DOI: 10.1126/scitranslmed.3004244 , 137rv6 (2012); 4 Sci Transl Med et al. Elaine Holmes Therapeutic Modulation of Microbiota-Host Metabolic Interactions http://stm.sciencemag.org/content/4/137/137rv6.full.html can be found at: and other services, including high-resolution figures, A complete electronic version of this article http://www.sciencemag.org/content/sci/336/6086/1262.full.html can be found online at: Related Resources for this article http://www.sciencemag.org/about/permissions.dtl in whole or in part can be found at: article permission to reproduce this of this article or about obtaining reprints Information about obtaining is a registered trademark of AAAS. Science Translational Medicine rights reserved. The title NW, Washington, DC 20005. Copyright 2012 by the American Association for the Advancement of Science; all last week in December, by the American Association for the Advancement of Science, 1200 New York Avenue (print ISSN 1946-6234; online ISSN 1946-6242) is published weekly, except the Science Translational Medicine on June 22, 2012 stm.sciencemag.org Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

DOI: 10.1126/scitranslmed.3004244, 137rv6 (2012);4 Sci Transl Med
et al.Elaine HolmesTherapeutic Modulation of Microbiota-Host Metabolic Interactions
http://stm.sciencemag.org/content/4/137/137rv6.full.htmlcan be found at:
and other services, including high-resolution figures,A complete electronic version of this article
http://www.sciencemag.org/content/sci/336/6086/1262.full.html can be found online at:Related Resources for this article
http://www.sciencemag.org/about/permissions.dtl in whole or in part can be found at: article
permission to reproduce this of this article or about obtaining reprintsInformation about obtaining
is a registered trademark of AAAS. Science Translational Medicinerights reserved. The title NW, Washington, DC 20005. Copyright 2012 by the American Association for the Advancement of Science; alllast week in December, by the American Association for the Advancement of Science, 1200 New York Avenue
(print ISSN 1946-6234; online ISSN 1946-6242) is published weekly, except theScience Translational Medicine
on
June
22,
201
2st
m.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om

REV I EW
METABOL I SM
Therapeutic Modulation of Microbiota-HostMetabolic InteractionsElaine Holmes,1 James Kinross,1 Glenn R. Gibson,2 Remy Burcelin,3 Wei Jia,4
Sven Pettersson,5,6 Jeremy K. Nicholson1*
The complex metabolic relationships between the host and its microbiota change throughout life and vary exten-sively between individuals, affecting disease risk factors and therapeutic responses through drug metabolism. Elu-cidating the biochemical mechanisms underlying this human supraorganism symbiosis is yielding new therapeuticinsights to improve human health, treat disease, and potentially modify human disease risk factors. Therapeuticoptions include targeting drugs to microbial genes or co-regulated host pathways and modifying the gut micro-biota through diet, probiotic and prebiotic interventions, bariatric surgery, fecal transplants, or ecological engi-neering. The age-associated co-development of the host and its microbiota provides a series of windows fortherapeutic intervention from early life through old age.
2
on Ju
ne 2
2, 2
01st
m.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om
INTRODUCTION
There is lifelong metabolic communication between the gut micro-biota and the mammalian host that develops and changes with time.There are many channels of chemical information exchange operatingbetween various microbial players and pathways in multiple hosttissue compartments [see Review by Nicholson et al. (1)]. These chan-nels operate in a similar way to hormonal communication in that spe-cific substances are released under certain physiological conditionsthat have targeted actions elsewhere in the body. In the case of thegut microbiota and the host, there are a number of metabolic axesabout which host and microbe interact (for example, metabolites ofbile acids produced by microbial enzyme action bind to host nuclearreceptors, linking microbes with cholesterol and steroid synthesis inthe host) (1).
As a deeper understanding of the multiple channels and mech-anisms of the metabolic (and linked immunological) axes of host-microbiota interactions emerges, so do new opportunities to exploitsuch knowledge for human health benefits through therapeutic target-ing of these microbe-host co-regulated pathways (1, 2). Attempts tomodulate the microbiota by controlling diet and using probioticsand prebiotics are well established (3), although underlying bio-chemical mechanisms are often lacking. Systems biology approachesare illuminating transgenomic cross-talk between the host genome(and signaling pathways) and the genomes (and metabolic products)of the microbiota and, in particular, how diet and the microbiotatogether influence host systemic biochemistry in animal models andhumans (4–8). However, there are other modalities for microbiota andmicrobiome manipulation (Fig. 1) that could yield new therapies fortreating a variety of common diseases caused by abnormalities of thegut microbiota and gut dysbiosis (where potentially hostile microbesproliferate at the expense of normal commensal and symbiont mi-
1Biomolecular Medicine, Department of Surgery and Cancer, Faculty of Medicine, ImperialCollege London, Exhibition Road, London SW7 2AZ, UK. 2Department of Food andNutritional Sciences, University of Reading, Whiteknights RG6 6AP, UK. 3INSERM, U1048,and Institut des Maladies Métaboliques et Cardiovasculaires I2MC, Hôpital Rangueil,BP84225, 31432 Toulouse, France. 4Department of Nutrition, University of North Carolinaat Greensboro, North Carolina Research Campus, Kannapolis, NC 28081, USA. 5Departmentof Microbiology, Tumor and Cell Biology, Karolinska Institutet, 117 77 Stockholm, Sweden.6School of Biological Sciences, Nanyang Technological University, 60 Nanyang Drive,Singapore 637551, Singapore.*To whom correspondence should be addressed. E-mail: [email protected]
www.S
crobes) (1). Such modalities include highly selective uses of targetedantibiotics (9) to remove or attenuate unwanted bacterial species,which may include nonpathogenic organisms that influence the manymicrobial niches of the gut. There are also more subtle approachesthat address the concept of the “druggable microbial genome” (2)where selective microbial activities are manipulated for host benefitwithout killing the bacteria themselves. This approach would capitalizeon knowledge of specific aspects of co-regulated signaling pathways inman and microbe, and drugs would be targeted specifically at micro-bial genes operating in key host-microbe metabolic and signalingpathways (1, 2) (Fig. 2). Microbial products, such as secondary meta-bolites that have drug-like activity, can also be harnessed for therapeu-tic purposes. Because molecular understanding of microbial actionadvances and new tools in synthetic biology become available, it is alsopossible to envisage ecological engineering of the microbial genome(microbiome) with modifications to genetic activities of individual or-ganisms or even microbial communities to benefit the host (Fig. 1)[see Review by Lemon et al. (9)]. However, these types of interventionsare still distant in terms of both our theoretical understanding andpractical implementation. The complex co-development of the gut mi-crobiota leads to different types of potential interventions at differentstages of a host’s life span (1, 5). Early interventions (before, say, 3years old) would be aimed at creating a long-term stable and “healthy”microbiota associated with disease prevention both in childhood andin adult life. Interventions later in life would be more geared totreating specific conditions, such as inflammatory bowel disease (1),but could also influence the longer-term health of the elderly as themicrobiota changes in old age (Fig. 1).
The evolution of drug-metabolizing enzymes has not been drivenby recent drug exposure, but rather by exposure of humans through-out history to plant toxins in the diet and microbial toxins and theneed to detoxify and eliminate them [see Perspective by Turnbaugh(10)] (2, 11). Indeed, there are multiple mechanisms of microbiota-host-drug interactions, and these can be modulated to enhance drugtherapy (for example, minimization of toxicity or enhancement of ef-ficacy). In the future, we can envisage how such modulation could beexploited for personalized healthcare (11, 12). In this Review, we ex-amine strategies for interventions at multiple levels of biomolecularorganization and host-microbiome “command and control” to opti-mize health, prevent disease, and develop new therapeutics (Fig. 1).
cienceTranslationalMedicine.org 6 June 2012 Vol 4 Issue 137 137rv6 1

REV I EW
Many of these interventions will not be practical without in-depthknowledge of the connections in the host-microbiota metabolic axes(1). The depth of association of gut microbiota abnormalities with in-creasingly common diseases such as inflammatory bowel disease, obe-sity, and diabetes makes the implementation of microbiome-basedtherapies an almost inevitable component of future personalizedand precision medicine (13).
on
June
22,
201
2st
m.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om
DRUG-MICROBIOTA-HOSTINTERACTIONS AND HUMANHEALTH
Because animals have evolved with spe-cific gut microbes and interdependent di-etary tolerances, so they have evolvedtailored detoxification systems to dealwith gut-derived toxins and xenobiotics(other compounds of nonmammalianorigin that enter the gut with the dietor are produced by the microbiota) (14).These detoxification systems include thecytochrome P450 enzymes of the hostgut, liver, and other organs that regulatephase 1 drug metabolism reactions (ox-idation, reduction, hydroxylation, etc.),which render foreign compounds morepolar and hence more easily eliminatedin the urine (13). So-called phase 2 drugmetabolism reactions are also used ex-tensively to modify gut microbial products.These involve conjugation of endogenousmetabolites or ions to foreign compoundsthrough reactions catalyzed by host en-zyme transfer systems that typically in-crease the polarity of foreign molecules,thus boosting their urinary excretion.These enzyme transfer systems includesulfotransferases that transfer sulfate tophenolic compounds and other hydrox-ylated species to form O-ether sulfates.For example, host sulfotransferases trans-fer sulfate to hydroxyl groups of phenoliccompounds, such as 4-cresol (which ismade from putrefaction of tyrosine inthe colon by microbes) that is then phase2–conjugated to form 4-cresyl sulfate, anabundant urinary microbial metabolite(14, 15). Other major phase 2 conjuga-tions use host uridine diphosphate–glucuronosyltransferase enzymes thattransfer D-glucuronic acid to phenolsand aromatic amines to form phenolicO-ether glucuronides or N-glucuronides,or to benzoic and phenylacetic acids toform O-ester glucuronides. Benzoateand phenylacetate can also be metabol-ically activated by host mitochondrial
Frewhwtyiaudsth
www.S
coenzyme A with subsequent enzymatic conjugation via aminoacid transferases with glycine and glutamine to form hippurateand phenylacetylglutamine, respectively, two of the most abundantgut microbial co-metabolites in man (16). The generation of benzoicand phenylacetic acids in the gut by microbial action may play an im-portant role in preventing excess systemic accumulation of the neuro-active amino acids glycine and glutamate (glutamate being theneuroactive deamination product of glutamine) (17).
Microbiotastatus
Health optimization
Disease prevention
• Probiotics• Prebiotics
• Probiotics• Prebiotics
Lifestyle
Bariatricsurgery
Environment:stress, allostatic
load
Dietarymodification
Druggingmicrobiome
Antibiotics
Ecologicalengineering
Targets:• Immune system • Endocannabinoids• Hormones• Bile acids• SCFAs• Biogenic amines• Xenometabolites
0yrs
• Brain development• Behavior
Synthetic biology
Pathogenresistance
Pathogentherapeutics
Helminth parasites
Fecal transplant
Energy
• Esophageal adenocarcinoma• Gastric ulcers• Nonalcoholic fatty liver disease• Metabolic syndrome• Drug toxicity
• Asthma• Atopy• Cystic fibrosis• Cardiovascular disease
Inflammatorybowel diseaseColon cancerObesitySepsisC. difficile
Maternal modulation
ss 12m 36m 12-18yr 25-35yr >36-59yr >60yr
Bact
eroi
dete
s
Firm
icut
es
Ther
apeu
tics
Hostgenome
Physiological programm
ing
Dysbiosis
Sym
bio
sis
ig. 1. Therapeutic modulation of the gut microbiota: from the cradle to the grave. The changinglationship between the gut microbiota and the human host throughout life offers a series of time
indows for therapeutic intervention. During infancy and early childhood, a healthy gut microbiotaelps to avoid dysbiosis that can lead to disease later in life. In adulthood, corrective strategies to dealith emergent dysbiosis and associated diseases such as inflammatory bowel disease, obesity, andpe 2 diabetes include modulating the microbiota using probiotics and prebiotics, antibiotics, bar-tric surgery, or by “drugging the microbiome.” Drugs targeted to microbial enzymes could also besed to boost the efficacy and reduce the toxicity of therapeutics. For extreme cases of gut-relatedisease, fecal transplantation or helminth therapy may prove beneficial. In the future, it may be pos-ible to prevent or treat abnormalities of the gut microbiota using modified organisms engineeredrough genetic or synthetic biology approaches.cienceTranslationalMedicine.org 6 June 2012 Vol 4 Issue 137 137rv6 2
REV I EW
on
June
22,
201
2st
m.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om
ebppbmsvthbmcbne
www.ScienceTranslationalMedicine.or
The phase 2 conjugating enzymes thatmodify gut microbial products alsoconjugate drugs that have certain func-tional groups. The commonly used drugacetaminophen (paracetamol, Tylenol) isextensively sulfated and glucuronidatedas are many other drugs that are also me-tabolized via cytochrome P450–basedphase 1 reactions. Thus, the enzymaticbasis of drug metabolism is directly linkedto this microbial metabolite detoxificationprocess, and so it is unsurprising to findthat gut microbial activities affect drugmetabolism and toxicity. Pharmaco-metabonomics is based on the notion thatpreinterventional metabolic signatures canbe used to predict post-interventional out-comes of drug treatments (18). Pharmaco-metabonomics has shown that the ratio ofsulfated to glucuronidated acetaminophen(when given at therapeutic doses) in humanurine correlates with the preinterventionalexcretion of the microbial co-metabolite4-cresyl sulfate. This implies that the dom-inant metabolic fate (glucuronidation andsulfation) of the drug acetaminophen de-pends on gut microbial activity that variesextensively between individuals (19). Thereare potentially hundreds of drugs that eitherhave hydroxyl groups or form hydrox-ylated metabolites that require sulfationfor detoxification and elimination. Thus,gut microbial activities in the distal colonthat produce 4-cresol from tyrosine couldaffect the metabolism and fate of manydrugs. This, in turn, may explain individ-ual variations in certain drug responsesthat are poorly predicted by pharmaco-genomics. This is just one example of adrug-microbial interaction (substrateconjugation competition) that affectsdrug metabolism and potentially toxicity.There are other gut microbial contribu-tions to host phenotype that can alterthe absorption, metabolism, and safetyof drugs and that may offer new directionsfor modulating drug activities [see Per-spective by Turnbaugh (10)]. These in-clude (i) competitive metabolic substrateutilization (as described above), (ii) pri-mary metabolism of orally administereddrugs by the gut microbiota, (iii) micro-bial secondary metabolite–mediated enzymeinduction, (iv) secondary metabolism of hu-man metabolites, and (v) gut bioavailability(local pH control and ionization state).
Such dependencies of conventionalxenobiotic metabolism and toxicity also
Viscerosensing
Leptin
Diet
EnvironmentAntibiotics
Bariatricsurgery
Druggingmicrobiome
Bile acids nuclear receptor axis
Endocannabinoid axis
Lifestyle
Aromatic acidphenolic axis
SCFA axis(e.g. Butyrate)
Mucosal angiogenesis
Immune systemaxis
Flavine monoxidasesystem
Inflammatory-metabolic axis
Neurotransmitteraxis
Choline-trimethylamineconversion axis
Ecological engineering
Parasite therapy
• Probiotics• Prebiotics• SynbioticsDietary
modulation
Renal excretion
Gut microbiota
Fecal transplant
Leptin receptor
Cannabinoidreceptor
Adipocytes
Cholesterol,steroid regulation
Insulin resistance
Drug discoverySynthetic
biology
anandamide
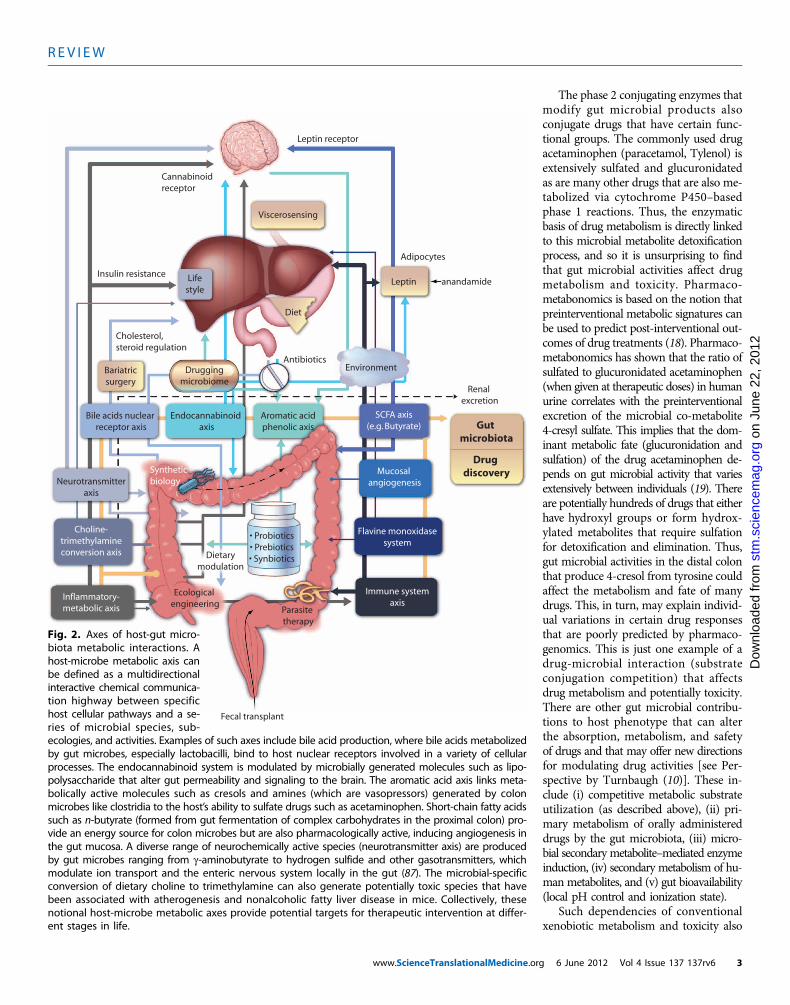
Fig. 2. Axes of host-gut micro-biota metabolic interactions. Ahost-microbe metabolic axis canbe defined as a multidirectionalinteractive chemical communica-tion highway between specifichost cellular pathways and a se-ries of microbial species, sub-
cologies, and activities. Examples of such axes include bile acid production, where bile acids metabolizedy gut microbes, especially lactobacilli, bind to host nuclear receptors involved in a variety of cellularrocesses. The endocannabinoid system is modulated by microbially generated molecules such as lipo-olysaccharide that alter gut permeability and signaling to the brain. The aromatic acid axis links meta-olically active molecules such as cresols and amines (which are vasopressors) generated by colonicrobes like clostridia to the host’s ability to sulfate drugs such as acetaminophen. Short-chain fatty acidsuch as n-butyrate (formed from gut fermentation of complex carbohydrates in the proximal colon) pro-ide an energy source for colon microbes but are also pharmacologically active, inducing angiogenesis ine gut mucosa. A diverse range of neurochemically active species (neurotransmitter axis) are producedy gut microbes ranging from g-aminobutyrate to hydrogen sulfide and other gasotransmitters, whichodulate ion transport and the enteric nervous system locally in the gut (87). The microbial-specificonversion of dietary choline to trimethylamine can also generate potentially toxic species that haveeen associated with atherogenesis and nonalcoholic fatty liver disease in mice. Collectively, theseotional host-microbe metabolic axes provide potential targets for therapeutic intervention at differ-nt stages in life.g 6 June 2012 Vol 4 Issue 137 137rv6 3

REV I EW
on J
une
22, 2
012
lead to the possibility of altering function or activities of the gut mi-crobiota deliberately to affect changes in drug activity to improve pa-tient safety. An excellent example is the deliberate modulation of gutmicrobiota activity to ameliorate toxicity of the anticancer drug CPT-11 that has severe gastrointestinal (GI) side effects. The drug itself isabsorbed from the gut and then partially detoxified via phase 2conjugation in the liver, forming a glucuronide conjugate. The glucu-ronide conjugate is then secreted into the gut in bile where the drugCPT-11 is regenerated due to the action of the b-glucuronidase en-zymes of gut commensal bacteria resulting in GI toxicity due to thefree drug. Wallace et al. (20) showed in mice that selective microbialb-glucuronidase inhibitors when given at the same time as CPT-11prevented deconjugation, hence minimizing the GI toxicity of thedrug. This study shows that designing drugs to modulate microbiotaactivities to reduce the side effects of therapeutics should be possible inpatients.
Host genes in concert with the gut microbiota, diet, and envi-ronmental stressors determine the metabolic phenotypes of indivi-duals (21, 22). There are specific axes of interaction that coupleparticular microbial activities to particular host pathways acting prin-cipally in the gut, liver, and brain (1). These axes are highly metabol-ically interconnected, vary in relative activity throughout the humanlife span as well as in response to diet and life-style, and provide spe-cific targets for therapeutic interventions (Fig. 2).
st
m.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om
PROBIOTIC AND PREBIOTIC INTERVENTIONS
Probiotics are defined as “live microorganisms that, when ad-ministered in adequate amounts, confer a health benefit on the host”(Food and Agriculture Organization of the United Nations, 2002).Probiotics must be safe, that is, meet the U.S. Food and Drug Admin-istration standard, should be amenable to industrial processes neces-sary for commercial production, must remain viable in the foodproduct and during storage, must persist in the GI tract long enoughto elicit an effect, and must improve host health (23–26). Consumersand healthcare workers are starting to appreciate the clinical impor-tance of our resident gut microbiota and the part that prebiotic andprobiotic microbial species can play. Abnormal gut microbial activityhas been implicated in a wide variety of diseases from inflammatorybowel disease to atherosclerosis, and manipulation of the microbiota isone approach to improve health and to treat disease (1, 3). Scientificresearch has focused on how to use probiotic bacterial species such asbifidobacteria and lactobacilli to alter the composition of the indige-nous gut microbiota (23). These probiotic bacterial species help toprotect the host by directly inhibiting the growth of harmful bacteria,and also benefit host health by reducing cholesterol levels, sustainingimmune responses, and synthesizing vitamins such as folate and B12(23–26).
Probiotics have been knowingly ingested by humans for hundredsof years in the form of live yogurt. In contrast, prebiotics—defined as“nondigestible food ingredients that are selectively metabolized by co-lonic bacteria that have the capacity to improve health”—were devel-oped only in the mid-1990s (3). Prebiotics are dietary ingredients suchas nondigestible oligosaccharides that can selectively enhance the ac-tivity of beneficial indigenous gut microbes such as lactobacilli andbifidobacteria. Unlike probiotics, prebiotics can be added to foodproducts that will be heated or cooked without destroying their effects.
www.S
Thus, prebiotic use is directed toward favoring beneficial changeswithin the indigenous gut microbial milieu. Prebiotics are distinctfrom most dietary fibers such as pectin, celluloses, and xylan, whichare not selectively metabolized in the gut. Criteria for classification as aprebiotic are as follows: (i) must resist gastric acidity, hydrolysis bymammalian enzymes, and GI absorption; (ii) must be readily fermentedby the gut microbiota; and (iii) must selectively stimulate the growthand activity of gut microbes associated with health (27). Any dietarycomponent that reaches the colon intact is a potential prebiotic; how-ever, it is the third criterion that is the most difficult to fulfill.
It is only recently that we have been able to sequence and annotatethe complete genomes of many gut microbes in any depth (28). Al-though the broad taxonomic classification of “good bacteria” has beenknown for some time (the restricted palette of probiotics has beendrawn from this), we have not established criteria for what is “normal”in man, let alone what is healthy in man. With increasing premorbid-ity due to obesity and insulin resistance, what is considered a normalgut microbiota may not necessarily be healthy for many people. Froma chemical perspective, much of the interest in the development ofprebiotics is aimed at nondigestible oligosaccharides such asfructooligosaccharides, trans-galactooligosaccharides, lactulose,isomaltooligosaccharides, xylooligosaccharides, soyoligosac-charides, and lactosucrose. In Europe, several human volunteertrials have succeeded in defining fructooligosaccharides, trans-galactooligosaccharides, and lactulose as prebiotics as evidencedby their ability to change the gut microbiota composition after ashort feeding period (29).
Both probiotics and prebiotics have been investigated for their abil-ity to alter the gut microbiota in a manner that improves humanhealth. Gastroenteritis, inflammatory conditions, atopic reactions,and intestinal cancers (30) are among the diseases that have been tar-geted (Fig. 1). Recent studies in mice and human twin pairs fed pro-biotic yogurt containing Lactobacillus rhamnosus and Lactobacillusparacasei have shown that although probiotic microbial species haveonly a slight impact on the gut microbiota composition (unsurprisinggiven the size of the indigenous microbial population), they can up-regulate expression of host genes involved in carbohydrate metabolism(31). Also, direct measurement of the metabolic effects of probioticsand prebiotics in germ-free mice seeded with the gut microbiota of ahuman baby (4, 6) indicates that both probiotics and prebiotics induceshifts in urinary and plasma metabolite profiles. These gnotobioticmice also showed markedly altered plasma lipid and lipoprotein pro-files measured by nuclear magnetic resonance (NMR) spectroscopyand multiple metabolic effects in different tissue compartments(4, 6, 32). Such systemic effects suggest that a metabolic amplificationof effect is occurring, with the probiotics and prebiotics changingmicrobe-host signaling properties. This may well be the case given themismatch between the minor effects of prebiotics and probiotics onspecies composition and their much larger observed effects on hostmetabolism. Furthermore, different probiotics and prebiotics and theircombinations may not be equivalent because they cause differentmetabolic effects in the host, suggesting that they are modulating dif-ferent signaling pathways (4, 6). Even in microbiologically and genet-ically homogeneous mouse models, a diverse response exists to thesame prebiotic-probiotic combination, suggesting that one size maynot fit all. Detailed functional assessments of probiotic and prebioticinterventions and the identification of specific microbial-metabolicconnections (33) will provide new multivariate metrics (in terms of
cienceTranslationalMedicine.org 6 June 2012 Vol 4 Issue 137 137rv6 4

REV I EW
shifts toward healthy host metabolic phenotypes) that will facilitate therational design of dietary interventions that are more finely targeted interms of their health attributes.
on
June
22,
201
2st
m.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om
THE HOST AND ITS MICROBIOTA DURING AGING
With a global impact on GI tract physiology, aging together with anassociated decline in immune system function and increase in chronicinflammation affects gut microbiota homeostasis, leading to changesin intestinal function and microbiota composition. Changes in thecomposition of the symbiotic microbiota with age may result inaltered fermentation (carbohydrate breakdown) and putrefaction(protein breakdown) in the colon and a greater susceptibility to dis-ease. Disease- or age-related physiological changes in the GI tract, aswell as modifications in life-style, diet, and immune system function,inevitably alter the metabolic pathways and small molecules involvedin host-microbiota co-metabolism in the gut (1). There is now interestin tailoring probiotic and prebiotic preparations, diets, drugs, and evenmicrobial engineering to populations of different ages and health sta-tus. The gut microbiota continues to change over life from infancy toold age (Fig. 1). During the first year of life, the infant’s gut ecosystemis colonized principally by opportunistic microorganisms to which thebaby is exposed (34). Aerobes such as staphylococci, streptococci, andEscherichia coli are usually the earliest colonizers followed withindays to weeks by strict anaerobes such as clostridia, bifidobacteria,ruminococci, and bacteroides. The microbiota of breast-fed infants islargely dominated by Bifidobacterium spp. An adult-like gut microbialprofile with a remarkable diversity develops gradually after weaning,driven by a solid diet and development of the gut mucosa itself.
It has been suggested that administration of probiotics, prebiotics,or synbiotics (a mixture of prebiotics and one or more probioticstrains) could counteract age-related gut microbial changes in humans(35–37). A clinical trial with 6-week-old healthy, full-term infants (38)showed that addition of a probiotic Bifidobacterium animalis sub-species lactis to infant formula increased intestinal immunity and re-duced inflammation of the gut mucosa. Improved immune responses,such as increased production of immunoglobulin A antibodies againstrotavirus and poliovirus after immunization, have been reported inCesarean-delivered, non–breast-fed infants. Prebiotic supplementationwith galactooligosaccharides and fructooligosaccharides (39) was welltolerated by formula-fed infants and showed beneficial effects includ-ing an increased abundance and proportion of bifidobacteria and re-duced pH of feces.
Autistic children have an unusual gut microbial composition(40, 41) with a higher abundance of clostridia and altered urinarymetabolic profiles with increases in phenylacetylglutamine, hippurate,and 4-cresyl sulfate compared to unaffected siblings or healthy chil-dren (42). 4-Cresyl sulfate appears to be elevated in the urine of se-verely autistic children (43), which may partly explain the poor abilityof these children to sulfate drugs such as paracetamol (acetamino-phen) (44), although the biological implications of these findings re-main to be elucidated.
In the elderly, the microbiota exhibits a lower microbial diversityand a lower abundance of bifidobacteria, an increase in opportunisticenvironmental facultative aerobes such as Staphylococcus, Streptococ-cus, and Enterobacteriaceae, and an increase in anaerobes such asClostridium groups and Bacteroides spp. (35). Increased antibiotic use
www.S
in older people results in an altered gut microbiota, although the meta-bolic implications of this are ill-defined (36). A recent study showedthat the fecal microbiota of the majority of an elderly cohort (>65 years)is dominated by a greater proportion of Bacteroides spp. and Clostridiumgroups compared to that of younger adults. A cohort of centenarians(>100 years old) in Northern Italy who were homogeneous regardinglife-style and diet showed a gut microbial signature very different fromthat of their younger (<70 years old) counterparts. The gut microbiotaof these centenarians is compromised by an altered Firmicutes pop-ulation and by enrichment of facultative anaerobes, notably pathogenicorganisms associated with increased inflammation. There was morethan a 10-fold increase in the anaerobe Eubacterium limosum and its rel-atives in the centenarians compared to that in the younger cohort (37).E. limosum is seen as a beneficial species because it generates acetate,which can be used by the host as an energy source. Supplementationof the diet of an elderly cohort with probiotic Bifidobacterium strainsincreased the numbers of these health-promoting bacteria in the colon(45). Ingestion of prebiotic trans-galactooligosaccharides, as well assynbiotic preparations, boosted the number of bifidobacteria in elderlyindividuals (46).
DISRUPTED MICROBE-IMMUNE INTERACTIONS,METABOLIC DISEASE, AND POTENTIAL TARGETS
A major challenge for effectively modulating the gut microbiota is theneed for new host and microbial markers and mediators of nutritionalstatus that characterize the integrity of the gut mucosal barrier andmucosal immunity. We are beginning to understand how hostsignaling pathways are regulated by microbial-host co-metabolitesand how these metabolites, which shuttle information between eu-karyotic and prokaryotic cells, modulate adaptive and innate immu-nity in the host. The worldwide epidemic of diabetes, obesity, andmetabolic disease has been linked to disruption of both the innate(47) and the adaptive (48–50) immune systems. It has been suggestedthat excessive production of the cytokines tumor necrosis factor–a,interleukin-6 (IL-6), and IL-1 by immune cells may contribute to in-sulin resistance and hyperglycemia, eventually resulting in diabetes(51–53). Deletion of genes encoding cytokines (53) and chemokines(54) or their corresponding receptors (55), or immune modulationthrough anti–T lymphocyte antibodies (50), could be used to attenuatethe impact of a fat-enriched diet on the development of metabolic dis-ease. In mice fed a high-fat diet, CD8+ effector T cells have been re-ported to infiltrate adipose tissue with a decrease in CD4+ helper T(TH) cells and regulatory T (Treg) cells (56). This study also showedthrough loss- and gain-of-function experiments and adoptive T celltransfer that Treg cells modulate the inflammatory state of adiposetissue, prompting the question of the origin of the antigens stimulatingthis immune response (56). The recent discovery that the phylotypicecology of the intestinal microbiota is markedly altered in response toa fat-enriched diet in rodents (57) and in obese individuals (58) or inpatients with type 2 diabetes (59) may help to answer this question.The marked improvement in metabolic characteristics in humans af-ter Roux-en-Y gastric bypass (bariatric) surgery used to treat obesity(or, as is increasingly the case, type 2 diabetes) has been associatedwith major and stable changes in the gut microbiota with increasesin g-Proteobacteria (60). Similar observations have been made in ro-dents after bariatric surgery and microbial metabolic analysis using
cienceTranslationalMedicine.org 6 June 2012 Vol 4 Issue 137 137rv6 5

REV I EW
on
June
22,
201
2st
m.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om
454 pyrosequencing and high-resolution NMR spectroscopic andmass spectrometric measurements of urine and plasma metabolites(61). These studies also revealed overgrowth of g-Proteobacteria especiallyEnterobacter hormaechei and extensive disruption of microbiota-hostmetabolic axes exemplified by marked shifts in bile acid metabolismafter Roux-en-Y gastric bypass surgery (61). A variety of other micro-bially generated compounds are altered after bariatric surgery includ-ing increases in 4-cresol, several vasoactive amines, g-aminobutyrate,acetate, and propionate (from fermentation). The microbial productsmodulated by bariatric surgery represent changes in nearly all of themetabolic axes shown in Fig. 2, suggesting that the microbiota is anessential part of the “gearbox” that connects the physical effects of bar-iatric surgery to the resulting beneficial effects, although determinationof the exact role of these pathways awaits further study in man.
The microbe antigenic repertoire can help to shape the immunesystem (62) and adapt the intestinal mucosa to the arrival of new com-mensals and symbionts. Bacterial antigens such as lipopolysaccharidestrigger the innate immune system through Toll-like receptors such asTLR4 (63). Female mice lacking TLR2 were protected from the ad-verse effects of a high-fat diet. They showed greater glucose tolerance,insulin sensitivity, and insulin secretion after 20 weeks on a high-fatdiet compared to wild-type mice fed the same diet (64). In the case ofTLR5, this receptor may protect against metabolic syndrome (that is,the opposite effect to TLR2 and TLR4) because mice genetically en-gineered to lack TLR5 exhibit hyperphagia and develop hallmarkfeatures of inflammation-mediated metabolic syndrome (65). Themetabolic phenotype is also “transmissible” because transplantationof the microbiota from TLR5-deficient mice to germ-free wild-typemice resulted in obesity and reduced insulin sensitivity in the recipientanimals. These findings also demonstrate that modulation of the im-mune systemmay affect hostmetabolism by altering the gutmicrobiota(Fig. 2) (1). The impact of the host immune system on the gut micro-biota has been shown in mice lacking Nod-like receptors NLRP3 andNLRP6, which control activation of the inflammasome (66, 67). Bacte-rial DNA fragments have been detected in the blood of obese mice, andit has been suggested that they could be used to predict diabetes (68, 69).In contrast to the increased ratio of Firmicutes toBacteroidetes observedin the intestine of obese mice (58), the blood of obese mice has DNAfragments of Proteobacteria (68), facultative anaerobes that can survivein the presence of oxygen. How these bacterial DNA fragments reachthe blood is unclear, although loosening of the tight junctions betweengut epithelial cells (57, 70) as well as colocalization of bacteria with in-testinal dendritic cells (69) have been reported. A reduction in mRNAsencoding zonula occludens-1 and occludin proteins that compose tightjunctions has been associated withmetabolic disease induced by a high-fat diet (57, 71). This mechanism correlates with changes to the gutmicrobiota independent of the genetic background of the mice or thediet they are fed (70). Antibiotic treatments (72), as well as prebioticsimprove intestinal permeability in mice through a mechanism that re-quires the secretion of glucagon-like peptide 2 (71).On the other hand, atranscellular mechanismmay be responsible for the continuous surveil-lance of changes in the gut microbiota that could alter immune systemfunction through changes in the intestinal antigenic repertoire. In obesityand type 2 diabetes, initial evidence suggests thatmucosal dysbiosis in theileum (69) could disrupt immune tolerance (73).
Dendritic cells expressing CX3CR1 that are able to access the gutlumen may be able to clear bacteria. In metabolic disease, the clear-ance of intestinal bacteria from the mucosa may be incomplete be-
www.S
cause bacteria have been reported to reach the mesenteric lymphnodes that are connected to adipose tissue (69). This mechanism in-volves NOD1 (the nucleotide-binding oligomerization-containingdomain protein 1), which promotes metabolic disease induced by ahigh-fat diet, but does not involve NOD2, which conversely protectsagainst diabetes and obesity (74). Such bacterial translocation gener-ates a state of “metabolic infection,” as also shown in streptozotocin-induced diabetic mice where bacteria that normally reside in the gut,lung, and skin were reported in the liver, spleen, kidneys, and mesen-teric lymph nodes (75). The notion is emerging that these mislocalizedbacteria might induce inflammation in, for example, adipose tissue,resulting in an influx of cells from the adaptive immune system. Thiswould create a complex milieu containing adipokines, chemokines, andtheir receptors (MCP-1, CCL7, CCL8/CCR2, CXCL14/CXCR2) andother molecules such as C3, factor B, adipsin, and angiopoietin-likeprotein 2 (76). Translocation of bacteria from intestinal or oral originhas also been described in atherosclerotic plaques where Proteobacteriarather than Firmicutes predominate (77). These bacteria belong to ourcommensal microbiota, which has “educated” the immune system frombirth and is thus seen by the immune system as “self.” A change in theintestinal microbiota repertoire induced by recent changes in Westernnutrition would constitute a rupture in this immune tolerance that hasevolved over millennia, potentially leading to an increase in inflam-mation and metabolic disease. One way to prevent and treat metabolicdisease may be to restore the microbiota-host immune relationship [seeReview by Blumberg and Powrie (78)].
ENGINEERING THE MICROBIOTA: A CHALLENGE FORTHE FUTURE
Engineering the microbiota to favor human health is a formidablechallenge that will require exquisite knowledge of co-metabolic regu-lation and ecological properties of the gut microbiota. For instance,engineering bacteria has shown potential for altering microbial meta-bolic pathways to enhance biofuel production in fermentationsystems. The same strategy could in principle be used to correct ab-normalities in metabolic or signaling pathways involved in diseasepathogenesis. Nicaise et al. have shown that a metabolic condition,hyperammonemia, which occurs constitutively or due to liver failure,can be treated successfully in mice by administration of Lactobacillusplantarum (79). A wild-type L. plantarum strain and two strains thatwere genetically engineered to consume ammonia or to lack the am-monia transporter were used in this study. The ammonia-consumingstrain lowered blood and fecal ammonia with improved survival of themice with hyperammonemia compared with the wild-type L. plantarumstrain. Similarly, Jones et al. (80) demonstrated that L. plantarum 80(pCBH1)–overexpressing bile salt hydrolase, when given to mice in amicroencapsulated form, efficiently degraded and removed glyco-deoxycholic acid and taurodeoxycholic acid, suggesting that it mayprove useful for lowering blood cholesterol. There are likely to bemore applications for engineered intestinal microbial strains includingthe direct consumption of toxic chemicals in the gut. The advantage ofthese interventions over other gut microbiota modulation strategies isthat they can be directed specifically at a toxic molecule or metabolicpathway rather than the entire microbiota. However, even subtlechanges in the microbiota and bacterial physiology can exert markedchanges in its ability to modulate the host immune system. A mechanistic
cienceTranslationalMedicine.org 6 June 2012 Vol 4 Issue 137 137rv6 6

REV I EW
on
June
22,
201
2st
m.s
cien
cem
ag.o
rg
understanding of such host-microbiota interactions can potentiallylead to new interventions through synthetic biology. For example, agenetically engineered defect in techoic acid biosynthesis in L. plantarumconverted a proinflammatory response in mice to the wild-type microbeto an anti-inflammatory response including increased IL-10 produc-tion (81). Yan et al. have described how protein p40 derived fromL. rhamnosus reduced epithelial cell apoptosis and suppressed inflam-mation in mouse models of inflammatory bowel disease by activatingthe receptor for epithelial growth factor (82). These findings exemplifyhow an understanding of the molecular basis of these microbiota-hostinteractions could lead to the development of new therapeutic strategiesfor treating a variety of disorders.
The involvement of the gut microbiota in diverse aspects of diseasepresents a multitude of opportunities for intervention (Fig. 1) (for ex-ample, treating obesity and metabolic syndrome with bariatric surgeryor modulating the gut microbiota by altering macronutrients in thediet or by administering prebiotics and probiotics) (83). For other dis-eases, more radical means of altering the gut microbiota have beenattempted including fecal transplantation from healthy donors (84)to treat infection with antibiotic-resistant Clostridium difficile (84).Other strategies include direct ecological manipulation of the gut mi-crobiota using microbe-microbe interactions through introduction ofsimilar but nonpathogenic bacterial strains and the use of helminths totreat asthma, Crohn’s disease, and ulcerative colitis (85). Treatmentwith helminth parasites such as Trichuris trichiura, Trichuris suis, orNecator americanus works on the principle that the parasite induces aTH2 cell–based immunoregulatory response that dampens the TH1 re-sponse in the host with a reduction in inflammation and improved gutmucosal barrier function. Animal models have shown marked pertur-bation of microbial metabolites after infection by parasites, indicatinga delicate equilibrium between the host, its resident microbiota, andacquired parasites (86), further emphasizing the metabolicallycomplex nature of the mammalian symbiotic supraorganism.
Dow
nloa
ded
from
FUTURE CHALLENGES
Understanding and then deliberately modulating microbiota-hostmetabolic and immune interactions to improve human health is oneof the greatest challenges of 21st century medicine. In addition to thetechnical complexity of the field, which involves state-of-the art deep ge-nome sequencing, advanced chemical technologies and bioinformatics,and computational integration of astonishing amounts of supraorganismsystems data, there is the discovery process of finding potential interven-tional or therapeutic targets, axes, and pathways within all of thiscomplexity. Advances in systems and computational biology will surelyenable us to measure and model many aspects of the dynamic inter-actions between the human host and its resident gut microbiota. Thebenefits of harnessing this knowledge for disease management and themodification of disease risk have great potential andwill help to underpinfuture developments in personalized healthcare and stratified medicineand ultimately in determining public health priorities.
REFERENCES AND NOTES1. J. K. Nicholson, E. Holmes, J. Kinross, R. Burcelin, G. Gibson, W. Jia, S. Pettersson, Host-gut
microbiota metabolic interactions. Science 336, 1262–1267 (2012).2. W. Jia, H. Li, L. Zhao, J. K. Nicholson, Gut microbiota: A potential new territory for drug
targeting. Nat. Rev. Drug Discov. 7, 123–129 (2008).
www.S
3. G. R. Gibson, M. B. Roberfroid, Dietary modulation of the human colonic microbiota: In-troducing the concept of prebiotics. J. Nutr. 125, 1401–1412 (1995).
4. F. P. Martin, Y. Wang, N. Sprenger, I. K. Yap, S. Rezzi, Z. Ramadan, E. Peré-Trepat, F. Rochat,C. Cherbut, P. van Bladeren, L. B. Fay, S. Kochhar, J. C. Lindon, E. Holmes, J. K. Nicholson,Top-down systems biology integration of conditional prebiotic modulated transgenomicinteractions in a humanized microbiome mouse model. Mol. Syst. Biol. 4, 205 (2008).
5. A. W. Walker, J. Ince, S. H. Duncan, L. M. Webster, G. Holtrop, X. Ze, D. Brown, M. D. Stares,P. Scott, A. Bergerat, P. Louis, F. McIntosh, A. M. Johnstone, G. E. Lobley, J. Parkhill, H. J. Flint,Dominant and diet-responsive groups of bacteria within the human colonic microbiota.ISME J. 5, 220–230 (2011).
6. F. P. Martin, Y. Wang, N. Sprenger, I. K. Yap, T. Lundstedt, P. Lek, S. Rezzi, Z. Ramadan,P. van Bladeren, L. B. Fay, S. Kochhar, J. C. Lindon, E. Holmes, J. K. Nicholson, Probiotic mod-ulation of symbiotic gut microbial-host metabolic interactions in a humanized microbiomemouse model. Mol. Syst. Biol. 4, 157 (2008).
7. X. Zheng, G. Xie, A. Zhao, L. Zhao, C. Yao, N. H. Chiu, Z. Zhou, Y. Bao, W. Jia, J. K. Nicholson,W. Jia, The footprints of gut microbial-mammalian co-metabolism. J. Proteome Res. 10,5512–5522 (2011).
8. J. R. Swann, E. J. Want, F. M. Geier, K. Spagou, I. D. Wilson, J. E. Sidaway, J. K. Nicholson,E. Holmes, Systemic gut microbial modulation of bile acid metabolism in host tissuecompartments. Proc. Natl. Acad. Sci. U.S.A. 108 (Suppl. 1), 4523–4530 (2011).
9. K. P. Lemon, G. C. Armitage, D. A. Relman, M. A. Fischbach, Microbiota-targeted therapies:An ecological perspective. Sci. Transl. Med. 4, 137rv5 (2012).
10. H. J. Haiser, P. J. Turnbaugh, Is it time for a metagenomic basis for therapeutics. Science336, 1253–1255 (2012).
11. J. K. Nicholson, E. Holmes, I. D. Wilson, Gut microorganisms, mammalian metabolism andpersonalized health care. Nat. Rev. Microbiol. 3, 431–438 (2005).
12. A. M. O’Hara, F. Shanahan, Gut microbiota: Mining for therapeutic potential. Clin. Gastroenterol.Hepatol. 5, 274–284 (2007).
13. R. Mirnezami, J. Nicholson, A. Darzi, Preparing for precision medicine. N. Engl. J. Med. 366,489–491 (2012).
14. J. K. Nicholson, I. D. Wilson, Opinion: Understanding ‘global’ systems biology: metabonomicsand the continuum of metabolism. Nat. Rev. Drug Discov. 2, 668–676 (2003).
15. J. L. Banker, E. G. Schmidt, Urinary phenols; effect of diet on the phenolic bodies of theurine. J. Biol. Chem. 165, 427–430 (1946).
16. A. Wijeyesekera, P. A. Clarke, M. Bictash, I. J. Brown, M. Fidock, T. Ryckmans, I. K. S. Yap, Q. Chan,J. Stamler, P. Elliott, E. Holmes, J. K. Nicholson, Quantitative UPLC-MS/MS analysis of the gutmicrobial co-metabolites phenylacetylglutamine, 4-cresyl sulphate and hippurate in humanurine: INTERMAP Study. Anal. Methods 4, 65–72 (2012).
17. D. Beyoğlu, R. L. Smith, J. R. Idle, Dog bites man or man bites dog? The enigma of theamino acid conjugations. Biochem. Pharmacol. 83, 1331–1339 (2012).
18. T. A. Clayton, J. C. Lindon, O. Cloarec, H. Antti, C. Charuel, G. Hanton, J. P. Provost, J. L. Le Net,D. Baker, R. J. Walley, J. R. Everett, J. K. Nicholson, Pharmaco-metabonomic phenotyping andpersonalized drug treatment. Nature 440, 1073–1077 (2006).
19. T. A. Clayton, D. Baker, J. C. Lindon, J. R. Everett, J. K. Nicholson, Pharmacometabonomicidentification of a significant host-microbiome metabolic interaction affecting humandrug metabolism. Proc. Natl. Acad. Sci. U.S.A. 106, 14728–14733 (2009).
20. B. D. Wallace, H. Wang, K. T. Lane, J. E. Scott, J. Orans, J. S. Koo, M. Venkatesh, C. Jobin, L. A. Yeh,S. Mani, M. R. Redinbo, Alleviating cancer drug toxicity by inhibiting a bacterial enzyme.Science 330, 831–835 (2010).
21. J. K. Nicholson, Global systems biology, personalized medicine and molecular epidemiol-ogy. Mol. Syst. Biol. 2, 52 (2006).
22. E. Holmes, I. D. Wilson, J. K. Nicholson, Metabolic phenotyping in health and disease. Cell134, 714–717 (2008).
23. G. R. Gibson, C. M. Williams, Eds., Functional Foods: Concept to Product (WoodheadPublishing, Cambridge, 2000).
24. T. Steer, H. Carpenter, K. Tuohy, G. R. Gibson, Perspectives on the role of the human gutmicrobiota and its modulation by pro- and prebiotics. Nutr. Res. Rev. 13, 229–254 (2000).
25. S. Kolida, D. Meyer, G. R. Gibson, A double-blind placebo-controlled study to establish thebifidogenic dose of inulin in healthy humans. Eur. J. Clin. Nutr. 61, 1189–1195 (2007).
26. S. Khani, H. M. Hosseini, M. Taheri, M. R. Nourani, A. A. Imani Fooladi, Probiotics as analternative strategy for prevention and treatment of human diseases: A review. Inflamm.Allergy Drug Targets 11, 79–89 (2012).
27. G. R. Gibson, K. P. Scott, R. A. Rastall, K. M. Tuohy, A. Hotchkiss, A. Dubert-Ferrandon, M. Gareau,E. F. Murphy, D. Saulnier, G. Loh, S. Macfarlane, N. Delzenne, Y. Ringel, G. Kozianowski,R. Dickmann, I. Lenoir-Wijnkoop, C. Walker, R. Buddington, Dietary prebiotics: Currentstatus and new definition. IFIS Funct. Foods Bull. 7, 1–19 (2011).
28. J. Qin, R. Li, J. Raes, M. Arumugam, K. S. Burgdorf, C. Manichanh, T. Nielsen, N. Pons, F. Levenez,T. Yamada, D. R. Mende, J. Li, J. Xu, S. Li, D. Li, J. Cao, B. Wang, H. Liang, H. Zheng, Y. Xie, J. Tap,P. Lepage, M. Bertalan, J. M. Batto, T. Hansen, D. Le Paslier, A. Linneberg, H. B. Nielsen,E. Pelletier, P. Renault, T. Sicheritz-Ponten, K. Turner, H. Zhu, C. Yu, S. Li, M. Jian, Y. Zhou, Y. Li,X. Zhang, S. Li, N. Qin, H. Yang, J. Wang, S. Brunak, J. Doré, F. Guarner, K. Kristiansen,
cienceTranslationalMedicine.org 6 June 2012 Vol 4 Issue 137 137rv6 7

REV I EW
on
June
22,
201
2st
m.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om
O. Pedersen, J. Parkhill, J. Weissenbach; MetaHIT Consortium, P. Bork, S. D. Ehrlich, J. Wang,A human gut microbial gene catalogue established by metagenomic sequencing. Nature464, 59–65 (2010).
29. G. R. Gibson, M. B. Roberfroid, Eds., A Handbook of Prebiotics (Taylor & Francis, Boca Raton,FL, 2008).
30. J. M. Kinross, A. W. Darzi, J. K. Nicholson, Gut microbiome-host interactions in health anddisease. Genome Med. 3, 14 (2011).
31. N. McNulty, T. Yatsunenko, A. Hsiao, J. J. Faith, B. D. Muegge, A. L. Goodman, B. Henrissat,R. Oozeer, S. Cools-Portier, G. Gobert, C. Chervaux, D. Knights, C. A. Lozupone, R. Knight,A. E. Duncan, J. R. Bain, M. J. Muehlbauer, C. B. Newgard, A. C. Heath, J. I. Gordon, Theimpact of a consortium of fermented milk strains on the gut microbiome of gnotobioticmice and monozygotic twins. Sci. Transl. Med. 3, 106ra106 (2011).
32. S. P. Claus, T. M. Tsang, Y. Wang, O. Cloarec, E. Skordi, F. P. Martin, S. Rezzi, A. Ross, S. Kochhar,E. Holmes, J. K. Nicholson, Systemic multicompartmental effects of the gut microbiome onmouse metabolic phenotypes. Mol. Syst. Biol. 4, 219 (2008).
33. J. K. Nicholson, J. Connelly, J. C. Lindon, E. Holmes, Metabonomics: A platform for studyingdrug toxicity and gene function. Nat. Rev. Drug Discov. 1, 153–161 (2002).
34. C. Palmer, E. M. Bik, D. B. DiGiulio, D. A. Relman, P. O. Brown, Development of the humaninfant intestinal microbiota. PLoS Biol. 5, e177 (2007).
35. E. Biagi, M. Candela, S. Fairweather-Tait, C. Franceschi, P. Brigidi, Ageing of the humanmetaorganism: The microbial counterpart. Age 34, 247–267 (2012).
36. K. Tiihonen, A. C. Ouwehand, N. Rautonen, Human intestinal microbiota and healthyageing. Ageing Res. Rev. 9, 107–116 (2010).
37. E. Biagi, L. Nylund, M. Candela, R. Ostan, L. Bucci, E. Pini, J. Nikkila, D. Monti, R. Satokari,C. Franceschi, P. Brigidi, W. De Vos, Through ageing, and beyond: Gut microbiota andinflammatory status in seniors and centenarians. PLoS One 5, e10667 (2010).
38. H. D. Holscher, L. A. Czerkies, P. Cekola, R. Litov, M. Benbow, S. Santema, D. D. Alexander,V. Perez, S. Sun, J. M. Saavedra, K. A. Tappenden, Bifidobacterium lactis Bb12 enhancesintestinal antibody response in formula-fed infants: A randomized, double-blind,controlled trial. JPEN J. Parenter. Enteral. Nutr. 36 (Suppl. 1), 106S–117S (2012).
39. H. D. Holscher, K. L. Faust, L. A. Czerkies, R. Litov, E. E. Ziegler, H. Lessin, T. Hatch, S. Sun,K. A. Tappenden, Effects of prebiotic-containing infant formula on gastrointestinal toleranceand fecal microbiota in a randomized controlled trial. JPEN J. Parenter. Enteral. Nutr. 36 (Suppl. 1),95S–105S (2012).
40. Y. Song, C. Liu, S. M. Finegold, Real-time PCR quantitation of clostridia in feces of autisticchildren. Appl. Environ. Microbiol. 70, 6459–6465 (2004).
41. S. M. Finegold, J. Downes, P. H. Summanen, Microbiology of regressive autism. Anaerobe18, 260–262 (2012).
42. I. K. Yap, M. Angley, K. A. Veselkov, E. Holmes, J. C. Lindon, J. K. Nicholson, Urinary meta-bolic phenotyping differentiates children with autism from their unaffected siblings andage-matched controls. J. Proteome Res. 9, 2996–3004 (2010).
43. L. Altieri, C. Neri, R. Sacco, P. Curatolo, A. Benvenuto, F. Muratori, E. Santocchi, C. Bravaccio,C. Lenti, M. Saccani, R. Rigardetto, M. Gandione, A. Urbani, A. M. Persico, Urinary p-cresolis elevated in small children with severe autism spectrum disorder. Biomarkers 16,252–260 (2011).
44. A. Alberti, P. Pirrone, M. Elia, R. H. Waring, C. Romano, Sulphation deficit in “low-functioning” autistic children: A pilot study. Biol. Psychiatry 46, 420–424 (1999).
45. M. Matsumoto, M. Sakamoto, Y. Benno, Dynamics of fecal microbiota in hospitalized elderlyfed probiotic LKM512 yogurt. Microbiol. Immunol. 53, 421–432 (2009).
46. J. Vulevic, A. Drakoularakou, P. Yaqoob, G. Tzortzis, G. R. Gibson, Modulation of the fecalmicroflora profile and immune function by a novel trans-galactooligosaccharide mixture(B-GOS) in healthy elderly volunteers. Am. J. Clin. Nutr. 88, 1438–1446 (2008).
47. J. C. Pickup, M. A. Crook, Is type II diabetes mellitus a disease of the innate immune sys-tem? Diabetologia 41, 1241–1248 (1998).
48. M. Feuerer, L. Herrero, D. Cipolletta, A. Naaz, J. Wong, A. Nayer, J. Lee, A. B. Goldfine, C. Benoist,S. Shoelson, D. Mathis, Lean, but not obese, fat is enriched for a unique population of reg-ulatory T cells that affect metabolic parameters. Nat. Med. 15, 930–939 (2009).
49. J. Liu, A. Divoux, J. Sun, J. Zhang, K. Clément, J. N. Glickman, G. K. Sukhova, P. J. Wolters, J. Du,C. Z. Gorgun, A. Doria, P. Libby, R. S. Blumberg, B. B. Kahn, G. S. Hotamisligil, G. P. Shi, Geneticdeficiency and pharmacological stabilization of mast cells reduce diet-induced obesity anddiabetes in mice. Nat. Med. 15, 940–945 (2009).
50. S. Nishimura, I. Manabe, M. Nagasaki, K. Eto, H. Yamashita, M. Ohsugi, M. Otsu, K. Hara,K. Ueki, S. Sugiura, K. Yoshimura, T. Kadowaki, R. Nagai, CD8+ effector T cells contributeto macrophage recruitment and adipose tissue inflammation in obesity. Nat. Med. 15,914–920 (2009).
51. G. S. Hotamisligil, Inflammation and metabolic disorders. Nature 444, 860–867 (2006).52. S. E. Shoelson, J. Lee, A. B. Goldfine, Inflammation and insulin resistance. J. Clin. Invest. 116,
1793–1801 (2006).53. G. S. Hotamisligil, P. Peraldi, A. Budavari, R. Ellis, M. F. White, B. M. Spiegelman, IRS-1-
mediated inhibition of insulin receptor tyrosine kinase activity in TNF-a-and obesity-inducedinsulin resistance. Science 271, 665–668 (1996).
www.S
54. H. Kanda, S. Tateya, Y. Tamori, K. Kotani, K. Hiasa, R. Kitazawa, S. Kitazawa, H. Miyachi, S. Maeda,K. Egashira, M. Kasuga, MCP-1 contributes to macrophage infiltration into adipose tissue, in-sulin resistance, and hepatic steatosis in obesity. J. Clin. Invest. 116, 1494–1505 (2006).
55. D. A. Gutierrez, A. X. Kennedy, J. S. Orr, E. K. Anderson, C. D. Webb, W. K. Gerrald, A. H. Hasty,Aberrant accumulation of undifferentiated myeloid cells in the adipose tissue of CCR2-deficient mice delays improvements in insulin sensitivity. Diabetes 60, 2820–2829 (2011).
56. S. Winer, Y. Chan, G. Paltser, D. Truong, H. Tsui, J. Bahrami, R. Dorfman, Y. Wang, J. Zielenski,F. Mastronardi, Y. Maezawa, D. J. Drucker, E. Engleman, D. Winer, H. M. Dosch, Normaliza-tion of obesity-associated insulin resistance through immunotherapy. Nat. Med. 15, 921–929(2009).
57. P. D. Cani, R. Bibiloni, C. Knauf, A. Waget, A. M. Neyrinck, N. M. Delzenne, R. Burcelin,Changes in gut microbiota control metabolic endotoxemia-induced inflammation inhigh-fat diet-induced obesity and diabetes in mice. Diabetes 57, 1470–1481 (2008).
58. R. E. Ley, F. Bäckhed, P. Turnbaugh, C. A. Lozupone, R. D. Knight, J. I. Gordon, Obesity altersgut microbial ecology. Proc. Natl. Acad. Sci. U.S.A. 102, 11070–11075 (2005).
59. X. Wu, C. Ma, L. Han, M. Nawaz, F. Gao, X. Zhang, P. Yu, C. Zhao, L. Li, A. Zhou, J. Wang,J. E. Moore, B. C. Millar, J. Xu, Molecular characterisation of the faecal microbiota inpatients with type II diabetes. Curr. Microbiol. 61, 69–78 (2010).
60. H. Zhang, J. K. DiBaise, A. Zuccolo, D. Kudrna, M. Braidotti, Y. Yu, P. Parameswaran,M. D. Crowell, R. Wing, B. E. Rittmann, R. Krajmalnik-Brown, Human gut microbiota in obesityand after gastric bypass. Proc. Natl. Acad. Sci. U.S.A. 106, 2365–2370 (2009).
61. J. V. Li, H. Ashrafian, M. Bueter, J. Kinross, C. Sands, C. W. le Roux, S. R. Bloom, A. Darzi,T. Athanasiou, J. R. Marchesi, J. K. Nicholson, E. Holmes, Metabolic surgery profoundly influ-ences gut microbial–host metabolic cross-talk. Gut 60, 1214–1223 (2011).
62. A. J. Macpherson, N. L. Harris, Interactions between commensal intestinal bacteria and theimmune system. Nat. Rev. Immunol. 4, 478–485 (2004).
63. P. D. Cani, J. Amar, M. A. Iglesias, M. Poggi, C. Knauf, D. Bastelica, A. M. Neyrinck, F. Fava,K. M. Tuohy, C. Chabo, A. Waget, E. Delmée, B. Cousin, T. Sulpice, B. Chamontin, J. Ferrières,J. F. Tanti, G. R. Gibson, L. Casteilla, N. M. Delzenne, M. C. Alessi, R. Burcelin, Metabolicendotoxemia initiates obesity and insulin resistance. Diabetes 56, 1761–1772 (2007).
64. J. A. Ehses, D. T. Meier, S. Wueest, J. Rytka, S. Boller, P. Y. Wielinga, A. Schraenen, K. Lemaire,S. Debray, L. Van Lommel, J. A. Pospisilik, O. Tschopp, S. M. Schultze, U. Malipiero,H. Esterbauer, H. Ellingsgaard, S. Rütti, F. C. Schuit, T. A. Lutz, M. Böni-Schnetzler,D. Konrad, M. Y. Donath, Toll-like receptor 2-deficient mice are protected from insulin resistanceand b cell dysfunction induced by a high-fat diet. Diabetologia 53, 1795–1806 (2010).
65. F. A. Carvalho, J. D. Aitken, A. T. Gewirtz, M. Vijay-Kumar, TLR5 activation induces secretoryinterleukin-1 receptor antagonist (sIL-1Ra) and reduces inflammasome-associated tissuedamage. Mucosal Immunol. 4, 102–111 (2011).
66. E. Elinav, T. Strowig, A. L. Kau, J. Henao-Mejia, C. A. Thaiss, C. J. Booth, D. R. Peaper, J. Bertin,S. C. Eisenbarth, J. I. Gordon, R. A. Flavell, NLRP6 inflammasome regulates colonic microbialecology and risk for colitis. Cell 145, 745–757 (2011).
67. J. Henao-Mejia, E. Elinav, C. Jin, L. Hao, W. Z. Mehal, T. Strowig, C. A. Thaiss, A. L. Kau,S. C. Eisenbarth, M. J. Jurczak, J. P. Camporez, G. I. Shulman, J. I. Gordon, H. M. Hoffman,R. A. Flavell, Inflammasome-mediated dysbiosis regulates progression of NAFLD andobesity. Nature 482, 179–185 (2012).
68. J. Amar, M. Serino, C. Lange, C. Chabo, J. Iacovoni, S. Mondot, P. Lepage, C. Klopp, J. Mariette,O. Bouchez, L. Perez, M. Courtney, M. Marre, P. Klopp, O. Lantieri, J. Doré, M. A. Charles,B. Balkau, R. Burcelin; D.E.S.I.R. Study Group, Involvement of tissue bacteria in the onsetof diabetes in humans: Evidence for a concept. Diabetologia 54, 3055–3061 (2011).
69. J. Amar, C. Chabo, A. Waget, P. Klopp, C. Vachoux, L. G. Bermúdez-Humarán, N. Smirnova,M. Bergé, T. Sulpice, S. Lahtinen, A. Ouwehand, P. Langella, N. Rautonen, P. J. Sansonetti,R. Burcelin, Intestinal mucosal adherence and translocation of commensal bacteria at theearly onset of type 2 diabetes: Molecular mechanisms and probiotic treatment. EMBOMol. Med. 3, 559–572 (2011).
70. M. Serino, E. Luche, S. Gres, A. Baylac, M. Bergé, C. Cenac, A. Waget, P. Klopp, J. Iacovoni,C. Klopp, J. Mariette, O. Bouchez, J. Lluch, F. Ouarné, P. Monsan, P. Valet, C. Roques, J. Amar,A. Bouloumié, V. Théodorou, R. Burcelin, Metabolic adaptation to a high-fat diet is associatedwith a change in the gut microbiota. Gut 61, 543–553 (2012).
71. P. D. Cani, S. Possemiers, T. Van de Wiele, Y. Guiot, A. Everard, O. Rottier, L. Geurts, D. Naslain,A. Neyrinck, D. M. Lambert, G. G. Muccioli, N. M. Delzenne, Changes in gut microbiota controlinflammation in obese mice through a mechanism involving GLP-2-driven improvement ofgut permeability. Gut 58, 1091–1103 (2009).
72. M. Membrez, F. Blancher, M. Jaquet, R. Bibiloni, P. D. Cani, R. G. Burcelin, I. Corthesy, K. Macé,C. J. Chou, Gut microbiota modulation with norfloxacin and ampicillin enhances glucosetolerance in mice. FASEB J. 22, 2416–2426 (2008).
73. L. Macia, A. N. Thorburn, L. C. Binge, E. Marino, K. E. Rogers, K. M. Maslowski, A. T. Vieira,J. Kranich, C. R. Mackay, Microbial influences on epithelial integrity and immune function asa basis for inflammatory diseases. Immunol. Rev. 245, 164–176 (2012).
74. J. D. Schertzer, A. K. Tamrakar, J. G. Magalhães, S. Pereira, P. J. Bilan, M. D. Fullerton, Z. Liu,G. R. Steinberg, A. Giacca, D. J. Philpott, A. Klip, NOD1 activators link innate immunity toinsulin resistance. Diabetes 60, 2206–2215 (2011).
cienceTranslationalMedicine.org 6 June 2012 Vol 4 Issue 137 137rv6 8

REV I EW
Jun
e 22
, 201
2
75. A. Imai, Y. Kurihara, Endogenous infection in mice with streptozotocin-induced diabetes.A feature of bacterial translocation. Can. J. Microbiol. 30, 1344–1348 (1984).
76. K. Lolmède, C. Duffaut, A. Zakaroff-Girard, A. Bouloumié, Immune cells in adipose tissue:Key players in metabolic disorders. Diabetes Metab. 37, 283–290 (2011).
77. A. R. Aquino, K. C. Lima, M. S. Paiva, I. N. Rôças, J. F. Siqueira Jr., Molecular survey of ath-eromatous plaques for the presence of DNA from periodontal bacterial pathogens, archaeaand fungi. J. Periodontal Res. 46, 303–309 (2011).
78. R. Blumberg, F. Powrie, Microbiota, disease, and back again to health: A metastable jour-ney. Sci. Transl. Med. 4, 137rv7 (2012).
79. C. Nicaise, D. Prozzi, E. Viaene, C. Moreno, T. Gustot, E. Quertinmont, P. Demetter, V. Suain,P. Goffin, J. Devière, P. Hols, Control of acute, chronic, and constitutive hyperammonemiaby wild-type and genetically engineered Lactobacillus plantarum in rodents. Hepatology48, 1184–1192 (2008).
80. M. L. Jones, H. Chen, W. Ouyang, T. Metz, S. Prakash, Microencapsulated genetically en-gineered Lactobacillus plantarum 80 (pCBH1) for bile acid deconjugation and its implica-tion in lowering cholesterol. J. Biomed. Biotechnol. 2004, 61–69 (2004).
81. C. Grangette, S. Nutten, E. Palumbo, S. Morath, C. Hermann, J. Dewulf, B. Pot, T. Hartung,P. Hols, A. Mercenier, Enhanced antiinflammatory capacity of a Lactobacillus plantarummutant synthesizing modified teichoic acids. Proc. Natl. Acad. Sci. U.S.A. 102, 10321–10326 (2005).
82. F. Yan, H. C. Cao, T. Cover, M. K. Washington, Y. Shi, L. S. Liu, R. Chaturvedi, R. M. Peek,K. T. Wilson, D. B. Polk, Colon-specific delivery of a probiotic-derived soluble protein amelio-rates intestinal inflammation in mice through an EGFR-dependent mechanism. J. Clin. Invest.121, 2242–2253 (2011).
83. R. S. Kootte, A. Vrieze, F. Holleman, G. M. Dallinga-Thie, E. G. Zoetendal, W. M. de Vos,A. K. Groen, J. B. Hoekstra, E. S. Stroes, M. Nieuwdorp, The therapeutic potential of manip-ulating gut microbiota in obesity and type 2 diabetes mellitus. Diabetes Obes. Metab. 14,112–120 (2012).
www.S
84. E. Gough, H. Shaikh, A. R. Manges, Systematic review of intestinal microbiota transplanta-tion (fecal bacteriotherapy) for recurrent Clostridium difficile infection. Clin. Infect. Dis. 53,994–1002 (2011).
85. F. Shanahan, Gut microbes: From bugs to drugs. Am. J. Gastroenterol. 105, 275–279 (2010).
86. Y. Wang, J. Utzinger, J. Saric, J. V. Li, J. Burckhardt, S. Dirnhofer, J. K. Nicholson, B. H. Singer,R. Brun, E. Holmes, Global metabolic responses of mice to Trypanosoma brucei brucei in-fection. Proc. Natl. Acad. Sci. U.S.A. 105, 6127–6132 (2008).
87. M. M. Gadalla, S. H. Snyder, Hydrogen sulfide as a gasotransmitter. J. Neurochem. 113, 14–26 (2010).
Acknowledgments: We acknowledge funding from the following: Imperial College Health-care Trust Biomedical Research Centre, Bill and Melinda Gates Foundation, U.K. MedicalResearch Council, The Wellcome Trust, and Fondation Merrieux (J.K.N., E.H., and J.K.); U.K. Bio-technology and Biological Sciences Research Council, Tate + Lyle, Procter and Gamble, Ganeden,Clasado (G.R.G.); Swedish Medical Research Council, COMBINE, EU-project TORNADO, Karolins-ka Institutet Inflammation Consortium, The Millennium Foundation Singapore, The NationalCancer Centre Singapore, and Nanyang Technical University (S.P.); Agence Nationale de laRecherche and European Framework Program FP7 (R.B.); U.S. National Institutes of Healthgrant # R01AA020212 (W.J.).
Submitted 30 April 2012Accepted 17 May 2012Published 6 June 201210.1126/scitranslmed.3004244
Citation: E. Holmes, J. Kinross, G. R. Gibson, R. Burcelin, W. Jia, S. Pettersson, J. K. Nicholson,Therapeutic modulation of microbiota-host metabolic interactions. Sci. Transl. Med. 4, 137rv6(2012).
on
cienceTranslationalMedicine.org 6 June 2012 Vol 4 Issue 137 137rv6 9
st
m.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om
Related Documents











