumstances, the protein induces insulin 58
resistance (35). 59
Association with the TTR-RBP complex allows the poorly soluble ROH to circulate in 60
blood but the vitamin dissociates from RBP prior to entering cells. It was proposed that, due to 61
its hydrophobic nature, ROH can readily move from extracellular RBP into cells by diffusion 62
across the plasma membranes at fluxes that are dictated by its extra-to-intra-cellular 63
on June 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

4
concentration gradient (10, 14, 20, 21). However, it has also been suggested that uptake of ROH 64
from circulating holo-RBP is mediated by a cell-surface receptor (13, 28). Indeed, a plasma 65
membrane protein termed STRA6 (Stimulated by Retinoic Acid 6) was found to bind holo-RBP 66
and transport ROH into cells (15). Our recent studies revealed that, in addition to its function as 67
an ROH transporter, STRA6 is a cytokine receptor. We thus found that binding of holo-RBP 68
triggers phosphorylation of a tyrosine residue in the cytosolic domain of STRA6, resulting in 69
recruitment and activation of the Janus kinas JAK2 and, in a cell-dependent manner, the 70
transcription factors STAT3 or STAT5. Holo-RBP thus activates STRA6-mediated signalling 71
that culminates in upregulation of STAT target genes (2, 4). As STAT target genes in white 72
adipose tissue (WAT) and muscle include Suppressor of cytokine signalling 3 (Socs3), a potent 73
inhibitor of insulin signalling (8), these findings suggested a rationale for understanding how 74
elevated serum levels of RBP in obese animals induce insulin resistance (35). Additional studies 75
showed that activation of STRA6 is triggered not simply by binding of holo-RBP but by a 76
STRA6-mediated translocation of ROH from extracellular holo-RBP to an intracellular acceptor, 77
the retinol-binding protein CRBP-I. Importantly, this movement was found to be critically linked 78
to the intracellular metabolism of ROH (5). The data further established that ROH uptake and 79
signalling by STRA6 are interdependent, i.e. that activation of a JAK2/STAT cascade by the 80
receptor requires ROH uptake and vice versus that phosphorylation of STRA6 is essential for 81
enabling ROH transport to proceed (5). 82
While these recent studies provided surprising new insights into the involvement of 83
STRA6 in vitamin A biology, the role that TTR, the binding partner of holo-RBP in blood, may 84
play in STRA6-mediated functions remained unknown. Here, we show that TTR blocks the 85
ability of holo-RBP to associate with STRA6 and thereby effectively suppresses both STRA6-86
on June 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

5
mediated ROH uptake and STRA6-initiated cell signalling. We show further that, consequently, 87
TTR protects mice from RBP-induced insulin resistance. The data indicate that, in addition to 88
preventing the loss of RBP by filtration in the kidney, TTR plays a central role in regulating 89
holo-RBP/STRA6 signalling. 90
91
Materials and Methods 92
Reagents. Human STRA6 was cloned as N-terminal hexahisitidine tagged protein in a 93
pReciever-M01 vector (Genecopoeia). Point mutations of RBP were generated using the 94
Stratagene Quikchange II mutagenesis kit. Lentiviruses harboring shRNA for STRA6, were 95
purchased from Openbiosystems. Antibodies against actin and hexahistidine were from Santa 96
Cruz biotechnology. Antibodies against JAK2, pJAK2, pAKT, pIR, IR, pSTAT5, STAT5 and 97
phospho-tyrosine were from Cell Signaling. RBP antibodies were purchased from Dako. 98
Retinol and bovine insulin were purchased from Sigma Chemical Co. and retinol was purchased 99
from Calbiochem. 100
Immunoblots and immunoprecipitations were performed as previously described (2). 101
Mouse Studies. Mice with a mixed C57BL/6-129/Sv (50%–50%) genetic background were 102
maintained on a 12 h light and dark cycle on a normal chow diet. Mice were housed according to 103
ARC protocol. The breeding diets (Diet #5P76 from LabDiet) contained 25,000-29,000 IU of 104
vitamin A per kg. The mice had access to water and diet ad libitum. 105
Cells. HepG2 and NIH3T3 cells were cultured in DMEM supplemented with 10% FBS and 10% 106
calf serum, respectively. NIH3T3-L1 preadipocytes were induced to differentiate and 107
differentiation was verified as previously described (3). 108
on June 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

6
Analysis of mRNA. RNA was extracted using TRIZOL (Molecular Research Center). cDNA 109
was generated using GeneAmp RNA PCR (Applied Biosystems). Q-PCR was carried out using 110
TaqMan chemistry and Assays-on-Demand probes (Applied Biosystems) for hSTRA6 111
(Hs00223621_m1), hSOCS3 (Hs02330328_g1), mSOCS3 (Mm01249143), mPPARγ 112
(Mm00440945_m1) and 18s rRNA (4352930). RNA was extracted from lipid tissues and 113
skeletal muscle using RNeasy lipid tissue minikit (Qiagen cat#74804) and RNeasy fibrous tissue 114
minikit (cat#74704). 115
Recombinant proteins. Bacterial expression vectors for histidine-tagged hRBP and hTTR were 116
respectively provided by Lawreen Connors, Boston University School of Medicine and Silke 117
Vogel, Columbia University. RBP and TTR were expressed in E. coli and purified as previously 118
described (see (34) for RBP and (16) for TTR. Purified proteins were dialyzed against 300 mM 119
NaCl, 100 mM Tris pH 7.4, and 5% glycerol. For RBP, the method generates holo-RBP, i.e. an 120
RBP-ROH complex at a 1:1 mole ratio. 121
Vitamin A uptake by cultured cells. Holo-RBP was incubated with 3H-retinol (~5000 122
cpm/nmole) for 2 h on ice. In experiments including TTR, 3H-labeled holo-RBP was 123
precomplexed with TTR prior to experimentation. Cultured cells were placed in serum free 124
media for 12 h prior to experimentation. Cells were then washed three times with phosphate-125
buffered saline (PBS), placed in serum free media and treated with 1 µM 3[H]-RBP-ROH or 126
3[H]-RBP-ROH-TTR for three minutes. Cells were washed twice with PBS and then placed in 127
100% ethanol for ten minutes. The ethanol phase was immediately counted using a scintillation 128
counter (Beckman Coulter LS6500). Protein content of each well was measured using the 129
Bradford assay and used for data normalization. 130
on June 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

7
Uptake of ROH from holo-RBP in vivo. Mice fed a regular chow diet were intraperitoneally 131
injected with 0.2 mg holo-RBP or holo-RBP-TTR containing 0.02 mCi 3H-ROH-RBP (total 132
volume 60 μl). Two hours post injection, a blood sample was collected and hearts were perfused 133
with 0.9% saline to remove blood containing labeled ROH. Tissues were isolated, weighed and 134
stored in liquid nitrogen. Tissues were homogenized in PBS, and homogenates placed in 135
scintillation fluid and counted (Beckman Coulter). Counts were normalized per gram tissue. 136
Glucose tolerance tests (GTT). Mice were fasted for 12 hours and injected intraperitoneally with 137
glucose (2 g kg-1 body weight). Blood was sampled from the tail vein and at 0, 15, 30, 60, and 138
120 minutes using an UltraTouch or Accu-Chek Performa glucometer. 139
Flurorescence titrations. Binding of ROH to RBP or its mutant was monitored by fluorescence 140
titrations as described (7). Prior to titrations, ROH was extracted from purified holo-RBP as 141
described (7). Ligand-binding was monitored by following the increase in ROH fluorescence 142
(λex=325; λem=480nm) that accompanies binding to the protein. Titration curves were 143
analyzed by fitting the data to an equation derived from simple binding theory (19) using the 144
software Origin (MicroCal). 145
Fluorescence anisotropy titrations. Association between holo-RBP and TTR was monitored by 146
by following the increase in fluorescence anisotropy of RBP-bound ROH (λex=325; 147
λem=480nm) that accompanies complex formation. Measurements were carried out using a 148
Photon Technology International Quantamaster spectrofluorometer equipped with Glan-149
Thompson polarizers. 150
151
RESULTS 152
on June 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

8
TTR inhibits STRA6-mediated uptake of ROH from holo-RBP. In most mammals, holo-RBP 153
circulates in blood in complex with transthyretin (TTR). To begin to examine the effect of TTR 154
on STRA6 function, hepatocarcinoma HepG2 cells, which endogenously express STRA6, were 155
used to compare cellular uptake of ROH from holo-RBP and from TTR-bound holo-RBP. 156
Recombinant RBP and TTR were expressed in E. coli and purified (see Materials and Methods). 157
HepG2 cells were treated with RBP complexed with 3H-retinol at a 1 μM concentration, similar 158
to the serum RBP level, or with 1 μM 3H-retinol-labeled RBP complexed with TTR at a 1:1 159
molar stoichiometry, similar to that found in blood (24). Media were removed, cells washed, 160
organic compounds extracted from the cells into ethanol, and the amount of 3H-retinol taken up 161
within the incubation period measured by scintillation counting. The rates of uptake of retinol 162
under the assay conditions were constant during the initial 5 min. (Fig. 1a) and subsequent 163
experiments were carried out with a single 3 min. time point, well within the initial linear rate. 164
The rate of ROH uptake from the holo-RBP-TTR complex was slower than uptake from holo-165
RBP alone (Fig. 1a). Moreover, increasing the TTR/RBP ratio by increasing the concentration of 166
TTR inhibited ROH uptake in a dose dependent manner (Fig. 1b). The dose response of the 167
initial rate of ROH transport from holo-RBP showed a two phase behavior comprised of an 168
initial saturable component, likely attributable to STRA6-mediated uptake, followed by a non-169
saturable phase, reflecting passive diffusion of ROH across the plasma membranes (Fig. 1c). In 170
contrast, uptake of ROH from the holo-RBP-TTR complex displayed a single, non-saturable 171
phase (Fig. 1c). These observations suggest that TTR does not impede the ability of ROH to 172
enter cells by passive diffusion but that it effectively blocks ROH transport mediated by STRA6. 173
In agreement with this notion, increasing the expression level of STRA6 in HepG2 cells (Fig. 1d) 174
facilitated ROH uptake from holo-RBP in a dose-responsive manner but had no effect on 175
on June 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

9
transport of ROH from TTR-bound holo-RBP (Fig. 1e). Also in agreement, decreasing the 176
expression of STRA6 in HepG2 cells (Fig. 1f) or in NIH3T3-L1 adipocytes (Fig. 1g) reduced the 177
rate of ROH uptake from holo-RBP but did not affect uptake from TTR-bound holo-RBP. The 178
observation that, in both cell lines, rates of uptake from the holo-RBP-TTR complex were similar 179
to those observed in the absence of STRA6 supports the conclusion that TTR specifically inhibits 180
STRA6-mediated transport. 181
The effect of TTR on ROH uptake from holo-RBP was then examined in vivo using our 182
newly generated STRA6-null mice (26). Twelve week old WT and STRA6-null male mice were 183
injected intraperitoneally with 3H-ROH-labeled holo-RBP or with holo-RBP complexed with 184
TTR, and ROH uptake into tissues was assessed 2 h later. Uptake of ROH into the STRA6-185
expressing tissues WAT, skeletal muscle, and the eye was modestly but significantly lower in 186
STRA6-null vs. WT mice (Fig. 1h), reflecting that the contribution of STRA6 to overall vitamin 187
A uptake by tissues in vivo is small. ROH uptake from TTR-bound holo-RBP was all but 188
identical to that observed in STRA6-/- animals (Fig. 1h). Neither ablation of STRA6 nor the 189
presence of TTR affected ROH uptake by the liver, an organ that does not express STRA6 (Fig. 190
1h). Hence, TTR specifically inhibits STRA6-mediated uptake of ROH in vivo. 191
192
TTR inhibits the association of holo-RBP with STRA6. STRA6 may bind the ternary RBP-193
ROH-TTR complex or, alternatively, it may recognize only free holo-RBP. To dissect between 194
these possibilities, we considered that, unlike in most mammals, holo-RBP in zebrafish (Danio 195
rerio) does not associate with TTR. Thus, presumably, zebrafish STRA6 does not contain a 196
TTR-binding region and, while ROH uptake by the mammalian STRA6 may involve recognition 197
of TTR, ROH uptake by zebrafish STRA6 (dSTRA6) will not. In these experiments, NIH3T3 198
on June 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

10
fibroblasts, which do not endogenously express STRA6, were used. We previously showed that 199
ROH metabolism is essential both for STRA6-mediated ROH transport and for holo-RBP-200
induced STRA6 signalling (5). Hence, to enable STRA6 action in these cells, an NIH3T3 line in 201
which ROH metabolism is enhanced by stably over-expressing lecithin:ROH-acyltransferase 202
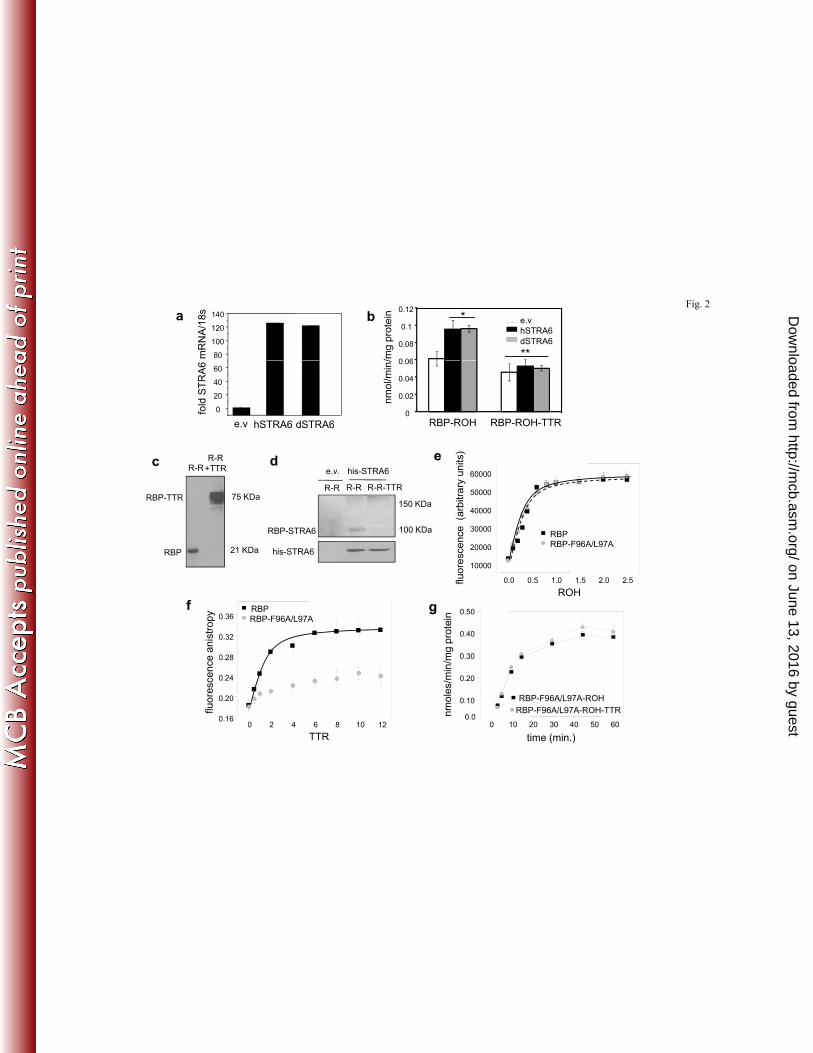
(LRAT), which catalyzes ROH esterification, was generated. Ectopic over-expression of either 203
hSTRA6 or dSTRA6 in LRAT-expressing NIH3T3 fibroblasts enhanced ROH uptake from holo-204
RBP to a similar extent and introduction of TTR similarly decreased the rate of uptake (Fig. 2a, 205
2b). The similarity of the response of dSTRA6, which is unlikely to contain a TTR-binding 206
capability, to that of hSTRA6 suggests that STRA6 in both species recognizes only free and not 207
TTR-bound holo-RBP. 208
The question of whether STRA6 binds free or TTR-bound holo-RBP was then directly 209
assessed. Recombinant holo-RBP was incubated alone or in the presence of TTR with the 210
chemical cross-linker Bis[sulfosuccinimidyl] suberate (0.5 mM, 14 h.), resulting in efficient 211
cross-linking of the holo-RBP-TTR complex (Fig. 2c). The mixtures and additional cross-linker 212
were added to NIH3T3 cells ectopically over-expressing histidine-tagged STRA6. STRA6 was 213
immunoprecipitated, precipitated proteins were resolved by SDS-PAGE, and immunoblotted for 214
RBP-containing complexes (Fig. 2d). Cross-linking of cells with holo-RBP resulted in the 215
appearance of a band with a molecular weight of ~100 KDa, corresponding to that of an RBP-216
bound STRA6. No such band was observed in cells cross-linked with the RBP-ROH-TTR 217
complex and no bands which may correspond to a STRA6-RBP-TTR (~150 KDa) appeared. The 218
data thus indicate that STRA6 associates only with free holo-RBP and that the presence of TTR 219
prevents the association. 220
on June 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

11
To further examine whether TTR inhibits STRA6-mediated ROH uptake by preventing 221
holo-RBP from binding to the receptor, an RBP mutant defective in its ability to bind TTR was 222
generated. The reported three dimensional crystal structure of the holo-RBP-TTR complex 223
suggests that the interactions between the two proteins are mediated by several residues 224
including Phe96 and Leu97 (18). An RBP mutant in which these residues were replaced with 225
alanines (RBP-F96A/L97A) was thus generated. The mutations did not alter the affinity of RBP 226
for retinol (Fig. 2e), indicating that the overall fold of the mutant is intact. As expected, the 227
F96A/L97A mutations disrupted the association of RBP with TTR (Fig. 2f). Measurements of 228
ROH uptake showed that, in contrast with its inhibitory activity on ROH uptake from WT-RBP, 229
TTR had no effect on ROH uptake from RBP-F96A/L97A (Fig. 2g). These observations further 230
establish that TTR inhibits STRA6-mediated ROH uptake by sequestering holo-RBP and not by 231
direct association with the receptor. 232
233
TTR inhibits holo-RBP-induced STRA6 signalling. The effect of TTR of RBP-induced 234
STRA6 signalling was then examined using NIH3T3-L1 adipocytes. We previously showed that 235
in, in these cells, activation of STRA6 by holo-RBP triggers a JAK2/STAT5 cascade to induce 236
the STAT target genes SOCS3 and PPARγ and inhibit insulin responses (2). Preadipocytes 237
NIH3T3-L1 cells were grown two days past confluence and induced to differentiate using a 238
standard hormone mix (10 µg/ml insulin, 0.5 mM 3-isobutyl-1-methylxanthine (IBMX), 0.25 239
mM dexamethasone). Three days later, media were replaced and cells grown for four days. 240
Differentiation was verified by monitoring lipid accumulation and by examining the expression 241
of the adipocyte marker FABP4 (3). As expected, treatment of differentiated adipocytes with 242
holo-RBP (R-R) increased the phosphorylation levels of JAK2 and STAT5 (Fig. 3a). In contrast, 243
on June 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

12
the holo-RBP-TTR complex did not alter the phosphorylation status of these proteins (Fig. 3a). 244
Accordingly, TTR-bound holo-RBP failed to induce the expression of SOCS3 and PPARγ (Fig. 245
3b). To examine the effect of TTR on the ability of holo-RBP to suppress insulin responses, 246
cells were pre-treated with holo-RBP or holo-RBP-TTR for 8 h., treated with insulin for 15 min., 247
and the levels of phosphorylation of the insulin receptor (IR) and its downstream effector AKT 248
were monitored. The data show that inhibition of insulin-induced phosphorylation of IR and 249
AKT by holo-RBP was blunted in the presence of TTR (Fig. 3c). TTR also inhibited the ability 250
of holo-RBP, but not of holo-RBP-F96A/L97A, defective in TTR binding, to trigger STAT5 251
phosphorylation (Fig. 3d) or to induce the expression of SOCS3 in NIH3T3-L1 adipocytes (Fig. 252
3e) or in HepG2 cells (Fig. 3f). 253
The effect of TTR on signaling by the zebrafish STRA6 was then examined. Notably, the 254
phosphotyrosine in the cytosolic domain of STRA6, the STAT recruitment site of the receptor, is 255
present in the dSTRA6, suggesting evolutionary conservation of STRA6 signalling (Fig. 3g). In 256
these experiments, NIH3T3 fibroblasts that ectopically over-express LRAT were transfected 257
with expression vectors for either hSTRA6 or dSTRA6. Treatment of cells expressing either 258
hSTRA6 or dSTRA6 with holo-RBP-induced phosphorylation of STAT3, the preferred STRA6-259
activated STAT in these cells (Fig. 3h) and upregulation of SOCS3 (Fig. 3i). TTR suppressed the 260
ability of holo-RBP to induce STAT3 phosphorylation and to upregulate SOCS3 expression in 261
cells expressing either hSTRA6 or dSTRA6 (Fig. 3h, 3i). 262
263
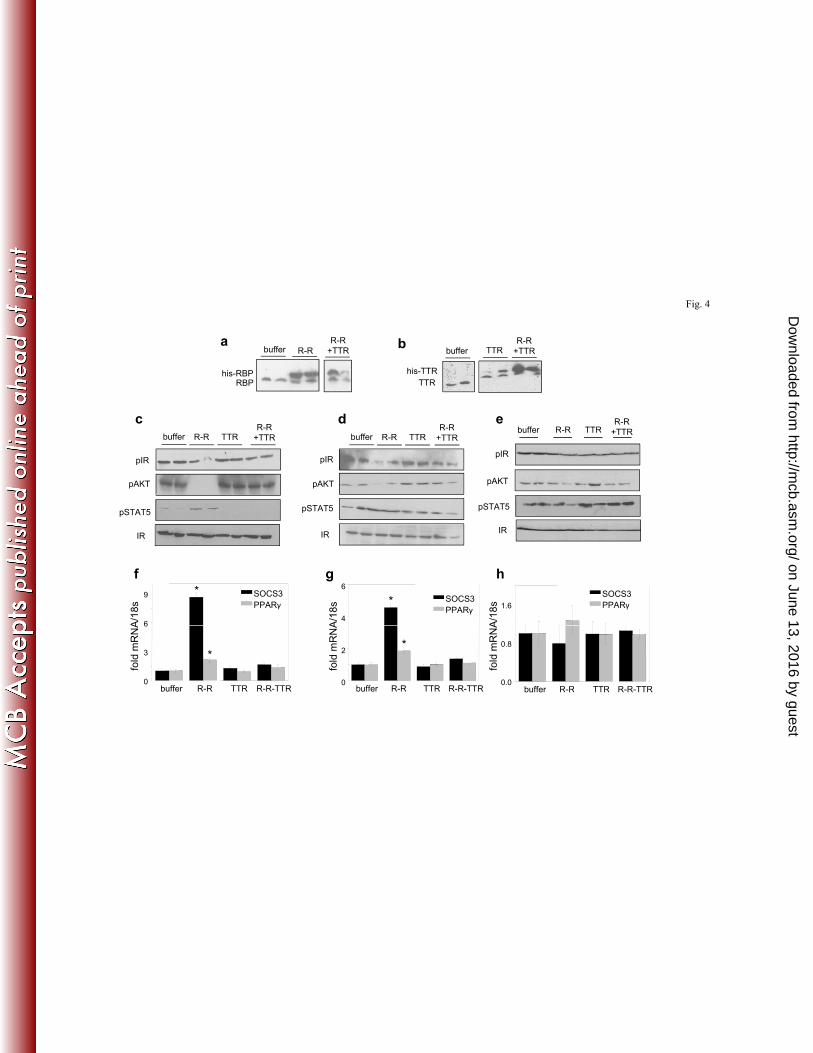
TTR inhibits the ability of holo-RBP to suppress insulin responses in vivo. The effect of TTR 264
on the ability of holo-RBP to promote insulin resistance in vivo was then investigated. Eight 265
week old mice were injected with recombinant holo-RBP, TTR, or holo-RBP-TTR. Mice were 266
on June 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

13
injected three times at 2 h. intervals and sacrificed an hour after the last injection. The 267
treatments resulted in respective elevation of serum levels of RBP, TTR, or both (Fig. 4a, 4b). 268
As expected, treatment of mice with holo-RBP reduced the phosphorylation levels of the insulin 269
receptor and AKT and induced the expression of SOCS3 and PPARγ in WAT (Fig. 4c, 4f) and 270
skeletal muscle (Fig. 4d, 4g) but not in liver (Fig. 4e, 4h). In contrast, treatment with RBP-ROH-271
TTR did not affect the phosphorylation of IR and AKT or the expression levels of the STAT 272
target genes (Fig. 4c-4h). 273
The observations that only free and not TTR-bound holo-RBP activates STRA6 suggest 274
that the serum RBP/TTR ratio is key for regulating STRA6 signalling. In agreement with the 275
report that expression of RBP in adipose tissue increases in obese rodents and humans, resulting 276
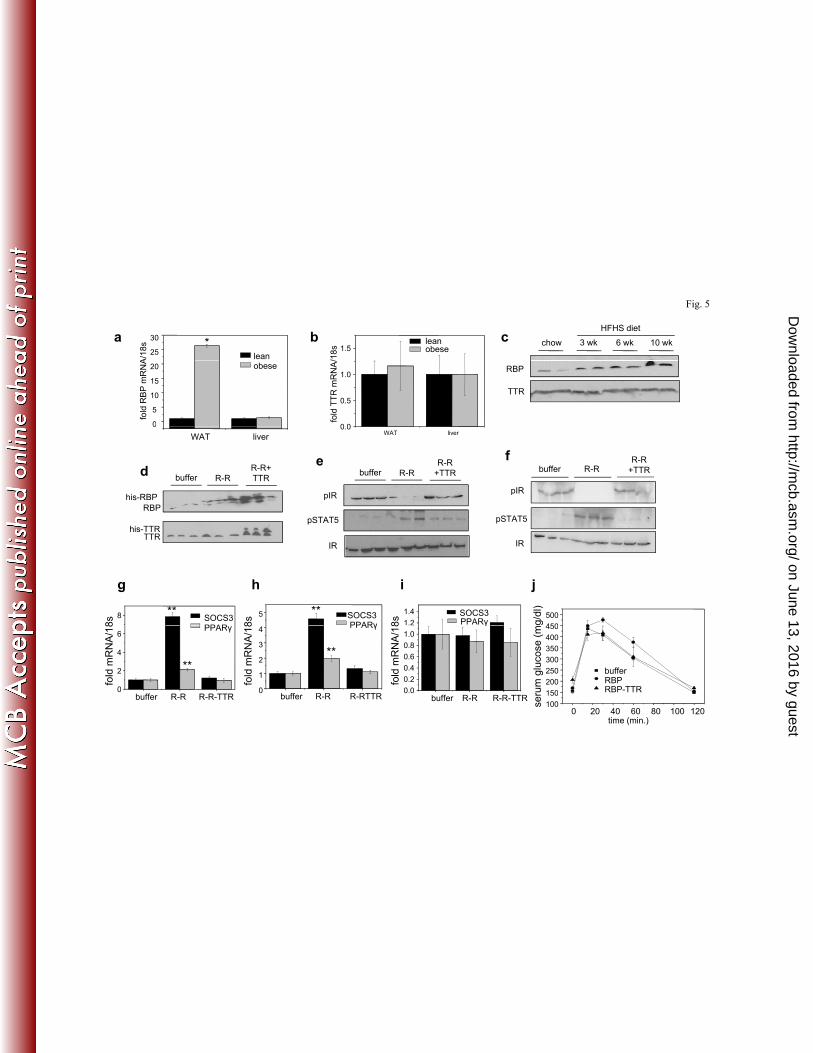
in elevation of serum RBP levels (35), feeding mice a high fat high sucrose (HFHS) diet for 10 277
weeks resulted in upregulation of the expression of RBP in WAT but not in liver (Fig. 5a). In 278
contrast, TTR expression in these organs was not affected by the diet (Fig. 5b). Accordingly, 279
serum level of RBP was markedly elevated while serum level of TTR remained unchanged in 280
obese mice (Fig. 5c). Hence, the RBP/TTR ratio is significantly higher in blood of obese vs. lean 281
mice. 282
To directly determine if TTR prevents holo-RBP- induced insulin resistance, mice were 283
treated with holo-RBP or holo-RBP-TTR for three weeks prior to carrying out glucose tolerance 284
tests (GTT). Mice were treated by implanting alzet osmotic pumps containing the appropriate 285
proteins (1 μM) thereby delivering constant amounts of proteins over the three week period. 286
Similarly to the short-term treatments (Fig. 4), three week treatment of mice with holo-RBP 287
induced phosphorylation of STAT5, reduced the activation level of IR and upregulated SOCS3 288
and PPARγ in WAT (Fig. 5e, 5g) and muscle (Fig. 5f, 5h) but not in the liver (Fig. 5i). In 289
on June 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

14
contrast, treatment with TTR-bound holo-RBP had no effect on the phosphorylation of STAT5 290
or IR and did not alter the expression levels of the STAT target genes (Fig. 5e-5i). Accordingly, 291
while holo-RBP treatment resulted in a sluggish response in GTT, reflecting the development of 292
insulin resistance, treatment with the holo-RBP-TTR complex did not alter the insulin responses 293
of the mice (Fig. 5j). Hence, association with TTR suppresses the ability of holo-RBP to 294
interfere with insulin signaling. 295
296
Discussion 297
298
Upon binding of extracellular holo-RBP, STRA6 transports ROH into cells and it activates a 299
signalling cascade culminating in induction of STAT target genes (4, 5). The observations 300
described here reveal that the binding partner of RBP in blood, TTR, effectively blocks 301
association of holo-RBP with STRA6. Consequently, STRA6 mediates cellular ROH uptake 302
only from free and not from TTR-bound holo-RBP. The data further show that, even in the 303
presence of free holo-RBP, STRA6-mediated ROH uptake by tissues comprises only a small 304
fraction of total uptake by target tissues in vivo (Fig. 1h). The observations thus support the 305
previously proposed model that supply of ROH from circulating holo-RBP or holo-RBP-TTR to 306
cells occurs primarily by diffusion through the plasma membranes (10, 14, 20, 21). Taken 307
together with the observations that ROH transport by STRA6 is critical for enabling activation of 308
STRA6 signalling (5), the data indicate that, with the exception of the eye (26), the main role of 309
ROH transport by STRA6 is not to provide the vitamin to cells but to couple sensing of 310
circulating free holo-ROH levels to cell signalling. It is worth noting that even in the eye, 311
on June 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

15
morphological changes and reduction in visual function in Stra6-null mice are mild, indicating 312
that STRA6 is not the only pathway by which ROH enters the retinal pigment epithelium (26). 313
The data reveal that, in addition to its function in preventing filtration of the 21 KDa RBP 314
in the kidney, TTR plays an important role in protecting cells from holo-RBP-induced signalling 315
mediated by STRA6. The observations that STRA6 “senses” only free and not TTR-bound RBP 316
establish that the receptor functions only under circumstances in which serum RBP level exceeds 317
that of TTR. Such circumstances are encountered, for example, in obese animals in which serum 318
level of RBP is elevated while TTR level is not (Fig. 5c). The circumstances in which the plasma 319
RBP concentration exceeds that of TTR in healthy lean animals remain to be clarified. It is 320
interesting to note in regard to this that it has long been known that insulin responsiveness varies 321
in a circadian fashion (17, 31). The molecular basis for these diurnal variations is incompletely 322
understood but the data presented here raise the intriguing possibility that they may arise from 323
diurnal variations in plasma RBP/TTR ratio. 324
The RBP/TTR ratio in blood may be altered by changes in the expression level of RBP, 325
or TTR, or both. TTR is expressed in the central nervous system and in the liver with the latter 326
serving as the main source for the protein in serum (9). Expression of hepatic TTR is 327
downregulated and, consequently, serum TTR level dramatically decreases during acute phase 328
response (APR), a process characterized by rapid reprogramming of gene expression and 329
metabolism in response to inflammatory cytokine signaling (1, 22). The low serum level of TTR 330
associated with APR may release holo-RBP thereby activating STRA6. Hence, STRA6 331
signalling may play a role in APR. It has also been reported that hepatic TTR expression is 332
regulated by sex hormones (12) and is directly controlled by hepatocyte nuclear factor 4α (HNF-333
4α) (30). The expression of RBP in brown adipose tissue and liver was reported to be regulated 334
on June 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

16
by cAMP-mediated pathways and by the nuclear receptors PPARα and PPARγ (6, 25). Whether, 335
by controlling TTR or RBP expression, these factors regulate the RBP-TTR ratio in blood and 336
thus STRA6 signalling remains to be clarified. 337
Notably, as free holo-RBP is rapidly excreted by glomeruli filtration, its lifetime in serum 338
is short. Holo-RBP thus functions like a classical cytokine: its availability to its membrane 339
receptor is tightly regulated and its signalling activities are constrained by a short half-life in the 340
circulation. These characteristics of the signalling activities of holo-RBP strikingly differ from 341
the characteristics of its role as a shuttling protein which mobilizes ROH from liver stores. 342
Unlike in the former capacity where holo-RBP functions on its own, delivery of ROH to target 343
tissues is mediated by the holo-RBP-TTR complex. The plasma level of this complex is under 344
tight homeostatic control and it provides ROH to target cells to support tissue requirement for 345
vitamin A without the need for a specialized receptor. 346
347
Acknowledgments 348
We are grateful to Michele Mumaw for help in early stages of this work. We thank Lawreen 349
Connors, Boston University School of Medicine, and Silke Vogel, Columbia University School 350
of Physicians and Surgeons, for the TTR and RBP expression constructs. This work was 351
supported by NIH grants DK060684 and CA107013 to N.N. The Mouse Metabolic Phenotyping 352
Center (MMPC) of Case Western Reserve University is supported by NIH grant DK59630. The 353
Stra6-null mouse line was established at the Mouse Clinical Institute (http://www-mci.u-354
strasbg.fr/) in the Genetic Engineering and Model Validation Department with Inserm and FRM 355
(DEQ20071210544) grants to NBG. 356
357
on June 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

17
References 358
359
1. Baumann, H., and J. Gauldie. 1994. The acute phase response. Immunol Today 15:74-80. 360 2. Berry, D. C., H. Jin, A. Majumdar, and N. Noy. 2011. Signaling by vitamin A and retinol-binding 361
protein regulates gene expression to inhibit insulin responses. Proc Natl Acad Sci U S A 362 108:4340-4345. 363
3. Berry, D. C., and N. Noy. 2009. All-trans-retinoic acid represses obesity and insulin resistance by 364 activating both peroxisome proliferation-activated receptor beta/delta and retinoic acid 365 receptor. Mol Cell Biol 29:3286-3296. 366
4. Berry, D. C., and N. Noy. 2012. Signaling by vitamin A and retinol-binding protein in regulation 367 of insulin responses and lipid homeostasis. Biochim Biophys Acta 1821:168-176. 368
5. Berry, D. C., S. M. O'Byrne, A. C. Vreeland, W. S. Blaner, and N. Noy. 2012. Cross-talk between 369 signalling and vitamin A transport by the retinol-binding protein receptor STRA6 Molecular and 370 Cellular Biology in press. 371
6. Bianconcini, A., A. Lupo, S. Capone, L. Quadro, M. Monti, D. Zurlo, A. Fucci, L. Sabatino, A. 372 Brunetti, E. Chiefari, M. E. Gottesman, W. S. Blaner, and V. Colantuoni. 2009. Transcriptional 373 activity of the murine retinol-binding protein gene is regulated by a multiprotein complex 374 containing HMGA1, p54 nrb/NonO, protein-associated splicing factor (PSF) and steroidogenic 375 factor 1 (SF1)/liver receptor homologue 1 (LRH-1). Int J Biochem Cell Biol 41:2189-2203. 376
7. Cogan, U., M. Kopelman, S. Mokady, and M. Shinitzky. 1976. Binding affinities of retinol and 377 related compounds to retinol binding proteins. Eur J Biochem 65:71-78. 378
8. Croker, B. A., H. Kiu, and S. E. Nicholson. 2008. SOCS regulation of the JAK/STAT signalling 379 pathway. Semin Cell Dev Biol 19:414-422. 380
9. Felding, P., and G. Fex. 1982. Cellular origin of prealbumin in the rat. Biochim Biophys Acta 381 716:446-449. 382
10. Fex, G., and G. Johannesson. 1988. Retinol transfer across and between phospholipid bilayer 383 membranes. Biochim Biophys Acta 944:249-255. 384
11. Germain, P., P. Chambon, G. Eichele, R. M. Evans, M. A. Lazar, M. Leid, A. R. De Lera, R. Lotan, 385 D. J. Mangelsdorf, and H. Gronemeyer. 2006. International Union of Pharmacology. LX. Retinoic 386 acid receptors. Pharmacol Rev 58:712-725. 387
12. Goncalves, I., C. H. Alves, T. Quintela, G. Baltazar, S. Socorro, M. J. Saraiva, R. Abreu, and C. R. 388 Santos. 2008. Transthyretin is up-regulated by sex hormones in mice liver. Mol Cell Biochem 389 317:137-142. 390
13. Heller, J. 1975. Interactions of plasma retinol-binding protein with its receptor. Specific binding 391 of bovine and human retinol-binding protein to pigment epithelium cells from bovine eyes. J Biol 392 Chem 250:3613-3619. 393
14. Hodam, J. R., and K. E. Creek. 1998. Comparison of the metabolism of retinol delivered to 394 human keratinocytes either bound to serum retinol-binding protein or added directly to the 395 culture medium. Exp Cell Res 238:257-264. 396
15. Kawaguchi, R., J. Yu, J. Honda, J. Hu, J. Whitelegge, P. Ping, P. Wiita, D. Bok, and H. Sun. 2007. 397 A membrane receptor for retinol binding protein mediates cellular uptake of vitamin A. Science 398 315:820-825. 399
16. Kingsbury, J. S., E. S. Klimtchuk, R. Theberge, C. E. Costello, and L. H. Connors. 2007. 400 Expression, purification, and in vitro cysteine-10 modification of native sequence recombinant 401 human transthyretin. Protein Expr Purif 53:370-377. 402
on June 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

18
17. Lee, A., M. Ader, G. A. Bray, and R. N. Bergman. 1992. Diurnal variation in glucose tolerance. 403 Cyclic suppression of insulin action and insulin secretion in normal-weight, but not obese, 404 subjects. Diabetes 41:750-759. 405
18. Naylor, H. M., and M. E. Newcomer. 1999. The structure of human retinol-binding protein (RBP) 406 with its carrier protein transthyretin reveals an interaction with the carboxy terminus of RBP. 407 Biochemistry 38:2647-2653. 408
19. Norris, A. W., and E. Li. 1998. Fluorometric titration of the CRABPs. Methods Mol Biol 89:123-409 139. 410
20. Noy, N., and Z. J. Xu. 1990. Interactions of retinol with binding proteins: implications for the 411 mechanism of uptake by cells. Biochemistry 29:3878-3883. 412
21. Noy, N., and Z. J. Xu. 1990. Kinetic parameters of the interactions of retinol with lipid bilayers. 413 Biochemistry 29:3883-3888. 414
22. Pepys, M. B., and M. L. Baltz. 1983. Acute phase proteins with special reference to C-reactive 415 protein and related proteins (pentaxins) and serum amyloid A protein. Adv Immunol 34:141-416 212. 417
23. Prapunpoj, P. 2009. Evolutionary changes to transthyretin. FEBS J 276:5329. 418 24. Raz, A., and D. S. Goodman. 1969. The interaction of thyroxine with human plasma prealbumin 419
and with the prealbumin-retinol-binding protein complex. J Biol Chem 244:3230-3237. 420 25. Rosell, M., E. Hondares, S. Iwamoto, F. J. Gonzalez, M. Wabitsch, B. Staels, Y. Olmos, M. 421
Monsalve, M. Giralt, R. Iglesias, and F. Villarroya. 2012. Peroxisome proliferator-activated 422 receptors-alpha and -gamma, and cAMP-mediated pathways, control retinol-binding protein-4 423 gene expression in brown adipose tissue. Endocrinology 153:1162-1173. 424
26. Ruiz, A., M. Mark, H. Jacobs, M. Klopfenstein, J. Hu, M. Lloyd, S. Habib, C. Tosha, R. A. Radu, N. 425 B. Ghyselinck, S. Nusinowitz, and D. Bok. 2012. Retinoid content, visual responses and ocular 426 morphology are compromised in the retinas of mice lacking the retinol-binding protein receptor, 427 STRA6. Invest Ophthalmol Vis Sci. 428
27. Schug, T. T., D. C. Berry, N. S. Shaw, S. N. Travis, and N. Noy. 2007. Opposing effects of retinoic 429 acid on cell growth result from alternate activation of two different nuclear receptors. Cell 430 129:723-733. 431
28. Shingleton, J. L., M. K. Skinner, and D. E. Ong. 1989. Characteristics of retinol accumulation 432 from serum retinol-binding protein by cultured Sertoli cells. Biochemistry 28:9641-9647. 433
29. Wald, G. 1968. The molecular basis of visual excitation. Nature 219:800-807. 434 30. Wang, Z., and P. A. Burke. 2010. Hepatocyte nuclear factor-4alpha interacts with other 435
hepatocyte nuclear factors in regulating transthyretin gene expression. FEBS J 277:4066-4075. 436 31. Whichelow, M. J., R. A. Sturge, H. Keen, R. J. Jarrett, L. Stimmler, and S. Grainger. 1974. Diurnal 437
variation in response to intravenous glucose. Br Med J 1:488-491. 438 32. Wilson, J. G., C. B. Roth, and J. Warkany. 1953. An analysis of the syndrome of malformations 439
induced by maternal vitamin A deficiency. Effects of restoration of vitamin A at various times 440 during gestation. Am J Anat 92:189-217. 441
33. Wolbach, S. B., and P. R. Howe. 1978. Nutrition Classics. The Journal of Experimental Medicine 442 42: 753-77, 1925. Tissue changes following deprivation of fat-soluble A vitamin. S. Burt Wolbach 443 and Percy R. Howe. Nutr Rev 36:16-19. 444
34. Xie, Y., H. A. Lashuel, G. J. Miroy, S. Dikler, and J. W. Kelly. 1998. Recombinant human retinol-445 binding protein refolding, native disulfide formation, and characterization. Protein Expr Purif 446 14:31-37. 447
35. Yang, Q., T. E. Graham, N. Mody, F. Preitner, O. D. Peroni, J. M. Zabolotny, K. Kotani, L. 448 Quadro, and B. B. Kahn. 2005. Serum retinol binding protein 4 contributes to insulin resistance 449 in obesity and type 2 diabetes. Nature 436:356-362. 450
on June 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

19
451
Figure legends 452
453
Figure. 1. TTR inhibits STRA6-mediated uptake of ROH from holo-RBP. a) Uptake of 3H-454
ROH by HepG2 cells treated with RBP-3H-ROH or RBP-3H-ROH-TTR (1 µM) for denoted 455
times. b) Uptake of 3H-ROH by HepG2 cells treated with the denoted concentrations of RBP-3H-456
ROH or RBP-3H-ROH-TTR for 3 min. c) Uptake of 3H-ROH by HepG2 cells following a 3 min. 457
incubation with 1 µM RBP-3H-ROH in the presence of denoted concentrations of TTR. d) 458
Levels of STRA6 mRNA in HepG2 cells transfected with varying amounts of STRA6 cDNA. e) 459
Effect of increasing the expression level of STRA6 in HepG2 cells on uptake of 3H-ROH from 460
RBP-3H-ROH or RBP-3H-ROH-TTR (1 µM, 3 min.). f) Top: expression level of STRA6 in 461
HepG2 cells transfected with an empty vector (e.v.) or vector harboring STRA6shRNA. Bottom: 462
Effect of decreasing the expression level of STRA6 in HepG2 cells on uptake of 3H-ROH from 463
RBP-3H-ROH or from RBP-3H-ROH-TTR (1 µM, 3 min.). g) Top: expression level of STRA6 in 464
NIH3T3-L1 cells transfected with an empty vector (e.v.) or vector harboring STRA6shRNA. 465
Effect of decreasing the expression level of STRA6 in NIH3T3-L1 adipocytes on uptake of 3H-466
ROH from RBP-3H-ROH or from RBP-3H-ROH-TTR (1 µM, 3 min.). h) 12 week old WT and 467
STRA6-null male mice were injected intraperitoneally with RBP-3H-ROH (100 μl, 0.1 mCi, 1 468
µM). 2 h later, tissues were isolated, weighed, homogenized, and 3H-ROH counted. Data are 469
means±SEM, *p<0.01 RBP-ROH- vs. RBP-ROH-TTR-treated groups. All p values were 470
calculated using a two-tailed student t-test. 471
Figure 2. STRA6 does not bind the holo-RBP-TTR complex. a) NIH3T3 cells stably over-472
expressing LRAT were transfected with an e.v, or with expression vectors encoding human 473
on June 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

20
(hSTRA6) or zebrafish (dSTRA6) STRA6, resulting in similar levels of mRNAs. b) Uptake of 474
3H-ROH from RBP-3H-ROH or RBP-3H-ROH-TTR (1 µM, 3 min.) by cells expressing hSTRA6 475
or dSTRA6. c) RBP-ROH or RBP-ROH-TTR (1 μM) was incubated with the chemical cross-476
linker Bis[sulfosuccinimidyl] suberate (0.5 mM) for 14 h. Proteins were resolved by SDS-PAGE 477
and visualized by Coomasie-blue staining. d) Cross-linked complexes and additional cross-linker 478
(0.5 mM) were added to HepG2 cells transfected with an e.v. or with a vector encoding histidine-479
tagged STRA6. Following a 15 min. incubation, his-STRA6 was immunoprecipitated using 480
antibodies against the tag and precipitated RBP and STRA6 visualized by immunoblots. e) 481
Fluorescence titrations of RBP and its F96A/L97A mutant (1 µM) with ROH. Progress of 482
titrations was monitored by following the increase in ROH fluorescence upon binding to the 483
protein (λex - 330 nm; λem - 460 nm). f) Fluorescence anisotropy titrations of holo-RBP and 484
holo-RBP-F96A/L97A (3 µM) with TTR. Progress of titrations was monitored by measuring the 485
fluorescence anisotropy of bound ROH (λex - 330 nm; λem - 460 nm). g) Uptake of 3H-ROH 486
from holo-RBP-F96A/L97A (1 µM, 3 min.) in the presence or absence of TTR. Data are 487
means±SEM, *p<0.01 vs. cells transfected with an empty vector. **p=0.01 vs. cells transfected 488
with an empty vector and treated with RBP-ROH. All p values were calculated using a two-489
tailed student t-test. 490
491
Figure 3. TTR blocks activation of STRA6 signaling by holo-RBP. a) NIH3T3-L1 adipocytes 492
were treated with 1 µM RBP-ROH, TTR, or RBP-ROH-TTR for 15 min. Cell were lyzed and 493
phosphorylated JAK2 (pJAK2) and STAT5 (pSTAT5) visualized by immunoblots. b) NIH3T3-494
L1 adipocytes cells were treated with 1 µM RBP-ROH, TTR, or RBP-ROH-TTR for 4 h and 495
levels of SOCS3 and PPARγ mRNA assessed by Q-PCR. Data are means±SEM. *p<0.001 vs. 496
on June 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

21
non-treated cells. c) NIH3T3-L1 adipocytes were pre-treated with 1 µM RBP-ROH, TTR or 497
RBP-ROH-TTR for 8 h and then treated with insulin (25 nM, 15 min.). Phosphorylated IR (pIR) 498
and AKT (pAKT) were visualized by immunoblots. Bottom panel: quantitation of band 499
intensities. Mean of two independent experiments. d) NIH3T3-L1 adipocytes were treated with 500
RBP-ROH or RBP-F96A/L97A-ROH (RBP96/97-R) in the presence or absence of TTR (1 µM 501
each, 15 min). Lysates were immunoblotted for pSTAT5. e) NIH3T3-L1 adipocytes were 502
treated with RBP-ROH or RBP-F96A/L97A-ROH in the presence or absence of TTR (1 µM 503
each, 4 h). Levels of SOCS3 mRNA were assessed by Q-PCR. Data are means±SEM. *p<0.001 504
vs. non-treated cells, **p<0.001 vs. R-R-TTR-treated cells. f) HepG2 cells were treated with 505
RBP-ROH in the presence or absence of TTR (1 µM each, 4 h). Levels of SOCS3 mRNA were 506
assessed by Q-PCR. Data are means±SEM. *p<0.001 vs. non-treated cells, g) The 507
phosphotyrosine motif in mouse, human and zebrafish STRA6 (mSTRA6, hSTRA6, dSTRA6). 508
h) NIH3T3 fibroblasts stably expressing LRAT were transfected with zebrafish and human 509
STRA6 and treated with 1 µM RBP-ROH or RBP-ROH-TTR for 15 min and lysates were 510
immunoblotted for pSTAT3. i) NIH3T3 fibroblasts stably over-expressing LRAT were 511
transfected with dSTRA6 or hSTRA6 and treated with 1 µM RBP-ROH or RBP-ROH-TTR for 4 512
h. Levels of SOCS3 mRNA were assessed by Q-PCR. Data are means±SEM. *p<0.001 vs. non-513
treated cells. All p values were calculated using a two-tailed student t-test. 514
515
Figure 4. TTR suppress activation of STRA6 by holo-RBP in vivo. Mice were injected three 516
times with 0.1 μmole RBP-ROH or 0.1 μmole RBP-ROH complexed with TTR and sacrificed 1 517
h after the last injection. a, b) Immunoblots of RBP (a) and TTR (b) in serum following 518
respective injections. Blots from 2 mice of each group are shown. c-e) Immunoblots of 519
on June 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

22
phosphorylated insulin receptor (pIR), AKT (pAKT) and STAT5 (pSTAT5) in WAT (c), skeletal 520
muscle (d) and liver (e) of mice treated as denoted. Total IR served as a loading control. f-h) 521
Levels of mRNA of SOCS3 and PPARγ in WAT (f), skeletal muscle (g) and liver (h) of treated 522
mice. Data are means ±SEM *p<0.001 buffer-treated vs. RBP-ROH treated mice. 523
524
Figure 5. TTR is protectives against holo-RBP-induced insulin resistance. a-b) Levels of 525
mRNA of RBP (a) and TTR (b) in WAT and liver of lean mice and of mice fed a HFHS diet for 526
10 weeks (obese). c) Immunoblots of RBP and TTR in serum of mice fed a HFHS diet for 0, 3, 527
6 and 10 weeks. d-j) Mice were implanted with an Alzet pump that contained buffer, 0.1 μM 528
holo-RBP or 0.1 μM holo-RBP complexed with TTR. Implants were replaced once a week for 3 529
weeks. d) Immunoblots of RBP (top) and TTR (bottom) in serum following 3 weeks of denoted 530
treatments. e, f) Immunoblots of pIR and pSTAT5 in WAT (e) and skeletal muscle (f) of mice 531
treated as denoted. g-i) Levels of SOCS3 mRNA in WAT (g), skeletal muscle (h) and liver (i) of 532
mice treated as denoted. j). Glucose tolerance tests carried out following 3 weeks of denoted 533
treatments. Data are means ±SEM. *p<0.001 lean vs. obese mice; **p<0.001 buffer-treated vs. 534
RBP-ROH-treated mice. All p values were calculated using a two-tailed student t-test. 535
on June 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from
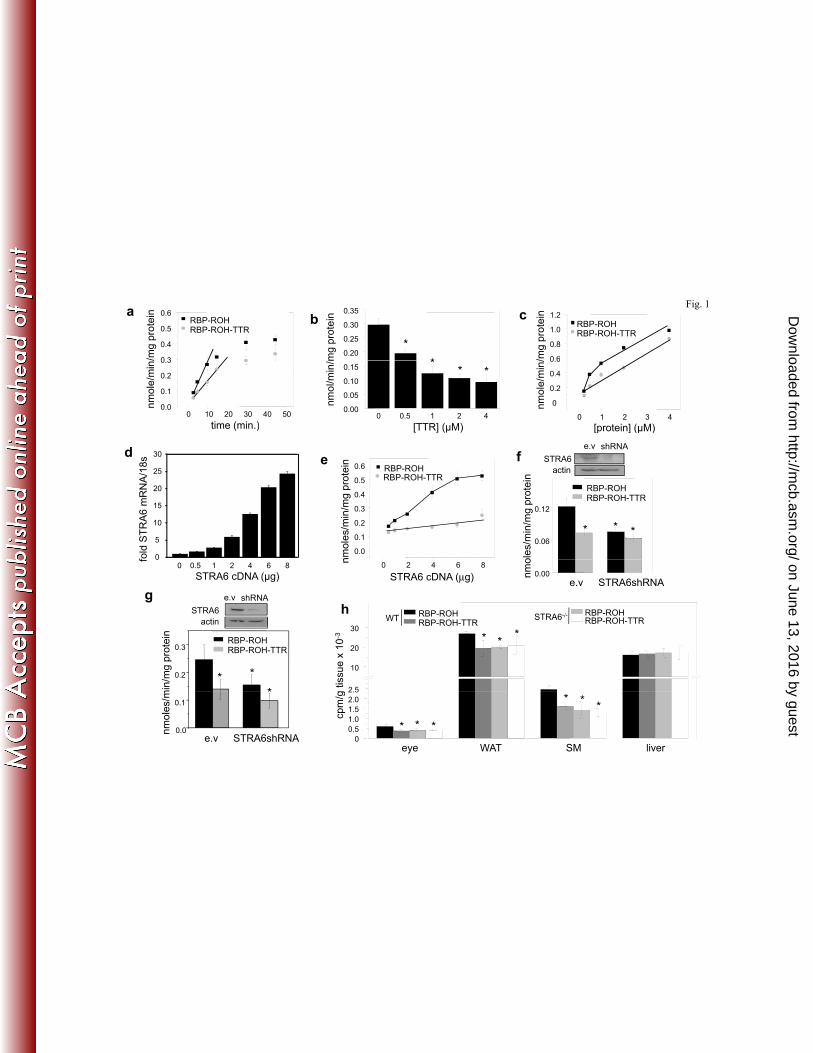
Fig. 1a
0.20
0.25
0.30
0.35
mg
prot
einb
0 3
0.4
0.5
0.6
/mg
prot
ein
RBP-ROHRBP-ROH-TTR
c
/mg
prot
ein
0 6
0.8
1.0
1.2RBP-ROHRBP-ROH-TTR
**
0 0.5 1 2 40.00
0.05
0.10
0.15
nmol
/min
/m
[TTR] (µM)0 10 20 30 40 50
0.0
0.1
0.2
0.3
nmol
e/m
in/
time (min ) [protein] (µM)
nmol
e/m
in/
0 1 2 3 4
0
0.2
0.4
0.6* * *
[TTR] (µM)time (min.) [protein] (µM)
0 4
0.5
0.6
prot
ein
RBP-ROHRBP-ROH-TTR
d f
20
25
30
RN
A/1
8s e STRA6actin
e.v shRNA
otei
n
RBP-ROH
0.0
0.1
0.2
0.3
0.4
nmol
es/m
in/m
g
0
5
10
15
fold
STR
A6 m
R
0.06
0.12
oles
/min
/mg
pro RBP-ROH-TTR
* **
30
RBP-ROHRBP-ROH-TTR
RBP-ROHRBP-ROH-TTRWT STRA6-/-h
0 2 4 6 8n
STRA6 cDNA (μg)
00 0.5 1 2 4 6 8
f
STRA6 cDNA (µg) e.v STRA6shRNA0.00nm
o
gSTRA6
actin
e.v shRNA
2.5
10
20
30RBP ROH TTR
g tis
sue
x 10
-3 * * *
0.2
0.3
min
/mg
prot
ein
RBP-ROHRBP-ROH-TTR
* **
eye WAT SM liver0
0.51.01.52.02.5
cpm
/g
** *
* * *0.0
0.1
nmol
es/m
e.v STRA6shRNA
on June 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from
0 06
0.08
0.1
0.12e.vhSTRA6dSTRA6
/mg
prot
einb
Fig. 2
80
100
120
140
mR
NA
/18sa *
**
0
0.02
0.04
0.06
RBP-ROH RBP-ROH-TTR
nmol
/min
/
e.v hSTRA6 dSTRA60
20
40
60
fold
STR
A6
e
50000
60000
ary
units
)
e.v hSTRA6 dSTRA6
c R-RR-R
+TTR
R-R R-R R-R-TTR
his-STRA6e.v.d
20000
30000
40000
50000
esce
nce
(arb
itr
RBPRBP-F96A/L97A
RBP
RBP-TTR
21 KDa
75 KDa
RBP-STRA6 100 KDa
his-STRA6
150 KDa
0.0 0.5 1.0 1.5 2.0 2.5
10000
fluor
e
ROH
0.36
opy RBPf
RBP-F96A/L97A0.50
eing
0.24
0.28
0.32
esce
nce
anis
tro
0.20
0.30
0.40
es/m
in/m
g pr
ote
0 2 4 6 8 10 120.16
0.20
fluor
e
TTR0 10 20 30 40 50 60
0.10
nmol
e
time (min.)
0.0
RBP-F96A/L97A-ROHRBP-F96A/L97A-ROH-TTR
on June 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from
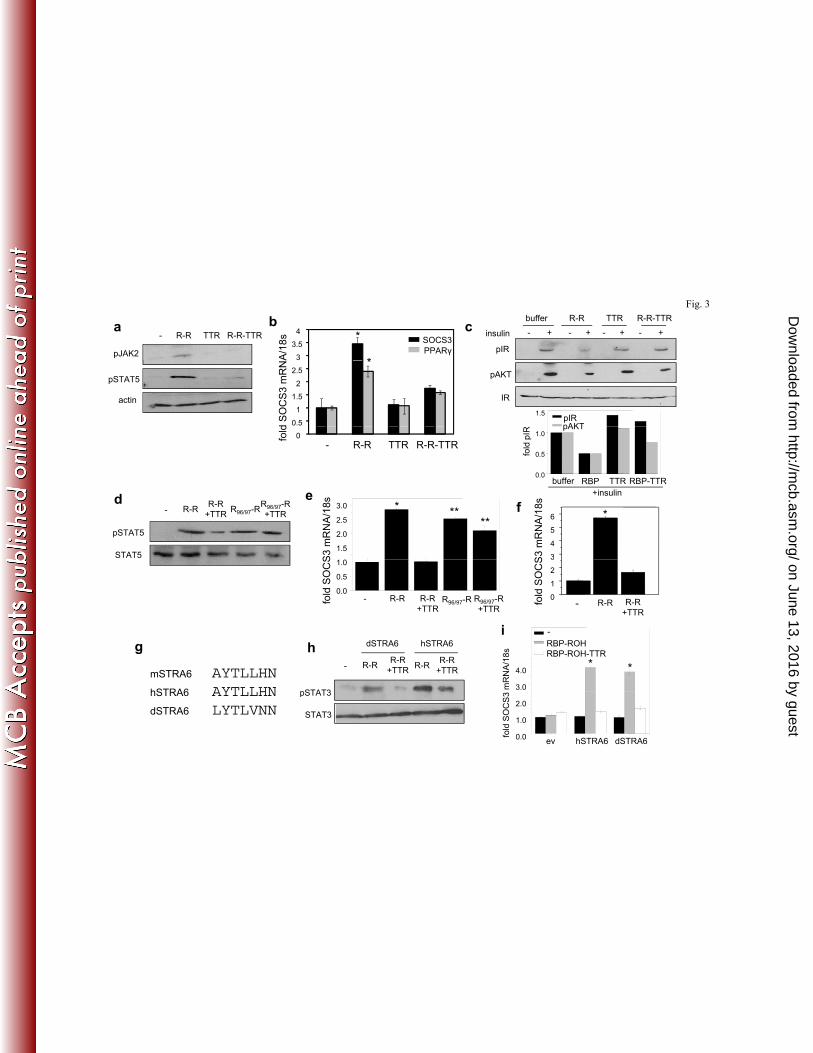
Fig. 3
pJAK2
- R-R TTR R-R-TTRa
3
3.5
4
A/18
s SOCS3PPARγ
b*
*
c - + - + - + - +
pIR
insulin
buffer R-R TTR R-R-TTR
pSTAT5
actin
0.5
1
1.5
2
2.5
d S
OC
S3
mR
NA *
pAKT
IR
1.5
R
pIRpAKT
0- R-R TTR R-R-TTR
fold
eRBP TTR RBP-TTR
0.0
0.5
1.0
fold
pIR
pAKT
+insulinbuffer
R-R R-R+TTR R96/97-R
R96/97-R+TTR-
pSTAT5
STAT5
d
1.5
2.0
2.5
3.0
S3 m
RN
A/1
8s
e* **
**
+insulin
3
4
5
6
S3 m
RN
A/1
8sf *
R96/97-R+TTR
- R-R R-R+TTR
0.0
0.5
1.0
fold
SO
CS
R96/97-R
i
- R-R R-R+TTR
0
1
2
3
fold
SO
CS
pSTAT3
R-R R-R+TTR- R-R R-R
+TTR
dSTRA6 hSTRA6h
3.0
4.0
mR
NA
/18s
-RBP-ROHRBP-ROH-TTR
i
* *mSTRA6 AYTLLHNhSTRA6 AYTLLHN
g
pSTAT3
STAT3
ev hSTRA6 dSTRA60.0
1.0
2.0
fold
SO
CS
3 hSTRA6 AYTLLHNdSTRA6 LYTLVNN
on June 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from
a
Fig. 4
R-R+TTRR-Rbuffer b R-R
+TTRTTRbuffer
b ff R R TTRR-RR R
c eR R
d
RBPhis-RBP his-TTR
TTR
pIR
buffer R-R TTR +TTRR-R+TTRTTRR-Rbuffer
pIR
pAKT
R-R+TTRTTRR-Rbuffer
pIR
pAKT pAKT
IR
pSTAT5
IR
pSTAT5
IR
pSTAT5
f g
6
9
A/18
s
SOCS3PPARγ
4
6
A/18
s SOCS3PPARγ 1.6
A/18
s
SOCS3PPARγ
h*
*
buffer R-R TTR R-R-TTR0
3
6
fold
mR
NA
buffer R-R TTR R-R-TTR0
2
fold
mR
NA
buffer R-R TTR R-R-TTR0.0
0.8
fold
mR
NA
**
buffer R R TTR R R TTR
on June 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from
chow 3 wk 6 wk 10 wkc25
30
A/18
s
lean
a1.5
/18s
leanobese
b
Fig. 5
*HFHS diet
RBP
TTR
0
5
10
15
20
fold
RB
Pm
RN
A obese
0 0
0.5
1.0
fold
TTR
mR
NA/
WAT liver0
WAT liver0.0
buffer R-RR-R
+TTR
IR
e f
pIR
buffer R-RR-R
+TTRbuffer R-R
R-R+TTRd
pIR
pSTAT5
IR
pIR
pSTAT5
IR
his-RBPRBP
his-TTRTTR
450500
g/dl
)
j
8
8s SOCS3PPAR
5
8s
SOCS3PPARγ 1.2
1.4
8s
SOCS3PPARγ
g h i
** **
150200250300350400450
um g
luco
se (m
bufferRBPRBP-TTR0
2
4
6
fold
mR
NA
/18 PPARγ
0
1
2
3
4
fold
mR
NA
/18 PPARγ
0 00.20.40.60.81.0
fold
mR
NA
/18 γ
****
0 20 40 60 80 100 120100150
seru
time (min.)
RBP TTRbuffer R-R R-R-TTR
0buffer R-R R-RTTR
0buffer R-R R-R-TTR
0.0
on June 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from
Related Documents











