Time-dependent fate of transplanted neural precursor cells in experimental autoimmune encephalomyelitis mice Angeliki Giannakopoulou a,b , Nikolaos Grigoriadis a, ⁎, Eleni Polyzoidou a , Athanasios Lourbopoulos a , Eleni Michaloudi b , Georgios C. Papadopoulos b a Second Department of Neurology, AHEPA University Hospital, Aristotle University of Thessaloniki, Macedonia, Greece b Laboratory of Anatomy, Histology and Embryology, Veterinary School, Aristotle University of Thessaloniki, Macedonia, Greece abstract article info Article history: Received 4 January 2010 Revised 23 March 2010 Accepted 12 April 2010 Available online xxxx Keywords: Neural stem cells EAE Multiple Sclerosis CNS Migration Inflammatory cytokines Transplantation TNFa TGFb INFγ Transplanted Neural Precursor Cells (NPCs) are capable of long-distance migration inside the inflamed CNS, but exhibit limited myelinating capacities in animal models of Multiple Sclerosis (MS). Inflammation seems to be both beneficial for the recruitment and migration of NPCs and restrictive for their terminal differentiation. In the present study, a set of transplantation experiments was applied in order to investigate the migratory potential, the differentiation pattern and long-term survival of NPCs in Experimental Autoimmune Encephalomyelitis (EAE) mice, the animal model of MS. The in vitro differentiation potential of NPCs in the presence of either pro- (TNFa, INFγ) or anti- (TGFb) inflammatory cytokines was also analyzed. According to the in vivo results obtained, at the acute phase of EAE only a small fraction of transplanted NPCs succeed to differentiate, whereas at chronic phase most of them followed a differentiation process to glial cell lineage along white matter tracts. However, this differentiation was not fully completed, since 8 months after their transplantation a number of NPCs remained as pre-oligodendrocytes. Glial differentiation of NPCs was also found to be inhibited or promoted following their treatment with TNFa or TGFb respectively, in vitro. Our findings suggest that inflammation triggers migration whereas the anti-inflammatory component is a prerequisite for NPCs to follow glial differentiation thereby providing myelinating oligodendrocytes. It is speculated that the fine balance between the pro- and anti-inflammatory determinants in the CNS may be a key factor for transplanted NPCs to exhibit a better therapeutic effect in EAE and MS. © 2010 Elsevier Inc. All rights reserved. Introduction Multiple Sclerosis (MS) is the most commonly diagnosed immune mediated demyelinating disease in young adults. Demyelination, axonal degeneration, neuronal dysfunction and apoptosis are key features in MS pathology and current evidence suggests that remyelination to some extent also occurs spontaneously (Lassmann et al., 1997; Patrikios et al., 2006). Adult brain contains Neural Precursor Cells (NPCs) in regional pools that are able to be engaged in endogenous repair (Galli et al., 2003). However, the size of the lesion may surpass the capacity of endogenous NPCs to repair the damage, or the same cells could become the targets of the pathological process with adverse repercussions on the innate ability of the CNS to self- repair (Pluchino and Martino, 2007). Therefore, in case the aetiology of disease progression resides in the host genome, transplantation of exogenous NPCs from a non-diseased source could be an option. Many studies have already proved that transplanted NPCs are capable of long-distance migration inside an inflamed CNS. They also exert immunosuppressive, anti-inflammatory and neurotrophic func- tions following their interaction with an inflammatory environment, in diseases such as EAE and MS (Einstein et al., 2006b; Pluchino et al., 2003). However, their myelinating capacities are limited, since a small percentage of transplanted NPCs differentiate into mature oligoden- drocytes. Although the inflammation seems to be important for the recruitment and migration of NPCs, (Picard-Riera et al., 2002) at the same it may be prohibitive for their terminal differentiation into cells of oligodendrocyte lineage. In animal models of MS, such as the Experimental Autoimmune Encephalomyelitis (EAE) transplantation experiments reveal that NPCs differentiate mainly into glia (Ben-Hur et al., 2003b, 2007; Einstein et al., 2003, 2006a,b; Pluchino et al., 2003). However, it is not known how the inflammatory environment dictates the differentia- tion fate of NPCs as disease develops and if there is any correlation between the differentiation of NPCs and the phase of disease or their localisation in the brain. Moreover, there is little knowledge about the long-term survival of transplanted NPCs after their integration into the host parenchyma as similar experiments were terminated maximum Experimental Neurology xxx (2010) xxx–xxx ⁎ Corresponding author. Second Department of Neurology, AHEPA University Hospital, Aristotle University of Thessaloniki, 1 Stilp. Kyriakidi str, 54636 Thessaloniki, Macedonia, Greece. E-mail address: [email protected] (N. Grigoriadis). YEXNR-10520; No. of pages: 11; 4C: 0014-4886/$ – see front matter © 2010 Elsevier Inc. All rights reserved. doi:10.1016/j.expneurol.2010.04.011 Contents lists available at ScienceDirect Experimental Neurology journal homepage: www.elsevier.com/locate/yexnr ARTICLE IN PRESS Please cite this article as: Giannakopoulou, A., et al., Time-dependent fate of transplanted neural precursor cells in experimental autoimmune encephalomyelitis mice, Exp. Neurol. (2010), doi:10.1016/j.expneurol.2010.04.011

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Experimental Neurology xxx (2010) xxx–xxx
YEXNR-10520; No. of pages: 11; 4C:
Contents lists available at ScienceDirect
Experimental Neurology
j ourna l homepage: www.e lsev ie r.com/ locate /yexnr
ARTICLE IN PRESS
Time-dependent fate of transplanted neural precursor cells in experimentalautoimmune encephalomyelitis mice
Angeliki Giannakopoulou a,b, Nikolaos Grigoriadis a,⁎, Eleni Polyzoidou a, Athanasios Lourbopoulos a,Eleni Michaloudi b, Georgios C. Papadopoulos b
a Second Department of Neurology, AHEPA University Hospital, Aristotle University of Thessaloniki, Macedonia, Greeceb Laboratory of Anatomy, Histology and Embryology, Veterinary School, Aristotle University of Thessaloniki, Macedonia, Greece
⁎ Corresponding author. Second Department of NHospital, Aristotle University of Thessaloniki, 1 Stilp. KyMacedonia, Greece.
E-mail address: [email protected] (N. Grigoriadi
0014-4886/$ – see front matter © 2010 Elsevier Inc. Adoi:10.1016/j.expneurol.2010.04.011
Please cite this article as: Giannakopoulou, Aencephalomyelitis mice, Exp. Neurol. (2010
a b s t r a c t
a r t i c l e i n f oArticle history:Received 4 January 2010Revised 23 March 2010Accepted 12 April 2010Available online xxxx
Keywords:Neural stem cellsEAEMultiple SclerosisCNSMigrationInflammatory cytokinesTransplantationTNFaTGFbINFγ
Transplanted Neural Precursor Cells (NPCs) are capable of long-distance migration inside the inflamed CNS,but exhibit limited myelinating capacities in animal models of Multiple Sclerosis (MS). Inflammation seemsto be both beneficial for the recruitment and migration of NPCs and restrictive for their terminaldifferentiation. In the present study, a set of transplantation experiments was applied in order to investigatethe migratory potential, the differentiation pattern and long-term survival of NPCs in ExperimentalAutoimmune Encephalomyelitis (EAE) mice, the animal model of MS. The in vitro differentiation potential ofNPCs in the presence of either pro- (TNFa, INFγ) or anti- (TGFb) inflammatory cytokines was also analyzed.According to the in vivo results obtained, at the acute phase of EAE only a small fraction of transplanted NPCssucceed to differentiate, whereas at chronic phase most of them followed a differentiation process to glial celllineage along white matter tracts. However, this differentiation was not fully completed, since 8 months aftertheir transplantation a number of NPCs remained as pre-oligodendrocytes. Glial differentiation of NPCs wasalso found to be inhibited or promoted following their treatment with TNFa or TGFb respectively, in vitro.Our findings suggest that inflammation triggers migration whereas the anti-inflammatory component is aprerequisite for NPCs to follow glial differentiation thereby providing myelinating oligodendrocytes. It isspeculated that the fine balance between the pro- and anti-inflammatory determinants in the CNS may be akey factor for transplanted NPCs to exhibit a better therapeutic effect in EAE and MS.
eurology, AHEPA Universityriakidi str, 54636 Thessaloniki,
s).
ll rights reserved.
., et al., Time-dependent fate of transplanted), doi:10.1016/j.expneurol.2010.04.011
© 2010 Elsevier Inc. All rights reserved.
Introduction
Multiple Sclerosis (MS) is the most commonly diagnosed immunemediated demyelinating disease in young adults. Demyelination,axonal degeneration, neuronal dysfunction and apoptosis are keyfeatures in MS pathology and current evidence suggests thatremyelination to some extent also occurs spontaneously (Lassmannet al., 1997; Patrikios et al., 2006). Adult brain contains NeuralPrecursor Cells (NPCs) in regional pools that are able to be engaged inendogenous repair (Galli et al., 2003). However, the size of the lesionmay surpass the capacity of endogenous NPCs to repair the damage, orthe same cells could become the targets of the pathological processwith adverse repercussions on the innate ability of the CNS to self-repair (Pluchino and Martino, 2007). Therefore, in case the aetiologyof disease progression resides in the host genome, transplantation ofexogenous NPCs from a non-diseased source could be an option.
Many studies have already proved that transplanted NPCs arecapable of long-distance migration inside an inflamed CNS. They alsoexert immunosuppressive, anti-inflammatory and neurotrophic func-tions following their interaction with an inflammatory environment,in diseases such as EAE and MS (Einstein et al., 2006b; Pluchino et al.,2003). However, their myelinating capacities are limited, since a smallpercentage of transplanted NPCs differentiate into mature oligoden-drocytes. Although the inflammation seems to be important for therecruitment and migration of NPCs, (Picard-Riera et al., 2002) at thesame it may be prohibitive for their terminal differentiation into cellsof oligodendrocyte lineage.
In animal models of MS, such as the Experimental AutoimmuneEncephalomyelitis (EAE) transplantation experiments reveal thatNPCs differentiate mainly into glia (Ben-Hur et al., 2003b, 2007;Einstein et al., 2003, 2006a,b; Pluchino et al., 2003). However, it is notknown how the inflammatory environment dictates the differentia-tion fate of NPCs as disease develops and if there is any correlationbetween the differentiation of NPCs and the phase of disease or theirlocalisation in the brain. Moreover, there is little knowledge about thelong-term survival of transplantedNPCs after their integration into thehost parenchyma as similar experiments were terminated maximum
neural precursor cells in experimental autoimmune

2 A. Giannakopoulou et al. / Experimental Neurology xxx (2010) xxx–xxx
ARTICLE IN PRESS
1–3 months after the graft implantation (Franklin et al., 1997; Shieldset al., 1999; Woodruff and Franklin, 1999).
In the present studywe studied i) the long-term survival,migrationand differentiation of transplanted NPCs in the inflammatory anddemyelinating environment of chronic EAE and ii) the in vitro differ-entiation potential of NPCs in the presence of either proinflammatory(TNFa, INFγ) or anti-inflammatory (TGFb) cytokines.
Materials and methods
Induction of chronic EAE and clinical evaluation of the animals
Animal experimentation received the approval of VeterinaryDirectorate of Thessaloniki and was conducted under compliancewith National Institutes of Health guidelines, Greek Governmentguidelines and the local ethics committee. All possible adequatemeasures were taken to minimize pain or discomfort of treatedanimals.
The induction of chronic EAEwas held in five to six-week-old C57Bl/6female mice, immunized subcutaneously with MOG 35–55 as describedpreviously by (Einstein et al., 2006b). Clinical signs of EAE appearedtypically after 10–12 days, reaching themaximum clinical score after 18–22 days (acutephase). Remissionperiodbegan25–30 dayspost induction(PI) of EAE with modest or little gradual clinical improvement thereafterup to the point where the animals entered a plateau (45–50 days PI,chronic phase). Animals were examined daily for neurological signs(score: 0: healthy or asymptomatic; 1: partial loss of tail tonicity; 2: tailparalysis, ataxia; 3: hind limbweakness and difficulty to roll over; 4: hindlimb paralysis; 5: tetraparesis; 6: death. Severely ill animals wererehydrated by intraperitoneal injections of saline until neurologicalimprovement that enabled them to feed themselves independently.
Isolation of NPCs from GFP+ newborn mice and growth of neurospheres
NPCs were isolated from the cerebral hemispheres of newbornGFP-expressing mice (“Green mouse FM131” C57BL/6 TgN(act-EGFP)OsbC14-Y01-FM131) (Okabe et al., 1997), courtesy of Dr. M. Okabe(Osaka University, Japan) as described previously (Einstein et al.,2006b). The cells were suspended in N2 medium, plated in T-75uncoated flask (107 cells per flask) and supplemented with 10 ng/mlbasic fibroblast growth factor (bFGF) (R&D) and 20 ng/ml epidermalgrowth factor (EGF) (R&D), added daily. Under these conditionsapproximately 0.2% of cells proliferated into clusters of small roundcells that grew into free-floating neurospheres within 7 days and thencollected and diluted in DMEM/F12 medium. A quantity of the spherepreparation was used for quantification of neurospheres and aftermechanical dissociation the total cell number and cell viability usingtrypan blue exclusion were measured. The neurosphere preparationfor transplantation experiments was diluted in DMEM/F12 to theconcentration of 500 neurospheres/μl approximately.
In vitro differentiation of mouse NPCs under inflammatory cytokines
To analyze the in vitro differentiation potential of neurospheres inthe presence of the inflammatory cytokines TNFa, INFγ, TGFb,approximately 300 floating spheres/150 μl N2 medium were adheredto 35 mm tissue culture dishes coated with 10 μg/ml poly-D-lysineand 10 μg/ml fibronectin (Sigma). Following washing with DMEM/F12, culture dishes were incubated in 2 ml N2 medium without FGF2and EGF for 2 and 5 days in 37 °C and 10% CO2. On the first dayfollowing the induction of differentiation, 500 IU/ml N2 medium ofTNFa (recombinant mouse tumor necrosis alpha), INFγ (recombinantmouse interferon γ), or TGF-b1 (recombinant human transformingGrowth Factor beta 1), all from R&D, was added separately in eachculture dish. The differentiation potential of these cells was assessedby double immunocytochemistry with anti-nestin (Chemicon, 1:200)
Please cite this article as: Giannakopoulou, A., et al., Time-dependent fateencephalomyelitis mice, Exp. Neurol. (2010), doi:10.1016/j.expneurol.2
and anti-GFAP (Dako, 1:800), anti-NG2 (Chemicon, 1:200) or anti-GalC (Chemicon, 1:100). Cells were incubated with primary anti-bodies overnight at 4 °C and exposed to the appropriate fluoro-chrome-labelled secondary antibodies for 1.5 h at RT (all from JacksonImmunoResearch Laboratories). Counterstaining was done with Dapi.Labelled cells were examined and photographed at appropriate wave-lengths using a Nikon fluorescent microscope.
Counting and statistical analysis of in vitro immunostained NPCs
Quantification was performed blinded from photomicrographssystematically acquired from three independent cultures. A total of60 digital photomicrographs for each cytokine and control and foreach pair of primary antibodies (20 of the first primary antibody,20 of the second primary antibody, and 20 of the Dapi-stainednuclei), were taken at 20× magnification. Using Adobe Photoshopsoftware, the images of immunoreactive cells were overlaid andquantified. The number of cells in which a nucleus was visible wascounted in each photomicrograph. Percentage numbers of positivecells that express each antigenic marker or coexpress both werecalculated for each photograph. All the percentages combined andaveraged to give a total percentage number for the antigenicmarkers and expressed as Mean±SE. Data was statisticallyevaluated using One Way ANOVA with Bonferroni multiplecomparison post-hoc testing between individual pairs of groupsand P values b0.05 were deemed significant.
Neurosphere transplantation
The immunized C57Bl/6 mice on day 7 after the induction of EAEwere used as recipients of GFP+ NPCs. Stereotactic surgery wasperformed under deep anesthesia with the combination of ketamine(100 mg/kg)/xylazin (10 mg/kg) and with the animals mounted in aminiaturized stereotactic frame. Each animal was injected with 5000spheres in a volume of 10 μl in both lateral ventricles (Coordinates:Lateral to Bregma=1 mm, Bregma zero, Ventral to Bregma=2.5 mm)(Paxinos and Franklin, 2001). Mice with EAE that received the in-jection of the DMEM/F12 medium without cells in the lateralventricles were used as controls.
Experimental schedule and brain fixation
Animals were divided into 2 groups: control and experimental, i.e.transplanted with NPCs. Each group was divided further into 3subgroups, according to the time of their sacrifice PI of EAE. Eachsubgroup of control group was consisted of 10 animals, while 1st, 2ndand 3rd subgroups of experimental group were consisted of 20, 40and 20 animals respectively. The 1st group was sacrificed at the peakof the disease (17 days PI), the 2nd group during the chronic phaseand the 3rd one, 8 months PI (Supplementary Fig. 1). Animals underdeep ether anesthesia received transcardial perfusion with 4%paraformaldehyde (PFA) solution in PBS pH 7.4. Brains were quicklyremoved, post fixed by immersion in the same fixative at 4 °C for 4 h.Afterwards tissues were placed in 30% sucrose solution in PBS, deepfrozen in isopentane and 6 μm sagittal and coronal serial sectionswere collected. For each section collected the next sectionwas thrownaway. All sections were examined with a fluorescent microscope forthe detection of GFP+ cells.
Measurement of transplanted cells migration
Spatial distribution of GFP+ cells was evaluated in a blindedmanner in each brain by microscopic visualization of serial sagittal orcoronal sections. The migratory potential of transplanted GFP+ cellswas estimated by composites of sagittal or coronal photomicrographs(at ×20 or ×10 magnification) comprising the entire migratory
of transplanted neural precursor cells in experimental autoimmune010.04.011

3A. Giannakopoulou et al. / Experimental Neurology xxx (2010) xxx–xxx
ARTICLE IN PRESS
pathway followed by transplanted cells. The distance covered by themigrating transplanted cells was measured from the edge of thelateral ventricle with the use Image of Pro Plus software and wasexpressed as a mean±SE for each region (e.g. corpus callosum,fimbria of hippocampus etc.) and for each animal group or phase ofdisease.
Immunofluorescent staining of NPCs in vivo
To determine whether transplanted cells were capable of differ-entiating along neural lineages, immunofluorescent stainings wereperformed. Section sampling was performed after the determinationof spatial distribution of transplanted cells and the definition of theserial section population. Moreover, the sagittal brain sectioning waschosen in order to include in one section all regions of interest (i.e.corpus callosum, fimbria of hippocampus, internal capsule, periven-tricular areas).
In line with this concept, brain area containing at least 20 GFP+cells per sagittal serial section were identified between L=0.49 mmand L=1.45 mm lateral to bregma. We used the principles of“systematic random sampling” based on which, serial sections (thefirst being randomly selected) were sampled in predeterminedintervals depending on the total number of sections per animal.Consequently, samples of 12 sections per animal per immunopheno-typic marker studied, were analyzed. Data were expressed as theaverage percentage of immunoreactive transplanted cells expressinga specific antigenic marker (per section)±SEM in any region ofinterest for each animal group.
Sections were incubated with the following primary antibodies:mouse IgG anti-Nestin (Chemicon, 1:200), mouse IgM anti-PSA-NCAM (Chemicon, 1:200), mouse IgG anti-NeuN (Chemicon, 1:500),mouse IgG anti-tubulin Beta III (Chemicon, 1:200), rabbit IgG Anti-NG2 (Chemicon, 1:200), mouse IgM anti-O4 (Chemicon, 1:100),mouse IgG anti-GalC (Chemicon, 1:100), mouse IgG anti-CNPase(Chemicon, 1:100) and rabbit IgG anti-GFAP Polyclonal (Dako,dilution 1:500). Secondary antibodies (all from Jackson ImmunoR-esearch Laboratories) were Rhodamine RedTM-X-conjugated andused at a dilution of 1:200. Counterstaining was done with Dapi.Labelled cells were examined and triads of digital microphotographs(one with fluorescent GFP+ cells, one with TRITC labelled primaryantibody, and one with Dapi-stained nuclei), were taken using aNikon fluorescent microscope. Under strict criteria and rigorousattention to detail, immunolabelling for each antigenic marker usedwere determined at numerous triads of microphotographs. GFPimmunohistochemistry was also performed in order to investigate ifyellowish cell like structures were necrotic or apoptotic GFP+ cells.Sections were incubated with rabbit IgG anti-GFP Polyclonal (Abcam,dilution 1:4000) diluted in PBS containing 5% NGS and 2.5% BSA for1 h at RT. After washes in PBS a labelled polymer-HRP anti-rabbit(Envision, dilution 1:200) was used for 30 min at RT and then DABwas added for 5 min to reveal GFP. After thorough wash with DW,hematoxylin (Merk) staining was performed for nuclear signalling.
Statistical analysis
All quantitative evaluations weremade in a uniform fashion by anobserver who was blinded to the experimental groups. Quantitativedata from several experiments were pooled to obtain the mean±SEfor each experimental group. Due to the restricted size of samples,Kruskal–Wallis test was implemented for each parameter. TheMann–Whitney U-test was used for comparisons between any twogroups. All analyses were conducted using the statistical softwareprogram SPSS v17. Differences were deemed statistically significantat Pb0.05.
Please cite this article as: Giannakopoulou, A., et al., Time-dependent fateencephalomyelitis mice, Exp. Neurol. (2010), doi:10.1016/j.expneurol.2
Results
Transplantation of NPCs attenuated the severity of clinical disease
The clinical score was evaluated daily in individual mice and thecumulative severity of disease “area under curve” was calculated atday 18 PI of EAE (peak of the acute phase), at day 50 PI (representingthe chronic phase) and 8 months PI. There was no difference in timeinitiation of the disease onset between controls and NPCs trans-planted animals. The decrease in disease severity by neurospheretransplantation was already noted at acute phase of disease, but themost significant attenuation of the clinical paralytic signs wasobserved at chronic phase (Supplementary Table 1).
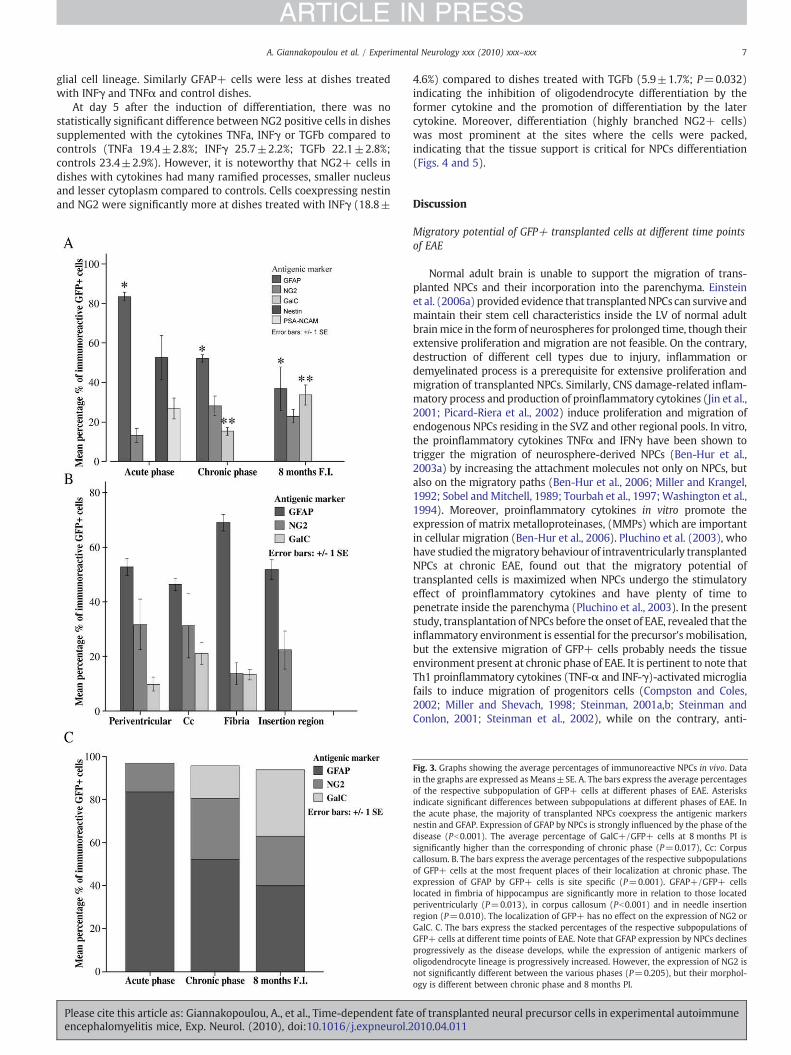
The spatial distribution and migratory potential of transplanted cells atdifferent phases of EAE
At the acute phase of the disease,many of the transplantedGFP+cellsremainedclusteredwithin theventricular space, attached to thewall of LVor trapped into the choroid plexus. A minority of them was found in theperiventricular parenchyma, up to a depth of several layers of cells.Extensive migration at this phase was limited toward inflammatoryinfiltrations adjacent to the LVs and in the border between thalamus andhippocampus (choroid fissure) (292.7±32.6 μm, Pb0.001 compared toperiventricular localization). Single GFP+ cells were also found along theneedle insertion site and nearby subpial surface due to a back-leakage ofsuspension.
At chronic phase, themigration of GFP+ cells wasmainly observedin white matter tracks lacking a clear cellular inflammatory processand especially in corpus callosum, ventricular hippocampal commis-sure, fimbria of hippocampus and internal capsule. Over 20 GFP+ cellsper section were detected between L=0.49 mm and L=1.45 mmlateral to bregma and B=−11.2 mm and B=0.6 mm rostra-caudal tobregma. However a percentage of GFP+ cells exhibited limited mi-gration, i.e. were found either to cover the LVs' wall or periventricularin proximity to the nuclei of septum and thalamus.
Eight months post transplantation; there were no transplantedGFP+ cells in the form of neurospheres or aggregates inside the LVs.All GFP+ cells integrated by the host parenchyma andwere located inthe same sites as their counterparts of chronic phase (Fig. 1). Compar-ison of the distances of GFP+ cells inside the brain parenchyma atdifferent time points indicates that there is statistical differencebetween acute and chronic phase (Kruskal–Wallis test, Pb0.001), butnot between chronic phase and 8 months PI. At chronic phase of EAEthe longest migratory pathwaywas inside the corpus callosum, wherethe greatest distance covered by GFP+ cells was found to be 1.4 mmat coronal sections (Table 1).
The differentiation potential of transplanted cells
The overall co-expression of the variousmarkers used is illustratedin Fig 2. Triads of images were used to identify GFP+ cells, simul-taneously immunoreactive for an individual marker and the respec-tive Dapi-stained nucleus (Supplementary Figs. 1, 2, 3). Used sectionswere thin enough not to permit overlapping of two different cells.
At the acute phase of EAE transplanted cells that remained in the formof aggregates of neurospheres expressed the antigenic markers of nestin(80.7±2.1%), glial fibrillary acid protein (GFAP, 76.2±3.9%) and thepolysialylated form of the neural adhesion molecule (PSA-NCAM, 30.8±7.5%). The average number of GFAP+/GFP+ and nestin+/GFP+ cellswere not significantly different (P=0.380), indicating that these twoantigenicmarkerswere coexpressed. TheGFP+cells that hadmoved intothe parenchyma in a depth of several cell layers acquired processes andmorphology of differentiating cells and lost the expression of nestin andPSA-NCAM. The GFAP was the major marker expressed by GFP+
of transplanted neural precursor cells in experimental autoimmune010.04.011
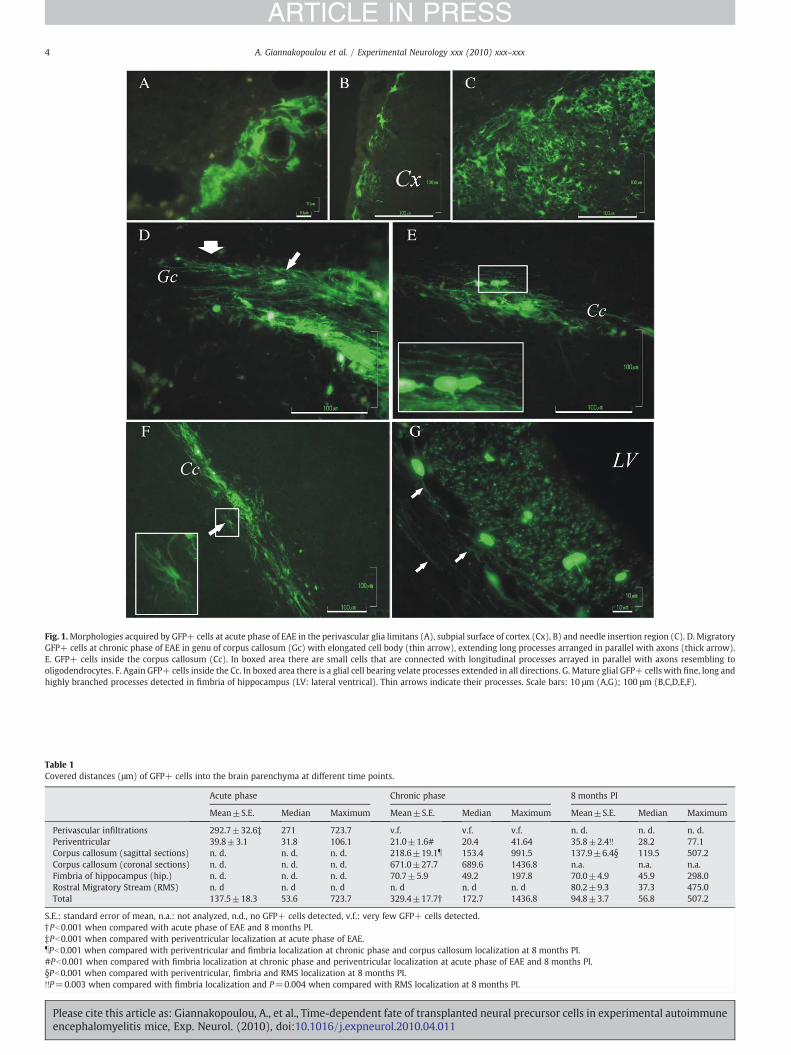
Fig. 1.Morphologies acquired by GFP+ cells at acute phase of EAE in the perivascular glia limitans (A), subpial surface of cortex (Cx), B) and needle insertion region (C). D. MigratoryGFP+ cells at chronic phase of EAE in genu of corpus callosum (Gc) with elongated cell body (thin arrow), extending long processes arranged in parallel with axons (thick arrow).E. GFP+ cells inside the corpus callosum (Cc). In boxed area there are small cells that are connected with longitudinal processes arrayed in parallel with axons resembling tooligodendrocytes. F. Again GFP+ cells inside the Cc. In boxed area there is a glial cell bearing velate processes extended in all directions. G. Mature glial GFP+ cells with fine, long andhighly branched processes detected in fimbria of hippocampus (LV: lateral ventrical). Thin arrows indicate their processes. Scale bars: 10 μm (A,G); 100 μm (B,C,D,E,F).
Table 1Covered distances (μm) of GFP+ cells into the brain parenchyma at different time points.
Acute phase Chronic phase 8 months PI
Mean±S.E. Median Maximum Mean±S.E. Median Maximum Mean±S.E. Median Maximum
Perivascular infiltrations 292.7±32.6‡ 271 723.7 v.f. v.f. v.f. n. d. n. d. n. d.Periventricular 39.8±3.1 31.8 106.1 21.0±1.6# 20.4 41.64 35.8±2.4!! 28.2 77.1Corpus callosum (sagittal sections) n. d. n. d. n. d. 218.6±19.1b 153.4 991.5 137.9±6.4§ 119.5 507.2Corpus callosum (coronal sections) n. d. n. d. n. d. 671.0±27.7 689.6 1436.8 n.a. n.a. n.a.Fimbria of hippocampus (hip.) n. d. n. d. n. d. 70.7±5.9 49.2 197.8 70.0±4.9 45.9 298.0Rostral Migratory Stream (RMS) n. d n. d n. d n. d n. d n. d 80.2±9.3 37.3 475.0Total 137.5±18.3 53.6 723.7 329.4±17.7† 172.7 1436.8 94.8±3.7 56.8 507.2
S.E.: standard error of mean, n.a.: not analyzed, n.d., no GFP+ cells detected, v.f.: very few GFP+ cells detected.†Pb0.001 when compared with acute phase of EAE and 8 months PI.‡Pb0.001 when compared with periventricular localization at acute phase of EAE.bPb0.001 when compared with periventricular and fimbria localization at chronic phase and corpus callosum localization at 8 months PI.#Pb0.001 when compared with fimbria localization at chronic phase and periventricular localization at acute phase of EAE and 8 months PI.§Pb0.001 when compared with periventricular, fimbria and RMS localization at 8 months PI.!!P=0.003 when compared with fimbria localization and P=0.004 when compared with RMS localization at 8 months PI.
4 A. Giannakopoulou et al. / Experimental Neurology xxx (2010) xxx–xxx
ARTICLE IN PRESS
Please cite this article as: Giannakopoulou, A., et al., Time-dependent fate of transplanted neural precursor cells in experimental autoimmuneencephalomyelitis mice, Exp. Neurol. (2010), doi:10.1016/j.expneurol.2010.04.011
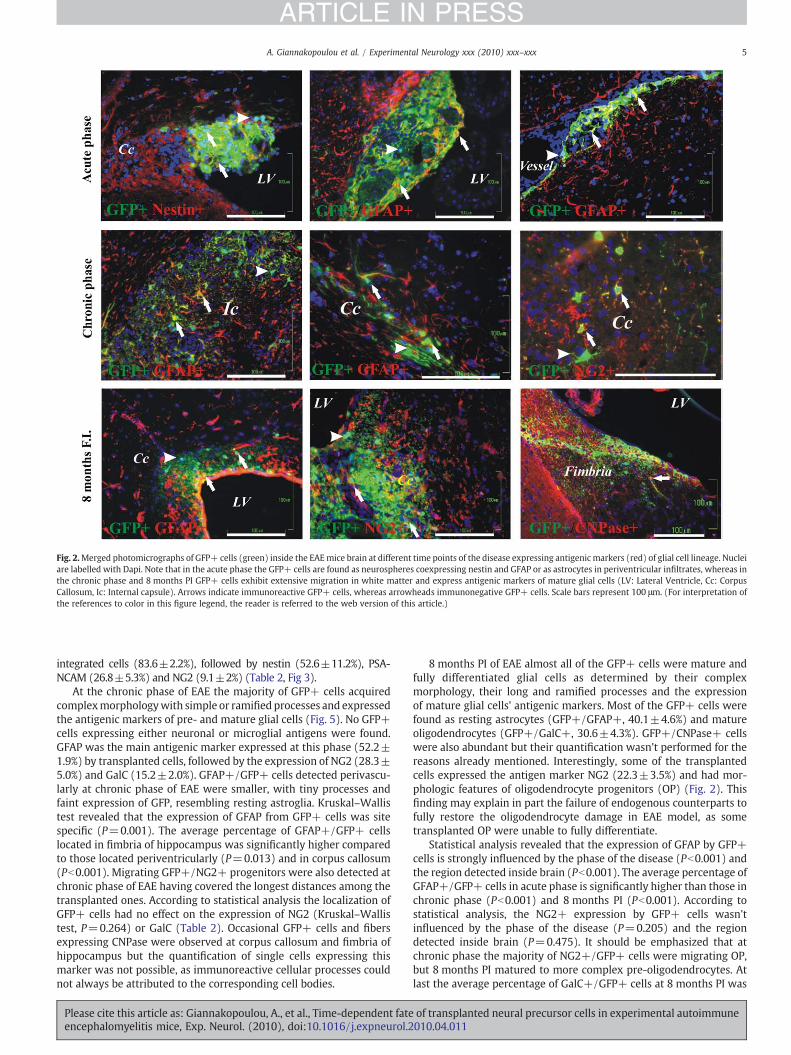
Fig. 2.Merged photomicrographs of GFP+ cells (green) inside the EAEmice brain at different time points of the disease expressing antigenic markers (red) of glial cell lineage. Nucleiare labelled with Dapi. Note that in the acute phase the GFP+ cells are found as neurospheres coexpressing nestin and GFAP or as astrocytes in periventricular infiltrates, whereas inthe chronic phase and 8 months PI GFP+ cells exhibit extensive migration in white matter and express antigenic markers of mature glial cells (LV: Lateral Ventricle, Cc: CorpusCallosum, Ic: Internal capsule). Arrows indicate immunoreactive GFP+ cells, whereas arrowheads immunonegative GFP+ cells. Scale bars represent 100 μm. (For interpretation ofthe references to color in this figure legend, the reader is referred to the web version of this article.)
5A. Giannakopoulou et al. / Experimental Neurology xxx (2010) xxx–xxx
ARTICLE IN PRESS
integrated cells (83.6±2.2%), followed by nestin (52.6±11.2%), PSA-NCAM (26.8±5.3%) and NG2 (9.1±2%) (Table 2, Fig 3).
At the chronic phase of EAE the majority of GFP+ cells acquiredcomplexmorphologywith simple or ramified processes and expressedthe antigenic markers of pre- and mature glial cells (Fig. 5). No GFP+cells expressing either neuronal or microglial antigens were found.GFAP was the main antigenic marker expressed at this phase (52.2±1.9%) by transplanted cells, followed by the expression of NG2 (28.3±5.0%) and GalC (15.2±2.0%). GFAP+/GFP+ cells detected perivascu-larly at chronic phase of EAE were smaller, with tiny processes andfaint expression of GFP, resembling resting astroglia. Kruskal–Wallistest revealed that the expression of GFAP from GFP+ cells was sitespecific (P=0.001). The average percentage of GFAP+/GFP+ cellslocated in fimbria of hippocampus was significantly higher comparedto those located periventricularly (P=0.013) and in corpus callosum(Pb0.001). Migrating GFP+/NG2+ progenitors were also detected atchronic phase of EAE having covered the longest distances among thetransplanted ones. According to statistical analysis the localization ofGFP+ cells had no effect on the expression of NG2 (Kruskal–Wallistest, P=0.264) or GalC (Table 2). Occasional GFP+ cells and fibersexpressing CNPase were observed at corpus callosum and fimbria ofhippocampus but the quantification of single cells expressing thismarker was not possible, as immunoreactive cellular processes couldnot always be attributed to the corresponding cell bodies.
Please cite this article as: Giannakopoulou, A., et al., Time-dependent fateencephalomyelitis mice, Exp. Neurol. (2010), doi:10.1016/j.expneurol.2
8 months PI of EAE almost all of the GFP+ cells were mature andfully differentiated glial cells as determined by their complexmorphology, their long and ramified processes and the expressionof mature glial cells' antigenic markers. Most of the GFP+ cells werefound as resting astrocytes (GFP+/GFAP+, 40.1±4.6%) and matureoligodendrocytes (GFP+/GalC+, 30.6±4.3%). GFP+/CNPase+ cellswere also abundant but their quantification wasn't performed for thereasons already mentioned. Interestingly, some of the transplantedcells expressed the antigen marker NG2 (22.3±3.5%) and had mor-phologic features of oligodendrocyte progenitors (OP) (Fig. 2). Thisfinding may explain in part the failure of endogenous counterparts tofully restore the oligodendrocyte damage in EAE model, as sometransplanted OP were unable to fully differentiate.
Statistical analysis revealed that the expression of GFAP by GFP+cells is strongly influenced by the phase of the disease (Pb0.001) andthe region detected inside brain (Pb0.001). The average percentage ofGFAP+/GFP+ cells in acute phase is significantly higher than those inchronic phase (Pb0.001) and 8 months PI (Pb0.001). According tostatistical analysis, the NG2+ expression by GFP+ cells wasn'tinfluenced by the phase of the disease (P=0.205) and the regiondetected inside brain (P=0.475). It should be emphasized that atchronic phase the majority of NG2+/GFP+ cells were migrating OP,but 8 months PI matured to more complex pre-oligodendrocytes. Atlast the average percentage of GalC+/GFP+ cells at 8 months PI was
of transplanted neural precursor cells in experimental autoimmune010.04.011
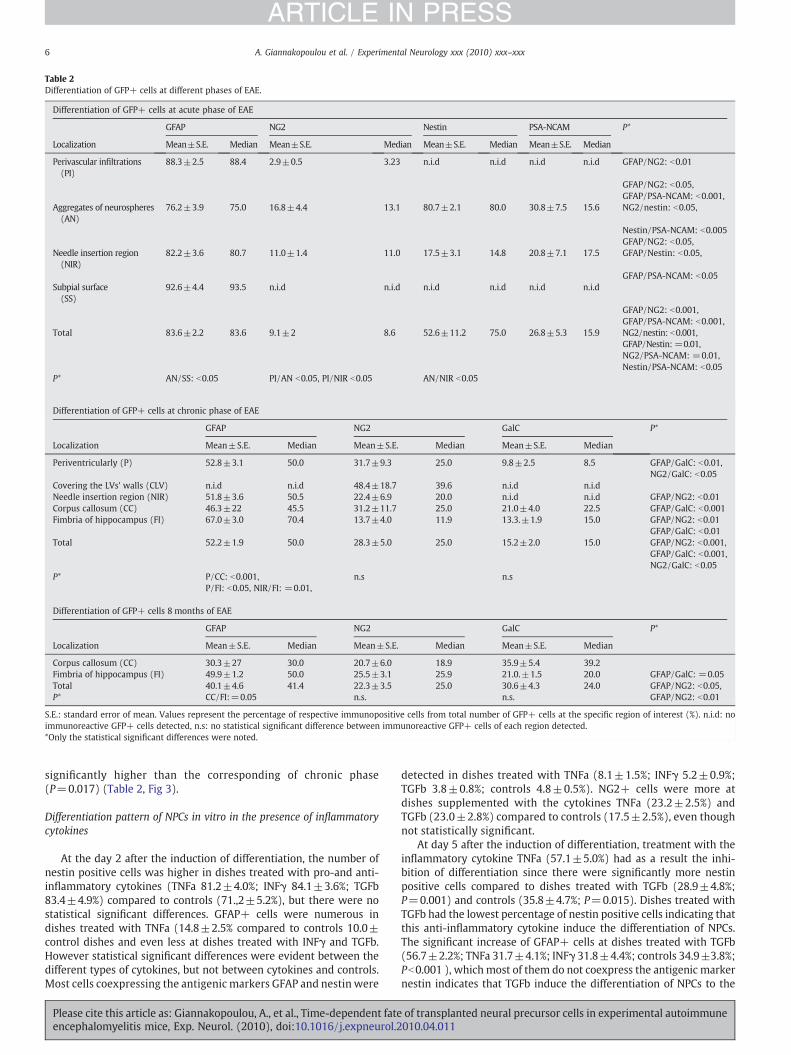
Table 2Differentiation of GFP+ cells at different phases of EAE.
Differentiation of GFP+ cells at acute phase of EAE
GFAP NG2 Nestin PSA-NCAM P*
Localization Mean±S.E. Median Mean±S.E. Median Mean±S.E. Median Mean±S.E. Median
Perivascular infiltrations(PI)
88.3±2.5 88.4 2.9±0.5 3.23 n.i.d n.i.d n.i.d n.i.d GFAP/NG2: b0.01
GFAP/NG2: b0.05,GFAP/PSA-NCAM: b0.001,
Aggregates of neurospheres(AN)
76.2±3.9 75.0 16.8±4.4 13.1 80.7±2.1 80.0 30.8±7.5 15.6 NG2/nestin: b0.05,
Nestin/PSA-NCAM: b0.005GFAP/NG2: b0.05,
Needle insertion region(NIR)
82.2±3.6 80.7 11.0±1.4 11.0 17.5±3.1 14.8 20.8±7.1 17.5 GFAP/Nestin: b0.05,
GFAP/PSA-NCAM: b0.05Subpial surface(SS)
92.6±4.4 93.5 n.i.d n.i.d n.i.d n.i.d n.i.d n.i.d
GFAP/NG2: b0.001,GFAP/PSA-NCAM: b0.001,
Total 83.6±2.2 83.6 9.1±2 8.6 52.6±11.2 75.0 26.8±5.3 15.9 NG2/nestin: b0.001,GFAP/Nestin: =0.01,NG2/PSA-NCAM: =0.01,Nestin/PSA-NCAM: b0.05
P* AN/SS: b0.05 PI/AN b0.05, PI/NIR b0.05 AN/NIR b0.05
Differentiation of GFP+ cells at chronic phase of EAE
GFAP NG2 GalC P*
Localization Mean±S.E. Median Mean±S.E. Median Mean±S.E. Median
Periventricularly (P) 52.8±3.1 50.0 31.7±9.3 25.0 9.8±2.5 8.5 GFAP/GalC: b0.01,NG2/GalC: b0.05
Covering the LVs' walls (CLV) n.i.d n.i.d 48.4±18.7 39.6 n.i.d n.i.dNeedle insertion region (NIR) 51.8±3.6 50.5 22.4±6.9 20.0 n.i.d n.i.d GFAP/NG2: b0.01Corpus callosum (CC) 46.3±22 45.5 31.2±11.7 25.0 21.0±4.0 22.5 GFAP/GalC: b0.001Fimbria of hippocampus (FI) 67.0±3.0 70.4 13.7±4.0 11.9 13.3.±1.9 15.0 GFAP/NG2: b0.01
GFAP/GalC: b0.01Total 52.2±1.9 50.0 28.3±5.0 25.0 15.2±2.0 15.0 GFAP/NG2: b0.001,
GFAP/GalC: b0.001,NG2/GalC: b0.05
P* P/CC: b0.001, n.s n.sP/FI: b0.05, NIR/FI: =0.01,
Differentiation of GFP+ cells 8 months of EAE
GFAP NG2 GalC P*
Localization Mean±S.E. Median Mean±S.E. Median Mean±S.E. Median
Corpus callosum (CC) 30.3±27 30.0 20.7±6.0 18.9 35.9±5.4 39.2Fimbria of hippocampus (FI) 49.9±1.2 50.0 25.5±3.1 25.9 21.0.±1.5 20.0 GFAP/GalC: =0.05Total 40.1±4.6 41.4 22.3±3.5 25.0 30.6±4.3 24.0 GFAP/NG2: b0.05,P* CC/FI:=0.05 n.s. n.s. GFAP/NG2: b0.01
S.E.: standard error of mean. Values represent the percentage of respective immunopositive cells from total number of GFP+ cells at the specific region of interest (%). n.i.d: noimmunoreactive GFP+ cells detected, n.s: no statistical significant difference between immunoreactive GFP+ cells of each region detected.*Only the statistical significant differences were noted.
6 A. Giannakopoulou et al. / Experimental Neurology xxx (2010) xxx–xxx
ARTICLE IN PRESS
significantly higher than the corresponding of chronic phase(P=0.017) (Table 2, Fig 3).
Differentiation pattern of NPCs in vitro in the presence of inflammatorycytokines
At the day 2 after the induction of differentiation, the number ofnestin positive cells was higher in dishes treated with pro-and anti-inflammatory cytokines (TNFa 81.2±4.0%; INFγ 84.1±3.6%; TGFb83.4±4.9%) compared to controls (71.,2±5.2%), but there were nostatistical significant differences. GFAP+ cells were numerous indishes treated with TNFa (14.8±2.5% compared to controls 10.0±control dishes and even less at dishes treated with INFγ and TGFb.However statistical significant differences were evident between thedifferent types of cytokines, but not between cytokines and controls.Most cells coexpressing the antigenic markers GFAP and nestin were
Please cite this article as: Giannakopoulou, A., et al., Time-dependent fateencephalomyelitis mice, Exp. Neurol. (2010), doi:10.1016/j.expneurol.2
detected in dishes treated with TNFa (8.1±1.5%; INFγ 5.2±0.9%;TGFb 3.8±0.8%; controls 4.8±0.5%). NG2+ cells were more atdishes supplemented with the cytokines TNFa (23.2±2.5%) andTGFb (23.0±2.8%) compared to controls (17.5±2.5%), even thoughnot statistically significant.
At day 5 after the induction of differentiation, treatment with theinflammatory cytokine TNFa (57.1±5.0%) had as a result the inhi-bition of differentiation since there were significantly more nestinpositive cells compared to dishes treated with TGFb (28.9±4.8%;P=0.001) and controls (35.8±4.7%; P=0.015). Dishes treated withTGFb had the lowest percentage of nestin positive cells indicating thatthis anti-inflammatory cytokine induce the differentiation of NPCs.The significant increase of GFAP+ cells at dishes treated with TGFb(56.7±2.2%; TNFa 31.7±4.1%; INFγ 31.8±4.4%; controls 34.9±3.8%;Pb0.001 ), whichmost of them do not coexpress the antigenic markernestin indicates that TGFb induce the differentiation of NPCs to the
of transplanted neural precursor cells in experimental autoimmune010.04.011
7A. Giannakopoulou et al. / Experimental Neurology xxx (2010) xxx–xxx
ARTICLE IN PRESS
glial cell lineage. Similarly GFAP+ cells were less at dishes treatedwith INFγ and TNFα and control dishes.
At day 5 after the induction of differentiation, there was nostatistically significant difference between NG2 positive cells in dishessupplemented with the cytokines TNFa, INFγ or TGFb compared tocontrols (TNFa 19.4±2.8%; INFγ 25.7±2.2%; TGFb 22.1±2.8%;controls 23.4±2.9%). However, it is noteworthy that NG2+ cells indishes with cytokines had many ramified processes, smaller nucleusand lesser cytoplasm compared to controls. Cells coexpressing nestinand NG2 were significantly more at dishes treated with INFγ (18.8±
Please cite this article as: Giannakopoulou, A., et al., Time-dependent fateencephalomyelitis mice, Exp. Neurol. (2010), doi:10.1016/j.expneurol.2
4.6%) compared to dishes treated with TGFb (5.9±1.7%; P=0.032)indicating the inhibition of oligodendrocyte differentiation by theformer cytokine and the promotion of differentiation by the latercytokine. Moreover, differentiation (highly branched NG2+ cells)was most prominent at the sites where the cells were packed,indicating that the tissue support is critical for NPCs differentiation(Figs. 4 and 5).
Discussion
Migratory potential of GFP+ transplanted cells at different time pointsof EAE
Normal adult brain is unable to support the migration of trans-planted NPCs and their incorporation into the parenchyma. Einsteinet al. (2006a) provided evidence that transplantedNPCs can survive andmaintain their stem cell characteristics inside the LV of normal adultbrainmice in the formof neurospheres for prolonged time, though theirextensive proliferation and migration are not feasible. On the contrary,destruction of different cell types due to injury, inflammation ordemyelinated process is a prerequisite for extensive proliferation andmigration of transplanted NPCs. Similarly, CNS damage-related inflam-matory process and production of proinflammatory cytokines (Jin et al.,2001; Picard-Riera et al., 2002) induce proliferation and migration ofendogenous NPCs residing in the SVZ and other regional pools. In vitro,the proinflammatory cytokines TNFα and IFNγ have been shown totrigger the migration of neurosphere-derived NPCs (Ben-Hur et al.,2003a) by increasing the attachment molecules not only on NPCs, butalso on the migratory paths (Ben-Hur et al., 2006; Miller and Krangel,1992; Sobel andMitchell, 1989; Tourbah et al., 1997;Washington et al.,1994). Moreover, proinflammatory cytokines in vitro promote theexpression of matrix metalloproteinases, (MMPs) which are importantin cellular migration (Ben-Hur et al., 2006). Pluchino et al. (2003), whohave studied themigratory behaviour of intraventricularly transplantedNPCs at chronic EAE, found out that the migratory potential oftransplanted cells is maximized when NPCs undergo the stimulatoryeffect of proinflammatory cytokines and have plenty of time topenetrate inside the parenchyma (Pluchino et al., 2003). In the presentstudy, transplantation of NPCs before the onset of EAE, revealed that theinflammatory environment is essential for the precursor's mobilisation,but the extensive migration of GFP+ cells probably needs the tissueenvironment present at chronic phase of EAE. It is pertinent to note thatTh1 proinflammatory cytokines (TNF-α and INF-γ)-activatedmicrogliafails to induce migration of progenitors cells (Compston and Coles,2002; Miller and Shevach, 1998; Steinman, 2001a,b; Steinman andConlon, 2001; Steinman et al., 2002), while on the contrary, anti-
Fig. 3. Graphs showing the average percentages of immunoreactive NPCs in vivo. Datain the graphs are expressed as Means±SE. A. The bars express the average percentagesof the respective subpopulation of GFP+ cells at different phases of EAE. Asterisksindicate significant differences between subpopulations at different phases of EAΕ. Inthe acute phase, the majority of transplanted NPCs coexpress the antigenic markersnestin and GFAP. Expression of GFAP by NPCs is strongly influenced by the phase of thedisease (Pb0.001). The average percentage of GalC+/GFP+ cells at 8 months PI issignificantly higher than the corresponding of chronic phase (P=0.017), Cc: Corpuscallosum. B. The bars express the average percentages of the respective subpopulationsof GFP+ cells at the most frequent places of their localization at chronic phase. Theexpression of GFAP by GFP+ cells is site specific (P=0.001). GFAP+/GFP+ cellslocated in fimbria of hippocampus are significantly more in relation to those locatedperiventricularly (P=0.013), in corpus callosum (Pb0.001) and in needle insertionregion (P=0.010). The localization of GFP+ has no effect on the expression of NG2 orGalC. C. The bars express the stacked percentages of the respective subpopulations ofGFP+ cells at different time points of EAE. Note that GFAP expression by NPCs declinesprogressively as the disease develops, while the expression of antigenic markers ofoligodendrocyte lineage is progressively increased. However, the expression of NG2 isnot significantly different between the various phases (P=0.205), but their morphol-ogy is different between chronic phase and 8 months PI.
of transplanted neural precursor cells in experimental autoimmune010.04.011
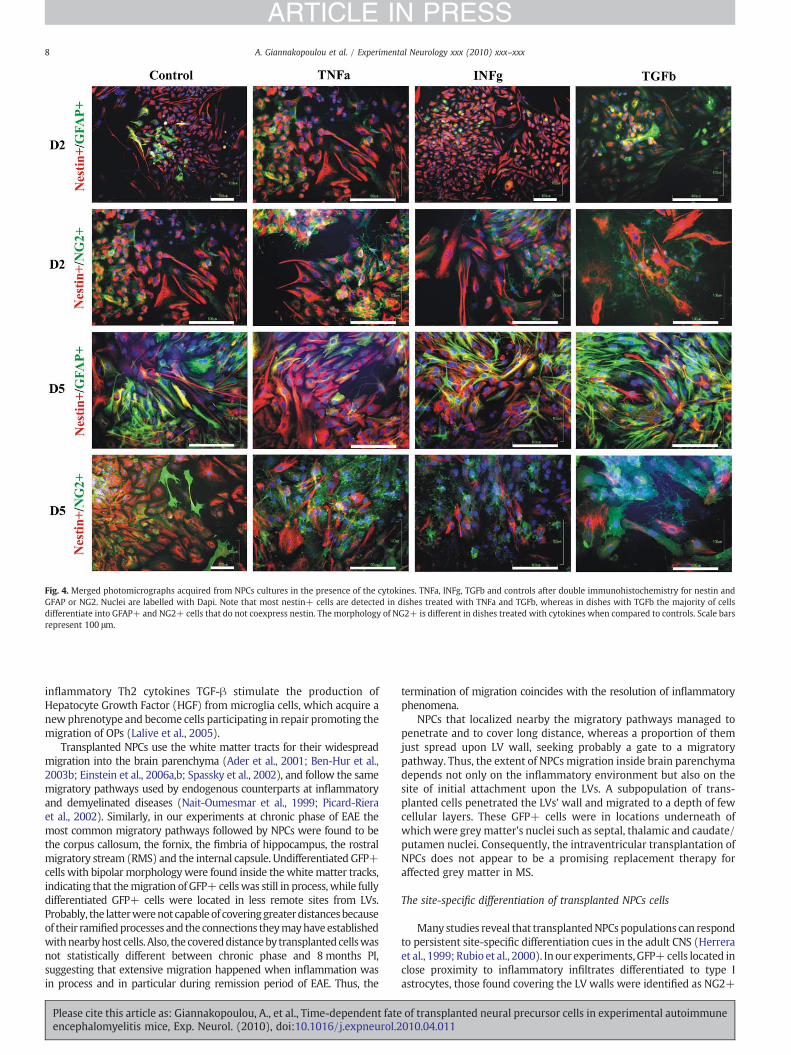
Fig. 4. Merged photomicrographs acquired from NPCs cultures in the presence of the cytokines. TNFa, INFg, TGFb and controls after double immunohistochemistry for nestin andGFAP or NG2. Nuclei are labelled with Dapi. Note that most nestin+ cells are detected in dishes treated with TNFa and TGFb, whereas in dishes with TGFb the majority of cellsdifferentiate into GFAP+ and NG2+ cells that do not coexpress nestin. The morphology of NG2+ is different in dishes treated with cytokines when compared to controls. Scale barsrepresent 100 μm.
8 A. Giannakopoulou et al. / Experimental Neurology xxx (2010) xxx–xxx
ARTICLE IN PRESS
inflammatory Th2 cytokines TGF-β stimulate the production ofHepatocyte Growth Factor (HGF) from microglia cells, which acquire anewphrenotype and become cells participating in repair promoting themigration of OPs (Lalive et al., 2005).
Transplanted NPCs use the white matter tracts for their widespreadmigration into the brain parenchyma (Ader et al., 2001; Ben-Hur et al.,2003b; Einstein et al., 2006a,b; Spassky et al., 2002), and follow the samemigratory pathways used by endogenous counterparts at inflammatoryand demyelinated diseases (Nait-Oumesmar et al., 1999; Picard-Rieraet al., 2002). Similarly, in our experiments at chronic phase of EAE themost common migratory pathways followed by NPCs were found to bethe corpus callosum, the fornix, the fimbria of hippocampus, the rostralmigratory stream (RMS) and the internal capsule. Undifferentiated GFP+cells with bipolar morphologywere found inside thewhitematter tracks,indicating that themigration of GFP+cellswas still in process, while fullydifferentiated GFP+ cells were located in less remote sites from LVs.Probably, the latterwerenot capableof coveringgreaterdistancesbecauseof their ramifiedprocesses and theconnections theymayhaveestablishedwithnearbyhost cells. Also, the covereddistanceby transplantedcellswasnot statistically different between chronic phase and 8 months PI,suggesting that extensive migration happened when inflammation wasin process and in particular during remission period of EAE. Thus, the
Please cite this article as: Giannakopoulou, A., et al., Time-dependent fateencephalomyelitis mice, Exp. Neurol. (2010), doi:10.1016/j.expneurol.2
termination of migration coincides with the resolution of inflammatoryphenomena.
NPCs that localized nearby the migratory pathways managed topenetrate and to cover long distance, whereas a proportion of themjust spread upon LV wall, seeking probably a gate to a migratorypathway. Thus, the extent of NPCs migration inside brain parenchymadepends not only on the inflammatory environment but also on thesite of initial attachment upon the LVs. A subpopulation of trans-planted cells penetrated the LVs' wall and migrated to a depth of fewcellular layers. These GFP+ cells were in locations underneath ofwhich were grey matter's nuclei such as septal, thalamic and caudate/putamen nuclei. Consequently, the intraventricular transplantation ofNPCs does not appear to be a promising replacement therapy foraffected grey matter in MS.
The site-specific differentiation of transplanted NPCs cells
Many studies reveal that transplantedNPCs populations can respondto persistent site-specific differentiation cues in the adult CNS (Herreraet al., 1999; Rubio et al., 2000). In our experiments, GFP+cells located inclose proximity to inflammatory infiltrates differentiated to type Iastrocytes, those found covering the LV walls were identified as NG2+
of transplanted neural precursor cells in experimental autoimmune010.04.011

9A. Giannakopoulou et al. / Experimental Neurology xxx (2010) xxx–xxx
ARTICLE IN PRESS
ependymal cells in agreement with their morphological characteristics,and a large proportion of GFP+ cells located inside the white mattertracks were positive for the antigenic markers of OP and matureoligodendrocytes. In corpus callosum, the transplanted cells differenti-ated in astrocytes and oligodendrocytes, with the percentage of thesetwo cell types having no statistical difference, while in the fimbria ofhippocampus they preferred to follow an astrocyte fate. Finally, thefailure of NPCs to neuronal differentiation should be attributed to their
Please cite this article as: Giannakopoulou, A., et al., Time-dependent fateencephalomyelitis mice, Exp. Neurol. (2010), doi:10.1016/j.expneurol.2
inability to migrate into grey matter and neurogenic structures such asRMS, olfactory bulb (OB) and cortex.
Differentiation arrest of transplanted NPCs at acute phase of EAE due toproinflammatory cytokines
At the inflammatory environment of acute phase of EAE most ofthe transplanted NPCs remained undifferentiated and only a smallfraction of them located in close proximity to inflammatory focidifferentiatedmainly into activated GFAP+/GFP+ astrocytes hedgingagainst inflammatory infiltration. Recently, Pluchino and Martino(2006, 2007) proposed that the inflammatory environment prevailingin EAE and MS should be accounted for the inability of endogenousNPCs derived from SVZs to differentiate into myelinating oligoden-drocytes. In an attempt to explain why most of the demyelinatedlesions in MS are detected periventicularly, suggested that the inflam-matory activity leads the NPCs of SVZ (concretely type B cells) toundergo symmetrical cellular divisions, thus preventing the produc-tion of glial precursors and their differentiation in myelinatingoligodendrocytes (Martino and Pluchino, 2006, 2007; Pluchino andMartino, 2007; Pluchino et al., 2008). This idea comes in agreementalso with our in vitro findings, where treatment of NPCs with theproinflammatory cytokines TNFa and INFγ results in the increase ofnestin positive cells 5 days later compared to controls.
Another likely explanation of differentiation arrest it could be thatthereweren't sufficient time forNPCs differentiation to oligodendrocytelineage. From transplantation to sacrifice the time interval was hardly11–14 days, while in similar experiments mature oligodendrocytes aredetected 30 days after transplantation of neurospheres or OP in vivo(Vitry et al., 2001). Besides, transplanted cells at the acute phase ofdisease had not yet been migrated in the parenchyma to interact withmicro-environment or demyelinated axons, so the initiative signallingfor their differentiation into oligodendrocytes was absent.
NPCs differentiation at chronic phase of EAE
At chronic phase of EAE, the lack of transplanted GFP+/nestin+cells contrasted the abundance of GFP+/NG2+ OP. This finding is inconsistence with the notion that Th2 cytokines, such as TGFβ, IL-4 andIL-10, which are also expressed during the effector phase of chronicEAE, are upregulated and peak during remission, resulting insuppression of immune response (Issazadeh et al., 1998). In thisline, the inflammatory cytokine TGFb has been proven here topromote in vitro the glial differentiation of NPCs. Th2 cytokines andneurotrophins, contrary to Th1 cytokines, are protective in experi-mental models of MS and induce the differentiation of NPCs and OPin vitro (Molina-Holgado et al., 2001). Chronic phase is alsocharacterized by decreased proportion of GFAP+/GFP+ cells andincreased proportion of NG2+/GFP+ cells. We believe that some
Fig. 5. Graphs showing the average percentages of immunoreactive NPCs in vitro aftertreatment with pro- and anti-inflammatory cytokines at D2 and D5 PID. Data in thegraphs are expressed as Means±SE. A. The bars express the average percentage ofnestin+ and GFAP+NPCS at D2 PID. No statistical significant differences were detectedin the expression of nestin by NPCs in different dishes. In terms of GFAP+ cells most ofthem were detected in dishes treated with TNFa. However, statistical significantdifferences were evident between the different types of cytokines, but not betweencytokine-treated and controls. Most cells coexpressing the antigenic markers GFAP andnestin were detected in dishes treated with TNFa. B. The bars express the averagepercentages of nestin+ and GFAP+ NPCs after treatment at D5 PID. Asterisks indicatesignificant differences between subpopulations at dishes treated with different types ofcytokines. There were significantly more nestin positive cells in dishes treated withTNFa compared to those treated with TGFb (P=0.001) and controls (P=0.015). Thesignificant increase of GFAP+ cells in dishes treated with TGFb (Pb0.001 ), which mostof them do not coexpress the antigenic marker nestin, indicates that TGFb induce thedifferentiation of NPCs to the glial cell lineage. C. The bars express the averagepercentages of NG2+ NPCs after treatment at D2 and D5 PID. No statistical significantdifferences were detected between cytokines treated and controls.
of transplanted neural precursor cells in experimental autoimmune010.04.011

10 A. Giannakopoulou et al. / Experimental Neurology xxx (2010) xxx–xxx
ARTICLE IN PRESS
GFAP+/GFP+ cells detected at the acute phase were not actuallydifferentiated astrocytes, but type B NPCs that later produce cells ofthe oligodendrocyte lineage (Doetsch et al., 1999; Menn et al., 2006).However, the fact that the majority of GFP+ cells in white mattertracks at chronic phase express the antigenic marker GFAP andpossess fine radiating processes calls for another explanation. It islikely that these cells represent embryonic astrocytes or have radialglial features in order to guide the endogenous NPCs migration, oralternatively constitute ‘isomorphic’ reactive astrocytes which allowdamage resolution (Williams et al., 2007). Blakemore et al. (2003)suggested that the influence of astrocytes on remyelination isdependent on their activation status and a distinction, therefore, hasto be made between the role of astrocytes in acute and chronic areasof demyelination.
The absence of intraparenchymalmigratory GFP+ cells eightmonthsafter their transplantation, in conjunction with the presence of GFP+/NG2+ cells showing morphological features of pre-oligodendrocytescould be attributed to the lack of either environmental cues calling forcellular differentiation or any more need for terminal differentiation tomature oligodendrocytes. In any way these GFP+/NG2+ cells, remain-ing as OP would probably be ready to form myelinating cells in a newdemyelinating attack. Unfortunately, this would be a short-termresponse, as there wouldn't be new NPCs to resupply them.
Therefore, it can be said that the inability of the transplanted (andprobably of the endogenous) NPCs to follow a differentiation processwhich might provide adequate myelinating oligodendrocytes is theresult of the persistent inflammatory environment prevailing in EAEand MS. Thus, the appropriate combination of immunoregulation andNPCs' transplantation might be considered as a therapeutic approachin MS. However, the possibility that anti-inflammatory or immuno-regulatory agends, particularly those currently used in MS treatment,might interfere with the kinetics and/or the behavior of transplantedNPCswithin CNS needs further clarification in future pharmacological-combined with cell-mediated- therapy studies.
Acknowledgments
We thank Ofira Einstein for experimental guidance, helpful con-sultations and constructive discussions. This work was supported byPENED 2001.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.expneurol.2010.04.011.
References
Ader, M., Schachner, M., Bartsch, U., 2001. Transplantation of neural precursor cells intothe dysmyelinated CNS of mutant mice deficient in the myelin-associatedglycoprotein and Fyn tyrosine kinase. Eur. J. NeuroSci. 14, 561–566.
Ben-Hur, T., Ben-Menachem, O., Furer, V., Einstein, O., Mizrachi-Kol, R., Grigoriadis, N.,2003a. Effects of proinflammatory cytokines on the growth, fate, and motility ofmultipotential neural precursor cells. Mol. Cell. Neurosci. 24, 623–631.
Ben-Hur, T., Ben-Yosef, Y., Mizrachi-Kol, R., Ben-Menachem, O., Miller, A., 2006.Cytokine-mediated modulation of MMPs and TIMPs in multipotential neuralprecursor cells. J. Neuroimmunol. 175, 12–18.
Ben-Hur, T., Einstein, O., Mizrachi-Kol, R., Ben-Menachem, O., Reinhartz, E., Karussis, D.,Abramsky, O., 2003b. Transplanted multipotential neural precursor cells migrateinto the inflamed white matter in response to experimental autoimmuneencephalomyelitis. Glia 41, 73–80.
Ben-Hur, T., van Heeswijk, R.B., Einstein, O., Aharonowiz, M., Xue, R., Frost, E.E., Mori, S.,Reubinoff, B.E., Bulte, J.W., 2007. Serial in vivo MR tracking of magnetically labeledneural spheres transplanted in chronic EAE mice. Magn. Reson. Med. 57, 164–171.
Blakemore, W.F., Gilson, J.M., Crang, A.J., 2003. The presence of astrocytes in areas ofdemyelination influences remyelination following transplantation of oligodendro-cyte progenitors. Exp. Neurol. 184, 955–963.
Compston, A., Coles, A., 2002. Multiple sclerosis. Lancet 359, 1221–1231.Doetsch, F., Caille, I., Lim, D.A., Garcia-Verdugo, J.M., Alvarez-Buylla, A., 1999.
Subventricular zone astrocytes are neural stem cells in the adult mammalianbrain. Cell 97, 703–716.
Please cite this article as: Giannakopoulou, A., et al., Time-dependent fateencephalomyelitis mice, Exp. Neurol. (2010), doi:10.1016/j.expneurol.2
Einstein, O., Ben-Menachem-Tzidon, O., Mizrachi-Kol, R., Reinhartz, E., Grigoriadis,N., Ben-Hur, T., 2006a. Survival of neural precursor cells in growth factor-poorenvironment: implications for transplantation in chronic disease. Glia 53,449–455.
Einstein, O., Grigoriadis, N., Mizrachi-Kol, R., Reinhartz, E., Polyzoidou, E., Lavon, I.,Milonas, I., Karussis, D., Abramsky, O., Ben-Hur, T., 2006b. Transplanted neuralprecursor cells reduce brain inflammation to attenuate chronic experimentalautoimmune encephalomyelitis. Exp. Neurol. 198, 275–284.
Einstein, O., Karussis, D., Grigoriadis, N., Mizrachi-Kol, R., Reinhartz, E., Abramsky, O.,Ben-Hur, T., 2003. Intraventricular transplantation of neural precursor cell spheresattenuates acute experimental allergic encephalomyelitis. Mol. Cell. Neurosci. 24,1074–1082.
Franklin, R.J., Gilson, J.M., Blakemore, W.F., 1997. Local recruitment of remyelinatingcells in the repair of demyelination in the central nervous system. J. Neurosci. Res.50, 337–344.
Galli, R., Gritti, A., Bonfanti, L., Vescovi, A.L., 2003. Neural stem cells: an overview. Circ.Res. 92, 598–608.
Herrera, D.G., Garcia-Verdugo, J.M., Alvarez-Buylla, A., 1999. Adult-derived neural precursorstransplanted into multiple regions in the adult brain. Ann. Neurol. 46, 867–877.
Issazadeh, S., Navikas, V., Schaub, M., Sayegh, M., Khoury, S., 1998. Kinetics ofexpression of costimulatory molecules and their ligands in murine relapsingexperimental autoimmune encephalomyelitis in vivo. J. Immunol. 161, 1104–1112.
Jin, K., Minami, M., Lan, J.Q., Mao, X.O., Batteur, S., Simon, R.P., Greenberg, D.A., 2001.Neurogenesis in dentate subgranular zone and rostral subventricular zone afterfocal cerebral ischemia in the rat. Proc. Natl. Acad. Sci. U. S. A. 98, 4710–4715.
Lalive, P.H., Paglinawan, R., Biollaz, G., Kappos, E.A., Leone, D.P., Malipiero, U., Relvas, J.B.,Moransard, M., Suter, T., Fontana, A., 2005. TGF-beta-treated microglia induceoligodendrocyte precursor cell chemotaxis through the HGF-c-Met pathway. Eur. J.Immunol. 35, 727–737.
Lassmann, H., Bruck, W., Lucchinetti, C., Rodriguez, M., 1997. Remyelination in multiplesclerosis. Mult. Scler. 3, 133–136.
Martino, G., Pluchino, S., 2006. The therapeutic potential of neural stem cells. Nat. Rev.Neurosci. 7, 395–406.
Martino, G., Pluchino, S., 2007. Neural stem cells: guardians of the brain. Nat. Cell Biol. 9,1031–1034.
Menn, B., Garcia-Verdugo, J.M., Yaschine, C., Gonzalez-Perez, O., Rowitch, D., Alvarez-Buylla, A., 2006. Origin of oligodendrocytes in the subventricular zone of the adultbrain. J. Neurosci. 26, 7907–7918.
Miller, M.D., Krangel, M.S., 1992. Biology and biochemistry of the chemokines: a familyof chemotactic and inflammatory cytokines. Crit. Rev. Immunol. 12, 17–46.
Miller, S.D., Shevach, E.M., 1998. Immunoregulation of experimental autoimmuneencephalomyelitis: editorial overview. Res. Immunol. 149, 753–759.
Molina-Holgado, E., Vela, J.M., Arevalo-Martin, A., Guaza, C., 2001. LPS/IFN-gammacytotoxicity in oligodendroglial cells: role of nitric oxide and protection by the anti-inflammatory cytokine IL-10. Eur. J. NeuroSci. 13, 493–502.
Nait-Oumesmar, B., Decker, L., Lachapelle, F., Avellana-Adalid, V., Bachelin, C., VanEvercooren, A.B., 1999. Progenitor cells of the adult mouse subventricular zoneproliferate, migrate and differentiate into oligodendrocytes after demyelination.Eur. J. NeuroSci. 11, 4357–4366.
Okabe, M., Ikawa, M., Kominami, K., Nakanishi, T., Nishimune, Y., 1997. ‘Greenmice’ as asource of ubiquitous green cells. FEBS Lett. 407, 313–319.
Patrikios, P., Stadelmann, C., Kutzelnigg, A., Rauschka, H., Schmidbauer, M., Laursen, H.,Sorensen, P.S., Bruck, W., Lucchinetti, C., Lassmann, H., 2006. Remyelination isextensive in a subset of multiple sclerosis patients. Brain 129, 3165–3172.
Paxinos, G., Franklin, K.B.J., 2001. TheMouse Brain in Stereotaxic Coordinates. AcademicPress.
Picard-Riera, N., Decker, L., Delarasse, C., Goude, K., Nait-Oumesmar, B., Liblau, R., Pham-Dinh, D., Evercooren, A.B., 2002. Experimental autoimmune encephalomyelitismobilizes neural progenitors from the subventricular zone to undergo oligoden-drogenesis in adult mice. Proc. Natl. Acad. Sci. U. S. A. 99, 13211–13216.
Pluchino, S., Martino, G., 2007. The therapeutic plasticity of neural stem/precursor cellsin multiple sclerosis. J. Neurol. Sci.
Pluchino, S., Muzio, L., Imitola, J., Deleidi, M., Alfaro-Cervello, C., Salani, G., Porcheri, C.,Brambilla, E., Cavasinni, F., Bergamaschi, A., Garcia-Verdugo, J.M., Comi, G., Khoury,S.J., Martino, G., 2008. Persistent inflammation alters the function of theendogenous brain stem cell compartment. Brain 131, 2564–2578.
Pluchino, S., Quattrini, A., Brambilla, E., Gritti, A., Salani, G., Dina, G., Galli, R., Del Carro,U., Amadio, S., Bergami, A., Furlan, R., Comi, G., Vescovi, A.L., Martino, G., 2003.Injection of adult neurospheres induces recovery in a chronic model of multiplesclerosis. Nature 422, 688–694.
Rubio, F.J., Bueno, C., Villa, A., Navarro, B., Martinez-Serrano, A., 2000. Geneticallyperpetuated human neural stem cells engraft and differentiate into the adultmammalian brain. Mol. Cell. Neurosci. 16, 1–13.
Shields, S.A., Gilson, J.M., Blakemore, W.F., Franklin, R.J., 1999. Remyelination occurs asextensively butmore slowly in old rats compared to young rats following gliotoxin-induced CNS demyelination. Glia 28, 77–83.
Sobel, R.A., Mitchell, M.E., 1989. Fibronectin in multiple sclerosis lesions. Am. J. Pathol.135, 161–168.
Spassky, N., de Castro, F., Le Bras, B., Heydon, K., Queraud-LeSaux, F., Bloch-Gallego, E.,Chedotal, A., Zalc, B., Thomas, J.L., 2002. Directional guidance of oligodendroglialmigration by class 3 semaphorins and netrin-1. J. Neurosci. 22, 5992–6004.
Steinman, L., 2001a. Blockade of gamma interferon might be beneficial in MS. MultScler. 7, 275–276.
Steinman, L., 2001b. Multiple sclerosis: a two-stage disease. Nat. Immunol. 2, 762–764.Steinman, L., Conlon, P., 2001. Antigen specific immunotherapy of multiple sclerosis.
J. Clin. Immunol. 21, 93–98.
of transplanted neural precursor cells in experimental autoimmune010.04.011

11A. Giannakopoulou et al. / Experimental Neurology xxx (2010) xxx–xxx
ARTICLE IN PRESS
Steinman, L., Martin, R., Bernard, C., Conlon, P., Oksenberg, J.R., 2002. Multiple sclerosis:deeper understanding of its pathogenesis reveals new targets for therapy. Annu.Rev. Neurosci. 25, 491–505.
Tourbah, A., Linnington, C., Bachelin, C., Avellana-Adalid, V., Wekerle, H., Baron-VanEvercooren, A., 1997. Inflammation promotes survival and migration of the CG4oligodendrocyte progenitors transplanted in the spinal cord of both inflammatoryand demyelinated EAE rats. J. Neurosci. Res. 50, 853–861.
Vitry, S., Avellana-Adalid, V., Lachapelle, F., Evercooren, A.B., 2001. Migration andmultipotentiality of PSA-NCAM+ neural precursors transplanted in the developingbrain. Mol. Cell. Neurosci. 17, 983–1000.
Please cite this article as: Giannakopoulou, A., et al., Time-dependent fateencephalomyelitis mice, Exp. Neurol. (2010), doi:10.1016/j.expneurol.2
Washington, R., Burton, J., Todd III, R.F., Newman, W., Dragovic, L., Dore-Duffy, P., 1994.Expression of immunologically relevant endothelial cell activation antigens onisolated central nervous systemmicrovessels from patients with multiple sclerosis.Ann. Neurol. 35, 89–97.
Williams, A., Piaton, G., Lubetzki, C., 2007. Astrocytes—friends or foes in multiplesclerosis? Glia 55, 1300–1312.
Woodruff, R.H., Franklin, R.J., 1999. Demyelination and remyelination of the caudalcerebellar peduncle of adult rats following stereotaxic injections of lysolecithin,ethidium bromide, and complement/anti-galactocerebroside: a comparativestudy. Glia 25, 216–228.
of transplanted neural precursor cells in experimental autoimmune010.04.011
Related Documents








![Myelin oligodendrocyte glycoprotein-specific antibodies from ......protein (MBP)] used to induce experimental autoimmune encephalomyelitis (EAE) in rodent models through induction](https://static.cupdf.com/doc/110x72/60ff0b7639f1f130b4007123/myelin-oligodendrocyte-glycoprotein-specific-antibodies-from-protein-mbp.jpg)


