This article was published in the above mentioned Springer issue. The material, including all portions thereof, is protected by copyright; all rights are held exclusively by Springer Science + Business Media. The material is for personal use only; commercial use is not permitted. Unauthorized reproduction, transfer and/or use may be a violation of criminal as well as civil law. ISSN 0018-8158, Volume 641, Number 1

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article was published in the above mentioned Springer issue.The material, including all portions thereof, is protected by copyright;all rights are held exclusively by Springer Science + Business Media.
The material is for personal use only;commercial use is not permitted.
Unauthorized reproduction, transfer and/or usemay be a violation of criminal as well as civil law.
ISSN 0018-8158, Volume 641, Number 1

PRIMARY RESEARCH PAPER
Transition from shallow lake to a wetland: a multi-proxycase study in Zalavari Pond, Lake Balaton, Hungary
Janos Korponai • Mihaly Braun • Krisztina Buczko •
Istvan Gyulai • Laszlo Forro • Judit Nedli •
Istvan Papp
Received: 10 July 2009 / Revised: 18 December 2009 / Accepted: 30 December 2009 / Published online: 16 January 2010
� Springer Science+Business Media B.V. 2010
Abstract Lake Balaton, the largest shallow lake in
Central Europe, has no natural outlet, therefore,
underwent water level changes during its 15,000–
17,000 years of history. The lake is very sensitive to
both climate changes and human impacts. Surround-
ings have been inhabited since the Stone Age;
however, heavy human impact can be recognized
during the past 6000 years. In this study, we estab-
lished three different stages for and reconstructed
water level changes of Lake Balaton by geochemical
data, subfossil Cladocera and diatom remains in the
sediments of the Zalavari Pond, a part of the Kis-
Balaton wetland. In 9900–8600 cal. year BP, climate
was dry, water level was low, and there was a
wetland in this area. Although organic matter content
was low in the sediment, the ratio of Fe/Mn was high.
Between 5600 and 5000 cal. year BP, water level
increased, Fe/Mn ratio shows that oxygen conditions
of sediments was improved in agreement with the
relatively low number of diatom remains and dense
chydorid remains. About 5000 cal. year BP, water
level of Lake Balaton decreased as indicated by high
organic content with low carbonate and high Fe/Mn
ratio in the sediments (oxygen depletion). At the
bottom of this section, high Fe and S concentrations
showed accumulation of pyrite (FeS2) that is common
in wetlands with very low redox potential. Low
abundance of Cladocera remains together with rich
and diverse diatom flora confirm the low water level
hypothesis. Our data support that the water level of
Handling editor: J. Padisak
J. Korponai (&)
West-Transdanubian District Water Authority,
Csik F. str. 4, 8360 Keszthely, Hungary
e-mail: [email protected]
J. Korponai
Department of Chemistry and Environmental Sciences,
University of West Hungary, Szombathely, Hungary
M. Braun
Department of Inorganic Chemistry, University of
Debrecen, 4010 Debrecen, Hungary
K. Buczko
Department of Botany, Hungarian Natural History
Museum, 1087 Budapest, Hungary
I. Gyulai
Department of Applied Ecology, University of Debrecen,
4010 Debrecen, Hungary
L. Forro � J. Nedli
Department of Zoology, Hungarian Natural History
Museum, 1088 Budapest, Hungary
I. Papp
Department of Mineralogy and Geology, University of
Debrecen, 4010 Debrecen, Hungary
123
Hydrobiologia (2010) 641:225–244
DOI 10.1007/s10750-009-0087-0
Author's personal copy

Lake Balaton was higher between 8600 and 5000 cal.
year BP than it is at present.
Keywords Lake Balaton � Shallow lake �Cladocera remains � Chydorids � Diatoms �Trace metals
Introduction
Studying the paleoenvironment of small closed basins
has a long tradition all over the World. Lacustrine
sediments of deep lakes can preserve almost undis-
turbed evidences of changes in the past; therefore,
they provide ideal objects for paleolimnological
studies, including taxonomical, evolutional, and pa-
leoenvironmental reconstruction. Datasets delivered
from deep lakes are also used frequently for making
‘‘climate models’’. The so-called shallow lakes (Pad-
isak & Reynolds, 2003) have great economic and
social importance. Their development and historical
ecological status are in the focus of many studies.
Paleolimnological experts fully appreciate the fact
that the sedimentation of the extended, shallow water
bodies is far from ideal; however, their investigations
are urgent tasks of limnology.
Sediments of shallow lakes show greater mixing
than those in deep waters due to wind effect or
activity of benthic organisms (Kearns et al., 1996).
Shallow lakes and wetlands are more sensitive to
climate and human impact (Whitmore et al., 1996;
Kenney et al., 2002; Moss et al., 2003; Valiranta
et al., 2005) since small water level fluctuations in
absolute terms are considerable changes in relative
water depth. It is especially true for such a large and
shallow lake as Lake Balaton is. The estimated age of
the lake is cca. 15,000 years (Cserny & Nagy-Bodor,
2000) and from then, Lake Balaton was almost
endorheic while sandbars interfered with the lake
water to flush toward Danube.
Starting at the end of the nineteenth century, the
lake and its surroundings have intensively been
studied by a great number of experts (see the
comprehensive studies by Virag, 1998, 2005). Pan-
tocsek (in Loczy, 1916), Zolyomi (1952, 1995), and
Sebestyen (1969a, b, 1970, 1971) conducted first
paleolimnological studies in Lake Balaton sediments.
Zolyomi took sediment cores from several places in
lakebed for pollen analysis while Sebestyen analyzed
subfossil Cladocera remains in large intervals and
characterized the Cladocera fauna of Lake Balaton as
Alona dominant. Sagi (1968) and Bendefy (1968)
reconstructed former water levels by historic maps and
archeological remains while the Geological Institute of
Hungary has performed geological survey by several
proxies as palynology, siliceous algae, trace elements,
and stable-isotope (d13C, d 18O) analysis of carbonates
since 1989 (Cserny & Nagy-Bodor, 2000; Tullner &
Cserny, 2003). These studies revealed that Lake
Balaton is 15,000–17,000 years old, and its develop-
ment started with formation of several shallow, cold
water lakes. Between 5800 and 2500 cal. year. BP,
warming climate and increased precipitation caused
rise of water level as a consequence of which a uniform
lake was formed (Cserny & Nagy-Bodor, 2000). In the
late Holocene, the water level varied approximately
between 104.6 and 112.5 m aAsl (above Adriatic sea
level) as a function of changing climate (Virag, 1998;
Cserny & Nagy-Bodor, 2000).
The sediment of Lake Balaton is rich in carbonate
and its content varies between 50 and 70% depending
on the basins. Carbonate content increases from South-
West (Keszthely) to North-East (Siofok) (Cserny &
Nagy-Bodor, 2000; Tullner & Cserny, 2003).
When water level decreased, Lake Balaton lost
large amount of its surface and the shallowest basins
became wetlands, while when water table lifted up,
area of open water extended and lacustrine conditions
settled. Studying the series of wetlands and lacustrine
deposits in the shallowest area of lakes is a good tool
to reconstruct the past water level changes of large
lake systems such as Lake Balaton. In this study, we
aim to perform reconstruction of changes in water
level of Lake Balaton by geochemistry and subfossils
of Cladocera and diatom remains in the sediment of
the Zalavari Pond as a part of Kis-Balaton wetland.
Our proxies (trace elements, cladocera, and diatom
remains) have been widely used as proxy data to study
history of aquatic ecosystems. Biological proxies as
diatoms (Cameron et al., 1999; Battarbee et al., 2001;
Koster et al., 2004) and cladocerans (Korhola &
Rautio, 2001; Jeppesen et al., 2002) have been shown
to respond sensitively to changes in the past lake water
levels (Korhola & Rautio, 2001; Korhola et al., 2005),
water temperature (Bloom et al., 2003), and eutrophi-
cation (Siver, 1999; Wessels et al., 1999); thus,
subfossil assemblages have become widely used in
226 Hydrobiologia (2010) 641:225–244
123
Author's personal copy

Europe (Buczko et al., 2009) and North America (e.g.,
Harmsworth, 1968; Hofmann, 2001, 2003) for the
reconstruction of paleoclimate.
Materials and methods
Study site
Lake Balaton is the largest shallow lake in Central
Europe. Its catchment area is 5,175 km2, and the major
inflow is the Zala River that discharges about half of the
total Balaton watershed. It has an average discharge of
about 5–10 m3s-1. The only outflow is the artificial Sio
canal that was constructed in the 1860s and connects
the lake to the Danube River. Zala River enters Lake
Balaton through the Kis-Balaton Wetland (KBW), a
large area which acts as a filter for suspended material
and nutrients that would otherwise be deposited in the
lake (Tatrai et al., 2000). The KBW comprises two
parallel lowland basins running north to south, divided
by a low ridge of hills (Figs. 1, 2).
Formerly, Kis-Balaton was the fifth basin of Lake
Balaton until the beginning of the nineteenth century.
From then, it has been called Kis-Balaton. Its open
water area decreased to 8% by the end of eighteenth
century and to 4% by the end of nineteenth century
(Domotorfy et al., 2003). River Zala flushed over the
wetland at high flooding. From the middle of the
nineteenth century, intense canalization was carried
out to form large farmlands in the area of Kis-
Balaton. As farming was not economically efficient,
canalization was abandoned in the 1960s. This
canalization resulted in two major changes:
(1) Only two small water bodies Zalavari (Table 1)
and Vorsi Ponds escaped from desiccation and had
open water areas. (2) The Zala river flew directly into
Lake Balaton instead of filtering through an extended
delta. As a result of increased phosphorus load, the
originally mesotrophic lake underwent rapid eutrophi-
cation: Keszthely basin became hypertrophic during
the 1970s. In the 1980s, large-scale eutrophication
management measures were initiated involving con-
struction of a large reservoir to improve water quality.
The reservoir occupies the former Kis-Balaton delta
area and is, in fact, an extended wetland including open
water and swamp habitats. Water quality of the lake has
been improved due to wetland reconstruction and
investments of drainage and sewage treatment around
the lake (Istvanovics et al., 2007; Hajnal & Padisak,
2008; Tatrai et al., 2008).
Field and laboratory methods
Soil and sediment were sampled at 30 sampling sites
in the Kis-Balaton wetland. Three random samples
Fig. 1 Lake Balaton and its
possible expansion at
several altitude (min. 104.8
recent shore line; max.
113 m aAsl)
Hydrobiologia (2010) 641:225–244 227
123
Author's personal copy

were taken at each sampling point and the uppermost
15 cm layers were used for determination of loss-on-
ignition (LOI550) (Heiri et al., 2001) and carbonate
(Scheibler method) content.
Two parallel, 165 and 148 cm long sediment cores
were taken with a Livingstone corer (Fig. 2). First
core was used for geochemical analysis and diatoms,
the other for identification of Cladocera remains.
Cores were matched stratigraphically by lithography,
organic matter (LOI550), and carbonate (LOI950)
content. Cores were subsampled (1 cm3) at every
5 cm for chemical, diatom, and Cladocera analyses
using a volumetric subsampler.
For loss-on-ignition analysis, samples were weighted
and ignited at 550�C for 4 h (Heiri et al., 2001) and at
950�C for 2 h. Trace element content was determined by
ICP-OES (Thermo Iris Intrepid II XSP). Macrophyte
and charred plant remains were selected from the core
and three radiocarbon dates (Table 3) were determined
at the Laboratory of Environmental Studies, Institute of
Nuclear Research, Hungarian Academy of Sciences in
Debrecen. Dates were calibrated using the software
CALIB REV5.0.0 (Stuiver et al., 2005).
Subsamples (1 cm3) for Cladocera analysis were
treated with 250 ml 10% KOH solution and heated at
80�C for at least 1 h. Subsamples were gently stirred
by glass-rod to deflocculate all organic matter
without damaging the remains and sieved through a
65-lm mesh. Quantitative slides were prepared by
pipetting 100 ll of each subsample on a microscope
slide and were mounted with glycerol jelly stained with
safranine. All slides were then examined under a
microscope at magnifications 100 and 400 and about
200–250 Cladocera remains were counted from each
sample (Korhola & Rautio, 2001). All Cladocera
remains were encountered (carapaces, head-shields,
post abdomen, ephippia, and post-abdominal claws)
separately, but only the most frequent body parts for
each taxon were used for estimating the species
abundance. Poorly fragmented remains were counted
if they contained a clear diagnostic feature as proposed
by Korhola & Rautio (2001). Total concentration per
cm3 was calculated and identification was carried out
by using Frey (1950, 1962, 1988, 1991), Goulden &
Frey (1963), Gulyas & Forro (1999), Sebestyen
(1969b, 1970, 1971), Whiteside et al. (1978).
Fig. 2 The location of
Zalavari Pond and sampling
site. The solid line is the
boundary of the watershed
of Lake Balaton (basins of
the lake: 1 Keszthely, 2Szigliget; 3 Szemes, 4Siofok)
Table 1 Main features of Zalavari Pond
Latitude (N) 46.6669�Longitude (E) 17.2184�Elevation (m aAsl) 106.5 m
Max. depth (m) 1.2
Mean depth (m) 1.1
Surface area (m2) 45,000
Catchments area (km2) 2,125
Mean pH 7.81
Conductivity 622 lS
Total phosphorus 1.95 mg/l
Total nitrogen 0.18 mg/l
228 Hydrobiologia (2010) 641:225–244
123
Author's personal copy
For diatom analysis, 1 cm3 subsamples were
treated with hot hydrogen-peroxide, nitric acid and
the cleaned material was mounted in Zorax (refrac-
tive index is 1.7). At least 300 diatom valves were
counted at each level. For light microscope analysis,
LEICA DM LB2 was used (100 X HCX PLAN
APO). Diatom identification followed the nomencla-
ture of Krammer and Lange-Bertalot (1986–1991),
and the volumes of Diatoms of Europe (Krammer,
2000; Lange-Bertalot, 2001), as well as some other
manuals (Lange-Bertalot & Krammer, 1989; Lange-
Bertalot, 1993; Lange-Bertalot & Metzeltin, 1996;
Houk, 2003; Kobayasi et al., 2006).
Major and trace element concentrations were
assessed by multivariate methods [cluster analysis,
principal component analysis (PCA) with SPSS
(SPSS Inc., 1998)]. Further multivariate analyses
[detrended correspondence analysis (DCA) and
redundancy analysis (RDA)] were carried out with R
(R Development Core Team, 2008), vegan (Oksanen
et al., 2007), and BiodiversityR (Kindt & Coe, 2005)
packages. Stratigraphic zones were determined by
psimpoll (CONISS clustering, Bennet, 2005). Con-
centrations and species data were log-transformed and
standardized for PCA, DCA, and RDA, and PCA
scores were used in cluster analysis (Euclidean
distance with Ward method).
Significance of each environmental variable in
explaining the variance in taxon data was determined
by testing the significance of the first canonical axis
by means of a Monte Carlo permutation test (1,000
random permutations).
Results
Soil, lithology, ages, major and trace elements
stratigraphy
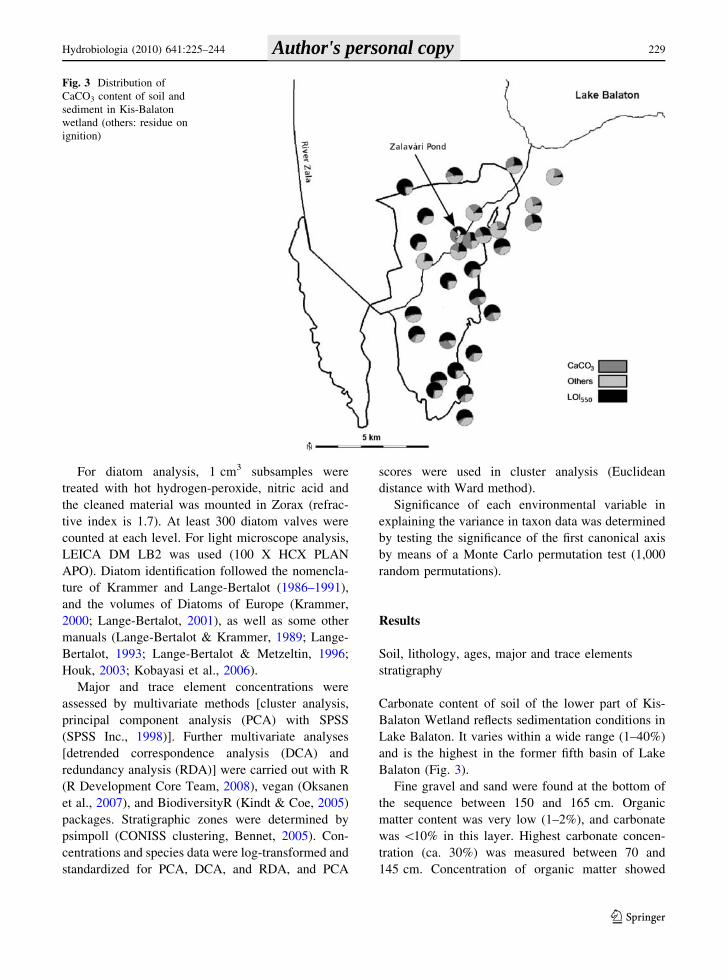
Carbonate content of soil of the lower part of Kis-
Balaton Wetland reflects sedimentation conditions in
Lake Balaton. It varies within a wide range (1–40%)
and is the highest in the former fifth basin of Lake
Balaton (Fig. 3).
Fine gravel and sand were found at the bottom of
the sequence between 150 and 165 cm. Organic
matter content was very low (1–2%), and carbonate
was \10% in this layer. Highest carbonate concen-
tration (ca. 30%) was measured between 70 and
145 cm. Concentration of organic matter showed
Fig. 3 Distribution of
CaCO3 content of soil and
sediment in Kis-Balaton
wetland (others: residue on
ignition)
Hydrobiologia (2010) 641:225–244 229
123
Author's personal copy
maxima (50–60%) and the lowest carbonate concen-
trations (6–7%) between 25 and 70 cm. The upper-
most sediment contained 20–30% organics and
20–25% carbonate, which was similar to the values
measured in surface soil samples in the area of the
fifth basin of Balaton. The core can be divided into 12
lithological units (Table 2) according to Troel-Smith
(1955). The high carbonate content and low organic
yield complicated the radiocarbon dating of the core.
No dates were obtained for the calcareous silts
Table 2 Lithological description of the core from Zalavari Pond
Depth (cm) Troel-Smith
categories
Description
0–5 Th3, Dh1 Dark, blackish-brown peat, containing macroscopically recognisable
plant remains. Organic content high
5–20 Th3, Lc1 Blackish-brown peat with high organic content. The amount of plant
remains is smaller, the amount of calcareous Mollusc shells is large
20–30 Th1, Lc1, Dg1, Dh1 Gray-black sediment, containing wood, Phragmites and Typha remains
30–55 Th2, Dl1, Dg1 Dark brown peat with wooden and herbaceous remains
55–70 Dl3, Dg1 Blackish sediment dominated by wood remains accompanied by
herbaceous remains
70–75 Dl1, Dg2, Ld1 Brownish peat containing charcoal grains
75–85 Lc2, Dg2 Calcareous gray silt with mollusc shells, containing visible plant remains
85–130 Lc3, Dg1 Calcareous gray silt containing less plant remains and numerous mollusc
shells
130–135 Dg1, Ld2, Lc1 Grayish-brown calcareous silt with mollusc shells
135–145 Dl1, Dg2, Ld1 Blackish peat with mollusc remains. Small amount of coarse sand and
small gravel appeared
145–150 Ga1, Ld1, Dl1, Blackish sediment with plant remains. The number of gravel significant
150–165 Ga1, Gg1, Ag2 Gray with black patches. Containing sand and gravel.
The number of 2–5 mm gravels large. Small amount of fibrous plant
remains
Table 3 Conventional,
calibrated radiocarbon ages
and d 13C for Zalavari Pond
Lab-code sample (cm) d 13C (PDB)
± 0.2 (%)
14C year BP Cal. year
BP (2r)
deb-13986 Zalavar 45–50 cm -27.82 1660 ± 35 1569 ± 69
deb-13996 Zalavar 65–70 cm -28.74 4135 ± 50 4680 ± 148
deb-13994 Zalavar 140–148 cm -28.24 7935 ± 60 8801 ± 184
Fig. 4 Age-depth model for Zalavari Pond
Table 4 Redundancy analysis of Cladocera remains and
major and trace elements partitioning of variance
Inertia Proportion
Total 14.145 1.0000
Constrained 7.792 0.5509
Unconstrained 6.535 0.4491
Axis RDA1 RDA2 RDA3 RDA4
Lambda 1.3123 1.1512 0.8356 0.7018
Variance explained % 20.6576 18.1211 13.1533 11.047
230 Hydrobiologia (2010) 641:225–244
123
Author's personal copy
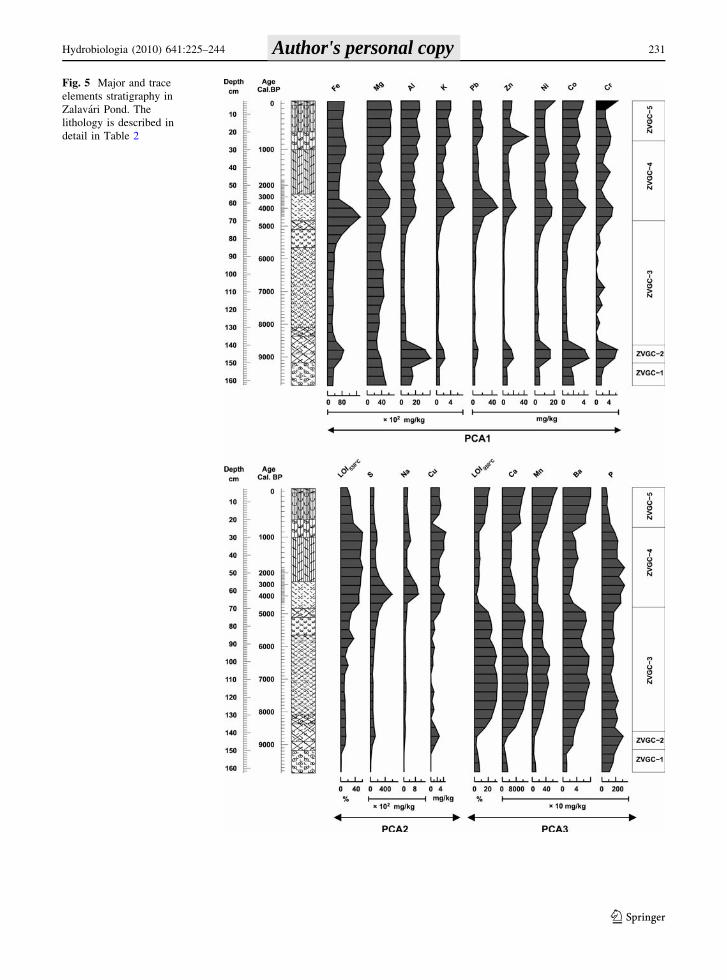
Fig. 5 Major and trace
elements stratigraphy in
Zalavari Pond. The
lithology is described in
detail in Table 2
Hydrobiologia (2010) 641:225–244 231
123
Author's personal copy

between 70 and 140 cm. Thus, the chronology of this
sequence is not ideal. The results of the three
radiocarbon dates are presented in Table 3. Relation-
ship between age and depth is not linear (Fig. 4).
Highest sedimentation rate (0.3 mm year-1) was
found in the youngest part (0–47.5 cm) of the core,
whereas the lowest (0.1 mm year-1) was between
47.5 and 67.5 cm.
Three principal components and five stratigraphic
zones were distinguished by the multivariate analysis
(Fig. 5). The first three principal components were
explained 89.6% of total variance. The trace elements
of sediment of the Zalavari Pond could be classified
into three groups. Elements used as indicators of
erosion (Al, K, Mg, Cr, Co, and Ni) and human
impact (Pb and Zn) were loaded into the first
principal component (PCA1). The redox indicator,
Fe was also loaded into the first principal component.
Each of these elements showed positive correlations,
and their concentrations are higher in the ZVGC-2
(140–155 cm) and the ZVGC-4 and ZVGC-5 (0–
70 cm) zones than they were in the others. The
organic matter (LOI550), sulfur, sodium, and copper
loaded into the second group (PCA2), refers to
wetland development. Concentration of these com-
ponents is higher in the upper part of the sequence
(20–70 cm). The third group (PCA3) indicates
lacustrine environment with higher concentrations
of Ca, Mn, and carbonate content (LOI950).
ZVGC-1 (9900–9100 cal. year BP; 165–150 cm)
Concentration of elements indicating erosion (PCA1
group) is high. The sediment is poor in carbonates
and organic matter. Appearance of fine gravels and
coarse sand confirms higher level of erosion. Low Ca/
Mg ratio shows that water level might be lower with
poor vegetation.
ZVGC-2 zone (9100–8600 cal. year BP;
150–140 cm)
Color of the sediment in this zone is darker than
either above or below. Carbonate concentration
remained low and Ca/Mg ratio decreased. Concen-
trations of Fe, Mn, S, Cu, P, and organic matter have
a local maximum at 9000 cal. year BP. The increase
of Fe is higher than that of Mn, which causes a high
Fe/Mn ratio. Sulfur concentration increased from 1 to
13 g kg-1. These phenomena may indicate shallow
water and reductive conditions and increase in the
trophic state of the lake. Intense erosion is indicated
by gravel, coarse sand, and high concentration of
aluminum and other elements loaded in PCA1.
ZVGC-3 zone (8600–4800 cal. year BP; 140–70 cm)
Color is grayish with high concentration of calcium,
manganese, barium, and carbonate. Amount of Mg is
also significant. Ca and carbonate concentrations
gradually increase toward the middle of the zone, and
then they decrease indicating water level changes.
Concentration of Al and all others in PCA1 group is
very low. Water level was high in the middle of the
zone. Later, organic matter content increased upward
the sequences and wetland phase started to be
established.
ZVGC-4 zone (4800–800 cal. year BP; 70–25 cm)
Organic content varies between 50 and 60%. The
high peak of Fe/Mn ratio indicates that redox
conditions turned to anoxic again. The average
concentrations of Al, K, Pb, Zn, Ni, Co, Cr, S, Na,
Cu, and P are higher than in ZVGC-3 zone, while the
amount of Ca, Mn, Ba, and carbonate decreases.
ZVGC-5 zone (800–0 cal. year BP; 25–0 cm)
The concentration of Fe, Al, Ni, Co, Cr, S, and Na is
similar to that of ZVGC-4. Organic content is lower,
while the concentration of Pb is slightly higher in this
zone. Ca, Mn, Ba, and carbonate content show a
strong increase. In the topmost 5 cm, the concentra-
tion of Ni, Co, and Cr slightly increases.
Cladocera
There were remains of 27 Cladocera species along the
core of Zalavari Pond. Species numbers varied
between 11 and 19 in the samples. These species are
very common in littoral habitats of eutrophic shallow
lakes with dense macrophytes, and in Lake Balaton as
well. Only ten species achieved dominant status
([10% in at least one subsample), of which nine were
chydorids, and only three long-term dominants were
232 Hydrobiologia (2010) 641:225–244
123
Author's personal copy
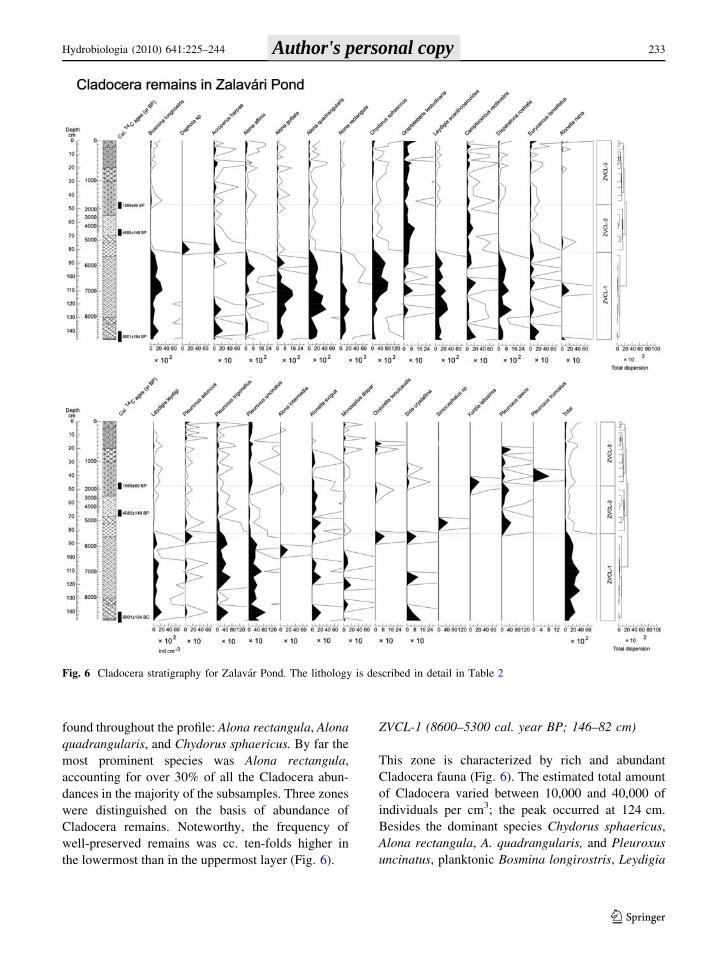
found throughout the profile: Alona rectangula, Alona
quadrangularis, and Chydorus sphaericus. By far the
most prominent species was Alona rectangula,
accounting for over 30% of all the Cladocera abun-
dances in the majority of the subsamples. Three zones
were distinguished on the basis of abundance of
Cladocera remains. Noteworthy, the frequency of
well-preserved remains was cc. ten-folds higher in
the lowermost than in the uppermost layer (Fig. 6).
ZVCL-1 (8600–5300 cal. year BP; 146–82 cm)
This zone is characterized by rich and abundant
Cladocera fauna (Fig. 6). The estimated total amount
of Cladocera varied between 10,000 and 40,000 of
individuals per cm3; the peak occurred at 124 cm.
Besides the dominant species Chydorus sphaericus,
Alona rectangula, A. quadrangularis, and Pleuroxus
uncinatus, planktonic Bosmina longirostris, Leydigia
Fig. 6 Cladocera stratigraphy for Zalavar Pond. The lithology is described in detail in Table 2
Hydrobiologia (2010) 641:225–244 233
123
Author's personal copy

leydigi, L. acanthocercoides, Alona affinis, Graptol-
eberis testudinaria, A. guttata, and Disparalona
rostrata were numerous.
ZVCL-2 (5300–1600 cal. year BP; 82–47 cm)
Frequency of remains of dominant Cladocera species
decreased in numbers in this zone. Total abundance
of cladocerans decreased; it varied between 700 and
2500 ind. cm-3. The pelagic Bosmina longirostris
were found in very low numbers. Besides the
dominant species, Alonella exigua, Pleuroxus trigo-
nellus, Camptocercus rectirostris, Graptoleberis
testudinaria, Pleuroxus uncinatus, and Acropterus
harpae were found in higher numbers. These species
occur in large number in shallow and densely
vegetated habitats. Increased frequency of these
species and Camptocercus rectirostris reflects the
expansion of vegetated bed. Some Daphnia ephippia
and Simocephalus claws were only found in the
bottom of this zone.
ZVCL-3 (1600–0 cal. year BP; 47–0 cm)
Number of Cladocera remains was low (500–4200 ind.
cm-3) but slightly increased toward the uppermost
layers, and reached maximum in (4200 ind. cm-3)
the topmost subsample. Number of the pelagic species
such as Bosmina longirostris was higher than in the
previous zone. Abundance of Pleuroxus uncinatus
slightly increased in this zone. Kurzia latissima and
Sida crystallina were found in the lower part of the
core (Fig. 6). Besides that the species composition of
this zone is similar to the previous one indicating
wetland phase, increasing number of remains of
Bosmina longirostris shows increased trophic levels.
Diatoms
The diatom flora of the core from Zalavari Pond was
diverse and variable with more than 150 taxa. Some of
the most frequent species were present through the
sequence like Amphora ovalis, Aulacoseira granulata,
A. ambigua, Pseudostaurosira brevistriata, and Staur-
oneis construens varying in abundances. Amphora
pediculus and Cocconeis placentula were also con-
stant members of the flora and they were missing
only from one sample. Summarizing the abundance,
Pseudostaurosira brevistriata, Stauroneis construens,
S. construens var. venter, and Staurosira pinnata were
the most characteristic members of the diatom assem-
blages of Zalavari pond. All of them are members of
the sand diatom flora; they are common benthic taxa
(Fig. 7). Mainly alkalophilous or indifferent diatom
species were recorded along the sequence, indicating
that there was no significant change in pH.
ZVD-1 (9900–7700 cal. year BP; 165–126 cm)
Staurosira construens–Staurosirella pinnata zone
Low diatom diversity in every aspect was the main
feature of this zone. The number of species was low
(10–20), uniformity of forms and dimensions were
characteristic, and mainly small sized (\20 lm)
diatoms were present. Some dissolved fragments of
larger diatoms were also recorded. Staurosira
construens and Staurosirella pinnata dominated the
assemblages. The extremely low abundance of
planktonic species indicated the lack of deep water.
Wind-disturbed shallow water without macrophytes
can be reconstructed on the basis of the mud diatom
flora.
ZVD-2 (77000–1900 cal. year BP; 64–50 cm)
Aulacoseira ambigua–A. granulata zone Between
126 and 64 cm, there was poor diatom preservation.
The diatom flora of this zone is characterized by high
diversity (the number of taxa in the samples varied
between 46 and 72). Planktonic forms, such as
Aulacoseira ambigua, A. granulata, A. granulata
var. angustissima, and Stephanodiscus species were
the most frequent in this zone. The ratio of planktonic
forms was[50%, indicating the possible existence of
extended open water. Cymbella neocistula was found
only in this zone.
ZVD-3 (1900–1300 cal. year BP; 50–38 cm)
Navicula tenelloides–Fallacia tenera zone: Abundant
diatom flora was of great diversity (Shannon–Weaver
diversity was between 4.81 and 5.04) with a decrease
in Aulacoseira spp. and an increase in benthic taxa.
Apart from Aulacoseira species, the ratio of other
planktonic taxa remained relatively high indicating a
gradual decrease in the depth of water. The nominal
species of this zone (Navicula tenelloides) attained a
234 Hydrobiologia (2010) 641:225–244
123
Author's personal copy

definite peak exactly in the middle of the zone (44 cm).
In this sample, 20% of the total diatom floras belong to
this species. Its abundance gradually decreased above
and below this level. The presence of Fallacia tenera
was characteristic for this zone. In addition,
Achnanthes delicatula and Platessa conspicua
Fig. 7 Diatom stratigraphy in Zalavari Pond. The lithology is described in detail in Table 2
Hydrobiologia (2010) 641:225–244 235
123
Author's personal copy

showed peaks in this zone further supporting the
shallowness of the lake.
ZVD-4 (1200–400 cal. year BP; 38–14 cm)
Staurosirella pinnata–Gyrosigma attenuatum zone: The
ratio of planktonic species further decreased and
benthic forms became dominant. Staurosirella
pinnata attained a peak at 32 cm. Amphora species
(Amphora ovalis and A. pediculus) were the most
frequent in this zone. Ulnaria ulna and Fragilariforma
virsecens also reached maximum abundances in this
zone. Diatom preservation was extremely poor
between 16 and 22 cm. This is indicated in Fig. 7 by
white shading. Some Gyrosigma fraction, broken
Epithemia valves, strongly silicified Aulacoseira
valves, and some small Fragilaria (sensu lato) were
found in these samples. These sporadic diatom remains
suggest that the flora and the composition of diatom
assemblages did not change; only the preservation was
unsatisfactory.
ZVD-5; (400–0 cal. year BP; 14–0 cm)
Pseudostaurosira brevistiata–Cocconeis placentula
zone: The diatom flora of this zone is characterized
by great taxonomic diversity and abundant
occurrence of benthic taxa (up to 90%). However,
the abundance of the planktonic forms gradually
decreased. Pseudostaurosira brevistiata was the most
frequent and abundant diatom in the upper part of the
zone. Staurosira construens and its varieties are also
frequent and abundant here. Increasing abundance of
the epiphytic Cocconeis placentula is detected in this
zone.
Redundancy analysis
As the ordination analyses applied to this study are
sensitive to rare species, only cladoceran taxa occurred
with Hill’s number [10. Of the initial 27 taxa, 18
fulfilled these criteria. Cladocera data yielded DCA
gradient lengths of 1.42372 and 1.04327 SD for axes 1
and 2, favoring the use of redundancy analysis (RDA).
Geochemical variables with high multicollinearity
[Variance Inflation Factor (VIFs [20)] were elimi-
nated one at time beginning with the variety having the
highest VIF, and the procedure was repeated until all
environmental VIFs were \20. Result of analysis on
the reduced datasets shows that 55% of total variance
of Cladoceran data were accounted by chemical data
and 45% remained unexplained. The eigenvalue of the
first two RDA axes, 5.38 and 0.91 captured 44% of the
cumulative variance in the Cladocera data, a slightly
larger proportion of variance than is explained by PCA
(38.77%) (Table 4). Monte Carlo permutation test
revealed that Na, Pb, and Fe contents were the most
explanatory environmental variables in the distribution
of Cladocera remains (Table 5). Since Fe has real
ecological value on the dispersion of cladoceran data,
as a redox indicator, and the PCA analysis revealed that
chemical variables were explained by PCA factors,
repeated RDA analysis were done on the cladoceran
dispersion with PCA scores of chemical data.
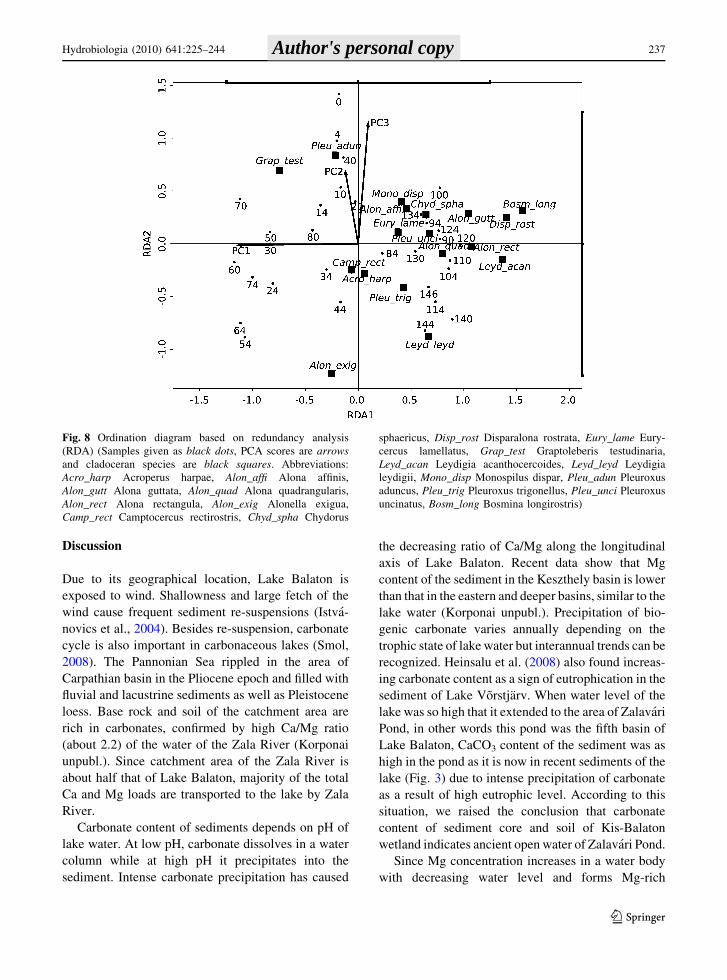
Result of the RDA is illustrated in Fig. 8 as a
Cladocera—chemical data biplot. On the biplot, PCA
factors are shown as arrows and the length of the
arrows indicates their relative importance in explain-
ing the species variation; whereas their orientation is
proportional to their approximate correlation to the
RDA axes. In this RDA, axis RDA1 is most clearly
related to those groups of elements that were found in
high concentration in wetland phase of Zalavari
Pond. This axis effectively separates the top sediment
layers. Cladocerans were found with low abundance
in these layers. RDA2 axis is related to those groups
of chemicals that contain carbonate, Ca, Mn, and P.
RDA2 axis also correlates with those groups of
elements that is explained by PC2 factor, of which
concentration is higher in the ZVGC-4 zone.
Table 5 Result of permutation test (1000 Monte Carlo per-
mutation) in constrained RDA of Cladocera data
Df Var F N. Perm Pr ([F)
Na 1 3.305 10.9259 100.000 0.01 **
Pb 1 1.214 4.0143 100.000 0.01 **
Zn 1 0.214 0.7080 100.000 0.72
Ba 1 0.644 2.1285 100.000 0.08 .
Fe 1 0.875 2.8940 100.000 0.04 *
LOI550 1 0.239 0.7904 100.000 0.60
P 1 0.495 1.6364 100.000 0.08 .
Cu 1 0.332 1.0961 100.000 0.30
Cr 1 0.473 1.5640 100.000 0.15
Residual 21 6.353
Signif. codes: 0 ‘***’ 0.001 ‘**’ 0.01 ‘*’ 0.05 ‘.’ 0.1 ‘’ 1
236 Hydrobiologia (2010) 641:225–244
123
Author's personal copy
Discussion
Due to its geographical location, Lake Balaton is
exposed to wind. Shallowness and large fetch of the
wind cause frequent sediment re-suspensions (Istva-
novics et al., 2004). Besides re-suspension, carbonate
cycle is also important in carbonaceous lakes (Smol,
2008). The Pannonian Sea rippled in the area of
Carpathian basin in the Pliocene epoch and filled with
fluvial and lacustrine sediments as well as Pleistocene
loess. Base rock and soil of the catchment area are
rich in carbonates, confirmed by high Ca/Mg ratio
(about 2.2) of the water of the Zala River (Korponai
unpubl.). Since catchment area of the Zala River is
about half that of Lake Balaton, majority of the total
Ca and Mg loads are transported to the lake by Zala
River.
Carbonate content of sediments depends on pH of
lake water. At low pH, carbonate dissolves in a water
column while at high pH it precipitates into the
sediment. Intense carbonate precipitation has caused
the decreasing ratio of Ca/Mg along the longitudinal
axis of Lake Balaton. Recent data show that Mg
content of the sediment in the Keszthely basin is lower
than that in the eastern and deeper basins, similar to the
lake water (Korponai unpubl.). Precipitation of bio-
genic carbonate varies annually depending on the
trophic state of lake water but interannual trends can be
recognized. Heinsalu et al. (2008) also found increas-
ing carbonate content as a sign of eutrophication in the
sediment of Lake Vorstjarv. When water level of the
lake was so high that it extended to the area of Zalavari
Pond, in other words this pond was the fifth basin of
Lake Balaton, CaCO3 content of the sediment was as
high in the pond as it is now in recent sediments of the
lake (Fig. 3) due to intense precipitation of carbonate
as a result of high eutrophic level. According to this
situation, we raised the conclusion that carbonate
content of sediment core and soil of Kis-Balaton
wetland indicates ancient open water of Zalavari Pond.
Since Mg concentration increases in a water body
with decreasing water level and forms Mg-rich
Fig. 8 Ordination diagram based on redundancy analysis
(RDA) (Samples given as black dots, PCA scores are arrowsand cladoceran species are black squares. Abbreviations:
Acro_harp Acroperus harpae, Alon_affi Alona affinis,
Alon_gutt Alona guttata, Alon_quad Alona quadrangularis,
Alon_rect Alona rectangula, Alon_exig Alonella exigua,
Camp_rect Camptocercus rectirostris, Chyd_spha Chydorus
sphaericus, Disp_rost Disparalona rostrata, Eury_lame Eury-
cercus lamellatus, Grap_test Graptoleberis testudinaria,
Leyd_acan Leydigia acanthocercoides, Leyd_leyd Leydigia
leydigii, Mono_disp Monospilus dispar, Pleu_adun Pleuroxus
aduncus, Pleu_trig Pleuroxus trigonellus, Pleu_unci Pleuroxus
uncinatus, Bosm_long Bosmina longirostris)
Hydrobiologia (2010) 641:225–244 237
123
Author's personal copy

calcite in the sediments (Cserny & Nagy-Bodor,
2000), high Mg concentration of sediments reflects
low water level in the given basin.
Changes in the plankton:periphyton ratio of dia-
tom assemblages indicate water level fluctuation
(Heinsalu et al., 2008). Different preservations of
diatom frustules take a bias into reconstruction of
paleoenvironment (Battarbee et al., 2005). Poorly
preserved diatom frustules in this region should
reflect to shallow water also, since temperature, pH,
salinity, grazing or bioturbation, and water depth
affect diatom preservation (Flower, 1993). Smol &
Boucherle (1985) and Newberry & Schelske (1986)
found poor preservation of frustules in early Holo-
cene, and speculated that enhanced dissolution during
this period was linked to the higher lake water pH at
the time of carbonate deposition from the core of the
Little Round Lake, Canada. Diatoms were poorly
preserved in calcareous sediments from Lake Malawi
(Gasse et al., 1995), and from ephemeral lakes
(Flower, 1993). Barker et al. (1994) found that the
pH is the most significant factor affecting silica
dissolution, and the dissolution increases exponen-
tially when pH is above 9.
Water pH of Lake Balaton is high, commonly
exceeding 8.5 in summer and this condition is
coinciding with high carbonate content (50–55%).
Under such conditions, Cserny et al. (1991) found
poorly preserved diatoms in the sediment of Lake
Balaton.
In the sediment of Zalavari Pond, high Ca/Mg was
found in the zone ZVGC-3 suggesting the water level
was relatively high (Fig. 9).
Ratio of planktonic (e.g., bosminids and daphnids)
to littoral (e.g., chydorids) Cladocera species is widely
used for paleolimnological reconstruction of lake
Fig. 9 Ca/Mg and Fe/Mn
ratio in sediment of Zalavari
Pond and zones of studied
proxies. The lithology is
described in detail in
Table 2
238 Hydrobiologia (2010) 641:225–244
123
Author's personal copy

water level changes (Sarmaja-Korjonen & Alhonen,
1999; Korhola et al., 2000; Sarmaja-Korjonen, 2001).
Different chitinisation of exosceletons of microcrust-
acens, comparing bosminids and daphnids, and ecol-
ogy could biased in reconstructed community.
Hofmann (1998) warned that using planktonic/littoral
ratio was not suitable for all cases. He found that
Bosmina longirostris was present at shallow periods,
and B. coregoni occurred in deep layers in Belauer See.
High carbonate content in water body has strong
effects on cladoceran community. Re-suspension and
intense carbonate precipitation make Lake Balaton a
stressful habitat for cladocerans especially for daph-
nids (G.-Toth, 1992). As a result, daphnids occur in
low numbers among recent zooplankton communities
and in the sediment as well. In the recent plankton, B.
longirostris and B. coregoni, Diaphanosoma brachiu-
rum are the dominant cladoceran species. Bosminids
are considered as planktonic species, but they occur
both in open water and in the littoral planktonic
habitats (Korhola & Rautio, 2001). Thus, amount of
bosminid remains may reflect extension of open
water (Gasiorowski & Kupryjanowicz, 2009).
Although Bosmina longirostris indicates rather eutro-
phic conditions than water level changing (Szeroc-
zynska pers. comm), we can consider this species as
good indicator of water level changes in shallow and
eutrophic ecosystems. Whenever water table of the
lake was elevated during the lake’s history, large
areas were inundated in Kis-Balaton and Nagy-Berek
areas; thus littoral zone increased in the western and
southern parts of Lake Balaton (Fig. 1). Since B.
longirostris prefers shallow habitats, its abundances
will increase with extension of littoral zone. We
found B. longirostris remains along the core both in
wetland and lacustrine sediments, but their density
was the highest in the 146–82 cm deep core section;
therefore, high abundances of B. longirostris indi-
cated higher water level in shallow environment.
Monospilus dispar is most abundant in the deepest,
Siofok basin while Bosmina longirostris is most
abundant in the largest Szigliget and the Keszthely
basins of Lake Balaton (Ponyi, 1971, Gyulai,
unpubl.).
Besides carbonate content, we found that redox
potential indicates water level changes, precisely, the
transition from lake to wetland as well. In the core of
Zalavari Pond, deviating trends are observed in
parameters often considered as indicators of redox
changes (Fe, Mn, and Fe/Mn ratio, Figs. 5, 9). Fe and
Mn exhibit very low solubility in oxidative environ-
ment, but Mn is more soluble than Fe; thus it is
mobilized at first when oxygen content decreases
(Engstrom & Wright, 1984). Concurrently, low Ca/
Mg and high Fe/Mn ratios indicate low water level
and wetland environment (Fig. 9).
Sedimentation was similar to that observed in
F}onyed and Zalavar (Medzihradszky, 2005; Juhasz
et al., 2007) and in the Keszthely basin of Lake
Balaton (Cserny and Nagy-Bodor, 2000). Cores from
F}onyed and Zalavar were taken close to the shoreline
of Kis-Balaton wetland and authors did not recognize
any sign of hiatus in sediment profiles.
At 10000–8600 cal. year BP years, the climate
was cold and dry and the water level of Lake Balaton
was low (Cserny & Nagy-Bodor, 2000; Juhasz et al.,
2007). Decreased runoff of rivers and creeks results
in the decrease of ionic concentrations in the lake
water. We found high Mg and trace metal concen-
tration in this zone, and low Ca/Mg ratio. These
phenomena indicate that water level was low and the
high Fe/Mg ratio reflects low redox environment
(Koinig et al., 2003), which infer the pond turned to
wetland (Fig. 9). Vegetation was sparse in the ancient
pond as shown by low organic matter content in the
sediment. The absence of planktonic diatom species
indicated the lack of deep water. Cladocera commu-
nity was characterized by phytophylous species, but
occurrence of Monospilus dispar proved large bare
lake bottom; macrophytes could be localized at
shoreline.
Between 8500 and about 5000 cal. year BP zone,
climate became warmer and wetter thus the water
level of the lake elevated (Cserny & Nagy-Bodor,
2000; Juhasz et al., 2007). In this period, dense
planktonic and littoral community developed simi-
larly to the recent Keszthely basin. This environment
was well oxygenated by activity of algae and
macrophytes and biogenic carbonate precipitation
increased Ca/Mg ratio and decreased Fe/Mg ratio in
the sediment.
The absence of diatoms between 8000 and 4000 cal.
year BP coincides with the high carbonate content in
the sediment due to high water level. Zalavari pond
must have been as large and deep as the recent
Keszthely basin as indicated by the poorly preserved
diatom zones; dissolution of silica frustules was
substantial in sediment in relation to strongly alkaline
Hydrobiologia (2010) 641:225–244 239
123
Author's personal copy

water with pH[ 9.0 or to the high carbonate content
(Barker et al., 1994; Reed, 1998). Water alkalinity also
depends on trophic level. Within eutrophic conditions,
pH can exceed 9 with high carbonate precipitation.
High amount of Cladocera remains and large number
of remains of Bosmina longirostris and the occurrence
of mud living Monospilus dispar also indicate deeper
water. Since chydorid fauna was dense and rich, we
could conclude that water level was high accomplished
with dense submerged vegetation along the shore in
large open water like the recent Keszthely basin is.
About 5000 cal. year BP, climate changed, con-
firmed intensive terrestrial erosion caused by human
deforestation (Sumegi et al., 2007). Human population
occupied the region in the pre-Neolithic, but the
agrarian population arrived around 7500 cal. year BP
as indicated by the relative increase of hazel pollen
(Juhasz et al., 2007). Strong human impact can be
observed from Middle Copper Age (Banffy, 2007)
when dominance of dock plantain and nettle pollen
indicated deforestation (Juhasz et al., 2007). Increasing
trace element concentrations from 5000 cal. year BP
coincide with intense deforestation while population
of Balaton-Lasinja culture had pastoral economy
(Banffy, 2007).
High organic matter content with low carbonate
and high Fe/Mn ratio (oxygen depletion) indicates the
shallowness and transition of the pond system to
wetland. At the bottom of ZVGC-4 section, high Fe
and S concentrations show bounding of pyrite (FeS2)
that is common in wetland with extremely low redox
environment. Pyrite can be found in the Holocene
sediment of Lake Balaton (Cserny et al., 1991).
In shallow environments, carbonate precipitation
is less intense due to decreased importance of
planktonic primary production. Among the cladoc-
erans, abundance of phytophylous species as Gra-
ptoleberis testudinaria, Camptocercus rectirostris,
Pleuroxus laevis increased. Water level in this period
had no effects on diatom communities, among which
the planktonic/tychoplanktonic diatoms were domi-
nant. We are able to solve the discrepancy between
diatoms and the other proxies if we assume the pelagic
habitat was so large among also extended littoral
(larger than in the historic eras) that pelagic diatoms
could develop but large emerged macrophyte i.e.,
reeds protected this basin from disturbance by wind.
Low carbonate content and cladoceran remains show
us the possible water level decrease. The gradually
decreasing but still relatively high water level per-
mitted the preservation of diatoms valves from
4000 cal. year BP.
Low concentration of Cladocera remains, rich and
divers diatom flora confirm the low water level
hypothesis. From about 1000 cal. year BP up to
recently, water level slightly increased as indicated
by higher carbonate content coinciding with lower
organic matter and slightly increasing Ca/Mg and
decreasing Fe/Mn ratios. We could reconstruct low
water level with dense macrophyte and high P
content since strong correlation exists between
submerged macrophyte and P accumulation (Rooney
et al., 2003), in ZVGC-2 and ZVGC-4 zones (Fig. 5).
Several studies proved uneven and mosaic-like
hydrological changes in the area of historical Lake
Balaton (e.g., Cserny & Nagy-Bodor, 2000; Jakab
et al., 2005). Jakab et al. (2005) found lacustrine
layers in 420–390 cm depth in the sediment of
Szigliget bay in the northern wetland of the lake, and
concluded that the water level was the highest around
5200 cal. year BP. It is consistent with the result of
Cserny & Nagy-Bodor (2000) who stated there was
humid climate, water level increased and modern lake
formed between 5800 and 2500 cal. year BP. Our
data, however, contradict these reconstructions since
lacustrine sediments were found in 140–70 cm depth;
it means that there was high water level between
8600 and 5000 cal. year BP.
This paleoenvironment of the pond is comparable
with the recent phase of Lake Balaton. In that time,
the pond was connected to the lake and it was
composed of a well-developed wetland with large
open water. In the Lake Balaton, waves induced by
weak wind (4 m s-1) are able to reach the bottom of
the lake and their energy dissipates in the surface of
the sediment disturbing it parallel. In that time, the
long wind fetch expanded into the Zalavari Pond. The
wind in the Lake Balaton results in almost permanent
mixing of the whole water column. Due to the
frequent re-suspension of highly calcareous (50–60%
carbonates) sediments and slow sedimentation of
precipitated carbonates, the water is always turbid
and phytoplankton and submerged vegetation become
light-limited. Although reduction of water level
increases light penetration and improve light climate
for submerged macrophytes at canopy level (Scheffer,
1998), vegetation remains weak at low water level of
the Lake Balaton due to the strong wind effect.
240 Hydrobiologia (2010) 641:225–244
123
Author's personal copy

According to Istvanovics et al. (2008), 12% of the
surface area is available for plant growth, and this
situation has been characteristic of Lake Balaton.
Virag (1998) concluded that the area of emergent
plants (mainly reed) has shown little variation
(11–18 km2, \3% of the surface area) from the end
of the eighteenth century, but the expansion was the
most striking in Keszthely Basin. Although the
formation of a macrophyte-dominated stable state
can be excluded in Lake Balaton irrespective of the
trophic conditions, patches that represent this state can
develop locally. Entz & Sebestyen (1946) demon-
strated that species richness of algae and invertebrate
communities of patches of Potamogeton perfoliatus
and Myriophyllum spicatum is much higher in all the
seasonal succession than the plant-free littoral. Bıro &
Gulyas (1974) found Alona affinis and Chydorus
sphaericus the most common taxa in Lake Balaton and
Alona rectangula, Disparalona rostrata, and Macro-
thrix laticornis were restricted to the reed-grass stand
of Keszthely basin. They also found Sida crystallina,
that occurred commonly in Lake Balaton. Reed-grass
vegetation of the Keszthely Bay at the inflow of Zala
River was characterized by the highest number of
species, both planktonic and littoral. In comparison,
the Cladocera community between 8600 and 5300 cal.
year BP in the Zalavari Pond and the recent fauna in
the Keszthely Basin of Lake Balaton we concluded
that they correspond to similar water level.
Water level has been recorded since 1803 and was
regularized since 1863. Datum level of the official
water gauge was established at 104.09 m aAsl. Since
water table is regulated between ?70 and ?110 cm
and the bottom of Keszthely basin is between 101.7
and 102.2 m aAsl, the maximum depth is cc. 3 m in
the Keszthely basin at the moderate water level.
Recently, the water table of Zalavari Pond stabilized
around 106.1 m aAsl. According to our measures, the
maximum depth of Zalavari Pond is 1.2 m; thus
bottom of Zalavari Pond is at higher altitude
(104.9 m aAsl). We found similar Cladocera com-
munity in Zalavari Pond between 8600 and 5300 BP
to recent Keszthely Basin. In this sense, water level
of this period in Zalavari Pond must be similar to the
recent level of Keszthely Basin, suggesting that it has
to be min. 3 m higher than the regulated level.
Result of new archeological excavations in the
southern shore of the Lake Balaton revealed the
archeological remains from Copper Age to Migration
period (7200–1500 cal. year BP) were found between
107.7 and 112.7 m aAsl in southern shore (Serlegi,
2007). This reconstructed water level changes are not
presented in the sediment records due to the short
duration of high water level stages. Our result
confirms that the water level of Lake Balaton was
higher between 8600 and 5000 cal. year BP than the
present.
Acknowledgments The authors thank their colleagues from
Laboratory of West-Transdanubian District Water Authority for
their help with data collection and collaboration. We would like
to express our special thanks for Kaarina Sarmaja-Korjonen for
her valuable comment. This study was financially supported by
Hungarian National Science Foundation, OTKA-T 049098 and
Hungarian National Research and Development Program
BALOKO 3B022/04.
References
Banffy, E., 2007. Settlement patterns in the Little Balaton
region and the Balaton Uplands. In Zlatyko, C., I. Juhasz
& P. Sumegi (eds), Environmental Archaeology in
Transdanubia. Varia Archaeologica Hungarica XX.
Budapest: 97–105.
Barker, P. A., N. Roberts, H. F. Lamb, S. Kaars & A. Ben-
kaddour, 1994. Interpretation of Holocene lake-level
change from diatom assemblages in Lake Sidi Ali, Middle
Atlas, Morocco. Journal of Paleolimnology 12: 223–234.
Battarbee, R. W., V. J. Jones, R. J. Flower, N. G. Cameron &
H. Bennion, 2001. Diatoms. In Smol, J. P., H. J. B. Birks
& W. M. Last (eds), Tracking Environmental Change
Using Lake Sediments, Vol. 3. Terrestrial, Algal and
Siliceous Indicators. Kluwer Academic Publishers,
Dordrecht, The Netherland: 155–202.
Battarbee, R. W., A. W. Mackay, D. H. Jewson, D. B. Ryves &
M. Sturm, 2005. Differential dissolution of Lake Baikal
diatoms: correction factors and implications for palaeo-
climatic reconstruction. Global and Planetary Change 46:
75–86.
Bendefy, L., 1968. A Balaton vızszintjenek valtozasai a neol-
iltikumtol napjainkig. (Water level changes of Lake
Balaton from Neolitic Era to The Present). Hidrologiai
Kozlony 48: 257–263. (in Hungarian with English
summary).
Bennet, K. D., 2005. Documentation for psimpoll 4.25 and
psimcomb 1.3. C program for plotting pollen diagram and
analysis pollen data.
Bıro, K. & P. Gulyas, 1974. Zoological investigations in the
open water Potamogeton perfoliatus stands of Lake Bal-
aton. Annales Instituti Biologici (Tihany) Hungariae
Academie Scientarium 41: 181–203.
Bloom, A. M., K. A. Moser, D. F. Porinchu & G. M. Mac-
Donald, 2003. Diatom-inference models for surface-water
temperature and salinity developed from a 57-lake cali-
bration set from the Sierra Nevada, California, USA.
Journal of Paleolimnology 29: 235–255.
Hydrobiologia (2010) 641:225–244 241
123
Author's personal copy

Buczko, K., E. K. Magyari, P. Bitusik & A. Wacnik, 2009.
Review of dated Late Quaternary palaeolimnological
records in the Carpatian Region, east-central Europe.
Hydrobiologia 631: 3–28.
Cameron, N. G., H. J. B. Birks, V. J. Jones, F. Berges, J.
Catalan, R. J. Flower, J. Garcia, B. Kawecka, K. A.
Koinig, A. Marchetto, P. Sanchez-Castillo, R. Schmidt,
M. Sisko, N. Solovieva, E. Stefkova & M. Toro, 1999.
Surface-sediment and epilithic diatom pH calibration sets
for remote European mountain lakes (AL:PE Project) and
their comparison with the Surface Waters Acidification
Programme (SWAP) calibration set. Journal of Paleo-
limnology 22: 291–317.
Cserny, T. & E. Nagy-Bodor, 2000. Limnogeology of Lake
Balaton (Hungary). In Gierlowski-Kordesch, E. H. &
K. R. Kelts (eds), Lake Basins Through Space and Time:
AAPG Studies in Geology, Vol 46. Tulsa, OK: 605–618.
Cserny, T., M. Foldvari, K. Ikrenyi, E. Nagy-Bodor, M. Hajos,
A. Szuromi-Korecz & I. Wojnarovits-Hrabka, 1991. A
Balaton aljzataba melyıtett To-24. sz. furas foldtani viz-
sgalatanak eredmenyei. (Geological investigations of the
lacustrine sediments of Lake Balaton based on the bore-
hole TO 24). Annual Report of the Geological Institute of
Hungary: 178-239. (in Hungarian with English summary).
Domotorfy, Z., D. Reeder & P. Pomogyi, 2003. Changes in the
macro-vegetation of the Kis-Balaton Wetlands over the
last two centuries: a GIS perspective. Hydrobiologia 506–
509: 671–679.
Engstrom, D. R. & H. E. Wright Jr., 1984. Chemical stratigraphy
of lake sediments as a record of environmental change. In
Haworth, E. Y. & J. W. G. Lund (eds), Lake Sediments and
Environmental History: Studies in Palaeolimnology and
Palaeoecology in Honour of Winifred Tutin, Chapter. 9.
Leicester University Press, Leicester: 11–67.
Entz, G. & O. Sebestyen, 1946. Das Leben des Balaton-Sees.
Arbeiten des Ungarischen Biologischen Forschungsinsti-
tutes 16: 179–411.
Flower, R. J., 1993. Diatom preservation: experiments and
observations on dissolution and breakage in modern and
fossil material. Hydrobiologia 269/270: 473–484.
Frey, D. G., 1950. The taxonomic and phylogenetic signifi-
cance of the head pores of the Chydoridae (Cladocera).
Internationale Revue der Gesamten Hydrobiologie 44:
27–50.
Frey, D. G., 1962. Cladocera from the Eemian interglacial of
Denmark. Journal of Paleontology 36: 1133–1154.
Frey, D. G., 1988. Littoral and offshore communities of dia-
toms, cladocerans and dipterous larvae, and their inter-
pretation in paleolimnology. Journal of Paleolimnology 1:
179–191.
Frey, D. G., 1991. First subfossil records of Daphnia head-
shields and shells (Anomopoda, Daphniidae) about
10,000 years old from northernmost Greenland, plus
Alona guttata (Chydoridae). Journal of Paleolimnology 6:
193–197.
Gasiorowski, M. & M. Kupryjanowicz, 2009. Lakepeat bog
transformation recorded in the sediments of the Stare
Biele mire (Northeastern Poland). Hydrobiologia 631:
143–154.
Gasse, F., S. Juggins & L. B. Khelifa, 1995. Diatombased
transfer functions for inferring past hydrochemical
characteristics of African lakes. Palaeogeography, Palae-
oclimatology, Palaeoecology 117: 31–54.
Goulden, C. E. & D. G. Frey, 1963. The occurrence and sig-
nificance of lateral head pores in the genus Bosmina
(Cladocera). Internationale Revue der Gesamten Hydro-
biologie 48: 513–522.
G.-Toth, L., 1992. Limiting effect of abioseston on food
ingestion, postembryonic development time and fecundity
of daphnids in Lake Balaton (Hungary). Journal of
Plankton Research 14: 435–446.
Gulyas, P. & L. Forro, 1999. Az agascsapu rakok (Cladocera)
kishatarozoja (A guide for the identification of Cladocera
occurring in Hungary). In Vızi Termeszet- es
Kornyezetvedelem (In Freshwater Nature Conservation
and Environmental Protection), Vol. 9. KGI, Budapest,
2nd edition. (in Hungarian).
Hajnal, E. & J. Padisak, 2008. Analysis of long-term ecological
status of Lake Balaton based on the ALMOBAL phyto-
plankton database. Hydrobiologia 599: 227–237.
Harmsworth, R. V., 1968. The developmental history of Blel-
ham Tarn (England) as shown by animal microfossils,
with special reference to the Cladocera. Ecological
Monographs 38: 223–241.
Heinsalu, A., H. Luup, T. Alliksaar, P. Noges & T. Noges,
2008. Water level changes in a large shallow lake as
reflected by the plankton:peryphyton-ratio of sedimentary
diatoms. Hydrobiologia 599: 23–30.
Heiri, O., A. F. Lotter & G. Lemcke, 2001. Loss on ignition as
a method for estimating organic and carbonate content in
sediments: reproducibility and comparability of results.
Journal of Paleolimnology 25: 101–110.
Hofmann, W., 1998. Cladocerans and chironomids as indica-
tors of lake level changes in north temperate lakes. Journal
of Paleolimnology 19: 55–62.
Hofmann, W., 2001. Late-Glacial/Holocene succession of the
chironomid and cladoceran fauna of the Soppensee
(Central Switzerland). Journal of Paleolimnology 25:
411–420.
Hofmann, W., 2003. The long-term succession of high-altitude
cladoceran assemblages: a 9000-year record from Sagi-
stalsee (Swiss Alps). Journal of Paleolimnology 30: 291–
296.
Houk, V., 2003. Atlas of freshwater centric diatoms with a brief
key and descriptions. Part I. Melosiraceae, Orthoseiraceae,
Paraliaceae and Aulacoseiraceae. In Czech Phycology
Supplement, Vol. 1. Czech Phycological Society, Praha,
Palacky University, Olomouc: 1–27.
Istvanovics, V., A. Osztoics & M. Honti, 2004. Dynamics and
ecological significance of daily internal load of phos-
phorus in shallow Lake Balaton, Hungary. Freshwater
Biology 49: 232–252.
Istvanovics, V., A. Clement, L. Somlyody, A. Specziar, L. G.
Toth & J. Padisak, 2007. Updating water quality targets
for shallow Lake Balaton (Hungary), recovering from
eutrophication. Hydrobiologia 581: 305–318.
Istvanovics, V., M. Honti, A. Kovacs & A. Osztoics, 2008.Distribution of submerged macrophytes along environ-
mental gradients in large, shallow Lake Balaton (Hungary).
Aquatic Botany 88: 317–330.
Jakab, G., P. Sumegi & Z. Szanto, 2005. Kes}o-glacialis
es holocen vızszintingadozasok a Szigligeti-obolben
242 Hydrobiologia (2010) 641:225–244
123
Author's personal copy

(Balaton) makrofosszılia vizsgalatok eredmenyei alapjan–
(Late Glacial and Holocene water level changes in the
Szigliget Bay, Lake Balaton based on macrofossil inves-
tigations). Foldtani Kozlony 135: 404–432. (in Hungarian).
Jeppesen, E., J. P. Jensen, S. Amsinck, F. Landkildehus, T.
Lauridsen & S. F. Mitchell, 2002. Reconstructing the
historical changes in Daphnia mean size and planktivo-
rous fish abundance in lakes from the size of Daphniaephippia in the sediment. Journal of Paleolimnology 27:
133–143.
Juhasz I., P. Sumegi, Zs. Szanto, E. Svingor, Molnar, M. &
G. Jakab, 2007. The Little Balaton region and the Balaton
uplands. In Zatyko, Cs., I. Juhasz & P. Sumegi (eds),
Environmental Archaeology in Transdanubia. Varia
Archaeologica Hungarica 20, Budapest: 27–175.
Kearns, C., N. Hairston & D. Kesler, 1996. Particle transport
by benthic invertebrates: its role in egg bank dynamics.
Hydrobiologia 332: 63–70.
Kenney, W. F., M. N. Waters, C. L. Schelske & M. Brenner,
2002. Sediment records of phosphorus-driven shifts to
phytoplankton dominance in shallow Florida lakes. Jour-
nal of Paleolimnology 27: 367–377.
Kindt, R. & R. Coe, 2005. Tree diversity analysis. A manual
and software for common statistical methods for ecolog-
ical and biodiversity studies. Nairobi (Kenya): World
Agroforestry Centre (ICRAF). ISBN 92-9059-179-X.
Kobayasi, H., M. Idei, S. Mayama, T. Nagumo & K. Osada,
2006. H. Kobayasi’s Atlas of Japanese Diatoms Based on
Electron Microscopy, Vol. 1. Uchida Rokakuho Publishing
Co., Ltd, Tokyo: 1–533.
Koinig, K. A., W. Shotyk, A. F. Lotter, C. Ohlendorf & M.
Sturm, 2003. 9000 years of geochemical evolution of
lithogenic major and trace elements in the sediment of an
alpine lake–the role of climate, vegetation, and land-use
history. Journal of Paleolimnology 30: 307–320.
Korhola, A. & M. Rautio, 2001. Cladocera and other Bran-
chiopod Crustaceans. In Smol, J. P., H. J. B. Birks &
W. M. Last (eds), Tracking Environmental Change Using
Lake Sediments, Vol. 4. Zoological Indicators. Kluwer
Academic Publishers, Dordrecht, The Netherland: 5–41.
Korhola, A., H. Olander & T. Blom, 2000. Cladoceran and
chironomid assemblages as qualitative indicators of water
depth in subarctic Fennoscandian lakes. Journal of
Paleolimnology 24: 43–54.
Korhola, A., M. Tikkanen & J. Weckstrom, 2005. Quantifica-
tion of Holocene lake-level changes in Finnish Lapland
using a cladocera—lake depth transfer model. Journal of
Paleolimnology 34: 175–190.
Koster, D., J. M. J. Racca & R. Pienitz, 2004. Diatom-based
inference models and reconstructions revisited: methods
and transformations. Journal of Paleolimnology 32:
233–246.
Krammer, K., 2000. The genus Pinnularia. In Diatoms of
Europe. Diatoms of the European Inland Waters and
Comparable Habitats, Vol. 1. A.R.G. Gantner Verlag,
K.G. Ruggell: 1–703.
Krammer, K. & H. Lange-Bertalot, 1986–1991. Bacillario-
phyceae. In Susswasserflora von Mitteleuropa., Vol. 1–4.
Gustav Fischer Verlag, Stuttgart, Jena.
Lange-Bertalot, H., 1993. 85 Neue Taxa und uber 100 weitere
neu definierte Taxa erganzend zur Susswasserflora von
Mitteleuropa, Vol. 2/1–4. Bibliotheca Diatomologica
27: 454.
Lange-Bertalot, H., 2001. Diatoms of Europe. Navicula sensustricto 10 Genera separated from Navicula sensu lato
Frustulia. In Lange-Bertalot, H. (ed.), Diatoms of the
European Inland Waters and Comparable Habitats, Vol. 2.
A.R.G. Gantner Verlag, K.G. Ruggell.
Lange-Bertalot, H. & K. Krammer, 1989. Achnanthes eine
Monographie der Gattung. In Lange-Bertalot, H. (ed.),
Bibliotheca Diatomologica, Vol. 18. J. Cramer, Berlin,
Stuttgart: 1–393.
Lange-Bertalot, H. & D. Metzeltin, 1996. Indicators of oli-
gotrophy. 800 taxa representative of three ecologically
distinct lake types. In Lange-Bertalot, H. (ed.), Iconog-
raphia Diatomologica, Annotated Diatom Micrographs,
Vol. 2. Koeltz Scientific Books, Konigstein: 1–90.
Loczy, L., 1916. Die geologischen Formationen der Bala-
tongegend und ihre regionale Tektonik (Erste Sektion).
Physische Geographie des Balatonsees und seiner Umge-
bung (Erster Band). Die Geomorphologie des Balatonsees
und seiner Umgebung (Erster Teil). Resultate der Wis-
senschaftlichen Erforschung des Balatonsees. Balaton-
ausshusse der ung. Geographischen Gesellschaft. Wien.
Medzihradszky, Zs., 2005. Holocene vegetation history and
human activity in the Kis-Balaton area, Western Hungary.
Studia Botanica Hungarica 36: 77–100.
Moss, B., D. Stephen, C. Alvarez, E. Becares, W. Van, S. E. de
Bund, E. Collings, E. Van Donk, T. De Eyto, C. Feld-
mann, M. Fernandez-Alaez, R. J. M. Fernandez-Alaez, F.
Franken, E. M. Garcia-Criado, M. Gross, L. A. Gyllstrom,
K. Hansson, A. Irvine, J. P. Jarvalt, E. Jensen, T. Jeppe-
sen, R. Kairesalo, T. Kornijow, H. Krause, A. Kunnap, E.
Laas, B. Lille, H. Lorens, M. R. Luup, P. Miracle, T.
Noges, M. Noges, I. Nykanen, W. Ott, E. T. H. M.
Peczula, G. Peeters, S. Phillips, V. Romo, J. Russell, M.
Salujoe, K. Scheffer, H. Siewertsen, C. Smal, H. Tesch, L.
Timm, I. Tuvikene, T. Tonno, E. Virro, D. Vicente &
Wilson, 2003. The determination of ecological status in
shallow lakes -a tested system (ECOFRAME) for imple-
mentation of the European Water Framework Directive.
Aquatic Conservation-Marine and Freshwater Ecosystems
13: 507–549.
Newberry, T. L. & C. L. Schelske, 1986. Biogenic silica record
in the sediments of Little Round Lake, Ontario. Hydro-
biologia 143: 293–300.
Oksanen, J., R. Kindt, P. Legendre, B. O’Hara & M. H. H.
Stevens, 2007, vegan: Community Ecology Package. R
package version 1.8-8.
Padisak, J. & C. S. Reynolds, 2003. Shallow lakes: the abso-
lute, the relative, the functional and the pragmatic.
Hydrobiologia 506–509: 1–11.
Ponyi, J. E., 1971. Investigation on crustacean and molluscan
remains in the upper sedimentary layer of Lake Balaton.
Annales Instituti Biologici (Tihany) Hungaricae Acade-
mie Scientarium 38: 183–197.
R Development Core Team, 2008. R: A Language and Envi-
ronment for Statistical Computing. R Foundation for Sta-
tistical Computing, Vienna, Austria. ISBN 3-900051-07-0.
Reed, J. M., 1998. Diatom preservation in the recent sediment
record of Spanish saline lakes: implications for palaeo-
climate study. Journal of Paleolimnology 19: 129–137.
Hydrobiologia (2010) 641:225–244 243
123
Author's personal copy

Rooney, N., J. Kalff & C. Habel, 2003. The role of submerged
macrophyte beds in phosphorus and sediment accumula-
tion in Lake Memphremagog, Quebec, Canada. Limnol-
ogy and Oceanography 48: 1927–1937.
Sagi, K., 1968. A Balaton vızallastendenciai 1863-i a tortenet
es kartografiai adatok tukreben (Water level trends of
Lake Balaton before 1863, as revealed by historical and
cartographical data. Veszprem Megyei Muzeumok
Kozlemenyei 7: 441–468. (in Hungarian).
Sarmaja-Korjonen, K., 2001. Correlation of fluctuations in
cladoceran planktonic: littoral ratio between three cores
from a small lake in southern Finland: Holocene water-
level changes. Holocene 11: 53–63.
Sarmaja-Korjonen, K. & P. Alhonen, 1999. Cladoceran and
diatom evidence of lake-level fluctuations from a Finnish
lake and the effect of aquatic-moss layers on microfossil
assemblages. Journal of Paleolimnology 22: 277–290.
Scheffer, M., 1998. Ecology of Shallow Lakes. Chapman and
Hall, London.
Sebestyen, O., 1969a. Kladocera tanulmanyok a Balatonon III.
Totorteneti el}otanulmanyok I—Cladocera studies in Lake
Balaton III. Preliminary studies for lake history investi-
gations. Annales Instituti Biologici (Tihany) Hungaricae
Academie Scientarium 36: 229–256. (in Hungarian with
English abstract).
Sebestyen, O., 1969b. Kladocera tanulmanyok a Balatonon IV.
Negyedkori maradvanyok a Balaton uledekeben I—
Cladocera studies in Lake Balaton IV. Quaternary remains
in the sediment of Lake Balaton I. Annales Instituti Bio-
logici (Tihany) Hungaricae Academie Scientarium 36:
229–256. (in Hungarian with English abstract).
Sebestyen, O., 1970. Kladocera tanulmanyok a Balatonon IV.
Negyedkori maradvanyok a Balaton uledekeben II—
Cladocera studies in Lake Balaton IV. Quaternary remains
in the sediment of Lake Balaton II. Annales Instituti Bi-
ologici (Tihany) Hungaricae Academie Scientarium 37:
247–279. (in Hungarian with English abstract).
Sebestyen, O., 1971. Kladocera tanulmanyok a Balatonon IV.
Negyedkori maradvanyok a Balaton uledekeben III—
Cladocera studies in Lake Balaton IV. Quaternary remains
in the sediment of Lake Balaton III. Annales Instituti
Biologici (Tihany) Hungaricae Academie Scientarium 38:
227–268. (in Hungarian with English abstract).
Serlegi, G., 2007. A balatonkereszturi ‘‘vızmerce’’:
Kornyezetregeszeti informaciok a Balaton deli partjanak
romai kori t}ortenetehez (The ‘‘stream gauge’’ of Bala-
tonkeresztur. Environmental archaeological data on the
Roman Period history of the southern shore of Lake Bal-
aton), In Bıro, Sz. (ed.), FiRKaK I. Fiatal Romai Koros
Kutatok I. konferncia kotete (First conference book of
young scientist of Roman Periods): 297–317. (in
Hungarian).
Siver, P. A., 1999. Development of paleolimnological inference
models for pH, total nitrogen and specific conductivity
based on planktonic diatoms. Journal of Paleolimnology 21:
45–60.
Smol, J. A., 2008. Pollution of lakes and rivers. A paleoenviron-
mental perspective, 2nd ed. Blackwell Publishing, Malden.
Smol, J. & M. M. Boucherle, 1985. Postglacial changes in algal
and cladocerans assemblages in Little Round Lake,
Ontario. Hydrobiologia 103: 25–49.
SPSS Inc., 1998. SPSS for Windows Rel. 8.0. SPSS Inc.,
Chicago.
Stuiver, M., P. J. Reimer & R. W. Reimer, 2005. CALIB 5.0.
http://www.calib.qub.ac.uk/crev50/.
Sumegi, P., E., Bodor, I. Juhasz, Z. Hunyadfalvi, K. Herbich,
S. Molnar & G. Timar, 2007. A Balaton deli partjan feltart
regeszeti lel}ohelyek kornyezettorteneti feldolgozasa
(Paleoenvironmental study of archeological sites in the
south shore of the lake Balaton). In Belenyesy K., Sz.
Honti & V. Kiss (eds), Rolling Time. Excavations on the
M7 Motorway on County Somogy Between Zamardi and
Ordacsehi. Somogy megyei Muzeumok Igazgatosaga—
MTA Regeszeti Intezete: 241–253. (in Hungarian).
Tatrai, I., K. Matyas, J. Korponai, G. Paulovits & P. Pomogyi,
2000. The role of Kis-Balaton Water protection system in
the control of water quality of Lake Balaton. Ecological
Engineering 16: 73–78.
Tatrai, I., V. Istvanovics, L. G. Toth & I. Kobor, 2008. Man-
agement measures and long-term water quality changes in
Lake Balaton (Hungary). Fundamental and Applied
Limnology (Archive fur Hydrobiologie) 172: 1–11.
Troel-Smith, J., 1955. Karakterisering af løse jordarter (Char-
acterisation of unconsolidated sediments). Danmarks
geologiske undersrgelse, Raekke 4: 1–73.
Tullner, T. & T. Cserny, 2003. New aspects of lake-level
changes: Lake Balaton, Hungary. Acta Geologica
Hungarica 46: 215–238.
Valiranta, M., S. Kultti, M. Nyman & K. Sarmaja-Korjonen,
2005. Holocene development of aquatic vegetation in
shallow Lake Njargajavri, Finnish Lapland, with evidence
of water-level fluctuations and drying. Journal of Paleo-
limnology 34: 203–215.
Virag, A., 1998. A Balaton multja es jelene (The Past and the
Present of Lake Balaton). Egri Nyomda Kft. (in Hungarian).
Virag, A., 2005. History of River Sio and Lake Balaton (1055–
2005). Kozlekedesi Dokumentacios Kft. (in Hungarian).
Wessels, M., K. Mohaupt, R. Kommerlin & A. Lenhard, 1999.
Reconstructing past eutrophication trends from diatoms
and biogenic silica in the sediment and the pelagic zone of
Lake Constance, Germany. Journal of Paleolimnology 21:
171–192.
Whiteside, M. C., J. B. Williams & C. P. White, 1978. Sea-
sonal Abundance and Pattern of Chydorid, Cladocera in
Mud and Vegetative Habitats. Ecology 59: 1177–1188.
Whitmore, T. J., M. Brenner & C. L. Schelske, 1996. Highly
variable sediment distribution in shallow, wind-stressed
lakes: a case for sediment-mapping surveys in paleo-
limnological studies. Journal of Paleolimnology 15: 207–
221.
Zolyomi, B., 1952. History of evolution of the vegetation coverin Hungary from the last ice age (in Hungarian). MTA
Biologiai Osztalyanak Kozlemenyei 1: 491–543.
Zolyomi, B., 1995. Opportunities for pollen stratigraphic
analysis of shallow lake sediments: the example of Lake
Balaton. GeoJournal 36: 237–241.
244 Hydrobiologia (2010) 641:225–244
123
Author's personal copy
Related Documents











