ELSEVIER FEMS Microbiology Ecology 21 (1996) 243-253 The role of polyglucose in oxygen-dependent respiration by a new strain of Desulfovibrio salexigens Ed W.J. van Niel a3 *, Teresa M. Pedro Gomes a, Anne Willems b, Matthew D. Collins b, Rudolf A. Prins a, Jan C. Gottschal a a Department of Microbiology, Unitiersity of Groningen, Kerklaan 30, 9751 NN Haren, The Netherlands h Institute of Food Research, Reading Laboratory, Earley Gate, Whiteknights Road, Reading RG6 6BZ, UK Received 17 May 1996; revised 5 August 1996: accepted 21 August 1996 Abstract Desulfooibrio salexigens strain Mast1 was isolated from the oxic/anoxic interface of a marine sediment. Growth under sulfate-reducing conditions was accompanied by polyglucose accumulation in the cell with every substrate tested. Highest polyglucose storage was found with glucose (0.8-1.0 g polyglucose (g protein)-‘), but the growth rate with this substrate was very low (0.015 h-l). Anaerobically grown cells of strain Mast1 exhibited immediate oxygen-dependent respiration. The endogenous oxygen reduction rate was proportional to the polyglucose content. The rate of aerobic respiration of pyruvate was also directly related to the polyglucose content indicating that this organism was only able to respire with oxygen as long as polyglucose was present. Maximum oxygen reduction rates were found at air saturating concentrations and were relatively low (3-50 nmol 0, min-’ (mg protein)- ’ ). Catalase was constitutively present in anaerobically grown cells. When batch cultures were exposed to oxygen, growth ceased immediately and polyglucose was oxidized to acetate within 40-50 h. Like the oxygen reduction activity, the nitro blue tetrazolium (NBT)-reduction activity in these cells was proportional to the polyglucose content. Under anaerobic starvation conditions there was no correlation between the NBT-reduction activity and polyglucose concentration and polyglucose was degraded slowly within 240 h. The ecological significance of aerobic polyglucose consumption is discussed. Keywords: DesulfoCbrio salexigens; Sulfate reduction; Glucose; Polyglucose; Oxygen-dependent respiration 1. Introduction Like many other ‘strict anaerobes’, several sul- fate-reducing bacteria (SRB) are capable of immedi- ate reduction of oxygen when this electron acceptor * Corresponding author. Tel.: +31 (50) 363 2036; fax: +31 (50) 363 2154; e-mail: [email protected] becomes available. Under such conditions sulfate reduction ceases or proceeds at low rates provided that the oxygen concentration remains low [ 1,2]. This is in contrast to results from field measurements indicating high rates of dissimilatory sulfate reduc- tion in oxic sediment layers [3-61. Immediate reduc- tion of oxygen by SRB suggests that they have constitutive oxygen-reducing enzymes. It has been demonstrated that a few redox carriers, like cy- 016%6496/96/$15.00 Copyright 0 1996 Federation of European Microbiological Societies. Published by Elsevier Science B.V. PII SO 168-6496(96)00060-8

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ELSEVIER FEMS Microbiology Ecology 21 (1996) 243-253
The role of polyglucose in oxygen-dependent respiration by a new strain of Desulfovibrio salexigens
Ed W.J. van Niel a3 * , Teresa M. Pedro Gomes a, Anne Willems b, Matthew D. Collins b, Rudolf A. Prins a, Jan C. Gottschal a
a Department of Microbiology, Unitiersity of Groningen, Kerklaan 30, 9751 NN Haren, The Netherlands
h Institute of Food Research, Reading Laboratory, Earley Gate, Whiteknights Road, Reading RG6 6BZ, UK
Received 17 May 1996; revised 5 August 1996: accepted 21 August 1996
Abstract
Desulfooibrio salexigens strain Mast1 was isolated from the oxic/anoxic interface of a marine sediment. Growth under sulfate-reducing conditions was accompanied by polyglucose accumulation in the cell with every substrate tested. Highest polyglucose storage was found with glucose (0.8-1.0 g polyglucose (g protein)-‘), but the growth rate with this substrate was very low (0.015 h-l). Anaerobically grown cells of strain Mast1 exhibited immediate oxygen-dependent respiration. The endogenous oxygen reduction rate was proportional to the polyglucose content. The rate of aerobic respiration of pyruvate was also directly related to the polyglucose content indicating that this organism was only able to respire with oxygen as long as polyglucose was present. Maximum oxygen reduction rates were found at air saturating concentrations and were relatively low (3-50 nmol 0, min-’ (mg protein)- ’ ). Catalase was constitutively present in anaerobically grown cells. When batch cultures were exposed to oxygen, growth ceased immediately and polyglucose was oxidized to acetate within 40-50 h. Like the oxygen reduction activity, the nitro blue tetrazolium (NBT)-reduction activity in these cells was proportional to the polyglucose content. Under anaerobic starvation conditions there was no correlation between the NBT-reduction activity and polyglucose concentration and polyglucose was degraded slowly within 240 h. The ecological significance of aerobic polyglucose consumption is discussed.
Keywords: DesulfoCbrio salexigens; Sulfate reduction; Glucose; Polyglucose; Oxygen-dependent respiration
1. Introduction
Like many other ‘strict anaerobes’, several sul- fate-reducing bacteria (SRB) are capable of immedi- ate reduction of oxygen when this electron acceptor
* Corresponding author. Tel.: +31 (50) 363 2036; fax: +31
(50) 363 2154; e-mail: [email protected]
becomes available. Under such conditions sulfate reduction ceases or proceeds at low rates provided that the oxygen concentration remains low [ 1,2]. This is in contrast to results from field measurements indicating high rates of dissimilatory sulfate reduc- tion in oxic sediment layers [3-61. Immediate reduc- tion of oxygen by SRB suggests that they have constitutive oxygen-reducing enzymes. It has been demonstrated that a few redox carriers, like cy-
016%6496/96/$15.00 Copyright 0 1996 Federation of European Microbiological Societies. Published by Elsevier Science B.V.
PII SO 168-6496(96)00060-8

244 E. W. J. wn Niel et al. / FEMS Microbiology Eco1og.v 2 I C 1996) 213-253
tochrome c3 [7] and rubredoxin oxidase [8], may use oxygen as an alternative electron acceptor. The pres- ence of protective enzymes against oxygen, such as catalase and superoxide dismutase, has often been reported [9,10] suggesting that these species may from time to time experience exposure to oxygen.
Most of the aerotolerant SRB tested oxidize dif- ferent substrates with oxygen, such as HZ, organic compounds and inorganic sulfur compounds, at rates comparable to those of aerobic bacteria [9.11]. Dur- ing respiration with oxygen ATP is generated, but in most cases is apparently not used for biomass pro- duction [ 11,121. Nevertheless, Marschall et al. [I] observed some growth in aerated cultures of Desul- fouibrio desulfuricans strains at oxygen concentra- tions of up to 1.5% in the gas phase. They could only establish such microaerobic growth in stabilized oxygen gradients. Krekeler and Cypionka [2] showed that D. desuljiiricans preferred oxygen to nitrate, which in turn is preferred to sulfate. In general, it has been suggested that preference for oxygen reduction by ‘strict anaerobes’ arises from scavenging mecha- nisms to detoxify oxygen [ 13,141. In this way oxy- gen-labile enzymes can be protected.
In the course of a study on the occurrence of amino acid-utilizing organisms at the oxic/anoxic interface of marine bacterial mats near hydrothermal gas seeps, a sulfate-reducing bacterium was isolated on alanine as the sole carbon source. The organism, strain Mastl, was identified as belonging to D. salexigens. Growth with various substrates under conditions of sulfate reduction and non-limiting nu- trient concentrations, resulted in intracellular polyglucose storage similar to that previously found with D. gigas [ 151. Recently, Santos et al. [12] showed that D. gigas could respire polyglucose with oxygen. They proposed that the presence of polyglu- case would enable its survival under oxic conditions for considerable periods of time. Therefore, we wanted to know how D. salexigens strain Mast1 would respond to oxygen exposure and whether polyglucose would play a role in this. The results show that this sulfate-reducing bacterium appears to reduce oxygen even up to air saturating concentra- tions; this makes it different from those microaerobic SRB reported thus far. In this organism intracellular polyglucose seemed to be an absolute requirement for aerobic respiration of exogenous energy sources.
2. Materials and methods
2. I. Source of organisms
The sulfate-reducing strain Mast 1 originated from the top layer (O-5 mm) of a marine sediment with a microbial mat at 10 m depth in the center of Paleo- hori Bay on the south shore of the Isle of Milos, Aegean Sea, Greece [16]. Throughout the microbial mat hydrothermal gas seeps were present. The organ- ism was isolated from an enrichment culture with alanine as the sole carbon and energy source. Desul- fovibrio salexigens DSM 2638 was obtained from the Deutsche Sammlung von Mikroorganismen (Braunschweig). D. gigas NCMIB 9332 was kindly provided by C.M.H. Hensgens.
2.2. Media and growth corlditions
The organisms were cultured in a buffered basal salts medium containing (g 1~ ’ demineralized water): NaCl, 30; MgC12. 6H20, 1.0; Na,SO,, 4.0: KCl, 0.7; CaCl, . 2H,O, 0.15; NH,Cl, 0.5; NaHCO,, 0.2; KBr, 0.1; SrCl, 6H,O, 0.04; MOPS, 2.09; re- sazurin, 0.001. Screw-cap tubes and rubber stop- pered serum bottles (300 ml) with basal salts medium for batch cultures were flushed with N?/CO? (80/20, v/v). After sterilizing, the medium (referred to as MPM-medium) was completed by adding (per liter): 80 ml of 0.2 mM Na,HPO,, 1 ml trace elements solution STE2 [17], 1 ml vitamins solution [ 181, 10 ml of a 10% yeast extract solution, 10 ml of 1.25% Na,S . 9H,O solution. The pH of the final medium was adjusted to 7.4 with 1 N HCl. Cultures were incubated under strictly anaerobic conditions in the dark at 30°C. The medium for continuous cul- tures was identical, except that the yeast extract concentration was lowered by a factor of 10 and no sulfide was added. Substrate concentrations are indi- cated in the text.
Chemostat cultures of D. salexigens strain Mast1 were grown at 30°C the pH was kept at 7.4 by automatic titration with 1 N NaOH. The chemostat had a working volume of 750 ml, was stirred by means of a magnetic stirrer, and the headspace of the culture vessel was flushed continuously with N,/CO, gas (80/20, v/v), freed of traces of 0, by passing over hot copper turnings.

E. W.J. uan Niel et al. / FEMS Microbiology Ecology 21 (19961 243-253 245
2.3. Isolation and maintenance of the strains
A slurry, made of the top layer of the sediment (ca. 3 ml) with 3 ml of basal medium, was used as a 5% inoculum for enrichment on alanine. A pure culture of the dominant organism was obtained by repeated application of deep-agar dilution series un- der N/CO, (80/20, v/v) [19]. Purity of the strain was checked on anaerobic agar plates and by micro- scopical observations of the cultures after growth in defined medium containing casamino acids, alanine and pyruvate. Stock cultures were transferred every 2-4 weeks and were stored at 4°C in the dark.
2.4. Electron microscopy
Washed cells of batch or continuous cultures were fixed with 0.1% glutaraldehyde in basal salts medium. Whole cells were negatively stained with 1% uranyl acetate. For sectioning, the cells were dehydrated in a graded series of ethanol solutions and embedded in Epon 8 12. The material was viewed in a Philips CM10 electron microscope. Polysaccha- ride granules of thin sections were stained on golden
grids with phosphotungstic acid [20].
2.5. 16s rRNA sequence analysis
DNA was extracted from cells by the method of Lawson et al. 1211. The 16s rRNA gene was ampli- fied as described previously [22] and purified using a Prep-A-Gene kit (Biorad) according to the manufac- turer’s instructions. The 16s rDNA fragment was sequenced using a Taq DyeDeoxy Terminator Cycle Sequencing kit (Applied Biosystems) and an auto- mated sequencer (Applied Biosystems, model 373A).
2.6. Determination of oxygen consumption rates and catalase acti@
Maximum specific oxygen consumption rates (QOyx- values) were determined polarographically in a biological oxygen monitor (BOM) (Yellow Springs Instruments, Yellow Springs, OH). Sub- strates and inhibitors were added with 500 ~1 Hamil- ton syringes. Cells were harvested by centrifugation for 10 min at 10,000 rpm and 4°C. After washing with basal salts buffer (pH 7.41, they were resus-
pended to a final protein concentration of 300-500
mg 1-l and kept on ice until use. Catalase activity was determined by adding 2.2
mM H,O, (final concentration) to an anaerobic cell suspension in the BOM. The initial oxygen produc- tion rate was taken as a measure for the catalase activity. The rate was corrected for the chemical decomposition rate.
2.7. Reduction of tetrazolium salts
The determination of activity in the cells was based on the ability of SRB to reduce tetrazolium salts as described by Fukui and Takii [23]. Two salts (Sigma, USA) were tested: triphenyltetrazolium chloride (TTC) and nitro blue tetrazolium (NBT). The reduction activity with TTC to formazan was very low, but with NBT maximum formazan produc- tion was seen within 40 min (at 30°C in the dark). Therefore the latter was used in the experiments. The effect of sulfate on the NBT-reduction activity was negligible, which supported the findings of Fukui and Takii [23]. Cells were withdrawn from cultures and 1 ml was exposed for a short time to air to oxidize sulfide, before addition to a sterile, anaerobic assay mixture in Hungate-tubes. The assay mixture consisted of 10 ml basal salts medium and 0.5 ml of 0.2% NBT (oxygen-free aqueous solution, 0.2 ,um filter sterilized). The reaction was terminated after 1 h by adding glutaraldehyde (final concentration 1 .O% w/w). Intracellular formazan was extracted by mix- ing 0.5 ml of the sample with 1 ml iso-amylalcohol, sonicating at 40 W for 4 min, and centrifuging. The absorbance of the alcohol layer was measured at 536 nm using a Shimadzu UV-1601 spectrophotometer.
2.8. Analytical techniques
Cells were counted microscopically with a Biirker-Tiirk counting chamber after appropriate di- lution of culture samples in basal salts medium containing 1% formaldehyde. Growth was followed by optical density determinations at 660 nm in a Vitalab calorimeter (Vital Scientific, Dieren) or with a Biotron calorimeter btr 101. Desulfoviridin was identified according to Widdel [24]. Desulfoviridin- positive D. gigas served as reference. Cytochromes were identified in the cytoplasmic fraction of soni-

246 E. W.J. ~‘an Niel et al. / FEMS Microbiology Ecology 21 (1996) 243-253
cated cell suspensions after centrifugation (36,000 X g, 60 min) by recording reduced versus oxidized difference spectra with an Aminco DW-2000 UV- VIS spectrophotometer (SLM Instruments). Organic acids and alcohols were analyzed by gas chromatog-
raphy, as described earlier 125,261. Sulfide was deter- mined calorimetrically [27]. Ammonium was mea- sured calorimetrically [ZS]. Glucose was measured with both the anthrone method [29] and with glucose oxidase (Boehringer, Germany), exclusively detect- ing glucose. Polyglucose in washed cells was deter-
mined as glucose equivalents by the anthrone method or after hydrolysis in 2 N H,S04 for 20 min at 120°C as glucose with glucose oxidase. Polyglucose was not removed from the cells prior to its determi- nation. Protein was measured according to the mi- croburette method [30] with bovine serum albumin as a standard.
3. Results
3.1. Isolation and characterization of strain Mast1
Repeated application of deep-agar dilution series with 10 mM alanine resulted in one type of colony in all the positive tubes of the highest dilution series (dilution 10e9). After growth of a pure culture on L-alanine, motile rods, sometimes in pairs, were seen. The cells were 2.5-3.5 pm in length and 0.5 pm in width. Spores were never observed. The cells stained gram-negative.
Growth occurred at temperatures ranging from 15°C to 37°C with an optimum specific growth rate at 31°C of 0.05 h-’ on alanine. The optimum pH range for growth was 7.0 to 7.5. Growth was ob- served over a salinity range from 1 to 8% of NaCl.
The cells contained desulfoviridin and the soluble extract exhibited the characteristic absorption bands of cytochrome cs with maxima at 553 nm and 524 nm after reduction with sodium dithionite. The oxi- dized extract showed an absorption peak at 420 nm.
The full 16s rRNA gene sequence of strain Mast1 has been deposited in the EMBL Data Library under accession number X82622. Comparative sequence analysis identified strain Mast 1 as a member of the genus Desulfovibrio and revealed more than 99% sequence similarity with the type strain of Desul-
fovibrio salexigens (ATCC 14822). Considerably lower levels of sequence relatedness (ca. 86-90%) were shown to other Desulfovibrio species. The isolate was therefore designated as D. salexigens strain Mast 1.
Strain Mast1 differed from the type strain with respect to electron donor usage [31,32] in that it was unable to grow on methanol or succinate. The sub- strates were incompletely oxidized to acetate and CO,. Pyruvate, fumarate and malate were fermented to acetate in the absence of sulfate. Besides sulfate, strain Mast1 was able to use thiosulfate as electron
acceptor.
3.2. Accumulation of polyglucose during growth in batch cultures
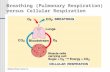
Like the type strain [32], strain mast1 was able to grow on glucose. The maximum specific growth rate of strain Mastl, as calculated from the increase in cell numbers, was low: 0.015 h-‘. Growth on glu- cose resulted in a culture with irregularly shaped cells widely differing in size (1.5 to 9 pm in length and 0.8 to 1.2 pm in width) (Fig. 1E). The cells contained electron-transparent granules (Fig. IC) which consisted of polyglucose as was determined by staining with phosphotungstic acid (Fig. ID). Formation of these granules started right from the beginning of the exponential phase when growing on
Table I
Protein concentrations (mg 1-l ) and polyglucose contents (g
polyglucose (g protein)-’ ) of Desulfouibrio salexigens strain
Mast1 and DSM 2638 grown in batch cultures on different
substrates (10 mM, except H,) and sulfate. Polyglucose was
measured as glucose equivalents with glucose oxidase
Strain Substrate Protein Po1yg1ucose
content
Mast1 glucose 77 0.88 pyruvate 30 0.59 alanine 46 0.37 glycerol 31 0.43 lactate 22 0.23 formate + acetate 19 0.18 H 2 + acetate 48 0.17
pyruvate, no sulfate 9 0.01
DSM 2638 alanine 55 0.29 glucose 116 0.0 1
E. W.J. um Niel ef al./ FEMS Microbiology Ecology 21 (1996) 243-253 247
glucose (Fig. 1B). No granules were observed when
grown on alanine (Fig. 1Al. However, polyglucose also accumulated in batch cultures of strain Mast1 growing on other substrates, provided that sulfate was present (Table 11. During fermentative growth on pyruvate, in the absence of sulfate, almost no polyglucose was stored (Table I). In batch culture strain D. salexigens strain DSM 2638T also showed
polyglucose accumulation when grown on alanine. The maximum growth rate of the type strain on glucose (0.007 h- ’ 1 was twofold lower than that of
strain Mast 1, but polyglucose was hardly produced (Table 1).
When grown on different concentrations of glu-
case (20-40 mM) strain Mast1 consumed maximally
9- 10 mM of this substrate. The pH dropped only 0.5 units from 7.4 to 6.9. It would appear that another component than glucose was limiting, but supple- menting the medium with trace elements, Fe-EDTA, yeast extract or vitamins, did not improve consump- tion of glucose. Neither was the concentration of sulfide inhibitory, because addition of 10 mM sulfide or regular flushing of the headspace with N,/CO, (80/20, v/v> during the mid-exponential phase did not alter the growth rate or cell density. Moreover, in fresh medium strain Mast1 grew on alanine with the
same prnax as in supematant of an outgrown glucose culture supplemented with trace elements, yeast ex-
Fig. 1. (A) Transmission electron micrograph of Desulfouibrio salexigens strain Mast1 grown on alanine. (B) Transmission electron
micrograph of strain Mast1 grown on glucose during the early exponential phase. (C) Transmission electron micrograph of strain Mast1 grown on 20 mM glucose. The white spots within the cells consist of polyglucose. (D) As (C), cell coupes were stained with
phosphotungstic acid. (El Phase contrast micrograph of strain Mast1 grown on 20 mM glucose. Bars in panels (A-D) represent 0.5 micron.
248 E. W.J. mn Niel et al. / FEMS Microbiology Ecology 21 f 1996) 213-253
tract, vitamins and phosphate. Apparently no in- hibitory compounds were present in the spent medium of the glucose culture (data not shown).
3.3. Polyglucose and oxygen-dependent respiration
Washed cell suspensions of anaerobically grown cultures of strain Mast1 exhibited immediate endoge-
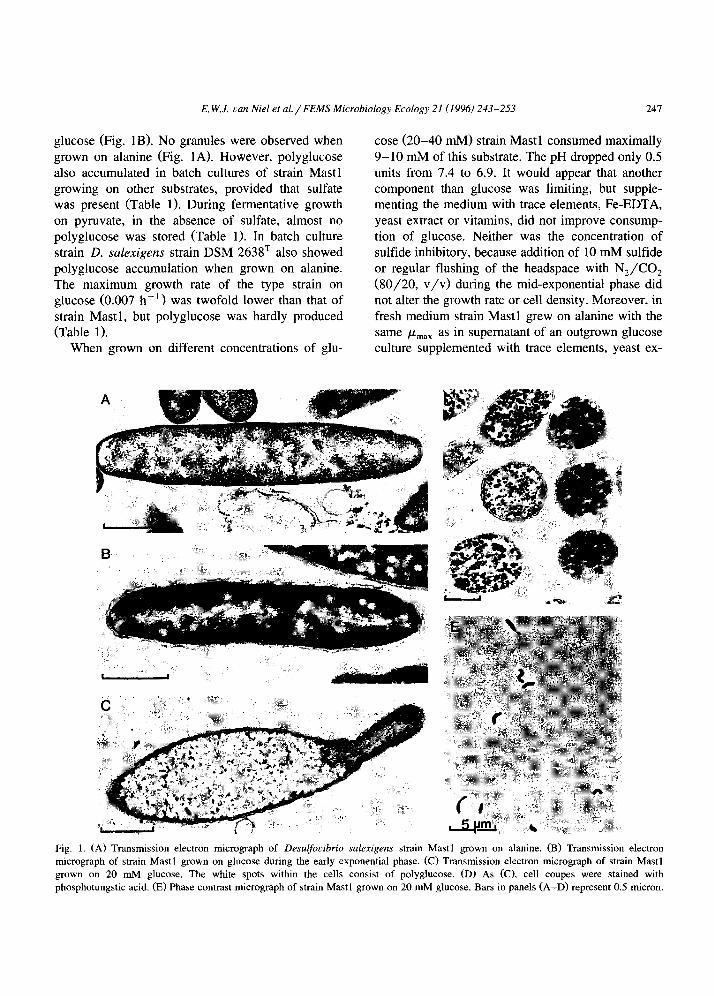
nous oxygen consumption. This oxygen consumption was identified as a biological activity, based on its linear dependence on the density of the cell suspen- sion and on the absence of detectable activity with autoclaved suspensions. Furthermore, it appeared that the initial endogenous respiration rate, at air satura- tion, was linearly correlated to the polyglucose con- tent of the cell (r = 0.98) (Fig. 2). Hence, cells without polyglucose were expected to have a very low or no endogenous respiration rate. Indeed. when strain Mast1 was grown on pyruvate without sulfate, almost no polyglucose was stored, and the endoge- nous respiration rate was in the order of O-3 nmol O? min-’ (mg protein) -’ (Fig. 2). A cell suspension of strain DSM 2638, grown anaerobically on alanine with a content of 0.29 g polyglucose (g protein)-‘, exhibited a similar activity (8.5 nmol O2 min- * (mg protein)- ’ ) as strain Mast 1, but no further data were obtained with cells containing different amounts of polyglucose. Interestingly, cells of strain Mast 1, grown on H2 with CO, and acetate showed an endogenous respiration rate much higher than ex- pected from the polyglucose content of the cells.
The substrates that could be oxidized aerobically were: alanine, glucose, pyruvate, glycerol, sulfide and thiosulfate, but not L(+)-lactate. All the organic substrates, including polyglucose, were stoichio- metrically oxidized to acetate. The oxidation rates with the external substrates were somewhat higher than with endogenous substrates: e.g. maximum spe- cific oxidation rates for pyruvate and glycerol were: 20 and 50 nmol O2 min-’ (mg protein)-‘, respec- tively. These figures are corrected for endogenous activity. Catalase was present constitutively in anaer- obically grown cells and its activity was 150 Fmol H,O, min-’ (mg protein)-‘.
Only small amounts of polyglucose (ca. 0.04 g (g protein)- ’ ) were stored when strain Mast 1 was grown in continuous culture with 50 mM pyruvate under sulfate-limitation (4 mM sulfate) at a dilution rate of
26
0v= I 0.0 0.2 0.4 0.8 0.8 1.0
polyglucose content (g (g protein)-‘)
Fig. 2. Relationship between the initial specific endogenous oxy-
gen respiration rate of Drsdfo~~ibrio snkrigrrz.s strain Mast 1 and
the polyglucose content of the cells. Both strains were anaerobi-
cally grown in batch culture on different substrates (see Table I ), washed and exposed to oxygen in the biological oxygen monitor.
Strain Mast1 pregrown on various substrates with sulfate in excesh
(0) and on pyruvate without sulfate (0).
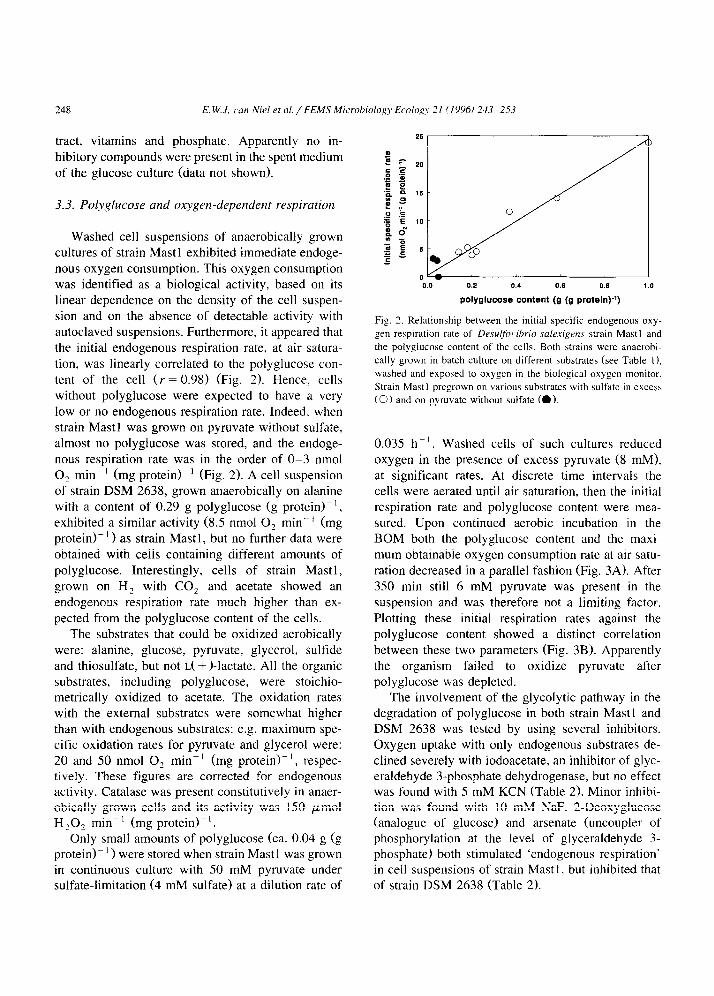
0.035 h-‘. Washed cells of such cultures reduced oxygen in the presence of excess pyruvate (8 mM), at significant rates. At discrete time intervals the cells were aerated until air saturation, then the initial respiration rate and polyglucose content were mea- sured. Upon continued aerobic incubation in the BOM both the polyglucose content and the maxi- mum obtainable oxygen consumption rate at air satu- ration decreased in a parallel fashion (Fig. 3A). After 350 min still 6 mM pyruvate was present in the suspension and was therefore not a limiting factor. Plotting these initial respiration rates against the polyglucose content showed a distinct correlation between these two parameters (Fig. 3B). Apparently the organism failed to oxidize pyruvate after polyglucose was depleted.
The involvement of the glycolytic pathway in the degradation of polyglucose in both strain Mast1 and DSM 2638 was tested by using several inhibitors. Oxygen uptake with only endogenous substrates de- clined severely with iodoacetate, an inhibitor of glyc- eraldehyde 3-phosphate dehydrogenase, but no effect was found with 5 mM KCN (Table 2). Minor inhibi- tion was found with 10 mM NaF. 2-Deoxyglucose (analogue of glucose) and arsenate (uncoupler of phosphorylation at the level of glyceraldehyde 3- phosphate) both stimulated ‘endogenous respiration’ in cell suspensions of strain Mast 1, but inhibited that of strain DSM 2638 (Table 2).
E. W.J. ran Niel et al. / FEM.7 Microbiology Ecology 21 (1996) 243-253 249
E t = 0.02 8 I
Time (min) Polyglucore content (Q (g protein)-I)
Fig. 3. (A) Initial respiration rates (0) and polyglucose concentrations (0) of washed cells of Desulfor,ibrio sakrigens strain Mast1
measured at discrete time intervals after sparging the cell suspension to air saturation. At the beginning the suspension was provided with 8
mM pyruvate. (B) Relation between the initial specific respiration rate on pyruvate with oxygen (not corrected for endogenous respiration)
and the polyglucose content of the cells.
3.4. Long term e.xposure to oxygen
Anaerobic batch cultures grown with 20 mM
pyruvate and excess sulfate were split in two at mid-exponential phase. One of them was further incubated with sterile air with the headspace contain- ing about 10% oxygen. The other one was main- tained anaerobic. These cultures were shaken contin-
L
g 80
0 8 2 E 40
I
0
0.6
8 5 0.4
c P :
0.2
0.0
B-
Cl ;
r
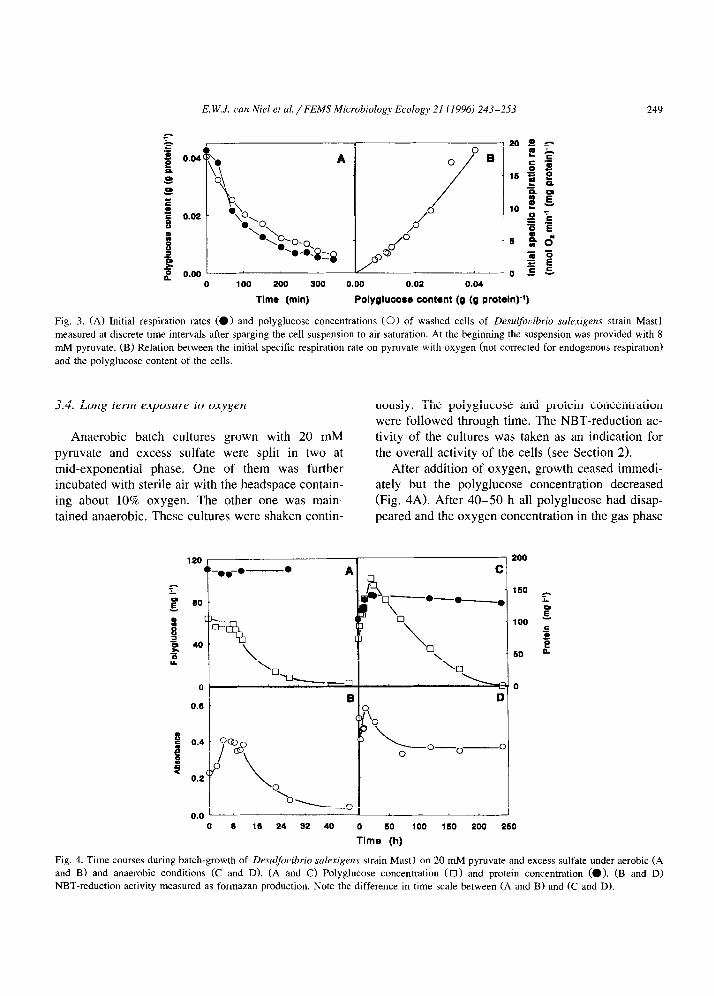
uously. The polyglucose and protein concentration were followed through time. The NBT-reduction ac- tivity of the cultures was taken as an indication for the overall activity of the cells (see Section 2).
After addition of oxygen, growth ceased immedi- ately but the polyglucose concentration decreased (Fig. 4A). After 40-50 h all polyglucose had disap- peared and the oxygen concentration in the gas phase
cl
200 i. ‘0
\c-•-•* --A 150 ,”
.L
P
\
100
f
“\, 60 E
0
D
3L 0
o-o-o
0 8 18 24 92 40 0 60 100 150 200 250
Time (h)
Fig. 4. Time courses during batch-growth of Desulfotibrio sakrigens strain Mast1 on 20 mM pyruvate and excess sulfate under aerobic (A and B) and anaerobic conditions (C and D). (A and C) Polyglucose concentration (0) and protein concentration (0). (B and D)
NBT-reduction activity measured as formazan production. Note the difference in time scale between (A and B) and (C and D).

250 E. W.J. uan Niel et al. / FEM.5 Microbiology Ecolog?; 21 (1996) 243-253
Table 2 4. Discussion Relative oxygen consumption rates of washed cells of Desdfouib-
rio sdexigens strain Mast1 and DSM 2638 with polyglucose as
the only electron donor under different conditions in the biological
oxygen monitor at 30°C. At the start of the measurements cell
suspensions were saturated with air. The endogenous rate mea-
sured without any addition was taken as 100%. All other rates
were related to that standard
Strain Condition O1 consumption
rate (%j
Mast1 no addition 100
iodoacetate ( 10 mM) 8-28
KCN (5 mM) 100
NaF (10 mM) 80
2-deoxyglucose (10 mM) 150-190
arsenate ( 10 mM) 180
DSM 2638 no addition 100
iodoacetate (10 mM) 13
2-deoxyglucose (10 mM) 72
arsenate (10 mM) 83
The isolated strain Mast1 exhibited the typical characteristics of Desulfovibrio salexigens. Strain Mast1 differed from the type strain in only a few properties, one of them being the high extent of polyglucose accumulation in cells during growth on glucose. Recently, several SRB were isolated from the oxic layer of a stratified marine water column and found to be closely related to D. salexigens [33]. Comparison of the 16s rRNA sequences of these isolates with those of D. salexigens ATCC 14822 and D. salexigens strain Mast 1 revealed sequence similarities of 92.5% and 93-95%, respectively, in- dicating that strain Mast1 is related more closely to D. salexigens than to the other isolates.
had dropped from 10 to 9%. The NBT-reduction capacity first increased about twofold in the first 4 h and then declined in parallel with the polyglucose concentration (Fig. 4B). During the time period that NBT-reduction rate declined this activity was pro- portional to the polyglucose concentration (r = 0.98, data not shown).
Like D. salexigens DSM 2638 [32], strain Mast1 consumed no more than 9-10 mM of glucose during growth in batch culture. No convincing explanation for this phenomenon is yet available. Results ob- tained revealed no limitation of medium components or toxicity by sulfide. Growth on glucose was slow for both D. salexigens strains suggesting that the restricted consumption in batch culture was some- how associated with the poor metabolism of this substrate.
In the anaerobic cultures growth was still ob- served during the first 10 h (Fig. 4C). The polyglu- case concentration of the culture parallelled the in- crease of the protein concentration. During the initial exponential phase the NBT-reduction capacity in- creased somewhat. After entering the stationary phase, this activity dropped within two days to about 67% of the maximum activity and remained at this value until the end of the experiment (Fig. 4D). Meanwhile polyglucose disappeared at a low rate over a period of about 240 h. The results indicated that the NBT-reduction activity was retained inde- pendently of the polyglucose content. As a control strain Mast1 was grown in batch cultures on pyru- vate without sulfate. These cultures did not accumu- late polyglucose. Washed cell suspensions of this culture hardly showed aerobic endogenous respira- tion and pyruvate was not respired with oxygen. Also, no formazan was produced from NBT during incubations of 1 h.
It appeared that on every growth substrate, strain Mast1 was able to produce polyglucose, with signifi- cant variation in the amounts stored for the various substrates (Table 1). The polymer was stored in the form of many small globular particles (Fig. 1C) rather than in the form of a few big granules. This means that the total volume of the globules (= polyglucose content) approaches a linear relationship to the total surface area of the globules. If it is assumed that enzymatic breakdown of polyglucose takes place at the surface of the particles, this may explain why the polyglucose degradation rate is pro- portional to the polyglucose content (Fig. 2).
Fig. 2 also indicates that there will be (almost) no aerobic endogenous respiration when the cells are depleted of polyglucose. But more surprisingly, when cell suspensions were drained of polyglucose, their aerobic respiration of pyruvate also declined. This would imply that strain Mast1 will cease to reduce oxygen as soon as all polyglucose has been con- sumed, irrespective of the presence of sufficient energy sources in the medium. A further indication

E. W.J. uan Niel et al. / FEMS Microbiology Ecology 21 (1996) 243-253 251
that polyglucose played a crucial role in the respira-
tion with oxygen is obtained from the NBT-reduc- tion activity measurements. This activity also ap- peared proportional to the polyglucose concentration in batch cultures exposed to oxygen (Fig. 4A and B). However, such a correlation between NBT-reduction activity and polyglucose content was absent in anaer- obic cultures under starvation conditions. Under these conditions breakdown of macromolecules other than polyglucose, such as proteins and RNA, will also provide the cell with energy for maintenance require- ments [34]. These processes together probably supply the reducing equivalents for the reduction of NBT in the anaerobic cultures. The NBT-reduction activity dropped to 67% within two days of starvation and remained at that value for a long time. This is similar to the observations made with anaerobic cultures of Desulfobacter latus under starvation conditions [35]. These authors also found that the mean 16s rRNA content decreased exponentially in two days, to 30% of the maximum value obtained for exponentially growing cells. Apparently with strain Mast1 these processes cannot be coupled to the reduction of oxygen. Most likely, certain enzymes involved in the metabolism of these macromolecules are sensitive to oxygen. This is probably the reason why this organ- ism would not grow at the expense of oxygen. This same paradox has been observed with other SRB [1,9] and other ‘strict’ anaerobes such as a non- sporulating strain of Clostridium acetobutylicum [ 131.
With respect to the reasons for the dependency of aerobic pyruvate oxidation activity on the polyglu- case degradation we can only speculate. As in D. gigas [12], aerobic conversion of polyglucose is me- diated by the glycolytic pathway as was suggested by the effect of iodoacetate on the endogenous metabolic activity (Table 21. In this process ATP is produced which may be necessary for the pyruvate uptake. In addition, NADH produced in glycolysis may be used to reduce oxygen to protect oxygen-la- bile enzymes such as pyruvate:ferredoxin oxido- reductase, to allow for continued pyruvate metabolism. Indeed, significant NADH oxidase ac- tivity has been found in cell-free extracts of strain Mast1 (E.W.J. van Niel, unpublished results). Chen et al. [8,36] described an electron transfer chain in D. gigas for the oxidation of NADH with oxygen, which was composed of NADH-rubredoxin oxido-
reductase, rubredoxin and a rubredoxin-oxygen ox-
idoreductase. Strain Mast1 was able to respire all the substrates
tested with oxygen, except L( + )-lactate. Stams and Hansen [37] demonstrated that L( + )-lactate dehydro- genase in D. desuljiiricans was very oxygen-sensi- tive. A similar oxygen-sensitive lactate dehydroge- nase may be present in strain Mastl.
The results obtained clearly show that D. salexi- gens strain Mast1 is distinct from the earlier reported microaerobic SRB, such as D. desulfiricuns [2,11], by its ability to reduce oxygen at high oxygen con- centrations, its high polyglucose accumulation and its relatively low specific respiration rates. With respect to the latter two characteristics, strain Mast1 is quite similar to D. gigas [12]. It must be con- cluded that SRB developed several different mecha- nisms to cope with oxic environments.
Strain Mast1 was isolated from the oxic/anoxic interface of a cyanobacterial mat. Thus this organism experiences periodic exposure to oxygen. Therefore, we postulate a model according to which organisms such as strain Mast 1 have adapted to alternating oxic/anoxic conditions. During darkness, under anaerobic conditions, strain Mast1 grows with con- comitant polyglucose accumulation by respiring vari- ous small organic compounds and hydrogen, pro- duced by fermentative organisms. During daytime excessive quantities of oxygen are produced by the cyanobacteria. As a consequence, growth of strain Mast1 would cease, but the activity of the cells can be maintained at a high level by aerobic respiration of polyglucose. Such a condition was termed ‘active dormancy’ by LeGall and Xavier [38] (as opposed to sporulation as ‘passive dormancy’). This mainte- nance of a high level of cell activity enables such cells to resume growth (and polyglucose production) immediately upon anaerobiosis. The experimental results obtained in this study are consistent with the proposed model, and studies to substantiate the valid- ity of this model should now also be undertaken in the field.
Acknowledgements
The authors are grateful to K.A. Sjollema for the preparation of electron micrographs and to Dr. T.A. Hansen for his constructive comments on the

252 E. W. J. wm Niel et al. / FEMS Microbiology Ecology 21 C 1996) 243-253
manuscript. This work was supported by grants from the EU, contract Nr. MAST II CT 930058 and CHRX CT93 0194; BIO-2-CT93-0119.
References
[II
Dl
[31
[41
151
[61
[71
[81
191
1101
[Ill
[I21
1131
1141
[151
Marschall. C., Frenzel, P. and Cypionka, H. (1993) Influence
of oxygen on sulfate reduction and growth of sulfate-reduc-
ing bacteria. Arch. Microbial. 159, 168-173.
Krekeler, D. and Cypionka, H. (1995) The preferred electron
acceptor of Desulfovibrio desulfuricans CSN. FEMS Micro-
biol. Ecol. 17, 271-278.
Canfield, D.E. and DesMarais. D.J. (1991) Aerobic sulfate
reduction in microbial mats. Science 25 1, 1471-1473.
Cohen, Y. (1989) Photosynthesis in cyanobacterial mats and
its relation to the sulfur cycle: A model for microbial sulfur
interactions. In: Microbial Mats: Physiological Ecology of
Benthic Microbial Communities (Cohen, Y. and Rosenberg,
E., Eds.), pp. 22-36. ASM, Washington, DC.
Friind, C. and Cohen, Y. (1992) Diurnal cycles of sulfate
reduction under oxic conditions in cyanobacterial mats. Appl.
Environ. Microbial. 58, 70-77.
Jflrgensen, B.B. and Bak, F. (1991) Pathways and micro-
biology of thiosulfate transformations and sulfate reduction
in a marine sediment (Kattegat, Denmark). Appl. Environ.
Microbial. 57. 847-856.
Postgate, J.R. ! 1956) Cytochrome cj and desulphoviridin:
pigments of the anaerobe Desulfoclibrio desuffuricuns. J.
Gen. Microbial. 14, 545-572.
Chen, L., Liu, M.-Y., LeGall, J., Fareleira, P., Santos, H. and
Xavier, A.V. (1993) Rubredoxin oxidase, a new flavo-
hemo-protein, is the site of oxygen reduction to water by the
‘strict anaerobe’ Desulforibrio gigas. Biochem. Biophys.
Res. Commun. 193, 100-105.
Abdollahi, H. and Wimpenny, J.W.T. (1990) Effects of
oxygen on the growth of Desulfoc,ibrio desulfuricarzs. J.
Gen. Microbial. 136, 1025- 1030.
Hardy, J.A. and Hamilton. A. (1981) The oxygen tolerance
of sulfate-reducing bacteria isolated from North Sea waters.
Curr. Microbial. 6, 259-262.
Dilling, W. and Cypionka, H. (1990) Aerobic respiration in
sulfate-reducing bacteria. FEMS Microbial. Lett. 7 I. 123-
128.
Santos, H., Fareleira, P., Xavier. A.V., Chen, L., Liu, M.-Y.,
LeGall, J. (1993) Aerobic metabolism of carbon reserves by
the ‘obligate anaerobe’ Dedfmibrio gigas. Biochem. Bio-
phys. Res. Commun. 195, 551-557. O’Brien, R.W. and Morris, J.G. (1971) Oxygen and the
growth and metabolism of Clostridium acetobu~licum. J.
Gen. Microbial. 68, 307-3 18. Bentzen, G. and Larsen. H. (1989) Oxygen activation and
defence against oxygen toxicity in a psychrophilic bac- teroidaceae. Arch. Microbial. 15 1, 95- 100.
Stams, A.J.M., Veenhuis, M.. Wenk, G.H. and Hansen, T.A.
(1983) Occurrence of polyglucose as a storage polymer in
Desuljkwibrio species and Desulfobulbus propionicus. Arch.
Microbial. 136. 54-59.
[16] Dando, P.R., Hughes, J.A. and Thiermann, F. (1995) Prelimi-
nary Observations on Shallow Hydrothermal Vents in the
Aegean Sea. In: Hydrothermal Vents and Processes, Vol. 87
(Parson. L.M., Walker, CL. and Dixon. D.-R.. Eds.), pp.
303-313. Special Publication Geological Society, London.
[17] Schut. F.. de Vries. E.J., Gottschal. J.C., Robertson. B.R.,
(181
[I91
LOI
El1
WI
1231
I241
[251
1261
1271
[281
Harder, W., Prins, R.A. and Button, D.K. (1993) Isolation of
typical marine bacteria by dilution culture: growth. mainte-
nance, and characteristics of isolates under laboratory condi-
tions. Appl. Environ. Microbial. 59, 2150-2160.
Heijthuijsen, J.H.F.G. and Hansen, T.A. (1986) Interspecies
hydrogen transfer in co-cultures of methanol-utilizing acido-
gens and sulphate-reducing or methanogenic bacteria. FEMS
Microbial. Ecol. 38, 57-64.
Pfennig, N., Widdel, F.. Triiper, H.G. (198 1) The dissimila-
tory sulfate-reducing bacteria. In: The Prokaryotes. Vol. 1 (Starr, M.P., Stolp, H., TrUper, H.G.. Balows, A. and
Schlegel, H.G.. Eds.), pp. 926-940. Springer-Verlag. Heidel-
berg.
Schade, H.A.R. (1973) On the staining of glycogen for
electron microscopy with polyacids of tungsten and molybde-
num. I. Direct staining of sections of osmium fixed and Epon
embedded mouse liver with aqueous solutions of phospho-
tungstic acid (PTA). In: Electron Microscopy and Cytochem-
istry (Wisse, E., Daems, W.Th., Molenaar, I. and Van Duyn.
P., Eds.), pp. 263-266. Elsevier. North Holland, Amsterdam.
Lawson, P.A., Gharbia, S.E., Shah. H.N. and Clark, D.R.
(1989) Recognition of Fusobacterium rtucleaturn subgroups
Fn-1, Fn-2 and Fn-3 by ribosomal RNA gene restriction
patterns. FEMS Microbial. Len. 65, 41-46.
Hutson. R.A., Thompson, D.E. and Collins, M.D. (1993)
Genetic interrelationships of saccharolytic Clostridiurn bo-
rulinum types B. E and F and related clostridia as revealed by
small subunit rRNA sequences. FEMS Microbial. Lett. 108.
103-l 10.
Fukui, M. and Takii, S. (1989) Reduction of tetrazolium salta
by sulfate-reducing bacteria. FEMS Microbial. Ecol. 62,
13-20.
Widdel, F. (1980) Anaerober Abbau von Fettsluren und
Benzoesgure durch neu isolierte Arten Sulfat-reduzierender Bakterien. Ph.D. thesis. University of Gijttingen, Germany.
Nanninga, H.J. and Gottschal, J.C. (1985) Amino acid fer-
mentation and hydrogen transfer in mixed cultures. FEMS
Microbial. Ecol. 31, 261-269.
Laanbroek, H.J.. Geerlings, H.J., Sijtsma. L. and Veldkamp,
H. (1984) Competition for sulfate and ethanol among Desul-
fobacter, Desulfobulbus, and Desulforibrio species isolated from intertidal sediments. Appi. Environ. Microbial. 47.
329-334. Triiper. H.G. and Schlegel. H.G. (1964) Sulphur metabolism
in Thiorhodaceae, I. Quantitatieve measurements on growing cells of Chrormtium okenii. Antonie van Leeuwenhoek 30,
225-238. Richterich, R. (1965) Klinische Chemie. Akademische Ver-
laggesellschaft, Frankfurt.

E. W.J. mn Niel ei al./ FEMS Microbiology Ecology 21 (1996) 243-253 253
1291 Trevelyan, W.E. and Harrison, J.S. (1952) Studies on yeast
metabolism I. Fractionation and microdetermination of cell
carbohydrates. Biochem. J. 50, 298-310.
[30] Goa. J. (1953) A microbiuret method for protein determina-
tion: determination of total protein in cerebrospinal fluid.
Stand. J. Clin. Lab. Invest, 5, 218-222.
[31] Postgate, J.R. (1984) The Sulphate-reducing Bacteria. 2nd
edn., 208 pp. Cambridge University Press, Cambridge.
[32] Zellner. P., Messner, P.. Kneifel, H. and Winter. J. (1989)
Desuljovibrio simpkx spec. nov.. a new sulfate-reducing
bacterium from a sour whey digester. Arch. Microbial. 152, 329-334.
1331 Teske, A., Wawer, C., Muyzer, G. and Ramsing, N.B. (1996)
Distribution of sulfate-reducing bacteria in a stratified fjord
(Mariager Fjord, Denmark) as evaluated by most-probable
number counts and denaturing gradient gel electrophoresis of
PCR-amplified ribosomal DNA fragments. Appl. Environ.
Microbial. 62, 1405-1415.
[34] Kaplan, R. and Apirion, D. (1975) The fate of ribosomes in
Escherichia coli cells starved for a carbon source. J. Biol.
Chem. 250. 1854-1863.
[35] Fukui, M., Suwa, Y. and Urushigawa, Y. (1996) High sur-
vival efficiency and ribosomal RNA decaying pattern of
Desulfobacter latus. a highly specific acetate-utilizing organ-
ism, during starvation. FEMS Micribiol. Ecol. 19, 17-25.
[36] Chen L., Liu, M.-Y.. Legall, J., Pareleira, P., Santos. H. and
Xavier. V. (1993) Purification and characterization of an
NADH-rubredoxin oxidoreductase involved in the utilization
of oxygen by Desuffot?bn’o gigas. Eur. J. Biochem. 216.
443-448.
1371 Stams, A.J.M. and Hansen, T.A. (1982) Oxygen labile
L( + )-lactate dehydrogenase activity in DesulfoL?brio desul-
furicans. FEMS Microbial. Lett. 13, 389-394.
[38] LeGall, J. and Xavier. A.V. (1996) Anaerobes response to
oxygen: the sulfate-reducing bacteria. Anaerobe 2. l-9.
Related Documents