Short communication The MitoDrome database annotates and compares the OXPHOS nuclear genes of Drosophila melanogaster, Drosophila pseudoobscura and Anopheles gambiae Domenica D’Elia a, * , Domenico Catalano a , Flavio Licciulli a , Antonio Turi a , Gaetano Tripoli b , Damiano Porcelli b , Cecilia Saccone a,c , Corrado Caggese b a Institute for Biomedical Technologies, CNR, Via Amendola 122/D, 70126 Bari, Italy b University of Bari, Department of Genetics and Microbiology, Via Amendola 165/A, 70126 Bari, Italy c University of Bari, Department of Biochemistry and Molecular Biology, Via Orabona 4, 70126 Bari, Italy Received 28 March 2006; received in revised form 6 July 2006; accepted 13 July 2006 Available online 21 July 2006 Abstract The oxidative phosphorylation (OXPHOS) is the primary energy-producing process of all aerobic organisms and the only cellular function under the dual control of both the mitochondrial and the nuclear genomes. Functional characterization and evolutionary study of the OXPHOS system is of great importance for the understanding of many as yet unclear aspects of nucleus-mitochondrion genomic co-evolution and co-regulation gene networks. The MitoDrome database is a web-based database which provides genomic annotations about nuclear genes of Drosophila melanogaster encoding for mitochondrial proteins. Recently, MitoDrome has included a new section annotating genomic information about OXPHOS genes in Drosophila pseudoobscura and Anopheles gambiae and their comparative anal- ysis with their Drosophila melanogaster and human counterparts. The introduction of this new comparative annotation section into Mito- Drome is expected to be a useful resource for both functional and structural genomics related to the OXPHOS system. Ó 2006 Elsevier B.V. and Mitochondria Research Society. All rights reserved. Keywords: Oxidative phosphorylation; Drosophila; Database; Mitochondria; Nuclear genes 1. Introduction Oxidative phosphorylation (OXPHOS) takes place in the mitochondrion and is the main system through which a respiring cell is able to produce the energy necessary for its maintenance and reproduction (Saraste, 1999). It has been estimated that there are approximately one thou- sand different polypeptides in the mitochondrion (Lopez et al., 2000) to which, the mitochondrial genome contrib- utes only 13 structural polypeptides, all taking part in the assembling of the OXPHOS complexes. Thus, the OXPHOS system is the only mitochondrial component under the dual control of both the mitochondrial and the nuclear genomes and as such represents the ideal study approach to challenging questions about nucleus-mito- chondrion genomic co-regulation (Garesse and Vallejo, 2001) and co-evolution (Andersson et al., 2003). The study of OXPHOS genes is necessary to understand the overall design principles and robustness of the cell energy-machin- ery and of those cellular functions which depend on ener- getic metabolism and oxidative stress, such as programmed cell death (Green and Reed, 1998), aging and aging-related disease (Nicholls, 2002). The availability of the complete genomic sequence of a growing number of model organisms has prompted comparative genomic stud- ies. Some years ago we carried out a systematic study to identify the Drosophila melanogaster counterparts of human genes encoded by the nucleus and targeting the 1567-7249/$ - see front matter Ó 2006 Elsevier B.V. and Mitochondria Research Society. All rights reserved. doi:10.1016/j.mito.2006.07.001 * Corresponding author. Tel.: +39 080 5929674; fax: +39 080 5929690. E-mail address: [email protected] (D. D’Elia). www.elsevier.com/locate/mito Mitochondrion 6 (2006) 252–257

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.elsevier.com/locate/mito
Mitochondrion 6 (2006) 252–257
Short communication
The MitoDrome database annotates and compares the OXPHOSnuclear genes of Drosophila melanogaster, Drosophila
pseudoobscura and Anopheles gambiae
Domenica D’Elia a,*, Domenico Catalano a, Flavio Licciulli a, Antonio Turi a,Gaetano Tripoli b, Damiano Porcelli b, Cecilia Saccone a,c, Corrado Caggese b
a Institute for Biomedical Technologies, CNR, Via Amendola 122/D, 70126 Bari, Italyb University of Bari, Department of Genetics and Microbiology, Via Amendola 165/A, 70126 Bari, Italy
c University of Bari, Department of Biochemistry and Molecular Biology, Via Orabona 4, 70126 Bari, Italy
Received 28 March 2006; received in revised form 6 July 2006; accepted 13 July 2006Available online 21 July 2006
Abstract
The oxidative phosphorylation (OXPHOS) is the primary energy-producing process of all aerobic organisms and the only cellularfunction under the dual control of both the mitochondrial and the nuclear genomes. Functional characterization and evolutionary studyof the OXPHOS system is of great importance for the understanding of many as yet unclear aspects of nucleus-mitochondrion genomicco-evolution and co-regulation gene networks. The MitoDrome database is a web-based database which provides genomic annotationsabout nuclear genes of Drosophila melanogaster encoding for mitochondrial proteins. Recently, MitoDrome has included a new sectionannotating genomic information about OXPHOS genes in Drosophila pseudoobscura and Anopheles gambiae and their comparative anal-ysis with their Drosophila melanogaster and human counterparts. The introduction of this new comparative annotation section into Mito-Drome is expected to be a useful resource for both functional and structural genomics related to the OXPHOS system.� 2006 Elsevier B.V. and Mitochondria Research Society. All rights reserved.
Keywords: Oxidative phosphorylation; Drosophila; Database; Mitochondria; Nuclear genes
1. Introduction
Oxidative phosphorylation (OXPHOS) takes place inthe mitochondrion and is the main system through whicha respiring cell is able to produce the energy necessaryfor its maintenance and reproduction (Saraste, 1999). Ithas been estimated that there are approximately one thou-sand different polypeptides in the mitochondrion (Lopezet al., 2000) to which, the mitochondrial genome contrib-utes only 13 structural polypeptides, all taking part in theassembling of the OXPHOS complexes. Thus, theOXPHOS system is the only mitochondrial component
1567-7249/$ - see front matter � 2006 Elsevier B.V. and Mitochondria Resea
doi:10.1016/j.mito.2006.07.001
* Corresponding author. Tel.: +39 080 5929674; fax: +39 080 5929690.E-mail address: [email protected] (D. D’Elia).
under the dual control of both the mitochondrial and thenuclear genomes and as such represents the ideal studyapproach to challenging questions about nucleus-mito-chondrion genomic co-regulation (Garesse and Vallejo,2001) and co-evolution (Andersson et al., 2003). The studyof OXPHOS genes is necessary to understand the overalldesign principles and robustness of the cell energy-machin-ery and of those cellular functions which depend on ener-getic metabolism and oxidative stress, such asprogrammed cell death (Green and Reed, 1998), agingand aging-related disease (Nicholls, 2002). The availabilityof the complete genomic sequence of a growing number ofmodel organisms has prompted comparative genomic stud-ies. Some years ago we carried out a systematic study toidentify the Drosophila melanogaster counterparts ofhuman genes encoded by the nucleus and targeting the
rch Society. All rights reserved.

D. D’Elia et al. / Mitochondrion 6 (2006) 252–257 253
mitochondrion. We systematically compared these humanprotein sequences, annotated in the SwissProt database,with the Drosophila genomic sequences, ESTs and cDNAdata available in the FlyBase database (FlyBase Consor-tium, 2003). This study involved the annotation of 278Drosophila genes, assumed to encode mitochondrial pro-teins, which have been collected and made publicly avail-able through the MitoDrome database (Sardiello et al.,2003). The recent availability of the genome sequence ofDrosophila pseudoobscura (Baylor College of Medicine:Drosophila Genome Project: http://www.hgsc.bcm.tmc.edu/projects/drosophila) and Anopheles gambiae (Holtet al., 2002), along with the D. melanogaster data drawnfrom our previous study, has allowed us to apply our expe-rience in the annotation of these genomes to the genes ofthe OXPHOS system (Tripoli et al., 2005). Structural andfunctional annotations of the predicted OXPHOS genesalong with the results of their comparative analyses havebeen recently annotated in the MitoDrome database underthe section ‘‘OXPHOS system’’. Information provided are:gene, transcript and protein sequences, upstream anddownstream sequences from the gene boundaries, geneexon/intron structural organization, alternative splicedproducts and amino acid sequence alignments; annotationof gene duplicates along with intra- and inter-species com-parison of members of each gene family; P-insertionmutant alleles of OXPHOS genes in D. melanogaster. Inthe present paper we describe the MitoDrome ‘‘OXPHOSsystem’’ section, the current MitoDrome content and itsnew graphical interface and usefulness.
2. Materials and methods
2.1. Genomic annotation and data sources
The strategy used for OXPHOS genome annotation hasalready been extensively described in a recent publication(Tripoli et al., 2005), here we only report it briefly to givean overview of the accuracy of the MitoDrome annota-tions. To identify the nuclear encoded OXPHOS genes inthe genomes of D. pseudoobscura and A. gambiae all theOXPHOS proteins, previously predicted in D. melanogas-
ter (Sardiello et al., 2003), were extracted from the Mito-Drome database and used for a similarity BLAST search(McGinnis and Madden, 2004) (tBLASTn option) againstthe draft genomic sequences and the available EST collec-tions of D. pseudoobscura and A. gambiae. The main crite-ria used to assign gene orthology were protein sequencehomology as well as conservation of gene structural orga-nization and of microsyntenic order. The validation ofthe BLAST based predictions with regard to exon/intronboundaries, splice sites, start/end codons, and UTRregions was obtained through the manual processing ofthe BLAST outputs and comparison with EST sequences(when available) and predicted CDSs. Multiple alignmentsof the amino acid and CDS sequences were obtained usingthe Multialign software (Corpet, 1988). Genome searching
for OXPHOS gene copies was carried out using the proteinsequence of each OXPHOS orthologous gene as a probe inthe investigated species. Only the reciprocal BLAST hitswith an E-value less than 10�20 in both directions andallowing amino acid sequence alignment along at least60% of their length were assumed to be putative duplicatedgenes. The additional criteria used were that the putativeprocessed proteins encoded had to have more than 40%identity and the amino-acid percentage similarity had tobe greater than the percentage identity. A list of D. melano-
gaster P-insertion mutant alleles of the OXPHOS geneswas compiled using information available from FlyBase(The FlyBase Consortium, 1999) and from the BDGP P-Element Gene Disruption Project (Spradling et al., 1999).Each of the P-element insertions was precisely localizedwithin the gene sequence by a BLASTN alignment of eachinsertion allele with the corresponding wild-type sequence.
2.2. Construction and content of the MitoDrome ‘‘OXPHOS
system’’ section
In order to integrate the new data produced and facili-tate their retrieval we have added a new section to theMitoDrome database, the ‘‘OXPHOS system’’ section,which has been implemented in a MySQL relational sche-ma. Data have been organized using four main criteria:species, protein function, gene homology (ortholog/paral-og clusters) and D. melanogaster P-insertion mutants. In
silico predictions have been integrated with informationabout genomic sources (e.g. scaffolds and/or contigs) usedfor gene assembly and with the product description fromthe Swiss Prot database (Bairoch and Boeckmann, 1991).Web links to related human entries in the Swiss Prot andGeneCards (Safran et al., 2002) databases and to the GOdatabase (Harris et al., 2004) have also been included. Datasubmission and updating/revision are carried out manuallyby means of an easy-to-use web-based interface developedusing the PHP Seagull Framework (http://seagull.php-kitchen.com/). This interface is accessible only by autho-rized individuals and the user profile was designed toselect the access to this database layer under usernameand password restrictions.
3. Results
3.1. Database content
Currently the database consists of two major sections,namely the ‘‘DROME LIST’’, and the ‘‘OXPHOS system’’section. The ‘‘DROME LIST’’ contains the annotations of285 D. melanogaster nuclear genes covering all mitochon-drial components as they were annotated in the first Mito-Drome release. In the ‘‘OXPHOS system’’ sectionMitoDrome annotates 78 OXPHOS orthologous genes ineach species investigated and 47 duplicates (20 in D. mela-
nogaster, 19 in D. pseudoobscura and eight in A. gambiae).The OXPHOS genes include 66 proteins known to be

254 D. D’Elia et al. / Mitochondrion 6 (2006) 252–257
components of the five respiratory complexes and 12 pro-teins involved in oxidative phosphorylation as accessoryproteins. For each of the identified genes the followingpre-computed and manual analyses are available:
I. gene sequence and structure,II. gene splice-site variants,III. structure and sequence of expressed transcript,IV. prediction of protein sequence,V. comparison (Clusters) of the gene structure (includ-ing splice-site variants, when present), protein sequencealignments (including the human counterpart) and phy-logenetic tree of the predicted orthologous- and paralo-gous-OXPHOS genes in the three species (D.melanogaster, D. pseudoobscura and A. gambiae),VI. annotations of 43 P-insertion mutant alleles ofOXPHOS D. melanogaster genes, grouped on the basisof their functional classification.
3.2. Database home page
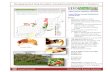
A new MitoDrome web-interface has been designed toallow easy consultation and retrieval of stored information,access to clustering data on orthologs and paralogs and theextraction of bio-sequences for any in silico analysis theusers wish to perform. The new web version of the Mito-Drome database is publicly available at the followingaddress: http://www2.ba.itb.cnr.it/MitoDrome/. In theupper part of the home page (Fig. 1) a navigation toolbarprovides direct access to the complete list of D. melanogas-
ter genes (DROME LIST), to the OXPHOS gene families(CLUSTER) and to the OXPHOS search and Blast pages(‘‘SEARCH’’ and ‘‘BLAST’’ buttons). A lateral frame,
Fig. 1. MitoDrom
on the left-hand side, provides access to the ‘‘OXPHOSsystem’’ section through pre-compiled queries of OXPHOSannotations and of P-insertion mutant alleles of OXPHOSgenes in D. melanogaster.
3.3. D. melanogaster gene collection and data retrieval
The D. melanogaster gene collection has conserved itsoriginal structure; data are organized in an ‘‘EMBL like’’flatfile format and can be retrieved through both the‘‘DROME LIST’’ page from the MitoDrome home pageand through SRS at our server (http://www.ba.itb.cnr.it/srs7/) under the section ‘‘Mitochondrial Databases’’. The‘‘DROME LIST’’ page provides the user with two differentaccesses, one to the table including all the genes alphabet-ically ordered according to the protein name, and onewhich allows the genes to be retrieved on the basis of theirfunctional classification. The ‘‘DROME LIST’’ page alsolets users download both the flatfile of the D. melanogaster
gene collection and the gene list.
3.4. OXPHOS system annotation format and availability
The annotations of OXPHOS genes, transcripts andproteins; clustering of orthologous- and paralogous-OXPHOS gene families and of the D. melanogaster P-inser-tion mutant alleles of OXPHOS genes are organized indatabase entries displayed as html pages. Each gene entryorganizes and integrates biological and molecular informa-tion on the gene and on its products, i.e. mRNA(s), CDS(s)and predicted translated product(s). Gene informationincludes the gene name, cytogenetic localization, and geno-mic source sequences from which the gene sequence andgene structure have been deduced, as well as contig or
e home page.

D. D’Elia et al. / Mitochondrion 6 (2006) 252–257 255
scaffold nucleotide coordinates. The gene sequence is partof the labelled ‘‘REFERENCE SEQUENCE’’ which,besides the gene sequence, includes stretches of the geneflanking regions (up to about 1000 bp upstream and down-stream of the first and last exon, respectively). When theprediction of alternative spliced products is available theseare indicated on the gene map and annotated as ‘‘AltForm’’ consecutively numbered in the CDS section of theentry. A colour code allows the easy localization of genefunctional elements. UTRs and exon sequences are shownin blue and red, respectively in the mRNA and in the‘‘REFERENCE SEQUENCE’’, while the remainingregions are shown in black. In addition, the polyadenyla-tion target site, when identified, is underlined in the ‘‘REF-ERENCE SEQUENCE’’ and indicated in the mRNAentry section. P-insertion mutant alleles in D. melanogaster
genes, when available, are visualized on the gene map attheir sequence position, as determined by the BLAST-Nanalysis, and reported in the relevant row of the ‘‘REFER-ENCE SEQUENCE’’ entry section, where a link to theexternal resource is also provided. Hyperlinks are providedinside the database entry to allow: (1) navigation from the‘‘REFERENCE SEQUENCE’’ frame to the sequence itself
Fig. 2. An example of ‘‘OXPHOS gene Cluster’’ entry. The figure shows the Cpseudoobscura indicating that duplication has occurred after Drosophila/Anoph
user can access the alignment of Cluster members including the human cousequence’’ button.
and vice versa, (2) dynamic connection to the associatedCluster of gene homologs in the three species of Dipteraand (3) direct access to a sequence export page for theextraction and export of genomic, mRNA, CDS and pro-tein sequences and sub-sequences.
3.5. OXPHOS gene Cluster annotation format and
availability
Data on clustering and comparison of the orthologous-and paralogous-OXPHOS gene families is organized inentries reporting the following information: the name ofthe protein, as given in the protein-name line of the SwissProt human entry, the SwissProt accession number (AC)and GeneCards database ID of the human protein, theprotein sequence alignment, the graphical representationof the genes maps (with the conserved exon/intronjunctions indicated by vertical dashed lines), the evolution-ary tree derived from the analysis of the protein sequencesalignment carried out using the ProtML program (http://ftp.cse.sc.edu/bioinformatics/molphy) and a clickable listof the OXPHOS entries belonging to the Cluster. An exam-ple of a Cluster entry is shown in Fig. 2.
OXB gene Cluster. Duplicates are present only in D. melanogaster and D.
eles divergence. Clicking on the ‘‘Protein sequence alignment’’ button thenterpart. Cluster sequences can be exported by clicking on the ‘‘Extract

256 D. D’Elia et al. / Mitochondrion 6 (2006) 252–257
3.6. How to browse and query OXPHOS system annotations
For browsing and querying the MitoDrome OXPHOSannotations the user can choose from three differentoptions either using pre-compiled queries, or using theadvanced Search form and the BLAST tool. The pre-com-piled queries, accessible through hyperlinks on the left ofthe database homepage, allow searches on the basis of gen-eral criteria such as organism, cluster of homologous genes,and availability of P-Insertion mutant alleles for genes ofD. melanogaster. The query result page returns the list ofentries matching the query criteria along with some keyinformation, which is differently organized depending onthe selected dataset. For example, the result page obtainedfor a ‘‘species’’ query gives a sortable entries list providingkey information including, organism name, MitoDromeentry name, gene name and its chromosomal location, pro-tein description and the hyperlink to the gene entry pageand the homology cluster entry to which the gene belongs.By selecting ‘‘Clusters’’ the user is presented with a sortabletable reporting the list of OXPHOS Clusters annotatedinto the database along with additional information suchas the SwissProt AC and GeneCard ID of the humanhomolog and the corresponding protein function. All thetable rows are hyperlinked to the relevant Cluster entry.In the case of a query for D. melanogaster ‘‘P-insertionmutant alleles’’ of OXPHOS genes, the system provides atable in which the genes are grouped on the basis of theirfunctional classification and the number of mutant allelesfor each functional category is indicated. As soon as afunctional category is chosen, the list of genes in that cate-gory is displayed along with the gene and protein name,cytogenetic map location and hyperlinks to the gene entryand to the mutant allele web resource. Through theadvanced Search form the user can perform more complexdatabase searches using the combination of different crite-ria such as gene name, chromosome location, protein func-tional classification and/or mitochondrial location, andapplying them selectively to OXPHOS genes from any ofthe organisms annotated, or to all of them.
3.7. Sequence export tool
From the query result page the user can access the geneentry, save the query result in a text and MS Excel readableformat and/or extract and export any type of sequence (i.e.,gene, upstream and downstream sequences from the geneboundaries, exon, intron, UTRs, transcript, CDS, protein)from selected entries or from all of the entries retrieved, byusing the sequence export page. This page is also accessiblefrom the Cluster entry page where it enables users to extractsequences from all the genes belonging to the same Cluster.
3.8. BLAST tool
The MitoDrome also provides an interface to an in-house BLAST server which enables users to search the
OXPHOS section of the database using a partial or anentire gene, transcript or protein sequence. This is especial-ly useful in database searches to elucidate the nature ofsequences of unknown function or to identify homologydomains and conserved features shared by differentsequences.
4. Discussion
The main concept of the MitoDrome database is to usethe Drosophila as a model to provide data that can be usefulfor the investigation of issues related to the cell energymetabolism and its pathological dysfunctions in human.The recent developments described in the present paper,expand this concept to a comparative approach directed atthe genome annotation of two dipteran species with signifi-cantly different evolutionary divergence times from D. mela-
nogaster, the D. pseudoobscura and the A. gambiae, andfocused on the genes of the ‘‘OXPHOS system’’. The objec-tive of this new release of the MitoDrome database has itsroots in emerging evidence from comparative studies. Thecomparative analysis of differently related species has beenproved to be the most effective instrument for functionalannotation of both coding and cis-regulatory sequencesand for the investigation of the evolutionary basis of cellularand developmental processes (Bergman et al., 2002). Com-parative analysis of the exon/intron organization of genesand their resulting protein structures is important to under-stand evolutionary relationships between different species,the rules of protein organization, and protein functionality.Furthermore, one of the most important aspects of genomeannotation is the evaluation of gene duplication and theanalysis of paralogous gene families. In this context, thenew MitoDrome annotations are expected to provide oppor-tunities for the evolutionary and functional characterizationof the OXPHOS system. These studies could provideinsights into the genetic regulatory mechanisms controllingenergy genes (Sardiello et al., 2005), the genetic and pheno-typic variation responsible for mitochondrial pathogenesis,genetic adaptation and speciation. In addition, they areexpected to constitute a state of the art resource which couldsignificantly facilitate OXPHOS genome annotation at leastfor species belonging to the Drosophila genus. In silico anal-ysis of the OXPHOS system is also expected to guide exper-imental research and contribute to revealing gene functionalinteractions related to mitochondrial function. To deal withthese challenging issues the MitoDrome database contrib-utes two important innovations. The ‘‘OXPHOS system’’annotations contain a wide number of analytical featureswhich are the result of targeted analyses. The annotationof ‘‘OXPHOS system’’ genes has been powered by humanintervention in the validation and integration of data deriv-ing from analyses of sequence homology, structural orthol-ogy (conservation of the number of introns, their locationin the coding sequence and conservation of the readingframe with respect to the flanking exons), spatial organiza-tion of genes and the presence of duplicates. The new data

D. D’Elia et al. / Mitochondrion 6 (2006) 252–257 257
structure facilitates data query and retrieval and provides aneasy-to-use tool to extract and export any type of sequence indifferent file formats.
Acknowledgments
This work has been supported by the Ministero dell’Ist-ruzione, dell’Universita e della Ricerca, Italy (projects:FIRB ‘‘Bioinformatica per la Genomica e la Proteomica’’,PON 2000-2006 ‘‘Project BIG’’, Cluster C03 – Progetto 2L. 488/92 and Contributi Straordinari del D.D. n.1105).
References
Andersson, S.G., Karlberg, O., Canback, B., Kurland, C.G., 2003. On theorigin of mitochondria: a genomics perspective. Philos. Trans. R. Soc.London B. Biol. Sci. 358, 165–179, Review.
Bairoch, A., Boeckmann, B., 1991. The SWISS-PROT protein sequencedata bank. Nucleic Acids Res. 19 (Suppl), 2247–2249.
Bergman, C.M., Pfeiffer, B.D., Rincon-Limas, D.E., Hoskins, R.A.,Gnirke, A., Mungall, C.J., Wang, A.M., Kronmiller, B., Pacleb, J.,Park, S., Stapleton, M., Wan, K., George, R.A., de Jong, P.J., Botas,J., Rubin, G.M., Celniker, S.E., 2002. Assessing the impact ofcomparative genomic sequence data on the functional annotation ofthe Drosophila genome. Genome Biol., 3 RESEARCH0086. Epub2002 Dec 30.
Corpet, F., 1988. Multiple sequence alignment with hierarchical clustering.Nucleic Acids Res. 16, 10881–10890.
FlyBase Consortium, 2003. The FlyBase database of the Drosophilagenome projects and community literature. Nucleic Acids Res., 31,172–175.
Garesse, R., Vallejo, C.G., 2001. Animal mitochondrial biogenesis andfunction: a regulatory cross-talk between two genomes. Gene 263, 1–16, Review.
Green, D.R., Reed, J.C., 1998. Mitochondria and apoptosis. Science 281,1309–1312.
Harris, M.A., Clark, J., Ireland, A., Lomax, J., Ashburner, M., Foulger,R., Eilbeck, K., Lewis, S., Marshall, B., Mungall, C., et al., 2004. TheGene Ontology (GO) database and informatics resource. NucleicAcids Res. 32 (Database issue), D258–D261.
Holt, R.A., Subramanian, G.M., Halpern, A., Sutton, G.G., Charlab, R.,Nusskern, D.R., Wincker, P., Clark, A.G., Ribeiro, J.M., Wides, R.,et al., 2002. The genome sequence of the malaria mosquito Anopheles
gambiae. Science 298, 129–149.Lopez, M.F., Kristal, B.S., Chernokalskaya, E., Lazarev, A., Shestopalov,
A.I., Bogdanova, A., Robinson, M., 2000. High-throughput profilingof the mitochondrial proteome using affinity fractionation andautomation. Electrophoresis 21, 3427–3440.
McGinnis, S., Madden, T.L., 2004. BLAST: at the core of a powerful anddiverse set of sequence analysis tools. Nucleic Acids Res. 32 (WebServer issue), W20–W25.
Nicholls, D.G., 2002. Mitochondrial function and dysfunction in the cell:its relevance to aging and aging-related disease. Int. J. Biochem. Cell.Biol. 34, 1372–1381, Review.
Safran, M., Solomon, I., Shmueli, O., Lapidot, M., Shen-Orr, S., Adato,A., Ben-Dor, U., Esterman, N., Rosen, N., Peter, I., Olender, T.,Chalifa-Caspi, V., Lancet, D., 2002. GeneCards 2002: towards acomplete, object-oriented, human gene compendium. Bioinformatics18, 1542–1543.
Saraste, M., 1999. Oxidative phosphorylation at the fin de siecle. Science5, 1488–1493, Review.
Sardiello, M., Licciulli, F., Catalano, D., Attimonelli, M., Maggese, C.,2003. MitoDrome: a database of Drosophila melanogaster nucleargenes encoding proteins targeted at the mitochondrion. Nucleic AcidsRes. 31, 322–324.
Sardiello, M., Tripoli, G., Romito, A., Minervini, C., Viggiano, L.,Caggese, C., Pesole, G., 2005. Energy biogenesis: one key forcoordinating two genomes. Trends Genet. 21, 12–16.
Spradling, A.C., Stern, D., Beaton, A., Rhem, E.J., Laverty, T., Mozden,N., Misra, S., Rubin, G.M., 1999. The Berkeley Drosophila GenomeProject gene disruption project: Single P-element insertions mutating25% of vital Drosophila genes. Genetics 153, 135–177.
Tripoli, G., D’Elia, D., Barsanti, P., Caggese, C., 2005. Comparison of theoxidative phosphorylation (OXPHOS) nuclear genes in the genomes ofDrosophila melanogaster, Drosophila pseudoobscura and Anopheles
gambiae. Genome Biol. 6, R11.
Related Documents