Blood, Vol 71, No 6 (June), 1988: pp 151 1-1517 1511 B L0 OD The American The J Society ournal of of Hematology VOL 71, NO 6 JUNE 1988 Transplantation of Human Hairy Cell Leukemia in Radiation-Preconditioned Nude Mice: Characterization of the Model by Histological, Histochemical, Phenotypic, and Tumor Kinetic Studies By Guy B. Faguet and Julia F. Agee Two cell lines (EH and HK) with hairy cell leukemia (HCL) immunophenotypes were recently derived from two HCL patients. Both cell lines were transplanted subcutaneously (2 x 10 or 2 x 10/mouse) in male BALB/c nu/nu mice (n = 1 28) with a 97% success rate when coimplanted with nonproliferative HT-1080 fibrosarcoma cells (2 x 101 mouse) in recipients preconditioned with total-body irra- diation (200 R weekly for 3 weeks). Tumors appeared five to ten days postimplant and reached up to 25% of body weight after a mean survival of 8 weeks (range, 30 to 90 days). Tumor histology suggested large cell lymphoma. Cytochemically and immunophenotypically. tumor cells were indistinguishable from their parent cells. Species and A NUMBER of human solid neoplasms are readily transplantable in athymic mice.’6 These xenotrans- plantation models provide in vivo environments for the initial assessment of novel approaches to chemotherapy, radiother- apy, or immunotherapy72 that are not ethically permissible in the human. These models also enable the study of metas- tases by using probes such as radiolabeled monoclonal anti- bodies (MoAbs)’3’5; however, the observation that xeno- grafted tumors are often less metastati&6”7 than the parent human neoplasms and that leukemia transplantation requires further immunosuppression ofthe host’822 limits the usefulness of xenotransplantation models. The increased natural killer (NK) cell activity23’24 and antitumor antibodies naturally occurring in nude mice25 have been postulated to play a role in the rejection of leukemia implants. An addi- tional factor might be the tendency of leukemia cells to infiltrate local tissues rather than to form nodules that further limit implantation. Such behavior might relate to a lack of growth-, angiogenesis-, or other implantation- promoting factors)7’26 In this report we present our experience with transplanta- tion of human hairy cell leukemia (HCL) cell lines developed in our laboratory27 in radiation-preconditioned, congenitally athymic mice. The transplantation yield was nearly 100%. Tumors grew locally and metastasized extensively to lym- phoid and extralymphoid tissues. This xenotransplantation model might be valuable for assessing the potential useful- ness of new approaches for the treatment of HCL. lineage derivation of tumor cells was confirmed by antibody probes against the mouse histocompatibility antigen H-2, human T and B lymphocyte antigens, and the HCL- associated common chronic lymphocytic leukemia antigen (cCLLa). In order of decreasing frequency, metastases occurred in the spleen, lungs. pleura. lymph nodes. bone marrow. and kidneys. Up to 12% of circulating lymphoid cells in mice were cCLLa-positive. which suggested hema- togenous tumor dissemination. This HCL xenotransplanta- tion model might be useful in preclinical studies for exploring novel experimental therapies for the manage- ment of human HCL. S 1988 by Grune & Stratton, Inc. Animals MATERIALS AND METHODS Athymic nude mice, BALB/c (nu/nu) male, 4 to 6 weeks of age, and weighing 20 to 35 g, were supplied by the Office of Mammalian Genetics and Animal Production, National Cancer Institute, Fred- crick Cancer Research Center, Frederick, MD. The mice were kept under sterile conditions in cages with fiberglass hoods in a room equipped with a negative air flow system and were fed a sterilized mouse diet and sterile water. All procedures were performed under strict sterile conditions. Cell Lines Molt-4,28 a human T cell line, and HT-1080,29 a fibrosarcoma cell line, were supplied by the American Type Culture Collection, Rockville, MD, and grown in RPMI 1640 medium (GIBCO, Grand From the Medical Research Services, Veterans Administration Medical Center, Augusta, GA. and Departments of Medicine and Cell and Molecular Biology, Medical College of Georgia. Submitted June 26, 1987; accepted January 13, 1988. Address reprint requests to Guy B. Faguet. MD, Veterans Administration Medical Center (I I IN). Augusta, GA 30912. The publication costs ofthis article were defrayed in part by page charge payment. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. section 1 734 solely to indicate this fact. © I 988 by Grune & Stratton, Inc. 0006-4971/88/7106-0026$3.00/0

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Blood, Vol 71, No 6 (June), 1988: pp 151 1-1517 1511
B L 0 OD The American
The JSociety
ournal of
of Hematology
VOL 71, NO 6 JUNE 1988
Transplantation of Human Hairy Cell Leukemia in Radiation-PreconditionedNude Mice: Characterization of the Model by Histological, Histochemical,
Phenotypic, and Tumor Kinetic Studies
By Guy B. Faguet and Julia F. Agee
Two cell lines (EH and HK) with hairy cell leukemia (HCL)
immunophenotypes were recently derived from two HCL
patients. Both cell lines were transplanted subcutaneously
(2 x 10 or 2 x 10/mouse) in male BALB/c nu/nu mice
(n = 1 28) with a 97% success rate when coimplanted with
nonproliferative HT-1080 fibrosarcoma cells (2 x 101
mouse) in recipients preconditioned with total-body irra-
diation (200 R weekly for 3 weeks). Tumors appeared five
to ten days postimplant and reached up to 25% of body
weight after a mean survival of 8 weeks (range, 30 to 90
days). Tumor histology suggested large cell lymphoma.
Cytochemically and immunophenotypically. tumor cells
were indistinguishable from their parent cells. Species and
A NUMBER of human solid neoplasms are readilytransplantable in athymic mice.’6 These xenotrans-
plantation models provide in vivo environments for the initial
assessment of novel approaches to chemotherapy, radiother-
apy, or immunotherapy7�2 that are not ethically permissible
in the human. These models also enable the study of metas-
tases by using probes such as radiolabeled monoclonal anti-
bodies (MoAbs)’3’5; however, the observation that xeno-
grafted tumors are often less metastati&6”7 than the parent
human neoplasms and that leukemia transplantation
requires further immunosuppression ofthe host’822 limits the
usefulness of xenotransplantation models. The increased
natural killer (NK) cell activity23’24 and antitumor antibodies
naturally occurring in nude mice25 have been postulated to
play a role in the rejection of leukemia implants. An addi-
tional factor might be the tendency of leukemia cells to
infiltrate local tissues rather than to form nodules that
further limit implantation. Such behavior might relate to a
lack of growth-, angiogenesis-, or other implantation-
promoting factors)7’26
In this report we present our experience with transplanta-
tion of human hairy cell leukemia (HCL) cell lines developedin our laboratory27 in radiation-preconditioned, congenitally
athymic mice. The transplantation yield was nearly 100%.
Tumors grew locally and metastasized extensively to lym-
phoid and extralymphoid tissues. This xenotransplantation
model might be valuable for assessing the potential useful-
ness of new approaches for the treatment of HCL.
lineage derivation of tumor cells was confirmed by antibody
probes against the mouse histocompatibility antigen H-2,
human T and B lymphocyte antigens, and the HCL-
associated common chronic lymphocytic leukemia antigen
(cCLLa). In order of decreasing frequency, metastases
occurred in the spleen, lungs. pleura. lymph nodes. bone
marrow. and kidneys. Up to 12% of circulating lymphoid
cells in mice were cCLLa-positive. which suggested hema-
togenous tumor dissemination. This HCL xenotransplanta-
tion model might be useful in preclinical studies for
exploring novel experimental therapies for the manage-
ment of human HCL.
S 1988 by Grune & Stratton, Inc.
Animals
MATERIALS AND METHODS
Athymic nude mice, BALB/c (nu/nu) male, 4 to 6 weeks of age,
and weighing 20 to 35 g, were supplied by the Office of MammalianGenetics and Animal Production, National Cancer Institute, Fred-crick Cancer Research Center, Frederick, MD. The mice were kept
under sterile conditions in cages with fiberglass hoods in a roomequipped with a negative air flow system and were fed a sterilizedmouse diet and sterile water. All procedures were performed under
strict sterile conditions.
Cell Lines
Molt-4,28 a human T cell line, and HT-1080,29 a fibrosarcoma cellline, were supplied by the American Type Culture Collection,
Rockville, MD, and grown in RPMI 1640 medium (GIBCO, Grand
From the Medical Research Services, Veterans Administration
Medical Center, Augusta, GA. and Departments of Medicine and
Cell and Molecular Biology, Medical College of Georgia.
Submitted June 26, 1987; accepted January 13, 1988.
Address reprint requests to Guy B. Faguet. MD, Veterans
Administration Medical Center (I I IN). Augusta, GA 30912.
The publication costs ofthis article were defrayed in part by page
charge payment. This article must therefore be hereby marked
“advertisement” in accordance with 18 U.S.C. section 1 734 solely to
indicate this fact.
©I 988 by Grune & Stratton, Inc.
0006-4971/88/7106-0026$3.00/0
1512 FAGUET AND AGEE
Island, NY) supplemented with 10% fetal calf serum (FCS) as
suspension and monolayer cultures, respectively. Hairy cell lines EH
and HK, developed in our laboratory27 from peripheral blood fromtwo patients with HCL, were grown as suspension cultures in a
medium (Flow Laboratories, Inglewood, CA) supplemented with15% FCS. All cultures were propagated in 75-cm2 tissue culture
flasks (Corning Glass Works, Corning, NY) in a 5% CO2, 95%
humidity incubator (Forma Scientific, Marietta, OH) with mediumreplacement twice weekly. Seeds were stored in liquid nitrogen.
Immunofluorescence Assays
Surface immunoglobulins (SIgs) were assessed by a direct immu-nofluorescence assay using fluorescein isothiocyanate(FITC)-con-
jugated antisera specific for each heavy and light chain. Othersurface determinants were assessed by indirect immunofluorescence(IFA)�#{176}32 by using FI�TC-conjugated goat antimouse Igs (Cappel
Laboratories, Cochranville, PA) and a panel of mouse antihumanMoAbs: OKT series (Ortho Diagnostics, Raritan, NJ); LEU series
(Becton Dickinson, Mountain View, CA); My4, Bi, MO2, and iS
(Coulter Immunology, Hialeah, FL); aHC1 and aHC2 (a gift fromDr D.N. Posnett, The Rockefeller University, New York); and
CLL- 1 and CLL-2,3’ antibodies specific against the common chronic
lymphocytic leukemia antigen (cCLLa), an antigen common to
HCL and related disorders.3’ Antimouse MoAbs Thyl .2 and H-
2Dk/H-2Kk/H-2Dd (Organon Teknika, Irving, TX) were also
used. Antibodies were used in concentrations sufficient to achieve
saturation binding. Cell fluorescence was determined by fluores-cence microscopy on a Leitz epifluorescence microscope (E. Leitz,Inc, Rockleigh, NY). Nonimmune mouse and rabbit antisera and
irrelevant antibodies and target cells were included as negative
controls.
Radiation
After a 2-week observation period to ensure health and acclimati-zation, mice to be irradiated were anesthetized lightly with anintraperitoneal (IP) injection of ketamine (35 �ag/g body weight)placed in 50-mL uncapped sterile plastic tubes covered with sterile
gauze for ventilation before they received three weekly doses of 200R total-body irradiation (TBI). TBI was administered by a thera-
peutic x-ray unit (courtesy of S. Reichard, Medical College of
Georgia, Augusta, GA) operated at 200 kV and 15 mA with a filter
of 0.5 mm Cu and 1 mm Al at dose rates of 40 to 100 R/min. The
half-value was equivalent to 0.9 mm Cu. The dose delivered to thecenter of the body area in roentgens per minute in air was measured
by a Victoreen dosimeter.
Transplantation
A series of pilot studies was first undertaken to assess the
feasibility of transplanting EH and HK cell lines. The transplanta-
tion protocol followed published guidelines including the use of
preirradiated nude mic&82’ and transplantation-promoting irra-diated HT-1080 fibrosarcoma cell lines as coimplants.18 This proto-
col confirmed the successful transplantation of leukemia-derivedMolt-4 cell lines when coimplanted with HT-1080 cells)8 Thus,Molt-4 cells coimplanted with HT-1080 cells were used as trans-plantable controls. Variables studied included cell line origin, route
of administration, size of inoculum, and the need for irradiationpreconditioning of recipient mice and for coimplanting transplanta-tion-promoting HT-l080 cells. Three days postirradiation or at
equivalent times in unirradiated animals, mice were given a singlesubcutaneous (SC) or intraperitoneal (IP) injection of 2 x I 0s or 2 x
106 cells from leukemic cell lines EH, 1-1K, or Molt-4 in the log phase
of growth that were admixed or not with 2 x 106 preirradiated
(5,000 rad) HT-1080 cells in 0.2 mL of RPMI 1640. Cell irradiationwas carried out in an x-ray unit operated at 90 kV and 10 mA at
doses of 1,000 R/min. After inoculation, SC tumor growth was
monitored from three-dimensional diameter measurements obtainedtwice weekly with vernier calipers. SC tumor volume was calculatedaccording to the following formula’: volume = [(4 x 3.14/3) x(L/2) x (W/2) x (D)]/2. IP tumor presence and growth were
assessed by abdominal palpation. SC tumor weights were measuredat autopsy. Blood samples were taken periodically from the tail veinand at autopsy from a heart puncture to monitor blood counts andthe circulation of tumor cells.
Tumor Histology and Immunophenotype
At various times after cell inoculation, mice were killed and
thorough autopsies performed. Tumors were dissected, weighed, andsectioned to assess histology. Tissue samples from tumors and allinternal organs were fixed in 4% formaldehyde fixative, embedded in
paraffin, stained with hematoxylin and eosin, and examined by lightmicroscopy. Cell suspensions were prepared from teased fresh tumorspecimens for assessing cytochemistry and immunophenotype.
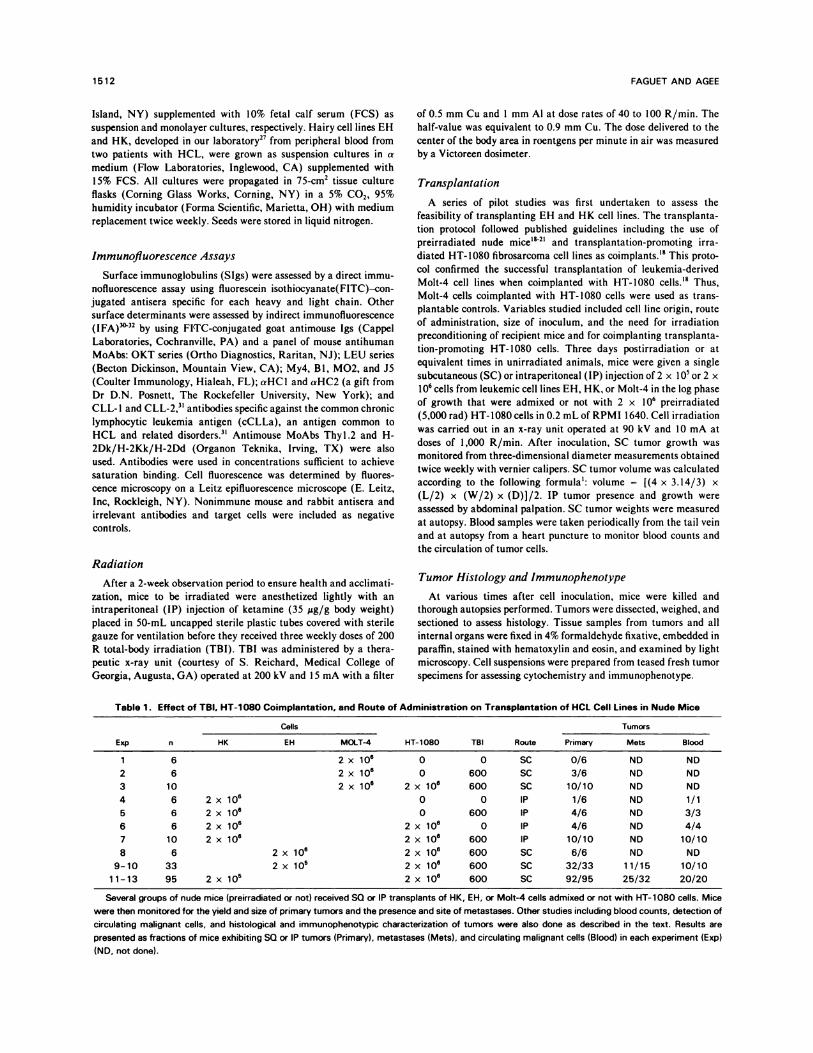
Table 1 . Effect o f TBI. HT-1 08 0 Coimplantati on. and Route of Administrat ion on Tra nsplantatio n of HCL ccl I Lines in Nu de Mice
Exp n
cells
HT-1080 TBI Route
Tumors
HK EH MOIT-4 Primary Mets BlOOd
1 6 2 x 10#{176} 0 0 SC 0/6 ND ND
2 6 2 x 1O� 0 600 SC 3/6 ND ND
3 10 2 x 10#{176} 2 x 1O� 600 SC 10/10 ND ND
4 6 2 x 106 0 0 IP 1/6 ND 1/1
5 6 2 x 106 0 600 P 4/6 ND 3/3
6 6 2 x 106 2 x 106 0 P 4/6 ND 4/4
7 10 2 x 106 2 x 106 600 IP 10/10 ND 10/10
8 6 2 x 10� 2 x 106 600 SC 6/6 ND ND
9-10 33 2 x i0� 2 x 10� 600 SC 32/33 11/15 10/10
11-13 95 2 x i0� 2 x 10#{176} 600 SC 92/95 25/32 20/20
Several groups of nude mice (preirradiated or not) received SQ or IP transplants of HK, EH, or Molt-4 cells admixed or not with HT-1080 cells. Mice
were then monitored for the yield and size of primary tumors and the presence and site of metastases. Other studies including blood counts, detection of
circulating malignant cells, and histological and immunophenotypic characterization of tumors were also done as described in the text. Results are
presented as fractions of mice exhibiting SQ or IP tumors (Primary), metastases (Mets), and circulating malignant cells (Blood) in each experiment (Exp)
(ND. not done).
RESULTS
Transplantability ofHCL Cell Lines
Preliminary experiments showed that SC or IP implanta-
tion of 2 x 106 Molt-4 (n = 12) or HK cells (n = 1 2) alone in
irradiated (n = 6) and nonirradiated mice (n = 6) resulted in
erratic or no tumor development, respectively (Table 1).
However, SC and IP transplantability of either cell line inirradiated mice was enhanced by coimplanting irradiation-
treated HT-1080 cells’8: HK cells were inoculated in miceexhibiting pristane-induced ascites to mimic the in vivogrowth environment optimal for hybridomas. This approachyielded abdominal tumors with diffuse peritoneal and mesen-
teric seeding in 68% of the animals (n = 28). In these mice,
up to 75% (mean, 36.3%; SEM, 8.4%) ofthe free ascitic fluid
cells expressed the characteristic hairy cell immunopheno-type (Table 2); likewise, 97% of 128 preirradiated mice
inoculated SC with either EH (n = 33) or HK (n = 95) cells
and 100% of ten control mice receiving Molt-4 cells devel-
oped SC tumors when coimplanted with irradiated HT-1080
cells. When this transplantation protocol was used, the two
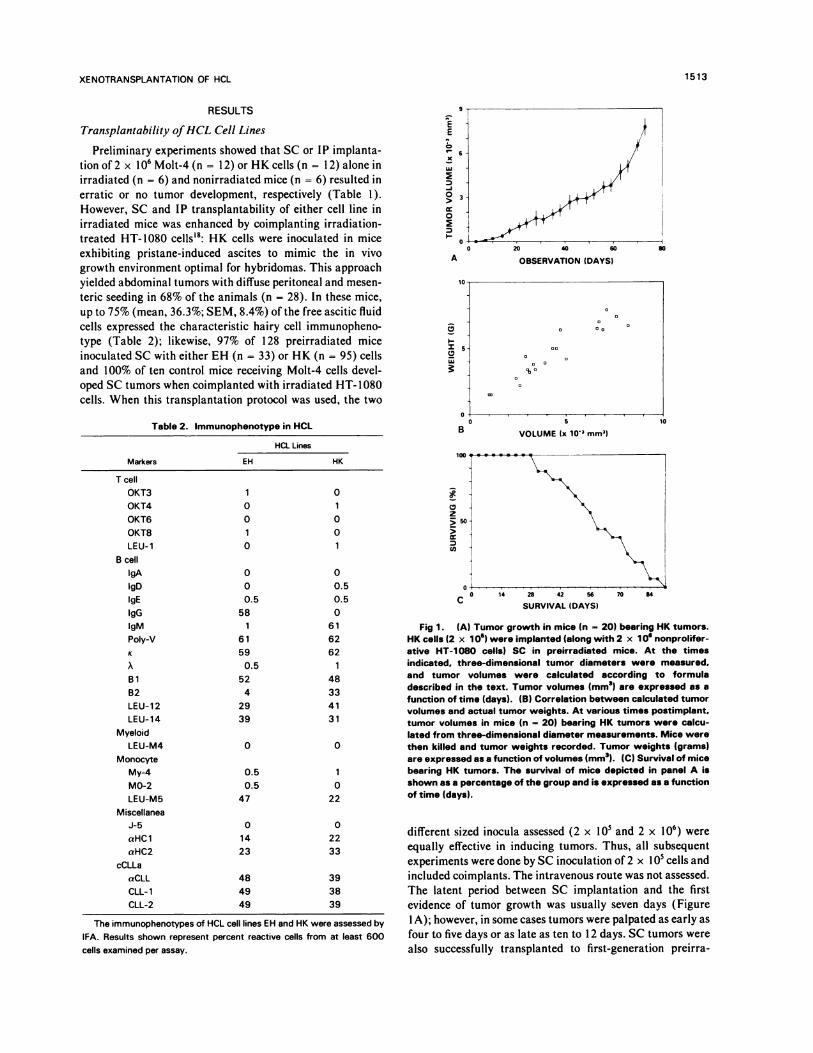
EE
K
Lu
-a0>0
I-A
I-IC,Lu
OBSERVATION (DAYS)
10
5 0�
0 5
B VOLUME lx 10” mm’)no
14
Table 2. Immunophenotype in HCL
Markers
HCL Lines
EH HK
Tce)l
OKT3 1 0
OKT4 0 1
OKT6 0 0
OKT8 1 0
LEU-1 0 1
B cell
IgA 0 0
lgD 0 0.5
gE 0.5 0.5
IgG 58 0
gM 1 61
Poly-V 6 1 62
K 59 62
A 0.5 1
Bi 52 48
B2 4 33
LEU-12 29 41
LEU-14 39 31
Myeloid
LEU-M4 0 0
Monocyte
My-4 0.5 1
M0-2 0.5 0
LEU-M5 47 22
Miscellanea
J-5 0 0
aHC1 14 22
aHC2 23 33
cCLLa
aCLL 48 39
CLL-1 49 38
CLL-2 49 39
28 42 56C�
SURVIVAL (DAYS)
70 84
The immunophenotypes of HCL cell lines EH and HK were assessed by
IFA. Results shown represent percent reactive cells from at least 600
cells examined per assay.
different sized inocula assessed (2 x l0� and 2 x 106) were
equally effective in inducing tumors. Thus, all subsequent
experiments were done by SC inoculation of 2 x 10� cells andincluded coimplants. The intravenous route was not assessed.The latent period between SC implantation and the firstevidence of tumor growth was usually seven days (Figure
1A); however, in some cases tumors were palpated as early as
four to five days or as late as ten to 1 2 days. SC tumors werealso successfully transplanted to first-generation preirra-
XENOTRANSPLANTATION OF HCL 1513
Fig 1 . (A) Tumor growth in mice (n 20) bearing HK tumors.HK cells (2 x 1 0) were implanted (along with 2 x 1 0’ nonprolifer-ative HT-1 080 cells) SC in preirradiated mice. At the timesindicated. three-dimensional tumor diameters were measured.and tumor volumes were calculated according to formuladescribed in the text. Tumor volumes (mm3) are expressed as afunction of time (days). (B) Correlation between calculated tumorvolumes and actual tumor weights. At various times postimplant.tumor volumes in mice (n - 20) bearing HK tumors were calcu-lated from three-dimensional diameter measurements. Mice werethen killed and tumor weights recorded. Tumor weights (grams)are expressed as a function of volumes (mm3). (C) Survival of micebearing HK tumors. The survival of mice depicted in panel A isshown as a percentage of the group and is expressed as a functionof time (days).
1514 FAGUET AND AGEE
diated mice (n = 6) without co-implants.No attempts were
made to assess the serial transplantation potential of SC
tumors or the minimum number of cells necessary for
implantation, and no correlation trends were seen between
inoculum size and tumor growth rates, metastases, or host
survival. No appreciable differences were observed between
EH and HK cell lines in terms of the implantation or growthrates of primary tumors or their ultimate size, development,
distribution or size of metastases, tumor histology, or host
survival.
Characterization of Tumors
Tumor growth. Tumor growth was initially slow butproceeded uninterruptedly until death. In one representative
experiment involving 20 mice bearing SC HK tumors, the
following observations were made: (a) there was an excellent
correlation (r = .96) between calculated tumor volume and
actual tumor weight measured at autopsy (Fig 1B). The
volume-weight ratio was I mm3/ I mg; (b) tumors grew to an
average calculated weight of approximately 2 and 4 g at 40
and 60 days postimplant, respectively (Fig 1A), and reached
an average weight of 5.09 g (SEM, I .7) after a mean survivalof 56 (SEM, 18) days; (c) attrition, appeared to nonran-
domly affect mice with larger tumors first, began in thesecond month postimplant and continued at a constant rateof approximately 12% per week (Fig I C); (d) the total body
weight at the end of each experiment showed only a slight
gain, which suggested that normal tissue weight loss approxi-
mately equivalent to that of the growing tumors had taken
place. Thus, after 30 to 45 days posttransplant, most animals
were cachectic, having lost 1 0% to I 5% of their initial body
weight in spite of adequate dietary intake.
Tumor dissemination. Primary tumors disseminated
extensively to lymphoid and nonlymphoid organs. Local
dissemination occurred by contiguous invasion of SC tissue
including fat and muscle. Diffuse infiltration of spleens with
splenomegaly was universal. Some but not all animals exhib-
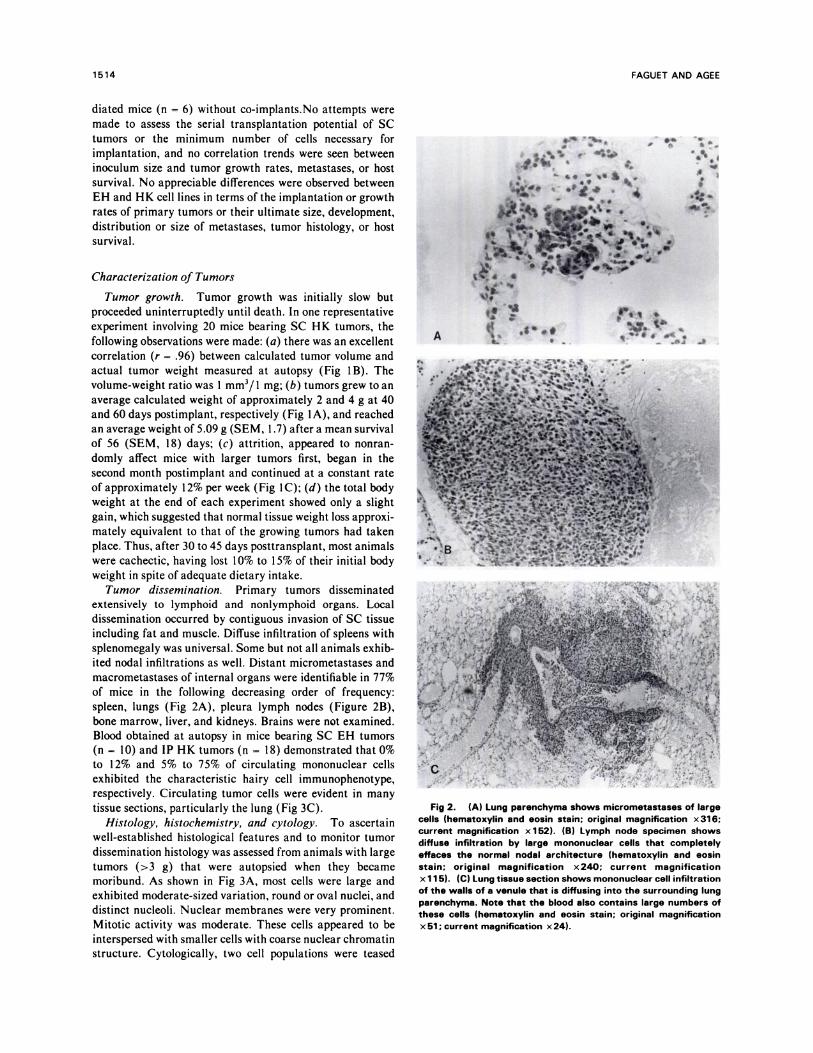
ited nodal infiltrations as well. Distant micrometastases andmacrometastases of internal organs were identifiable in 77%
of mice in the following decreasing order of frequency:
spleen, lungs (Fig 2A), pleura lymph nodes (Figure 2B),
bone marrow, liver, and kidneys. Brains were not examined.
Blood obtained at autopsy in mice bearing SC EH tumors
(n = 10) and IP HK tumors (n = 18) demonstrated that 0%
to 12% and 5% to 75% of circulating mononuclear cells
exhibited the characteristic hairy cell immunophenotype,
respectively. Circulating tumor cells were evident in many
tissue sections, particularly the lung (Fig 3C).
Histology, histochemistry, and cytology. To ascertain
well-established histological features and to monitor tumor
dissemination histology was assessed from animals with large
tumors (>3 g) that were autopsied when they became
moribund. As shown in Fig 3A, most cells were large and
exhibited moderate-sized variation, round or oval nuclei, and
distinct nucleoli. Nuclear membranes were very prominent.
Mitotic activity was moderate. These cells appeared to be
interspersed with smaller cells with coarse nuclear chromatin
structure. Cytologically, two cell populations were teased
Fig 2. (A) Lung parenchyma shows micrometastases of largecells (hematoxylin and eosin stain; original magnification x 316;current magnification x 1 52). (B) Lymph node specimen showsdiffuse infiltration by large mononuclear cells that completelyeffaces the normal nodal architecture (hematoxylin and eosinstain; original magnification x240; current magnificationx 1 1 5). (C) Lung tissue section shows mononuclear cell infiltration
of the walls of a venule that is diffusing into the surrounding lungparenchyma. Note that the blood also contains large numbers ofthese cells (hematoxylin and eosin stain; original magnificationx 51 ; current magnification x 24).

XENOTRANSPLANTATION OF HCL 1515
Table 3. Lineage and Species Derivation of EH
and Molt-4 Tumors
Tumors
Antibodies EH Molt-4
Antihuman
T cells
OKT6 ND 46
LEU-1 1.5 45.5
B cells:
Bi 43.5 ND
Poly-V 40.5 0.5
HCL cells
CLL-1 47 ND
Antimouse:
T cells:
Thyl.2 16 20
H2
H-2Dk/H-2Kk/H-2Dd 38.5 33
Preirradiated (200 R/wk for 3 weeks) mice were implanted with EH
and MoIt-4 cells. When the tumors reached >2 g, cells were teased and
assayed by IFA with the antihuman antibodies OKT6, LEU-1, Bi,
anti-Igs. and CLL-1; and antimouse T cell (Thyl.2) and antimouse H2
complex anti-H-2Dk/H-2Kk/H-2Dd antibodies. Reactive cells are
expressed as a percentage of the 600 cells examined per assay.#{149}Poly..Vrefers to polyvalent antihuman Ig antibody.
H-2Dk/H-2Kk/H-2Dd-positive small cells (33%), more
than half of which also reacted with the MoAb Thyl .2.
Blood Counts
Fig 3. (A) Histological section of an SC HK tumor shows afairly homogenous population of large lymphoid cells with thicknuclear membranes and prominent nucleoli (hematoxylin andeosin stain; original magnification x 31 6; current magnificationx 1 52). (B) A cytocentrifuge spread of cells teased from the same
tumor shows moderate anisocytosis with a preponderance of
large. nucleolated lymphoid cells (May-Gr#{252}nwald-Giemsa stain;
original magnification x 400; current magnification x 192).
from primary tumors (Fig 3B): one, made oflarge immature
lymphoid cells; the other, reminiscent of small, mature
lymphocytes. Most but not all large cells exhibited tartrate
resistant acid phosphatase (TRAP) reactivity. Coimplanted
HT-1080 cells, recognizable histologically by their eosino-
philic cytoplasm and association with fibrous trabeculae,’8were absent in tumors. These nonproliferative cells appear
not to survive past 2 or 3 weeks postimplant.
Tumor immunophenotype. The species and lineage den-vation of cells teased from EH tumors was assessed by IFA
on two mice bearing tumors ranging in weight from 2 to 5 g.
Phenotypically, two cell populations were identified (Table
3): one of preponderantly large cells that reacted with
antihuman SIg (41%), B) (43%) and anti-cCLLa (47%)antibodies; the other made largely of small cells reactingwith antimouse MoAbs Thyl.2 (16%) and H-2Dk/H-2Kk/
H-2Dd (38%). Only background reactivity (2%) was
observed with the antihuman T cell antibody LEU-l. Like-
wise, primary tumors of two mice bearing Molt-4 tumors (2
to 4 g in weight) exhibited a preponderant population
(>45%) of OKT6- and LEU-l-positive large cells and
Complete counts from tail vein or heart puncture blood
were obtained in a group of ketamine-anesthetized nude
mice (n = I 2) before and three days after irradiation and 4
to 6 weeks after tumor implantation, respectively. TB!
induced 89% and 75% reductions in average circulating
lymphocyte (679/�zL v 4,096/�L) and platelet ( I 3 1 ,000/�L
V 478,000/j�L) counts, respectively (Table 4). Averages for
hemoglobin levels (15.6 v 14.0 g/zL) and granulocyte
(2,704/zL v l,777/jsL) and monocyte (135/zL v l05/�L)
counts were unaffected. Blood counts obtained at autopsy
(mean tumor size, 2.8 g) revealed a return to normal platelet
counts (mean, 584,000/�L), with little change in granulo-
cyte counts but the presence of anemia (mean, 1 2.Og/.zL)
and persistent lymphopenia (mean, 523/�L).
DISCUSSION
A preliminary experiment confirmed the widely held viewthat direct transplantation of human hematopoietic and
lymphoid malignant cells in nude mice is unsuccessful.
Naturally occurring cytotoxic antitumor antibodies25 and the
increased NK cell activity observed in nude mice23’24 havebeen implicated in the rejection ofsuch implants. In addition,
the tendency of leukemic cells to infiltrate local tissues rather
than form nodules may also impede implantation. However,xenotransplantation has been reportedly facilitated by host
preconditioning with TB!; coimplantation of nonprolifera-
tive, transplantation-promoting HT-l080 cells; or by both.’8’
21.29 Analysis of our cumulative data supports these findings.
Indeed, the overall frequency of “takes” increased from 8.3%
1516 FAGUET AND AGEE
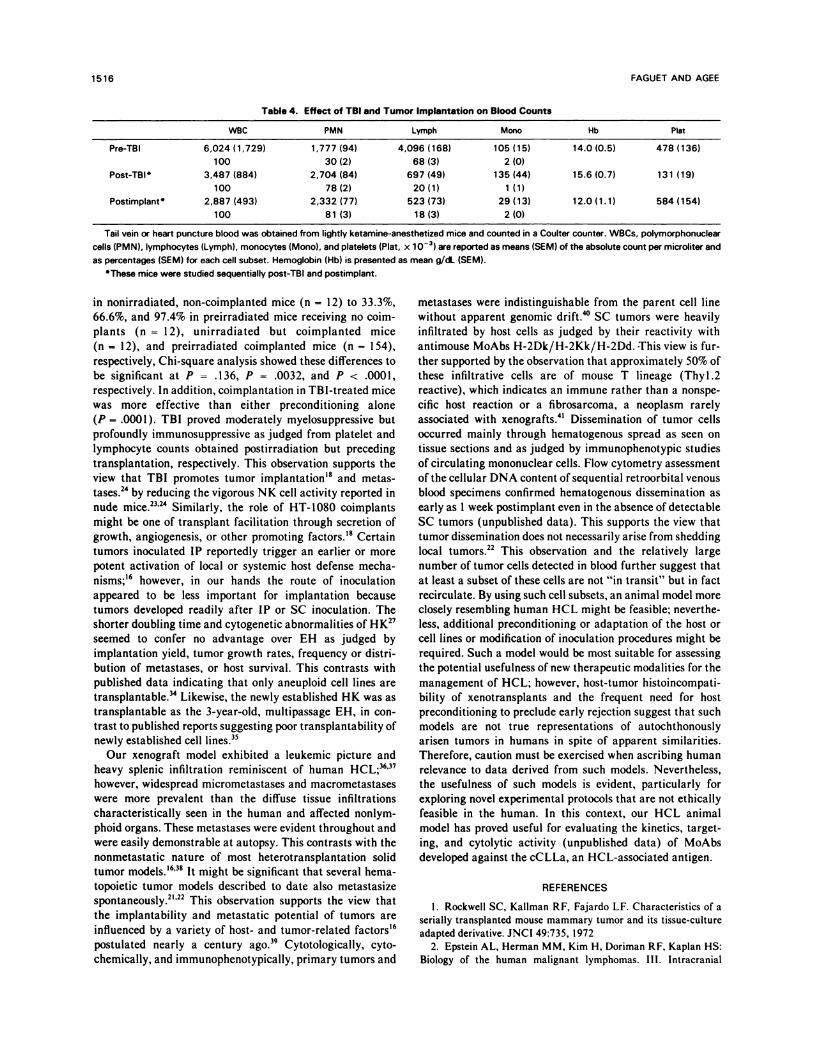
Table 4. Effect of TBI and Tumor Implantation on Blood Counts
WBC PMN Lymph Mono Hb P)at
Pre-TBI 6,024 (1,729)
100
1,777 (94)
30(2)
4,096 (168)
68(3)
105 (15)
2(0)
14.0 (0.5) 478 (136)
Post-TBI 3,487(884)
100
2,704(84)
78(2)
697(49)
20(1)
135(44)
1(1)
15.6(0.7) 131 (19)
Postimplant 2,887 (493)
100
2,332 (77)
81 (3)
523 (73)
18 (3)
29 (13)
2 (0)
12.0 (1.1) 584 (154)
Tail vein or heart puncture blood was obtained from lightly ketamine-anesthetized mice and counted in a Coulter counter. WBCs. polymorphonuclear
cells (PMN). lymphocytes (Lymph), monocytes (Mono), and platelets (Plat, x 10�) are reported as means (SEM) of the absolute count per microliter and
as percentages (SEM) for each cell subset. Hemoglobin (Hb) is presented as mean g/dL (SEM).
‘These mice were studied sequentially post-TBI and postimplant.
in nonirradiated, non-coimplanted mice (n = 12) to 33.3%,
66.6%, and 97.4% in preirradiated mice receiving no coim-
plants (n = 12), unirradiated but coimplanted mice
(n = 12), and preirradiated coimplanted mice (n = 154),respectively, Chi-square analysis showed these differences to
be significant at P = .136, P = .0032, and P < .0001,
respectively. In addition, coimplantation in TBI-treated mice
was more effective than either preconditioning alone
(P = .0001). TB! proved moderately myelosuppnessive butprofoundly immunosuppressive as judged from platelet and
lymphocyte counts obtained postirradiation but preceding
transplantation, respectively. This observation supports the
view that TB! promotes tumor implantation’#{176} and metas-
tases.24 by reducing the vigorous NK cell activity reported in
nude mice.23’24 Similarly, the role of HT-1080 coimplantsmight be one of transplant facilitation through secretion of
growth, angiogenesis, or other promoting � Certaintumors inoculated IP reportedly trigger an earlier or more
potent activation of local or systemic host defense mecha-
nisms;’6 however, in our hands the route of inoculation
appeared to be less important for implantation because
tumors developed readily after IP or SC inoculation. Theshorter doubling time and cytogenetic abnormalities of HK27
seemed to confer no advantage over EH as judged by
implantation yield, tumor growth rates, frequency or distni-
bution of metastases, or host survival. This contrasts with
published data indicating that only aneuploid cell lines are
transplantable.TM Likewise, the newly established HK was astransplantable as the 3-year-old, multipassage EH, in con-
trast to published reports suggesting poor transplantability of
newly established cell lines.35
Our xenograft model exhibited a leukemic picture andheavy splenic infiltration reminiscent of human HCL;36’37
however, widespread micrometastases and macrometastaseswere more prevalent than the diffuse tissue infiltrationscharacteristically seen in the human and affected nonlym-
phoid organs. These metastases were evident throughout and
were easily demonstrable at autopsy. This contrasts with the
nonmetastatic nature of most heterotransplantation solidtumor models.’6’38 It might be significant that several hema-topoietic tumor models described to date also metastasize
spontaneously.21’22 This observation supports the view thatthe implantability and metastatic potential of tumors areinfluenced by a variety of host- and tumor-related factors’6
postulated nearly a century ago.3’ Cytotologically, cyto-
chemically, and immunophenotypically, primary tumors and
metastases were indistinguishable from the parent cell line
without apparent genomic dnift.�#{176}SC tumors were heavily
infiltrated by host cells as judged by their reactivity with
antimouse MoAbs H-2Dk/H-2Kk/H-2Dd. ‘This view is fur-
then supported by the observation that approximately 50% of
these infiltrative cells are of mouse T lineage (Thyl .2
reactive), which indicates an immune rather than a nonspe-
cific host reaction or a fibrosarcoma, a neoplasm rarely
associated with xenografts.4’ Dissemination of tumor cells
occurred mainly through hematogenous spread as seen on
tissue sections and as judged by immunophenotypic studies
of circulating mononuclear cells. Flow cytometry assessmentof the cellular DNA content of sequential retroorbital venousblood specimens confirmed hematogenous dissemination as
early as 1 week postimplant even in the absence of detectable
SC tumors (unpublished data). This supports the view thattumor dissemination does not necessarily arise from shedding
local tumors.22 This observation and the relatively large
number of tumor cells detected in blood further suggest that
at least a subset of these cells are not “in transit” but in fact
recirculate. By using such cell subsets, an animal model moreclosely resembling human HCL might be feasible; neverthe-
less, additional preconditioning or adaptation of the host or
cell lines or modification of inoculation procedures might be
required. Such a model would be most suitable for assessing
the potential usefulness of new therapeutic modalities for the
management of HCL; however, host-tumor histoincompati-
bility of xenotransplants and the frequent need for host
preconditioning to preclude early rejection suggest that such
models are not true representations of autochthonously
arisen tumors in humans in spite of apparent similarities.
Therefore, caution must be exercised when ascribing humanrelevance to data derived from such models. Nevertheless,
the usefulness of such models is evident, particularly forexploring novel experimental protocols that are not ethically
feasible in the human. In this context, our HCL animal
model has proved useful for evaluating the kinetics, target-
ing, and cytolytic activity (unpublished data) of MoAbs
developed against the cCLLa, an HCL-associated antigen.
REFERENCES
I . Rockwell SC, Kallman RF, Fajardo LF. Characteristics of aserially transplanted mouse mammary tumor and its tissue-culture
adapted derivative. JNCI 49:735, 19722. Epstein AL, Herman MM, Kim H, Doriman RF, Kaplan HS:
Biology of the human malignant lymphomas. III. Intracranial

XENOTRANSPLANTATION OF HCL 1517
heterotransplantation in the nude, athymic mouse. Cancer 37:2 158,1976
3, Rousseau-Merck M, Bigel P. Mouly H, Flamant F, ZuckeriM, Wache AC, Nezelof C: Transplantability in nude mice of
embryonic and other childhood tumours. Br i Cancer 52:279, 1985
4. Friedlander ML, Russell P. Taylor 1W, Tattersall MH: Ovar-
ian tumour xenografts in the study of the biology of human epithelial
ovarian cancer. Br J Cancer 5 1 :3 1 9, 1985
5. Giovanella BC, Stehlin iS, Williams Li, Lee 55, Shepard RC:Heterotransplantation of human cancers into nude mice. A model
system for human cancer chemotherapy. Cancer 42:2269, 19786. Noguchi DP, Wallace R, iohnson i, Early EM, O’Brien 5,
Ferrone 5, Pellegrino MA, Milstein i, Needy C, Browne W,
Petricciani JC: Characterization of WiDr: A human colon carci-
noma cell line. In Vitro 15:401, 19797. Hill iH, Plant RL, Harria DM, Grossweiner LI, Rok B, Seter
AJ: The nude mouse xenograft system: A model for photodetection
and photodynamic therapy in head and neck squamous cell carci-noma. Am i Otolaryngol 7:17, 1986
8. Lindenberger i, Hermeking H, Kummermehr i, Denekamp i:
Response of human tumour xenografts to fractionated X-irradiation.
Radiother Oncol 6: 1 5, 1986
9. Azar I-IA, Fernandez SB, Bros LM, Specter SC: Human
tumor xenografts in nude mice: Current experience and chemother-apy trials. Pathol Annu 19:245, 1984
10. Jones DH, Goldman A, Gordon I, Pritchard J, Gregory Bi,
Kemshead iT: Therapeutic application of a radiolabelled mono-
clonal antibody in nude mice xenografted with human neuroblasto-
ma: Tumoricidal effects and distribution studies. Int i Cancer35:715, 1985
1 1 . Weil-Hillman G, Uckun FM, Manske JM, Vallera D: Com-
bined immunochemotherapy of human solid tumors in nude mice.
Cancer Res 47:579, 1987I 2. Latif ZA, Lozzio BB, Lozzio CB, Herberman RB, Wust Ci:
Abrogation of the proliferation of human leukemia cells in nude
mice by a xenoantiserum. Leuk Res 3:371, 1979
1 3. Ballou B, Levine G, Hakala TR, Solter D: Tumor location
detection with radioactively labeled monoclonal antibody and exter-nal scintigraphy. Science 206:844, 1979
14. Giovanella BC, Yim SD, Morgan AC, Stehlin iS, Williams
Li ir: Metastases of human melanomas transplanted in “nude”
mice. JNCI 50:1051, 1973
15. Graham SD, Mickey DD, Paulson DF: Detection of meta-
static tumors in nude mice. JNCF 60:7 1 5, 1978
I 6. Fidler Ii: Rationale and methods for the use of nude mice to
study the biology and therapy of human cancer metastasis. Cancer
Metastasis Rev 5:29, 198617. Ueyama Y, Morita K, Kondo Y, Sato N, Asno 5, Ohsawa N,
Sakurai M, Nagumo F, lijima K, Tamaoki N: Direct and serialtransplantation of a Ph’ +ve human myeloblastoid tumor into nude
mice. Br i Cancer 36:523, 1977
1 8. Ziegler HW, Fnizzera G, Bach FH: Successful transplanta-
tion of a human leukemia cell line into nude mice: Conditions
optimizing graft acceptance. JNCI 68:15, 1982
19. Toolan HW: Successful subcutaneous growth and transplan-tation of human tumors in X-irradiated laboratory animals. Proc Soc
Exp Biol Med 77:572, 1951
20. Watanabe 5, Shimosato Y, Juroki M, Sato Y, Nakajima T:Transplantability of human lymphoid cell line, lymphoma and
leukemia in splenectomized and/or irradiated nude mice. Cancer
Res4O:2588, 1980
21 . Watanabe 5, Shimosato Y, Kameya T, Kuroki M, Kitahara
T, Monato K, Shimoyama M: Leukemia distribution of a humanacute lymphocytic leukemia cell line (Ichikawa strain) in nude miceconditioned with whole-body irradiation. Cancer Res 38:3494, 1978
22. Lozzio BB, Machado EA, Lair SV, Lozzio CB: Reproduciblemetastatic growth of K-562 human myelogenous leukemia cells innude mice. JNCI 63:295, 1979
23. Herberman RB: Natural cell-mediated cytotoxicity in nude
mice, in Fogh i, Giovanella BC (eds): The Nude Mouse in Experi-
mental and Clinical Research. San Diego, Academic, 1978, p 13524. Hanna N: Role of natural killer cell in control of cancer
metastasis. Cancer Metastasis Rev 1:45, 1982
25. Martin WH, Martin SE: Naturally occurring cytotoxic anti-tumor antibodies in sera of congenitally athymic (nude) mice.
Nature 249:564, 1974
26. Brem S, Cotran R, Folkman i: Tumor angiogenesis: Aquantitative method for histologic grading. JNC! 48:347, 1972
27. Faguet GB, Satya-Prakash KL, Agee iF. Cytochemical,cytogenetic, immunphenotypic and tumorigenic characterization of
two hairy cell lines. Blood 69:422, 198728. Minowada i, Ohnuma T, Moore GE: Rosette-forming human
lymphoid cell lines. I. Establishment and evidence for origin of
thymus-derived lymphocytes. JNCI 49:891, 197229. Rasheed 5, Nelson-Rees WA, ioth EM, Arnstein P, Gardner
MD: Characterization of a newly derived sarcoma cell line (HT-1080). Cancer 33:1027, 1974
30. Faguet GB: Mechanisms of lymphocyte activation: The roleof suppressor cells in the proliferative responses of chronic lymphatic
leukemia lymphocytes. i Clin Invest 63:67, 19793 1 . Agee iF, Garver F, Faguet GB: An antigen common to
chronic lymphocytic and hairy cell leukemia not shared by normal
lymphocytes or by other leukemic cells. Blood 68:62, 1986
32. Martin Pi, Hansen iA, Nowisski RC, Brown MA: A new
human T-cell differentiated antigen: Unexpected on chronic lym-phocytic leukemia cells. Immunogenetics I 1:429, 1980
33. Faguet GB, Agee iF: Monoclonal antibodies against the
chronic lymphatic leukemia antigen, cCLLa: Characterization and
reactivity. Blood 70:437, 198734. Diehl V, Krause P, Hellniengel KP, Busche M, Schedel I,
Laskewitz E: Lymphoid cell lines: In vitro markers in correlation totumorigenicity in nude mice, in Thierfelder 5, Rodth H, Thiel E
(eds): Hematology and Blood Transfusion, vol 20. Berlin, Springer-Verlag, 1977, p 289
35. Nilsson K, Giovanella BC, Stehlin iS, Klein G: Tumorigenic-
ity of human hematopoietic cell lines in athymic nude mice. Int i
Cancer 19:337, 1974
36. Bouroncle BA: Leukemia reticuloendotheliosis, hairy cellleukemia. Blood 53:412, 1979
37, Golomb HM, Catovsky D, Golde DW: Hairy cell leukemia, a
five-year update on seventy-one patients. Ann Intern Med 99:485,
1983
38. Bather R, Becker BC, Contreras G, Furesz i: Heterotrans-plantation studies with tissue culture cell lines in various animal and
in vitro host systems. i Biol Stand 13:13, 1985
39. Paget 5: The distribution of secondary growths in cancer ofthebreast. Lancet 1:571, 1898
40. Sharkey FE, Spicer iH, Fogh i: Changes in histologicaldifferentiation of human tumors transplanted to athymic nude mice:
A morphometric study. Exp Cell Biol 53:100, 1985
41 . Sparrow 5, Jones M, Billington 5, Stace B: The in vivo
malignant transformation of mouse fibroblasts in the presence ofhuman tumour xenografts. Br i Cancer 53:793, 1986
Related Documents











