This article was downloaded by: [UNIVERSITY OF ADELAIDE LIBRARIES] On: 22 July 2013, At: 19:45 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Archives of Agronomy and Soil Science Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/gags20 Transgenic plants for abiotic stress tolerance: current status Syed Sarfraz Hussain a , Hasnain Raza a , Irfan Afzal b & Mahmood Akhtar Kayani c a Institute for Molecular Physiology and Biotechnology of Plants (IMBIO-Bartels), University of Bonn, Bonn, Germany b Department of Crop Physiology, University of Agriculture, Faisalabad, Pakistan c Department of Biosciences, COMSATS Institute of Information Technology (CIIT), Islamabad, Pakistan Published online: 16 Jun 2011. To cite this article: Syed Sarfraz Hussain , Hasnain Raza , Irfan Afzal & Mahmood Akhtar Kayani (2012) Transgenic plants for abiotic stress tolerance: current status, Archives of Agronomy and Soil Science, 58:7, 693-721, DOI: 10.1080/03650340.2010.540010 To link to this article: http://dx.doi.org/10.1080/03650340.2010.540010 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. Terms &

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article was downloaded by: [UNIVERSITY OF ADELAIDE LIBRARIES]On: 22 July 2013, At: 19:45Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Archives of Agronomy and Soil SciencePublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/gags20
Transgenic plants for abiotic stresstolerance: current statusSyed Sarfraz Hussain a , Hasnain Raza a , Irfan Afzal b & MahmoodAkhtar Kayani ca Institute for Molecular Physiology and Biotechnology of Plants(IMBIO-Bartels), University of Bonn, Bonn, Germanyb Department of Crop Physiology, University of Agriculture,Faisalabad, Pakistanc Department of Biosciences, COMSATS Institute of InformationTechnology (CIIT), Islamabad, PakistanPublished online: 16 Jun 2011.
To cite this article: Syed Sarfraz Hussain , Hasnain Raza , Irfan Afzal & Mahmood Akhtar Kayani(2012) Transgenic plants for abiotic stress tolerance: current status, Archives of Agronomy and SoilScience, 58:7, 693-721, DOI: 10.1080/03650340.2010.540010
To link to this article: http://dx.doi.org/10.1080/03650340.2010.540010
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the“Content”) contained in the publications on our platform. However, Taylor & Francis,our agents, and our licensors make no representations or warranties whatsoever as tothe accuracy, completeness, or suitability for any purpose of the Content. Any opinionsand views expressed in this publication are the opinions and views of the authors,and are not the views of or endorsed by Taylor & Francis. The accuracy of the Contentshould not be relied upon and should be independently verified with primary sourcesof information. Taylor and Francis shall not be liable for any losses, actions, claims,proceedings, demands, costs, expenses, damages, and other liabilities whatsoever orhowsoever caused arising directly or indirectly in connection with, in relation to or arisingout of the use of the Content.
This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden. Terms &

Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3

Transgenic plants for abiotic stress tolerance: current status
Syed Sarfraz Hussaina*, Hasnain Razaa, Irfan Afzalb andMahmood Akhtar Kayanic
aInstitute for Molecular Physiology and Biotechnology of Plants (IMBIO-Bartels), Universityof Bonn, Bonn, Germany; bDepartment of Crop Physiology, University of Agriculture,Faisalabad, Pakistan; cDepartment of Biosciences, COMSATS Institute of Information
Technology (CIIT), Islamabad, Pakistan
(Received 7 May 2010; final version received 1 November 2010)
Abiotic stress is one of the primary causes of crop losses worldwide (Bray et al.2000. Responses to abiotic stresses. In: Buchanana BB, Gruissem W, Jones RL,editors. Biochemistry and Molecular Biology of Plants. Rockville (MD):American Society of Plant Physiologists. p. 1158–1249). To cope with thedetrimental effects of stress, plants have evolved many biochemical and molecularmechanisms. One of the well-documented stress responses in plants is accumula-tion of osmolytes during stress. Although their actual roles in plant-stresstolerance remain controversial, these molecules are thought to have positiveeffects on enzyme and membrane integrity, along with adaptive roles in mediatingosmotic adjustment in plants grown under drought conditions. Recent studieshave demonstrated that the manipulation of genes involved in the biosynthesis ofthese osmolytes have improved plant tolerance to drought and salinity in anumber of crops. There is renewed hope of understanding the molecular basis ofosmolyte accumulation under stress and manipulating these processes via geneticengineering. For future work on generating transgenic plants with still higherlevels of tolerance, the new knowledge may be used via guided genetic engineeringof multiple genes to create crop plants with significantly increased productivityunder drought stress. This review surveys the current advances in engineeringabiotic stress-tolerant plants, particularly the genetic engineering of osmolytegenes (osmoprotectants) for imparting drought stress tolerance in plants.
Keywords: osmolytes; drought stress; transgenic plants; metabolic engineering
Introduction: a brief account of effects of drought on plants
Crop plants are affected by a variety of abiotic stresses, but drought represents someof the most significant constraints on agricultural productivity (Capell et al. 2004).Drought is the major abiotic stress severely affecting the distribution andproductivity of crop plants. For example, up to 45% of the world’s agriculturallands, wherein 38% of the world human population resides, are subjected tocontinuous or frequent drought (Hussain et al. 2008). Water is essential at everystage of plant growth from seed germination to plant maturation. A shortage ofwater is the single most important factor that reduces global crop yields, with far-reaching socio-economic implications (Castiglioni et al. 2008). Any degree of water
*Corresponding author. Email: [email protected]
Archives of Agronomy and Soil Science
Vol. 58, No. 7, July 2012, 693–721
ISSN 0365-0340 print/ISSN 1476-3567 online
� 2012 Taylor & Francis
http://dx.doi.org/10.1080/03650340.2010.540010
http://www.tandfonline.com
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3

imbalance may produce deleterious effects on crop growth and development. Plantsexhibit a variety of responses to environmental stresses that enable them to surviveor tolerate adverse conditions. Such plants respond to drought by making changes atthe cellular, metabolic and molecular levels to cope with the stress (Zhu 2001;Shinozaki and Yamaguchi-Shinozaki 2007). Although the general effects of droughton plant growth and development are fairly well known, the primary effects of waterdeficit at the cellular and molecular levels are not well understood (Zhu 2002;Chaitanya et al. 2003; Chaves and Oliveira 2004).
In order to improve the agricultural productivity within the limited landresources, it is imperative to ensure high crop yields against unfavorableenvironmental stresses such as drought. Three main approaches have been used inbreeding drought tolerant varieties: (1) select for high yield potential under optimalconditions; (2) select for maximum yield in target environments; and (3) manipulatethe genetic parameters of drought stress tolerance in the breeding and selectionschemes. Based on these approaches, the success in obtaining tolerant plants viaconventional breeding strategies has been useful in some cases to limited extent.However, in general, the success rate has not been encouraging in the case of droughtstress because of the multigenic nature of the stress. Therefore, researchers feel lessmotivated to employ these approaches, which are also time-consuming andlaborious (Bartels and Hussain 2008).
Currently, there are no economically viable technological means to facilitate cropproduction under stress conditions (Ashraf and Foolad 2007) using breedingmethods. However, the development of crop plants tolerant to drought stress isconsidered a promising approach, which may help satisfy growing food demandsfrom both developing and under-developed countries. By contrast, improvement ofstress tolerance by genetic engineering overcomes the bottlenecks of plant breedingmethods. Transgenic approaches can be used in combination with conventionalbreeding strategies to create crops with enhanced drought tolerance (Capell et al.2004). The current genetic engineering strategies rely on the transfer to the targetplant of one or several genes that are either involved in signaling and/or regulatorypathways, or that encode enzymes present in pathways leading to the synthesis offunctional and structural protectants, such as osmolytes and antioxidants, orpathways that encode stress tolerance-conferring proteins (Wang et al. 2003; Vinocurand Altman 2005; Valliyodan and Nguyen 2006; Bhatnagar-Mathur et al. 2007;Kathuria et al. 2007; Sreenivasulu et al. 2007).
Engineering plants with osmoprotectant pathways
Improving crop resistance to drought stress is a long-standing goal of agriculturalbiotechnology (McCue and Hanson 1990; Jain and Selvaraj 1997). Plants haveevolved various mechanisms to cope with stress conditions and these include theshifts in the physiology of the plant and the expression of stress-associated genes,leading to the formation of a wide variety of low molecular mass metabolitescollectively known as compatible solutes such as proline, glycine betaine, sugars suchas sucrose, trehalose and fructans and sugar alcohols like sorbitol, mannitol,ononitol, pinitol and polyols (Bohnert and Jensen 1996; Rajam et al. 1998; Bohnertand Shen 1999; Kumar et al. 2006). These osmolytes are uniformly neutral withrespect to the perturbation of cellular functions even when present at high
694 S.S. Hussain et al.
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3

concentrations (Yancy et al. 1982; Serraj and Sinclair 2002). Accumulation of thesemolecules either actively or passively helps plants to retain water within cells andprotects cellular compartments from injury caused by dehydration or maintainsturgor pressure during water stress (Table 1–5). Moreover, these molecules stabilizethe structure and function of certain macromolecules, signaling functions orinduction of adaptive pathways and scavenge reactive oxygen species (Hasegawaet al. 2000; Chen and Murata 2002). However, the molecular and cellularinteractions of these solutes are not completely understood.
To date, tolerance to abiotic stresses has mainly been achieved throughengineering for increased cellular levels of compatible solutes (Flowers 2004; KumarS et al. 2004; Chinnusamy et al. 2005). Another noteworthy point is that increasedlevels of solutes often have enhanced tolerance for drought, salt and cold stress at thesame time (Garg et al. 2002; Jang et al. 2003; Park et al. 2007), implying that geneticengineering by altering osmolytes is a fruitful approach for obtaining combinedtolerance to different abiotic stresses. In fact, simple, one-step transformationstrategies have been designed to increase the accumulation of these osmolytes,especially in plant species, in which these molecules do not accumulate naturally. Inmost cases, the introduction of a single foreign gene into a transgenic plant has led toa moderate increase in tolerance with modest accumulation of osmoprotectants.
In this review, we focus on results published in recent years regarding theinvolvement of osmolytes in improving plant stress tolerance via transgenicapproach.
Amino acid-derived osmoprotectants and their role in drought tolerance
Metabolic acclimation via the accumulation of osmolytes/compatible solutes isregarded as a basic strategy for the protection and survival of plants in extremeenvironments. Accumulation of compatible solutes is ubiquitous in plants and isoften referred to as osmoprotection. In stress-tolerant transgenic plants, manygenes involved in the synthesis of osmoprotectants, such as amino acids (proline),quaternary compounds (glycine betaine) and a variety of sugars and sugaralcohols (mannitol, trehalose and galactinol) that accumulate during stressadjustment, have been used (Chen and Murata 2002; Chaves and Oliveira2004; Vinocur and Altman 2005). Although there are many examples of aminoacid-derived osmoprotectants, to date, metabolic engineering studies have mainlyfocused on increasing proline and betaine accumulation (Holmberg and Bulow1998; McNeil et al. 1999).
Proline
Proline is an osmoprotecting molecule that accumulates in many organisms,including bacteria, fungi and plants, in response to drought and salinity (Kumaret al. 2003; Claussen 2005). Proline is one of the most commonly found compatibleosmolytes in water-stressed plants (Delauney and Verma 1993; Yoshiba et al. 1997).Pyrroline-5-carboxylate synthetase (P5CS) is encoded by a nuclear gene that hasbeen cloned from several different plants, for example, Vigna aconitifolia (Hu et al.1992), Arabidopsis thaliana (Strizhov et al. 1997), Medicago sativa (Ginzberg et al.1998), Lycopersicum esculentum (Fujita et al. 1998), Medicago truncatula (Armen-gaud et al. 2004) and Phaseolus vulgaris (Chen et al. 2009).
Archives of Agronomy and Soil Science 695
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3
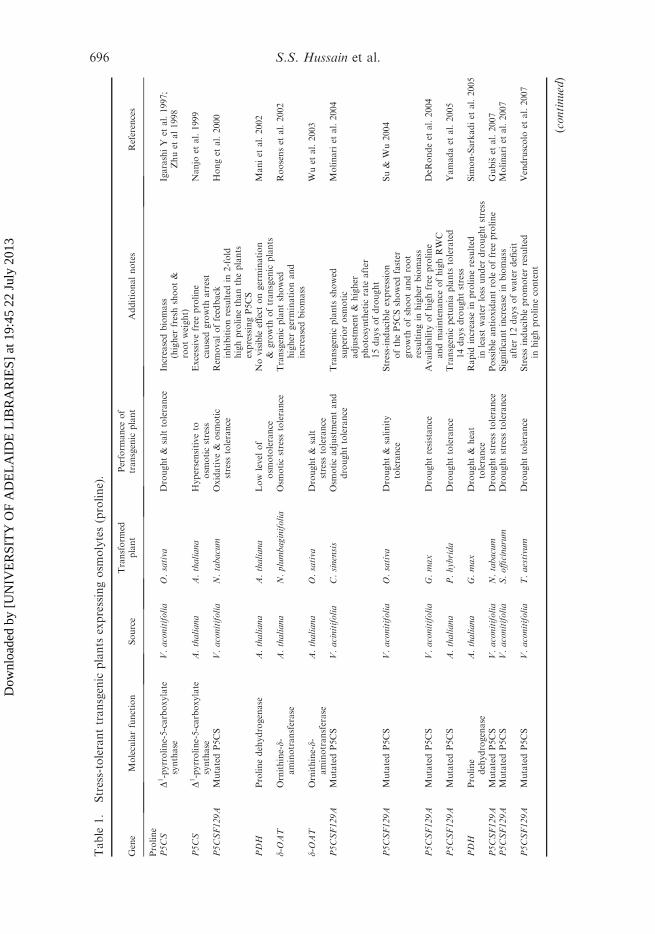
Table
1.
Stress-toleranttransgenic
plants
expressingosm
olytes(proline).
Gene
Molecularfunction
Source
Transform
edplant
Perform
ance
of
transgenic
plant
Additionalnotes
References
Proline
P5CS
D1-pyrroline-5-carboxylate
synthase
V.aconitifolia
O.sativa
Drought&
salttolerance
Increasedbiomass
(higher
freshshoot&
rootweight)
IgarashiY
etal.1997;
Zhuet
al1998
P5CS
D1-pyrroline-5-carboxylate
synthase
A.thaliana
A.thaliana
Hypersensitiveto
osm
oticstress
Excessivefree
proline
causedgrowth
arrest
Nanjo
etal.1999
P5CSF129A
MutatedP5CS
V.aconitifolia
N.tabacum
Oxidative&
osm
otic
stress
tolerance
Rem
ovaloffeedback
inhibitionresulted
in2-fold
highprolinethantheplants
expressingP5CS
Honget
al.2000
PDH
Prolinedehydrogenase
A.thaliana
A.thaliana
Low
level
of
osm
otolerance
Novisible
effectongermination
&growth
oftransgenic
plants
Maniet
al.2002
d-OAT
Ornithine-d-
aminotransferase
A.thaliana
N.plumbaginifolia
Osm
oticstress
tolerance
Transgenic
plantshowed
higher
germinationand
increasedbiomass
Roosenset
al.2002
d-OAT
Ornithine-d-
aminotransferase
A.thaliana
O.sativa
Drought&
salt
stress
tolerance
Wuet
al.2003
P5CSF129A
MutatedP5CS
V.acinitifolia
C.sinensis
Osm
oticadjustmentand
droughttolerance
Transgenic
plants
showed
superiorosm
otic
adjustment&
higher
photosynthetic
rate
after
15daysofdrought
Molinariet
al.2004
P5CSF129A
MutatedP5CS
V.aconitifolia
O.sativa
Drought&
salinity
tolerance
Stress-inducible
expression
oftheP5CSshowed
faster
growth
ofshootandroot
resultingin
higher
biomass
Su&
Wu2004
P5CSF129A
MutatedP5CS
V.aconitifolia
G.max
Droughtresistance
Availabilityofhighfree
proline
andmaintenance
ofhighRWC
DeR
ondeet
al.2004
P5CSF129A
MutatedP5CS
A.thaliana
P.hybrida
Droughttolerance
Transgenic
petunia
plants
tolerated
14daysdroughtstress
Yamadaet
al.2005
PDH
Proline
dehydrogenase
A.thaliana
G.max
Drought&
heat
tolerance
Rapid
increase
inprolineresulted
inleast
waterloss
under
droughtstress
Sim
on-Sarkadiet
al.2005
P5CSF129A
MutatedP5CS
V.aconitifolia
N.tabacum
Droughtstress
tolerance
Possible
antioxidantrole
offree
proline
Gubis
etal.2007
P5CSF129A
MutatedP5CS
V.aconitifolia
S.officinarum
Droughtstress
tolerance
Significantincrease
inbiomass
after
12daysofwaterdeficit
Molinariet
al.2007
P5CSF129A
MutatedP5CS
V.aconitifolia
T.aestivum
Droughttolerance
Stressinducible
promoterresulted
inhighprolinecontent
Vendruscolo
etal.2007
(continued)
696 S.S. Hussain et al.
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3
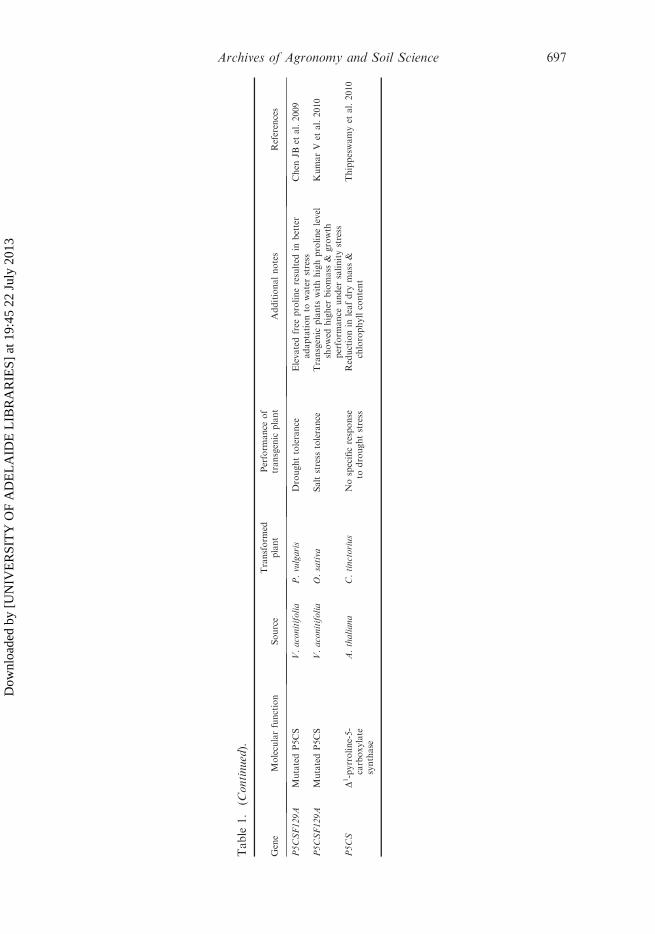
Table
1.
(Continued).
Gene
Molecularfunction
Source
Transform
edplant
Perform
ance
of
transgenic
plant
Additionalnotes
References
P5CSF129A
MutatedP5CS
V.aconitifolia
P.vulgaris
Droughttolerance
Elevatedfree
prolineresulted
inbetter
adaptationto
waterstress
Chen
JBet
al.2009
P5CSF129A
MutatedP5CS
V.aconitifolia
O.sativa
Saltstress
tolerance
Transgenic
plants
withhighprolinelevel
showed
higher
biomass
&growth
perform
ance
under
salinitystress
KumarV
etal.2010
P5CS
D1-pyrroline-5-
carboxylate
synthase
A.thaliana
C.tinctorius
Nospecificresponse
todroughtstress
Reductionin
leafdry
mass
&chlorophyllcontent
Thippeswamyet
al.2010
Archives of Agronomy and Soil Science 697
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3

Although the exact function of proline is still under investigation, prolineaccumulation can influence stress tolerance in multiple ways (Szabados and Savoure2009). Proline may serve as a hydroxyl radical scavenger (Smirnoff and Cumbes1989), thus reducing the acidity of the cell (Venekamp et al. 1989). In addition, itmay also function as an osmolyte (Kishor et al. 1995) and molecular chaperone,being able to protect protein integrity and enhance the activities of different enzymes(Iyer and Caplan 1998). Proline biosynthesis from the amino acid glutamate can beachieved by using the P5CS gene from mothbean (Vigna aconitifolia), which encodesa bifunctional enzyme containing the catalytic activities of g-glutamyl kinase andglutamic-g-semialdehyde dehydrogenase (Holmberg and Bulow 1998).
Several studies have used this gene to produce transgenic plants (Table 1) withincreased proline production and subsequent stress tolerance (Kishor et al. 1995;Zhu et al. 1998; Roosens et al. 2002; Sawahel and Hassan 2002; Verdoy et al. 2006).Mani et al. (2002) generated transgenic Arabidopsis containing either sense orantisense proline dehydrogenase (PDH). Transgenic plants carrying sense PDH didnot show significant levels of osmotolerance under stress conditions. However,exogenous application of proline increased the tolerance of sense transgenic plants toosmotic stress. Indeed, transgenic Arabidopsis plants with antisense (Mani et al.2002) or knockout (Nanjo et al. 2003) PDH resulted in increased prolineaccumulation and better growth performance under stress (Simon-Sarkadi et al.2005).
It has also been reported that transgenic petunia plants that overexpress P5CSgene from Arabidopsis and P5CS gene from rice can withstand drought conditionsfor longer than wild-type plants (Yamada et al. 2005). Also, the expression of P5CSresulted in drought tolerance in soybean (DeRonde et al. 2004). Similarly, Chen et al.(2009) noticed a higher transcript level (2.5 times) of PvP5CS gene in common beanunder drought. This showed that P5CS from common bean was a stress-induciblegene regulating the accumulation of proline in plants subjected to drought stress.Transgenic wheat plants expressing P5CS gene from Vigna aconitifolia under thecontrol of a stress-induced promoter complex AIPC resulted in enhanced prolineaccumulation as well as drought tolerance under water deficit for 15 days(Vendruscolo et al. 2007). The results of a series of experiments showed that prolinemight confer drought stress tolerance to transgenic wheat plants by increasing theantioxidant system rather than as osmotic adjustment mediator.
However, P5CS, a rate-limiting enzyme in proline biosynthesis, is subjected tofeedback inhibition by proline, and earlier reports suggested that prolineaccumulation in plants under stress might involve the loss of feedback regulationdue to a conformational change in the P5CS protein (Hong et al. 2000). Therefore,Hong et al. (2000) removed this feedback inhibition by site-directed mutagenesis andthe resulting gene P5CSF129A was recently used in two studies, which clearlydemonstrated the involvement of proline in response to drought and salt (Gubiset al. 2007; Chen et al. 2009). Transgenic tobacco carrying mutagenizedP5CSA129A gene showed no negative effects on plant growth or physiologyand accumulated 17–22-fold more proline than wild-type plants under controlconditions. Similarly, plants also exhibited drought tolerance by further elevatingproline content under drought stress (Gubis et al. 2007). However, despite this,there are only a few reports in which researchers have used this mutated versionof P5CS gene (Hong et al. 2000; Molinari et al. 2004; Bhatnagar-Mathur et al.2009; Kumar et al. 2010).
698 S.S. Hussain et al.
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3

Improvement in the drought or salt tolerance of crop plants via engineeringproline metabolism is a current possibility (Vendruscolo et al. 2007; Yamchi et al.2007; Bhatnagar-Mathur et al. 2009; Kumar et al. 2010) and should be exploredmore extensively. However, further research is required to study the feasibility ofengineering stress tolerance in plants via targeted engineering of proline metabolism.
Glycine betaine
Glycine betaine (GB) is a quaternary ammonium compound that occurs in a varietyof plants, animals and microorganisms (Rhodes and Hanson 1993). GB accumulatesin the chloroplasts and plastids of many halotolerant plants and increases thetolerance of such plants to various types of abiotic stresses (Chen and Murata 2008).The physiological role of GB in alleviating osmotic stress was proposed on the basisof its enhanced accumulation in some stressed plants (Wyn Jones 1984; Sakamotoand Murata 2001). This compound is one of several such compatible solutes(Dorffling et al. 1990) that have an osmoprotection function. These solutes are alsoknown to protect proteins and enzyme activities under water deficits (Mohanty et al.2002; Ashraf and Foolad 2007), and even to stabilize membranes during freezing(Zhao et al. 1992; Rhodes and Hanson 1993; Papageorgiou and Murata 1995). Thecryoprotective effect of GB appears to come from its compatibility withmacromolecular structure and function (Xing and Rajashekar 2001). It has beensuggested that glycine betaine can help stabilize the protein tertiary structure andprevent or reverse disruption of the tertiary structure of these proteins caused bynon-compatible (perturbing) solutes. Based upon this observation, it has beensuggested that GB has a chaperone function (Sakamoto and Murata 2000; Chen andMurata 2002).
GB-encoding genes have been cloned from various microorganisms and plants,and transgenic plants of various species have been produced that express one orseveral of these genes (Sakamoto et al. 1998; Nuccio et al. 1998; Huang et al. 2000;McNeil et al. 2000, 2001; Mohanty et al. 2002; Shen et al. 2002). A moderate stresstolerance was noted in some, but not all, such GB-producing transgenic lines, withvariable responses to drought, salinity and freezing (Table 2). In some cases, theexogenous supply of GB helps the engineered compound to improve the stresstolerance of the plant (Quan et al. 2004b) by regulating expression of specific stress-responsive genes and by blocking reactive oxygen species (ROS) accumulationduring stress, for example, chilling stress (Einset et al. 2007; 2008; Einset andConnolly 2009). Waditee et al. (2003) found an alternative biosynthetic pathway ofbetaine from glycine, which is catalyzed by two N-methyltransferase enzymes.Overexpression of these enzymes in Arabidopsis resulted in the accumulation ofsignificant level of GB (Waditee et al. 2005).
Despite the encouraging results regarding the effect of GB accumulation ondrought tolerance in transgenic plants, poor accumulation of GB has been observedin some instances due to a limited choline supply (Nuccio et al. 1998) and impairedimport of choline into the chloroplast (Nuccio et al. 2000). Many agronomicallyimportant crops such as wheat, potato and tomato are non-accumulators of GB andare therefore, potential targets for engineering GB biosynthesis to increase theirstress tolerance via the expression of GB biosynthesis genes. (Bartels and Hussain2008). Shirasawa et al. (2006) reported that transgenic rice plants expressing spinachcholine monooxygenase (CMO) accumulated GB at lower levels than those reported
Archives of Agronomy and Soil Science 699
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3
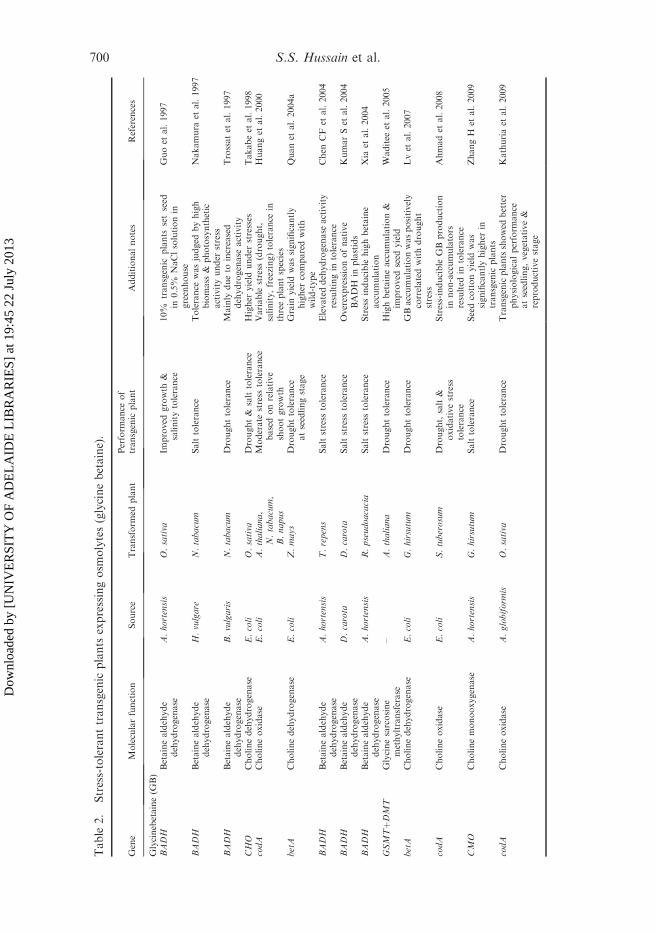
Table
2.
Stress-toleranttransgenic
plants
expressingosm
olytes(glycinebetaine).
Gene
Molecularfunction
Source
Transform
edplant
Perform
ance
of
transgenic
plant
Additionalnotes
References
Glycinebetaine(G
B)
BADH
Betainealdehyde
dehydrogenase
A.hortensis
O.sativa
Improved
growth
&salinitytolerance
10%
transgenic
plants
setseed
in0.5%
NaClsolutionin
greenhouse
Guoet
al.1997
BADH
Betainealdehyde
dehydrogenase
H.vulgare
N.tabacum
Salttolerance
Tolerance
wasjudged
byhigh
biomass
&photosynthetic
activityunder
stress
Nakamura
etal.1997
BADH
Betainealdehyde
dehydrogenase
B.vulgaris
N.tabacum
Droughttolerance
Mainly
dueto
increased
dehydrogenase
activity
Trossatet
al.1997
CHO
Cholinedehydrogenase
E.coli
O.sativa
Drought&
salttolerance
Higher
yield
under
stresses
Takabeet
al.1998
codA
Cholineoxidase
E.coli
A.thaliana,
N.tabacum,
B.napus
Moderate
stress
tolerance
basedonrelative
shootgrowth
Variable
stress
(drought,
salinity,freezing)tolerance
inthreeplantspecies
Huanget
al.2000
betA
Cholinedehydrogenase
E.coli
Z.mays
Droughttolerance
atseedlingstage
Grain
yield
wassignificantly
higher
comparedwith
wild-type
Quanet
al.2004a
BADH
Betainealdehyde
dehydrogenase
A.hortensis
T.repens
Saltstress
tolerance
Elevateddehydrogenase
activity
resultingin
tolerance
Chen
CFet
al.2004
BADH
Betainealdehyde
dehydrogenase
D.carota
D.carota
Saltstress
tolerance
Overexpressionofnative
BADH
inplastids
KumarSet
al.2004
BADH
Betainealdehyde
dehydrogenase
A.hortensis
R.pseudoacacia
Saltstress
tolerance
Stressinducible
highbetaine
accumulation
Xia
etal.2004
GSMTþDMT
Glycinesarcosine
methyltransferase
–A.thaliana
Droughttolerance
Highbetaineaccumulation&
improved
seed
yield
Waditee
etal.2005
betA
Cholinedehydrogenase
E.coli
G.hirsutum
Droughttolerance
GBaccumulationwaspositively
correlatedwithdrought
stress
Lvet
al.2007
codA
Cholineoxidase
E.coli
S.tuberosum
Drought,salt&
oxidativestress
tolerance
Stress-inducible
GB
production
innon-accumulators
resulted
intolerance
Ahmadet
al.2008
CMO
Cholinemonooxygenase
A.hortensis
G.hirsutum
Salttolerance
Seedcottonyield
was
significantlyhigher
intransgenic
plants
ZhangH
etal.2009
codA
Cholineoxidase
A.globiform
isO.sativa
Droughttolerance
Transgenicplantsshowed
better
physiologicalperform
ance
atseedling,vegetative&
reproductivestage
Kathuriaet
al.2009
700 S.S. Hussain et al.
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3

by Sakamoto et al. (1998) and Mohanty et al. (2002). The evidence for a limitedinternal choline supply pointed to the need to enhance choline synthesis (Nuccioet al. 1998).
Recently, two independent studies addressed the role of GB in non-accumulatingplants. Lv et al. (2007) investigated the high GB accumulation under stress in fivehomozygous transgenic cotton lines at three development stages. The resultsdemonstrated that transgenic plants accumulated higher GB levels than wild-typeplants either before or after drought stress. Similarly, high GB levels in transgenicplants were found to be positively correlated with drought tolerance. Incontinuation, transgenic potato plants expressing a bacterial choline oxidase (betA) gene under the control of an oxidative stress-inducible SWPA2 promoteraccumulated high levels of GB under drought stress (Ahmad et al. 2008). In additionto high GB levels, transgenic plants also maintained high water content andaccumulated high levels of vegetative biomass compared with non-transgenic (NT)plants under drought. These results suggested that GB not only protects cellmembranes and enzymes under water stress, but is also involved in osmoticadjustment in transgenic plants. By contrast, the effectiveness of stress tolerance alsodepends on the cellular compartment in which GB accumulates, as shown in the caseof transgenic tomato plants in which chloroplastic accumulation is more effectivethan cytosolic accumulation (Park et al. 2007; Ahmad et al. 2008). Similarly, theprotective effects of GB in the reproductive organs of plants have been observed inArabidopsis, maize and tomato under different abiotic stresses (Sakamoto et al. 2000;Park et al. 2004; Quan et al. 2004a, 2004b). This protection might have beenmediated by the high levels of GB that accumulate in reproductive organs as aconsequence of translocation from other organs, such as leaves. These resultsrevealed that the accumulation of GB in reproductive organs can effectively protectplants from reproductive failure under stress.
Sugars and sugar alcohols and their role in drought tolerance
Accumulation of sugar-related compounds, such as sucrose, raffinose, sorbitol andfructans, has been observed repeatedly in response to osmotic stress in a variety oforganisms. Many of these compounds are believed to stabilize the membranes andproteins during dehydration. According to Crowe et al. (1992), sugars can replacethe water molecules and thus stabilize the proteins or membranes in a physical statesimilar to that in the presence of water molecules. An alternative function has beenproposed for the disaccharides sucrose and trehalose, which can form a glass phasein the dry state (Crowe et al. 1998). A glass is a liquid of high viscosity that is capableof slowing down chemical reactions, which could lead to long-term stability in aliving system (Bruni and Leopold 1992).
Trehalose
Accumulation of trehalose, a rare non-reducing sugar, occurs in many bacteria,yeast, fungi and in a few desiccation-tolerant plants, so-called resurrection plants.During desiccation stress, trehalose protects the biological molecules in response todifferent stress conditions, but it does not accumulate to high enough levels in mostplants, probably because of the presence of trehalase activity (Goddijn andSmeekens 1998; Singer and Lindquist 1998; Goddijn and van Dun 1999; Iturriaga
Archives of Agronomy and Soil Science 701
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3

et al. 2000). Trehalase contributes to trehalose degradation (Paul et al. 2008) and inthe presence of a trehalase inhibitor (validamycin A), trehalose accumulates toseveral fold in concentration (Goddijn and van Dun 1999). Trehalose acts as astorage carbohydrate and possesses the unique feature of reversible water-absorptionthat results in protection of the biological molecules from desiccation-induceddamage; this sugar appears to be superior to other sugars in conferring protection todamage (Rontein et al. 2002).
Trehalose is synthesized in two steps from glucose-6-phosphate and uridinediphosphoglucose, via trehalose-6-phosphate. The first step is catalyzed by trehalosephosphate synthase (TPS), and the second by trehalose-6-phosphatase (TPP)(Bartels and Hussain 2008).
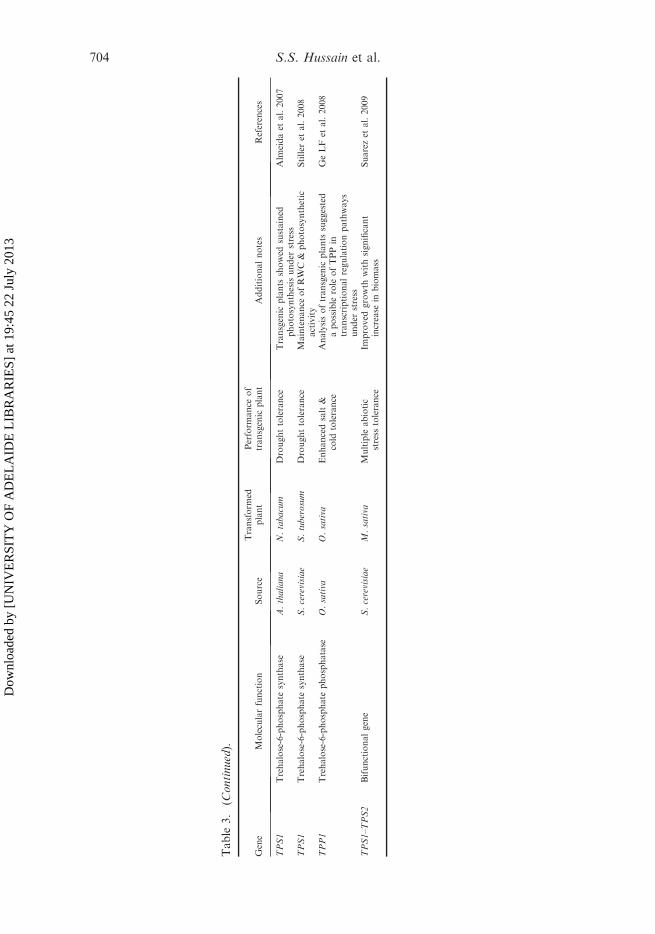
Multiple studies have linked trehalose to stress tolerance in plants (Table 3). Thegenes encoding the trehalose biosynthesis enzymes from different sources weretransferred into plants to obtain transgenic plants with relatively higher levels oftrehalose. Different groups have generated stress-tolerance plants by transferringmicrobial trehalose biosynthetic genes in tobacco (Holmstrom et al. 1996; Goddijnet al. 1997; Romero et al. 1997; Pilon-Smits et al. 1998; Lee et al. 2003; Han et al.2005; Karim et al. 2007), rice (Garg et al. 2002; Jang et al. 2003), tomato (Cortinaand Culianez-Macia 2005), potato (Goddijn et al. 1997), and Arabidopsis (Karimet al. 2007; Miranda et al. 2007). Tobacco plants overexpressing the yeast trehalosephosphate synthase gene (TPS1) showed significant tolerance to drought. However,the plants displayed reduced growth under non-stress conditions and multiplephenotypic alterations (Holmstrom et al. 1996; Romero et al. 1997). Tobacco plantstransformed with bacterial OtsA (TPS1) genes have a greater ability to retain water,whereas plants with OtsB (TPP) have reduced levels of photosynthesis. Both types oftransgenic tobacco plants accumulate only a small amount of trehalose byconstitutively expressing the genes (Pilon-Smits et al. 1998). Although plants withimproved stress tolerance were obtained (Table 3), no direct correlation wasobserved between trehalose level and stress tolerance (Bartels and Hussain 2008).This discrepancy and the observation that modification of the trehalose levels oftenproduced altered growth and morphogenic phenotypes led to the discovery thattrehalose-6-phosphate, a metabolic intermediate of trehalose synthesis, is a signalingmolecule regulating sugar and starch metabolism, which links these pathways toother metabolic networks like photosynthesis and hormonal homeostasis (Goddijnand van Dun 1999; Paul 2007). Nevertheless, subsequent studies circumvent thisproblem by using a microbial TPS–TPP fusion gene in tobacco under the control ofthe Arabidopsis ribulose-1,5-bisphosphate carboxylase (RuBisCO) promoter. Thiseliminates the growth problem, while retaining the improved drought tolerance(Karim et al. 2007). A new development in this direction has been the over-production of trehalose using stress-inducible or tissue-specific expression of abifunctional trehalose-6-phosphate synthase/phosphatase (TPSP) fusion gene (OtsAand OtsB) in rice transgenic plants (Garg et al. 2002). Such transgenic rice plantsexhibited increased tolerance to drought, salt and low-temperature stresses. Inaddition, these transgenic plants also exhibited improved photosystem function andincreased photosynthetic activity under non-stressed conditions.
Recently, Alfalfa transgenic plants expressing bifunctional TPS–TPP enzymeexhibited tolerance to multiple abiotic stresses, however if the CaMV 35S promoterwas used the plants were moderately stunted. By contrast, expression under RD29Astress-inducible promoter led to large plants with significant increase in biomass
702 S.S. Hussain et al.
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3
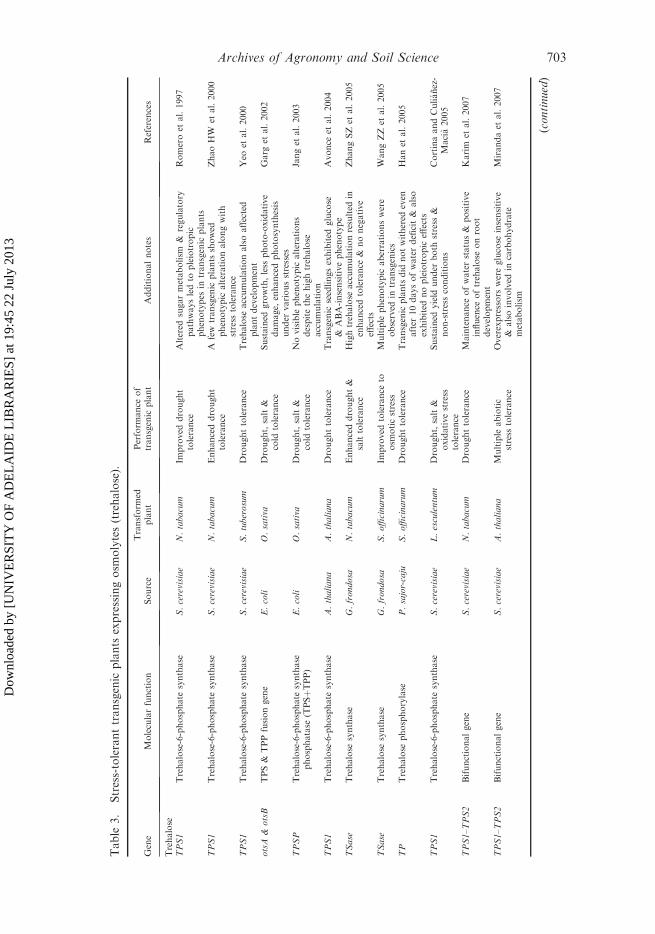
Table
3.
Stress-toleranttransgenic
plants
expressingosm
olytes(trehalose).
Gene
Molecularfunction
Source
Transform
edplant
Perform
ance
of
transgenic
plant
Additionalnotes
References
Trehalose
TPS1
Trehalose-6-phosphate
synthase
S.cerevisiae
N.tabacum
Improved
drought
tolerance
Altered
sugarmetabolism
®ulatory
pathwaysledto
pleiotropic
phenotypes
intransgenic
plants
Romeroet
al.1997
TPS1
Trehalose-6-phosphate
synthase
S.cerevisiae
N.tabacum
Enhanceddrought
tolerance
Afew
transgenic
plants
showed
phenotypic
alterationalongwith
stress
tolerance
ZhaoHW
etal.2000
TPS1
Trehalose-6-phosphate
synthase
S.cerevisiae
S.tuberosum
Droughttolerance
Trehalose
accumulationalsoaffected
plantdevelopment
Yeo
etal.2000
otsA
&otsB
TPS&
TPPfusiongene
E.coli
O.sativa
Drought,salt&
cold
tolerance
Sustained
growth,less
photo-oxidative
damage,
enhancedphotosynthesis
under
variousstresses
Garg
etal.2002
TPSP
Trehalose-6-phosphate
synthase
phosphatase
(TPSþTPP)
E.coli
O.sativa
Drought,salt&
cold
tolerance
Novisible
phenotypic
alterations
despitethehightrehalose
accumulation
Janget
al.2003
TPS1
Trehalose-6-phosphate
synthase
A.thaliana
A.thaliana
Droughttolerance
Transgenic
seedlingsexhibited
glucose
&ABA-insensitivephenotype
Avonce
etal.2004
TSase
Trehalose
synthase
G.frondosa
N.tabacum
Enhanceddrought&
salttolerance
Hightrehalose
accumulationresulted
inenhancedtolerance
&nonegative
effects
ZhangSZ
etal.2005
TSase
Trehalose
synthase
G.frondosa
S.officinarum
Improved
tolerance
toosm
oticstress
Multiple
phenotypic
aberrationswere
observed
intransgenics
WangZZ
etal.2005
TP
Trehalose
phosphorylase
P.sajor-caju
S.officinarum
Droughttolerance
Transgenicplants
did
notwithered
even
after
10daysofwaterdeficit&
also
exhibited
nopleiotropic
effects
Hanet
al.2005
TPS1
Trehalose-6-phosphate
synthase
S.cerevisiae
L.esculentum
Drought,salt&
oxidativestress
tolerance
Sustained
yield
under
both
stress
&non-stressconditions
CortinaandCulianez-
Macia2005
TPS1–TPS2
Bifunctionalgene
S.cerevisiae
N.tabacum
Droughttolerance
Maintenance
ofwaterstatus&
positive
influence
oftrehalose
onroot
development
Karim
etal.2007
TPS1–TPS2
Bifunctionalgene
S.cerevisiae
A.thaliana
Multiple
abiotic
stress
tolerance
Overexpressors
wereglucose
insensitive
&alsoinvolved
incarbohydrate
metabolism
Mirandaet
al.2007
(continued)
Archives of Agronomy and Soil Science 703
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3
Table
3.
(Continued).
Gene
Molecularfunction
Source
Transform
edplant
Perform
ance
of
transgenic
plant
Additionalnotes
References
TPS1
Trehalose-6-phosphate
synthase
A.thaliana
N.tabacum
Droughttolerance
Transgenic
plants
showed
sustained
photosynthesisunder
stress
Alm
eidaet
al.2007
TPS1
Trehalose-6-phosphate
synthase
S.cerevisiae
S.tuberosum
Droughttolerance
Maintenance
ofRWC&
photosynthetic
activity
Stiller
etal.2008
TPP1
Trehalose-6-phosphate
phosphatase
O.sativa
O.sativa
Enhancedsalt&
cold
tolerance
Analysisoftransgenic
plants
suggested
apossible
role
ofTPPin
transcriptionalregulationpathways
under
stress
GeLF
etal.2008
TPS1–TPS2
Bifunctionalgene
S.cerevisiae
M.sativa
Multiple
abiotic
stress
tolerance
Improved
growth
withsignificant
increase
inbiomass
Suarezet
al.2009
704 S.S. Hussain et al.
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3

(Suarez et al. 2009). Subsequently, it should also be possible to modulate theincreased trehalose accumulation by downregulating the plant trehalase activity orby expressing the trehalose biosynthetic genes under tissue- or stress-specificregulation (Penna 2003; Stiller et al. 2008). Tobacco chloroplasts transformed with(Saccharomyces cerevisiae) ScTPS1 have generated plants with enhanced droughttolerance without the growth defects observed in nuclear-transformed ScTPS1transgenic plants. In such plants, trehalose accumulation was almost 25-fold higherin chloroplast of the transgenic plants compared with the nuclear transgenic plants(Lee et al. 2003). Tobacco plants transformed with trehalose synthase (TSase) fromGrifola frondosa Fr. showed no growth retardation, but rather enhanced droughtand salt tolerance (Zhang et al. 2005). Similarly, Han et al. (2005) used a differenttype of trehalose biosynthetic gene (trehalose phosphorylase) that bypasses thetrehalose-6-phosphate.
To date, the introduction of trehalose biosynthetic genes into several plants hasimproved their drought tolerance without having any negative effect on plant growthor grain yield (Pilon-Smits et al. 1999; Abebe et al. 2003). Miranda et al. (2007)reported construction and overexpression of the TPS1–TPS2 gene fusion inArabidopsis. Their results demonstrated that the transgenic Arabidopsis plantsshowed a significant increase in drought, freezing, salt and heat tolerance, with nosignificant morphological or growth alterations. Recently, Suzuki et al. (2008)revealed the involvement of yet another TPS, AtTPS5, in plant stress tolerance (heatstress). Although using trehalose to confer protection against multiple abioticstresses could potentially be of great use in improving the abiotic stress tolerance ofstress-sensitive crops, our understanding of how it interacts and acts upon stress isfar from complete. Hence, the regulatory network connected with trehalosemetabolism needs to be studied in detail before genetically engineering trehaloselevels can be used in practice.
Fructans and other carbohydrates
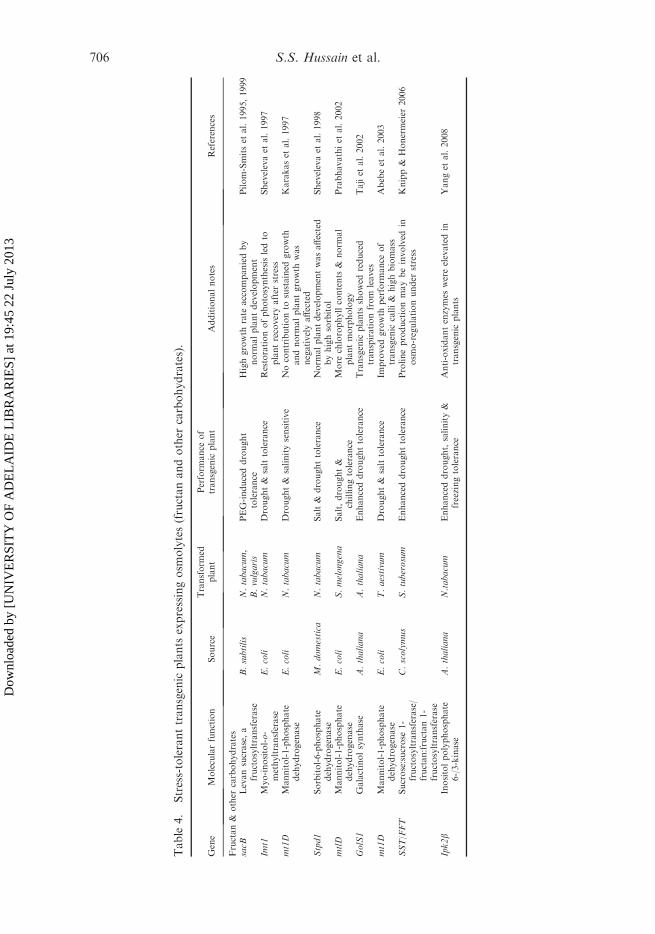
Fructan or polyfructose molecules serve as the main storage carbohydrate in manyplant species. Fructan-accumulating plants are globally distributed in temperateclimate zones, which are characterized by seasonal drought, cold or frost; such plantsare almost absent in the tropical regions (Hendry and Wallace 1993). In plants, thephysiological role of fructans is not clear and constitutes one of the most activeresearch areas involving transgenic fructan production (Livingston et al. 2009).However, fructans are thought to be involved in abiotic stress tolerance by virtue oftheir presence in vacuoles (Pilon-Smits et al. 1995). Fructans are also thought toprotect plants against drought and cold stress (Wiemken et al. 1995); evidence of thiswas derived from transgenic plants (Table 4).
To date, the SacB gene (encoding Bacillus subtilis levansucrase) has been usedmost extensively in such studies. Tobacco plants transformed with a bacterial genefor the fructan biosynthesis were more resistant to polyethylene glycol (PEG)-induced drought stress, as determined by changes in their growth properties andbiomass accumulation (Ebskamp et al. 1994; Pilon-Smits et al. 1995). Thesetransgenic plants performed better than controls under drought conditions,demonstrating rapid growth with increased fresh and dry weight accumulation(Chen and Murata 2002). Similarly, transgenic sugarbeet plants had significantlybetter growth statistics than wild-type plants under drought stress (Pilon-Smits et al.
Archives of Agronomy and Soil Science 705
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3
Table
4.
Stress-toleranttransgenic
plants
expressingosm
olytes(fructanandother
carbohydrates).
Gene
Molecularfunction
Source
Transform
edplant
Perform
ance
of
transgenic
plant
Additionalnotes
References
Fructan&
other
carbohydrates
sacB
Levansucrase,a
fructosyltransferase
B.subtilis
N.tabacum,
B.vulgaris
PEG-induceddrought
tolerance
Highgrowth
rate
accompaniedby
norm
alplantdevelopment
Pilom-Smitset
al.1995,1999
Imt1
Myo-inositol-o-
methyltransferase
E.coli
N.tabacum
Drought&
salttolerance
Restorationofphotosynthesisledto
plantrecoveryafter
stress
Shevelevaet
al.1997
mt1D
Mannitol-1-phosphate
dehydrogenase
E.coli
N.tabacum
Drought&
salinitysensitive
Nocontributionto
sustained
growth
andnorm
alplantgrowth
was
negativelyaffected
Karakaset
al.1997
Stpd1
Sorbitol-6-phosphate
dehydrogenase
M.domestica
N.tabacum
Salt&
droughttolerance
Norm
alplantdevelopmentwasaffected
byhighsorbitol
Shevelevaet
al.1998
mtlD
Mannitol-1-phosphate
dehydrogenase
E.coli
S.melongena
Salt,drought&
chillingtolerance
More
chlorophyllcontents
&norm
al
plantmorphology
Prabhavathiet
al.2002
GolS1
Galactinolsynthase
A.thaliana
A.thaliana
Enhanceddroughttolerance
Transgenic
plants
showed
reduced
transpirationfrom
leaves
Tajiet
al.2002
mt1D
Mannitol-1-phosphate
dehydrogenase
E.coli
T.aestivum
Drought&
salttolerance
Improved
growth
perform
ance
of
transgenic
calli&
highbiomass
Abebeet
al.2003
SST/FFT
Sucrose:sucrose
1-
fructosyltransferase/
fructan:fructan1-
fructosyltransferase
C.scolymus
S.tuberosum
Enhanceddroughttolerance
Prolineproductionmaybeinvolved
inosm
o-regulationunder
stress
Knipp&
Honermeier
2006
Ipk2b
Inositolpolyphosphate
6-/3-kinase
A.thaliana
N.tabacum
Enhanceddrought,salinity&
freezingtolerance
Anti-oxidantenzymes
wereelevatedin
transgenic
plants
Yanget
al.2008
706 S.S. Hussain et al.
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3

1999). It is still unclear exactly how the fructans contribute to the stress tolerance ofthe transformed plants. It is speculated that, like other sugars, fructans act asregulators or signal molecules, thus influencing plant metabolism, as scavengers ofROS (Shen et al. 1997). However, it seems to be clear that fructans promote theprocess of root branching, thus increasing the root surface and subsequent wateruptake (Datta 2002).
Drought stress also affects the metabolism of other important carbohydrates inplants. In addition to fructans, the raffinose family oligosaccharides (RFO) such asraffinose, stachyose and galactinol play important roles in drought stress tolerance inplants and seeds (Seki et al. 2007). Seven galactinol synthase (GolS)-related geneshave been characterized in the Arabidopsis genome, but information on the roles ofthese genes in conferring drought stress tolerance to these plants is still missing (Tajiet al. 2002; Valliyodan and Nguyen 2006). Transgenic Arabidopsis plants, whichoverexpress the drought-inducible galactinol synthase genes (AtGolS1/AtGolS2)exhibited enhanced drought tolerance because of the accumulation of galactinol andraffinose molecules (Taji et al. 2002). In addition, DREB1A/CBF3-expressingtransgenic plants showed tolerance to drought and cold owing to their accumulationof more galactinol and raffinose than non-transgenic plants (Taji et al. 2002; Sekiet al. 2007). These studies emphasize the importance of galactinol and raffinose instress tolerance and demonstrate that the modification of drought stress tolerance bymanipulation of these sugars is a viable alternative in plants.
Polyols
The accumulation of sugar alcohol/polyols is related to drought and salinity stresstolerance in many plant species (Bohnert and Jensen 1996). Some plants (Tobaccoand Arabidopsis) are non-accumulators of these compounds (e.g. mannitol),therefore, accumulation of these sugar alcohols such as mannitol (Tarczynskiet al. 1992; Thomas et al. 1995; Shen et al. 1997; Karakas et al. 1997), D-ononitol(Vernon et al. 1993; Sheveleva et al. 1997), inositol (Smart and Flores 1997) andsorbitol (Tao et al. 1995; Sheveleva et al. 1998) constitute another class ofcompounds with osmoprotective activities that have been the target of molecularengineering studies in plants (Table 4). However, it should be kept in mind thattransgenic Arabidopsis plants accumulating inositol did not show any significantdifference in stress tolerance, but the transgenic plants accumulating mannitol, D-ononitol and sorbitol had enhanced tolerance to drought or salt stress. Transgenictobacco plants expressing IMT1 gene coding for the myo-inositol-o-methyltransfer-ase enzyme were more drought and salt tolerant than wild-type tobacco plants(Sheveleva et al. 1997). Similarly, inositol phosphates (IPs) have been implicated inimportant roles in stress signaling. Many genes encoding inositol polyphosphatekinases have been identified but their physiological functions have not been fullyunderstood. Recently, Yang et al. (2008) showed that constitutive expression ofArabidopsis IPK2b gene in transgenic tobacco enhanced its tolerance towardsvarious abiotic stresses including drought, salt and freezing stresses. Although theprecise mode of action of AtIPK2b in plant responses to stress remains elusive, theseresults provided direct evidence that tolerance to various abiotic stresses can beachieved by alteration of AtIPK2b expression in transgenic plants. In addition,mannitol-producing tobacco plants exhibited increased tolerance to high salinity.Mannitol-expressing seeds of Arabidopsis were able to germinate in media with high
Archives of Agronomy and Soil Science 707
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3

concentrations of NaCl, compared with control seeds, which ceased to germinate ata much lower NaCl concentration (Thomas et al. 1995). Expression of the MT1Denzyme in the chloroplasts of the tobacco plants confers enhanced resistance tooxidative stress induced by the presence of methyl viologen (Shen et al. 1997). Suchresistance was attributed to an increased capacity to scavenge the hydroxyl radicals.Similarly, when the MT1D gene was engineered in wheat, it improved tolerance todrought and salinity stress in wheat primarily through osmotic adjustment (Abebeet al. 2003). However, the amount of accumulated mannitol was too small to protectagainst stress through osmotic adjustment. Therefore, the osmoprotective role ofthese compounds cannot be attributed solely to mere osmotic adjustment; they mayalso protect plants against damage by scavenging ROS and thus maintaining proteinstructure and function (Wang et al. 2003).
Studies involving the transfer of genes involved in the biosynthetic pathway ofpolyols were performed during early attempts to produce stress-tolerant plants.Unfortunately, engineering some of these compounds had a number of undesirableeffects. One such problem was observed when a new pathway for sorbitol synthesiswas engineered in tobacco plants, which resulted in growth defects and tissuenecrosis (Sheveleva et al. 1998). This study is a good example of a case in which acompetitive or antagonistic effect is elicited between the transgene activity and thehost plant metabolism. A reasonable explanation for the negative effects of polyolsin transgenic plants could be that a competition is set up for one or the same productbetween the host and the transgene.
Another case in which a positive effect is generated is that of the engineering of anew pathway for D-ononitol accumulation in the tobacco plants. In this case, D-ononitol production has been engineered to be stress dependent and thetransformants recovered faster than wild-type plants after rewatering (Shevelevaet al. 1997). These studies highlight the importance of conducting the metabolicanalyses of the transgenic stressed plants, in order to predict the in vivo pathwayfluxes and the pool sizes of the intermediate compounds (Nuccio et al. 1999). Oncesuch data has been obtained it could be incorporated into the metabolic models andcan be used to estimate the impact on other pathways when a transgene is insertedinto the plant of interest.
Polyamines and their role in drought tolerance
In plants the polyamines (PA), mainly diamine putrescine (Put), triamine spermidine(Spd) and tetraamine spermine (Spm), are the major components. These polyaminesare not only involved in the fundamental cellular processes, for example cell division,morphogenesis, chromatin organization, mRNA translation, ribosome biogenesis,cell proliferation, secondary metabolism, senescene and programmed cell dealth(Igarashi and Kashiwagi 2000; Thomas T and Thomas TJ 2001; Davies 2004; Kuehnand Phillips 2005; Kumar et al. 2006), but also play a pivotal role in plant responsesto abiotic and biotic stresses (Rajam 1997; Bouchereau et al. 1999; Galston 2001;Walters 2003; Ma et al. 2005; Bhattacharya and Rajam 2006; Kumar et al. 2006; Liuet al. 2006; Yamaguchi et al. 2007). A general phenomenon observed is that the PAscan alter their titers in response to various types of environmental stresses such aswater stress (Capell et al. 2004; Kasukabe et al. 2004; Ma et al. 2005), low and hightemperatures (Song et al. 2002; Hummel et al. 2004; Imai et al. 2004) and salinity(Maiale et al. 2004; Roy et al. 2005; Liu et al. 2006). Genes encoding these PA
708 S.S. Hussain et al.
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3

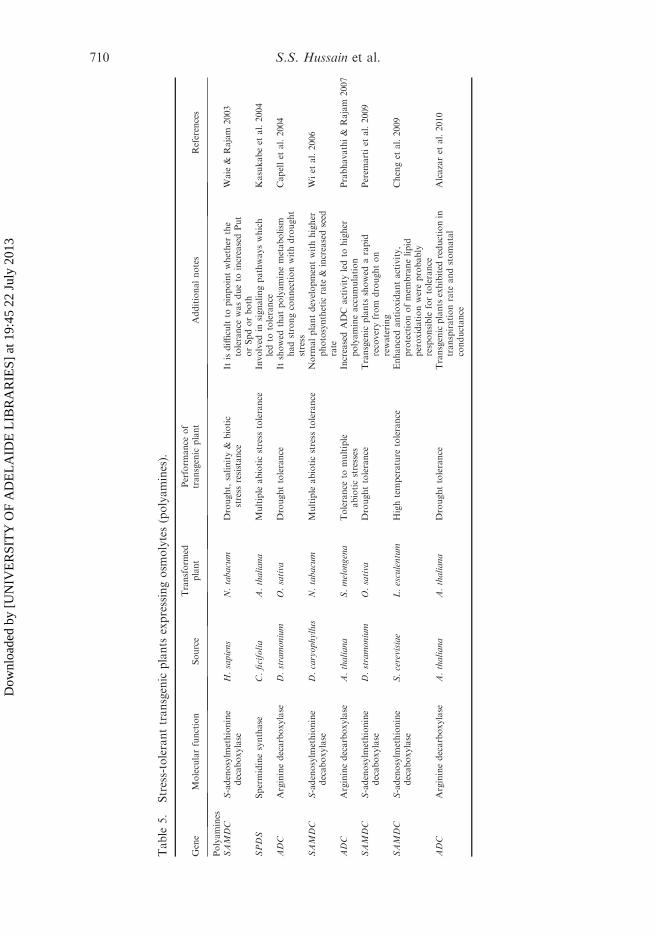
biosynthetic enzymes have been isolated from a variety of plant species (Malmberget al. 1998; Bagni and Tassoni 2001). A recent study with Arabidopsis mutant plantshas demonstrated that the cellular polyamine content often changes in response toabiotic stresses; however, its physiological relevance is unknown. It was alsoobserved that the PA biosynthesis is enhanced in drought-stressed plants(Bouchereau et al. 1999) and Spm content also increases, which suggests a role forthe Spm in drought stress tolerance (Yamaguchi et al. 2007). In the model plantArabidopsis, all of the pathway genes have been identified, these are: ADC-encodinggenes (ADC1 and ADC2, Watson and Malmberg 1996; Watson et al. 1997); theagmatine iminohydrolase gene (Janowitz et al. 2003); the N-carbamoylputrescineamidohydrolase gene and four genes for AdoMetDC (Franceschetti et al. 2001; Ge Cet al. 2006); two genes (SPDS1 and SPDS2) of the Spd synthase enzyme (Hanzawaet al. 2002); and two genes (ACL5 and SPMS) of the Spm synthase enzyme(Hanzawa et al. 1997, 2000). It has been reported that overexpression of PAbiosynthetic genes like the ADC (Capell et al. 1998, 2004; Roy and Wu 2001), ODC(Kumria and Rajam 2002), SAMDC (Roy and Wu 2002; Waie and Rajam 2003) andSPD SYN (Franceschetti et al. 2004; Kasukabe et al. 2004, 2006) in plants like rice,tobacco, Arabidopsis and sweet potato, has resulted in increased tolerance to variousabiotic stresses.
There have been several reports on creating transgenic plants harboring thepolyamine biosynthetic genes in an attempt to enhance stress tolerance (Table 5).For example, in a recent study, transgenic rice plants expressing the Daturastramonium ADC (arginine decarboxylase) gene produced higher levels of Put underdrought stress than wild-type plants, which led to higher levels of Spd and Spm andimproved the drought tolerance, owing to which the transgenic plants exhibited lesschlorophyll loss and leaf curling than wild-type plants (Capell et al. 2004; Peremartiet al. 2009). Another example is introduction of the Tritordeum SAMDC (S-adenosylmethionine decarboxylase) gene into the rice plant, leading to a three- tofour-fold increase in the Spd and Spm levels in transformed plants, which then hadnormal growth and development, even under NaCl stress (Roy and Wu 2002). Inanother example, transgenic tobacco plants overexpressing a human SAMDC genehad higher Spd and Put levels and exhibited tolerance to drought and salt stress(Waie and Rajam 2003). Recently, Peremarti et al. (2009) generated transgenic riceplants constitutively expressing heterologous SAMDC gene from Datura stramoniumto distinguish between the roles of Put and the higher polyamines Spd andSpm. Both transgenic and wild-type plants showed identical symptoms whenexposed to drought stress, but transgenic plants recovered much more quickly onrewatering.
In another set of experiments, ADC-expressing transgenic rice plants producedhigher levels of Put, Spd and Spm and exhibited drought tolerance. These resultsconfirmed the involvement of polyamines in drought stress and further attributedindividual roles to Put, Spd and Spm. Interestingly, introduction of a singlepolyamine biosynthesis gene has been shown to confer tolerance to multiple stresses.Examples of these are when Kasukabe et al. (2004, 2006) and Wi et al. (2006)reported that the overexpression of the SPDS (spermidine synthase) from Curcurbitaficifolia in Arabidopsis, sweet potato (Ipomoea batatas) or tobacco enhanced thetolerance of these plants to drought, chilling, freezing, salinity and oxidative stress.Similarly, Prabhavathi and Rajam (2007) have demonstrated that the transgeniceggplant harboring the oat ADC gene exhibited increased tolerance to drought,
Archives of Agronomy and Soil Science 709
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3
Table
5.
Stress-toleranttransgenic
plants
expressingosm
olytes(polyamines).
Gene
Molecularfunction
Source
Transform
edplant
Perform
ance
of
transgenic
plant
Additionalnotes
References
Polyamines
SAMDC
S-adenosylm
ethionine
decaboxylase
H.sapiens
N.tabacum
Drought,salinity&
biotic
stress
resistance
Itisdiffi
cultto
pinpointwhether
the
tolerance
wasdueto
increasedPut
orSpdorboth
Waie
&Rajam
2003
SPDS
Spermidinesynthase
C.ficifolia
A.thaliana
Multiple
abioticstress
tolerance
Involved
insignalingpathwayswhich
ledto
tolerance
Kasukabeet
al.2004
ADC
Argininedecarboxylase
D.stramonium
O.sativa
Droughttolerance
Itshowed
thatpolyaminemetabolism
hadstrongconnectionwithdrought
stress
Capellet
al.2004
SAMDC
S-adenosylm
ethionine
decaboxylase
D.caryophyllus
N.tabacum
Multiple
abioticstress
tolerance
Norm
alplantdevelopmentwithhigher
photosynthetic
rate
&increasedseed
rate
Wiet
al.2006
ADC
Argininedecarboxylase
A.thaliana
S.melongena
Tolerance
tomultiple
abioticstresses
IncreasedADC
activityledto
higher
polyamineaccumulation
Prabhavathi&
Rajam
2007
SAMDC
S-adenosylm
ethionine
decaboxylase
D.stramonium
O.sativa
Droughttolerance
Transgenic
plants
showed
arapid
recoveryfrom
droughton
rewatering
Perem
artiet
al.2009
SAMDC
S-adenosylm
ethionine
decaboxylase
S.cerevisiae
L.esculentum
Hightemperature
tolerance
Enhancedantioxidantactivity,
protectionofmem
branelipid
peroxidationwereprobably
responsible
fortolerance
Chenget
al.2009
ADC
Argininedecarboxylase
A.thaliana
A.thaliana
Droughttolerance
Transgenicplantsexhibited
reductionin
transpirationrate
andstomatal
conductance
Alcazaret
al.2010
710 S.S. Hussain et al.
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3

salinity, low and high temperature, heavy metal and were also found to be resistanceagainst the fungal wilt disease.
Conclusion
Abiotic stress is one of the primary causes of crop losses worldwide. Crop yieldunder abiotic stress depends not only on the mere survival of plants under stressconditions, but also on the phenological and developmental plasticity of plants.Food security is unsustainable without dramatic yield increases in marginalenvironments, especially drought- and salinity-prone areas (Bartels and Hussain2008; Gill and Tuteja 2010). Drought stress affects practically every aspect of plantgrowth and metabolism. Much progress has been made in unraveling the complexstress-response mechanisms, particularly in the identification of several crucialfactors related to the plant response to drought stress. Model plants like Arabidopsisand tobacco have been mostly used to raise transgenic plants with various genes, butonce their role has been established, stress tolerance could be conferred toeconomically important plants using these genes. It is these types of studies,addressing the function and potentiality of the stress-responsive genes using thetransgenic approach, that we have reviewed here. In future, it is expected that otherapproaches, in particular gene knockouts and RNAi techniques, will further aid inour understanding of the role of various other unknown genes.
Recent progress in the field of genomics, proteomics and metabolomics providesample opportunities for the isolation and characterization of novel drought-responsive genes and pathways. The evolutionary conservation of the stress responsehas shown that functionally analogous stress-associated determinants exist in bothunicellular and multicellular organisms. Therefore, elucidation of the components ofcomplex stress-responsive pathways in simpler systems like bacteria and yeast mayhelp in finding overlapping mechanisms in higher plants. So, coordinated efforts areneeded to understand the illusive genes and the mechanisms of drought tolerance.
A variety of genes has been employed to generate transgenic plants thataccumulate osmoprotectants and are tolerant to drought and other abiotic stresses.It may, however, be noted that these successes reflect the performance of plantsunder controlled greenhouse conditions. In fact, no scientific reports on extensivefield tests have yet been published. Only field trials can show which approaches arepromising for the further development of varieties able to meet economical demands.However, transgenic plants often accumulate low or insufficient osmolytes toimprove plant stress tolerance. Further research is currently underway to improvethe efficiency of this approach by taking into account factors like substrateavailability and metabolic fluxes in transgenic plants, which may result in highaccumulation of these osmolytes. Hence, future studies should involve both areductionist approach, to understand the molecular basis of stress tolerance, and anintegrative approach towards analyzing the benefits of stress-tolerant transgenicplants.
Acknowledgements
We thank Dr Bujun Shi of ACPFG (Australia) for critical reading and English correction ofthe manuscript. SSH is supported by an Endeavour Research Fellowship from Department ofEducation, Employment and Work Relation (DEEWR), Australia.
Archives of Agronomy and Soil Science 711
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3

References
Abebe T, Guenzi AC, Martin B, Cushman JC. 2003. Tolerance of mannitol accumulatingtransgenic wheat to water stress and salinity. Plant Physiol. 131:1748–1755.
Ahmad R, Kim MD, Back KH, Kim HS, Lee HS, Kwon SY, Murata N, Chung WII, KwakSS. 2008. Stress-induced expression of choline oxidase in potato plant chloroplasts confersenhanced tolerance to oxidative, salt, and drought stresses. Plant Cell Rep. 27:687–698.
Alcazar R, Planas J, Saxena T, Zarza X, Bortolotti C, Cuevas J, Bitrian M, Tiburcio AF,Altabella T. 2010. Putrescine accumulation confers drought tolerance in transgenicArabidopsis plants over-expressing the homologous arginine decarboxylase 2 gene. PlantPhysiol Biochem. 48:547–552.
Almeida AM, Silva AB, Araujo SS, Cardoso LA, Santos DMJ, Torne JM, Silva JM, Paul MJ,Fevereiro PS. 2007. Responses to water withdrawal of tobacco plants geneticallyengineered with the AtTPS1 gene: a special reference to photosynthetic parameters.Euphytica. 154:113–126.
Armengaud P, Thiery L, Buhot N, March GG, Savoure A. 2004. Transcriptional regulation ofproline biosynthesis in Medicago truncatula reveals developmental and environmentalspecific features. Plant Physiol. 120:442–450.
Ashraf M, Foolad MR. 2007. Roles of glycinebetaine and proline in improving plant abioticstress resistance. Environ Exp Bot. 59:206–216.
Avonce N, Leyman B, Mascorro Gallardo JO, van Dijck P, Thevelein J, Itturiaga G. 2004.The Arabidopsis trehalose-6-P synthase AtTPS1 gene is a regulator of glucose, abscisicacid, and stress signaling. Plant Physiol. 136:3649–3659.
Bagni N, Tassoni A. 2001. Biosynthesis, oxidation and conjugation of aliphatic polyamines inhigher plants. Amino Acids. 20:301–317.
Bartels D, Hussain SS. 2008. Current status and implications of engineering drought tolerancein plants using transgenic approaches. CAB Rev Persp Agr Vet Sci Nutri Natu Sci. 3:020.
Bhatnagar-Mathur P, Vadez V, Shrama KK. 2007. Transgenic approaches for abiotic stresstolerance in plants: retrospect and prospects. Plant Cell Rep. 27:411–424.
Bhatnagar-Mathur P, Vincent V, Devi MJ, Lavanya M, Vani G, Sharma KK. 2009. Geneticengineering of chickpea (Cicer arietinum L.) with the P5CSF129A gene for osmoregulationwith implications on drought tolerance. Mol Breed. 23:591–606.
Bhattacharya E, Rajam MV. 2006. Polyamine biosynthesis as a novel target for engineeringcrop plants for abiotic stress tolerance. J Plant Biol. 33:1–7.
Bohnert HJ, Jensen RG. 1996. Strategies for engineering water-stress tolerance in plants.Trends Biotechnol. 14:89–97.
Bohnert HJ, Shen B. 1999. Transformation and compatible solutes. Sci Hort. 78:237–260.Bouchereau A, Aziz A, Larher F, Martin-Tanguy J. 1999. Polyamines and environmental
challenges: recent development. Plant Sci. 140:103–125.Bray EA, Bailey-Serres J, Weretilnyk E. 2000. Responses to abiotic stresses. In: Buchanana
BB, Gruissem W, Jones RL, editors. Biochemistry and Molecular Biology of Plants.Rockville (MD): American Society of Plant Physiologists. p. 1158–1249.
Bruni F, Leopold AC. 1992. Cytoplasmic glass formation in maize embryos. Seed Sci Res.2:251–253.
Capell T, Bassie L, Christou P. 2004. Modulation of the polyamine biosynthetic pathway intransgenic rice confers tolerance to drought stress. Proc Natl Acad Sci USA. 101:9909–9914.
Capell T, Escobar C, Liu H, Burtin D, Lepri O, Christou P. 1998. Over-expression of oatarginine decarboxylase cDNA in transgenic rice (Oryza sativa L.) affects normaldevelopment pattern in vitro and results in putrescine accumulation in transgenic plants.Theoret Appl Genet. 97:246–254.
Castiglioni P, Warner D, Bensen RJ, Anstrom DC, Harrison J, Stoecker M, Abad M, KumarG, Salvador S, D’ordine R, Navarro S, Back S, Fernandes M, Targolli J, Dasgupta S,Bonin C, Luethy MH, Heard JE. 2008. Bacterial RNA chaperones confer abiotic stresstolerance in plants and improved grain yield in maize under water limited conditions.Plant Physiol. 147:446–455.
Chaitanya KV, Sundar D, Jutur PP, Reddy RA. 2003. Water stress effects on photosynthesisin different mulberry cultivars. Plant Growth Regul. 40:75–80.
712 S.S. Hussain et al.
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3

Chaves MM, Oliveira MM. 2004. Mechanisms underlying plant resistance to water deficits:prospects for water-saving agriculture. J Exp Bot. 55:2365–2384.
Chen CF, Li YW, Chen Y, Bai JR, Li H, Zhu YF, Chen SY, Jia X. 2004. Saline tolerant whiteclover transformed with the betaine aldehyde dehyrogenase gene by Agrobacteriumtumefaciens. Acta Genet Sin. 31:97–101.
Chen JB, Wang SM, Jing RL, Mao XG. 2009. Cloning the PvP5CS gene from common bean(Phaseolus vulgaris) and its expression patterns under abiotic stresses. J Plant Physiol.166:12–19.
Chen TH, Murata N. 2002. Enhancement of tolerance of abiotic stress bymetabolic engineering of betaines and other compatible solutes. Curr Opin Plant Biol.5:250–257.
Chen TH, Murata N. 2008. Glycine betaine: an effective protectant against abiotic stresses inplants. Trends Plant Sci. 13:499–505.
Cheng L, Zou Y, Ding S, Zhang J, Yu X, Cao J, Lu G. 2009. Polyamine accumulation intransgenic tomato enhances the tolerance to high temperature stress. J Integr Plant Biol.51:489–499.
Chinnusamy V, Jagendorf A, Zhu JK. 2005. Understanding and improving salt tolerance inplants. Crop Sci. 45:437–438.
Claussen W. 2005. Proline as a measure of stress in tomato plants. Plant Sci. 168:241–248.Cortina C, Culianez-Macia F. 2005. Tomato abiotic stress enhanced tolerance by trehalose
synthesis. Plant Sci. 169:75–82.Crowe JH, Carpenter JF, Crowe LM. 1998. The role of vitrification in anhydrobiosis. Annu
Rev Physiol. 60:73–103.Crowe JH, Hoekstra FA, Crowe LM. 1992. Anhydrobiosis. Annu Rev Physiol. 54:579–599.Datta SK. 2002. Recent developments in transgenics for abiotic stress tolerance in rice.
JIRCAS Working Report. Japan International Research Centre for Agricultural Sciences(JIRCAS) p. 431–53.
Davies PJ. 2004. The plant hormones: their nature, occurrence and function. In: Davies PJ,editor. Plant Hormones, Biosynthesis, Signal Transduction, Action. Dordrecht (TheNetherlands): Kluwer Academic. p. 1–15.
Delauney AJ, Verma DPS. 1993. Proline biosynthesis and osmoregulation in plants. Plant J.4:215–223.
DeRonde JAD, Cress WA, Krugerd GHJ, Strasserd RJ, Van Staden J. 2004. Photosyntheticresponse of transgenic soybean plants, containing an Arabidopsis P5CS gene, during heatand drought stress. J Plant Physiol. 161:1211–1224.
Dorffling K, Schulenburg S, Lesselich G, Dorffling H. 1990. Abscisic acid and proline levels incold hardened winter wheat leaves in relation to variety-specific differences in freezingresistance. J Agron Crop Sci. 165:230–239.
Ebskamp MJM, van der Meer IM, Spronk BA, Weisbeek PJ, Smeekens SJM. 1994.Accumulation of fructose polymers in transgenic tobacco. Biotechnology. 12:272–275.
Einset J, Connolly EL. 2009. Glycine betaine enhances extra cellular processes blocking ROSsignaling during stress. Plant Signal Behav. 4:197–199.
Einset J, Winge P, Bones AM. 2007. ROS signaling pathway in chilling stress. Plant SignalBehav. 2:365–367.
Einset J, Winge P, Bones AM, Connolly EL. 2008. The FRO2 ferric reductase is required forglycine betaine’s effect on chilling tolerance in Arabidopsis roots. Plant Physiol. 134:334–341.
Flowers TJ. 2004. Improving crop salt tolerance. J Exp Bot. 55:307–319.Franceschetti M, Fornale S, Tassoni A, Zuccherelli K, Mayer MJ, Bagni N. 2004. Effects of
spermidine synthase over-expression on polyamine biosynthetic pathway in tobaccoplants. J Plant Physiol. 161:989–1001.
Franceschetti M, Hanfrey C, Scaramagli S, Torrigiani P, Bagni N, Burtin D, Michael AJ.2001. Characterization of monocot and dicot plant S-adenosyl-methionine decarboxylasegene families including identification in the mRNA of a highly conserved pair of upstreamoverlapping open reading frames. J Biochem. 353:403–409.
Fujita T, Maggio A, Garcia-Rios M, Bressan RA, Csonka LN. 1998. Comparative analysis ofthe regulation of expression and structures of two evolutionarily divergent genes from D1-pyrroline-5-carboxylate synthetase from tomato. Plant Physiol. 118:661–674.
Archives of Agronomy and Soil Science 713
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3

Galston AW. 2001. Plant biology: retrospect and prospect. Curr Sci. 80:143–152.Garg AK, Kim J-K, Owens TG, Ranwala AP, Choi YD, Kochian LV, Wu RJ. 2002.
Trehalose accumulation in rice plants confers high tolerance levels to different abioticstresses. Proc Natl Acad Sci USA. 99:15898–15903.
Ge C, Cui X, Wang Y, Hu Y, Fu Z, Zhang D, Cheng Z, Li J. 2006. BUD2, encoding an S-adenosylmethionine decarboxylase, is required for Arabidopsis growth and development.Cell Res. 16:446–456.
Ge LF, Chao DY, Shi M, Zhu MZ, Gao JP, Lin HX. 2008. Over-expression of the trehalose-6-phosphate phosphatase gene OsTPP1 confers stress tolerance in rice and results inactivation of stress responsive genes. Planta. 228:191–201.
Gill SS, Tuteja N. 2010. Polyamines and abiotic stress tolerance in plants. Plant Signal Behav.5:26–33.
Ginzberg I, Stein H, Kapulink Y, Szabados L, Strizhov H, Schnell J, Koncz C, Zilberstein A.1998. Isolation and characterization of two different cDNAs of D1-pyrroline-5-carboxylatesynthetase in alfalfa, transcriptionally induced upon salt stress. Plant Mol Biol. 38:755–764.
Goddijn O, Smeekens S. 1998. Sensing trehalose biosynthesis in plants. Plant J. 14:143–146.Goddijn OJM, van Dun K. 1999. Trehalose metabolism in plants. Trends Plant Sci. 4:315–319.Goddijn OJM, Verwoerd TC, Voogd E, Krutwagen RWHH, de Graaf PTHM, Poels J, Van
Dun K, Ponstein AS, Damm B, Pen J. 1997. Inhibition of trehalase activity enhancestrehalose accumulation in transgenic plants. Plant Physiol. 113:181–190.
Gubis J, Vankova R, Cervena V, Dragunova MM, Hudcovicova M, Lichtnerova HT,Dokupil T, Jurekova Z. 2007. Transformed tobacco plants with increased tolerance todrought. South Afr J Bot. 73:505–511.
Guo Y, Zhang L, Ziao G, Cao SY, Cu DM, Tian WZ, Shouyi C. 1997. Expression of betainealdehyde dehydrogenase gene and salinity tolerance in rice transgenic plants. Sci ChinaSer. 40:496–501.
Han SE, Park SR, Kwon HB, Yi BY, Lee GB, Byun MO. 2005. Genetic engineering ofdrought-resistant tobacco plants by introducing the trehalose phosphorylase (TP) genefrom Pleurotus sajor-caju. Plant Cell Tissue Organ Cult. 82:151–158.
Hanzawa Y, Imai A, Michael AJ, Komeda Y, Takahashi T. 2002. Characterization of thespermidine synthase-related gene family in Arabidopsis thaliana. FEBS Lett. 527:176–180.
Hanzawa Y, Takahashi T, Komeda Y. 1997. ACL5: an Arabidopsis gene required forinternodal elongation after flowering. Plant J. 12:863–874.
Hanzawa Y, Takahashi T, Michael AJ, Burtin D, Long D, Pineiro M, Coupland G, KomedaY. 2000. ACAULIS5, an Arabidopsis gene required for stem elongation, encodes aspermine synthase. EMBO J. 19:4248–4256.
Hasegawa PM, Bressan R, Zhu JK, Bohnert HJ. 2000. Plant cellular and molecular responsesto high salinity. Annu Rev Plant Physiol Plant Mol Biol. 51:463–499.
Hendry GAF, Wallace RK. 1993. The origin, distribution, and evolutionary significance offructans. In: Suzuki M, Chatterton NJ, editors. Science and Technology of Fructans. BocaRaton (FL): CRC Press. p. 119–139.
Holmberg N, Bulow L. 1998. Improving stress tolerance in plants by gene transfer. TrendsPlant Sci. 3:61–66.
Holmstrom K-O, Mantyla E, Welin B, Mandal A, Palva ET, Tunnela OE, Londesborough J.1996. Drought tolerance in tobacco. Nature. 379:683–684.
Hong ZL, Lakkineni K, Zhang ZM, Verma DPS. 2000. Removal of feedback inhibition ofdelta(1)-pyrroline-5-carboxylate synthetase results in increased proline accumulation andprotection of plants from osmotic stress. Plant Physiol. 122:1129–1136.
Hu CAA, Delauney AJ, Verma DPS. 1992. A bifunctional enzyme (D1-pyrroline-5-carboxylate synthetase) catalyzes the first two steps in praline biosynthesis in plants.Proc Natl Acad Sci USA. 89:9354–9358.
Huang J, Hirji R, Adam L, Roazwadowski KL, Hammerlindi JK, Keller WA, Selvaraj G.2000. Genetic engineering of glycinebetaine production towards enhancing stress tolerancein plants: metabolic limitations. Plant Physiol. 122:747–756.
Hummel I, Gouesbet G, El Amrani A, Ainouche A, Couee I. 2004. Characterization of thetwo arginine decarboxylase (polyamine biosynthesis) paralogues of the endemicsubantarctic cruciferous species Pringlea antiscorbutica and analysis of their differentialexpression during development and response to environmental stress. Gene. 342:199–209.
714 S.S. Hussain et al.
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3

Hussain M, Farooq M, Jabran K, Rehman H, Akram M. 2008. Exogenous glycinebetaineapplication improves yield under water limited conditions in hybrid sunflower. ArchAgron Soil Sci. 54:557–567.
Igarashi K, Kashiwagi K. 2000. Polyamines: mysterious modulators of cellular functions.Biochem Biophys Res Commun. 271:559–564.
Igarashi Y, Yoshiba Y, Sanada Y, Yamaguchi-Shinozaki K, Wada K, Shinozaki K. 1997.Characterization of the gene D1-pyrroline-5-carboxylate synthetase and correlationbetween the expression of the gene and salt tolerance in Oryza sativa L. Plant Mol Biol.33:857–865.
Imai R, Ali A, Pramanik HR, Nakaminami K, Sentoku N, Kato H. 2004. A distinctive classof spermidine synthase is involved in chilling response in rice. J Plant Physiol. 161:883–886.
Iturriaga G, Gaff GF, Zentella R. 2000. New desiccation tolerant plants, including a grass inthe central highlands of Mexico, accumulate trehalose. Aust J Bot. 48:153–158.
Iyer S, Caplan A. 1998. Products of praline catabolism can induce osmotically regulated genesin rice. Plant Physiol. 116:203–211.
Jain RK, Selvaraj G. 1997. Molecular genetic improvement of salt tolerance in plants. AnnuRev Biotechnol. 3:245–267.
Jang IC, Oh SJ, Seo JS, Choi WB, Song SI, Kim CH, Kim YS, Seo H-S, Choi YD, Nahm BH,Kim J-K. 2003. Expression of a bifunctional fusion of the Escherichia coli genes fortrehalose-6-phosphate synthase and trehalose-6-phosphate phosphate in transgenic riceplants increases trehalose accumulation and abiotic stress tolerance without stuntinggrowth. Plant Physiol. 131:516–524.
Janowitz T, Kneifel H, Piotrowski M. 2003. Identification and characterization of plantagmatine iminohydrolase, the last missing link in polyamine biosynthesis of plants. FEBSLett. 544:258–261.
Karakas B, Ozias-Akins P, Stushnoff C, Suefferheld M, Rieger M. 1997. Salinity and droughttolerance of mannitol-accumulating transgenic plants. Plant Cell Environ. 20:609–616.
Karim S, Aronsson H, Ericson H, Pirhonen M, Leyman B, Welin B, Mantyla E, Palva ET,van Dijck P, Holstrom KO. 2007. Improved drought tolerance without undesired sideeffects in transgenic plants producing trehalose. Plant Mol Biol. 64:371–386.
Kasukabe Y, He L, Nada K, Misawa S, Ihara I, Tachibana S. 2004. Overexpression ofspermidine synthase enhances tolerance to multiple environmental stresses and up-regulates the expression of various stress-regulated genes in transgenic Arabidopsisthaliana. Plant Cell Physiol. 45:712–722.
Kasukabe Y, He L, Watakabe Y, Otani M, Shimada T, Tachibana S. 2006. Improvement ofenvironmental stress tolerance of sweet potato by introduction of genes for spermidinesynthase. Plant Biotechnol. 23:75–83.
Kathuria H, Giri J, Nataraja KN, Murata N, Udayakumar M, Tyagi AK. 2009.Glyvinebetaine-induced water stress tolerance in codA-expressing transgenic indica riceis associated with up-regulation of several stress responsive genes. Plant Biotechnol J.7:512–526.
Kathuria H, Giri J, Tyagi H, Tyagi AK. 2007. Advances in transgenic rice biotechnology. CritRev Plant Sci. 26:65–103.
Kishor PBK, Hong Z, Miao GH, Hu CAA, Verma DPS. 1995. Overexression of delta 1-pyrroline-5carboxylate synthetase increases proline production and confers osmotolerancein transgenic plants. Plant Physiol. 108:1387–1394.
Knipp G, Honermeier B. 2006. Effect of water stress on proline accumulation of geneticallymodified potatoes (Solanum tuberosum L.) generating fructans. J Plant Physiol. 163:392–397.
Kuehn GD, Phillips GC. 2005. Roles of polyamines in apoptosis and other recent advances inplant polyamines. Crit Rev Plant Sci. 24:123–130.
Kumar S, Dhingra A, Daniell H. 2004. Plastid-expressed betaine aldehyde dehydrogenasegene in carrot cultured cells, roots, and leaves confer enhanced salt tolerance. PlantPhysiol. 136:2843–2854.
Kumar SG, Reddy AM, Sudhakar C. 2003. NaCl effects on proline metabolism in two highyielding genotypes of mulberry (Morus alba L.) with contrasting salt tolerance. Plant Sci.165:1245–1251.
Archives of Agronomy and Soil Science 715
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3

Kumar SV, Sharma ML, Rajam MV. 2006. Polyamine biosynthetic pathway as a novel targetfor potential applications in plant biotechnology. Physiol Mol Biol Plant. 12:13–28.
Kumar V, Shriram V, Kavi Kishor PB, Jawali N, Shitole MG. 2010. Enhanced prolineaccumulation and salt stress tolerance of transgenic indica rice by overexpressingP5CSF129A gene. Plant Biotechnol Rep. 4:37–48.
Kumria R, Rajam MV. 2002. Ornithine decarboxylase transgene in tobacco affects polyaminemetabolism, in vitro morphogenesis and response to salt stress. J Plant Physiol. 159:983–990.
Lee KS, Choi WY, Ko JC, Kim TS, Gregorio GB. 2003. Salinity tolerance of japonica andindica rice (Oryza sativa L.) at the seedling stage. Planta. 216:1043–1046.
Liu JH, Nada K, Honda C, Kitashiba H, Wen XP, Pang XM, Moriguchi T. 2006. Polyaminebiosynthesis of apple callus under salt stress: importance of arginine decarboxylasepathway in stress response. J Exp Bot. 57:2589–2599.
Livingston DP III, Hincha DK, Heyer AG. 2009. Fructan and its relationship to abiotic stresstolerance in plants. Cell Mol Life Sci. 66:2007–2023.
Lv S, Yang A, Zhang K, Wang L, Zhang J. 2007. Increase of glycinebetaine synthesisimproves drought tolerance in cotton. Mol Breed. 20:233–248.
Ma R, Zhang M, Li B, Du G, Wang J, Chen J. 2005. The effects of endogenous Ca2þ onendogenous polyamine levels and drought-resistant traits of spring wheat grown underarid conditions. J Arid Environ. 63:177–190.
Maiale S, Sanchez DH, Guirda A, Vidal A, Ruiz O. 2004. Spermine accumulation under saltstress. J Plant Physiol. 161:35–42.
Malmberg RL, Watson MB, Galloway GL, Yu W. 1998. Molecular genetic analysis of plantpolyamines. Crit Rev Plant Sci. 17:199–224.
Mani S, van de Cotte B, Montagu M, Verbruggen N. 2002. Altered levels of prolinedehydrogenase cause hypersensitivity to praline and its analogs in Arabidopsis. PlantPhysiol. 128:73–83.
McCue KF, Hanson AD. 1990. Drought and salt tolerance: toward understanding andapplication. Trends Biotechnol. 8:358–362.
McNeil SD, Nuccio ML, Hanson AD. 1999. Betaines and related osmoprotectants. Targetsfor metabolic engineering of stress resistance. Plant Physiol. 120:945–949.
McNeil SD, Nuccio ML, Rhodes D, Shachar-Hill Y, Hanson AD. 2000. Radiotracer andcomputer modeling evidence that phosphobase methylation in the main route of cholinesynthesis in tobacco. Plant Physiol. 123:371–380.
McNeil SD, Nuccio ML, Ziemak MJ, Hanson AD. 2001. Enhanced synthesis of choline andglycine betaine in transgenic tobacco plants that overexpress phosphoethanolamine N-methyltransferase. Proc Natl Acad Sci USA. 98:10001–10005.
Miranda JA, Avonce N, Suarez R, Thevelein JM, Dijck PV, Iturriaga G. 2007. A bifunctionalTPS–TPP enzyme from yeast confers tolerance to multiple and extreme abiotic conditionsin transgenic Arabidopsis. Planta. 226:1411–1421.
Mohanty A, Kathuria H, Ferjani A, Sakamoto A, Mohanty P, Murata N, Tyagi AK. 2002.Transgenic of an elite indica rice variety Pusa Basmati 1 harbouring the codA gene ishighly tolerant to salt stress. Theor Appl Genet. 106:51–57.
Molinari HBC, Marur CJ, Daros E, de Campos MKF, Rodrigues JF, de Carvalho P, FilhoJCB, Pereira LFP, Vieira LGE. 2007. Evaluation of the stress-inducible production ofproline in transgenic sugarcane (Saccharum spp.): osmotic adjustment, chlorophyllfluorescence and oxidative stress. Plant Physiol. 130:218–229.
Molinari HBC, Marur CJ, Filho JCB, Kobayashi AK, Pileggi M, Junior RPL, Pereira LFP,Vieira LGE. 2004. Osmotic adjustment in transgenic citrus rootstock Carrizo citrange(Citrus sinensis Osb. 6 Poncirus trifoliata L. Raf.) overproducing proline. Plant Sci.167:1375–1381.
Nakamura T, Yokota S, Muramoto Y, Tsutsui K, Oguri Y, Fukui K, Takabe T. 1997.Expression of a betaine aldehyde dehydrogenase gene in rice, a glycine betainenonaccumulator, and possible localization of its protein in peroxisomes. Plant J.11:1115–1120.
Nanjo T, Fujita M, Seki M, Kato T, Tabata S, Shinozaki K. 2003. Toxicity of free prolinerevealed in an Arabidopsis T-DNA-tagged mutant deficient in proline dehydrogenase.Plant Cell Physiol. 44:541–548.
716 S.S. Hussain et al.
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3

Nanjo T, Kobayashi M, Yoshiba Y, Kakubari Y, Yamaguchi-Shinozaki K, Shinozaki K.1999. Antisense suppression of praline degradation improves to freezing and salinity inArabidopsis thaliana. FEBS Lett. 461:205–210.
Nuccio ML, McNeil SD, Ziemak MJ, Hanson AD, Jain RK, Selvaraj G. 2000. Choline importinto chloroplasts limits glycine betaine synthesis in tobacco: analysis of plants engineeredwith a chloroplastic or a cytosolic pathway. Metab Eng. 2:300–311.
Nuccio ML, Rhodes D, McNeil SD, Hanson AD. 1999. Metabolic engineering of plants forosmotic stress resistance. Curr Opin Plant Biol. 2:128–134.
Nuccio ML, Russell BL, Nolte KD, Rathinasabapathi B, Gage DA, Hanson AD. 1998. Theendogenous choline supply limits glycine betaine synthesis in transgenic tobaccoexpressing choline monooxygenase. Plant J. 16:487–496.
Papageorgiou GC, Murata N. 1995. The unusually strong stabilizing effects of glycinebetaineon the structure and function of oxygen-evolving photosystem II complex. PhotosynthRes. 44:243–252.
Park EJ, Jeknic Z, Pino MT, Murata N, Chen THH. 2007. Glycinebetaine accumulation ismore effective in chloroplasts than in the cytosol for protecting transgenic tomato plantsagainst abiotic stress. Plant Cell Environ. 30:994–1005.
Park EJ, Jeknic Z, Sakamoto A, Denoma Y, Yawansiri R, Murata N, Chen THH. 2004.Genetic engineering of glycine betaine synthesis in tomato protects seeds, plants, andflowers from chilling damage. Plant J. 40:474–487.
Paul JM, Primavesi LF, Jhurreea D, Zhang Y. 2008. Trehalose metabolism and signaling.Annu Rev Plant Biol. 59:417–441.
Paul M. 2007. Trehalose-6-phosphate. Curr Opin Plant Biol. 10:303–309.Penna S. 2003. Building stress tolerance through over-producing trehalose in transgenic
plants. Trends Plant Sci. 8:355–357.Peremarti A, Bassie L, Christou P, Capell T. 2009. Spermine facilitates recovery from drought
but does not confer drought tolerance in transgenic rice plants expressing Daturastramonium S-adenosylmethionine decarboxylase. Plant Mol Biol. 70:253–264.
Pilon-Smits EAH, Ebskamp MJM, Paul MJ, Jeuken MJW, Weisbeek PJ, Smeekens SCM.1995. Improved performance of transgenic fructan-accumulating tobacco under droughtstress. Plant Physiol. 107:125–130.
Pilon-Smits EAH, Terry N, Sears T, Kim H, Zayed A, Hwang S, Van Dun K, Voogd E,Verwoerd TC, Krutwagen RWHH, Giddijn JM. 1998. Trehalose-producing transgenictobacco plants show improved growth performance under drought stress. J Plant Physiol.152:525–532.
Pilon-Smits EAH, Terry N, Sears T, van Dunn K. 1999. Enhanced drought resistance infructan-producing sugar beet. Plant Physiol Biochem. 37:313–317.
Prabhavathi VR, Rajam MV. 2007. Polyamine accumulation in transgenic eggplant enhancestolerance to multiple abiotic stresses and fungal resistance. Plant Biotechnol. 24:273–282.
Prabhavathi VR, Yadav JS, Kumar PA, Rajam MV. 2002. Abiotic stress tolerance intransgenic eggplant (Solanum melongena L.) by introduction of bacterial mannitolphosphodehydrogenase gene. Mol Breed. 9:137–147.
Quan R, Shang M, Zhang H, Zhao Y, Zhang J. 2004a. Engineering of enhanced glycinebetaine synthesis improves drought tolerance in maize. Plant Biotechnol J. 2:477–486.
Quan R, Shang M, Zhang H, Zhao Y, Zhang J. 2004b. Improved chilling tolerance bytransformation with beta gene for the enhancement of glycinebetaine synthesis in maize.Plant Sci. 166:141–149.
Rajam MV. 1997. Polyamines. In: Prasad MNV, editor. Plant Ecophysiology. New york:Wiley. p. 343–374.
Rajam MV, Dagar S, Waie B, Yadav JS, Kumar PA, Shoeb F, Kumria R. 1998. Geneticengineering of polyamine and carbohydrate metabolism for osmotic stress tolerance inhigher plants. J Biosci. 23:473–482.
Rhodes D, Hanson AD. 1993. Quaternary ammonium and tertiary sulfonium compounds inhigher plants. Annu Rev Plant Physiol Plant Mol Biol. 44:357–384.
Romero C, Belles JM, Vaya JL, Serrano R, Culianez-Macia FA. 1997. Expression of the yeasttrehalose-6-phosphate synthase gene in transgenic tobacco plants: pleiotropic phenotypesincludedrought tolerance. Planta. 201:293–297.
Archives of Agronomy and Soil Science 717
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3

Rontein D, Basset G, Hanson AD. 2002. Metabolic engineering of osmoprotectantaccumulation in plants. Metab Eng. 4:49–56.
Roosens NH, Al Bitar F, Loenders K, Angenon G, Jacob M. 2002. Overexpression ofornithine-delta-aminotransferase increaees proline biosynthesis and confers osmotoler-ance in transgenic plants. Mol Breed. 9:73–80.
Roy M, Wu R. 2001. Arginine decarboxylase transgene expression and analysis ofenvironmental stress tolerance in transgenic rice. Plant Sci. 160:869–875.
Roy M, Wu R. 2002. Overexpression of S-adenosylmethionine enhances sodium chloride-stress tolerance. Plant Sci. 163:987–992.
Roy P, Niyogi K, Sen Gupta DN, Ghosh B. 2005. Spermidine treatment to rice seedlingsrecovers salinity stress-induced damage of plasma membrane and PM-bound-Hþ-ATPasein salt tolerant and salt-sensitive rice cultivars. Plant Sci. 168:583–591.
Sakamoto A, Murata N. 2000. Genetic engineering of glycinebetaine synthesis in plants:current status and implications for enhancement of stress tolerance. J Exp Bot. 51:81–88.
Sakamoto A, Murata N. 2001. The use of choline oxidase, a glycinebetaine-synthesizingenzyme, to create stress resistant transgenic plants. Plant Physiol. 125:180–188.
Sakamoto A, Alia, Murata N. 1998. Metabolic engineering of rice leading to biosynthesis ofglycine betaine and tolerance to salt and cold. Plant Mol Biol. 38:1011–1019.
Sakamoto A, Valverde R, Alia, Chen THH, Murata N. 2000. Transformation of Arabidopsiswith the codA gene for choline oxidase enhances freezing tolerance of plants. Plant J.22:449–453.
Sawahel WA, Hassan AH. 2002. Generation of transgenic wheat plants producing high levelsof osmoprotectant proline. Biotechnol Lett. 24:721–725.
Seki M, Umezawa T, Urano K, Shinozaki K. 2007. Regulatory metabolic networks in droughtstress responses. Curr Opin Plant Biol. 10:296–302.
Serraj R, Sinclair TR. 2002. Osmolyte accumulation: can it really help increase crop yieldunder drought conditions? Plant Cell Environ. 25:333–341.
Shen B, Jensen RG, Bohnert HJ. 1997. Increased resistance to oxidative stress in transgenicplants by targeting mannitol biosynthesis to chloroplasts. Plant Physiol. 113:1177–1183.
Shen YG, Du BX, Zhang WK, Zhang JS, Chen SY. 2002. AhCMO, regulated by stresses inAtriplex hortensis, can improve drought tolerance in transgenic tobacco. Theor ApplGenet. 105:815–821.
Sheveleva EV, Chmara W, Bohnert HJ, Jensen RG. 1997. Increased salt and droughttolerance by D-ononitol production in transgenic Nicotiana tabacumL. Plant Physiol.115:1211–1219.
Sheveleva EV, Marquez S, Chmara W, Zegeer A, Jensen RG, Bohnert HJ. 1998. Sorbitol-6-phosphate dehydrogenase expression in transgenic tobacco. High amounts of sorbitol leadto necrotic lesions. Plant Physiol. 117:831–839.
Shinozaki K, Yamaguchi-Shinozaki K. 2007. Gene networks involved in drought stressresponse and tolerance. J Exp Bot. 58:221–227.
Shirasawa K, Takabe T, Takabe T, Kishitani S. 2006. Accumulation of glycinebetaine in riceplants that overexpress choline monooxygenase from spinach and evaluation of theirtolerance to abiotic stress. Ann Bot. 98:565–571.
Simon-Sarkadi L, Kocsy G, Varhegyi A, Galiba G, de Ronde JA. 2005. Genetic manipulationof proline accumulation influences the concentrations of other amino acids in soybeansubjected to simultaneous drought and heat stress. J Agr Food Chem. 53:7512–7517.
Singer MA, Lindquist S. 1998. Thermotolerance in Saccharomyces cerevisiae: the Yin andYang of trehalose. Trends Biotechnol. 16:460–468.
Smart CC, Flores S. 1997. Overexpression of D-myo-inositol-3-phosphate synthase leads toelevated levels of inositol in Arabidopsis. Plant Mol Biol. 33:811–820.
Smirnoff N, Cumbes QJ. 1989. Hydroxyl radical scavenging activity of compatible solutes.Phytochemistry. 28:1057–1060.
Song J, Nada K, Tachibana S. 2002. Suppression of S-adenosylmethionine decarboxylaseactivities is a major cause for hightemperature inhibition of pollen germination and tubegrowth in tomato (Lycopersicon esculentum Mill.). Plant Cell Physiol. 43:619–627.
Sreenivasulu N, Sopory SK, Kavi Kishor PB. 2007. Deciphering the regulatory mechanisms ofabiotic stress tolerance in plants by genomic approaches. Gene. 388:1–13.
718 S.S. Hussain et al.
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3

Stiller I, Dulai S, Kondrak M, Tarnai R, Szabo L, Toldo O, Banfalvi Z. 2008. Effects ofdrought on water content and photosynthetic parameters in potato plants expressing thetrehalose-6-phosphate synthase gene of Saccharomyces cerevisiae. Planta. 227:299–308.
Strizhov N, Abraham E, Okresz L, Blichling S, Zilberstein A, Schell J, Koncz A, Szabados L.1997. Differential expression of two P5CS genes controlling praline accumulation duringsalt stress requires ABA and is regulated by ABA1,ABI1 and AXR2 in Arabidopsis. PlantJ. 12:557–569.
Su J, Wu R. 2004. Stress-inducible synthesis of proline in transgenic rice confers faster growthunder stress conditions than that with constitutive synthesis. Plant Sci. 166:941–948.
Suarez R, Calderon C, Itturiaga G. 2009. Enhanced tolerance to multiple abiotic stresses intransgenic alfalfa accumulating trehalose. Crop Sci. 49:1791–1799.
Suzuki N, Bajad S, Shuman J, Shulaev V, Mittler R. 2008. The transcriptional co-activatormbf1c is a key regulator of the thermotolerance in Arabidopsis thaliana. J Biol Chem.283:9269–9275.
Szabados L, Savoure A. 2009. Proline: a multifunctional amino acid. Trends Plant Sci. 15:89–97.
Taji T, Ohsumi C, Luchi S, Seki M, Kasuga M, Kobayashi M, Yamaguchi-Shinozaki K,Shinozaki K. 2002. Important role of drought and cold-inducible genes for galactinolsynthase in stress tolerance in Arabidopsis thaliana. Plant J. 29:417–426.
Takabe T, Hayashi Y, Tanaka A, Kishitani S. 1998. Evaluation of glycine betaineaccumulation for stress tolerance in transgenic rice plants. Proceedings of theInternational Workshop on Breeding and Biotechnology for environmental stress inRice. Hokkaido Agricultural Experiment Station, Sapporo, Japan. p. 63–68.
Tao R, Uratsu SL, Dandekar AM. 1995. Sorbitol synthesis in transgenic tobacco with applecDNA encoding NADP-dependent sorbitol-6-phosphate dehydrogenase. Plant CellPhysiol. 366:525–532.
Tarczynski MC, Jensen RG, Bohnert HJ. 1992. Expression of a bacterial mtlD gene intransgenic tobacco leads to production and accumulation of mannitol. Proc Natl Acad SciUSA. 89:2600–2604.
Thippeswamy M, Chandraobulreddy P, Sinilal B, Shivakumar M, Sudhakar C. 2010. Prolineaccumulation and the expression of D1-pyrroline-5-carboxylate synthase in two safflowercultivars. Biol Plant. 54:386–390.
Thomas JC, Sepahi M, Arendall B, Bohnert HJ. 1995. Enhancement of seed germination inhigh salinity by engineering mannitol expression in Arabidopsis thaliana. Plant CellEnviron. 18:801–806.
Thomas T, Thomas TJ. 2001. Polyamines in cell growth and cell death: molecular mechanismsand therapeutic applications. Cell Mol Life Sci. 58:244–58.
Trossat C, Rathinasabapathi B, Hanson AA. 1997. Transgenically expressed betaine aldehydedehydrogenase efficiently catalyses oxidation of dimethylsulfonioproprion-aldehyde andomega-aminoaldehydes. Plant Physiol. 113:1457–1461.
Valliyodan B, Nguyen H. 2006. Understanding regulatory networks and engineering forenhanced drought tolerance in plants. Curr Opin Plant Biol. 9:1–7.
Vendruscolo ECG, Schuster I, Pileggi M, Scapim CA, Molinari HBC, Marur CJ, Vieira LGE.2007. Stress-induced synthesis of proline confers tolerance to water deficit in transgenicwheat. J Plant Physiol. 164:1367–1376.
Venekamp JH, Lampe JEM, Koot JTM. 1989. Organic acids as source of drought inducibleproline synthesis in field bean plants Vicia faba L. J Plant Physiol. 133:654–659.
Verdoy D, Coba De La Pena T, Redondo FJ, Lucas MM, Pueyo JJ. 2006. TransgenicMecicago truncatula plants that acumulate proline display nitrogen-fixing activity withenhanced tolerante to osmotic stress. Plant Cell Environ. 29:1913–1923.
Vernon DM, Tarczynski MC, Jensen RG, Bohnert HJ. 1993. Cyclitol production in transgenictobacco. Plant J. 4:199–205.
Vinocur B, Altman A. 2005. Recent advances in engineering plant tolerance to abiotic stress:achievements and limitations. Curr Opin Biotechnol. 16:123–132.
Waditee R, Bhuiyan MN, Rai V, Aoki K, Tanaka Y, Hibino T, Suzuki S, Takano J,Jagendorf AT, Takabe T, Takabe T. 2005. Genes for direct methylation of glycine providehigh levels of glycinebetaine and abiotic-stress tolerance in Synechococcus and Arabidopsis.Proc Natl Acad Sci USA. 102:1318–1323.
Archives of Agronomy and Soil Science 719
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3

Waditee R, Tanaka Y, Aoki K, Hibino T, Jikuya H, Takano J, Takabe T, Takabe T. 2003.Isolation and functional characterization of N-methyltransferases that catalyze betainesynthesis grom glycine in a holotolerant photosynthetic organism Aphanothece halophy-tica. J Biol Chem. 278:4932–4942.
Waie B, Rajam MV. 2003. Effect of increased polyamine biosynthesis on stress responses intransgenic tobacco by introduction of human S-adenosylmethionine gene. Plant Sci.164:727–734.
Walters DR. 2003. Polyamines and plant disease. Phytochemistry. 64:97–107.Wang WX, Vinocur B, Altman A. 2003. Plant responses to drought, salinity and extreme
temperatures: towards genetic engineering for stress tolerance. Planta. 219:1–14.Wang ZZ, Zhang SZ, Yang BP, Li YR. 2005. Trehalose synthase gene transfer mediated by
Agrobacterium tumefaciens enhances resistance to osmotic stress in sugarcane. Sugar Tech.7:49–54.
Watson MB, Malmberg RL. 1996. Regulation of Arabidopsis thaliana (L.) Heynh argininedecarboxylase by potassium deficiency stress. Plant Physiol. 111:1077–1083.
Watson MB, Yu W, Galloway G, Malmberg RL. 1997. Isolation and characterization of asecond arginine decarboxylase cDNA from Arabidopsis (Accession No. AF009647). PlantPhysiol. 114:1569.
Wi SJ, Kim WT, Park KY. 2006. Overexpression of carnation S-adenosylmethioninedecarboxylase gene generates a broad spectrum tolerance to abiotic stresses in transgenictobacco plants. Plant Cell Rep. 25:1111–1121.
Wiemken A, Sprenger N, Boller T. 1995. Fructans – an extension of sucrose by sucrose. In:Pontis HG, Salerno GL, Echeverria EJ, editors. Current Topics in Plant Physiology, Vol14: Sucrose Metabolism, Biochemistry, Physiology and Molecular Biology. Rockville(MD): ASPP. p. 178–189.
Wu LQ, Fan ZM, Guo L, Li YQ, Zhang WJ, Qu LJ, Chen ZL. 2003. Overexpression of anArabidopsis delta OAT gene enhances salt and drought tolerance in transgenic rice. ChinSci Bull. 48:2594–2600.
Wyn Jones RG. 1984. Phytochemical aspects of osmotic adaptation. Rec Adv Phytochem.18:55–78.
Xia Y, Liang HM, Chen SY, Sun ZX, Wang TM, Li XY, Fu LL, Huang J, Yan LP, Liu D.2004. Betaine aldehyde dehydrogenase (BADH) gene transformation of tetraploid clone ofblack locust mediated by Agrobacterium. Sci Agr Sin. 37:1208–1211.
Xing W, Rajashekar CB. 2001. Glycine betaine involvement in freezing tolerance and waterstress in Arabidopsis thaliana. Environ Exp Bot. 46:21–28.
Yamada M, Morishita H, Urano K, Shiozaki N, Yamaguchi-Shinozaki K, Shinozaki K,Yoshiba Y. 2005. Effects of free proline accumulation in petunias under drought stress. JExp Bot. 56:1975–1981.
Yamaguchi K, Takahashi Y, Berberich T, Imai A, Takahashi T, Michael AJ, Kusano T. 2007.A protective role for the polyamine spermine against drought stress in Arabidopsis.Biochem Biophys Res Commun. 352:486–490.
Yamchi A, Jazii FR, Mousav A, Karkhane AA, Renu. 2007. Proline accumulation intransgenic tobacco as a result of expression of Arabidopsis D1-pyrroline-5-carboxylatesynthetase (p5cs) during osmotic stress. J Plant Biochem Biotechnol. 16:9–15.
Yancy PH, Clark ME, Hand SC, Bowlus RD, Somero GN. 1982. Living with water stress:evolution of osmolyte system. Science. 217:1214–1222.
Yang L, Tang R, Zhu J, Liu H, Mueller-Roeber B, Xia H, Zhang H. 2008. Enhancementof stress tolerance in transgenic tobacco plants constitutively expressing AtIpk2b, aninositol polyphosphate 6-/3-kinase from Arabidopsis thaliana. Plant Mol Biol. 66:329–343.
Yeo ET, Kwon HB, Han SE, Lee JT, Ryu JC, Byun MO. 2000. Genetic engineering ofdrought resistant potato plants by introduction of the trehalose-6-phosphate synthase(TPS1) gene from Saccharomyces cerevisiae. Mol Cell. 10:263–268.
Yoshiba Y, Kiyosue T, Nakashima K, Yamaguchi-Shinozaki K, Shinozaki K. 1997.Regulation of levels of proline as an osmolyte in plants under water stress. Plant CellPhysiol. 38:1095–1102.
Zhang H, Dong H, Li W, Sun Y, Chen S, Kong X. 2009. Increased glycinebetaine synthesisand salinity tolerance in AhCMO transgenic cotton lines. Mol Breed. 23:289–298.
720 S.S. Hussain et al.
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3

Zhang SZ, Yang BP, Feng CL, Tang HL. 2005. Genetic transformation of tobacco with thetrehalose synthase gene from Grifola frondosa Fr. enhances the resistance to drought andsalt in tobacco. J Integ Plant Biol. 47:579–587.
Zhao HW, Chen YJ, Hu YL, Gao Y, Lin ZP. 2000. Construction of a trehalose-6-phosphatesynthase gene driven by drought responsive promoter and expression of drought resistancein transgenic tobacco. Acta Bot Sin. 42:616–619.
Zhao Y, Aspinall D, Paleg LG. 1992. Protection of membrane integrity in Medicago sativa L.by glycinebetaine against the effects of freezing. J Plant Physiol. 140:541–543.
Zhu B, Su J, Chang M, Verma DPS, Fan YL, Wu R. 1998. Overexpression of a D1-pyrroline-5- carboxylate synthetase gene and analysis of tolerance to water- and salt-stress intransgenic rice. Plant Sci. 139:41–48.
Zhu JK. 2001. Plant salt tolerance. Trends Plant Sci. 6:66–71.Zhu JK. 2002. Salt and drought stress signal transduction in plants. Annu Rev Plant Biol.
53:247–273.
Archives of Agronomy and Soil Science 721
Dow
nloa
ded
by [
UN
IVE
RSI
TY
OF
AD
EL
AID
E L
IBR
AR
IES]
at 1
9:45
22
July
201
3
Related Documents











