Transcriptional Regulation of Scar Gene Expression in Primary Astrocytes PAUL GRIS, 1,2 ALLYSON TIGHE, 1,3 DAVID LEVIN, 1 RAHUL SHARMA, 1 AND ARTHUR BROWN 1,2,3 * 1 The Spinal Cord Injury Team, BioTherapeutics Research Group, Robarts Research Institute, London, Ontario, Canada 2 Program in Neuroscience, The University of Western Ontario, London, Ontario, Canada 3 The Department of Physiology and Pharmacology, The University of Western Ontario, London, Ontario, Canada KEY WORDS CSPG; SOX9; xylosyltransferase; laminin; fibronectin ABSTRACT The failure of the adult injured spinal cord to support axo- nal regeneration is in part attributed to the glial scar. Reac- tive astrocytes constitute a major cellular component of the glial scar and are heterogeneous with respect to the extrac- ellular matrix proteins that they secrete. Astrocytes may produce antiregenerative molecules such as chondroitin sul- phate proteoglycans (CSPGs) or proregenerative molecules such as laminin and fibronectin. While many different CSPGs are expressed after spinal cord injury (SCI) they all rely on the same enzymes, xylosyltransferase-I and -II (XT- I, XT-II) and chondroitin 4-sulfotransferase (C4ST) to add the repulsive chondroitin sulfate side chains to their core proteins. We show that XT-I, XT-II, and C4ST are part of a CSPG biosynthetic gene (CBG) battery. Using primary astrocyte cultures and quantitative PCR we demonstrate that TGFb2, PDGF, and IL-6 induce the expression of CBGs, laminin and fibronectin by several-fold. We further show that over-expression of the transcription factor SOX9 also strongly induces the expression of CBGs but does not increase the expression of laminin or fibronectin. Corre- spondingly, SOX9 knock-down in primary astrocytes causes a decrease in CBG and an increase in laminin and fibronec- tin mRNA levels. Finally, we show that the in vivo expres- sion profiles of TGFb2, PDGF, IL-6, and SOX9 are consist- ent with their potential roles in differentially regulating CBGs, laminin and fibronectin gene expression in the injured spinal cord. This work suggests that SOX9 levels may be pivotal in determining the balance of pro- and anti- regenerative extracellular matrix molecules produced by astrocytes. V V C 2007 Wiley-Liss, Inc. INTRODUCTION The absence of axonal regeneration after spinal cord injury (SCI) has been attributed in part to the nonpermis- sive environment of the glial scar that inhibits axonal growth (Fawcett and Asher, 1999). Although macro- phages, microglia, oligodendrocytes, invading Schwann cells and meningeal fibroblasts contribute to the glial scar, astrocytes predominate (Fawcett and Asher, 1999). Reactive astrocytes in the injured CNS are heterogeneous with respect to their production of scar proteins (Fitch and Silver, 1997). Whereas in the majority of cases the extracellular matrix molecules (ECM) produced by astro- cytes have been shown to inhibit axonal regeneration (Bahr et al., 1995; Davies et al., 1999; McKeon et al., 1991; Reier and Houle, 1988), astrocytes also have been shown to secrete ECM molecules that promote axonal growth (McKeon et al., 1991). Thus astrocytes may pro- mote or inhibit regeneration after SCI depending upon the balance of growth-inhibiting and growth-promoting ECM molecules that they produce. Chondroitin sulfate proteoglycans (CSPGs) are prob- ably the most important of the inhibitory molecules pro- duced by reactive astrocytes (Eddleston and Mucke, 1993; Fawcett and Asher, 1999; Silver and Miller, 2004). In vivo and in vitro studies have shown that neurons cease to extend their axons into areas rich in CSPGs (Davies et al., 1997, 1999; McKeon et al., 1991; Zuo et al., 1998). CSPGs share a common structure comprising a central core protein with a number of chondroitin sulfate side chains (Morgenstern et al., 2002). Chondroitin sulfate side chain synthesis is initiated by the addition of xylose onto a serine moiety of the core protein. This rate-limiting step in the generation of chondroitin sulfate side chains function is carried out by the enzyme xylosyltransferase (XT) that exists in two isoforms encoded by two different genes XT-I and XT-II (Gotting et al., 2000; Kearns et al., 1991; Schwartz, 1977). These side chains are subse- quently sulfated by either chondroitin 4-sulfotransferase (C4ST) (Yamauchi et al., 2000) or chondroitin 6-sulfo- transferase (Fukuta et al., 1995) although, in astrocytes, C4ST predominates (Gallo and Bertolotto, 1990). Astrocytes can also produce an array of growth promot- ing molecules including laminin (Liesi and Silver, 1988), N-cadherin (Tomaselli et al., 1988), neural cell adhesion molecule (NCAM) (Neugebauer et al., 1988), and fibronec- tin (Matthiessen et al., 1989). Laminin and fibronectin have been shown to be good substrates for neurite exten- sion in in vitro models of axon growth (Costa et al., 2002; Fok-Seang et al., 1995; Hammarback et al., 1988; McKeon et al., 1991; Rogers et al., 1983, 1987). In vivo models dem- onstrate that sensory axon regeneration is dependent on astrocyte-associated fibronectin (Davies et al., 1997, 1999; Grant sponsors: Canadian Institutes of Health Research (CIHR), Krembil Foun- dation (Toronto). *Correspondence to: Dr. Arthur Brown, The Robarts Research Institute, 100 Perth Drive, London, Ontario, N6A 5K8 Canada. E-mail: [email protected] Received 6 March 2007; Revised 7 May 2007; Accepted 1 June 2007 DOI 10.1002/glia.20537 Published online 27 June 2007 in Wiley InterScience (www.interscience. wiley.com). GLIA 55:1145–1155 (2007) V V C 2007 Wiley-Liss, Inc.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Transcriptional Regulation of Scar Gene Expressionin Primary Astrocytes
PAUL GRIS,1,2 ALLYSON TIGHE,1,3 DAVID LEVIN,1 RAHUL SHARMA,1 AND ARTHUR BROWN1,2,3*1The Spinal Cord Injury Team, BioTherapeutics Research Group, Robarts Research Institute, London, Ontario, Canada2Program in Neuroscience, The University of Western Ontario, London, Ontario, Canada3The Department of Physiology and Pharmacology, The University of Western Ontario, London, Ontario, Canada
KEY WORDSCSPG; SOX9; xylosyltransferase; laminin; fibronectin
ABSTRACTThe failure of the adult injured spinal cord to support axo-nal regeneration is in part attributed to the glial scar. Reac-tive astrocytes constitute a major cellular component of theglial scar and are heterogeneous with respect to the extrac-ellular matrix proteins that they secrete. Astrocytes mayproduce antiregenerative molecules such as chondroitin sul-phate proteoglycans (CSPGs) or proregenerative moleculessuch as laminin and fibronectin. While many differentCSPGs are expressed after spinal cord injury (SCI) they allrely on the same enzymes, xylosyltransferase-I and -II (XT-I, XT-II) and chondroitin 4-sulfotransferase (C4ST) to addthe repulsive chondroitin sulfate side chains to their coreproteins. We show that XT-I, XT-II, and C4ST are part of aCSPG biosynthetic gene (CBG) battery. Using primaryastrocyte cultures and quantitative PCR we demonstratethat TGFb2, PDGF, and IL-6 induce the expression ofCBGs, laminin and fibronectin by several-fold. We furthershow that over-expression of the transcription factor SOX9also strongly induces the expression of CBGs but does notincrease the expression of laminin or fibronectin. Corre-spondingly, SOX9 knock-down in primary astrocytes causesa decrease in CBG and an increase in laminin and fibronec-tin mRNA levels. Finally, we show that the in vivo expres-sion profiles of TGFb2, PDGF, IL-6, and SOX9 are consist-ent with their potential roles in differentially regulatingCBGs, laminin and fibronectin gene expression in theinjured spinal cord. This work suggests that SOX9 levelsmay be pivotal in determining the balance of pro- and anti-regenerative extracellular matrix molecules produced byastrocytes. VVC 2007 Wiley-Liss, Inc.
INTRODUCTION
The absence of axonal regeneration after spinal cordinjury (SCI) has been attributed in part to the nonpermis-sive environment of the glial scar that inhibits axonalgrowth (Fawcett and Asher, 1999). Although macro-phages, microglia, oligodendrocytes, invading Schwanncells and meningeal fibroblasts contribute to the glialscar, astrocytes predominate (Fawcett and Asher, 1999).Reactive astrocytes in the injured CNS are heterogeneouswith respect to their production of scar proteins (Fitchand Silver, 1997). Whereas in the majority of cases theextracellular matrix molecules (ECM) produced by astro-cytes have been shown to inhibit axonal regeneration
(Bahr et al., 1995; Davies et al., 1999; McKeon et al.,1991; Reier and Houle, 1988), astrocytes also have beenshown to secrete ECM molecules that promote axonalgrowth (McKeon et al., 1991). Thus astrocytes may pro-mote or inhibit regeneration after SCI depending uponthe balance of growth-inhibiting and growth-promotingECM molecules that they produce.
Chondroitin sulfate proteoglycans (CSPGs) are prob-ably the most important of the inhibitory molecules pro-duced by reactive astrocytes (Eddleston and Mucke, 1993;Fawcett and Asher, 1999; Silver and Miller, 2004). In vivoand in vitro studies have shown that neurons cease toextend their axons into areas rich in CSPGs (Davieset al., 1997, 1999; McKeon et al., 1991; Zuo et al., 1998).CSPGs share a common structure comprising a centralcore protein with a number of chondroitin sulfate sidechains (Morgenstern et al., 2002). Chondroitin sulfateside chain synthesis is initiated by the addition of xyloseonto a serine moiety of the core protein. This rate-limitingstep in the generation of chondroitin sulfate side chainsfunction is carried out by the enzyme xylosyltransferase(XT) that exists in two isoforms encoded by two differentgenes XT-I and XT-II (Gotting et al., 2000; Kearns et al.,1991; Schwartz, 1977). These side chains are subse-quently sulfated by either chondroitin 4-sulfotransferase(C4ST) (Yamauchi et al., 2000) or chondroitin 6-sulfo-transferase (Fukuta et al., 1995) although, in astrocytes,C4ST predominates (Gallo and Bertolotto, 1990).
Astrocytes can also produce an array of growth promot-ing molecules including laminin (Liesi and Silver, 1988),N-cadherin (Tomaselli et al., 1988), neural cell adhesionmolecule (NCAM) (Neugebauer et al., 1988), and fibronec-tin (Matthiessen et al., 1989). Laminin and fibronectinhave been shown to be good substrates for neurite exten-sion in in vitro models of axon growth (Costa et al., 2002;Fok-Seang et al., 1995; Hammarback et al., 1988; McKeonet al., 1991; Rogers et al., 1983, 1987). In vivomodels dem-onstrate that sensory axon regeneration is dependent onastrocyte-associated fibronectin (Davies et al., 1997, 1999;
Grant sponsors: Canadian Institutes of Health Research (CIHR), Krembil Foun-dation (Toronto).
*Correspondence to: Dr. Arthur Brown, The Robarts Research Institute, 100Perth Drive, London, Ontario, N6A 5K8 Canada. E-mail: [email protected]
Received 6 March 2007; Revised 7 May 2007; Accepted 1 June 2007
DOI 10.1002/glia.20537
Published online 27 June 2007 in Wiley InterScience (www.interscience.wiley.com).
GLIA 55:1145–1155 (2007)
VVC 2007 Wiley-Liss, Inc.

Tom et al., 2004) and that intrathecal administration oflaminin-g1 promotes regeneration in a rat model of SCI(Wiksten et al., 2004).
The goal of this study was to identify pathways that dif-ferentially regulate the expression of XT-I, XT-II, andC4ST on the one hand and laminin and fibronectin on theother. We herein report that TGFb2, IL-6, and PDGF, incombination with SOX9, can differentially regulate theexpression of these genes in primary astrocytes and maybe responsible for their different expression profiles afterSCI.
MATERIALS AND METHODSAnimals and Surgeries
All protocols for these experiments were approved bythe University of Western Ontario Animal Care Commit-tee in accordance with the policies established in theGuide to Care and Use of Experimental Animals preparedby the Canadian Council on Animal Care. Thirty-twofemale Wistar Rats (Charles River) weighing 250–300 gwere premedicated with diazepam (3.5 mg/kg, i.p.) and at-ropine (0.05 mg/kg s.c.). Anesthesia was induced with 4%halothane and maintained with 1–1.5% halothane. A lam-inectomy was performed to expose the T4 spinal segmentand a modified aneurysm clip calibrated to a 50 g weight,was passed extradurally around the cord. Severe spinalcord compression was achieved by releasing the clip andallowing it to remain closed for 1 min (Fehlings and Tator,1995). The surgical wounds were closed and the rats weregiven 5 mg/kg of Baytril (Bayer), 5 mL of 0.9% salinetwice daily for 3 days and buprenophine (0.01 mg/kg s.c.)as needed. Bladders were manually emptied twice daily.After 12 h or 3, 7, 21, or 42 days, the rats were anesthe-tized with 1:2 ratio of ketamine: xylazine (0.13 mL/100 g)the injured segment of the spinal cord was removed, im-mediately homogenized in ice cold Trizol solution (Invi-trogen) and the RNA extracted as described (Carmelet al., 2001).
Primary Cell Culture
Primary astrocyte cultures were prepared from new-born rats at postnatal day 1 (P1) (Wilson and Dixon,1989). The upper portion of the skull was removed andthe meninges carefully dissected away to avoid contami-nation of the culture with fibroblasts. The neocorticeswere removed, placed into serum-free advanced D-MEM(Dulbecco’s Modified Eagle Medium, Invitrogen), homoge-nized by pipeting and gravity-filtered through 70-lm cellstrainer (Falcon). The cells were plated onto 6-well dishes(Falcon). The percentage of GFAP-expressing cells inthese cultures was found to be >95%. Cytokine treat-ments of primary astrocytes with PDGF, IL-6, TNFa, andTGFb2 (R and D systems) and bFGF2 (Invitrogen) werecarried out for 12 h. RNAwas extracted using the Qiagen(Germany) RNA-easy kit following manufacturer’s speci-fications. The transfections of primary astrocytes with
SiRNAs and the CMV-SOX9 expression construct wereconducted using Lipofectamine 2000 (Invitrogen) accord-ing to manufacturer’s specifications in 6-well dishes(Falcon). The siRNA (AAAGUUGUCGCUCCCACUGAA-GUUU) was used at a concentration of 150 pM. The uni-versal negative control scrambled SiRNA was usedaccording to manufacturer’s (Invitrogen) specifications.Transfection efficiencies with a fluorescently tagged con-trol SiRNA was 35–40%. The CMV-SOX9 construct haspreviously been described (Foster et al., 1994; Lefebvreet al., 1997) and plasmid transfection efficiency estimatedby cotransfection with a CMV-GFP construct was approxi-mately 10%.
RNA In Situ Hybridization
Rats were perfused with 4% paraformaldehyde 21 or 42days after SCI, their spinal cords removed and cryostat-sectioned horizontally at 16 lm. RNA in situ hybridiza-tion for XT-I expression was carried out using standardprocedures (Schaeren-Wiemers and Gerfin-Moser, 1993).A 491 bp fragment from nucleotides 226–717 (NCBIaccession number XM 341912.1) of the rattus XT-I genewas amplified by reverse transcription PCR, and subcl-oned into pGEM-T Easy (Promega). An antisense ribop-robe was generated using the T7 RNA polymerase anddigoxigenin-labeled UTP. The riboprobe signal wasdetected using an antidigoxigenin alkaline phosphatase-conjugated antibody (1:500; Roche) and 4-nitro blue tetra-zolium chloride with 5-bromo-4-chloro-3-indolyl-phos-phate (NBT-BCIP; Roche). Sense riboprobes were used asnegative controls.
Immunohistochemistry
Rats were perfused with 4% paraformaldehyde 21or 42 days after SCI, their spinal cords removed andcryostat-sectioned horizontally at 16 lm. Slides wereprocessed for immunohistochemistry using anti-GFAPantibodies (BD Pharmigen) at a 1:200 dilution toidentify reactive astrocytes, anti-CD11b antibodies(Sigma) at a 1:200 dilution to identify macrophagesor with an antibody, CS56 (Sigma), that recognizesthe terminal portions of chondroitin sulfate-4 or -6side chains and thus detects a variety of CSPGs(Avnur and Geiger, 1984; Fawcett and Asher, 1999) ata 1:50 dilution.
Slot Blot Analysis
Tissue and cell samples were lysed in RIPA buffer[20 mM, Tris-HCl (pH 7.6), 150 mM NaCl, 0.5% sodiumdeoxycholate, 1% Triton X-100, 0.1% SDS]. Then the pro-teins (3 lg/well) were transferred to polyvinylidene difluo-ride membranes (Millipore, Mississauga, ON) using Bio-Dot slot blot apparatus (BioRad, Mississauga, ON). Themembranes were first blocked in 10% nonfat powdered
1146 GRIS ET AL.
GLIA DOI 10.1002/glia

milk and then incubated with primary antibody at 1:200dilution overnight for CS-56 (Sigma, Missouri, USA).Following the incubation with HRP-conjugated donkeyanti-mouse antibody (1:10,000), membranes were incu-bated in ECL plus Western blotting detection reagents(Amersham, Buckinghamshire, UK) according to themanufacturer’s specifications. Immunoreactive bandswere scanned by an imaging densitometer (BioRad GF-700 Imaging Densitometer, Mississauga, ON); and resultswere quantified using Multi-Analist software (BioRad,Mississauga, ON). All values were normalized by dividingthe densitometric values for expression by the values forexpression of b-actin (anti-b-actin antibody from Sigma1:10,000 dilution).
In Silico Analysis of Putative Promoter Regions
The putative promoter regions of CBGs were identifiedusing ELDORADO software. Transcription start siteswere automatically assigned to the genes using databasesintegrated in to the promoter identification programELDORADO (Cohen et al., 2006). Promoter nucleotidesequences were analyzed using DIALIGN software tool(Genomatix Software).
Microarray Hybridization and Data Analysis
All GeneChips were processed at the London Re-gional Genomics Centre (Robarts Research Institute,London, ON, Canada). RNA quality was assessedusing the Agilent 2100 Bioanalyzer (Agilent Technolo-gies, Palo Alto, CA) and the RNA 6000 Nano kit (Cali-per Life Sciences, Mountain View, CA). RNA wasextracted from a 2 mm portion of spinal cord centeredon T4 (at the epicenter of the lesion in the spinal cordinjured rats). Biotinylated complimentary RNA (cRNA)was prepared from 10 lg of total RNA as per the Affy-metrix GeneChip Technical Analysis Manual (Affyme-trix, Santa Clara, CA). Double-stranded cDNA wassynthesized using SuperScriptII (Invitrogen, Carlsbad,CA) and oligo (dT)24 primers. Biotin-labeled cRNA wasprepared by in vitro transcription using the BioArrayHigh-Yield RNA Transcript Labeling kit (Enzo Bio-chem, New York) incorporating biotinylated UTP andCTP. 10 lg of labeled cRNA was hybridized toRAE230A GeneChips for 16 h at 45�C. GeneChipswere scanned with the Affymetrix GeneChip Scanner3000 (Affymetrix, Santa Clara, CA). Probe signalintensities were generated with GCOS1.3 (Affymetrix,Santa Clara, CA) using default values for the Statisti-cal Expression algorithm parameters and a TargetSignal of 150 for all probe sets and a NormalizationValue of 1. Gene expression level data was generatedusing the RMA preprocessor in GeneSpring GX 7.0(Agilent Technologies, Palo Alto, CA). Data were thentransformed, (measurements less than 0.01 set to0.01) and normalized per chip to the 50th percentile,and per gene to median.
Quantitative Polymerase Chain Reaction
First strand cDNA was synthesized from 1 lg RNA percondition (cell culture or animal tissue) using the HighCapacity cDNA Archive Kit according to the manufacturerprotocol (Applied Biosystems Foster City CA). The primerprobe sets, optical adhesive covers, and PCR plates werepurchased from Applied Biosystems (Foster City, CA). Theprobes were labeled with 50 FAM and with 30TAMRA asquencher with the exception of the ribosomal probe, whichwas labeled with 50 VIC. For Taq Man assays the thermalcycler conditions were 10 min at 95�C followed by 40 cyclesof 30 s at 95�C followed by 30 s at 60�C. A standard curveof cycle thresholds using cDNA serial dilutions was estab-lished and used to calculate abundance of each targetmRNA. Technical triplicates and at least biological tripli-cates were run on all conditions tested. Values were nor-malized to the amounts of 18S mRNA as determined byquantitative polymerase chain reaction (Q-PCR). The datawere analyzed by a two way ANOVA following by a Bon-ferroni test with Dunn’s correction for multiple compari-sons or Dunnet’s procedure when comparisons were madewith a single variable (control). Student’s t-test was usedwhen only two groups were compared.
Primer-probe sets for TaqMan gene expression assays:
RESULTSExpression Profiles of XT-I, XT-II, and
C4ST After SCI
We have previously shown that genes with relatedfunction may be regulated together as classes or bat-teries after SCI (Gris et al., 2003; Nelander et al., 2005).We hypothesized that, in astrocytes, genes that promoteaxon regeneration and genes that inhibit axon regenera-tion would be differentially regulated as gene classes.We thus predicted that XT-I, XT-II, and C4ST wouldform part of a CSPG biosynthetic gene (CBG) batteryand that there would be important differences in theexpression patterns and transcriptional regulation ofCBGs on the one hand and laminin and fibronectin onthe other. To evaluate the possibility that XT-I, XT-II,and C4ST are part of a gene battery we profiled theirexpression by Q-PCR after SCI in the rat. XT-I, XT-II,and C4ST all showed similar patterns of gene expressionafter SCI as detected by Q-PCR (Figs. 1A–C). ThemRNA levels of these genes peak from 12 h to 3 days fol-lowing SCI, then return to baseline levels by 7 dayspostinjury. XT-I, XT-II, and C4ST increase their expres-
Target Gene Probe and Primer catalog number
18S 4308329XT-I 1391062AXT-II Mm00517563_m1C4ST Mm00517563_m1Laminin-g1 Mm00711808_m1SOX9 Mm 0048840_m1TGb2 Rn00579674_m1IL-6 Rn00561420_m1
1147SOX9 AND CYTOKINES REGULATE SCAR GENES
GLIA DOI 10.1002/glia
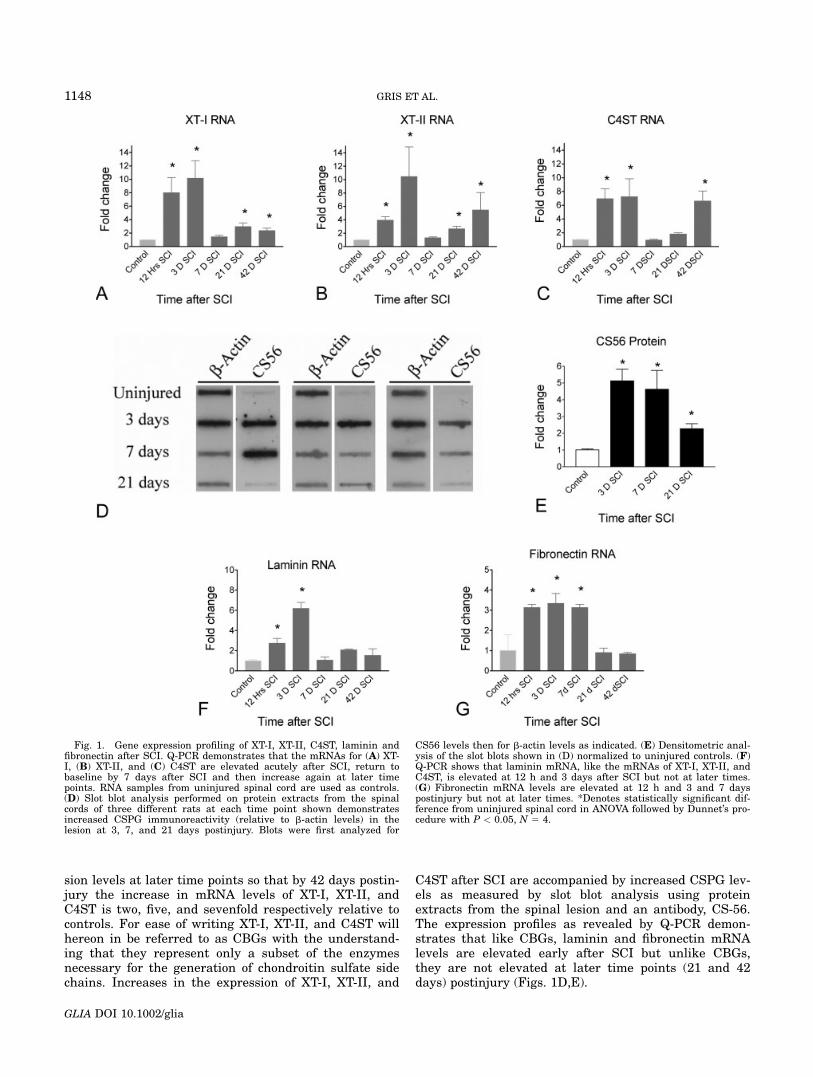
sion levels at later time points so that by 42 days postin-jury the increase in mRNA levels of XT-I, XT-II, andC4ST is two, five, and sevenfold respectively relative tocontrols. For ease of writing XT-I, XT-II, and C4ST willhereon in be referred to as CBGs with the understand-ing that they represent only a subset of the enzymesnecessary for the generation of chondroitin sulfate sidechains. Increases in the expression of XT-I, XT-II, and
C4ST after SCI are accompanied by increased CSPG lev-els as measured by slot blot analysis using proteinextracts from the spinal lesion and an antibody, CS-56.The expression profiles as revealed by Q-PCR demon-strates that like CBGs, laminin and fibronectin mRNAlevels are elevated early after SCI but unlike CBGs,they are not elevated at later time points (21 and 42days) postinjury (Figs. 1D,E).
Fig. 1. Gene expression profiling of XT-I, XT-II, C4ST, laminin andfibronectin after SCI. Q-PCR demonstrates that the mRNAs for (A) XT-I, (B) XT-II, and (C) C4ST are elevated acutely after SCI, return tobaseline by 7 days after SCI and then increase again at later timepoints. RNA samples from uninjured spinal cord are used as controls.(D) Slot blot analysis performed on protein extracts from the spinalcords of three different rats at each time point shown demonstratesincreased CSPG immunoreactivity (relative to b-actin levels) in thelesion at 3, 7, and 21 days postinjury. Blots were first analyzed for
CS56 levels then for b-actin levels as indicated. (E) Densitometric anal-ysis of the slot blots shown in (D) normalized to uninjured controls. (F)Q-PCR shows that laminin mRNA, like the mRNAs of XT-I, XT-II, andC4ST, is elevated at 12 h and 3 days after SCI but not at later times.(G) Fibronectin mRNA levels are elevated at 12 h and 3 and 7 dayspostinjury but not at later times. *Denotes statistically significant dif-ference from uninjured spinal cord in ANOVA followed by Dunnet’s pro-cedure with P < 0.05, N 5 4.
1148 GRIS ET AL.
GLIA DOI 10.1002/glia

Identification of the Cellular Source of CBGmRNA by In Situ Hybridization
To determine the cellular source of CBG mRNA afterSCI we carried out RNA in situ hybridization analysis onsections of rat injured spinal cords with a XT-I antisenseriboprobe. Immunohistochemistry on these same sectionsusing an anti-CD11b mAb to detect macrophages and ananti-GFAP antibody to detect astrocytes indicated that6 weeks after injury both these cell types express XT-I inthe lesion (Fig. 2).
IL-6, PDGF, and TGFb2 Are PutativeRegulators of XT-I, XT-II, and C4ST
The first strategy used to identify potential positiveregulators of XT-I, XT-II, and C4ST gene transcriptionwas based on the premise that a subset of moleculesthat are able to up-regulate their expression will showexpression patterns similar to the Q-PCR-delineatedexpression patterns of XT-I, XT-II, and C4ST asdescribed earlier. For example, an inducer of XT-I, XT-II,and C4ST expression will itself show elevated levels ofexpression when XT-I, XT-II, and C4ST expression levelsare high and low levels of expression when XT-I, XT-II,and C4ST expression levels are low. Thus we analyzedgene expression profiles in the rat injured spinal cordthat we obtained using an Affymetrix platform (the Affy-metrix Rat 230A gene chip) after a clip compression SCIin the rat. Since inflammation is known to be a key reg-ulator of fibrosis and scarring, we restricted our interestto known cytokine mediators of inflammation. From thisGroup 3 cytokines, IL-6, TGFb2, and PDGF, withexpression profiles most similar to the expression pro-files of XT-I, XT-II, and C4ST were identified. The micro-array-delineated expression profile for two of these cyto-kines, TGFb2 and IL-6, was verified by Q-PCR onmRNA isolated from lesion sites at 12 h, 3, 7, 21, and 42days after SCI (Figs. 3A,B). In agreement with themicroarray data, these cytokines demonstrated rapid
increases in mRNA levels 12 h after SCI followed by adecrease to baseline levels by 7 days after SCI and asubsequent increase inTGFb2 but not IL-6 mRNA at 21and 42 days after SCI. Thus these cytokines and XT-I,XT-II, and C4ST have similar patterns of expressionafter SCI.
In Silico Analysis of Promoter Regionsof XT-I, XT-II, and C4ST
The second strategy used to identify regulators of XT-I,XT-II, and C4ST was to identify transcription factor bind-ing sites in common to the promoters of all three genes.This strategy was on the basis of the premise that, if XT-I,XT-II, and C4ST constitute part of a gene battery, thenthey would be controlled by an overlapping set of tran-scription factors. We defined the putative promoterregions of XT-I, XT-II, and C4ST using Genomatix suitesoftware (Genomatix Software). To reduce the possibilityof incorrectly identifying putative transcription factorbinding sites, we compared the promoter sequences of
Fig. 2. XT-I is expressed in macro-phages and astrocytes at the lesion epi-center 6 weeks after SCI. After under-going RNA in situ hybridization for XT-Iexpression (signal appears blue) the sec-tions were processed for immunohisto-chemistry for (A) GFAP or (B) CD11bprotein expression (signal appearswhite). Bar 5 0.1 mm.
Fig. 3. Q-PCR confirmation of the expression profiles of candidateregulators of CBG genes. Microarray analysis suggested that TGFb2,IL-6, and PDGF have expression profiles that are consistent with a roleas candidate transcriptional regulators of XT-I, XT-II, C4ST. Q-PCRwas used to confirm this pattern of expression in the spinal lesion for(A) TGFb2 and (B) IL-6. The mRNAs for these genes are elevated earlyafter SCI and then return to baseline levels between 3 and 7 days afterSCI. TGFb2 mRNA levels subsequently increase again at later times.*Denotes statistically significant difference from uninjured spinal cordin ANOVA followed by Dunnet’s procedure with P < 0.05, N 5 4.
1149SOX9 AND CYTOKINES REGULATE SCAR GENES
GLIA DOI 10.1002/glia
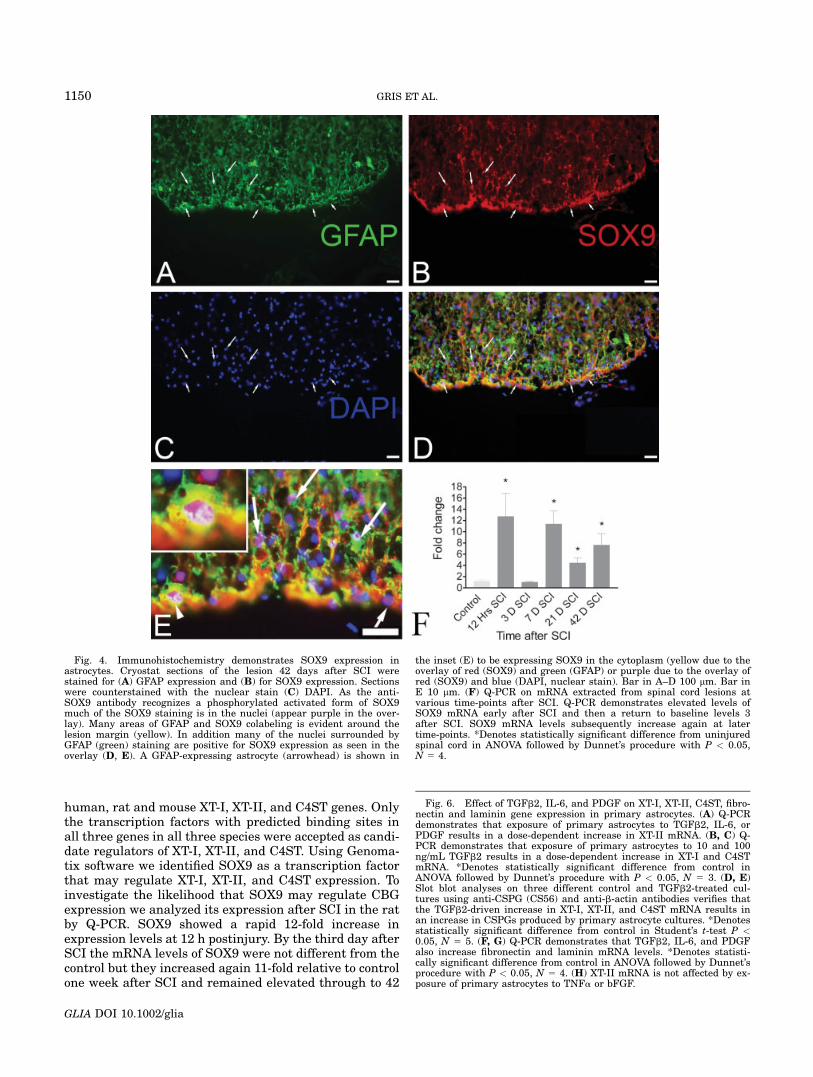
human, rat and mouse XT-I, XT-II, and C4ST genes. Onlythe transcription factors with predicted binding sites inall three genes in all three species were accepted as candi-date regulators of XT-I, XT-II, and C4ST. Using Genoma-tix software we identified SOX9 as a transcription factorthat may regulate XT-I, XT-II, and C4ST expression. Toinvestigate the likelihood that SOX9 may regulate CBGexpression we analyzed its expression after SCI in the ratby Q-PCR. SOX9 showed a rapid 12-fold increase inexpression levels at 12 h postinjury. By the third day afterSCI the mRNA levels of SOX9 were not different from thecontrol but they increased again 11-fold relative to controlone week after SCI and remained elevated through to 42
Fig. 4. Immunohistochemistry demonstrates SOX9 expression inastrocytes. Cryostat sections of the lesion 42 days after SCI werestained for (A) GFAP expression and (B) for SOX9 expression. Sectionswere counterstained with the nuclear stain (C) DAPI. As the anti-SOX9 antibody recognizes a phosphorylated activated form of SOX9much of the SOX9 staining is in the nuclei (appear purple in the over-lay). Many areas of GFAP and SOX9 colabeling is evident around thelesion margin (yellow). In addition many of the nuclei surrounded byGFAP (green) staining are positive for SOX9 expression as seen in theoverlay (D, E). A GFAP-expressing astrocyte (arrowhead) is shown in
the inset (E) to be expressing SOX9 in the cytoplasm (yellow due to theoverlay of red (SOX9) and green (GFAP) or purple due to the overlay ofred (SOX9) and blue (DAPI, nuclear stain). Bar in A–D 100 lm. Bar inE 10 lm. (F) Q-PCR on mRNA extracted from spinal cord lesions atvarious time-points after SCI. Q-PCR demonstrates elevated levels ofSOX9 mRNA early after SCI and then a return to baseline levels 3after SCI. SOX9 mRNA levels subsequently increase again at latertime-points. *Denotes statistically significant difference from uninjuredspinal cord in ANOVA followed by Dunnet’s procedure with P < 0.05,N 5 4.
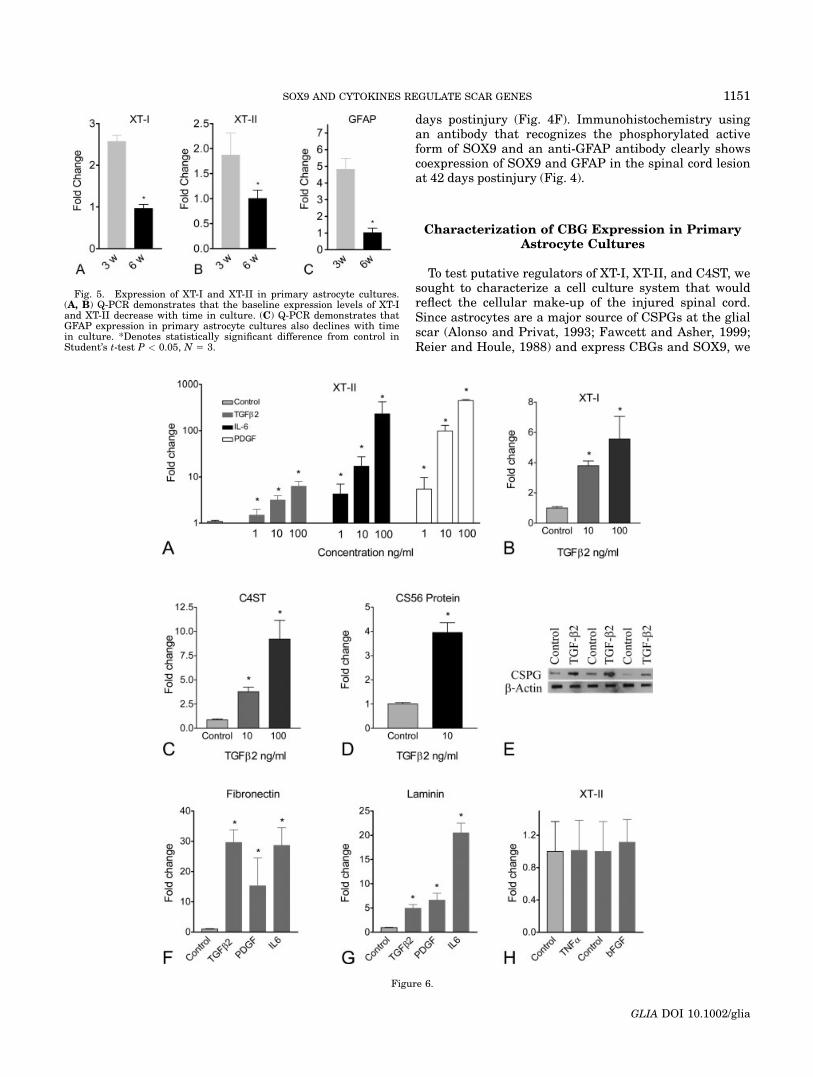
Fig. 6. Effect of TGFb2, IL-6, and PDGF on XT-I, XT-II, C4ST, fibro-nectin and laminin gene expression in primary astrocytes. (A) Q-PCRdemonstrates that exposure of primary astrocytes to TGFb2, IL-6, orPDGF results in a dose-dependent increase in XT-II mRNA. (B, C) Q-PCR demonstrates that exposure of primary astrocytes to 10 and 100ng/mL TGFb2 results in a dose-dependent increase in XT-I and C4STmRNA. *Denotes statistically significant difference from control inANOVA followed by Dunnet’s procedure with P < 0.05, N 5 3. (D, E)Slot blot analyses on three different control and TGFb2-treated cul-tures using anti-CSPG (CS56) and anti-b-actin antibodies verifies thatthe TGFb2-driven increase in XT-I, XT-II, and C4ST mRNA results inan increase in CSPGs produced by primary astrocyte cultures. *Denotesstatistically significant difference from control in Student’s t-test P <0.05, N 5 5. (F, G) Q-PCR demonstrates that TGFb2, IL-6, and PDGFalso increase fibronectin and laminin mRNA levels. *Denotes statisti-cally significant difference from control in ANOVA followed by Dunnet’sprocedure with P < 0.05, N 5 4. (H) XT-II mRNA is not affected by ex-posure of primary astrocytes to TNFa or bFGF.
1150 GRIS ET AL.
GLIA DOI 10.1002/glia
days postinjury (Fig. 4F). Immunohistochemistry usingan antibody that recognizes the phosphorylated activeform of SOX9 and an anti-GFAP antibody clearly showscoexpression of SOX9 and GFAP in the spinal cord lesionat 42 days postinjury (Fig. 4).
Characterization of CBG Expression in PrimaryAstrocyte Cultures
To test putative regulators of XT-I, XT-II, and C4ST, wesought to characterize a cell culture system that wouldreflect the cellular make-up of the injured spinal cord.Since astrocytes are a major source of CSPGs at the glialscar (Alonso and Privat, 1993; Fawcett and Asher, 1999;Reier and Houle, 1988) and express CBGs and SOX9, we
Fig. 5. Expression of XT-I and XT-II in primary astrocyte cultures.(A, B) Q-PCR demonstrates that the baseline expression levels of XT-Iand XT-II decrease with time in culture. (C) Q-PCR demonstrates thatGFAP expression in primary astrocyte cultures also declines with timein culture. *Denotes statistically significant difference from control inStudent’s t-test P < 0.05, N 5 3.
Figure 6.
1151SOX9 AND CYTOKINES REGULATE SCAR GENES
GLIA DOI 10.1002/glia
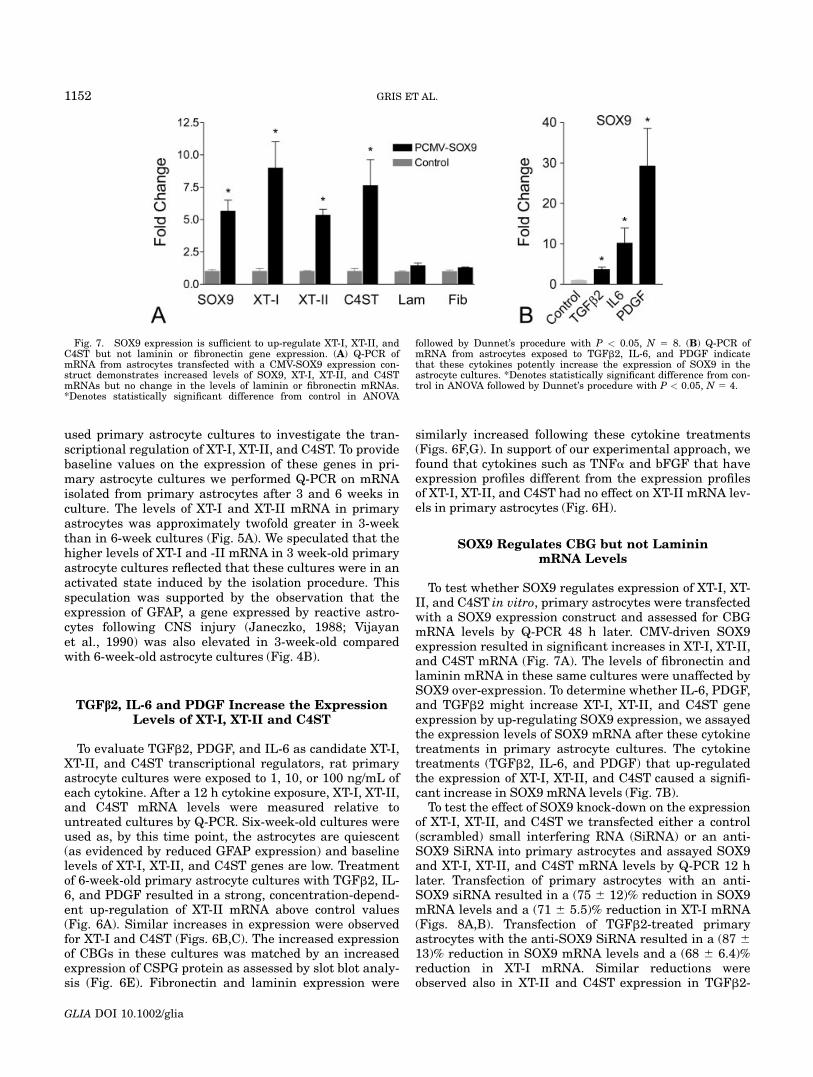
used primary astrocyte cultures to investigate the tran-scriptional regulation of XT-I, XT-II, and C4ST. To providebaseline values on the expression of these genes in pri-mary astrocyte cultures we performed Q-PCR on mRNAisolated from primary astrocytes after 3 and 6 weeks inculture. The levels of XT-I and XT-II mRNA in primaryastrocytes was approximately twofold greater in 3-weekthan in 6-week cultures (Fig. 5A). We speculated that thehigher levels of XT-I and -II mRNA in 3 week-old primaryastrocyte cultures reflected that these cultures were in anactivated state induced by the isolation procedure. Thisspeculation was supported by the observation that theexpression of GFAP, a gene expressed by reactive astro-cytes following CNS injury (Janeczko, 1988; Vijayanet al., 1990) was also elevated in 3-week-old comparedwith 6-week-old astrocyte cultures (Fig. 4B).
TGFb2, IL-6 and PDGF Increase the ExpressionLevels of XT-I, XT-II and C4ST
To evaluate TGFb2, PDGF, and IL-6 as candidate XT-I,XT-II, and C4ST transcriptional regulators, rat primaryastrocyte cultures were exposed to 1, 10, or 100 ng/mL ofeach cytokine. After a 12 h cytokine exposure, XT-I, XT-II,and C4ST mRNA levels were measured relative tountreated cultures by Q-PCR. Six-week-old cultures wereused as, by this time point, the astrocytes are quiescent(as evidenced by reduced GFAP expression) and baselinelevels of XT-I, XT-II, and C4ST genes are low. Treatmentof 6-week-old primary astrocyte cultures with TGFb2, IL-6, and PDGF resulted in a strong, concentration-depend-ent up-regulation of XT-II mRNA above control values(Fig. 6A). Similar increases in expression were observedfor XT-I and C4ST (Figs. 6B,C). The increased expressionof CBGs in these cultures was matched by an increasedexpression of CSPG protein as assessed by slot blot analy-sis (Fig. 6E). Fibronectin and laminin expression were
similarly increased following these cytokine treatments(Figs. 6F,G). In support of our experimental approach, wefound that cytokines such as TNFa and bFGF that haveexpression profiles different from the expression profilesof XT-I, XT-II, and C4ST had no effect on XT-II mRNA lev-els in primary astrocytes (Fig. 6H).
SOX9 Regulates CBG but not LamininmRNA Levels
To test whether SOX9 regulates expression of XT-I, XT-II, and C4ST in vitro, primary astrocytes were transfectedwith a SOX9 expression construct and assessed for CBGmRNA levels by Q-PCR 48 h later. CMV-driven SOX9expression resulted in significant increases in XT-I, XT-II,and C4ST mRNA (Fig. 7A). The levels of fibronectin andlaminin mRNA in these same cultures were unaffected bySOX9 over-expression. To determine whether IL-6, PDGF,and TGFb2 might increase XT-I, XT-II, and C4ST geneexpression by up-regulating SOX9 expression, we assayedthe expression levels of SOX9 mRNA after these cytokinetreatments in primary astrocyte cultures. The cytokinetreatments (TGFb2, IL-6, and PDGF) that up-regulatedthe expression of XT-I, XT-II, and C4ST caused a signifi-cant increase in SOX9 mRNA levels (Fig. 7B).
To test the effect of SOX9 knock-down on the expressionof XT-I, XT-II, and C4ST we transfected either a control(scrambled) small interfering RNA (SiRNA) or an anti-SOX9 SiRNA into primary astrocytes and assayed SOX9and XT-I, XT-II, and C4ST mRNA levels by Q-PCR 12 hlater. Transfection of primary astrocytes with an anti-SOX9 siRNA resulted in a (75 6 12)% reduction in SOX9mRNA levels and a (71 6 5.5)% reduction in XT-I mRNA(Figs. 8A,B). Transfection of TGFb2-treated primaryastrocytes with the anti-SOX9 SiRNA resulted in a (87 6
13)% reduction in SOX9 mRNA levels and a (68 6 6.4)%reduction in XT-I mRNA. Similar reductions wereobserved also in XT-II and C4ST expression in TGFb2-
Fig. 7. SOX9 expression is sufficient to up-regulate XT-I, XT-II, andC4ST but not laminin or fibronectin gene expression. (A) Q-PCR ofmRNA from astrocytes transfected with a CMV-SOX9 expression con-struct demonstrates increased levels of SOX9, XT-I, XT-II, and C4STmRNAs but no change in the levels of laminin or fibronectin mRNAs.*Denotes statistically significant difference from control in ANOVA
followed by Dunnet’s procedure with P < 0.05, N 5 8. (B) Q-PCR ofmRNA from astrocytes exposed to TGFb2, IL-6, and PDGF indicatethat these cytokines potently increase the expression of SOX9 in theastrocyte cultures. *Denotes statistically significant difference from con-trol in ANOVA followed by Dunnet’s procedure with P < 0.05, N 5 4.
1152 GRIS ET AL.
GLIA DOI 10.1002/glia
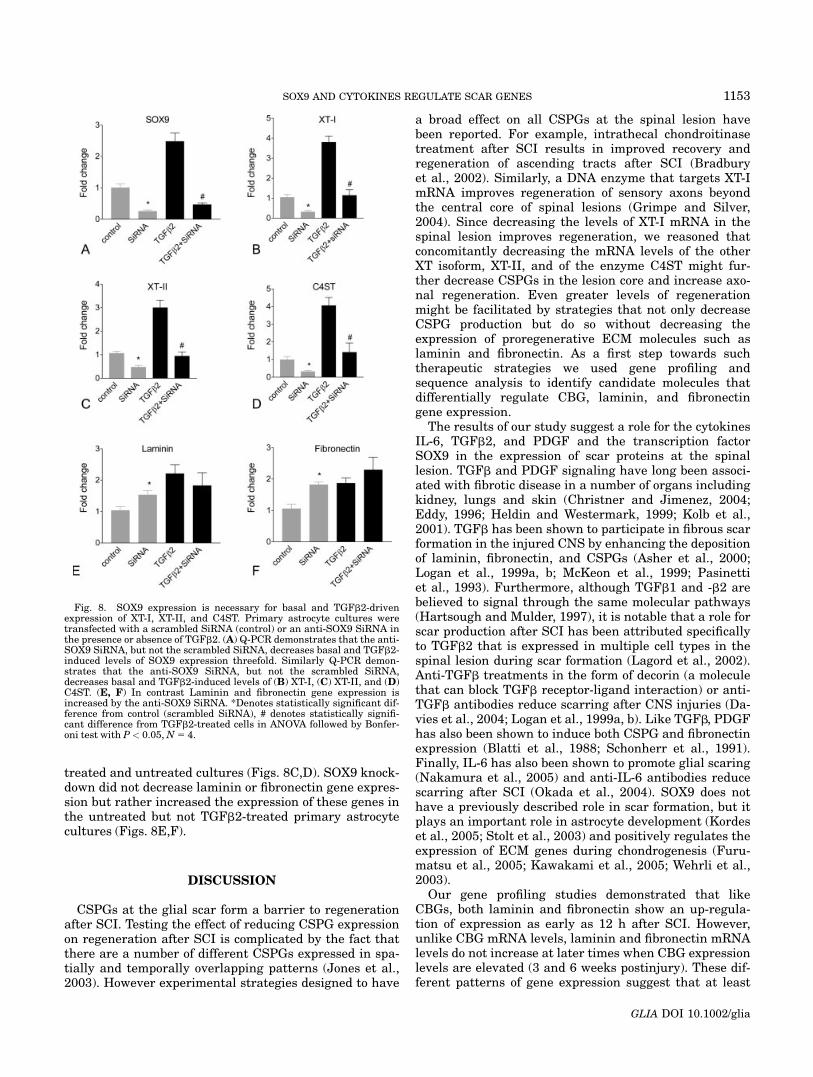
treated and untreated cultures (Figs. 8C,D). SOX9 knock-down did not decrease laminin or fibronectin gene expres-sion but rather increased the expression of these genes inthe untreated but not TGFb2-treated primary astrocytecultures (Figs. 8E,F).
DISCUSSION
CSPGs at the glial scar form a barrier to regenerationafter SCI. Testing the effect of reducing CSPG expressionon regeneration after SCI is complicated by the fact thatthere are a number of different CSPGs expressed in spa-tially and temporally overlapping patterns (Jones et al.,2003). However experimental strategies designed to have
a broad effect on all CSPGs at the spinal lesion havebeen reported. For example, intrathecal chondroitinasetreatment after SCI results in improved recovery andregeneration of ascending tracts after SCI (Bradburyet al., 2002). Similarly, a DNA enzyme that targets XT-ImRNA improves regeneration of sensory axons beyondthe central core of spinal lesions (Grimpe and Silver,2004). Since decreasing the levels of XT-I mRNA in thespinal lesion improves regeneration, we reasoned thatconcomitantly decreasing the mRNA levels of the otherXT isoform, XT-II, and of the enzyme C4ST might fur-ther decrease CSPGs in the lesion core and increase axo-nal regeneration. Even greater levels of regenerationmight be facilitated by strategies that not only decreaseCSPG production but do so without decreasing theexpression of proregenerative ECM molecules such aslaminin and fibronectin. As a first step towards suchtherapeutic strategies we used gene profiling andsequence analysis to identify candidate molecules thatdifferentially regulate CBG, laminin, and fibronectingene expression.
The results of our study suggest a role for the cytokinesIL-6, TGFb2, and PDGF and the transcription factorSOX9 in the expression of scar proteins at the spinallesion. TGFb and PDGF signaling have long been associ-ated with fibrotic disease in a number of organs includingkidney, lungs and skin (Christner and Jimenez, 2004;Eddy, 1996; Heldin and Westermark, 1999; Kolb et al.,2001). TGFb has been shown to participate in fibrous scarformation in the injured CNS by enhancing the depositionof laminin, fibronectin, and CSPGs (Asher et al., 2000;Logan et al., 1999a, b; McKeon et al., 1999; Pasinettiet al., 1993). Furthermore, although TGFb1 and -b2 arebelieved to signal through the same molecular pathways(Hartsough and Mulder, 1997), it is notable that a role forscar production after SCI has been attributed specificallyto TGFb2 that is expressed in multiple cell types in thespinal lesion during scar formation (Lagord et al., 2002).Anti-TGFb treatments in the form of decorin (a moleculethat can block TGFb receptor-ligand interaction) or anti-TGFb antibodies reduce scarring after CNS injuries (Da-vies et al., 2004; Logan et al., 1999a, b). Like TGFb, PDGFhas also been shown to induce both CSPG and fibronectinexpression (Blatti et al., 1988; Schonherr et al., 1991).Finally, IL-6 has also been shown to promote glial scaring(Nakamura et al., 2005) and anti-IL-6 antibodies reducescarring after SCI (Okada et al., 2004). SOX9 does nothave a previously described role in scar formation, but itplays an important role in astrocyte development (Kordeset al., 2005; Stolt et al., 2003) and positively regulates theexpression of ECM genes during chondrogenesis (Furu-matsu et al., 2005; Kawakami et al., 2005; Wehrli et al.,2003).
Our gene profiling studies demonstrated that likeCBGs, both laminin and fibronectin show an up-regula-tion of expression as early as 12 h after SCI. However,unlike CBG mRNA levels, laminin and fibronectin mRNAlevels do not increase at later times when CBG expressionlevels are elevated (3 and 6 weeks postinjury). These dif-ferent patterns of gene expression suggest that at least
Fig. 8. SOX9 expression is necessary for basal and TGFb2-drivenexpression of XT-I, XT-II, and C4ST. Primary astrocyte cultures weretransfected with a scrambled SiRNA (control) or an anti-SOX9 SiRNA inthe presence or absence of TGFb2. (A) Q-PCR demonstrates that the anti-SOX9 SiRNA, but not the scrambled SiRNA, decreases basal and TGFb2-induced levels of SOX9 expression threefold. Similarly Q-PCR demon-strates that the anti-SOX9 SiRNA, but not the scrambled SiRNA,decreases basal and TGFb2-induced levels of (B) XT-I, (C) XT-II, and (D)C4ST. (E, F) In contrast Laminin and fibronectin gene expression isincreased by the anti-SOX9 SiRNA. *Denotes statistically significant dif-ference from control (scrambled SiRNA), # denotes statistically signifi-cant difference from TGFb2-treated cells in ANOVA followed by Bonfer-oni test with P< 0.05,N5 4.
1153SOX9 AND CYTOKINES REGULATE SCAR GENES
GLIA DOI 10.1002/glia

some elements regulating CBG expression are differentfrom those that regulate the expression of laminin and fi-bronectin. Our results clearly show that SOX9 expressionis both necessary and sufficient for CBG expression in pri-mary astrocytes and suggests that cytokine up-regulationof CBG expression is SOX9 dependent. Laminin and fibro-nectin gene expression are also positively regulated byIL-6, TGFb2, and PDGF. The role of SOX9 in regulatinglaminin and fibronectin expression is as yet unclear. Theanti-SOX9 SiRNA transfections suggest that laminin andfibronectin expression are in fact negatively regulated bySOX9. These results were not supported by the CMV-SOX9 transfections that would have been predicted toreduce laminin and fibronectin mRNA levels. We suggestthat the basal levels of laminin and fibronectin mRNA in6-week-old primary astrocyte cultures is sufficiently lowthat further reductions in expression are difficult todetect especially given that the CMV-SOX9 transfectionefficiencies were approximately 10% of cells.
Thus our in vitro functional data and our in vivoexpression data suggest a model by which the increase inlaminin, fibronectin, and CBG expression early after SCImay be regulated by cytokines such as TGFb2, IL-6, orPDGF or directly by SOX9 in the case of CBGs. SOX9expression may itself be up-regulated after injury bythese same cytokines. At subchronic and chronic timepoints the expression levels of laminin and fibronectinmay fall relative to the levels of CBGs because cytokinelevels decrease but SOX9 levels remain high. Thus wesuggest that the predominance of CSPGs at the spinallesion site may depend in part on the expression levels ofthe transcription factor SOX9 which differentially regu-lates regeneration-inhibiting and promoting ECM pro-teins expressed in the injured spinal cord. ThereforeSOX9 may be an important target for regeneration strat-egies after SCI.
ACKNOWLEDGMENTS
Dr. A. Brown was supported by a New InvestigatorAward from the Heart and Stroke Foundation of Canadaand P. Gris was supported by studentships from theCIHR, the Ontario Neurotrauma Foundation and the On-tario Graduate Scholarship Program. We would like tothank Dr. Michael Underhill for the generous gift of theCMV-SOX9 expression plasmid.
REFERENCES
Alonso G, Privat A. 1993. Reactive astrocytes involved in the formationof lesional scars differ in the mediobasal hypothalamus and in otherforebrain regions. J Neurosci Res 34:523–538.
Asher RA, Morgenstern DA, Fidler PS, Adcock KH, Oohira A, Brais-tead JE, Levine JM, Margolis RU, Rogers JH, Fawcett JW. 2000.Neurocan is upregulated in injured brain and in cytokine-treatedastrocytes. J Neurosci 20:2427–2438.
Avnur Z, Geiger B. 1984. Immunocytochemical localization of nativechondroitin-sulfate in tissues and cultured cells using specific mono-clonal antibody. Cell 38:811–822.
Bahr M, Przyrembel C, Bastmeyer M. 1995. Astrocytes from adult ratoptic nerves are nonpermissive for regenerating retinal ganglion cellaxons. Exp Neurol 131:211–220.
Blatti SP, Foster DN, Ranganathan G, Moses HL, Getz MJ. 1988.Induction of fibronectin gene transcription and mRNA is a primaryresponse to growth-factor stimulation of AKR-2B cells. Proc NatlAcad Sci USA 85:1119–1123.
Bradbury EJ, Moon LD, Popat RJ, King VR, Bennett GS, Patel PN,Fawcett JW, McMahon SB. 2002. Chondroitinase ABC promotes func-tional recovery after spinal cord injury. Nature 416:636–640.
Carmel JB, Galante A, Soteropoulos P, Tolias P, Recce M, Young W,Hart RP. 2001. Gene expression profiling of acute spinal cord injuryreveals spreading inflammatory signals and neuron loss. PhysiolGenomics 7:201–213.
Christner PJ, Jimenez SA. 2004. Animal models of systemic sclerosis:Insights into systemic sclerosis pathogenesis and potential therapeu-tic approaches. Curr Opin Rheumatol 16:746–752.
Cohen CD, Klingenhoff A, Boucherot A, Nitsche A, Henger A, BrunnerB, Schmid H, Merkle M, Saleem MA, Koller KP, Werner T, GroneHJ, Nelson PJ, Kretzler M. 2006. Comparative promoter analysisallows de novo identification of specialized cell junction-associatedproteins. Proc Natl Acad Sci USA 103:5682–5687.
Costa S, Planchenault T, Charriere-Bertrand C, Mouchel Y, Fages C,Juliano S, Lefrancois T, Barlovatz-Meimon G, Tardy M. 2002. Astro-glial permissivity for neuritic outgrowth in neuron-astrocyte cocul-tures depends on regulation of laminin bioavailability. Glia 37:105–113.
Davies JE, Tang X, Denning JW, Archibald SJ, Davies SJ. 2004.Decorin suppresses neurocan, brevican, phosphacan and NG2 expres-sion and promotes axon growth across adult rat spinal cord injuries.Eur J Neurosci 19:1226–1242.
Davies SJ, Fitch MT, Memberg SP, Hall AK, Raisman G, Silver J. 1997.Regeneration of adult axons in white matter tracts of the centralnervous system. Nature 390:680–683.
Davies SJ, Goucher DR, Doller C, Silver J. 1999. Robust regenerationof adult sensory axons in degenerating white matter of the adult ratspinal cord. J Neurosci 19:5810–5822.
Eddleston M, Mucke L. 1993. Molecular profile of reactive astrocytes—Implications for their role in neurologic disease. Neuroscience 54:15–36.
Eddy AA. 1996. Molecular insights into renal interstitial fibrosis. J AmSoc Nephrol 7:2495–2508.
Fawcett JW, Asher RA. 1999. The glial scar and central nervous systemrepair. Brain Res Bull 49:377–391.
Fehlings MG, Tator CH. 1995. The relationships among the severity ofspinal cord injury, residual neurological function, axon counts, andcounts of retrogradely labeled neurons after experimental spinal cordinjury. Exp Neurol 132:220–228.
Fitch MT, Silver J. 1997. Activated macrophages and the blood-brainbarrier: Inflammation after CNS injury leads to increases in putativeinhibitory molecules. Exp Neurol 148:587–603.
Fok-Seang J, Smith-Thomas LC, Meiners S, Muir E, Du JS, HousdenE, Johnson AR, Faissner A, Geller HM, Keynes RJ, and others. 1995.An analysis of astrocytic cell lines with different abilities to promoteaxon growth. Brain Res 689:207–223.
Foster JW, Dominguez-Steglich MA, Guioli S, Kowk G, Weller PA,Stevanovic M, Weissenbach J, Mansour S, Young ID, GoodfellowPN, and others. 1994. Campomelic dysplasia and autosomal sex re-versal caused by mutations in an SRY-related gene. Nature 372:525–530.
Fukuta M, Uchimura K, Nakashima K, Kato M, Kimata K, ShinomuraT, Habuchi O. 1995. Molecular cloning and expression of chick chon-drocyte chondroitin 6-sulfotransferase. J Biol Chem 270:18575–18580.
Furumatsu T, Tsuda M, Taniguchi N, Tajima Y, Asahara H. 2005.Smad3 induces chondrogenesis through the activation of SOX9 viaCREB-binding protein/p300 recruitment. J Biol Chem 280:8343–8350.
Gallo V, Bertolotto A. 1990. Extracellular matrix of cultured glial cells:Selective expression of chondroitin 4-sulfate by type-2 astrocytes andtheir progenitors. Exp Cell Res 187:211–223.
Gotting C, Kuhn J, Zahn R, Brinkmann T, Kleesiek K. 2000. Molecularcloning and expression of human UDP-d-Xylose:proteoglycan coreprotein b-d-xylosyltransferase and its first isoform XT-II. J Mol Biol304:517–528.
Grimpe B, Silver J. 2004. A novel DNA enzyme reduces glycosamino-glycan chains in the glial scar and allows microtransplanted dorsalroot ganglia axons to regenerate beyond lesions in the spinal cord.J Neurosci 24:1393–1397.
Gris P, Murphy S, Jacob JE, Atkinson I, Brown A. 2003. Differentialgene expression profiles in embryonic, adult-injured and adult-unin-jured rat spinal cords. Mol Cell Neurosci 24:555–567.
Hammarback JA, McCarthy JB, Palm SL, Furcht LT, Letourneau PC.1988. Growth cone guidance by substrate-bound laminin pathways iscorrelated with neuron-to-pathway adhesivity. Dev Biol 126:29–39.
1154 GRIS ET AL.
GLIA DOI 10.1002/glia

Hartsough MT, Mulder KM. 1997. Transforming growth factor-b signal-ing in epithelial cells. Pharmacol Ther 75:21–41.
Heldin CH, Westermark B. 1999. Mechanism of action and in vivo roleof platelet-derived growth factor. Physiol Rev 79:1283–1316.
Janeczko K. 1988. The proliferative response of astrocytes to injury inneonatal rat brain. A combined immunocytochemical and autoradio-graphic study. Brain Res 456:280–285.
Jones LL, Margolis RU, Tuszynski MH. 2003. The chondroitin sulfateproteoglycans neurocan, brevican, phosphacan, and versican are dif-ferentially regulated following spinal cord injury. Exp Neurol 182:399–411.
Kawakami Y, Tsuda M, Takahashi S, Taniguchi N, Esteban CR, Zem-myo M, Furumatsu T, Lotz M, Belmonte JC, Asahara H. 2005. Tran-scriptional coactivator PGC-1a regulates chondrogenesis via associa-tion with Sox9. Proc Natl Acad Sci USA 102:2414–2419.
Kearns AE, Campbell SC, Westley J, Schwartz NB. 1991. Initiation ofchondroitin sulfate biosynthesis: A kinetic analysis of UDP-D-xylose:-core protein b-D-xylosyltransferase. Biochemistry 30:7477–7483.
Kolb M, Margetts PJ, Galt T, Sime PJ, Xing Z, Schmidt M, Gauldie J.2001. Transient transgene expression of decorin in the lung reducesthe fibrotic response to bleomycin. Am J Respir Crit Care Med 163(3,Part 1):770–777.
Kordes U, Cheng YC, Scotting PJ. 2005. Sox group E gene expressiondistinguishes different types and maturational stages of glial cells indeveloping chick and mouse. Brain Res Dev Brain Res 157:209–213.
Lagord C, Berry M, Logan A. 2002. Expression of TGFb2 but notTGFb1 correlates with the deposition of scar tissue in the lesionedspinal cord. Mol Cell Neurosci 20:69–92.
Lefebvre V, Huang W, Harley VR, Goodfellow PN, de Crombrugghe B.1997. SOX9 is a potent activator of the chondrocyte-specific enhancerof the pro a1(II) collagen gene. Mol Cell Biol 17:2336–2346.
Liesi P, Silver J. 1988. Is astrocyte laminin involved in axon guidancein the mammalian CNS? Dev Biol 130:774–785.
Logan A, Baird A, Berry M. 1999a. Decorin attenuates gliotic scar for-mation in the rat cerebral hemisphere. Exp Neurol 159:504–510.
Logan A, Green J, Hunter A, Jackson R, Berry M. 1999b. Inhibition ofglial scarring in the injured rat brain by a recombinant human mono-clonal antibody to transforming growth factor-b2. Eur J Neurosci11:2367–2374.
Matthiessen HP, Schmalenbach C, Muller HW. 1989. Astroglia-releasedneurite growth-inducing activity for embryonic hippocampal neuronsis associated with laminin bound in a sulfated complex and free fibro-nectin. Glia 2:177–188.
McKeon RJ, Jurynec MJ, Buck CR. 1999. The chondroitin sulfate pro-teoglycans neurocan and phosphacan are expressed by reactive astro-cytes in the chronic CNS glial scar. J Neurosci 19:10778–10788.
McKeon RJ, Schreiber RC, Rudge JS, Silver J. 1991. Reduction of neu-rite outgrowth in a model of glial scarring following CNS injury iscorrelated with the expression of inhibitory molecules on reactiveastrocytes. J Neurosci 11:3398–3411.
Morgenstern DA, Asher RA, Fawcett JW. 2002. Chondroitin sulphate pro-teoglycans in the CNS injury response. Prog Brain Res 137:313–332.
Nakamura M, Okada S, Toyama Y, Okano H. 2005. Role of IL-6 in spinalcord injury in a mouse model. Clin Rev Allergy Immunol 28:197–204.
Nelander S, Larsson E, Kristiansson E, Mansson R, Nerman O, Sig-vardsson M, Mostad P, Lindahl P. 2005. Predictive screening for regu-lators of conserved functional gene modules (gene batteries) in mam-mals. BMC Genomics 6:68.
Neugebauer KM, Tomaselli KJ, Lilien J, Reichardt LF. 1988. N-cad-herin, NCAM, and integrins promote retinal neurite outgrowth onastrocytes in vitro. J Cell Biol 107:1177–1187.
Okada S, Nakamura M, Mikami Y, Shimazaki T, Mihara M, Ohsugi Y,Iwamoto Y, Yoshizaki K, Kishimoto T, Toyama Y, and others. 2004.Blockade of interleukin-6 receptor suppresses reactive astrogliosisand ameliorates functional recovery in experimental spinal cordinjury. J Neurosci Res 76:265–276.
Pasinetti GM, Nichols NR, Tocco G, Morgan T, Laping N, Finch CE.1993. Transforming growth factor b1 and fibronectin messenger RNAin rat brain: Responses to injury and cell-type localization. Neuro-science 54:893–907.
Reier PJ, Houle JD. 1988. The glial scar: Its bearing on axonal elonga-tion and transplantation approaches to CNS repair. Adv Neurol47:87–138.
Rogers SL, Letourneau PC, Palm SL, McCarthy J, Furcht LT. 1983.Neurite extension by peripheral and central nervous system neuronsin response to substratum-bound fibronectin and laminin. Dev Biol98:212–220.
Rogers SL, Letourneau PC, Peterson BA, Furcht LT, McCarthy JB.1987. Selective interaction of peripheral and central nervous systemcells with two distinct cell-binding domains of fibronectin. J Cell Biol105:1435–1442.
Schaeren-Wiemers N, Gerfin-Moser A. 1993. A single protocol to detecttranscripts of various types and expression levels in neural tissueand cultured cells: In situ hybridization using digoxigeni-labeledcRNA probes. Histochemistry 100:431–440.
Schonherr E, Jarvelainen HT, Sandell LJ, Wight TN. 1991. Effects ofplatelet-derived growth factor and transforming growth factor-b1 onthe synthesis of a large versican-like chondroitin sulfate proteoglycanby arterial smooth muscle cells. J Biol Chem 266:17640–17647.
Schwartz NB. 1977. Regulation of chondroitin sulfate synthesis. Effectof b-xylosides on synthesis of chondroitin sulfate proteoglycan, chon-droitin sulfate chains, and core protein. J Biol Chem 252:6316–6321.
Silver J, Miller JH. 2004. Regeneration beyond the glial scar. Nat RevNeurosci 5:146–156.
Stolt CC, Lommes P, Sock E, Chaboissier MC, Schedl A, Wegner M.2003. The Sox9 transcription factor determines glial fate choice inthe developing spinal cord. Genes Dev 17:1677–1689.
Tom VJ, Doller CM, Malouf AT, Silver J. 2004. Astrocyte-associated fi-bronectin is critical for axonal regeneration in adult white matter.J Neurosci 24:9282–9290.
Tomaselli KJ, Neugebauer KM, Bixby JL, Lilien J, Reichardt LF. 1988.N-cadherin and integrins: Two receptor systems that mediate neuro-nal process outgrowth on astrocyte surfaces. Neuron 1:33–43.
Vijayan VK, Lee YL, Eng LF. 1990. Increase in glial fibrillary acidicprotein following neural trauma. Mol Chem Neuropathol 13:107–118.
Wehrli BM, Huang W, De Crombrugghe B, Ayala AG, Czerniak B.2003. Sox9, a master regulator of chondrogenesis, distinguishes mes-enchymal chondrosarcoma from other small blue round cell tumors.Hum Pathol 34:263–269.
Wiksten M, Vaananen AJ, Liebkind R, Liesi P. 2004. Regeneration ofadult rat spinal cord is promoted by the soluble KDI domain of g1laminin. J Neurosci Res 78:403–410.
Wilson JX, Dixon SJ. 1989. Ascorbic acid transport in mouse and ratastrocytes is reversibly inhibited by furosemide, SITS, and DIDS.Neurochem Res 14:1169–1175.
Yamauchi S, Mita S, Matsubara T, Fukuta M, Habuchi H, Kimata K,Habuchi O. 2000. Molecular cloning and expression of chondroitin 4-sulfotransferase. J Biol Chem 275:8975–8981.
Zuo J, Neubauer D, Dyess K, Ferguson TA, Muir D. 1998. Degradationof chondroitin sulfate proteoglycan enhances the neurite-promotingpotential of spinal cord tissue. Exp Neurol 154:654–662.
1155SOX9 AND CYTOKINES REGULATE SCAR GENES
GLIA DOI 10.1002/glia
Related Documents











