Dynamics and ecological significance of daily internal load of phosphorus in shallow Lake Balaton, Hungary VERA ISTVA ´ NOVICS,* ANDRA ´ S OSZTOICS † AND MA ´ RK HONTI* *Water Resources Management Group of the Hungarian Academy of Sciences, Department of Sanitary and Environmental Engineering, Budapest University of Technology and Economics, Budapest, Hungary † Department of Civil and Environmental Engineering, Budapest University of Technology and Economy, Budapest, Hungary SUMMARY 1. As supported by field data, turbidity recorded by light scattering sensors could reliably be converted into concentration of suspended particulate matter (SPM) and coefficient of vertical light attenuation (K d ) in Lake Balaton. 2. Autocorrelation analysis revealed that proper determination of SPM concentration and K d required daily sampling. To approximate daily rate of resuspension, 15 min or more frequent measurements were needed. Thus, routine monitoring provides very little insight into environmental variability of shallow lakes as habitats for phytoplankton. 3. The internal P load was estimated from daily rate of resuspension and P desorption capacity of sediments. The latter was assumed to be proportionate to the potentially mobile inorganic P content of SPM. A comparison with net primary production and nutrient status of phytoplankton showed that the proposed method of estimating time series of internal P load captured seasonal trends. 4. The daily rate of resuspension was high whereas that of internal P load was low in Lake Balaton relative to other shallow lakes. The latter reflects favourable behaviour of the calcite-rich sediments. As a consequence, carrying capacity of Basin 1 of Lake Balaton was P-determined. 5. The timing of external and internal loads was radically different. While the former showed mostly seasonal changes, large pulses characterised the latter. As a consequence, internal load may supply more P to phytoplankton growth during the critical summer months than external load. However, the relative importance of these sources may show substantial interannual variability. 6. Large resuspension events often followed each other during periods of 10–15 days. It has been shown that disturbances in this frequency range are of key importance in maintaining the diversity of phytoplankton. We propose that resuspension can be perceived not only as a disturbance factor but also as a factor that periodically relaxes nutrient stress. The former feature may dominate the instantaneous effect, whereas the latter may determine the persistent effect of resuspension on succession of phytoplankton. Keywords: light attenuation, nutrient status of algae, P desorption, pulses in P load, resuspension, turbidity Introduction In most shallow lakes, internal P load is the principal component of P supply for phytoplankton growth during summer. When the carrying capacity of a lake is P-determined (Reynolds, 1992), internal load is a key factor in regulating year-to-year variability of Correspondence: Vera Istva ´novics, Department of Sanitary and Environmental Engineering, Budapest University of Technology and Economics, H-1111 Budapest, Mu ˆ egyetem rkp. 3., Hungary. E-mail: [email protected] Freshwater Biology (2004) 49, 232–252 232 Ó 2004 Blackwell Publishing Ltd

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Dynamics and ecological significance of daily internalload of phosphorus in shallow Lake Balaton, Hungary
VERA ISTVANOVICS,* ANDRAS OSZTOICS † AND MARK HONTI*
*Water Resources Management Group of the Hungarian Academy of Sciences, Department of Sanitary and Environmental
Engineering, Budapest University of Technology and Economics, Budapest, Hungary†Department of Civil and Environmental Engineering, Budapest University of Technology and Economy, Budapest, Hungary
SUMMARY
1. As supported by field data, turbidity recorded by light scattering sensors could reliably
be converted into concentration of suspended particulate matter (SPM) and coefficient of
vertical light attenuation (Kd) in Lake Balaton.
2. Autocorrelation analysis revealed that proper determination of SPM concentration and
Kd required daily sampling. To approximate daily rate of resuspension, 15 min or more
frequent measurements were needed. Thus, routine monitoring provides very little insight
into environmental variability of shallow lakes as habitats for phytoplankton.
3. The internal P load was estimated from daily rate of resuspension and P desorption
capacity of sediments. The latter was assumed to be proportionate to the potentially mobile
inorganic P content of SPM. A comparison with net primary production and nutrient
status of phytoplankton showed that the proposed method of estimating time series of
internal P load captured seasonal trends.
4. The daily rate of resuspension was high whereas that of internal P load was low in Lake
Balaton relative to other shallow lakes. The latter reflects favourable behaviour of the
calcite-rich sediments. As a consequence, carrying capacity of Basin 1 of Lake Balaton was
P-determined.
5. The timing of external and internal loads was radically different. While the former
showed mostly seasonal changes, large pulses characterised the latter. As a consequence,
internal load may supply more P to phytoplankton growth during the critical summer
months than external load. However, the relative importance of these sources may show
substantial interannual variability.
6. Large resuspension events often followed each other during periods of 10–15 days.
It has been shown that disturbances in this frequency range are of key importance in
maintaining the diversity of phytoplankton. We propose that resuspension can be
perceived not only as a disturbance factor but also as a factor that periodically relaxes
nutrient stress. The former feature may dominate the instantaneous effect, whereas the
latter may determine the persistent effect of resuspension on succession of phytoplankton.
Keywords: light attenuation, nutrient status of algae, P desorption, pulses in P load, resuspension,turbidity
Introduction
In most shallow lakes, internal P load is the principal
component of P supply for phytoplankton growth
during summer. When the carrying capacity of a lake
is P-determined (Reynolds, 1992), internal load is a
key factor in regulating year-to-year variability of
Correspondence: Vera Istvanovics, Department of Sanitary and
Environmental Engineering, Budapest University of Technology
and Economics, H-1111 Budapest, Muegyetem rkp. 3., Hungary.
E-mail: [email protected]
Freshwater Biology (2004) 49, 232–252
232 � 2004 Blackwell Publishing Ltd

algal biomass (van der Molen & Boers, 1994; Istva-
novics & Somlyody, 2001). At the same time, internal
P load has been recognised to be a major cause of the
resistance of shallow lakes to recover from eutrophi-
cation (Sas, 1989). Thus, internal P load is of prime
importance from the viewpoint of both phytoplankton
ecology and eutrophication management. An order-
of-magnitude or a time-integrated (annual, seasonal,
monthly) estimate of net internal load normally
suffices for managerial purposes and the majority of
studies on internal P load satisfy this need (Lijklema
et al., 1986; Sas, 1989; Kozerski & Kleeberg, 1998;
Søndergaard, Jensen & Jeppesen, 1999; Istvanovics &
Somlyody, 2001). Evaluation of the role of internal P
load in algal dynamics, however, requires temporal
resolution that matches specific growth rates of
phytoplankton. Although numerous direct and indir-
ect approaches have been developed for estimating
internal P load in shallow lakes, none of these
methods allow the appropriate time scale over
prolonged periods of time to be achieved.
As Bostrom, Jansson & Forsberg (1982) have
pointed out, internal P load is a function of various
mobilisation and transport processes. The relative
importance of different P mobilisation mechanisms
differs among lakes in accordance with the geochem-
ical composition of their sediments (Bostrom, 1984).
Wind induced sediment resuspension is the most
important transport process in wind-exposed shallow
lakes (Lijklema et al., 1986; Søndergaard, Kristensen
& Jeppesen, 1992; Evans, 1994; Ogilvie & Mitchell,
1998). Mobilisation and transport of sedimentary P
can be characterised by different temporal scales.
While mobilisation results in relatively smooth
changes over several days to a few months, sediment
resuspension may fluctuate widely over hours.
Considering fast kinetics of P desorption from
suspended sediments with initial rate constants of
<1 min)1 (Williams, Syers & Harris, 1970; Li et al.,
1972; Kuo & Lotse, 1974), the internal load via P
desorption must closely follow the dynamics of
sediment resuspension. Consequently, if the aim is
to relate algal growth and/or phytoplankton succes-
sion to P supply originating mostly from P de-
sorption, conventional studies on P mobilisation and
P release from sediments are less relevant than
studies on the dynamics of sediment resuspension.
Resuspension has manifold and contrasting influ-
ences on algal dynamics. Reduced light availability
has been shown to retard algal growth (Somlyody &
Koncsos, 1991). Simultaneously, however, transport
of meroplanktonic/benthic species may temporarily
increase the biomass of phytoplankton (Padisak,
G.-Toth & Rajczy, 1988, 1990). Besides instantaneous
effects, increased nutrient availability upon resuspen-
sion may stimulate algal growth (Rijkeboer, De Bles
& Gons, 1991; Ogilvie & Mitchell, 1998). In a number
of lakes, large resuspension events are followed by a
measurable change in the concentration of soluble
reactive P (SRP) (Søndergaard et al., 1992; Hamilton
& Mitchell, 1997; Ogilvie & Mitchell, 1998). Depend-
ing on the equilibrium concentration between sedi-
ment and water, this change may either be negative
or positive. In highly calcareous Lake Balaton,
resuspension fails to detectably increase typically
low concentrations (<5 mg P m)3) of SRP. The P
uptake experiments, however, have suggested that P
deficient phytoplankton in the lake could not main-
tain net P uptake for longer than a few tens of
minutes in the absence of a desorptive P flux from
resuspended sediments (Istvanovics & Herodek,
1995).
In the present study, we examined sediment
resuspension in Lake Balaton by high frequency
recording of turbidity during two vegetation periods.
Using additional information about phosphorus
desorption capacity of the sediments, we estimated
the internal P load and related internal P supply to the
nutrient status of phytoplankton.
Methods
Sampling site
Lake Balaton is a large (596 km2), shallow
(zmean ¼ 3.2 m), elongated lake that can be divided
into four basins (Fig. 1). The largest tributary, the Zala
River (mean flow at the mouth section is 7.8 m)3 s)1)
enters the smallest (38 km2) and shallowest
(zmean ¼ 2.3 m) Basin 1. The only outflow connects
Basin 4 with the Danube. Basin 1 has become
hypertrophic during the 1970s (Herodek, 1986) and
recovered surprisingly quickly after a series of man-
agement measures taken from the mid-1980s (Istva-
novics & Somlyody, 2001).
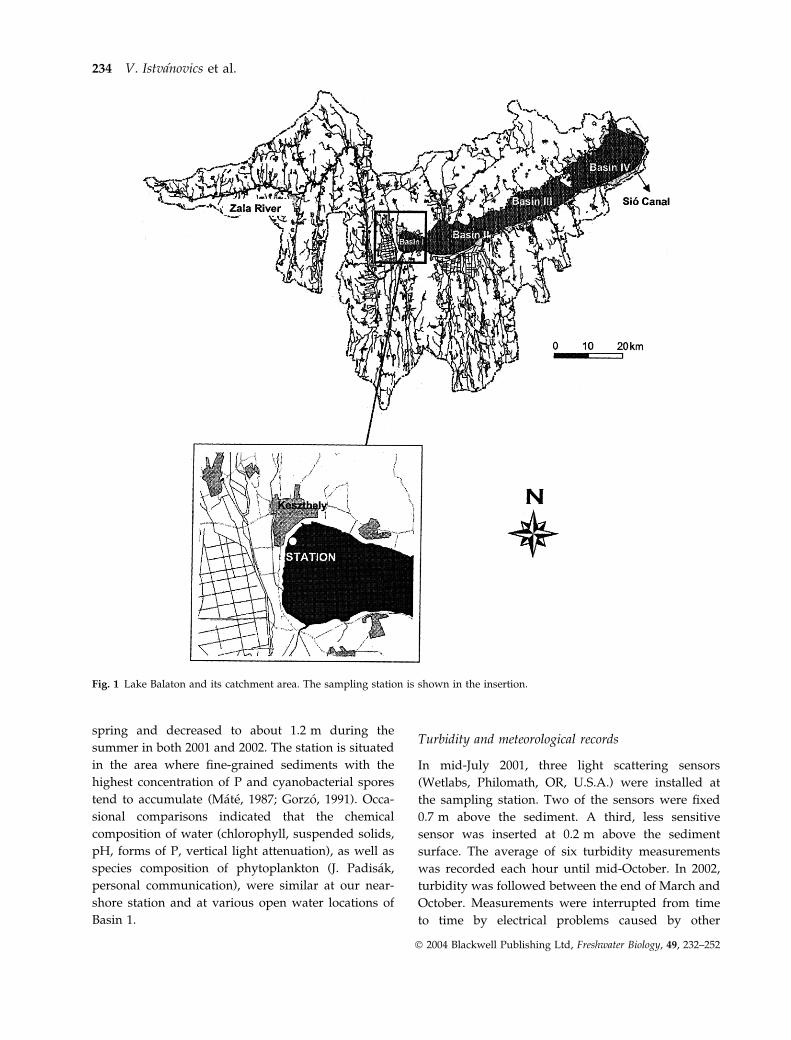
We built a permanent sampling station 100 m
offshore at the outskirts of the town of Keszthely
(Fig. 1). The water depth was about 1.5 m in early
Dynamics of internal P load 233
� 2004 Blackwell Publishing Ltd, Freshwater Biology, 49, 232–252
spring and decreased to about 1.2 m during the
summer in both 2001 and 2002. The station is situated
in the area where fine-grained sediments with the
highest concentration of P and cyanobacterial spores
tend to accumulate (Mate, 1987; Gorzo, 1991). Occa-
sional comparisons indicated that the chemical
composition of water (chlorophyll, suspended solids,
pH, forms of P, vertical light attenuation), as well as
species composition of phytoplankton (J. Padisak,
personal communication), were similar at our near-
shore station and at various open water locations of
Basin 1.
Turbidity and meteorological records
In mid-July 2001, three light scattering sensors
(Wetlabs, Philomath, OR, U.S.A.) were installed at
the sampling station. Two of the sensors were fixed
0.7 m above the sediment. A third, less sensitive
sensor was inserted at 0.2 m above the sediment
surface. The average of six turbidity measurements
was recorded each hour until mid-October. In 2002,
turbidity was followed between the end of March and
October. Measurements were interrupted from time
to time by electrical problems caused by other
Fig. 1 Lake Balaton and its catchment area. The sampling station is shown in the insertion.
234 V. Istvanovics et al.
� 2004 Blackwell Publishing Ltd, Freshwater Biology, 49, 232–252

instruments operated at the station. Until mid-April,
sensors were installed at about 0.1, 0.7 and 1 m above
the sediment. Thereafter five sensors were applied at
about 0.05, 0.1, 0.4, 0.7 and 0.9 m above the sediment.
The least sensitive sensor was always the one closest
to the sediment surface. Turbidity was recorded every
15 min. The sensors were cleaned with ethanol every
3–4 days, a procedure that prevented biofilm forma-
tion on the measuring surface. When the measuring
window of a sensor was found to be overgrown, data
from between two cleanings were excluded.
The maximum output signal of each sensor was
5 V. This corresponded to a maximum turbidity of
150–170 NTU in the case of the sensitive sensors and
about 500 NTU in the case of the low sensitivity
sensor. Sensors were calibrated against the concen-
tration of suspended particulate matter (SPM) and the
coefficient of downward vertical light attenuation
(Kd, m)1) in the laboratory (see below).
Turbidity data were collected using a Keithley
(USA) multiplexer programmed by TestPoint (Capital
Equipment Corporation, Billerica, MA, U.S.A.). From
September 2002, the system could also be checked and
controlled remotely.
In order to calibrate light scattering sensors, 20
sediment cores were taken in a 100 m2 area around
the sampling station. The upper 1 cm layer of the
cores was pooled into a single sample. During the first
calibration trial, small amounts of known weight of
sediment were added incrementally to 5.3 L of What-
man GF/F filtered lake water in a large, black
container. The suspension was continuously stirred
with a magnetic stirrer. The water content of the
original sediment was known, and the concentration
of SPM calculated in the suspension. Light scattering
was measured after each addition at a low surface
light intensity. Photosynthetically active radiation
(PAR, lmol quanta m)2 s)1) was detected with a 4punderwater quantum sensor (Li-Cor, Lincoln, NE,
U.S.A.) at two fixed depths. During the latter meas-
urement, incident surface radiation was 180 lmol -
quanta m)2 s)1. During the second calibration trial,
the slowly sedimenting fraction of the sediment was
collected by suspending about 100 mL of the bulk
sediment in 500 mL Whatman GF/F filtered lake
water in a measuring cylinder. Water was siphoned
off carefully from above the settled material after
10 min. The sedimented material was resuspended
again in filtered water and this procedure was
repeated until an insignificant amount of sediment
remained in suspension. To concentrate slowly sedi-
menting fraction (vsed < 12–14 m day)1), water with
the resuspended matter from all successive resuspen-
sions was centrifuged at 1500 g for 20 min. Calibra-
tion was performed as during the first trial, except
that aliquots of the concentrated fine sediment with
known content of dry matter were pipetted into the
measuring vessel.
Continuous turbidity records were converted into
SPM concentration using the regression obtained
during the second calibration trial. The SPM time
series was smoothed by calculating time weighted 7
point moving averages. Weights were taken from the
autocorrelation function of SPM (see Results), and the
SPM content of the water column per unit surface area
was computed. Because of the lack of appropriate
information, we neglected changes in water depth.
Resuspension was estimated as an increase in the SPM
content of the water column between two subsequent
records. Daily rates of resuspension (Rd, g m)2 day)1)
were summed from the individual resuspension
events. The P desorption from resuspended sediments
was calculated using the assumptions that (i) rate of P
desorption from the resuspended sediments was a
linear function of the sum of the NH4Cl-RP and NaOH-
RP fractions of SPM and (ii) time-averaged concentra-
tion of these fractions corresponded to a desorption of
5 lg P g)1 dry mass (cf. Lijklema et al., 1986). The
fractional composition of P in SPM was determined
according to Hieltjes & Lijklema (1980; see below).
A small meteorological station was set up at the
sampling station by the HWI Elektronika Company,
Hungary. Global radiation, wind speed, wind direc-
tion and water temperature at three to five depths
were measured every 10 s. Mean values were saved at
15-min intervals on a data logger.
Manual sampling programme
Manual sampling started in mid-March and contin-
ued until mid-October.
Photosynthetically active radiation was measured
weekly with a 4p underwater quantum sensor in the
air and at 10 cm depth intervals in the water between
07:00 and 08:00 hours. Simultaneously, water samples
were taken from the middle of the water column with
a Meyer flask. Processing of samples started immedi-
ately after transportation to the shoreline laboratory of
Dynamics of internal P load 235
� 2004 Blackwell Publishing Ltd, Freshwater Biology, 49, 232–252

the West Transdanubian Water Authority (less than
half an hour). Unless indicated otherwise, Whatman
GF/F glass fibre filters (Whatman, Maidstone, U.K.)
were used for filtration.
The concentration of SPM was determined gravimet-
rically after filtering an appropriate volume of water
(200–800 mL) onto dried, preweighed filters (105 �C,
24 h). Organic carbon content of SPM was approxima-
ted by measuring total C content after fuming the filters
with concentrated HCl for 10 min. The C concentration
was determined with a CHN autoanalyser.
Suspended particulate matter was concentrated for
sequential extraction of P from 800 to 2000 mL of lake
water by centrifugation at 1500 g for 20 min. Loss of
SPM (usually <5%) was determined gravimetrically
by filtering the supernatant onto preweighed filters.
Visual inspection showed that the loss could primar-
ily be attributed to poorly sedimenting cyanobacteria.
Fractional distribution of P was determined in
duplicates of the concentrated SPM sample according
to the sequential extraction scheme of Hieltjes &
Lijklema (1980). Total P content of the NaOH extract
was measured according to the method of Menzel &
Corwin (1965). When calculating the P concentration
of various fractions (lg P g)1 dry mass), we corrected
for the loss of SPM during centrifugation. Fractional
composition of P was determined in triplicates from
the uppermost 1 cm sediment layer collected monthly
in the same manner as described above. In 2002, the
fine-grained, slowly sedimenting fraction of this layer
was also extracted sequentially.
The concentration of chlorophyll a was measured
spectrophotometrically after extracting the filters in
90% cold acetone in the dark and correcting for
phaeopigments by acidification (Lorenzen, 1967).
Forms of P were measured in triplicate. Concentration
of SRP was determined according to Murphy & Riley
(1962) using 10 cm vials. Total P and total dissolved P
were analysed according to Menzel & Corwin (1965).
Surplus P (SP) content of the phytoplankton was
estimated by filtering 10–30 mL of water onto pre-
washed cellulose acetate membrane filters of 0.2 lm
pore diameter (Whatman) and extracting the filters in
boiling distilled water for 1 h (Fitzgerald & Nelsson,
1966). Concentrations of ammonium, nitrite and
nitrate were measured according to standard methods
(FBA, 1978). Total N and total dissolved N were
determined in triplicate by second order derivative
spectroscopy after oxidation to nitrate with alkaline
persulfate solution (Crumpton, Isenhart & Mitchell,
1992). Concentrations of total inorganic carbon, Ca2+
and Mg2+ were obtained by titration.
Light dependence of steady-state photosynthesis (PI
curve) was determined biweekly using the 14C method
of Lewis & Smith (1983). Two MBq of NaH14CO3
(Amersham, Little Chalfont, U.K.) were added to
300 mL of lake water at a PAR of about 0.2 lmol
quanta m)2 s)1. The sample was distributed in 5 mL
portions into 43 original glass scintillation vials
(Wheaton, Millville, NJ, U.S.A.), and these were
incubated for 30 min at photon flux density between
0.6 and 700 lmol quanta m)2 s)1 in a photosynthetron
that was thermostated at the ambient water tempera-
ture (±1.5 �C). Five additional background samples
were prepared by immediately poisoning 5 mL aliqu-
ots with 0.1 mL of buffered formalin. Photosynthesis
was stopped by poisoning the samples. Unused 14C
was removed by vigorously shaking the samples for
2 h with 0.25 mL of 6 NN HCl. Following neutralisation,
10 mL of Optiphase Highsafe scintillator (Wallac,
Boston, MA, U.S.A.) was added. Five total count
samples were prepared by adding 0.1 mL of labelled
water to a mixture of 5 mL unlabelled lake water and
10 mL scintillator that contained 0.2 mL of phenetil-
amin. Radioactivity was detected in a RackBetaII
liquid scintillation counter for 10 min. Efficiency of
counting was determined by the channel ratio method.
Parameters of the PI curves were obtained by non-
linear fitting of the Jassby & Platt (1976) model to the
data:
PBðIÞ ¼ PBmax � tanh aB � I=PB
max
� �� RB
where PB and PBmax [lg C (lg chl a))1 h)1] are the
actual and the maximum rates of biomass-specific
photosynthesis, respectively; I (lmol quanta m)2 s)1)
is the photon flux density; aB [lg C (lg chl a))1 h)1
(lmol quanta m)2 s)1))1] is the slope of the initial
linear portion of the PI curve and RB [lg C (lg chl
a))1 h)1] is the biomass specific rate of dark respir-
ation assumed to be independent of light conditions.
In order to estimate area-specific net primary
production, turbidity was converted into Kd based
on the regression obtained during the second calibra-
tion trial. Incident PAR was calculated as 47% of
global radiation with the assumption that 1 mol
photon in the visible range was equivalent to 218 kJ
(Reynolds, 1997). From the ratio of PAR measured in
236 V. Istvanovics et al.
� 2004 Blackwell Publishing Ltd, Freshwater Biology, 49, 232–252
the air and immediately below the water surface, we
assumed a mean albedo of 25% that could be due to
wave action. Photon flux density was calculated every
15 min at depth intervals of 10 cm. PBmax and concen-
tration of chlorophyll a were changed half way
between two measurements, whereas aB was estima-
ted from the regression obtained between this value
and Kd. Respiration was assumed to equal 2% of
biomass specific maximal photosynthesis (cf. Falkow-
ski & Raven, 1997; Reynolds, 1997).
Daily external load data from the Zala River were
obtained from the West Transdanubian Water
Authority. This component represents 90–95% of
total nutrient load to Basin 1 of Lake Balaton (Istva-
novics, Somlyody & Clement, 2002).
Results
Sediment resuspension and internal P load
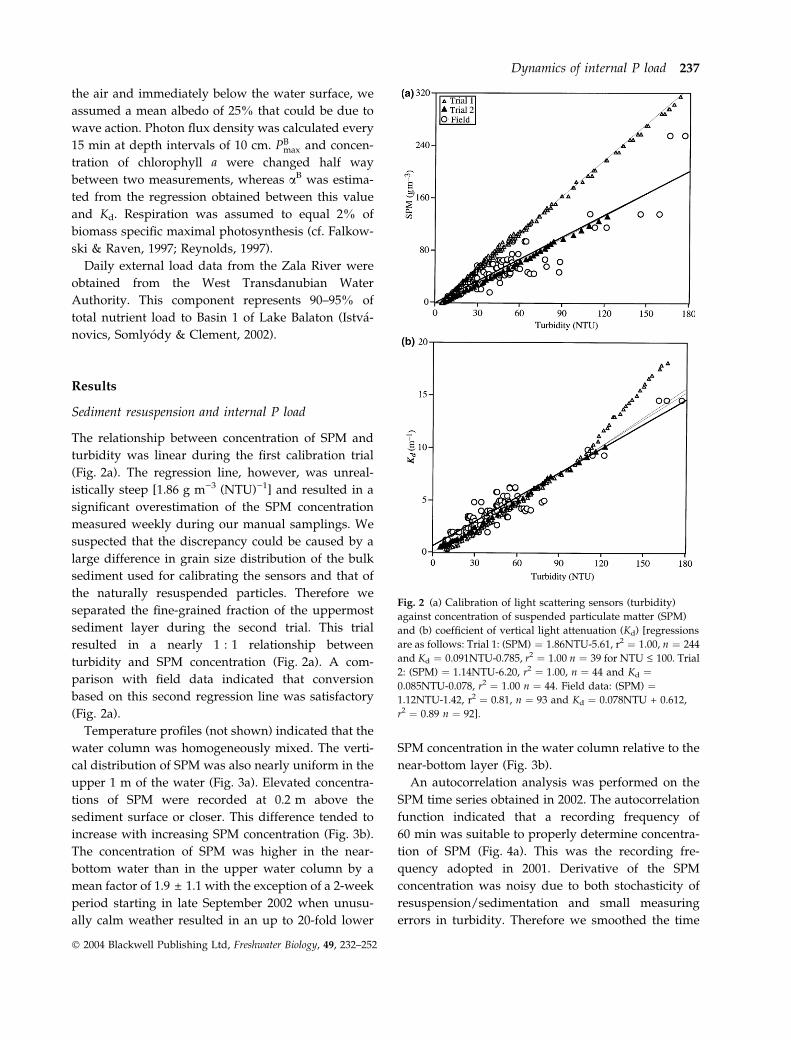
The relationship between concentration of SPM and
turbidity was linear during the first calibration trial
(Fig. 2a). The regression line, however, was unreal-
istically steep [1.86 g m)3 (NTU))1] and resulted in a
significant overestimation of the SPM concentration
measured weekly during our manual samplings. We
suspected that the discrepancy could be caused by a
large difference in grain size distribution of the bulk
sediment used for calibrating the sensors and that of
the naturally resuspended particles. Therefore we
separated the fine-grained fraction of the uppermost
sediment layer during the second trial. This trial
resulted in a nearly 1 : 1 relationship between
turbidity and SPM concentration (Fig. 2a). A com-
parison with field data indicated that conversion
based on this second regression line was satisfactory
(Fig. 2a).
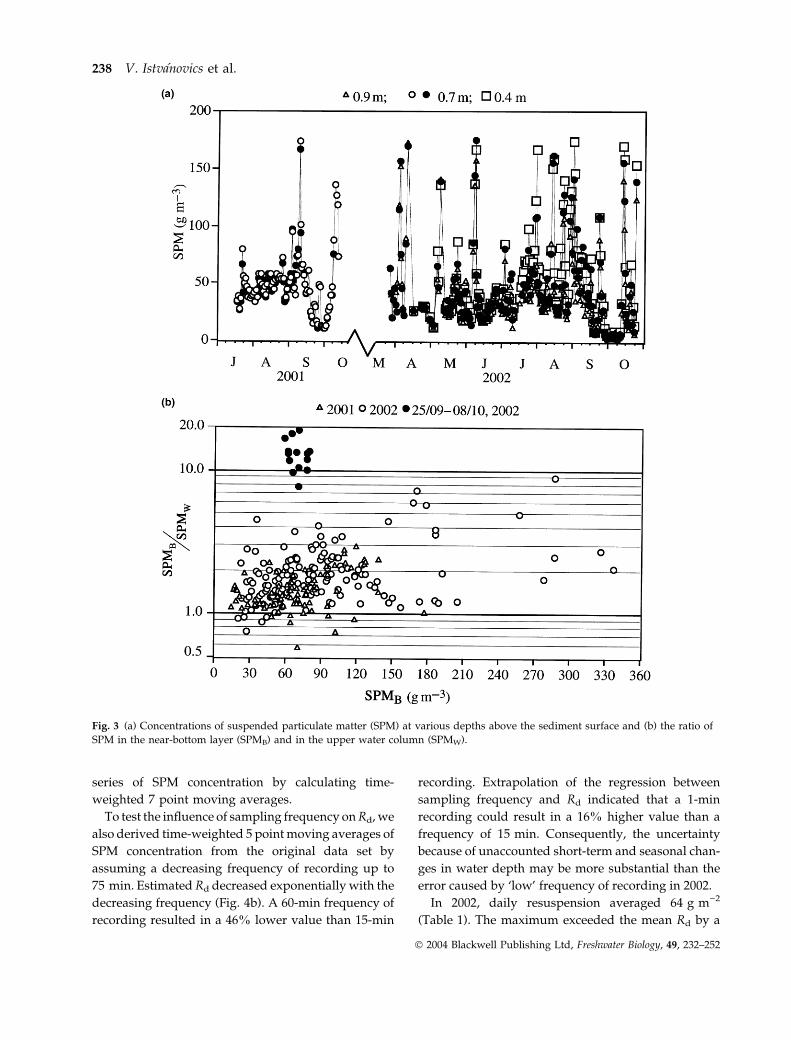
Temperature profiles (not shown) indicated that the
water column was homogeneously mixed. The verti-
cal distribution of SPM was also nearly uniform in the
upper 1 m of the water (Fig. 3a). Elevated concentra-
tions of SPM were recorded at 0.2 m above the
sediment surface or closer. This difference tended to
increase with increasing SPM concentration (Fig. 3b).
The concentration of SPM was higher in the near-
bottom water than in the upper water column by a
mean factor of 1.9 ± 1.1 with the exception of a 2-week
period starting in late September 2002 when unusu-
ally calm weather resulted in an up to 20-fold lower
SPM concentration in the water column relative to the
near-bottom layer (Fig. 3b).
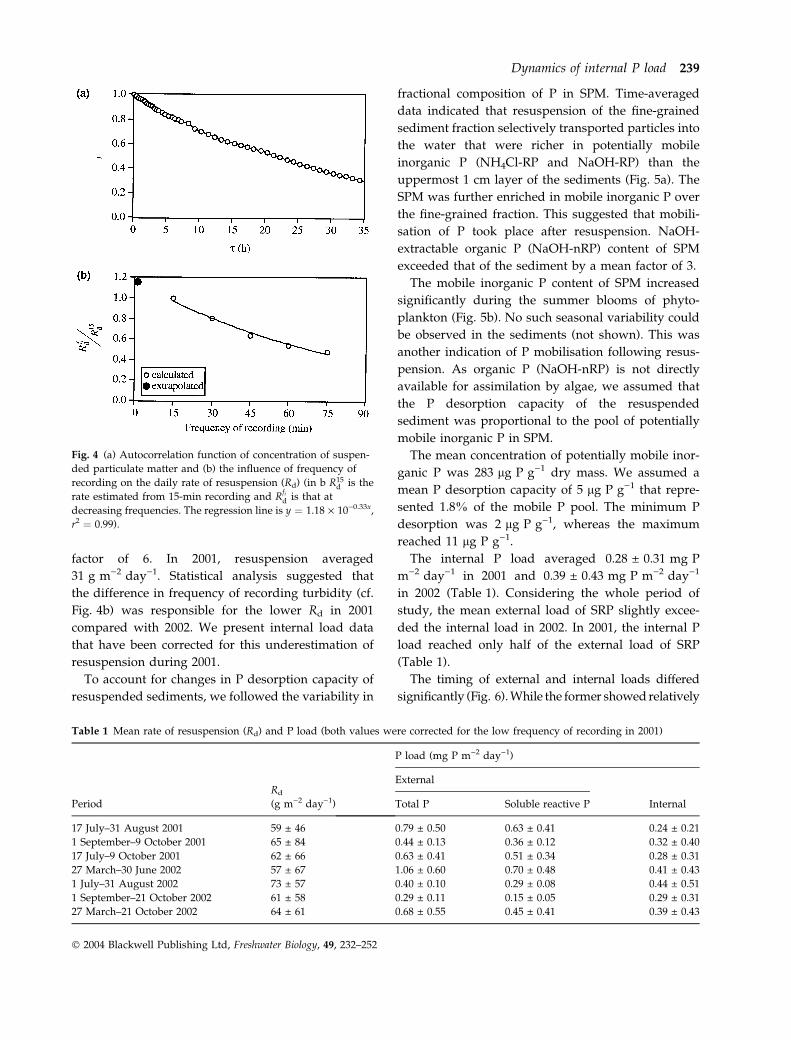
An autocorrelation analysis was performed on the
SPM time series obtained in 2002. The autocorrelation
function indicated that a recording frequency of
60 min was suitable to properly determine concentra-
tion of SPM (Fig. 4a). This was the recording fre-
quency adopted in 2001. Derivative of the SPM
concentration was noisy due to both stochasticity of
resuspension/sedimentation and small measuring
errors in turbidity. Therefore we smoothed the time
Fig. 2 (a) Calibration of light scattering sensors (turbidity)
against concentration of suspended particulate matter (SPM)
and (b) coefficient of vertical light attenuation (Kd) [regressions
are as follows: Trial 1: (SPM) ¼ 1.86NTU-5.61, r2 ¼ 1.00, n ¼ 244
and Kd ¼ 0.091NTU-0.785, r2 ¼ 1.00 n ¼ 39 for NTU £ 100. Trial
2: (SPM) ¼ 1.14NTU-6.20, r2 ¼ 1.00, n ¼ 44 and Kd ¼0.085NTU-0.078, r2 ¼ 1.00 n ¼ 44. Field data: (SPM) ¼1.12NTU-1.42, r2 ¼ 0.81, n ¼ 93 and Kd ¼ 0.078NTU + 0.612,
r2 ¼ 0.89 n ¼ 92].
Dynamics of internal P load 237
� 2004 Blackwell Publishing Ltd, Freshwater Biology, 49, 232–252
series of SPM concentration by calculating time-
weighted 7 point moving averages.
To test the influence of sampling frequency on Rd, we
also derived time-weighted 5 point moving averages of
SPM concentration from the original data set by
assuming a decreasing frequency of recording up to
75 min. Estimated Rd decreased exponentially with the
decreasing frequency (Fig. 4b). A 60-min frequency of
recording resulted in a 46% lower value than 15-min
recording. Extrapolation of the regression between
sampling frequency and Rd indicated that a 1-min
recording could result in a 16% higher value than a
frequency of 15 min. Consequently, the uncertainty
because of unaccounted short-term and seasonal chan-
ges in water depth may be more substantial than the
error caused by ‘low’ frequency of recording in 2002.
In 2002, daily resuspension averaged 64 g m)2
(Table 1). The maximum exceeded the mean Rd by a
Fig. 3 (a) Concentrations of suspended particulate matter (SPM) at various depths above the sediment surface and (b) the ratio of
SPM in the near-bottom layer (SPMB) and in the upper water column (SPMW).
238 V. Istvanovics et al.
� 2004 Blackwell Publishing Ltd, Freshwater Biology, 49, 232–252
factor of 6. In 2001, resuspension averaged
31 g m)2 day)1. Statistical analysis suggested that
the difference in frequency of recording turbidity (cf.
Fig. 4b) was responsible for the lower Rd in 2001
compared with 2002. We present internal load data
that have been corrected for this underestimation of
resuspension during 2001.
To account for changes in P desorption capacity of
resuspended sediments, we followed the variability in
fractional composition of P in SPM. Time-averaged
data indicated that resuspension of the fine-grained
sediment fraction selectively transported particles into
the water that were richer in potentially mobile
inorganic P (NH4Cl-RP and NaOH-RP) than the
uppermost 1 cm layer of the sediments (Fig. 5a). The
SPM was further enriched in mobile inorganic P over
the fine-grained fraction. This suggested that mobili-
sation of P took place after resuspension. NaOH-
extractable organic P (NaOH-nRP) content of SPM
exceeded that of the sediment by a mean factor of 3.
The mobile inorganic P content of SPM increased
significantly during the summer blooms of phyto-
plankton (Fig. 5b). No such seasonal variability could
be observed in the sediments (not shown). This was
another indication of P mobilisation following resus-
pension. As organic P (NaOH-nRP) is not directly
available for assimilation by algae, we assumed that
the P desorption capacity of the resuspended
sediment was proportional to the pool of potentially
mobile inorganic P in SPM.
The mean concentration of potentially mobile inor-
ganic P was 283 lg P g)1 dry mass. We assumed a
mean P desorption capacity of 5 lg P g)1 that repre-
sented 1.8% of the mobile P pool. The minimum P
desorption was 2 lg P g)1, whereas the maximum
reached 11 lg P g)1.
The internal P load averaged 0.28 ± 0.31 mg P
m)2 day)1 in 2001 and 0.39 ± 0.43 mg P m)2 day)1
in 2002 (Table 1). Considering the whole period of
study, the mean external load of SRP slightly excee-
ded the internal load in 2002. In 2001, the internal P
load reached only half of the external load of SRP
(Table 1).
The timing of external and internal loads differed
significantly (Fig. 6). While the former showed relatively
Fig. 4 (a) Autocorrelation function of concentration of suspen-
ded particulate matter and (b) the influence of frequency of
recording on the daily rate of resuspension (Rd) (in b R15d is the
rate estimated from 15-min recording and Rfid is that at
decreasing frequencies. The regression line is y ¼ 1.18 · 10)0.33x,
r2 ¼ 0.99).
Table 1 Mean rate of resuspension (Rd) and P load (both values were corrected for the low frequency of recording in 2001)
Period
Rd
(g m)2 day)1)
P load (mg P m)2 day)1)
External
Total P Soluble reactive P Internal
17 July–31 August 2001 59 ± 46 0.79 ± 0.50 0.63 ± 0.41 0.24 ± 0.21
1 September–9 October 2001 65 ± 84 0.44 ± 0.13 0.36 ± 0.12 0.32 ± 0.40
17 July–9 October 2001 62 ± 66 0.63 ± 0.41 0.51 ± 0.34 0.28 ± 0.31
27 March–30 June 2002 57 ± 67 1.06 ± 0.60 0.70 ± 0.48 0.41 ± 0.43
1 July–31 August 2002 73 ± 57 0.40 ± 0.10 0.29 ± 0.08 0.44 ± 0.51
1 September–21 October 2002 61 ± 58 0.29 ± 0.11 0.15 ± 0.05 0.29 ± 0.31
27 March–21 October 2002 64 ± 61 0.68 ± 0.55 0.45 ± 0.41 0.39 ± 0.43
Dynamics of internal P load 239
� 2004 Blackwell Publishing Ltd, Freshwater Biology, 49, 232–252
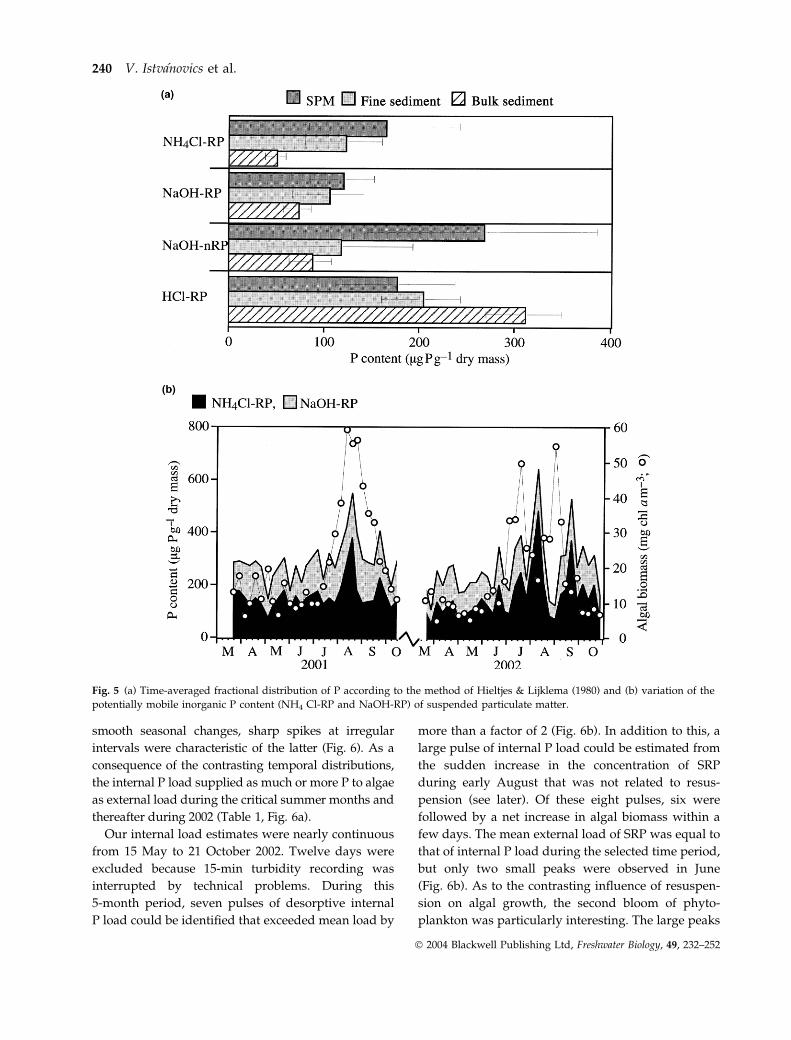
smooth seasonal changes, sharp spikes at irregular
intervals were characteristic of the latter (Fig. 6). As a
consequence of the contrasting temporal distributions,
the internal P load supplied as much or more P to algae
as external load during the critical summer months and
thereafter during 2002 (Table 1, Fig. 6a).
Our internal load estimates were nearly continuous
from 15 May to 21 October 2002. Twelve days were
excluded because 15-min turbidity recording was
interrupted by technical problems. During this
5-month period, seven pulses of desorptive internal
P load could be identified that exceeded mean load by
more than a factor of 2 (Fig. 6b). In addition to this, a
large pulse of internal P load could be estimated from
the sudden increase in the concentration of SRP
during early August that was not related to resus-
pension (see later). Of these eight pulses, six were
followed by a net increase in algal biomass within a
few days. The mean external load of SRP was equal to
that of internal P load during the selected time period,
but only two small peaks were observed in June
(Fig. 6b). As to the contrasting influence of resuspen-
sion on algal growth, the second bloom of phyto-
plankton was particularly interesting. The large peaks
Fig. 5 (a) Time-averaged fractional distribution of P according to the method of Hieltjes & Lijklema (1980) and (b) variation of the
potentially mobile inorganic P content (NH4 Cl-RP and NaOH-RP) of suspended particulate matter.
240 V. Istvanovics et al.
� 2004 Blackwell Publishing Ltd, Freshwater Biology, 49, 232–252
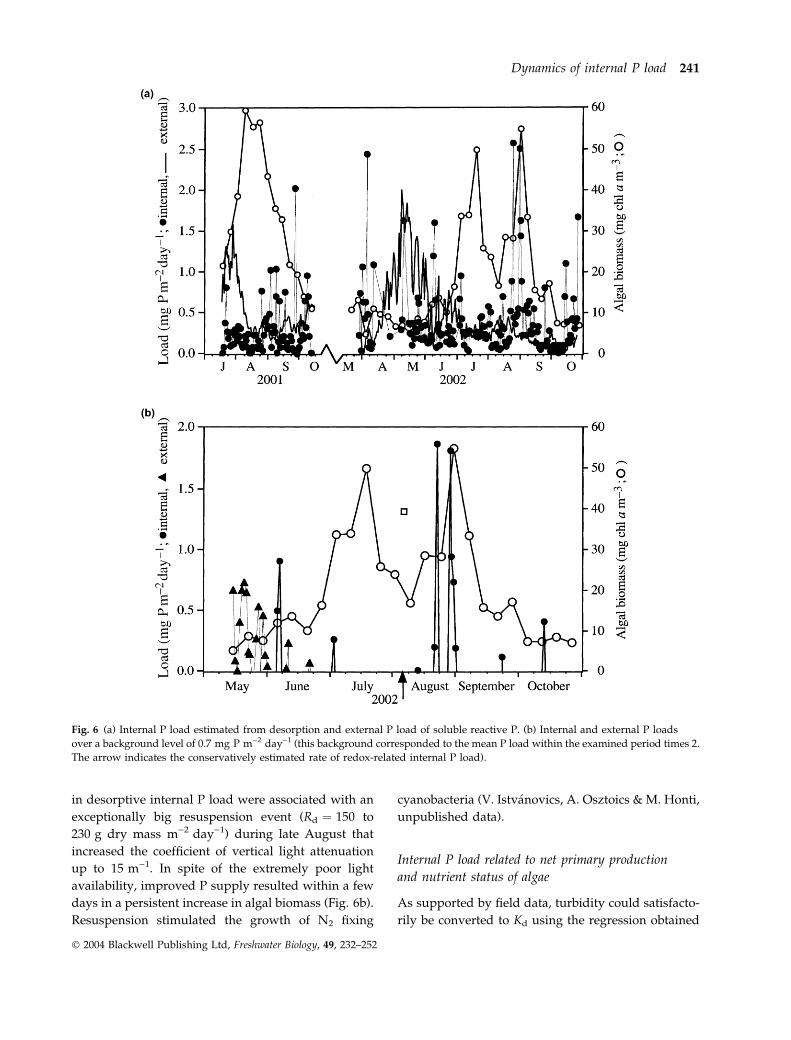
in desorptive internal P load were associated with an
exceptionally big resuspension event (Rd ¼ 150 to
230 g dry mass m)2 day)1) during late August that
increased the coefficient of vertical light attenuation
up to 15 m)1. In spite of the extremely poor light
availability, improved P supply resulted within a few
days in a persistent increase in algal biomass (Fig. 6b).
Resuspension stimulated the growth of N2 fixing
cyanobacteria (V. Istvanovics, A. Osztoics & M. Honti,
unpublished data).
Internal P load related to net primary production
and nutrient status of algae
As supported by field data, turbidity could satisfacto-
rily be converted to Kd using the regression obtained
Fig. 6 (a) Internal P load estimated from desorption and external P load of soluble reactive P. (b) Internal and external P loads
over a background level of 0.7 mg P m)2 day)1 (this background corresponded to the mean P load within the examined period times 2.
The arrow indicates the conservatively estimated rate of redox-related internal P load).
Dynamics of internal P load 241
� 2004 Blackwell Publishing Ltd, Freshwater Biology, 49, 232–252
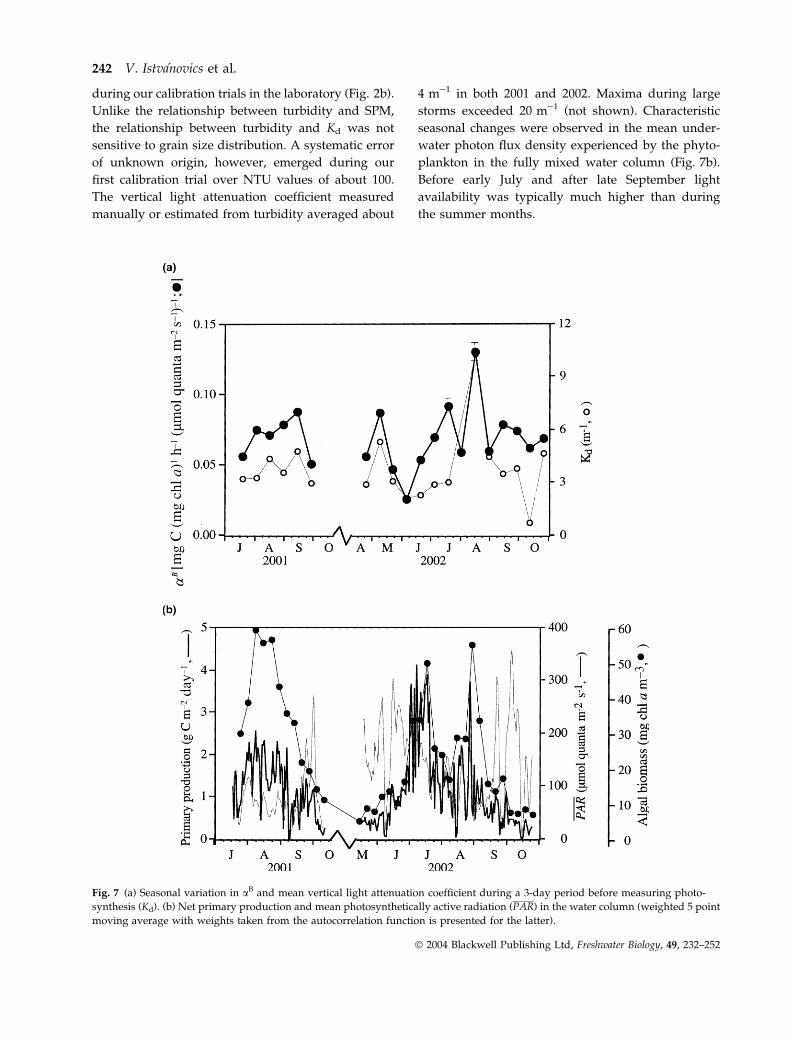
during our calibration trials in the laboratory (Fig. 2b).
Unlike the relationship between turbidity and SPM,
the relationship between turbidity and Kd was not
sensitive to grain size distribution. A systematic error
of unknown origin, however, emerged during our
first calibration trial over NTU values of about 100.
The vertical light attenuation coefficient measured
manually or estimated from turbidity averaged about
4 m)1 in both 2001 and 2002. Maxima during large
storms exceeded 20 m)1 (not shown). Characteristic
seasonal changes were observed in the mean under-
water photon flux density experienced by the phyto-
plankton in the fully mixed water column (Fig. 7b).
Before early July and after late September light
availability was typically much higher than during
the summer months.
Fig. 7 (a) Seasonal variation in aB and mean vertical light attenuation coefficient during a 3-day period before measuring photo-
synthesis (Kd). (b) Net primary production and mean photosynthetically active radiation (PAR) in the water column (weighted 5 point
moving average with weights taken from the autocorrelation function is presented for the latter).
242 V. Istvanovics et al.
� 2004 Blackwell Publishing Ltd, Freshwater Biology, 49, 232–252
Maximum rate of biomass specific photosynthesis
varied between 4 and 9 mg C (mg chl a))1 h)1 with
peak values during the summer (not shown). aB
ranged from 0.03 to 0.13 lg C (lg chl))1 h)1 (lmol -
quanta m)2 s)1))1. Changes in aB were best correlated
with the mean of vertical light attenuation coefficients
during a 3-day period preceding the measurement of
photosynthesis (Fig. 7a; r2 ¼ 0.59, n ¼ 19). Net pri-
mary production was estimated as 103 and 91 g C m)2
in the period 17 July to 9 October in 2001 and 2002,
respectively. During the period 15 May and 21 October
2002, net primary production reached 174 g C m)2.
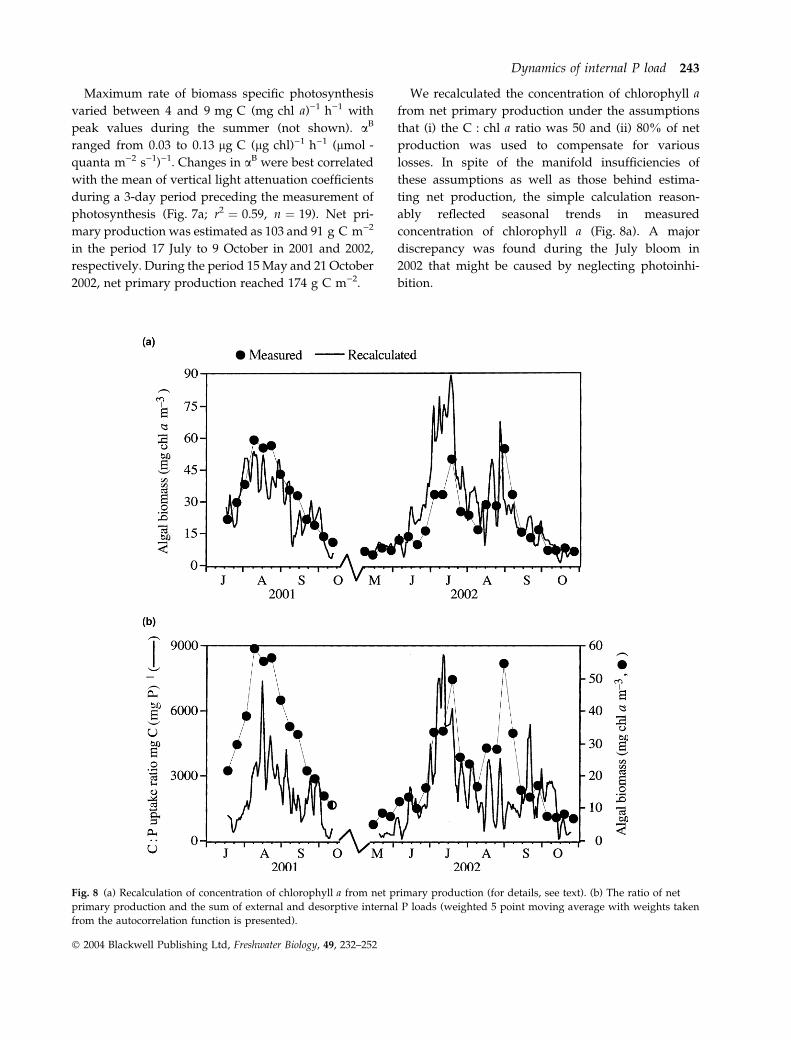
We recalculated the concentration of chlorophyll a
from net primary production under the assumptions
that (i) the C : chl a ratio was 50 and (ii) 80% of net
production was used to compensate for various
losses. In spite of the manifold insufficiencies of
these assumptions as well as those behind estima-
ting net production, the simple calculation reason-
ably reflected seasonal trends in measured
concentration of chlorophyll a (Fig. 8a). A major
discrepancy was found during the July bloom in
2002 that might be caused by neglecting photoinhi-
bition.
Fig. 8 (a) Recalculation of concentration of chlorophyll a from net primary production (for details, see text). (b) The ratio of net
primary production and the sum of external and desorptive internal P loads (weighted 5 point moving average with weights taken
from the autocorrelation function is presented).
Dynamics of internal P load 243
� 2004 Blackwell Publishing Ltd, Freshwater Biology, 49, 232–252
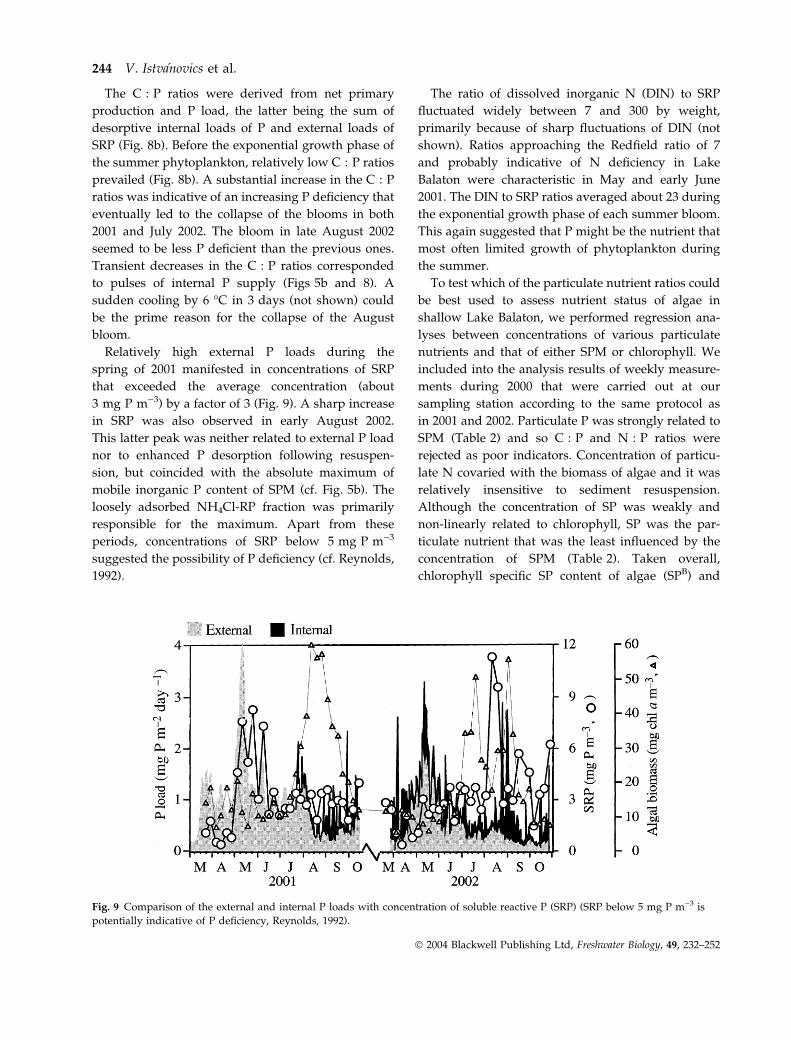
The C : P ratios were derived from net primary
production and P load, the latter being the sum of
desorptive internal loads of P and external loads of
SRP (Fig. 8b). Before the exponential growth phase of
the summer phytoplankton, relatively low C : P ratios
prevailed (Fig. 8b). A substantial increase in the C : P
ratios was indicative of an increasing P deficiency that
eventually led to the collapse of the blooms in both
2001 and July 2002. The bloom in late August 2002
seemed to be less P deficient than the previous ones.
Transient decreases in the C : P ratios corresponded
to pulses of internal P supply (Figs 5b and 8). A
sudden cooling by 6 �C in 3 days (not shown) could
be the prime reason for the collapse of the August
bloom.
Relatively high external P loads during the
spring of 2001 manifested in concentrations of SRP
that exceeded the average concentration (about
3 mg P m)3) by a factor of 3 (Fig. 9). A sharp increase
in SRP was also observed in early August 2002.
This latter peak was neither related to external P load
nor to enhanced P desorption following resuspen-
sion, but coincided with the absolute maximum of
mobile inorganic P content of SPM (cf. Fig. 5b). The
loosely adsorbed NH4Cl-RP fraction was primarily
responsible for the maximum. Apart from these
periods, concentrations of SRP below 5 mg P m)3
suggested the possibility of P deficiency (cf. Reynolds,
1992).
The ratio of dissolved inorganic N (DIN) to SRP
fluctuated widely between 7 and 300 by weight,
primarily because of sharp fluctuations of DIN (not
shown). Ratios approaching the Redfield ratio of 7
and probably indicative of N deficiency in Lake
Balaton were characteristic in May and early June
2001. The DIN to SRP ratios averaged about 23 during
the exponential growth phase of each summer bloom.
This again suggested that P might be the nutrient that
most often limited growth of phytoplankton during
the summer.
To test which of the particulate nutrient ratios could
be best used to assess nutrient status of algae in
shallow Lake Balaton, we performed regression ana-
lyses between concentrations of various particulate
nutrients and that of either SPM or chlorophyll. We
included into the analysis results of weekly measure-
ments during 2000 that were carried out at our
sampling station according to the same protocol as
in 2001 and 2002. Particulate P was strongly related to
SPM (Table 2) and so C : P and N : P ratios were
rejected as poor indicators. Concentration of particu-
late N covaried with the biomass of algae and it was
relatively insensitive to sediment resuspension.
Although the concentration of SP was weakly and
non-linearly related to chlorophyll, SP was the par-
ticulate nutrient that was the least influenced by the
concentration of SPM (Table 2). Taken overall,
chlorophyll specific SP content of algae (SPB) and
Fig. 9 Comparison of the external and internal P loads with concentration of soluble reactive P (SRP) (SRP below 5 mg P m)3 is
potentially indicative of P deficiency, Reynolds, 1992).
244 V. Istvanovics et al.
� 2004 Blackwell Publishing Ltd, Freshwater Biology, 49, 232–252
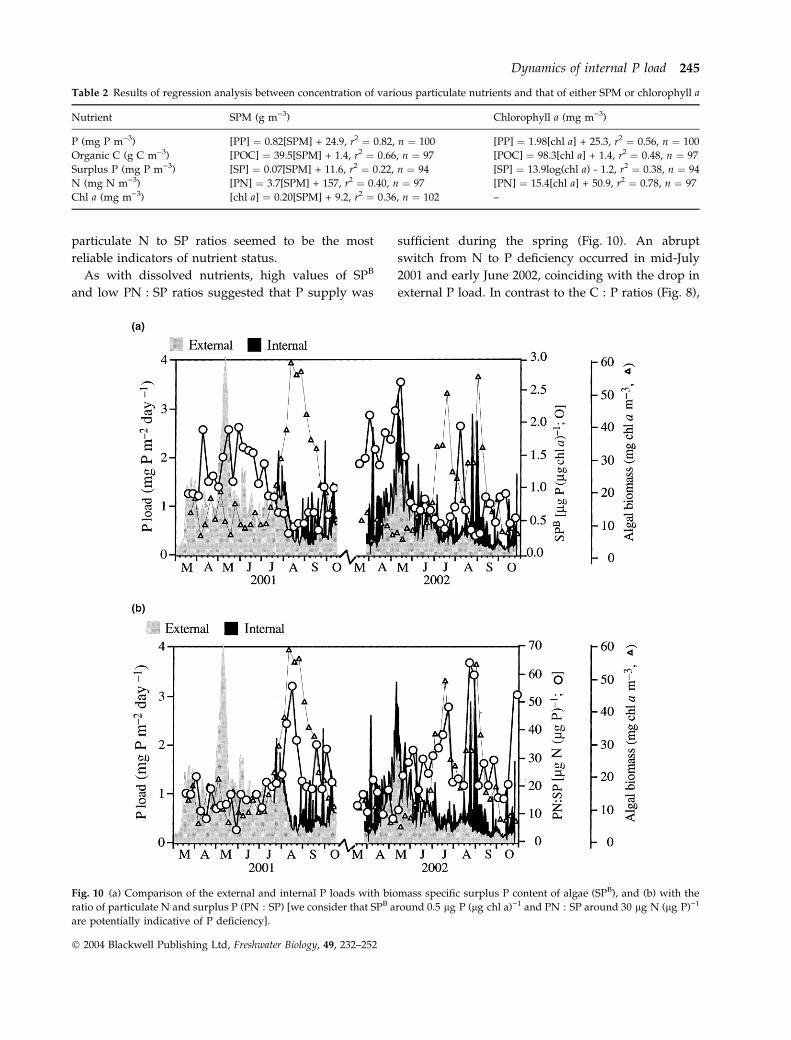
particulate N to SP ratios seemed to be the most
reliable indicators of nutrient status.
As with dissolved nutrients, high values of SPB
and low PN : SP ratios suggested that P supply was
sufficient during the spring (Fig. 10). An abrupt
switch from N to P deficiency occurred in mid-July
2001 and early June 2002, coinciding with the drop in
external P load. In contrast to the C : P ratios (Fig. 8),
Fig. 10 (a) Comparison of the external and internal P loads with biomass specific surplus P content of algae (SPB), and (b) with the
ratio of particulate N and surplus P (PN : SP) [we consider that SPB around 0.5 lg P (lg chl a))1 and PN : SP around 30 lg N (lg P))1
are potentially indicative of P deficiency].
Table 2 Results of regression analysis between concentration of various particulate nutrients and that of either SPM or chlorophyll a
Nutrient SPM (g m)3) Chlorophyll a (mg m)3)
P (mg P m)3) [PP] ¼ 0.82[SPM] + 24.9, r2 ¼ 0.82, n ¼ 100 [PP] ¼ 1.98[chl a] + 25.3, r2 ¼ 0.56, n ¼ 100
Organic C (g C m)3) [POC] ¼ 39.5[SPM] + 1.4, r2 ¼ 0.66, n ¼ 97 [POC] ¼ 98.3[chl a] + 1.4, r2 ¼ 0.48, n ¼ 97
Surplus P (mg P m)3) [SP] ¼ 0.07[SPM] + 11.6, r2 ¼ 0.22, n ¼ 94 [SP] ¼ 13.9log(chl a) - 1.2, r2 ¼ 0.38, n ¼ 94
N (mg N m)3) [PN] ¼ 3.7[SPM] + 157, r2 ¼ 0.40, n ¼ 97 [PN] ¼ 15.4[chl a] + 50.9, r2 ¼ 0.78, n ¼ 97
Chl a (mg m)3) [chl a] ¼ 0.20[SPM] + 9.2, r2 ¼ 0.36, n ¼ 102 –
Dynamics of internal P load 245
� 2004 Blackwell Publishing Ltd, Freshwater Biology, 49, 232–252

neither SPB nor PN : SP ratios revealed difference in P
supply during the three summer blooms. Transient
relaxation of P deficiency due to pulses of internal P
load could not be observed in these indicators, with the
exception of the presumably redox-related enhance-
ment of internal P load during early August, 2002.
Discussion
Resuspension and internal P load
The light scattering sensors turned out to be surpris-
ingly reliable tools for estimating both the concentra-
tion of SPM and vertical light attenuation coefficient
in Lake Balaton. This was not a priori evident, as the
same approach has failed in other lakes. Thus, van
Duin (1992) reported that in polymictic Markemeer,
the Netherlands, turbidity sensors overestimated
concentration of SPM in the field by a factor of 5–50.
At the same time, Markensten & Pierson (2003)
reported that SPM concentrations could reliably be
approximated in shallow Lake Malaren, Sweden,
using transmissometer sensors. Composition and
grain size distribution of sediments are the likely
reason for such intersystem differences. In this
respect, the most influential features of Balaton
sediments are high magnesian calcite content (50–
65% of dry mass), very low organic content (<4%),
most particles are <100 lm and mean particle size is
10–40 lm (Mate, 1987).
Because we recorded turbidity at various depths
but at a single station, the influence of horizontal
transport on changes in SPM remains unknown.
Wind-induced sediment resuspension has been mod-
elled with success in Lake Balaton assuming local
equilibrium with the wind and negligible horizontal
transport (Luettich, Harleman & Somlyody, 1990).
Occasional samplings during variable wind condi-
tions indicated that SPM concentrations were not
significantly different between our permanent sam-
pling station and various locations in Basin 1. Thus,
horizontal transport was not likely to seriously distort
our estimates of resuspension.
The mean rate of resuspension was about
60 g m)2 day)1 in both years after correcting for the
50% underestimation of Rd in 2001 because of low
frequency (60 min) of recording. One must, however,
bear in mind that the water depth at our sampling
station (1.2 m) was only half of the mean depth of
Basin 1 of Lake Balaton (2.3 m). As in our station, a
significant increase in the SPM concentration was
restricted to a near-bottom layer of 0.2 m or so in
deeper areas of Lake Balaton (Somlyody, 1984). Thus,
basin-wide daily resuspension could be about twice as
high as that estimated for our shallow station.
Modelling sediment resuspension in Lake Balaton,
Luettich et al. (1990) found that a few millimetre thick
surface layer of the sediments behaved in a fully non-
cohesive manner. In agreement with this, the rate of
sediment resuspension was much higher in Lake
Balaton than in many other shallow lakes (Aalderink
et al., 1985; Evans, 1994; Kozerski & Kleeberg, 1998).
Luettich et al. (1990) speculated that bioturbation
and wind-induced agitation might be responsible for
the non-cohesivity. Extrapolation of our data (Fig. 3)
to a 2.3 m deep column showed that the mean SPM
content was 116 g dry mass m)2 with maxima
approaching 490 g m)2. On the assumption that water
content of the sediments subject to resuspension was
95%, stirring up of a 2.3 mm thick layer accounted for
the mean SPM content. The maximum corresponded
to 9.5 mm. Extrapolating again to the mean depth of
2.3 m, rate of daily resuspension would also corres-
pond to a 2.3 mm thick layer. Thus, the turnover of
the non-cohesive layer could be estimated as 1 day)1.
There are two obvious consequences of this rapid
turnover. First, it suggests that resuspension itself is a
crucial factor in maintaining non-cohesivity of surface
sediments. Secondly, replenishment of P desorbed
upon resuspension must be rapid provided that P
desorption capacity remains constant. Indeed,
Søndergaard et al. (1992) found that the rate of P
desorption was constant during repeated experimen-
tal resuspension of sediments and concluded that
replenishment of P was fast in Arresø, Denmark. No
comparable evidence is available from Lake Balaton.
However, intense microbial activity at the sediment/
water interface (Zlinszky, 1987) as well as bioturba-
tion and metabolic activity of macrozoobenthos sug-
gest that fast replenishment may also occur in our
lake.
The fine-grained sediment fraction was separated
from the bulk sediment on the basis of its slow
sedimentation. Fractional composition of P was
clearly indicative of selective resuspension of particles
that were enriched in potentially mobile inorganic P
and depleted in less mobile HCl-RP relative to the
bulk sediment (Fig. 5). The concentration of mobile
246 V. Istvanovics et al.
� 2004 Blackwell Publishing Ltd, Freshwater Biology, 49, 232–252

inorganic P further increased upon resuspension. This
can be explained by the finding that an increase in pH
was the most important factor of P mobilisation in
Lake Balaton (Pettersson & Bostrom, 1986; Istvanov-
ics, 1988).
The central idea of our study was that high
resolution internal load estimates can be derived from
the continuously recorded rate of sediment resuspen-
sion and nutrient desorption capacity of resuspended
sediments. It is likely that desorption of ammonium
occurs upon resuspension (Ogilvie & Mitchell, 1998).
However, as no information was available on ammo-
nium desorption in Lake Balaton, we have not paid
attention to the internal load of N.
We must admit that our assumptions of P
desorption of sediments were based on relatively
weak experimental evidence. These assumptions
were as follows: (i) P desorption was proportionate
to the potentially mobile inorganic P fraction of SPM
and (ii) mean capacity was 5 lg P g)1 dry mass.
Assumption (i) was likely to cause less uncertainty, as
variability in Rd highly exceeded that in mobile
inorganic P content of SPM as indicated by the
coefficients of variation (0.97 and 0.36, respectively).
As a consequence, temporal changes in desorptive P
load were more strongly influenced by the dynamics
of resuspension than by slower, mostly seasonal
changes in mobile inorganic P content of SPM. The
experimental basis for assumption (ii) was far from
strong because of the difficulty in detecting changes
accurately in low SRP concentration during P desorp-
tion experiments that are carried out within the
realistic range of SPM concentrations (Lijklema et al.,
1986). Lijklema et al. (1986) concluded that the P
desorption capacity of sediments varied between
5 and 10 lg P g)1 in Lake Balaton. Our assumptions
resulted in a reasonably similar range (2 and
11 lg P g)1). Nevertheless, the choice of a different
mean value of P desorption capacity would induce a
proportionate change in the absolute value of internal
P load. These uncertainties led us to testing our
estimate against net primary production and nutrient
status of phytoplankton.
Net primary production was a rough approxima-
tion, as several features of dynamic photosynthetic
response of phytoplankton were neglected (cf. Fal-
kowski & Raven, 1997). We only accounted for
photoacclimation by changing the value of aB accord-
ing to the relationship between measured aB and
mean Kd during a 3-day period preceding the meas-
urement of photosynthesis (cf. Kiefer & Mitchell,
1983). In spite of these oversimplifications, the
estimate agreed well with other estimates of primary
production in Basin 1 of Lake Balaton (L. Voros,
personal communication). Moreover, the seasonal
trend of measured concentrations of chlorophyll a
could reasonably be recalculated from our estimate.
Thus, seasonal changes in C : P ratios derived from
net primary production and from the sum of external
and internal P loads are likely to reflect real changes in
P availability. Absolute values of C : P ratios were not
conclusive because of the substantial recycling of P
within the water column (Herodek, 1986; Voros,
V-Balogh & Herodek, 1996).
The comparison of internal P load with indicators
of nutrient status obtained from chemical measure-
ments was no less problematic than the above
approach. In general, dissolved nutrients and their
ratios are inferior indicators of nutrient status of
phytoplankton relative to ratios of particulate nu-
trients. In Lake Balaton, however, the interference of
resuspension cannot be neglected. Regression analy-
sis revealed that only particulate N and SP were
more strongly related to changes in algal bio-
mass than to changes in inorganic SPM (Table 2).
Pettersson (1980) suggested from his measurements
in stratified Lake Erken, Sweden, that SPB values
around 0.2 lg P (lg chl a))1 were indicative of P
deficiency. We suggest that this threshold was about
0.5 lg P (lg chl a))1 in shallow Lake Balaton. Simi-
larly, the PN : SP ratio varied between 10 and
20 lg N (lg P))1 in P sufficient cultures of Cylindro-
spermopsis raciborskii. Increasing P deficiency induced
a tenfold increase in this ratio (V. Istvanovics,
A. Osztoics & M. Honti, unpublished data). This N2
fixing cyanobacterium was dominant during the
bloom in 2001 and co-dominant in 2002 (J. Padisak,
personal communication). In the field, PN : SP ratio
ranged from 10 to 50 lg N (lg P))1. Values over
30 lg N (lg P))1 might be indicative of P deficiency.
As a consequence of all of the above-mentioned
uncertainties, seasonal trends rather than absolute
values of internal P load could be tested against
independent data sets.
Various indicators (Figs 8–10) resulted in a similar
conclusion about seasonal changes in the nutrient
status of phytoplankton. The external load during the
spring was high enough to sustain P sufficiency of the
Dynamics of internal P load 247
� 2004 Blackwell Publishing Ltd, Freshwater Biology, 49, 232–252

low algal biomass present, and even to increase the
SRP concentration in 2001. Internal P load together
with the diminished external load during the summer
was too low to maintain P sufficiency. Thus, summer
blooms were P deficient, and the yield was P-deter-
mined sensu Reynolds (1992). The agreement between
C : P ratios and nutrient ratios suggested that our
method of estimating internal P load captured the
seasonal trends correctly.
We must note that within the frame of the present
study, diurnal changes in photosynthetic activity of
various algal groups were followed using a novel
technique, the on-line delayed fluorescence excita-
tion spectroscopy (Gerhardt & Bodemer, 2000). We
plan to test our internal load estimates against this
data set that has the same temporal resolution as
our turbidity time series. However, the results
obtained by DF spectroscopy first require a critical
evaluation.
The internal P load averaged 0.39 mg P m)2 day)1
in 2002 and 0.28 mg P m)2 day)1 in 2001. The ampli-
tude of larger pulses varied between 1 and
2.5 mg P m)2 day)1. Release rates of P reached the
amplitude of these pulses from both batch systems
(Pettersson & Bostrom, 1986) and intact sediment
cores (Istvanovics, 1988). As discussed previously,
basin-wide Rd and in turn, internal P load were about
twice as high as in our shallow station. Nevertheless,
the difference between the rate of desorptive P load
and release rates measured in laboratory remained
surprisingly small. For example, Søndergaard et al.
(1992) estimated an internal P load of 60–
70 mg P m)2 day)1 in Arresø, Denmark from resus-
pension experiments in laboratory. This load was
20–30 times greater than P release from undisturbed
sediment cores. Of resuspended P, 12% was
desorbed. Desorption experiments with sediments of
Lake Balaton showed an order of magnitude lower P
desorption capacity (Lijklema et al., 1986) than sedi-
ments of Arresø. Accordingly, the rate of internal P
load was also very much lower (Table 1, Fig. 6). This
is another indication of the favourable behaviour of
highly calcareous sediments of Lake Balaton that has
been inferred from the evaluation of long-term data
by Istvanovics & Somlyody (2001).
Unlike our estimates of internal P load, area-specific
external P load is independent of water depth.
Consequently, basin-wide mean internal P load might
be similar to the external load of total P during the
study period in 2002, and exceeded the latter by a
factor of 2 during summer (Table 1). Previous experi-
mental and modelling studies (Pettersson & Bostrom,
1986; Somlyody, 1986; Istvanovics, 1988) also conclu-
ded that on an annual basis external and internal
loads were similar. The comparison suggests that
the magnitude of our internal P load estimate was
realistic.
The proportion of external to internal P load may
show considerable interannual fluctuations. Our
study period was too short to assess long-term
variability in internal P load. The external load in
2001 reached only 42% of the mean load during the
period 1993–2002. In 2002, the respective value was
30%. The reason was the extreme deficit in precipi-
tation during the last 3 years, and closure of the weir
at the mouth of the Zala River in the summer. This
intervention resulted in the sudden drop of external P
load in mid-July 2001 and early June 2002 (Figs 6, 9
and 10). Internal load together with the diminished
external load failed to ease the increasing P deficiency
of summer blooms (Figs 9 and 10). Thus, closure of
the weir efficiently reduced carrying capacity of Basin
1. In years of mean or high flow, the relative
importance of internal P load is expected to be lower.
At the same time, evaluation of long-term data
suggested that in years of extreme blooms of C.
raciborskii, internal load exceeded external one during
summers (Istvanovics & Somlyody, 2001; Istvanovics
et al., 2002).
Although P desorption following sediment resus-
pension is the key process of internal P load in
wind-exposed shallow lakes (Lijklema et al., 1986;
Søndergaard et al., 1992), it is not the only component.
During the present study, we also followed nutrient
gradients in the interstitial water by the dialysis
technique. Cylindrical rod samplers separated into
nine chambers of 3 cm length were inserted into the
sediment at monthly intervals at our station. The
upward diffusive flux of P peaked at
0.2 mg P m)2 day)1 in the upper 10 cm sediment
layer during the summer. The simultaneous flux of
iron, however, was high enough to retain P flux from
deeper sediments provided that Fe(III) could precipi-
tate in the oxidised surface layer (cf. Lofgren, 1987;
Lofgren & Bostrom, 1989). The SRP concentration at
1.5 cm below the sediment surface exceeded that at
1.5 cm above the surface by a factor of 2–3. Thus,
neither advection of pore water nor diffusion of P
248 V. Istvanovics et al.
� 2004 Blackwell Publishing Ltd, Freshwater Biology, 49, 232–252

could, in general, significantly contribute to internal P
load. The first week of August 2002 was an exception.
In that week, SRP concentration suddenly increased in
the water by 8 mg P m)3. Such a phenomenon has
never been reported during summer in Lake Balaton.
We immediately checked and found that SRP showed
a similar peak at various locations of Basin 1. The
most likely explanation was that extreme weather
conditions led to the reduction of sediment surface.
This event was an obviously important source of
internal P load that temporarily eased the P deficiency
of phytoplankton. We certainly underestimated the
rate of internal load during this period because of
both P assimilation by algae and the possibility that
enhanced turbulent diffusion was restricted to a few
days instead of a week.
Implications for phytoplankton ecology
Autocorrelation analysis of SPM time series revealed
that an appropriate characterisation of SPM concen-
tration required daily sampling. The same holds true
for another derivative of turbidity, Kd. Considering
that (i) short-term variability in underwater light
field induces a series of photoacclimation reactions
(Falkowski & Raven, 1997), (ii) variability over
1–2 weeks repeatedly disrupts phytoplankton succes-
sion (Padisak, G.-Toth & Rajczy, 1988; Padisak et al.,
1990) and (iii) seasonal changes represent a major
driving force for succession (Reynolds, 1997), one
must conclude that routine monitoring programmes
yield very little insight into shallow lakes as habitats
for phytoplankton. Moreover, sufficiently precise
estimates of sediment resuspension and the associ-
ated desorptive flux of P require an even higher
frequency of sampling (15 min or higher in Lake
Balaton). This is because the higher the frequency of
recording, the closer we approximate gross resuspen-
sion. Automatic recording of turbidity or other
parameters convertible to SPM and Kd are the only
means that will provide sufficient information about
growth conditions of algae in wind-exposed shallow
lakes.
Apart from hypertrophic lakes where self-shading
of phytoplankton significantly contributes to vertical
light attenuation, maximal light availability is expec-
ted to occur during the summer. In this respect, the
only difference between a stratified deep lake and a
polymictic shallow lake is that frequent resuspension
induces sharp oscillations in light in the latter. In
his lake typology, Reynolds (1997) referred to Lake
Balaton as one of the illustrative examples of
polymictic lakes. However, our data do not support
this expectation. Mean photon flux density in the
water column was lowest, albeit widely fluctuating,
between early July and late September in both 2001
and 2002. This increase in vertical light attenuation
might have two causes. First, calcite precipitation is
very intense in this highly calcareous lake. Biogen-
ically induced carbonate precipitation is proportion-
ate to photosynthesis that peaks in summer.
Concentration of Ca was depleted from about 60 to
30 g m)3 during the summer of both years. Sedi-
mentation of freshly precipitated, small calcite crys-
tals is presumably extremely slow (2 m day)1 or less,
cf. Somlyody & Koncsos, 1991). Secondly, mean
sediment resuspension was somewhat higher during
the summer than during the cold seasons at least
in 2002. Diminished light availability during the
summer must be an important factor in selection
for shade tolerant cyanobacteria in Lake Balaton
(Padisak & Reynolds, 1998), as well as in other
calcareous lakes.
Strong resuspension events induce well defined
restructuring of phytoplankton assemblages in Lake
Balaton (Padisak et al., 1988, 1990; Padisak, 1993): the
immediate effect includes an increase in the biomass
of meroplanktonic species and, above an unspecified
threshold effect of wind, a substantial decrease in the
biomass of filamentous cyanobacteria. Resuspension
is followed by an outburst of small, shade tolerant,
r-selected species, including picoalgae. After a few
days, these are gradually replaced by larger, slowly
growing K-selected species. These patterns can best be
identified when large resuspension events follow each
other in 10–15 days. Our data showed that the
frequency of large resuspension events often matched
this criterion. We also observed that out of eight
pulses of internal P load between May and October
2002, six were followed by a net biomass increase
within a few days. Either the pulses were too low, or
the sampling frequency was too long, the enhanced P
supply failed to induce detectable changes in nutrient
status of phytoplankton. Seasonal changes in C : P
ratios (Fig. 8b), however, suggested that exponential
development of summer blooms was only possible at
the expense of previously stored P. Our findings
together with those of Padisak (1993) and Padisak
Dynamics of internal P load 249
� 2004 Blackwell Publishing Ltd, Freshwater Biology, 49, 232–252

et al. (1988, 1990) indicate that pulses of P were large
enough to select for K species with a high P storage
capacity. Thus, sediment resuspension can be per-
ceived not only as a disturbance factor but also as a
factor that periodically relaxes nutrient stress. The
former feature may dominate the instantaneous effect,
whereas the latter may determine the persistent effect
of resuspension on succession of phytoplankton.
Acknowledgments
This study was financially supported by the grant
EVK1-CT-1999-00037 ‘Phytoplankton-om-line’ and by
the Balaton Project of the Office of the Prime Minister.
We are indebted to Dr Laszlo Somlyody, Dr Kurt
Pettersson and two anonymous referees for criti-
cally reading this manuscript. We thank the West-
Transdanubian Water Authority and the Balaton
Limnological Research Institute of the Hungarian
Academy of Sciences for their permission to use their
laboratory facilities. We are indebted to Dr Volkmar
Gerhardt for installing the light scattering sensors at
our station. Mr Norbert Turay and Mr Gabor Poor
provided excellent technical assistance.
References
Aalderink R.H., Lijklema L., Brenkelman J., van
Raaphorst W. & Brinkman A.G. (1985) Quantification
of wind induced resuspension in a shallow lake. Water
Science and Technology, 17, 943–954.
Bostrom B. (1984) Potential mobility of phosphorus in
different types of lake sediments. Internazionale Revue
der gesamten Hydrobiologie, 69, 457–474.
Bostrom B., Jansson M. & Forsberg C. (1982) Phosphorus
release from lake sediments. Archiv fur Hydrobiologie
Beiheft Ergebnisse der Limnologie, 18, 5–59.
Crumpton W.G., Isenhart T.M. & Mitchell P.D. (1992)
Nitrate and organic nitrogen analyses with second-
derivative spectroscopy. Limnology and Oceanography,
37, 907–913.
van Duin E.H.S. (1992) Sediment Transport, Light and Algal
Growth in the Markemeer, PhD thesis. Department of
Nature Conservation, Agricultural University Wagen-
ingen, The Netherlands.
Evans R.D. (1994) Empirical evidence of the importance
of sediment resuspension in lakes. Hydrobiologia, 284,
5–12.
Falkowski P.G. & Raven J.A. (1997) Aquatic Photosyn-
thesis. Blackwell Science, New York.
FBA (1978) Water Analysis: Some Revised Methods for
Limnologists (Eds F.J.H. Mackereth, J. Heron &
J.F. Talling). Freshwater Biological Association,
Ambleside, U.K.
Fitzgerald C.P. & Nelsson T. (1966) Extractive and
enzymatic analyses for limiting or surplus phosphorus
in algae. Journal of Phycology, 2, 32–37.
Gerhardt V. & Bodemer U. (2000) Delayed fluorescence
excitation spectroscopy: a method for determining
phytoplankton composition. Archiv fur Hydrobiologie
Spec. Issues Advanc. Limnol., 55, 101–120.
Gorzo Gy. (1991) Studies for determining technology of
sediment dredging (in Hungarian). Vızugyi Kozleme-
nyek, 78, 186–193.
Hamilton D.P. & Mitchell S.F. (1997) Wave-induced shear
stresses, plant nutrients and chlorophyll in seven
shallow lakes. Freshwater Biology, 38, 159–168.
Herodek S. (1986) Phytoplankton changes during eutro-
phication and P and N metabolism. In: Modeling and
Managing Shallow Lake Eutrophication (Eds L. Somlyody
& G. Straten van), pp. 183–204. Springer-Verlag, Berlin.
Hieltjes A.H.M. & Lijklema L. (1980) Fractionation of
inorganic phosphates in calcareous sediments. Journal
of Environmental Quality, 9, 405–407.
Istvanovics V. (1988) Seasonal variation of phosphorus
release from the sediments of shallow Lake Balaton
(Hungary). Water Research, 22, 1473–1481.
Istvanovics V. & Herodek S. (1995) Estimation of net
uptake and leakage rates of orthophosphate from 32P
uptake kinetics by a force-flow model. Limnology and
Oceanography, 40, 17–32.
Istvanovics V. & Somlyody L. (2001) Factors influencing
lake recovery from eutrophication – the case of Basin 1
of Lake Balaton. Water Research, 35, 729–735.
Istvanovics V., Somlyody L. & Clement A. (2002)
Cyanobacteria-mediated internal eutrophication in
shallow Lake Balaton after load reduction. Water
Research, 36, 3314–3322.
Jassby A.D. & Platt T. (1976) Mathematical formulation of the
relationship between photosynthesis and light for phyto-
plankton. Limnology and Oceanography, 21, 520–547.
Kiefer D.A. & Mitchell B.C. (1983) A simple steady state
description of phytoplankton growth based on absorp-
tion cross spectra and quantum efficiency. Limnology
and Oceanography, 28, 770–776.
Kozerski H.-P. & Kleeberg A. (1998) The sediments and
benthic-pelagic exchange in the shallow Lake Muggel-
see (Berlin, Germany). Internazionale Revue der gesamten
Hydrobiologie, 83, 77–112.
Kuo S. & Lotse G. (1974) Kinetics of phosphate adsorp-
tion and desorption by lake sediments. Soil Science
Society of America Proceedings, 38, 50–54.
250 V. Istvanovics et al.
� 2004 Blackwell Publishing Ltd, Freshwater Biology, 49, 232–252

Lewis M.R. & Smith C.R. (1983) A small volume, short
incubation-time method for the measurement of
photosynthesis as a function of incident irradiance.
Marine Ecology Progress Series, 13, 211–221.
Li W.C., Armstrong D.E., Williams J.D.H., Harris R.F. &
Syers J.K. (1972) Rate and extent of inorganic phos-
phate exchange in lake sediments. Soil Science Society of
America Proceedings, 36, 279–285.
Lijklema L., Gelencser P., Szilagyi F. & Somlyody L.
(1986) Sediment and its interaction with water. In:
Modeling and Managing Shallow Lake Eutrophication (Eds
L. Somlyody & G. Straten van), pp. 156–183. Springer-
Verlag, Berlin.
Lofgren S. (1987) Phosphorus Retention in Sediments –
Implications for Aerobic Phosphorus Release in Shallow
Lakes, PhD thesis. Uppsala University, Sweden.
Lofgren S. & Bostrom B. (1989) Interstitial water
concentrations of phosphorus, iron and manganese
in a shallow eutrophic Swedish lake – implications
for phosphorus cycling. Water Research, 23, 1115–
1125.
Lorenzen C.J. (1967) Determination of chlorophyll and
phaeo-pigments: spectrophotometric equations. Lim-
nolology and Oceanography, 12, 343–346.
Luettich R.A., Harleman D.R.F. & Somlyody L. (1990)
Dynamic behavior of suspended sediment concen-
trations in a shallow lake perturbed by episodic
wind events. Limnology and Oceanography, 35, 1050–1067.
Markensten H. & Pierson D.C. (2003) A dynamic model
for flow and wind driven sediment resuspension in a
shallow basin. Hydrobiologia, 494, 305–311.
Mate F. (1987) Mapping of recent sediments in Lake
Balaton (In Hungarian). In: A Magyar Allami Foldtani
Intezet evi jelentese az 1985. evrol, pp. 367–379.
Menzel D.H. & Corwin N. (1965) The measurement of
total phosphorus in seawater based on the liberation of
organically bound fractions by persulfate oxidation.
Limnology and Oceanography, 10, 280–282.
van der Molen D.T. & Boers P.C.M. (1994) Influence of
internal loading on phosphorus concentration in
shallow lakes before and after reduction of the external
loading. Hydrobiologia, 275/276, 379–389.
Murphy J. & Riley J.P. (1962) A modified single solution
method for the determination of phosphate in natural
waters. Analytica Chemica Acta, 27, 31–36.
Ogilvie B.G. & Mitchell S.F. (1998) Does sediment
resuspension have persistent effects on phytoplank-
ton? Experimental studies in three shallow lakes.
Freshwater Biology, 49, 51–63.
Padisak J. (1993) The influence of different timescale
disturbances on the species richness, diversity and
equitability of phytoplankton in shallow lakes. Hydro-
biologia, 249, 135–156.
Padisak J. & Reynolds C.S. (1998) Selection of phyto-
plankton associations in Lake Balaton, Hungary, in
response to eutrophication and restoration measures,
with special reference to the cyanoprokaryotes. Hydro-
biologia, 384, 41–53.
Padisak J., G.-Toth L. & Rajczy M. (1988) The role of
storms in the summer succession of phytoplankton in a
shallow lake (Lake Balaton, Hungary). Journal of
Plankton Research, 10, 249–265.
Padisak J., G.-Toth L. & Rajczy M. (1990) Stir-up effect of
wind on a more-or-less stratified shallow lake phyto-
plankton community, Lake Balaton, Hungary. Hydro-
biologia, 191, 249–254.
Pettersson K. (1980) Alkaline phosphatase activity and
algal surplus phosphorus as phosphorus deficiency
indicators in Lake Erken. Archiv fur Hydrobiologie, 89,
54–87.
Pettersson K. & Bostrom B. (1986) Phosphorus exchange
between sediment and water in Lake Balaton. In:
Sediments and Water Interactions (Ed. P.G. Sly), pp. 427–
435. Springer-Verlag, New York.
Reynolds C.S. (1992) Eutrophication and the manage-
ment of planktonic algae: what Vollenweider couldn’t
tell us. In: Eutrophication: Research and Application to
Water Supply (Eds D.W. Sutcliffe & J.G. Jones), pp. 4–
29. Freshwater Biological Association, Ambleside, U.K.
Reynolds C.S. (1997) Vegetation Processes in the Pelagic: A
Model for Ecosystem Theory. Ecology Institute, Olden-
dorf/Luhe.
Rijkeboer M., De Bles F. & Gons H.J. (1991) Role of sestonic
detritus as a P-buffer. Mem. Ist. ital. Idrobiol., 48, 251–260.
Sas H. (1989) Lake Restoration and Reduction of Nutrient
Loading: Expectations, Experiences, Extrapolations. Acad-
emia Verlag Richarz, St Augustin.
Somlyody L. (1984) Water Quality Management in Shallow
Lakes (in Hungarian), DSc thesis. University of Tech-
nology and Economy, Budapest.
Somlyody L. (1986) Eutrophication management models.
In: Modeling and Managing Shallow Lake Eutrophication
(Eds L. Somlyody & G. Straten van), pp. 69–110.
Springer-Verlag, Berlin.
Somlyody L. & Koncsos L. (1991) Influence of sediment
resuspension on the light conditions and algal growth
in Lake Balaton. Ecological Modelling, 57, 173–192.
Søndergaard M., Kristensen P. & Jeppesen E. (1992)
Phosphorus release from resuspended sediment in the
shallow and wind-exposed Lake Arresø, Denmark.
Hydrobiologia, 228, 91–99.
Søndergaard, M., Jensen J.P. & Jeppesen E. (1999):
Internal phosphorus loading in shallow Danish lakes.
Hydrobiologia, 408/409, 145–152.
Dynamics of internal P load 251
� 2004 Blackwell Publishing Ltd, Freshwater Biology, 49, 232–252

Voros L., V-Balogh K. & Herodek S. (1996) Microbial
food web in a large, shallow lake (Lake Balaton,
Hungary). Hydrobiologia, 339, 57–65.
Williams J.D.H., Syers J.K. & Harris R.F. (1970) Adsorp-
tion and desorption of inorganic phosphorus by lake
sediments in a 0.1 M NaCl system. Environmental
Science and Technology, 4, 517–519.
Zlinszky J. (1987) The Biologically Available Nutrient
Loading and Bacterial Production in Lake Balaton (in
Hungarian), PhD thesis. Balaton Limnological Research
Institute, Tihany, Hungary.
(Manuscript accepted 15 December 2003)
252 V. Istvanovics et al.
� 2004 Blackwell Publishing Ltd, Freshwater Biology, 49, 232–252
Related Documents











