EARLY EMBRYOLOGY, FATE DETERMINATION, AND PATTERNING IN DROSOPHILA NEELAM DEVPURA (M.Sc. ,NET) CSIR-UGC JRF Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

EARLY EMBRYOLOGY, FATEDETERMINATION, AND
PATTERNING IN DROSOPHILA
NEELAM DEVPURA (M.Sc. ,NET)CSIR-UGC JRF
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15
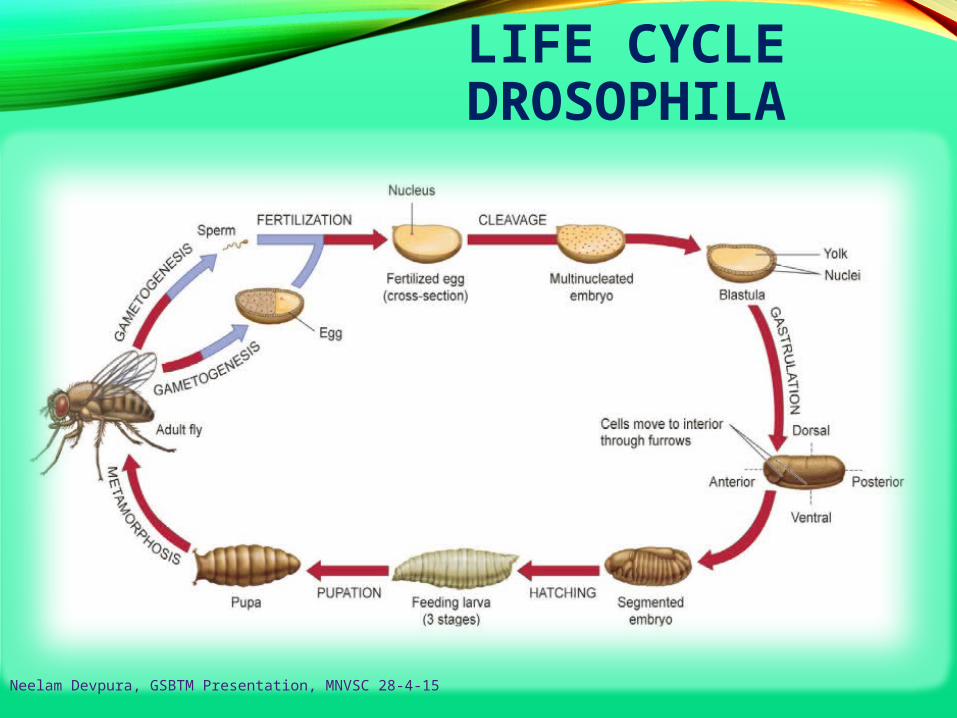
LIFE CYCLE DROSOPHILA
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

DROSOPHILA LIFE CYCLE
• Life cycle by daysDay 0: Female lays eggs
Day 1: Eggs hatch
Day 2: First instar (one day in length)
Day 3: Second instar (one day in length)
Day 5: Third and final instar (two days in length)
Day 7: Larvae begin roaming stage. Pupariation (pupa
formation) occurs 120 hours after egg laying
Day 11-12: Eclosion (adults emerge from the pupa case).
Females become sexually mature 8-10 hours after
eclosion
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15
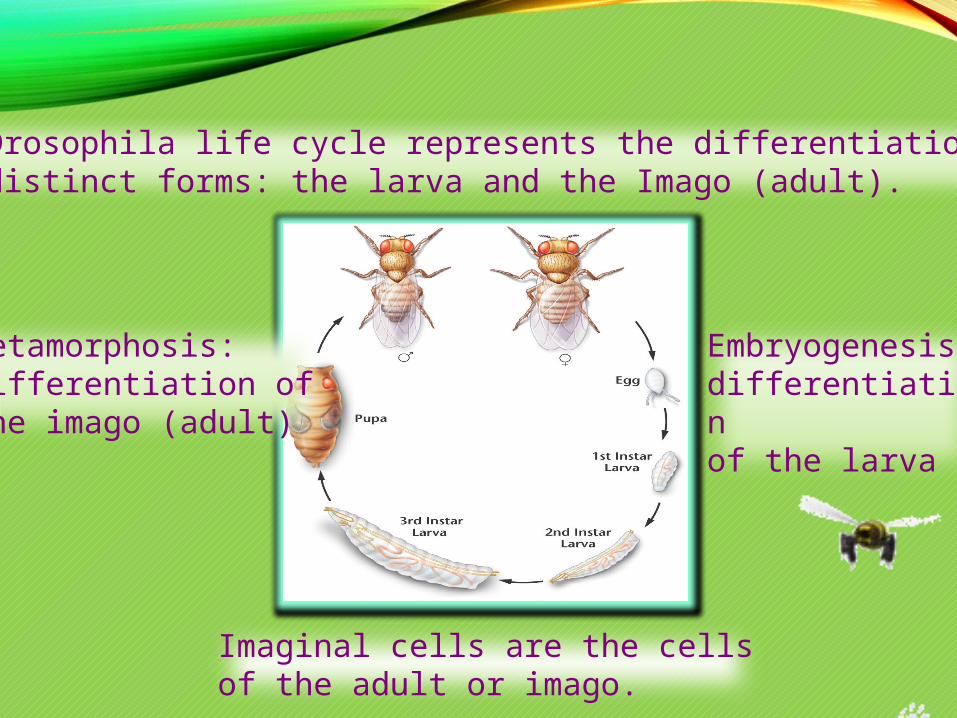
The Drosophila life cycle represents the differentiation of two distinct forms: the larva and the Imago (adult).
Embryogenesis: differentiationof the larva
Metamorphosis:differentiation ofthe imago (adult)
Imaginal cells are the cellsof the adult or imago.

DROSOPHILA DEVELOPMENT
¤ Embryonic development in Drosophila is an orderly sequence of
change and is controlled by the differential expression of genes.
¤ Drosophila display a holometabolous method of development,
meaning that they have three distinct stages of their post –
embryonic life cycle, each with radically different body plans:
larva, pupa and finally, adult(imago
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

Neelam Devpura, GSBTM Presentation, MNVSC
DROSOPHILA DEVELOPMENT - OVERVIEW
Cleavage
Fertilization
Gastrulation Drosophila body plan
Oocyte formation Genetic control of axis specification
Anterior-posterior Dorsal-ventral
Segmentation genes Homeotic genes
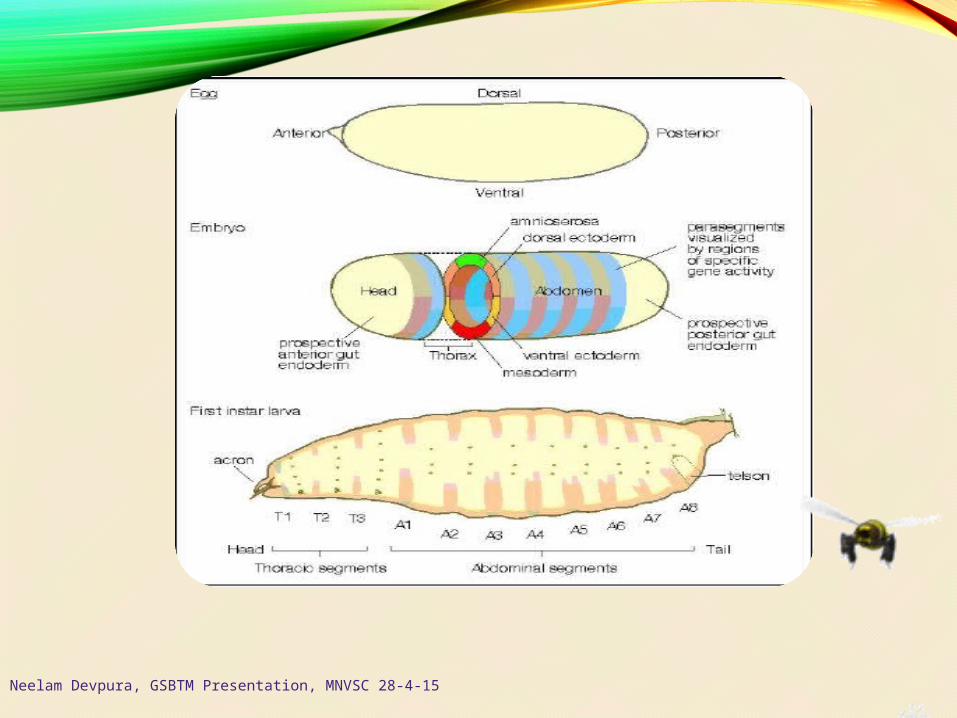
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15
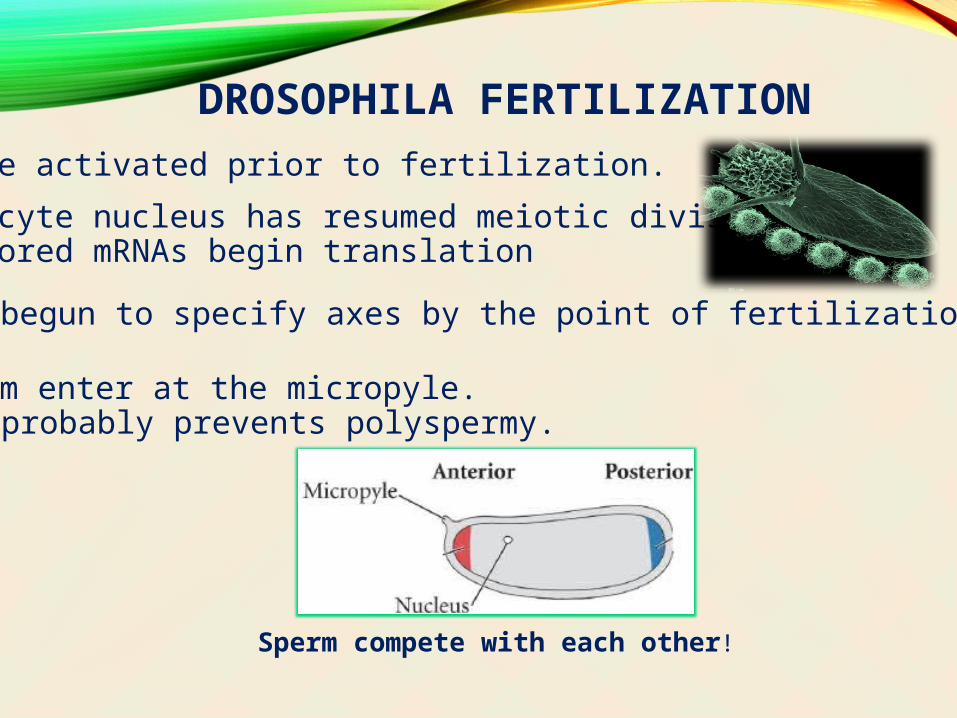
DROSOPHILA FERTILIZATIONEggs are activated prior to fertilization.
- oocyte nucleus has resumed meiotic division- stored mRNAs begin translation
Eggs have begun to specify axes by the point of fertilization.
Sperm enter at the micropyle.- probably prevents polyspermy.
Sperm compete with each other!

EARLY DEVELOPMENT OF DROSOPHILA
• Egg is centrolecithal
• After fertilization, series of superficial cleavages
• Blastoderm is syncytial until 13th cleavage (256 nuclei!)
• Nuclei begin dividing centrally, migrate toward the edges
• Several nuclei migrate to posterior end, form cell membranes (pole cells)
• Give rise to the adult gametes• What cells are like this in mammals?
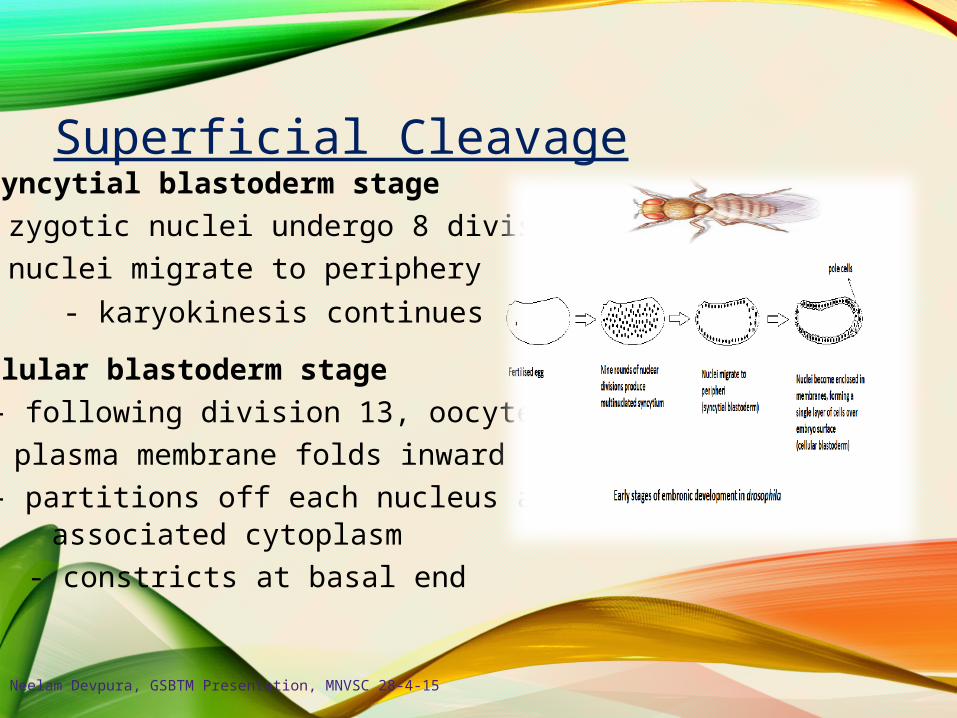
Superficial CleavageSyncytial blastoderm stage
- zygotic nuclei undergo 8 divisions
- nuclei migrate to periphery
- karyokinesis continues
Cellular blastoderm stage
- following division 13, oocyte
plasma membrane folds inward
- partitions off each nucleus andassociated cytoplasm
- constricts at basal end
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15
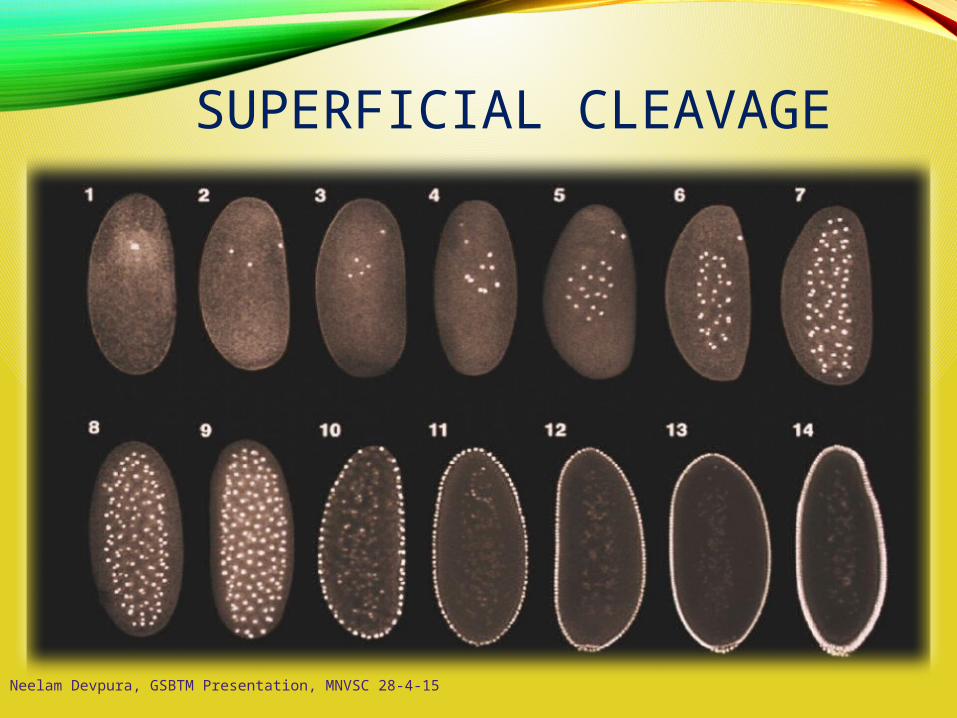
SUPERFICIAL CLEAVAGE
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

Although nuclei share the same cytoplasm, the cytoplasm is not
uniform in its makeup
• Maternal molecules are distributed differently
Eventually cells will form plasma membranes and the embryo
will consist of a cellular blastoderm
Mid-blastula transition occurs slowly, increasing transcription of
zygotic genes
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

GASTRULATION
• At MBT, gastrulation begins, forming mesoderm, endoderm, ectoderm
• Cells fold inward to form ventral furrow
• Embryo bends to from cephalic furrow
• Pole cells are internalized, endoderm invaginates
• Ectoderm converges and extends along midline to form Germ Band
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

GERM BAND
• Wraps around the embryo
• As it wraps around the dorsal surface, the A-P
axis of the embryo is laid down
• Body segments begin to form
• At the end of germ band extension
• Organs are beginning to form
• Body segmentation is set-up
• Groups of cells called imaginal discs are set
aside, these cells will form adult structuresNeelam Devpura, GSBTM Presentation, MNVSC 28-4-15

DROSOPHILA LARVAE
• During metamorphosis
• 3 “instar” larvae
• Pupae
• Adult
• After gastrulation, 1st instar larvae is formed
• Has head and tail end
• Repeating segments along axis
• Generally the same type of body plan as adult
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

DROSOPHILA BODY PLAN
• 3 thoracic segments
• Each different from each other
• 8 abdominal segments
• Each different from each other
• Able to tell the difference in the larvae based on
cuticle
• Covering of the embryo
• Correspond to the adult segmentsNeelam Devpura, GSBTM Presentation, MNVSC 28-4-15

Segments form along the anterior-posterior axis, then become specialized.
Specification of tissues depends on their position along the primary
axes.
A/P and D/V axes established by interactions between the developingoocyte and its surrounding follicle cells
T1- legs
T2 – legs & wings
T3 – legs & halteres
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

GENETICS OF AXIS SPECIFICATION IN
DROSOPHILA
• Controlled by a variety of genes
• Maternal effect genes
• Gap genes
• Pair-rule genes
• Segment polarity genes
• Homeotic selector genes
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

Anterior-Posterior Body PlanDrosophila use a hierarchy of gene expression to
establish the anterior-posterior body plan.
2. Gap genes: first zygotic genes expressedDivide embryo into regions- expressed in broad, partially
overlapping domains (~ 3 segments wide)- code for transcription factors; transient
-activated or repressed by maternal effect genes-
Hunchbackoverlap
bicoid gradient
Kruppel
1. Maternal effect genes (e.g. bicoid, nanos)Establish polarity:- mRNAs differentially placed in eggs- transcriptional or translational
regulatory proteins; transient- diffuse through syncytial cytoplasm
- activate or repress zygotic genes
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

- even-skipped (red),fuschi tarazu (black)
engrailed
3. Pair-rule genes;Establish segmental plan- regulated by combinations of gap genes- code for transcription factors; transient- divide the embryo into periodic units
- pattern of seven transverse bands delimit parasegments4. Segment polarity genes;
Set boundaries of segments(i.e. establish A-P for each segment)- activated by pair-rule genes- code for variety of proteins; stable- divide embryo into 14 segmental units
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

-
5. Homeotic selector genes;Provide segmental identity- interactions of gap, pair-rule,
and segment polarity proteins- determines developmental fat
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

Segmentation Genes
Cell fate commitment:Phase 1 – specificationPhase 2 – determination
- early in development cell fate depends on interactionsamong protein gradients
- specification is flexible; it can alter in response to signalsfrom other cells
- eventually cells undergo transition from loose commitmentto irreversible determination
The transition from specification to determination in Drosophila ismediated by the segmentation genes.
- these divide the early embryo into a repeating series of segmentalprimordia along the anterior-posterior axis
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15
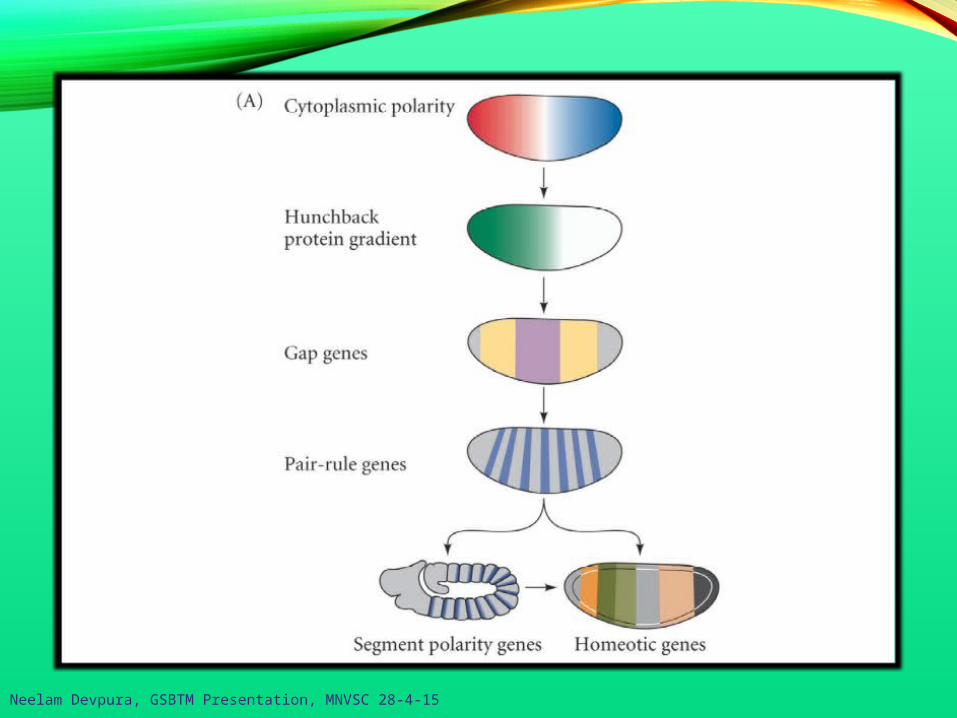
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15
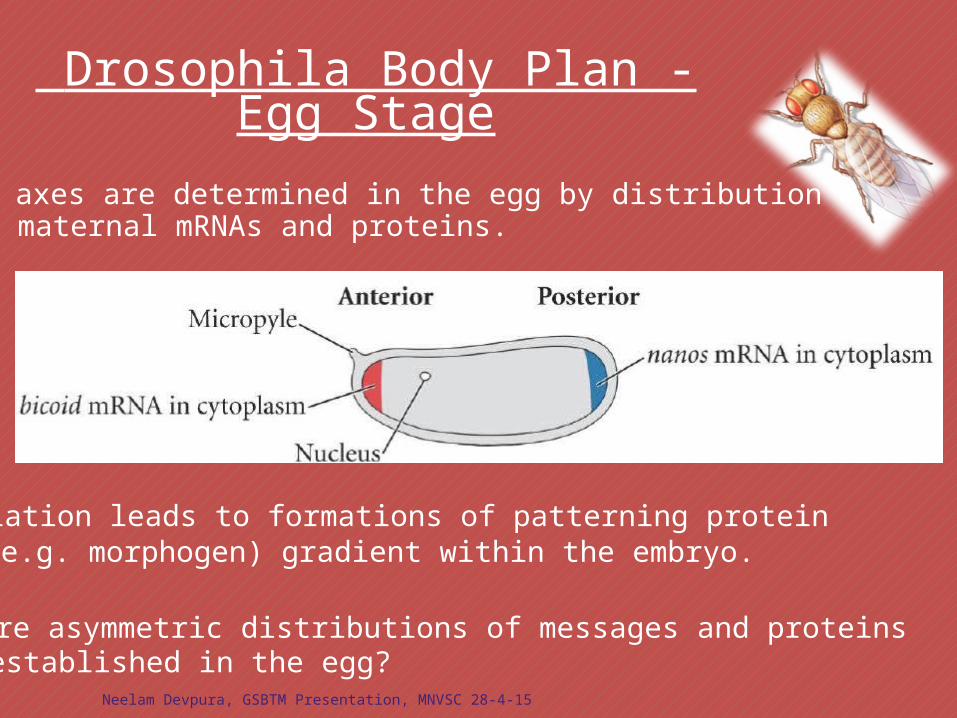
Drosophila Body Plan - Egg Stage
Translation leads to formations of patterning protein(e.g. morphogen) gradient within the embryo.
Body axes are determined in the egg by distribution ofmaternal mRNAs and proteins.
How are asymmetric distributions of messages and proteinsestablished in the egg?
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15
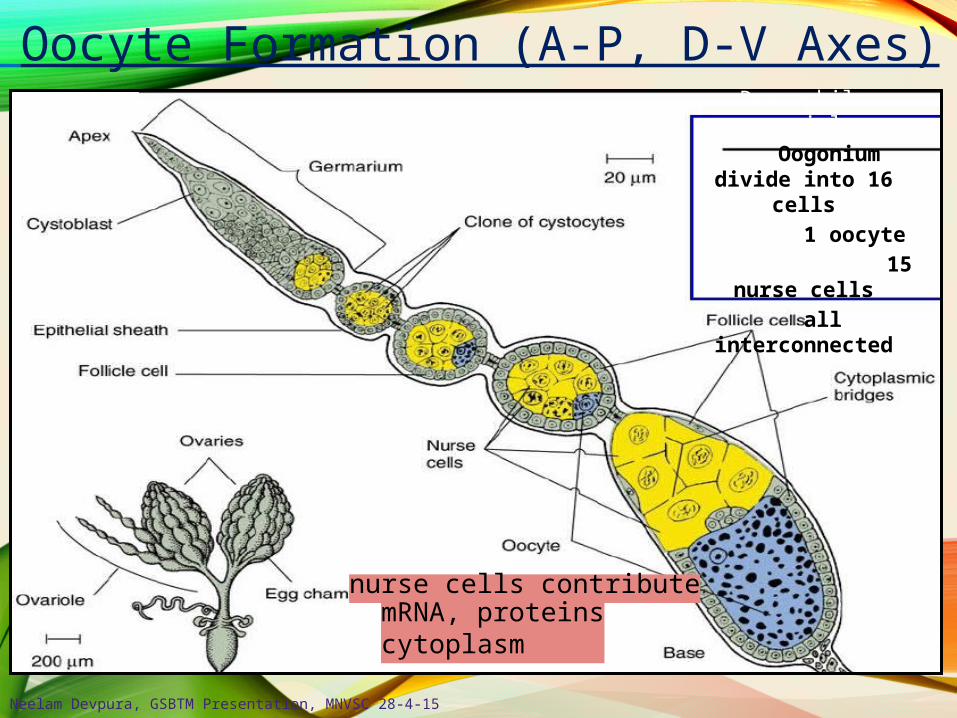
Oocyte Formation (A-P, D-V Axes)
nurse cells contributemRNA, proteinscytoplasm
Drosophila ovariole Oogonium divide
into 16 cells 1 oocyte
15 nurse cells all
interconnected
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

Anterior-Posterior Axis Formation
Torpedo (RTK Gurken receptor) present on follicular cells
Gurken binding results in “posteriorization” of follicles- posteriorized follicles re-organize egg microtubules; (-) = anterior
Gurken protein localized between nucleus and cell membrane- Note – Gurken diffuses only a short distance
Nurse cells synthesize gurken (TGF-β family)- gurken mRNA transported toward oocyte nucleus (in posterior region)

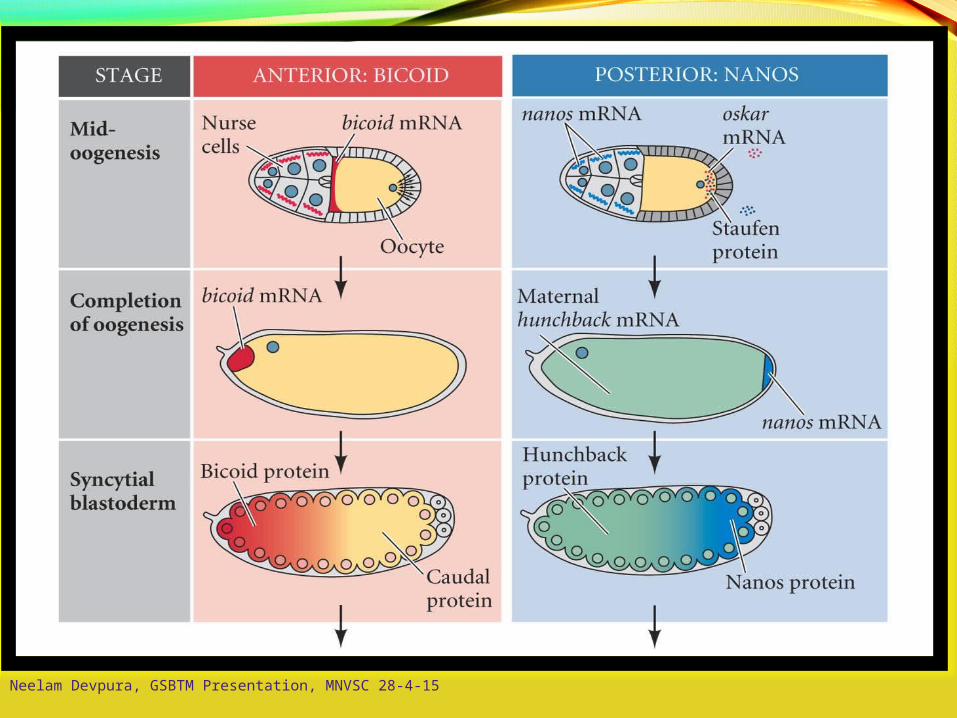
A-P Axis: bicoid / Oskar / nanos
Nurse cells manufacturebicoid and nanos mRNA- deliver cytoplasminto oocyte
bicoid binds to dynein- moves to non-growing
(-) end of microtubules
oscar mRNA formscomplex with kinesin I- moves toward growing
(+) end of microtubules
Oskar binds nanos mRNA- retains nanos in
posterior end
“posteriorized” folliclesproduce organized (+/-)microtubules
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

Posterior Specification - 1
Nanos preventshunchback translation
Oskar binds nanos;remains in posterior
At least 9 maternal genes make up the posterior patterning group; including
nanos trap:
Staufen allowsoskar translation
staufen, oskar, nanos
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

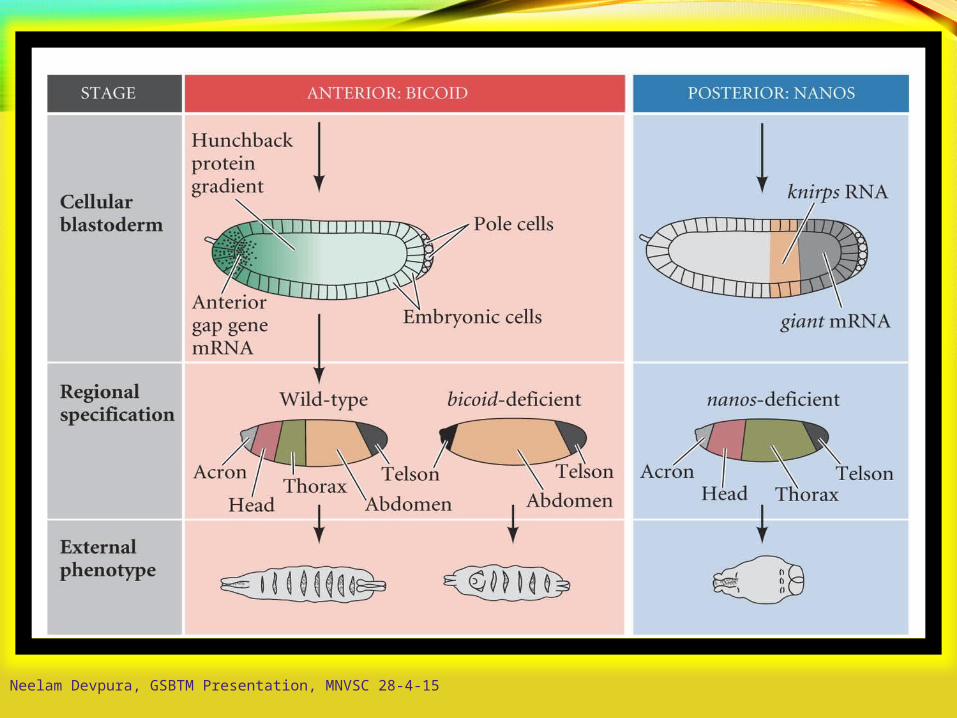
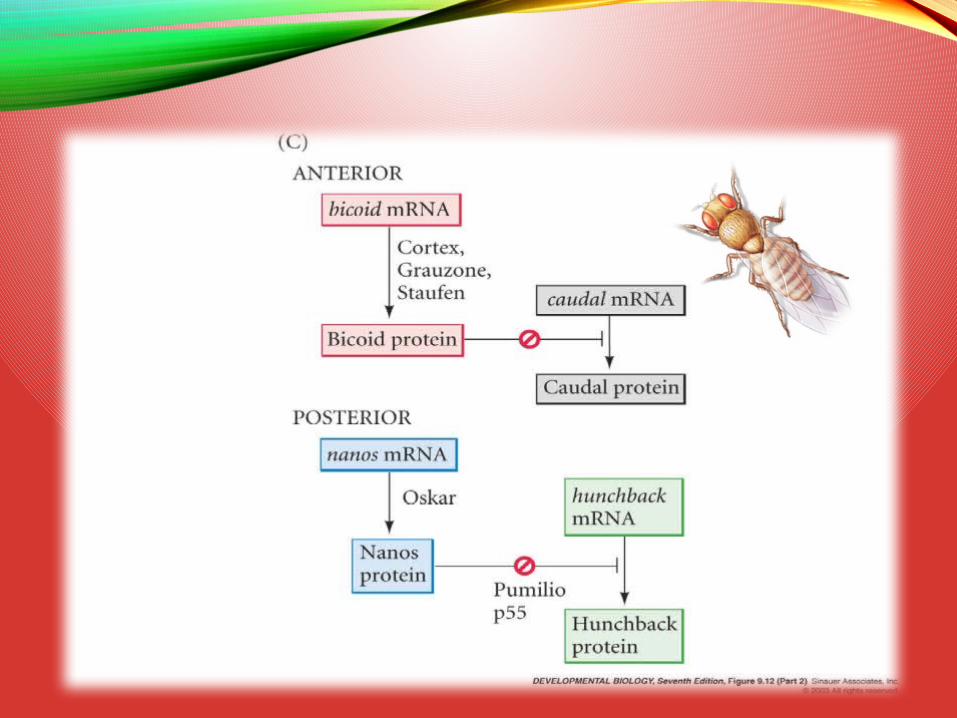
Model of Anterior-Posterior Patterning
mRNA in oocytes(maternal messages)
Early cleavageembryo proteins
hunchback translationrepressed by Nanos
caudal translationrepressed by Bicoid
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15
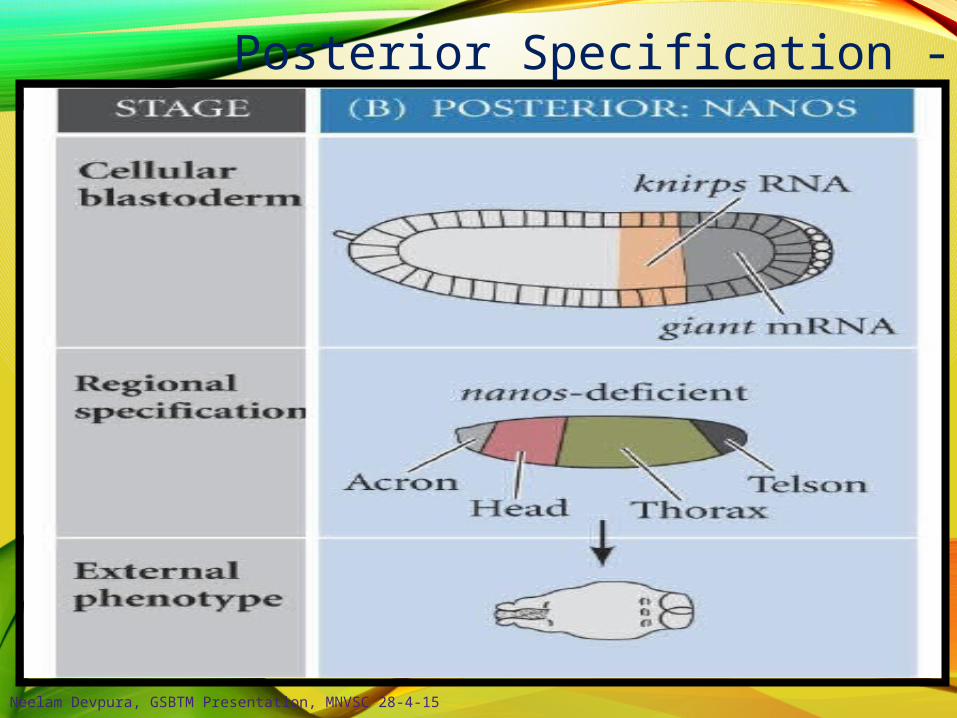
Posterior Specification - 2
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15
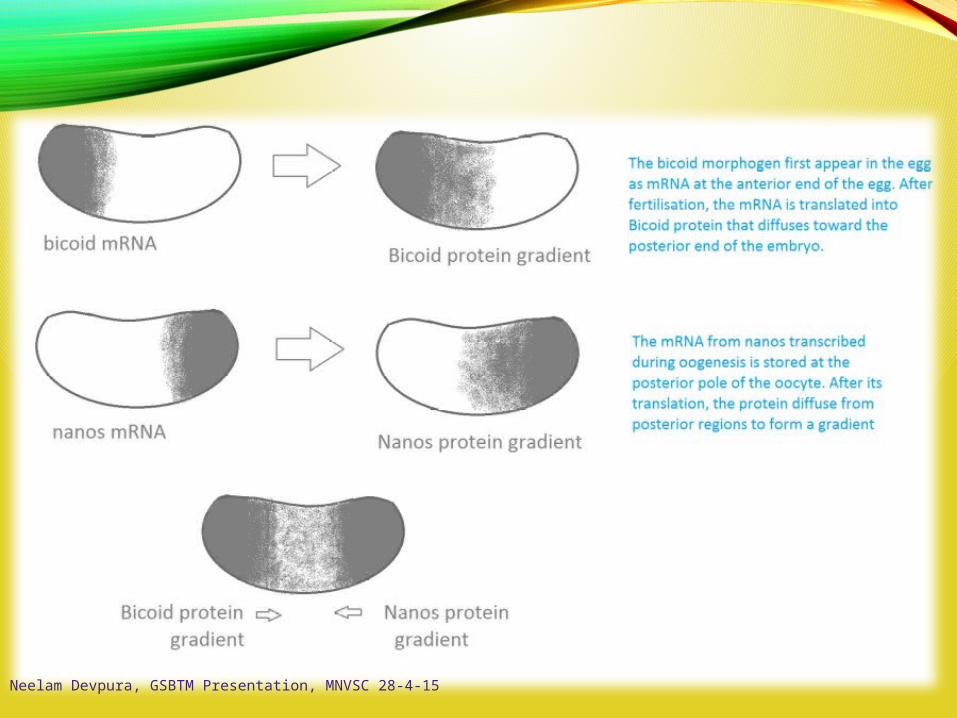
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15
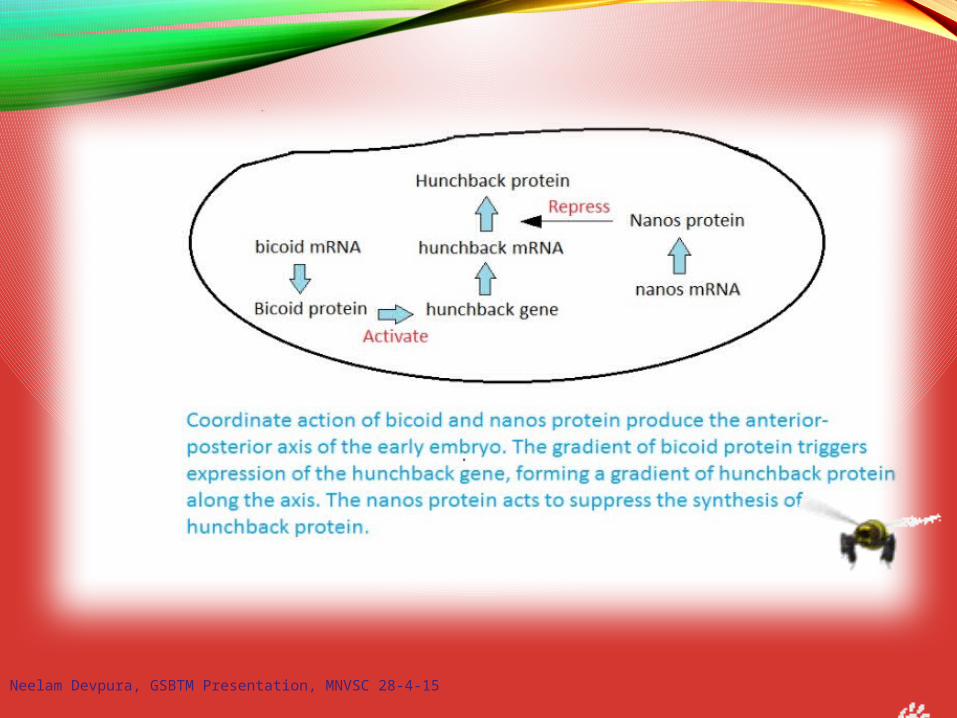
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

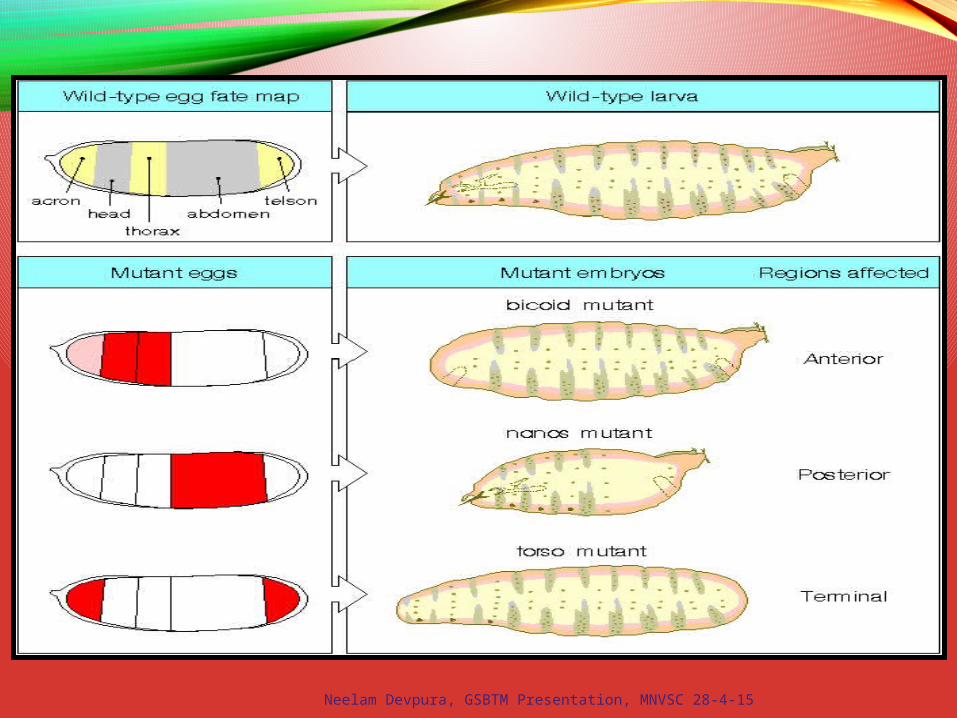
Bicoid Mutants
Martin Klingler
Bicoid – homeodomaintranscription factor;morphogen
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

Manipulating Bicoid
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15
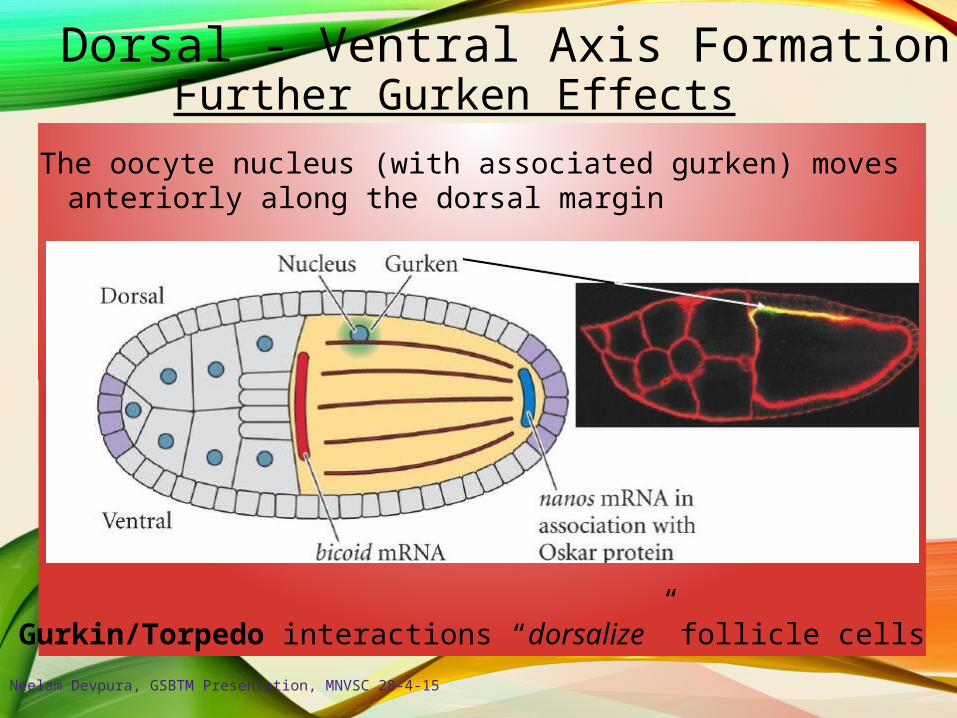
Dorsal - Ventral Axis FormationFurther Gurken Effects
The oocyte nucleus (with associated gurken) movesanteriorly along the dorsal margin
Gurkin/Torpedo interactions “dorsalize” follicle cells
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15
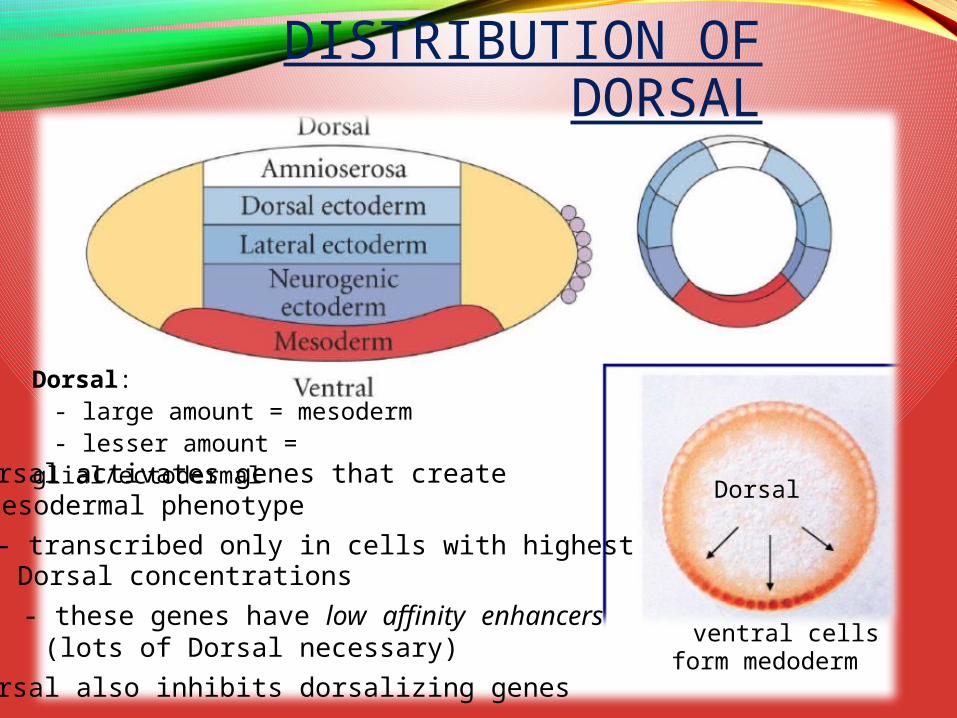
Dorsal activates genes that createmesodermal phenotype
- transcribed only in cells with highestDorsal concentrations
- these genes have low affinity enhancers(lots of Dorsal necessary)
Dorsal also inhibits dorsalizing genes
Dorsal
ventral cellsform medoderm
DISTRIBUTION OF DORSAL
Dorsal:- large amount = mesoderm- lesser amount = glial/ectodermal

Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

Zygotic Patterning Genes
decapentaplaegic (dpp),zerknüllt (zen), tolloid aredorsal patterning genes- repressed by Dorsal
Intermediate dorsal activatesrhomboid (no Twist or snail)- determines neural ectoderm- rhomboid + twist = glial cells
Intermediate dorsal activates fgf8- fgf8 repressed by snail
- promotes mesodermal ingressiondorsal
High Dorsal – activates twist and snail (low affinity enhancers)- mesoderm determinants
Dorsal (TF) – expressed ventrally; establishes diffusion gradient dorsally
~ 30 genes directly affected by Dorsal Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15
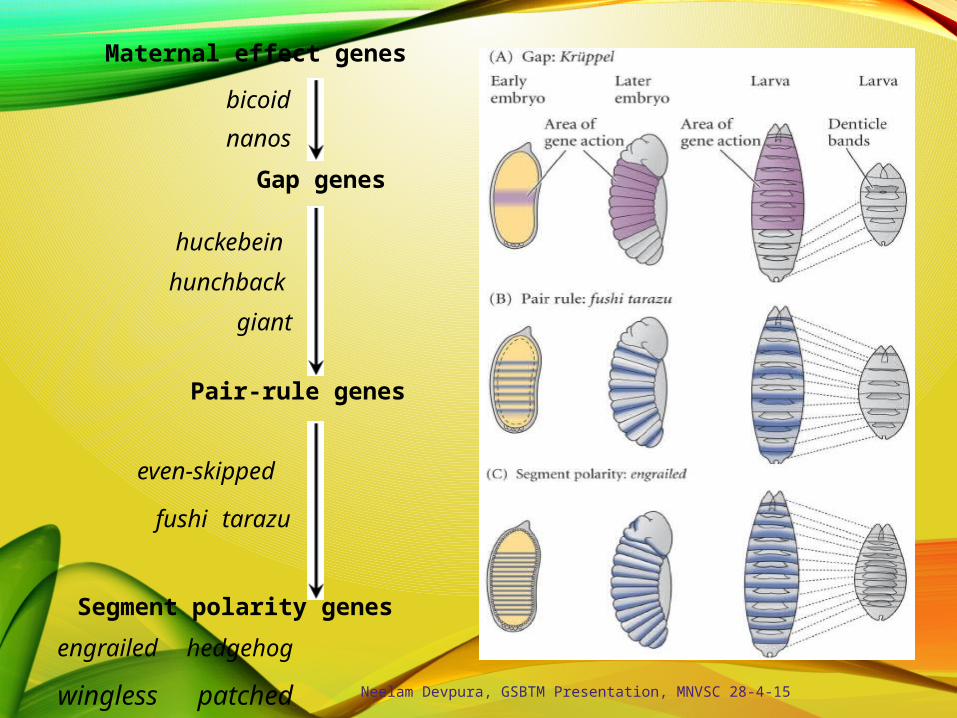
wingless patched
Maternal effect genes
bicoid
nanos
Gap genes
huckebein
hunchback
giant
Pair-rule genes
even-skipped
fushi tarazu
Segment polarity genes
engrailed hedgehog
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

GAP GENE EXPRESSION
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

Krüppel Specification by Hunchback
Wolpert, 2007
Threshold activation/repression
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

Pair-Rule Gene Regulation
e.g. – even-skipped
- each stripe regulatedby a different setof enhancers
- expression patterns arestabilized by interactionsamong other gene products
e.g. even-skippedexpression limitedby Giant
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

fushi tarazu – pair-rule gene
Segments and Parasegments
Segments and parasegmentsorganized fromA/P compartmentsout of phase
Cells of adjacentcompartmentsdo not mix
Expression patterns in early embryos are not delineatedby segmental boundaries, but by
- parasegments:fundamental units of embryonic gene expression
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15
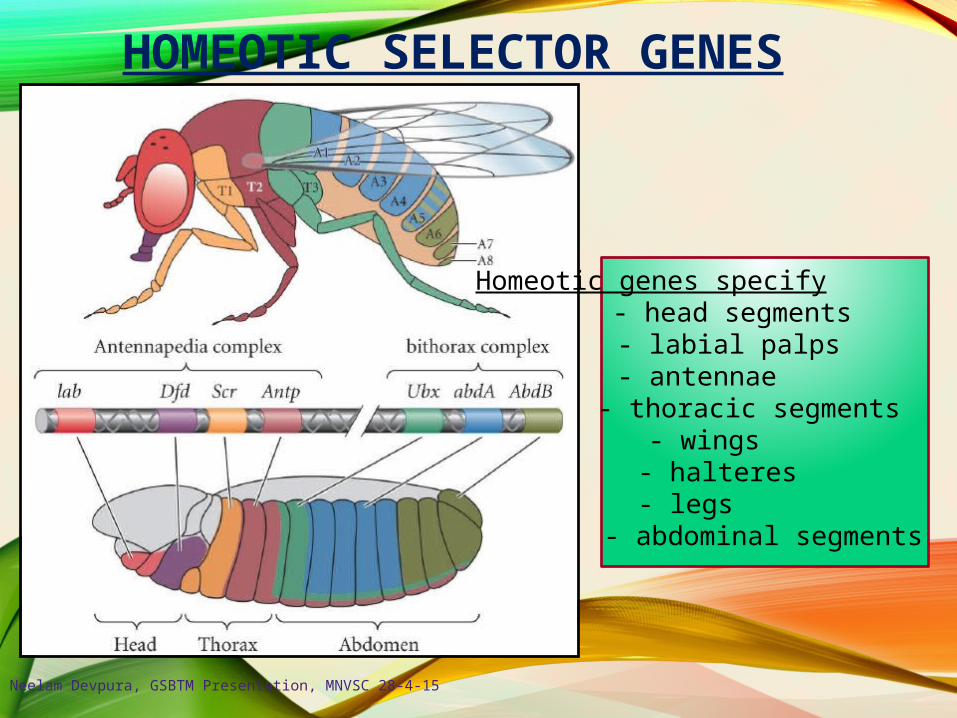
HOMEOTIC SELECTOR GENES
Homeotic genes specify :- head segments
- labial palps- antennae
- thoracic segments- wings- halteres- legs
- abdominal segments
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

Eight Genes Regulate the Identity of Within the Adult and Embryo
labial (lab)
proboscipedia (pb)
Deformed (Dfd)
Sex combs reduced (Scr)
Antennapedia (Antp)
Ultrabithorax (Ubx)
abdominal A (abd-A)
Abdominal B (Abd-B)
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

Homeotic genes encode nuclear proteins containing a DNA-binding motif called a homeodomain.
The products are transcription factors that specify segment identity by activating multiple gene expression events.
The genes are initially activated imprecisely by the concentration gradients of gap gene products.
e.g. Ubx is switched on between certain concentrations of hunchback to give a broad band of expression near the middle of the embryo. Later, fushi tarazu and even skipped sharpen the limits of Ubx expression which comes into register with the anterior boundaries of specific parasegments.
The BX-C and ANT-C genes have extensive non-coding sequences (introns) that are critical in regulating their individual expression.
Homeotic-Selector Genes
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

Homeotic Gene Expression
distal-less – jaws, limbs
Antennapeida –thoracic
Ultrabithorax –abdomen
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

Figure . The patterns of expression compared to the chromosomal locations of the genes of the HOM complex. The sequence of genes in each of the two subdivisions of the chromosomal complex corresponds to the spatial sequence in which the genes are expressed. Note that most of the genes are expressed at a high level throughout one parasegment (dark color) and at a lower level in some adjacent parasegments (medium color) where the presence of the transcripts is necessary for a normal phenotype, light color where it is not). In regions where the expression domains overlap, it is usually the most "posterior" of the locally active genes that determines the local phenotype. The drawings in the lower part of the figure represent the gene expression patterns in embryos at the extended germ band stage, about 5 hours after fertilization.
Patterns of Expression
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

•The direction of homeotic transformations depends on whether the mutation
causes loss of homeotic gene function where the gene normally acts or gain of
function where the gene normally does not act.
•Ultrabithorax (Ubx) acts in the haltere to promote haltere development and
repress wing development. Loss of function mutations in Ubx transform the
haltere into a wing.
•Dominant mutations that cause Ubx to gain function in the wing transform that
structure into a haltere.
•In antenna-to-leg transformations of Antennapedia the mutants reflect a
dominant gain of Antennapedia gene function in the antennae.
Homeotic Mutations
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

Figure :. Contribution of BX-C genes — Ubx, abdA, and AbdB—to determination of parasegment identity. The numbers above each larva indicate the parasegments; those below, the corresponding segments. The cuticular pattern of larvae is used to assign an identity to each parasegment (PS), which is indicated by color, as depicted in the wild type at the top. Red PS and segment labels indicate abnormal patterns that do not correspond exactly to any found in wild-type larvae. [Adapted from P. A. Lawrence, 1992, The Making of a Fly: Genetics of Animal Design, Blackwell Scientific Publications.]
Effects of Mutations in Bithorax Complex
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15
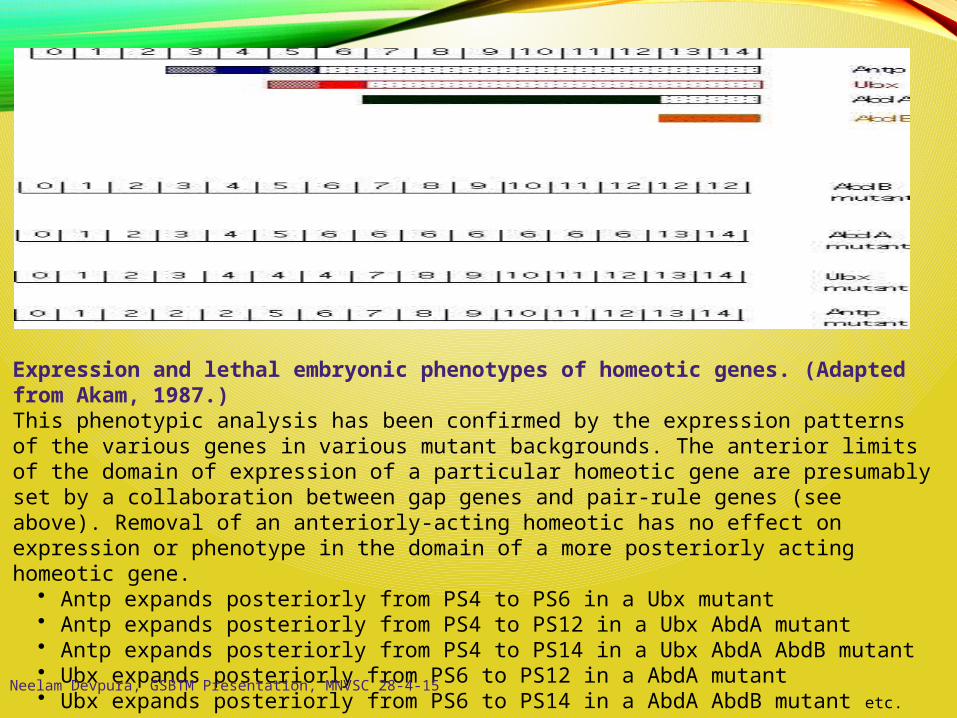
Expression and lethal embryonic phenotypes of homeotic genes. (Adapted from Akam, 1987.)This phenotypic analysis has been confirmed by the expression patterns of the various genes in various mutant backgrounds. The anterior limits of the domain of expression of a particular homeotic gene are presumably set by a collaboration between gap genes and pair-rule genes (see above). Removal of an anteriorly-acting homeotic has no effect on expression or phenotype in the domain of a more posteriorly acting homeotic gene.
• Antp expands posteriorly from PS4 to PS6 in a Ubx mutant • Antp expands posteriorly from PS4 to PS12 in a Ubx AbdA mutant • Antp expands posteriorly from PS4 to PS14 in a Ubx AbdA AbdB mutant • Ubx expands posteriorly from PS6 to PS12 in a AbdA mutant • Ubx expands posteriorly from PS6 to PS14 in a AbdA AbdB mutant etc. Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

Homeotic Gene Expression in Mutant Embryos
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

Maintaining Hox Gene Expression The transcription-control regions of some Hox genes contain binding
sites for their encoded proteins – autoregulatory loop (e.g. lab and Dfd).
A second mechanism requires proteins that modulate chromatin structure. There are two classes – the trithorax group and the polycomb group.
Early patterning requires repression as well as activation of gene expression. Polycomb proteins have a repressive effect on the expression of Hox genes.
Polycomb proteins bind multiple chromosomal locations to form large macromolecular complexes and this becomes “locked in”.
Trithorax proteins maintain the expression of many Hox genes. These also form large multiprotein complexes at multiple chromosomal sites, but mainain an open chromatin structure and stimulate gene expression.
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15
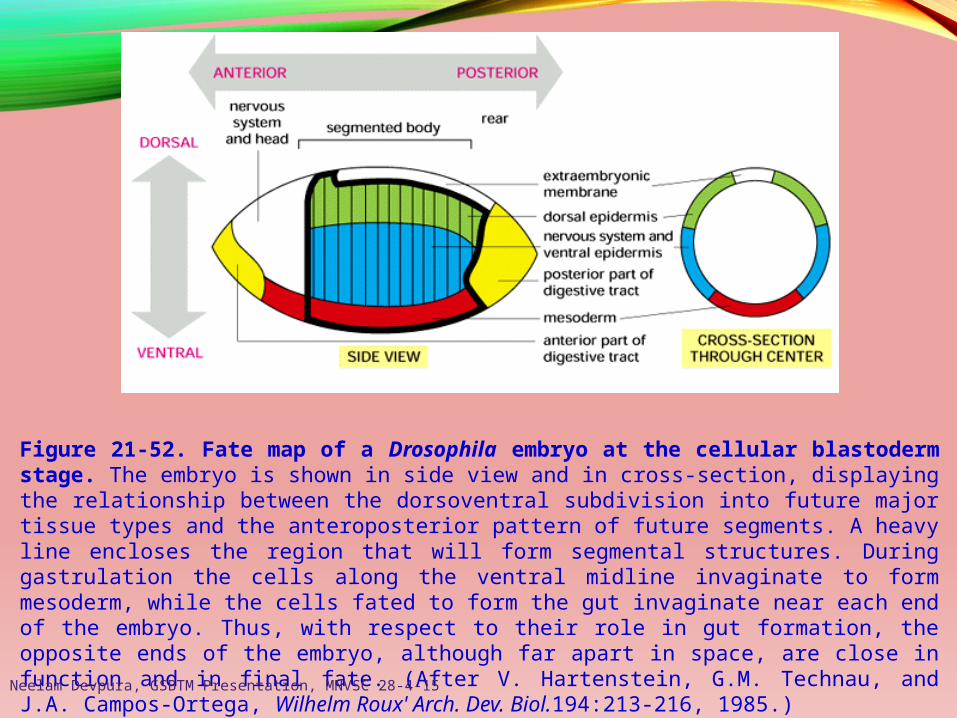
Figure 21-52. Fate map of a Drosophila embryo at the cellular blastoderm stage. The embryo is shown in side view and in cross-section, displaying the relationship between the dorsoventral subdivision into future major tissue types and the anteroposterior pattern of future segments. A heavy line encloses the region that will form segmental structures. During gastrulation the cells along the ventral midline invaginate to form mesoderm, while the cells fated to form the gut invaginate near each end of the embryo. Thus, with respect to their role in gut formation, the opposite ends of the embryo, although far apart in space, are close in function and in final fate. (After V. Hartenstein, G.M. Technau, and J.A. Campos-Ortega, Wilhelm Roux' Arch. Dev. Biol.194:213-216, 1985.)Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15
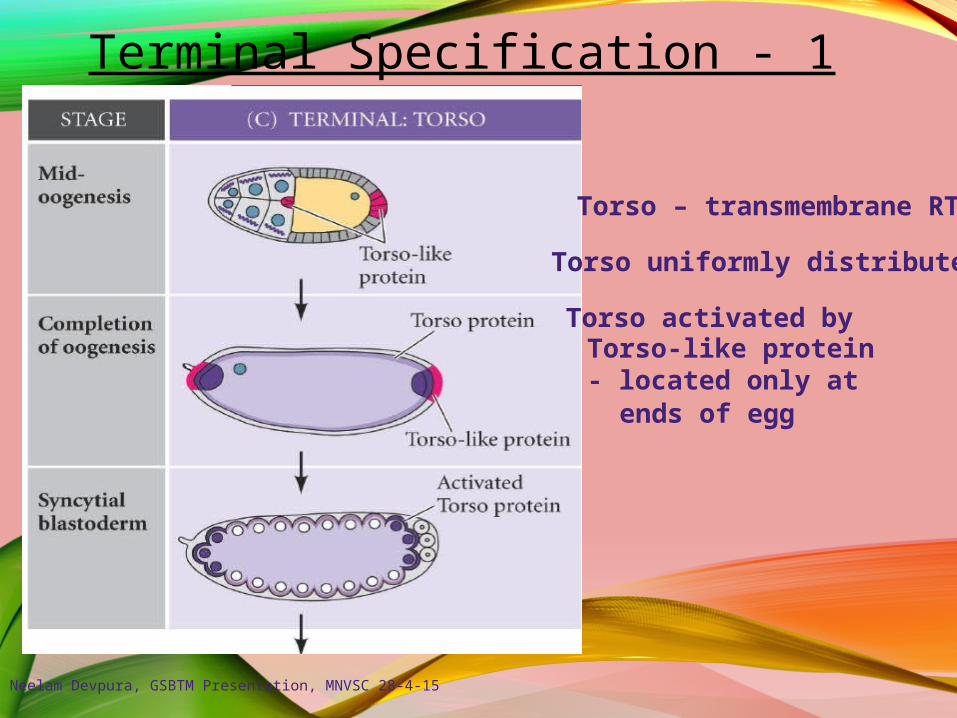
Terminal Specification - 1
Torso – transmembrane RTK
Torso uniformly distributed
Torso activated byTorso-like protein- located only at
ends of egg
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15
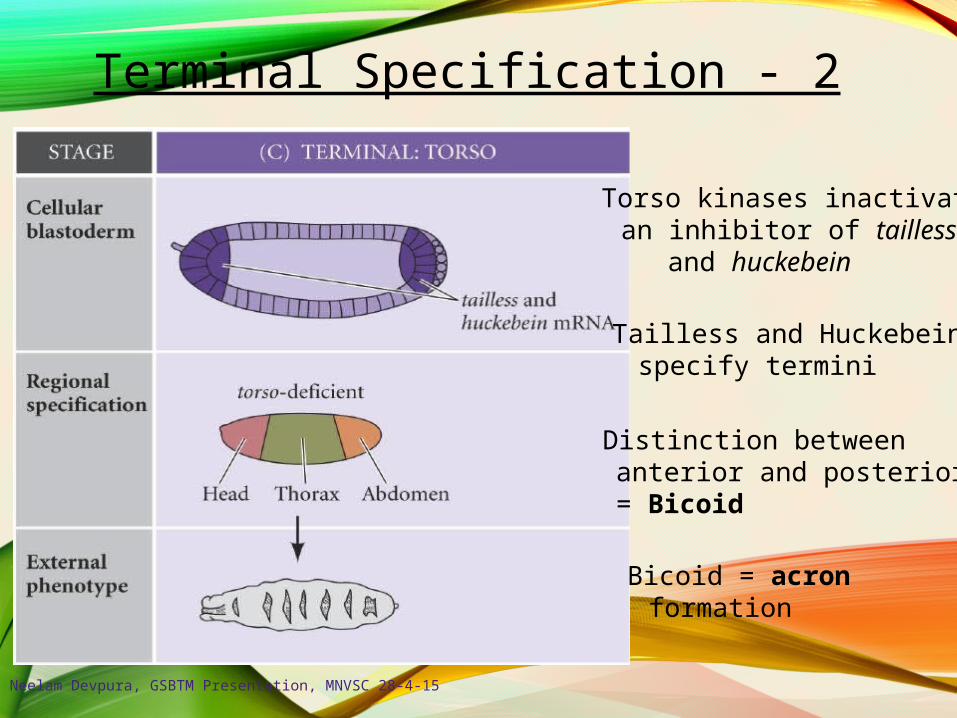
Terminal Specification - 2
Distinction betweenanterior and posterior= Bicoid
Bicoid = acronformation
Torso kinases inactivatean inhibitor of taillessand huckebein
Tailless and Huckebeinspecify termini
Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15

Neelam Devpura, GSBTM Presentation, MNVSC 28-4-15
Thank You
Discussion Time
Related Documents