International Journal of Medical Microbiology 297 (2007) 513–523 Transcriptome analyses extend understanding of Streptococcus pyogenes regulatory mechanisms and behavior toward immunomodulatory substances Catur Riani 1 , Kerstin Standar 1 , Somboon Srimuang 2 , Cordula Lembke, Bernd Kreikemeyer, Andreas Podbielski Department of Medical Microbiology, Virology, and Hygiene, University Hospital Rostock, Schillingallee 70, D-18057 Rostock, Germany Received 30 August 2006; received in revised form 2 April 2007; accepted 4 April 2007 Abstract The recent genome sequencing of several Streptococcus pyogenes serotype strains allowed the design of corresponding DNA microarrays and their usage for specific transcriptome analyses. In the present study, we employed transcriptomics together with functional tests to investigate the impact of the CiaH sensor gene of the CiaRH two-component regulator on gene expression and virulence traits of serotype M49 S. pyogenes strains CS101 and 591. In parallel, we studied the effects of the immunostimulatory substance Luivac s on the serotype M49 S. pyogenes transcriptome and several biological features of serotype M1, M2, M3, M6, M18, and M49 S. pyogenes strains. Overall, the transcriptome analyses allowed a swift identification of differences in transcript abundance apparently associated with the observed strain-specific changes in matrix protein binding, eukaryotic cell interactions, or biofilm formation of the ciaH mutants and of wild-type strains exposed to a commercially available substance used for preventing upper respiratory tract infections. r 2007 Elsevier GmbH. All rights reserved. Keywords: Streptococcus pyogenes; Transcriptomics; Cia regulator; Luivac immunostimulator; Virulence Introduction Purulent infections of the upper respiratory tract and skin caused by Streptococcus pyogenes (group A streptococci, GAS) are extremely frequent, with about 730 million yearly incidences worldwide. In addition, these bacteria trigger 940,000 yearly cases of post- infection sequelae and 660,000 yearly cases of life- threatening infections (Carapetis et al., 2005). Unlike for other similarly or less important pathogens, vaccination against S. pyogenes is currently not available. This situation stimulated a substantial funding primarily in the USA for sequencing the genomes of several S. pyogenes serotype strains to facilitate the understanding of crucial pathogenicity traits of these bacteria and to identify promising targets for vaccine development. As a consequence, presently there are 12 strains of 8 ARTICLE IN PRESS www.elsevier.de/ijmm 1438-4221/$ - see front matter r 2007 Elsevier GmbH. All rights reserved. doi:10.1016/j.ijmm.2007.04.005 Corresponding author. Tel.: +49 38 1494 5900; fax: +49 38 1494 5902. E-mail address: [email protected] (A. Podbielski). 1 Both authors contributed equally to this publication. 2 Present address: Research Ctr., Ramathibodi Hospital, Mahidol University, Rama 6 Road, Bangkok 10400, Thailand.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ARTICLE IN PRESS
1438-4221/$ - se
doi:10.1016/j.ijm
�Correspondfax: +4938 149
E-mail addr
(A. Podbielski)1Both author2Present add
University, Ram
International Journal of Medical Microbiology 297 (2007) 513–523
www.elsevier.de/ijmm
Transcriptome analyses extend understanding of Streptococcus pyogenesregulatory mechanisms and behavior toward immunomodulatory
substances
Catur Riani1, Kerstin Standar1, Somboon Srimuang2, Cordula Lembke,Bernd Kreikemeyer, Andreas Podbielski�
Department of Medical Microbiology, Virology, and Hygiene, University Hospital Rostock, Schillingallee 70, D-18057 Rostock,
Germany
Received 30 August 2006; received in revised form 2 April 2007; accepted 4 April 2007
Abstract
The recent genome sequencing of several Streptococcus pyogenes serotype strains allowed the design ofcorresponding DNA microarrays and their usage for specific transcriptome analyses. In the present study, weemployed transcriptomics together with functional tests to investigate the impact of the CiaH sensor gene of theCiaRH two-component regulator on gene expression and virulence traits of serotype M49 S. pyogenes strains CS101and 591. In parallel, we studied the effects of the immunostimulatory substance Luivacs on the serotype M49 S.
pyogenes transcriptome and several biological features of serotype M1, M2, M3, M6, M18, and M49 S. pyogenes
strains. Overall, the transcriptome analyses allowed a swift identification of differences in transcript abundanceapparently associated with the observed strain-specific changes in matrix protein binding, eukaryotic cell interactions,or biofilm formation of the ciaH mutants and of wild-type strains exposed to a commercially available substance usedfor preventing upper respiratory tract infections.r 2007 Elsevier GmbH. All rights reserved.
Keywords: Streptococcus pyogenes; Transcriptomics; Cia regulator; Luivac immunostimulator; Virulence
Introduction
Purulent infections of the upper respiratory tract andskin caused by Streptococcus pyogenes (group Astreptococci, GAS) are extremely frequent, with about
e front matter r 2007 Elsevier GmbH. All rights reserved.
m.2007.04.005
ing author. Tel.: +4938 1494 5900;
4 5902.
ess: [email protected]
.
s contributed equally to this publication.
ress: Research Ctr., Ramathibodi Hospital, Mahidol
a 6 Road, Bangkok 10400, Thailand.
730 million yearly incidences worldwide. In addition,these bacteria trigger 940,000 yearly cases of post-infection sequelae and 660,000 yearly cases of life-threatening infections (Carapetis et al., 2005). Unlike forother similarly or less important pathogens, vaccinationagainst S. pyogenes is currently not available. Thissituation stimulated a substantial funding primarily inthe USA for sequencing the genomes of several S.
pyogenes serotype strains to facilitate the understandingof crucial pathogenicity traits of these bacteria and toidentify promising targets for vaccine development. As aconsequence, presently there are 12 strains of 8

ARTICLE IN PRESSC. Riani et al. / International Journal of Medical Microbiology 297 (2007) 513–523514
serotypes for which the complete genome sequenceshave been elucidated (see Beres et al. (2006) for completereference list).
Based on one to several of these genome sequences,several transcriptome analyses were performed thataddressed the activity of the two-component regulatorsCovRS, Ihk/Irs, and SptRS on the expression of the S.
pyogenes genes (Graham et al., 2002; Shelburne et al.,2005; Voyich et al., 2003), and the behavior of wild-typeS. pyogenes strains when exposed to different tempera-tures (Smoot et al., 2001) as well as environments suchas human blood and saliva or animal model infections(Cho and Caparon, 2005; Shelburne et al., 2005;Sitkiewicz and Musser, 2006). As a consequence, S.
pyogenes regulatory networks began to prevail. Addi-tionally, factors were identified that assist bacterialsurvival during defined steps of the pathogenesis process(Musser and DeLeo, 2006).
Supported by a grant of the German Ministry ofResearch and Technology, we sequenced major sectionsof the genome of the S. pyogenes serotype M49 strain591 and developed a DNA microarray based on thegenome sequences of S. pyogenes serotypes M1, M3,M18, and M49. Employing these microarrays, westudied the growth phase-associated S. pyogenes tran-scriptome (Beyer-Sehlmeyer et al., 2005) and thetranscriptomes of the AraC-type regulator MsmR(Nakata et al., 2005), the RALP-type stand-aloneregulator Nra (Kreikemeyer et al., submitted forpublication), and the three-component regulator Fas-ABC (Kreikemeyer et al., manuscript in preparation).Results showed growth phase-dependent shifts inexpression of sugar and amino acid transport systems,and an interplay between the MsmR and Nra regula-tors. Furthermore, we identified another Nra-dependentRALP-type regulator, which in turn controls the epf
plasminogen-binding surface protein and sagA hemoly-sin genes of a novel pathogenicity island.
In the present study, we show the applicability oftranscriptomics for basic and applied research byexamining the genes affected by the CiaRH two-component sensor–regulator pair and by an immuno-stimulatory substance, Luivacs, which is therapeuticallyadministered to prevent recurrent upper respiratorytract infections.
The CiaRH sensor–regulator (CiaH–CiaR) pair is oneof 12 two-component systems which are encoded byevery S. pyogenes genome sequenced so far. Homol-ogous (71–48% and 85–77% amino acid sequenceidentity for CiaH and CiaR, respectively) two-compo-nent systems have been identified in Streptococcus
pneumoniae, S. mitis, S. agalactiae, S. suis, S. mutans,and S. thermophilus genome sequences. CiaH pneumo-coccal mutants were initially identified by their increasedresistance to cefotaxime, a second-line drug used to treatinfections by penicillin-resistant pneumococci, and
simultaneously, by their impaired natural competence(Guenzi et al., 1994; Zahner et al., 1996). Since then,complex activities of CiaRH have been revealed. Thesystem is involved in competence development via Ca2+
sensing and pathways involving oxygen stress orComCDE regulation (Echenique et al., 2000; Echeniqueand Trombe, 2001; Giammarinaro et al., 1999; Zahneret al., 2002). CiaRH also influences the pneumococcalability to form protoplasts and the susceptibility todesoxycholate lysis as well as to late cell wall synthesisinhibitors (Giammarinaro et al., 1999; Mascher et al.,2006). In the meantime, the CiaRH transcriptome wasdescribed in three pneumococcal strains (Dagkessa-manskaja et al., 2004; Mascher et al., 2003; Sebert et al.,2002). It comprises up to 20 genes/operons involved incompetence development. In addition several stress-response genes are members of the CiaRH regulon,insinuating that competence development imposes stressupon pneumococcal cells that is balanced by the CiaRHregulatory activity (Ibrahim et al., 2004). Finally, thepneumococal capability for nasopharyngeal coloniza-tion is apparently associated with the interactionbetween CiaRH and the stress-induced protease HtrA(Ibrahim et al., 2004; Sebert et al., 2002).
In GAS, the function of the CiaRH homologue isunclear. Since the majority of GAS cells miss thecomplete set of genes for competence development, theCiaRH regulon in GAS most probably differs from thatof pneumococci. CiaRH was found to be part of theCovRS two-component regulator regulon in GAS.CovRS negatively affects expression of most virulencegenes and many stress-response genes in a mouse skininfection model, but not in culture media, the expressionof ciaRH (Graham et al., 2002).
Studying the transcriptome of regulators involvesanalysis of isogenic mutant strains. For the character-ization of transcriptomes associated with the activity oftherapeutic agents, several wild-type strains should beexamined. While the activities of antibiotics on GAS atinhibitory and sub-inhibitory doses have been identified,to our knowledge, the intrinsic antibacterial activities ofimmunostimulatory substances were not addressed sofar. The chemical nature of such a substance, Luivacs,is complex. It is prepared from seven lyophylizedbacterial species, i.e. Staphylococcus aureus, Streptococ-
cus mitis, S. pyogenes, Streptococcus pneumoniae,Klebsiella pneumoniae, Moraxella (Branhamella) catar-
rhalis, and Haemophilus influenzae. This preparation isorally administered as tablets and absorbed in the ileumand colon. Its clinical efficacy in the respiratory tract hasbeen demonstrated in several randomized, prospectivestudies (Jareoncharsri et al., 2003; Ruah et al., 2001;Steurer-Stey et al., 2004). Besides exerting a general notwell-understood effect on macrophage and T-helper cellactivity and increasing g-interferon and secretory IgAproduction (Ruedl et al., 1994; Spisek et al., 2004), the

ARTICLE IN PRESSC. Riani et al. / International Journal of Medical Microbiology 297 (2007) 513–523 515
complex substance could include compounds withgrowth-inhibitory and/or gene expression-modulatingeffects for S. pyogenes isolates.
In the present report, we extend our data collectionresulting from the BMBF-funded S. pyogenes transcrip-tome analyses by studying a defined S. pyogenes
regulator mutant and the effects of a commercialsubstance used for preventing recurrent S. pyogenes
infections. In both cases, this approach helped toidentify genes that are involved in the specific pheno-types demonstrated for the ciaRH mutants and theLuivacs-exposed cells.
Materials and methods
Bacterial strains and culture conditions
The GAS serotype M49 strains CS101 and 591 wereobtained from P.P. Cleary (Minneapolis, USA) and R.Lutticken (Aachen, Germany). The GAS serotypestrains M1, M2, M3, M6, and M18 were gifts fromthe Lancefield strain collection of the RockefellerUniversity (Podbielski et al., 1994). The Escherichia coli
strain DH5a (Gibco-BRL) was used as a host forplasmids pFW11 and pFW5-luc (Podbielski et al., 1999).All E. coli strains were cultured in Luria–Bertanimedium at 37 1C with agitation. The GAS wild-typestrains and isogenic derivatives were cultured inTodd–Hewitt broth (Invitrogen) supplemented with0.2% yeast extract (Invitrogen) (THY medium) at37 1C under a 5% CO2–20% O2 atmosphere, unlessotherwise specified. For selection and maintenance ofmutants, spectinomycin was added to the media at60 mg/ml each for E. coli and GAS.
DNA techniques
Purification of chromosomal and plasmid DNA,transformation of GAS, and other conventional manip-ulations were essentially done as described previously(Podbielski et al., 1999).
Construction of recombinant vectors
For the construction of the ciaH mutants and ciaRH-luciferase transcriptional fusions, fragments of theciaRH operon or the ciaRH promoter region wereamplified from M49 chromosomal template DNA withprimers cia-ko-FOR (50-GAT GAC GCT AGC GTGCTG ATG ATT ATC TGA CA-30), cia-ko-REV (50-GAT GAC GGA TCC GTT TTT CTG CTC CACTTG-30), cia-luc-FOR (50-GAT GAC GTC GAC CCTTAG TCA CTG AAC TTG-30), cia-luc-REV (50-GATGAC GGA TCC ATT GTT CTT AAA TAA AGT
AAC-30), and, via NheI/BamHI and SalI/BamHIextensions, subsequently cloned into the pFW11 orpFW5-luc vectors, respectively. Resulting recombinantplasmids were electroporated into GAS and the specificintegration of the constructs into the GAS chromosomewas confirmed by Southern blot hybridizations and site-specific PCRs.
Microarray analysis
For microarray analysis, two independent cultures ofeach S. pyogenes wild-type or mutant strain were grownto transition phase (OD600nm ¼ 1.0). Total RNA wasextracted with the RNeasy mini kit and RNase-freeDNase set (Qiagen) according to the manufacturer’sinstructions. RNA quality and integrity was assessed byelectrophoretic analysis and measurement of the A260/A280 ratios. Then, amino allyl-modified cDNA wassynthesized and labeled with CyDye using a CyScribePost-Labeling Kit (Amersham) according to the manu-facturer’s instructions. The DNA microarrays, contain-ing 50mer oligos designed from open reading framesequences of a partial serotype M49 genome sequence(NZ_AAFV00000000) and previously reported com-plete serotypes M1, M3, and M18 sequences, wasconstructed by MWG (Beyer-Sehlmeyer et al., 2005).Oligo chip slide processing for hybridization, scanning,image and statistical analyses were carried out aspreviously reported (Beyer-Sehlmeyer et al., 2005).
Quantitative assays for luciferase activity
The GAS luc reporter strains were cultured in THY orin chemically defined medium (CDM) in ambient air,under 5% CO2–20% O2 atmosphere, or under anaerobicconditions using the Anoxomat anaerobic culturesystem (Mart Microbiology). For different oxygen stressconditions, cultures in ambient air were performed asstanding cultures or with vigorous shaking at 200rotations per minute (rpm).
To measure luminescence, aliquots from bacterialcultures were withdrawn at 1-h intervals and processedas described previously (Podbielski et al., 1999). Formeasuring the influence of divalent cations on ciaRH
transcription rates, Cu, Co, Fe, Mg, Mn, and Zn ionswere added to the CDM at 3–6 different finalconcentrations as indicated in the results section.
Determination of antibiotic resistance/activity
For measuring the resistance of GAS wild-type andciaR-mutant strains to selected antibiotics, E-tests wereperformed according to the instructions of the manu-facturer (Sigma).

ARTICLE IN PRESSC. Riani et al. / International Journal of Medical Microbiology 297 (2007) 513–523516
The inherent antibiotic activity of Luivacs wasexamined by dissolving this substance in phosphate-buffered saline (PBS), pH 7.4, to 100mg/ml and addingappropriate aliquots from this stock solution to theculture medium prior to inoculation with the testbacteria. Bacterial numbers used for seeding into thetest wells and subsequently encountered over the growthcycle were determined by drawing aliquots at hourlyintervals, preparing serial dilutions in PBS, plating100 ml aliquots onto Columbia sheep blood agar, andcounting colonies after overnight incubation.
In parallel, potential antibacterial and anti-virulenceactivities of Luivacs were assessed by modified bacte-riocin assays. Therefore, 100 ml aliquots of Luivacs
dissolved in PBS to final concentrations of 1, 0.1, and0.001 g/l were applied in lines onto Columbia sheepblood agar. After drying of these lines of dissolvedLuivacs for 1 h at room temperature, 10-ml aliquotsfrom overnight cultures containing the suspended testbacteria were placed in perpendicular lines onto theagar. The agar plates were incubated overnight under a5% CO2–20% O2 atmosphere, and inspected thefollowing day for inhibition zones and changes inhemolysin production at the intersections of the lines.
Matrix protein-binding assays
The binding of GAS strains to immobilized humanmatrix proteins (Sigma) was measured as previouslydescribed (Kreikemeyer et al., 2004).
Eukaryotic cell adherence and internalization
Human epithelial HEp-2 (ATCC CCL23) cells weremaintained in Dulbecco’s modified Eagle medium(Gibco) supplemented with 10% fetal bovine serum(Gibco). Adherence to and internalization by HEp-2cells was assessed by the antibiotic protection assay andby the double immunofluorescence staining assay asspecified by Kreikemeyer et al. (2004). In each case,results were obtained from at least three independentassays.
Where indicated, viability of the eukaryotic host cellsexposed to either GAS strains or Luivacs was assessedby using the Live/Dead staining kit (Molecular Probes,MoBiTec) according to the manufacturer’s instructions.
Biofilm assays
Biofilm formation and organization of GAS serotypestrains M2, M6, and M18 in the absence and presence ofLuivacs at 0.001, 0.1, 1, and 10 g/l were observed understatic conditions using uncoated (M6), collagen type I-coated (M18) or fibronectin-coated (M2) polystyrene24-well plates or coverslips (Cellstar, Greiner bio-one;
Thermanox, Nunc), brain–heart infusion medium, and aCO2-enriched atmosphere for periods up to 72 h ofincubation. Biofilm density and ultrastructure weredetermined by safranin staining assays and scanningelectron microscopy (SEM) as described by Lembkeet al. (2006).
Statistical analysis
When indicated, statistical analysis was performedwith Student’s t-test using StatView software (SASInstitute). A confidence interval of po0.05 was con-sidered significant.
Results
CiaRH expression profiles
In order to identify the peak expression of ciaRH forsubsequent analyses of its maximum impact on S.
pyogenes gene expression and biological functions and,additionally, to record potential effects of environmen-tal conditions on the ciaRH expression profile, aluciferase reporter gene box was introduced into theciaRH promoter region of serotype M49 GAS strainsCS101 and 591. As a consequence, the ciaRH promoterregion in these strains was duplicated, leaving ciaRH
under undisturbed control of its orthologous promoter.The cia-luc fusion strains were incubated as batch
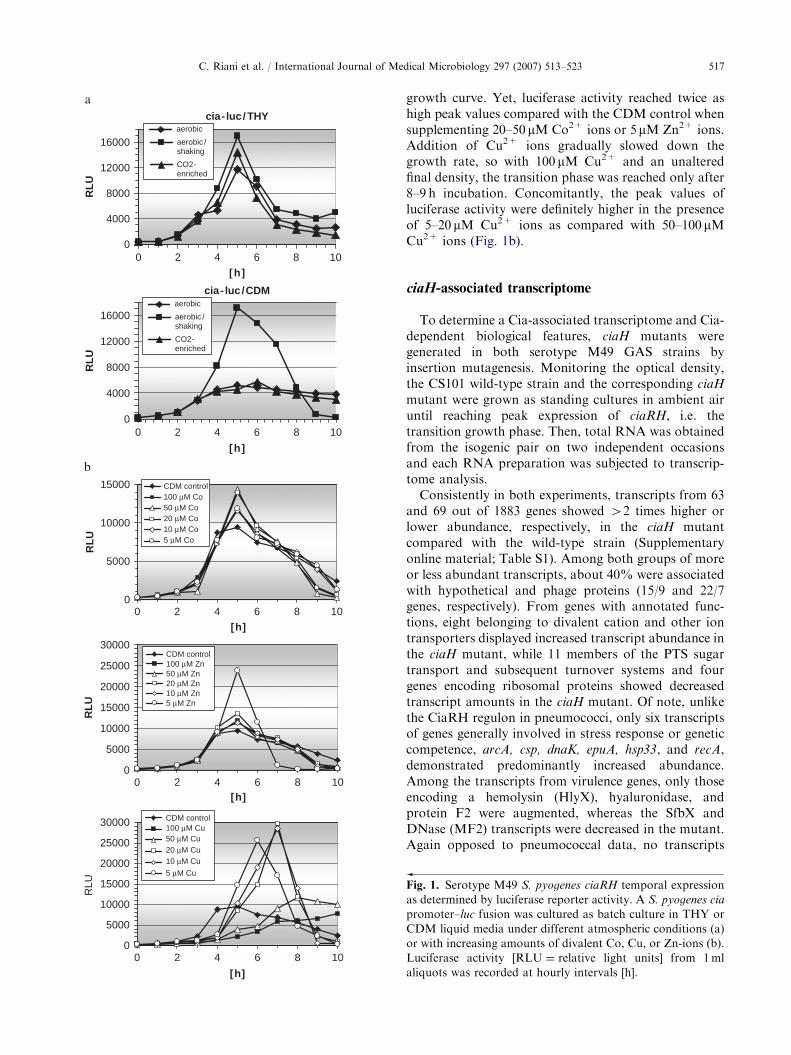
cultures under aerobic, CO2-enriched, and anaerobicatmospheres. Within the reproducibility range of thismethod, there was no difference between the luciferaseexpression profiles in both strains and under thedifferent atmospheric conditions except for strainsgrown in CDM with vigorous shaking (200 rpm) underan aerobic atmosphere (Fig. 1a). Using both THY andCDM broths as growth medium, the luciferase reporteractivity displayed a relatively sharp increase to max-imum activity in the transition growth phase after 4–5 hof incubation and subsequently in the stationary phaseup to 10 h of incubation, a constant decrease to abouthalf to a fifth of the peak values. However, growth inTHY broth led to luciferase peak values three timeshigher than in CDM (Fig. 1a).
Since Ca2+ was shown to influence the pneumococcalCiaRH activity, the influence of divalent cations onGAS ciaRH expression was measured by incubating thecia-luc fusion strains in CDM with defined supplementsof selected divalent cations. Fe2+ ions in concentrationsup to 500 mM as well as Mg2+ and Mn2+ ions inconcentrations up to 72 mM had no effects both ongrowth and on ciaRH expression as measured byluciferase activity (data not shown). Co2+ and Zn2+
ions in concentrations up to 100 mM did not affect the
ARTICLE IN PRESS
cia-luc / THY
0
4000
8000
12000
16000
0 2 6 8 10
[h]
RL
U
0
4000
8000
12000
16000
RL
U
aerobic
aerobic /shaking
CO2-enriched
cia- luc /CDM
0
5000
10000
15000
RL
UR
LU
CDM control
100 µM Co
50 µM Co
20 µM Co
10 µM Co
5 µM Co
0
5000
10000
15000
20000
25000
30000
0 2 4 6 8 10
RLU
0
5000
10000
15000
20000
25000
30000
0 2 4 6 8 10
[h]
[h]
[h]
CDM control
100 µM Zn
50 µM Zn
20 µM Zn 10 µM Zn 5 µM Zn
CDM control
100 µM Cu
50 µM Cu
20 µM Cu
10 µM Cu
5 µM Cu
4
0 2 6 8 10
[h]
4
0 2 6 8 104
aerobic
aerobic /shaking
CO2-enriched
a
b
C. Riani et al. / International Journal of Medical Microbiology 297 (2007) 513–523 517
growth curve. Yet, luciferase activity reached twice ashigh peak values compared with the CDM control whensupplementing 20–50 mM Co2+ ions or 5 mM Zn2+ ions.Addition of Cu2+ ions gradually slowed down thegrowth rate, so with 100 mM Cu2+ and an unalteredfinal density, the transition phase was reached only after8–9 h incubation. Concomitantly, the peak values ofluciferase activity were definitely higher in the presenceof 5–20 mM Cu2+ ions as compared with 50–100 mMCu2+ ions (Fig. 1b).
ciaH-associated transcriptome
To determine a Cia-associated transcriptome and Cia-dependent biological features, ciaH mutants weregenerated in both serotype M49 GAS strains byinsertion mutagenesis. Monitoring the optical density,the CS101 wild-type strain and the corresponding ciaH
mutant were grown as standing cultures in ambient airuntil reaching peak expression of ciaRH, i.e. thetransition growth phase. Then, total RNA was obtainedfrom the isogenic pair on two independent occasionsand each RNA preparation was subjected to transcrip-tome analysis.
Consistently in both experiments, transcripts from 63and 69 out of 1883 genes showed 42 times higher orlower abundance, respectively, in the ciaH mutantcompared with the wild-type strain (Supplementaryonline material; Table S1). Among both groups of moreor less abundant transcripts, about 40% were associatedwith hypothetical and phage proteins (15/9 and 22/7genes, respectively). From genes with annotated func-tions, eight belonging to divalent cation and other iontransporters displayed increased transcript abundance inthe ciaH mutant, while 11 members of the PTS sugartransport and subsequent turnover systems and fourgenes encoding ribosomal proteins showed decreasedtranscript amounts in the ciaH mutant. Of note, unlikethe CiaRH regulon in pneumococci, only six transcriptsof genes generally involved in stress response or geneticcompetence, arcA, csp, dnaK, epuA, hsp33, and recA,demonstrated predominantly increased abundance.Among the transcripts from virulence genes, only thoseencoding a hemolysin (HlyX), hyaluronidase, andprotein F2 were augmented, whereas the SfbX andDNase (MF2) transcripts were decreased in the mutant.Again opposed to pneumococcal data, no transcripts
Fig. 1. Serotype M49 S. pyogenes ciaRH temporal expression
as determined by luciferase reporter activity. A S. pyogenes cia
promoter–luc fusion was cultured as batch culture in THY or
CDM liquid media under different atmospheric conditions (a)
or with increasing amounts of divalent Co, Cu, or Zn-ions (b).
Luciferase activity [RLU ¼ relative light units] from 1ml
aliquots was recorded at hourly intervals [h].
ARTICLE IN PRESS
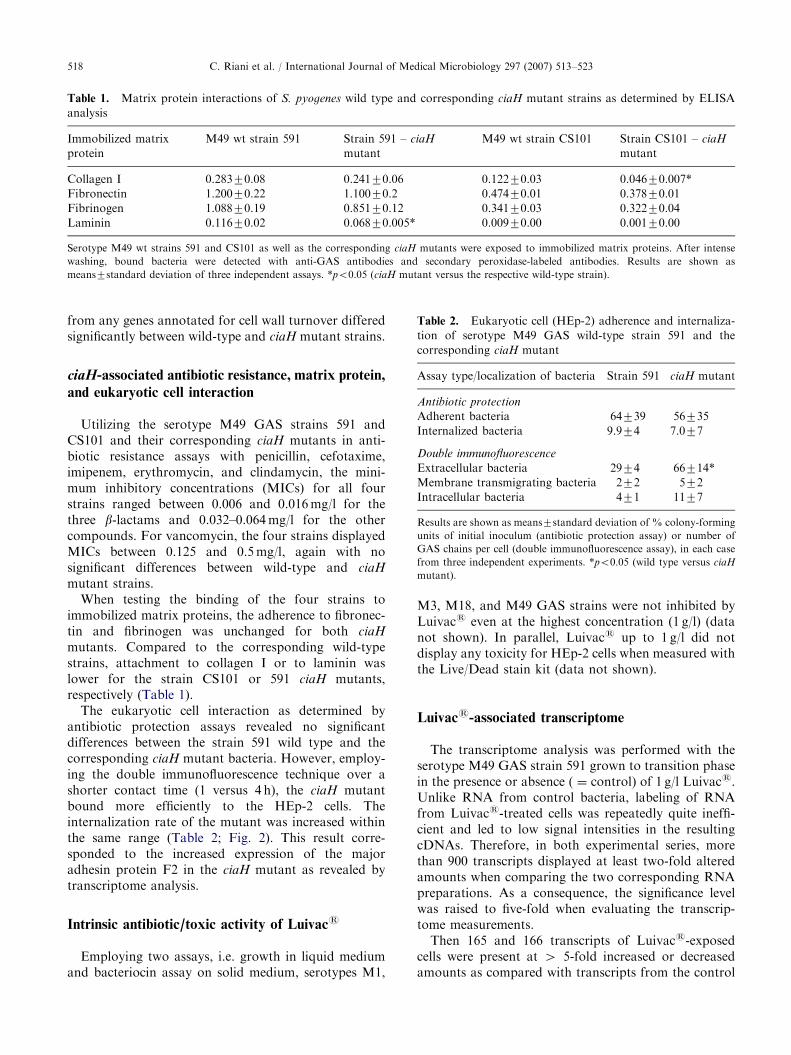
Table 1. Matrix protein interactions of S. pyogenes wild type and corresponding ciaH mutant strains as determined by ELISA
analysis
Immobilized matrix
protein
M49 wt strain 591 Strain 591 – ciaH
mutant
M49 wt strain CS101 Strain CS101 – ciaH
mutant
Collagen I 0.28370.08 0.24170.06 0.12270.03 0.04670.007*
Fibronectin 1.20070.22 1.10070.2 0.47470.01 0.37870.01
Fibrinogen 1.08870.19 0.85170.12 0.34170.03 0.32270.04
Laminin 0.11670.02 0.06870.005* 0.00970.00 0.00170.00
Serotype M49 wt strains 591 and CS101 as well as the corresponding ciaH mutants were exposed to immobilized matrix proteins. After intense
washing, bound bacteria were detected with anti-GAS antibodies and secondary peroxidase-labeled antibodies. Results are shown as
means7standard deviation of three independent assays. *po0.05 (ciaH mutant versus the respective wild-type strain).
Table 2. Eukaryotic cell (HEp-2) adherence and internaliza-
tion of serotype M49 GAS wild-type strain 591 and the
corresponding ciaH mutant
Assay type/localization of bacteria Strain 591 ciaH mutant
Antibiotic protection
Adherent bacteria 64739 56735
Internalized bacteria 9.974 7.077
Double immunofluorescence
Extracellular bacteria 2974 66714*
Membrane transmigrating bacteria 272 572
Intracellular bacteria 471 1177
Results are shown as means7standard deviation of % colony-forming
units of initial inoculum (antibiotic protection assay) or number of
GAS chains per cell (double immunofluorescence assay), in each case
from three independent experiments. *po0.05 (wild type versus ciaH
mutant).
C. Riani et al. / International Journal of Medical Microbiology 297 (2007) 513–523518
from any genes annotated for cell wall turnover differedsignificantly between wild-type and ciaH mutant strains.
ciaH-associated antibiotic resistance, matrix protein,
and eukaryotic cell interaction
Utilizing the serotype M49 GAS strains 591 andCS101 and their corresponding ciaH mutants in anti-biotic resistance assays with penicillin, cefotaxime,imipenem, erythromycin, and clindamycin, the mini-mum inhibitory concentrations (MICs) for all fourstrains ranged between 0.006 and 0.016mg/l for thethree b-lactams and 0.032–0.064mg/l for the othercompounds. For vancomycin, the four strains displayedMICs between 0.125 and 0.5mg/l, again with nosignificant differences between wild-type and ciaH
mutant strains.When testing the binding of the four strains to
immobilized matrix proteins, the adherence to fibronec-tin and fibrinogen was unchanged for both ciaH
mutants. Compared to the corresponding wild-typestrains, attachment to collagen I or to laminin waslower for the strain CS101 or 591 ciaH mutants,respectively (Table 1).
The eukaryotic cell interaction as determined byantibiotic protection assays revealed no significantdifferences between the strain 591 wild type and thecorresponding ciaH mutant bacteria. However, employ-ing the double immunofluorescence technique over ashorter contact time (1 versus 4 h), the ciaH mutantbound more efficiently to the HEp-2 cells. Theinternalization rate of the mutant was increased withinthe same range (Table 2; Fig. 2). This result corre-sponded to the increased expression of the majoradhesin protein F2 in the ciaH mutant as revealed bytranscriptome analysis.
Intrinsic antibiotic/toxic activity of Luivacs
Employing two assays, i.e. growth in liquid mediumand bacteriocin assay on solid medium, serotypes M1,
M3, M18, and M49 GAS strains were not inhibited byLuivacs even at the highest concentration (1 g/l) (datanot shown). In parallel, Luivacs up to 1 g/l did notdisplay any toxicity for HEp-2 cells when measured withthe Live/Dead stain kit (data not shown).
Luivacs-associated transcriptome
The transcriptome analysis was performed with theserotype M49 GAS strain 591 grown to transition phasein the presence or absence ( ¼ control) of 1 g/l Luivacs.Unlike RNA from control bacteria, labeling of RNAfrom Luivacs-treated cells was repeatedly quite ineffi-cient and led to low signal intensities in the resultingcDNAs. Therefore, in both experimental series, morethan 900 transcripts displayed at least two-fold alteredamounts when comparing the two corresponding RNApreparations. As a consequence, the significance levelwas raised to five-fold when evaluating the transcrip-tome measurements.
Then 165 and 166 transcripts of Luivacs-exposedcells were present at 4 5-fold increased or decreasedamounts as compared with transcripts from the control

ARTICLE IN PRESS
Fig. 2. Eukaryotic (HEp-2) cell interaction of a S. pyogenes
wild-type strain (a) and corresponding ciaH mutant (b) as
determined by double immunofluorescence microscopy. The
staining and visualization was performed according to the
protocol described by Kreikemeyer et al. (2004). Intracellular
and extracellular bacteria are stained red or light green,
respectively. Nuclei of HEp-2 cells appear as blue due to DAPI
staining.
C. Riani et al. / International Journal of Medical Microbiology 297 (2007) 513–523 519
bacteria. Among the augmented mRNA molecules,virulence-associated mRNA species included thoseencoding the superantigens SpeI and Mf4, cysteinprotease SpeB, and the global positive regulator Mga.Decreased virulence messages included capsule synthesisgene hasA, plasminogen-binding GAPDH and enolase,superantigen SpeA, collagen-like protein Scl1, and aRALP-type regulator (Supplementary online material,Table S2).
Luivacs-associated eukaryotic cell interaction
For testing eukaryotic cell adherence and internaliza-tion by the antibiotic protection assay, Luivacs was
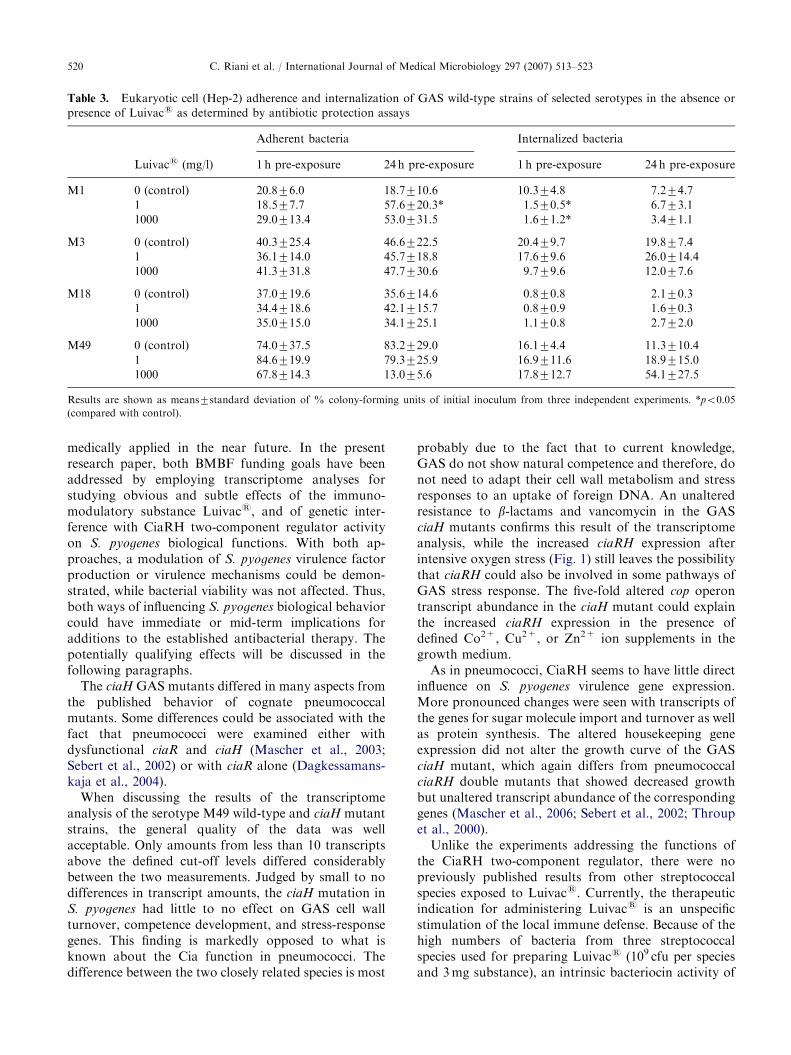
added at 0.001 and 1 g/l to the eukaryotic cell culturemedium 1 or 24 h before exposure to serotype M1, M3,M18, and M49 GAS strains. The resulting picture wascomplex. While pre-exposure to Luivacs had noinfluence on the eukaryotic cell interaction of theserotypes M18 and M49 S. pyogenes strains, the M1and M3 strains displayed increased adherence ordecreased internalization rates, respectively (Table 3).For the M1 and M3 strains, the effects were morepronounced after 24 or 1 h Luivacs pre-exposure,respectively.
Luivacs-associated GAS biofilms
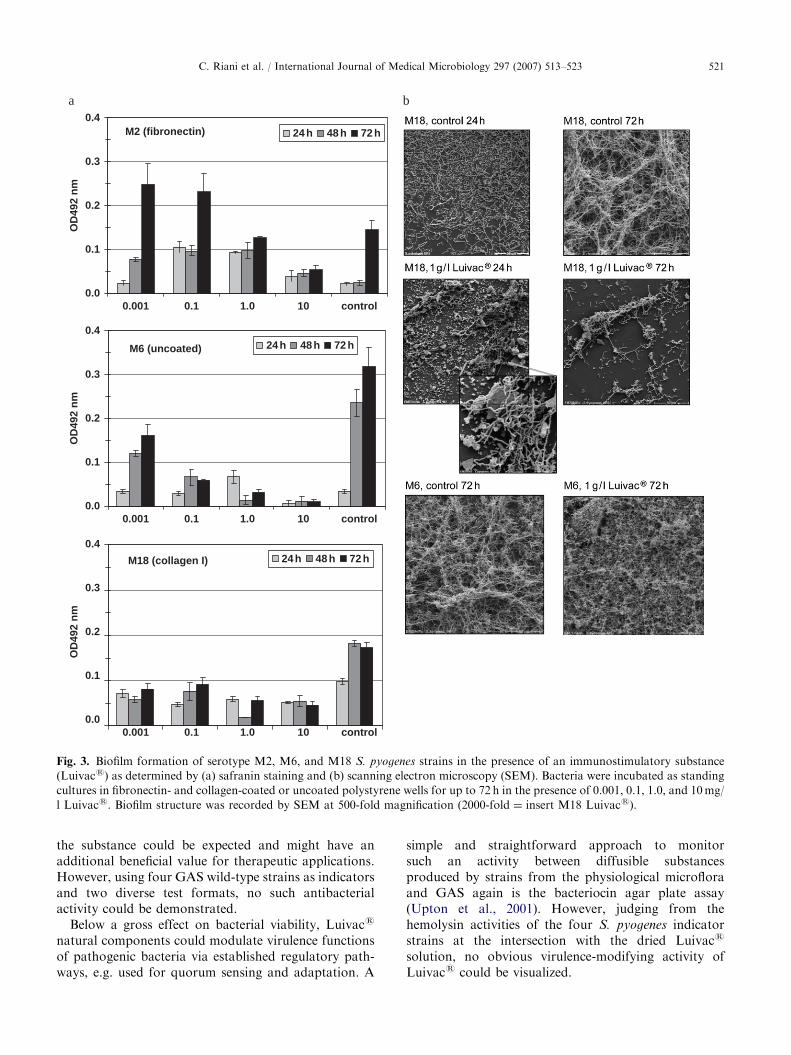
Using Luivacs at concentrations up to 10 g/l, a dose-dependent decrease in biofilm mass could be visualizedespecially after 72 h of incubation by measuring safraninstaining intensity for the serotype M6 GAS strain(Fig. 3a). Similar but less pronounced effects could beshown for the serotype M2 GAS strain with Luivacs atconcentrations of 1 and 10 g/l. Concomitantly, thinnerbiofilms were seen by SEM especially after 72 hincubation (Fig. 3b), confirming the assessments ofsafranin-stained biofilms.
The M18 isolate displayed decreased biofilm amountseven at the earliest time point and the lowest Luivacs
concentration. The finding that these biofilms wereextremely sensitive to washing procedures and the SEM-based demonstration of incompletely degraded bacteriafrom the Luivacs preparation integrated into thebiofilm (insert at 2000-fold magnification) insinuated adose-independent interference of Luivacs with serotypeM18 GAS biofilm formation (Fig. 3b).
Discussion
The pathogenic role of Streptococcus pyogenes inhuman infectious diseases is currently reduced byadministration of antibiotics with little to no selectivitytoward the related physiological microflora. Specificmanipulation of the bacterial virulence mechanismswithout dramatic effects on bacterial viability could bean attractive alternative for the treatment of patientswith a functional immune defense. Substances with sucheffects either still need to be developed or may befeasible but yet await characterization of this desirablefeature.
Progress for resolving this problem could come fromBMBF-funded research, since a primary goal of BMBFfunding is to stimulate investigations focused on thetransformation of results from basic research into atechnical or medical application. As a secondary goal,BMBF funding will also support basic research if it isdirected to produce results that could be technically or
ARTICLE IN PRESS
Table 3. Eukaryotic cell (Hep-2) adherence and internalization of GAS wild-type strains of selected serotypes in the absence or
presence of Luivacs as determined by antibiotic protection assays
Adherent bacteria Internalized bacteria
Luivacs (mg/l) 1 h pre-exposure 24 h pre-exposure 1 h pre-exposure 24 h pre-exposure
M1 0 (control) 20.876.0 18.7710.6 10.374.8 7.274.7
1 18.577.7 57.6720.3* 1.570.5* 6.773.1
1000 29.0713.4 53.0731.5 1.671.2* 3.471.1
M3 0 (control) 40.3725.4 46.6722.5 20.479.7 19.877.4
1 36.1714.0 45.7718.8 17.679.6 26.0714.4
1000 41.3731.8 47.7730.6 9.779.6 12.077.6
M18 0 (control) 37.0719.6 35.6714.6 0.870.8 2.170.3
1 34.4718.6 42.1715.7 0.870.9 1.670.3
1000 35.0715.0 34.1725.1 1.170.8 2.772.0
M49 0 (control) 74.0737.5 83.2729.0 16.174.4 11.3710.4
1 84.6719.9 79.3725.9 16.9711.6 18.9715.0
1000 67.8714.3 13.075.6 17.8712.7 54.1727.5
Results are shown as means7standard deviation of % colony-forming units of initial inoculum from three independent experiments. *po0.05
(compared with control).
C. Riani et al. / International Journal of Medical Microbiology 297 (2007) 513–523520
medically applied in the near future. In the presentresearch paper, both BMBF funding goals have beenaddressed by employing transcriptome analyses forstudying obvious and subtle effects of the immuno-modulatory substance Luivacs, and of genetic inter-ference with CiaRH two-component regulator activityon S. pyogenes biological functions. With both ap-proaches, a modulation of S. pyogenes virulence factorproduction or virulence mechanisms could be demon-strated, while bacterial viability was not affected. Thus,both ways of influencing S. pyogenes biological behaviorcould have immediate or mid-term implications foradditions to the established antibacterial therapy. Thepotentially qualifying effects will be discussed in thefollowing paragraphs.
The ciaH GAS mutants differed in many aspects fromthe published behavior of cognate pneumococcalmutants. Some differences could be associated with thefact that pneumococci were examined either withdysfunctional ciaR and ciaH (Mascher et al., 2003;Sebert et al., 2002) or with ciaR alone (Dagkessamans-kaja et al., 2004).
When discussing the results of the transcriptomeanalysis of the serotype M49 wild-type and ciaH mutantstrains, the general quality of the data was wellacceptable. Only amounts from less than 10 transcriptsabove the defined cut-off levels differed considerablybetween the two measurements. Judged by small to nodifferences in transcript amounts, the ciaH mutation inS. pyogenes had little to no effect on GAS cell wallturnover, competence development, and stress-responsegenes. This finding is markedly opposed to what isknown about the Cia function in pneumococci. Thedifference between the two closely related species is most
probably due to the fact that to current knowledge,GAS do not show natural competence and therefore, donot need to adapt their cell wall metabolism and stressresponses to an uptake of foreign DNA. An unalteredresistance to b-lactams and vancomycin in the GASciaH mutants confirms this result of the transcriptomeanalysis, while the increased ciaRH expression afterintensive oxygen stress (Fig. 1) still leaves the possibilitythat ciaRH could also be involved in some pathways ofGAS stress response. The five-fold altered cop operontranscript abundance in the ciaH mutant could explainthe increased ciaRH expression in the presence ofdefined Co2+, Cu2+, or Zn2+ ion supplements in thegrowth medium.
As in pneumococci, CiaRH seems to have little directinfluence on S. pyogenes virulence gene expression.More pronounced changes were seen with transcripts ofthe genes for sugar molecule import and turnover as wellas protein synthesis. The altered housekeeping geneexpression did not alter the growth curve of the GASciaH mutant, which again differs from pneumococcalciaRH double mutants that showed decreased growthbut unaltered transcript abundance of the correspondinggenes (Mascher et al., 2006; Sebert et al., 2002; Throupet al., 2000).
Unlike the experiments addressing the functions ofthe CiaRH two-component regulator, there were nopreviously published results from other streptococcalspecies exposed to Luivacs. Currently, the therapeuticindication for administering Luivacs is an unspecificstimulation of the local immune defense. Because of thehigh numbers of bacteria from three streptococcalspecies used for preparing Luivacs (109 cfu per speciesand 3mg substance), an intrinsic bacteriocin activity of
ARTICLE IN PRESS
0.0
0.1
0.2
0.3
0.4
0.001 0.1 1.0 10 control
OD
492 n
m
0.0
0.1
0.2
0.3
0.4
0.001 0.1 1.0 10 control
OD
492 n
m
0.0
0.1
0.2
0.3
0.4
0.001 0.1 1.0 10 control
OD
492 n
m
24 h 48 h 72 h
24 h 48 h 72 h
24 h 48 h 72 h
M6 (uncoated)
M18 (collagen I)
M2 (fibronectin)
a b
Fig. 3. Biofilm formation of serotype M2, M6, and M18 S. pyogenes strains in the presence of an immunostimulatory substance
(Luivacs) as determined by (a) safranin staining and (b) scanning electron microscopy (SEM). Bacteria were incubated as standing
cultures in fibronectin- and collagen-coated or uncoated polystyrene wells for up to 72 h in the presence of 0.001, 0.1, 1.0, and 10mg/
l Luivacs. Biofilm structure was recorded by SEM at 500-fold magnification (2000-fold ¼ insert M18 Luivacs).
C. Riani et al. / International Journal of Medical Microbiology 297 (2007) 513–523 521
the substance could be expected and might have anadditional beneficial value for therapeutic applications.However, using four GAS wild-type strains as indicatorsand two diverse test formats, no such antibacterialactivity could be demonstrated.
Below a gross effect on bacterial viability, Luivacs
natural components could modulate virulence functionsof pathogenic bacteria via established regulatory path-ways, e.g. used for quorum sensing and adaptation. A
simple and straightforward approach to monitorsuch an activity between diffusible substancesproduced by strains from the physiological microfloraand GAS again is the bacteriocin agar plate assay(Upton et al., 2001). However, judging from thehemolysin activities of the four S. pyogenes indicatorstrains at the intersection with the dried Luivacs
solution, no obvious virulence-modifying activity ofLuivacs could be visualized.

ARTICLE IN PRESSC. Riani et al. / International Journal of Medical Microbiology 297 (2007) 513–523522
It took the panel of transcriptome analysis andfunctional tests, such as eukaryotic cell interaction andbiofilm formation, to demonstrate that Luivacs exerteda more subtle activity on the altogether six tested GASstrains and affected more complex biological functions.
When monitoring eukaryotic cell adherence andinternalization of GAS in the presence of Luivacs, itbecame obvious that the substance exerted its effects in astrain-specific manner. The most pronounced differ-ences were seen with the serotype M1 strain, whichbound at much higher levels to Luivacs-pretreated cells,but internalized with much lower efficiency. Accordingto the results of the transcriptome analysis, this is mostprobably achieved by priming the eukaryotic target cellsand not by influencing the bacterial adhesin expression,although a direct modifying activity of Luivacs onexposed GAS surface proteins cannot be excluded onthe basis of the experimental setup.
Another GAS strain-specific effect of Luivacs couldbe observed when examining its influence on biofilmformation. Since only a subset of GAS strains is able toform biofilms, the panel of tested strains is not identicalto that of the eukaryotic cell interaction trials. This time,the observed effects appear to depend on a directactivity of Luivacs on the single GAS strains, since theSEM pictures independently from Luivacs absence orpresence showed an even initial adherence of the M6and M2 isolates to the crude polystyrene or fibronectin-coated supporting material, respectively, but subse-quently decreased biofilm formation in both settings.This direct effect of Luivacs on GAS may complementits immunostimulatory function, since GAS biofilmscould be associated with recurrent, antibiotic therapy-resistant respiratory infections (Baldassarri et al.,2006).
Unlike previous transcriptome analyses on selectedGAS strains, the presence of Luivacs altered theamount and/or quality of the obtained total RNA inaddition to influencing the abundance of several crucialvirulence factors. The mechanistic background of thisaction is unclear. However, the effect could indicateanother intrinsic activity of this substance against GAS.So in addition to its immunomodulatory effects,Luivacs could contribute to directly fight infectionscaused by a subset of S. pyogenes strains.
In conclusion, both sets of analyses document thattranscriptome analyses reveal specific effects of muta-tions or growth medium supplements, i.e. immuno- orvirulence-modulatory substances, that would be misseddue to the absence of an obvious phenotype in standardassays for biological functions. Thus, transcriptomicscould direct researchers to implement more specificfunctional tests. Of course, transcriptome analysis alsosupports the understanding of a molecular backgroundin case functional assays had already revealed mutation-or growth supplement-associated differences.
Acknowledgments
The authors thank Ms. Jana Normann for herconsistently excellent technical assistance. We are alsograteful to the Research Institute for the Biology ofFarm Animals, Dummerstorf, and L. Jonas for grantingaccess to the microarray reader and scanning electronmicroscope, respectively. The work of C. Lembke, A.Podbielski, and K. Standar was supported by the BMBFPathoGenomik grant PTJ-BIO/0313134. In addition, C.Riani was supported by the DAAD grant A/04/42567and S. Srimuang by the DAAD grant A/02/06858.
Appendix A. Supplementary materials
The online version of this article contains addi-tional supplementary data. Please visit doi:10.1016/j.ijmm.2007.04.005.
References
Baldassarri, L., Creti, R., Recchia, S., Imperi, M., Facinelli,
B., Giovanetti, E., Pataracchia, M., Alfarone, G., Orefici,
G., 2006. Therapeutic failures of antibiotics used to treat
macrolide-susceptible Streptococcus pyogenes infections
may be due to biofilm formation. J. Clin. Microbiol. 44,
2721–2727.
Beres, S.B., Richter, E.W., Nagiec, M.J., Sumby, P., Porcella,
S.F., DeLeo, F.R., Musser, J.M., 2006. Molecular genetic
anatomy of inter- and intraserotype variation in the human
bacterial pathogen group A Streptococcus. Proc. Natl.
Acad. Sci. USA 103, 7059–7064.
Beyer-Sehlmeyer, G., Kreikemeyer, B., Horster, A., Podbiels-
ki, A., 2005. Analysis of the growth phase-associated
transcriptome of Streptococcus pyogenes. Int. J. Med.
Microbiol. 295, 161–177.
Carapetis, J.R., Steer, A.C., Mulholland, E.K., Weber, M.,
2005. The global burden of group A streptococcal diseases.
Lancet Infect. Dis. 5, 685–694.
Cho, K.H., Caparon, M.G., 2005. Patterns of virulence gene
expression differ between biofilm and tissue communities of
Streptococcus pyogenes. Mol. Microbiol. 57, 1545–1556.
Dagkessamanskaja, A., Moscoso, M., Henard, V., Guiral, S.,
Overweg, K., Reuter, M., Martin, B., Wells, J., Claverys,
J.P., 2004. Interconnection of competence, stress and CiaR
regulons in Streptococcus pneumoniae: competence triggers
stationary phase autolysis of ciaR mutant cells. Mol.
Microbiol. 51, 1071–1086.
Echenique, J.R., Trombe, M.C., 2001. Competence modula-
tion by the NADH oxidase of Streptococcus pneumoniae
involves signal transduction. J. Bacteriol. 183, 768–772.
Echenique, J.R., Chapuy-Regaud, S., Trombe, M.C., 2000.
Competence regulation by oxygen in Streptococcus pneu-
moniae: involvement of ciaRH and comCDE. Mol. Micro-
biol. 36, 688–696.

ARTICLE IN PRESSC. Riani et al. / International Journal of Medical Microbiology 297 (2007) 513–523 523
Giammarinaro, P., Sicard, M., Gasc, A.M., 1999. Genetic and
physiological studies of the CiaH-CiaR two-component
signal-transducing system involved in cefotaxime resistance
and competence of Streptococcus pneumoniae. Microbiol-
ogy 145, 1859–1869.
Graham, M.R., Smoot, L.M., Migliaccio, C.A.L., Virtaneva,
K., Sturdevant, D.E., Porcella, S.F., Federle, M.J., Adams,
G.J., Scott, J.R., Musser, J.M., 2002. Virulence control in
group A Streptococcus by a two-component gene regula-
tory system: global expression profiling and in vivo
infection modeling. Proc. Natl. Acad. Sci. USA 99,
13855–13860.
Guenzi, E., Gasc, A.M., Sicard, M.A., Hakenbeck, R., 1994. A
two-component signal-transducing system is involved in
competence and penicillin susceptibility in laboratory
mutants of Streptococcus pneumoniae. Mol. Microbiol. 12,
505–515.
Ibrahim, Y.M., Kerr, A.R., McCluskey, J., Mitchell, T.J.,
2004. Control of virulence by the two-component system
CiaR/H is mediated via HtrA, a major virulence factor of
Streptococcus pneumoniae. J. Bacteriol. 186, 5258–5266.
Jareoncharsri, P., Bunnag, C., Tunsuriyawong, P., Assana-
sane, P., Voraprayoon, S., Pinkaew, B., 2003. An open-
label, prospective study of an oral polyvalent bacterial
lysate (Luivac) in the treatment of recurrent respiratory
tract infections in Thai patients. Asian Pac. J. Allergy
Immunol. 21, 223–230.
Kreikemeyer, B., Oehmcke, S., Nakata, M., Hofrogge, R.,
Podbielski, A., 2004. Streptococcus pyogenes fibronectin-
binding protein F2: expression profile, binding character-
istics and impact on eukaryotic cell interactions. J. Biol.
Chem. 279, 15850–15859.
Lembke, C., Podbielski, A., Hidalgo-Grass, C., Jonas, L.,
Hanski, E., Kreikemeyer, B., 2006. Characterization of
biofilm formation by clinically relevant serotypes of
group A streptococci. Appl. Environ. Microbiol. 72,
2864–2875.
Mascher, T., Zahner, D., Merai, M., Balmelle, N., de Saizieu,
A., Hakenbeck, R., 2003. The Streptococcus pneumoniae cia
regulon: Cia target sites and transcription profile analysis.
J. Bacteriol. 185, 60–70.
Mascher, T., Heintz, M., Zahner, D., Merai, M., Hakenbeck,
R., 2006. The CiaRH system of Streptococcus pneumoniae
prevents lysis during stress induced by treatment with cell
wall inhibitors and by mutations in pbp2x involved in beta-
lactam resistance. J. Bacteriol. 188, 1959–1968.
Musser, J.M., DeLeo, F.R., 2006. Toward a genome-wide
systems biology analysis of host-pathogen interactions in
group A Streptococcus. Am. J. Pathol. 167, 1461–1471.
Nakata, M., Podbielski, A., Kreikemeyer, B., 2005. MsmR, a
specific positive regulator of the Streptococcus pyogenes
FCT pathogenicity region and cytolysin-mediated translo-
cation system genes. Mol. Microbiol. 57, 786–803.
Podbielski, A., Weber-Heynemann, J., Cleary, P.P., 1994.
Immunoglobulin-binding FcrA and Enn proteins, and M
proteins of group A streptococci evolved independently
from a common ancestral protein. Med. Microbiol.
Immunol. 183, 33–42.
Podbielski, A., Woischnik, M., Leonard, B.A.B., Schmidt,
K.H., 1999. Characterization of nra, a global negative
regulator gene in group A streptococci. Mol. Microbiol. 31,
1051–1064.
Ruah, S.B., Ruah, C., van Aubel, A., Abel, S., Elsasser, U.,
2001. Efficacy of a polyvalent bacterial lysate in children
with recurrent respiratory tract infections. Adv. Ther. 18,
151–162.
Ruedl, C., Fruhwirth, M., Wick, G., Wolf, H., 1994. Immune
response in the lungs following oral immunization with
bacterial lysates of respiratory pathogens. Clin. Diagn. Lab.
Immunol. 1, 150–154.
Sebert, M.E., Palmer, L.M., Rosenberg, M., Weiser, J.N.,
2002. Microarray-based identification of htrA, a Strepto-
coccus pneumoniae gene that is regulated by the CiaRH
two-component system and contributes to nasopharyngeal
colonization. Infect. Immun. 70, 4059–4067.
Shelburne, S.A., Granville, C., Tokuyama, M., Sitkiewicz, I.,
Patel, P., Musser, J.M., 2005. Growth characteristics of and
virulence factor production by group A Streptococcus
during cultivation in human saliva. Infect. Immun. 73,
4723–4731.
Sitkiewicz, I., Musser, J.M., 2006. Expression microarray and
mouse virulence analysis of four conserved two-component
gene regulatory systems in group A Streptococcus. Infect.
Immun. 74, 1339–1351.
Smoot, L.M., Smoot, J.C., Graham, M.R., Somerville, G.A.,
Sturdevant, D.E., Migliaccio, C.A.L., Sylva, G.L., Musser,
J.M., 2001. Global differential gene expression in response
to growth temperature alteration in group A Streptococcus.
Proc. Natl. Acad. Sci. USA 98, 10416–10421.
Spisek, R., Brazova, J., Rozkova, D., Zapletova, K., Sediva,
A., Bartunkova, J., 2004. Maturation of dendritic cells by
bacterial immunomodulators. Vaccine 22, 2761–2768.
Steurer-Stey, C., Bachmann, L.M., Steurer, J., Tramer, M.R.,
2004. Oral purified bacterial extracts in chronic bronchitis
and COPD: systematic review. Chest 126, 1645–1655.
Throup, J.P., Koretke, K.K., Bryant, A.P., Ingraham, K.A.,
Chalker, A.F., Ge, Y., Marra, A., Wallis, N.G., Brown,
J.R., Holmes, D.J., Rosenberg, M., Burnham, M.K., 2000.
A genomic analysis of two-component signal transduction
in Streptococcus pneumoniae. Mol. Microbiol. 35, 566–576.
Upton, M., Tagg, J.R., Wescombe, P., Jenkinson, H.F., 2001.
Intra- and interspecies signaling between Streptococcus
salivarius and Streptococcus pyogenes mediated by SalA
and SalA1 antibiotic peptides. J. Bacteriol. 183, 3931–3938.
Voyich, J.M., Sturdevant, D.E., Braughton, K.R., Kobayashi,
S.D., Lei, B., Virtaneva, K., Dorward, D.W., Musser, J.M.,
DeLeo, F.R., 2003. Genome-wide protective response used
by group A Streptococcus to evade destruction by human
polymorphonuclear leukocytes. Proc. Natl. Acad. Sci. USA
100, 1996–2001.
Zahner, D., Grebe, T., Guenzi, E., Krauß, J., van der Linden,
M., Terhune, K., Stock, J.B., Hakenbeck, R., 1996.
Resistance determinants for beta-lactam antibiotics in
laboratory mutants of Streptococcus pneumoniae that are
involved in genetic competence. Microb. Drug Res. 2,
187–191.
Zahner, D., Kaminski, K., van der Linden, M., Mascher, T.,
Merai, M., Hakenbeck, R., 2002. The ciaR/ciaH regulatory
network of Streptococcus pneumoniae. J. Mol. Microbiol.
Biotechnol. 4, 211–216.
Related Documents











