NATURE | VOL 406 | 13 JULY 2000 | www.nature.com 151 articles The genome sequence of the plant pathogen Xylella fastidiosa The Xylella fastidiosa Consortium of the Organization for Nucleotide Sequencing and Analysis*, Sa ˜ o Paulo, Brazil * A full list of authors appears at the end of this paper ............................................................................................................................................................................................................................................................................ Xylella fastidiosa is a fastidious, xylem-limited bacterium that causes a range of economically important plant diseases. Here we report the complete genome sequence of X. fastidiosa clone 9a5c, which causes citrus variegated chlorosis—a serious disease of orange trees. The genome comprises a 52.7% GC-rich 2,679,305-base-pair (bp) circular chromosome and two plasmids of 51,158 bp and 1,285 bp. We can assign putative functions to 47% of the 2,904 predicted coding regions. Efficient metabolic functions are predicted, with sugars as the principal energy and carbon source, supporting existence in the nutrient-poor xylem sap. The mechanisms associated with pathogenicity and virulence involve toxins, antibiotics and ion sequestration systems, as well as bacterium–bacterium and bacterium–host interactions mediated by a range of proteins. Orthologues of some of these proteins have only been identified in animal and human pathogens; their presence in X. fastidiosa indicates that the molecular basis for bacterial pathogenicity is both conserved and independent of host. At least 83 genes are bacteriophage-derived and include virulence-associated genes from other bacteria, providing direct evidence of phage-mediated horizontal gene transfer. Citrus variegated chlorosis (CVC), which was first recorded in Brazil in 1987, affects all commercial sweet orange varieties 1 . Symptoms include conspicuous variegations on older leaves, with chlorotic areas on the upper side and corresponding light brown lesions, with gum-like material on the lower side. Affected fruits are small, hardened and of no commercial value. A strain of Xylella fastidiosa was first identified as the causal bacterium in 1993 (ref. 2) and found to be transmitted by sharpshooter leafhoppers in 1996 (ref. 3). CVC control is at present limited to removing infected shoots by pruning, the application of insecticides and the use of healthy plants for new orchards. In addition to CVC, other strains of X. fastidiosa cause a range of economically important plant diseases including Pierce’s disease of grapevine, alfalfa dwarf, phony peach disease, periwinkle wilt and leaf scorch of plum, and are also associated with diseases in mulberry, pear, almond, elm, sycamore, oak, maple, pecan and coffee 4 . The triply cloned X. fastidiosa 9a5c, sequenced here, was derived from the pathogenic culture 8.1b obtained in 1992 in Bordeaux (France) from CVC-affected Valencia sweet orange twigs collected in Macaubal (Sa ˜o Paulo, Brazil) on May 21, 1992 (ref. 2). Strain 9a5c produces typical CVC symptoms on inoculation into experimental citrus plants 5 , and into Nicotiana tabacum (S. A. Lopes, personal communication) and Catharantus roseus (P. Brant- Monteiro, personal communication)—two novel experimental hosts. General features of the genome The basic features of the genome are listed in Table 1 and a detailed map is shown in Fig. 1. The conserved origin of replication of the large chromosome has been identified in a region between the putative 50S ribosomal protein L34 and gyrB genes containing dnaA, dnaN and recF 6 . The Escherichia coli DnaA box consensus sequence TTATCCACA is found on both DNA strands close to dnaA. In addition, there are typical 13-nucleotide (ACCACCAC- CACCA) and 9-nucleotide (two TTTCATTGG and two TTTTA- TATT) sequences in other intergenic sequences of this region. This region is coincident with the calculated GC-skew signal inversion 7 . We have designated base 1 of the X. fastidiosa genome as the first Tof the only TTTTAT sequence found between the ribosomal protein L34 gene and dnaA. The overall percentage of open reading frames (ORFs) for which a putative biological function could be assigned (47%) was slightly below that for other sequenced genomes such as Thermotoga maritima 8 (54%), Deinococcus radiodurans 9 (52.5%) and Neisseria meningitidis 10 (53.7%). This may reflect the lack of previous complete genome sequences from phytopathogenic bacteria. Plas- mid pXF1.3 contains only two ORFs, one of which encodes a replication-associated protein. Plasmid pXF51 contains 64 ORFs, of which 5 encode proteins involved in replication or plasmid stability and 20 encode proteins potentially involved in conjugative transfer. One ORF encodes a protein similar to the virulence- associated protein D (VapD), found in many other bacterial pathogens 11 . Four regions of pXF51 present significant DNA simi- larity to parts of transposons found in plasmids from other bacteria, suggesting interspecific horizontal exchange of genetic material. The principal paralogous families are summarized in Table 2. The complete list of ORFs with assigned function is shown in Table 3. Seventy-five proteins present in the 21 completely sequenced genomes in the COG database 12 (as of 15th March 2000) were also found in X. fastidiosa. Each of these sequences was used to Table 1 General features of the Xylella fastidiosa 9a5c genome Main chromosome Length (bp) 2,679,305 G+C ratio 52.7% Open reading frames (ORFs) 2,782 Coding region (% of chromosome size) 88.0% Average ORF length (bp) 799 ORFs with functional assignment 1,283 ORFs with matches to conserved hypothetical proteins 310 ORFs without significant data base match 1,083 Ribosomal RNA operons 2 (16SrRNA-Ala-TGC-tRNA-Ile-GAT- tRNA-23SrRNA-5SrRNA) tRNAs 49 (46 different sequences corresponding to all 20 amino acids) tmRNA 1 ............................................................................................................................................................................. Plasmid pXF51 Length (bp) 51,158 G+C ratio 49.6% Open reading frames (ORFs) 64 Protein coding region (% of plasmid size) 86.9% ORFs with functional assignment 30 ORFs with matches to conserved hypothetical proteins 8 ORFs without significant data base match 24 ............................................................................................................................................................................. Plasmid pXF1.3 Length (bp) 1,285 G+C ratio 55.6% Open reading frames (ORFs) 2 ORFs with functional assignment 1 ............................................................................................................................................................................. © 2000 Macmillan Magazines Ltd

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

NATURE | VOL 406 | 13 JULY 2000 | www.nature.com 151
articles
The genome sequence of the plantpathogen Xylella fastidiosaThe Xylella fastidiosa Consortium of the Organization for Nucleotide Sequencing and Analysis*, Sao Paulo, Brazil
* A full list of authors appears at the end of this paper............................................................................................................................................................................................................................................................................
Xylella fastidiosa is a fastidious, xylem-limited bacterium that causes a range of economically important plant diseases. Here wereport the complete genome sequence of X. fastidiosa clone 9a5c, which causes citrus variegated chlorosis—a serious disease oforange trees. The genome comprises a 52.7% GC-rich 2,679,305-base-pair (bp) circular chromosome and two plasmids of51,158 bp and 1,285 bp. We can assign putative functions to 47% of the 2,904 predicted coding regions. Efficient metabolicfunctions are predicted, with sugars as the principal energy and carbon source, supporting existence in the nutrient-poor xylemsap. The mechanisms associated with pathogenicity and virulence involve toxins, antibiotics and ion sequestration systems, aswell as bacterium–bacterium and bacterium–host interactions mediated by a range of proteins. Orthologues of some of theseproteins have only been identified in animal and human pathogens; their presence in X. fastidiosa indicates that the molecularbasis for bacterial pathogenicity is both conserved and independent of host. At least 83 genes are bacteriophage-derived andinclude virulence-associated genes from other bacteria, providing direct evidence of phage-mediated horizontal gene transfer.
Citrus variegated chlorosis (CVC), which was first recorded in Brazilin 1987, affects all commercial sweet orange varieties1. Symptomsinclude conspicuous variegations on older leaves, with chloroticareas on the upper side and corresponding light brown lesions, withgum-like material on the lower side. Affected fruits are small,hardened and of no commercial value. A strain of Xylella fastidiosawas first identified as the causal bacterium in 1993 (ref. 2) and foundto be transmitted by sharpshooter leafhoppers in 1996 (ref. 3). CVCcontrol is at present limited to removing infected shoots by pruning,the application of insecticides and the use of healthy plants for neworchards. In addition to CVC, other strains of X. fastidiosa cause arange of economically important plant diseases including Pierce’sdisease of grapevine, alfalfa dwarf, phony peach disease, periwinklewilt and leaf scorch of plum, and are also associated with diseases inmulberry, pear, almond, elm, sycamore, oak, maple, pecan andcoffee4. The triply cloned X. fastidiosa 9a5c, sequenced here, wasderived from the pathogenic culture 8.1b obtained in 1992 inBordeaux (France) from CVC-affected Valencia sweet orangetwigs collected in Macaubal (Sao Paulo, Brazil) on May 21, 1992(ref. 2). Strain 9a5c produces typical CVC symptoms on inoculationinto experimental citrus plants5, and into Nicotiana tabacum (S. A.Lopes, personal communication) and Catharantus roseus (P. Brant-Monteiro, personal communication)—two novel experimentalhosts.
General features of the genomeThe basic features of the genome are listed in Table 1 and a detailedmap is shown in Fig. 1. The conserved origin of replication of thelarge chromosome has been identified in a region between theputative 50S ribosomal protein L34 and gyrB genes containingdnaA, dnaN and recF6. The Escherichia coli DnaA box consensussequence TTATCCACA is found on both DNA strands close todnaA. In addition, there are typical 13-nucleotide (ACCACCAC-CACCA) and 9-nucleotide (two TTTCATTGG and two TTTTA-TATT) sequences in other intergenic sequences of this region. Thisregion is coincident with the calculated GC-skew signal inversion7.We have designated base 1 of the X. fastidiosa genome as the first Tofthe only TTTTAT sequence found between the ribosomal proteinL34 gene and dnaA.
The overall percentage of open reading frames (ORFs) for which aputative biological function could be assigned (47%) was slightlybelow that for other sequenced genomes such as Thermotogamaritima8 (54%), Deinococcus radiodurans9 (52.5%) and Neisseria
meningitidis10 (53.7%). This may reflect the lack of previouscomplete genome sequences from phytopathogenic bacteria. Plas-mid pXF1.3 contains only two ORFs, one of which encodes areplication-associated protein. Plasmid pXF51 contains 64 ORFs,of which 5 encode proteins involved in replication or plasmidstability and 20 encode proteins potentially involved in conjugativetransfer. One ORF encodes a protein similar to the virulence-associated protein D (VapD), found in many other bacterialpathogens11. Four regions of pXF51 present significant DNA simi-larity to parts of transposons found in plasmids from other bacteria,suggesting interspecific horizontal exchange of genetic material.
The principal paralogous families are summarized in Table 2. Thecomplete list of ORFs with assigned function is shown in Table 3.Seventy-five proteins present in the 21 completely sequencedgenomes in the COG database12 (as of 15th March 2000) werealso found in X. fastidiosa. Each of these sequences was used to
Table 1 General features of the Xylella fastidiosa 9a5c genome
Main chromosomeLength (bp) 2,679,305G+C ratio 52.7%Open reading frames (ORFs) 2,782Coding region (% of chromosome size) 88.0%Average ORF length (bp) 799ORFs with functional assignment 1,283ORFs with matches to conservedhypothetical proteins
310
ORFs without significant data base match 1,083Ribosomal RNA operons 2
(16SrRNA-Ala-TGC-tRNA-Ile-GAT-tRNA-23SrRNA-5SrRNA)
tRNAs 49 (46 different sequencescorresponding to all 20 amino acids)
tmRNA 1.............................................................................................................................................................................
Plasmid pXF51Length (bp) 51,158G+C ratio 49.6%Open reading frames (ORFs) 64Protein coding region (% of plasmid size) 86.9%ORFs with functional assignment 30ORFs with matches to conservedhypothetical proteins
8
ORFs without significant data base match 24.............................................................................................................................................................................
Plasmid pXF1.3Length (bp) 1,285G+C ratio 55.6%Open reading frames (ORFs) 2ORFs with functional assignment 1.............................................................................................................................................................................
© 2000 Macmillan Magazines Ltd

generate a phylogenetic tree of the 22 organisms. In 69% of suchtrees, X. fastidiosa was grouped with Haemophilus influenzae andE. coli, consistent with a phylogenetic analysis undertaken with the16S rRNA gene13.
One ORF, a cytosine methyltransferase (XF1774), is interruptedby a Group II intron. The intron was identified on the basis of thepresence of a reverse transcriptase-like gene (as in other Group IIintrons), conserved splice sites, conserved sequence in structure Vand conserved elements of secondary structure14. Group II intronsare rare in prokaryotes, but have been found in different evolutivelineages including E. coli, cyanobacteria and proteobacteria15.
Transcription, translation and repairThe basic transcriptional and translational machinery of X. fasti-diosa is similar to that of E. coli16. Recombinational repair, nucleo-tide and base-excision repair, and transcription-coupled repair arepresent with some noteworthy features. For example, no photolyasewas found, indicating exclusively dark repair. Although the maingenes of the SOS pathway, recA and lexA, are present, ORFscorresponding to the three DNA polymerases induced by SOS inE. coli (DNA polymerases II, IVand V)17 are missing, indicating thatthe mutational pathway itself may be distinct.
Energy metabolismEven though X. fastidiosa is, as its name suggests, a fastidiousorganism, energy production is apparently efficient. In additionto all the genes for the glycolytic pathway, all genes for thetricarboxylic acid cycle and oxidative and electron transportchains are present. ATP synthesis is driven by the resulting chemi-osmotic proton gradient and occurs by an F-type ATP synthase.Fructose, mannose and glycerol can be utilized in addition toglucose in the glycolytic pathway. There is a complete pathway forhydrolysis of cellulose to glucose, consisting of 1,4-b-cellobiosidase,endo-1,4-b-glucanase and b-glucosidase, suggesting that cellulosebreakdown may supplement the often low concentrations of mono-saccharides in the xylem18. Two lipases are encoded in the genome,but there is no b-oxidation pathway for the hydrolysis of fatty acids,presumably precluding their utilization as an alternative carbon andenergy source. Likewise, although enzymes required for the break-down of threonine, serine, glycine, alanine, aspartate and glutamateare present, pathways for the catabolism of the other naturallyoccurring amino acids are incomplete or absent.
The gluconeogenesis pathway appears to be incomplete. Phos-phoenolpyruvate carboxykinase and the gluconeogenic enzymefructose-1,6-bisphosphatase, which are required to bypass theirreversible step in glycolysis, are not present. The absence of thefirst is compensated by the presence of phosphoenolpyruvatesynthase and malate oxidoreductase, which together can generatephosphoenolpyruvate from malate. There appears, however, to beno known compensating pathway for the absence of fructose-1,6-bisphosphatase. It is possible that among the large number ofunidentified X. fastidiosa genes there are non-homologous genesthat compensate for steps in such critical pathways. Barring thispossibility, however, the absence of a functional gluconeogenesispathway implies a strict dependence on carbohydrates both as a
source of energy and anabolic precursors. The glyoxylate cycle isabsent and the pentose phosphate pathway is incomplete. In thelatter pathway, genes for neither 6-phosphogluconic dehydrogenasenor transaldolase were identified.
Small molecule metabolismX. fastidiosa exhibits extensive biosynthetic capabilities, pre-sumably an absolute requirement for a xylem-dwelling bacterium.Most of the genes found in E. coli necessary for the synthesis of allamino acids from chorismate, pyruvate, 3-phosphoglycerate, glu-tamate and oxaloacetic acid16 were identified. However, some genesin X. fastidiosa are bi-functional, such as phosphoribosyl-AMPcyclohydrolase/phosphoribosyl-ATP pyrophosphatase (XF2213),aspartokinase/homoserine dehydrogenase I (XF2225), imidazole-glycerolphosphate dehydratase/histidinol-phosphate phosphatase(XF2217) and a new diaminopimelate decarboxylase/aspartatekinase (XF1116) that would catalyse the first and the last steps oflysine biosynthesis. In addition, the gene for acetylglutamate kinase(XF1001) has an acetyltransferase domain at its carboxy-terminalend that would compensate for the missing acetyltransferase in thearginine biosynthesis pathway. Other missing genes include phos-phoserine phosphatase, cystathionine b-lyase, homoserine O-suc-cinyltransferase and 2,4,5-methyltetrahydrofolate-homocysteinemethyltransferase. The first two enzymes are also absent in theBacillus subtilis genome, the third is absent in Haemophilus influ-enzae and the fourth is missing in both genomes12. We thus presumethat alternative, unidentified enzymes complete the biosyntheticpathways in these organisms and in X. fastidiosa.
The pathways for the synthesis of purines, pyrimidines andnucleotides are all complete. X. fastidiosa is also apparently capableof both synthesizing and elongating fatty acids from acetate. Again,however, some E. coli enzymes were not found, such as holo acyl-carrier-protein synthase (also absent in Synechocystis sp., H. influenzaeand Mycoplasma genitalium) and enoyl-ACP reductase (NADPH)(FabI) (also absent from M. genitalium, Borrelia burgdorferi andTreponema pallidum)12.
X. fastidiosa appears to be capable of synthesizing an extensivevariety of enzyme cofactors and prosthetic groups, including biotin,folic acid, pantothenate and coenzyme A, ubiquinone, glutathione,thioredoxin, glutaredoxin, riboflavin, FMN, FAD, pyrimidinenucleotides, porphyrin, thiamin, pyridoxal 59-phosphate and lipo-ate. In a number of the synthetic pathways, one or more of theenzymes present in E. coli are absent, but this is also true for at leastone other sequenced Gram-negative bacterial genome in each case12.We therefore again infer that the missing enzymes are either notessential or replaced by unknown proteins with novel structures.
Transport-related proteinsA total of 140 genes encoding transport-related proteins wereidentified, representing 4.8% of all ORFs. For comparison, E. coli,B. subtilis and M. genitalium have around 10% of genes encodingtransport proteins, whereas Helicobacter pylori, Synechocystis sp. andMethanococcus jannaschii have 3.5–5.4% (ref. 19). Transport sys-tems are central components of the host–pathogen relationship(Fig. 2). There are a number of ion transporters and transporters forthe uptake of carbohydrates, amino acids, peptides, nitrate/nitrite,sulphate, phosphate and vitamin B12. Many different transport
articles
152 NATURE | VOL 406 | 13 JULY 2000 | www.nature.com
Table 2 Largest families of paralogous genes
Family(total number of families = 312)
Number of genes(total number of genes = 853)
.............................................................................................................................................................................
ATP-binding subunits of ABC transporters 23Reductases/dehydrogenases 12Two-component system, regulatory proteins 12Hypothetical proteins 10Transcriptional regulators 9Fimbrial proteins 9Two-component system, sensor proteins 9.............................................................................................................................................................................
Figure 1 Linear representation of the main chromosome and plasmids pXF51 andpXF1.3 of the Xylella fastidiosa genome. Genes are coloured according to theirbiological role. Arrows indicate the direction of transcription. Genes with frameshift andpoint mutations are indicated with an X. Ribosomal RNA genes, the tmRNA, the principalrepeats, prophages and the group II intron are indicated by coloured lines. Transfer RNAsare identified by a single letter identifying the amino acid. Pie chart represents thedistribution of the number of genes according to biological role. The numbers belowprotein-producing genes correspond to gene IDs.
Q
© 2000 Macmillan Magazines Ltd

families are represented and include both small and large mechano-sensitive conductance ion channels, a monovalent cation:protonantiporter (CAP-2) and a glycerol facilitator belonging to themajor intrinsic protein (MIP) family. In addition, 23 ABCtransport systems comprising 41 genes can be identified. X. fasti-diosa appears to possess a phosphotransferase system (PTS) thattypically mediates small carbohydrate uptake. There are both theenzyme I and HPr components of this system, as well as a genesupposedly involved in its regulation (pstK or hprK); however, thereis no PTS permease—an essential component of the phosphotrans-ferase complex. The functionality of the system therefore remains inquestion.
There are five outer membrane receptors, including siderophores,ferrichrome-iron and haemin receptors, which are all associatedwith iron transport. The energizing complexes, TonB–ExbB–ExbDand the paralogous TolA–TolR–TolQ, essential for the functioningof the outer membrane receptors, are also present. In all, 67 genesencode proteins involved in iron metabolism. We propose that in X.fastidiosa the uptake of iron and possibly of other transition metalions such as manganese causes a reduction in essential micro-nutrients in the plant xylem, contributing to the typical symptomsof leaf variegation.
The X. fastidiosa genome encodes a battery of proteins thatmediate drug inactivation and detoxification, alteration of potentialdrug targets, prevention of drug entry and active extrusion ofdrugs and toxins. These include ABC transporters and transportprocesses driven by a proton gradient. Of the latter, eight belong tothe hydrophobe/amphiphile efflux-1 (HAE1) family, which act asmultidrug resistance factors.
AdhesionX. fastidiosa is characteristically observed embedded in an extra-cellular translucent matrix in planta20. Clumps of bacteria formwithin the xylem vessels leading to their blockage and symptoms ofthe disease such as water-stress leaf curling. We deduce, from ouranalysis of the complete genome sequence, that the matrix iscomposed of extracellular polysaccharides (EPSs) synthesized byenzymes closely related to those of Xanthomonas campestris pvcampestris (Xcc) that produce what is commercially known asxanthan gum. In comparison with Xcc, however, we did not findgumI (encoding glycosyltransferase V, which incorporates theterminal mannose), gumL (encoding ketalase which adds pyruvateto the polymer) or gumG (encoding acetyltransferase which addsacetate), suggesting that Xylella gum may be less viscous than itsXanthomonas counterpart.
Positive regulation of the synthesis of extracelullar enzymes andEPS in Xanthomomas is effected by proteins coded by the rpf(regulation of pathogenicity factors) gene cluster21. Mutations inany of these genes in Xanthomomas results in failure to synthesizethe EPS. In consequence, the strain becomes non-pathogenic21.X. fastidiosa contains genes that encode RpfA, RpfB, RpfC and RpfF,suggesting that both bacteria may regulate the synthesis of patho-genic EPS factors through similar mechanisms.
Fimbria-like structures are readily apparent upon electron micro-scopical observation of X. fastidiosa within both its plant and insecthosts22. Because of the high velocity of xylem sap passing throughnarrow portions of the insect foregut, fimbria-mediated attachmentmay be essential for insect colonization. Indeed, in the insectmouthparts the bacteria are attached in ordered arrays, indicatingspecific and polarized adhesion23. In addition, fimbriae are thoughtto be involved in both plant–bacterium and bacterium–bacteriuminteractions during colonization of the xylem itself. We identified 26genes encoding proteins responsible for the biogenesis and functionof Type 4 fimbria filaments. This type of fimbria is found at the polesof a wide range of bacterial pathogens where they act to mediateadhesion and translocation along epithelial surfaces24. The genesinclude pilS and pilR homologues, which encode a two-component
system controlling transcription of fimbrial subunits, presumably inresponse to host cues, and pilG, H, I, J and chpA, which encode achemotactic system transducing environmental signals to the pilusmachinery.
In addition to the EPS and fimbriae, which are likely to havecentral roles in the clumping of bacteria and in adhesion to thexylem walls, we also identified outer membrane protein homo-logues for afimbrial adhesins. Although fimbrial adhesins are wellcharacterized as crucial virulence factors in both plant and humanpathogens25, afimbrial adhesins, which are directly associated withthe bacterial cell surface, have been hitherto associated only withhuman and animal pathogens, where they promote adherence toepithelial tissue. Of the three putative adhesins of this kind iden-tified in X. fastidiosa, two exhibit significant similarity to eachother (XF1981, XF1529) and to the hsf and hia gene products ofH. influenzae26. The third (XF1516) is similar to the uspA1 geneproduct of Moraxella catarrhalis27. All these proteins share thecommon C-terminal domain of the autotransporter family28.Direct experimentation will be required to establish whether theseadhesins promote binding to plant cell structures or components ofthe insect vector foregut, or both. Nevertheless, their presence in theX. fastidiosa genome adds to the increasing evidence for thegenerality of mechanisms of bacterial pathogenicity, irrespectiveof the host organism29.
We also identified three different haemagglutinin-like genes.Again, similar genes have not previously been identified in plantpathogens. These genes (XF2775, XF2196, XF0889) are the largest inthe genome and exhibit highest similarity to a Neisseria meningitidisputative secreted protein10.
Intervessel migrationMovement between individual xylem vessels is crucial for effectivecolonization by X. fastidiosa. For this to occur, degradation of the pitmembrane of the xylem vessel is required. Of the known pectolyticenzymes capable of this function, a polygalacturonase precursorand a cellulase were identified, although the former contains anauthentic frameshift. These genes exhibited highest similarity toorthologues in Ralstonia solanacearum—which causes wilt diseasein tomatoes—where the polygalacturonase genes are required forwild-type virulence.
ToxicityWe identified five haemolysin-like genes: haemolysin III (XF0175),which belongs to an uncharacterized protein family, and four others(XF0668, XF1011, XF2407, XF2759) which belong to the RTX toxinfamily that contains tandemly repeated glycine-rich nonapeptidemotifs at the C-terminal domain. One of these ORFs is closelyrelated to bacteriocin, an RTX toxin also found in the plantbacterium Rhizobium leguminosarum30. RTX or RTX-like proteinsare important virulence factors widely distributed among Gram-negative pathogenic bacteria31.
There are two Colicin-V-like precursor proteins. Colicin V is anantibacterial polypeptide toxin produced by E. coli, which actsagainst closely related sensitive bacteria32. The precursors consistof 102-amino-acid peptides (XF0262, XF0263) that have the typicalconserved leader 15-amino-acid motif, and have some similaritywith Colicin V from E. coli at the remaining C-terminal portion.The necessary apparatus for Colicin biosynthesis and secretion isalso present. Interestingly, in E. coli most of the genes necessary forbiogenesis and export of Colicin V are in a gene cluster present in aplasmid, whereas in X. fastidiosa these genes are dispersed in thechromosome.
We found four genes that may function in polyketide biogenesis:polyketide synthase (PKS), pteridine-dependent deoxygenase,daunorubicin C-13 ketoreductase and a NonF-related protein.These genes belong to the synthesis pathways of frenolicin, rapa-mycin, daunorubicin and nonactin, respectively. These pathways
articles
NATURE | VOL 406 | 13 JULY 2000 | www.nature.com 153© 2000 Macmillan Magazines Ltd
articles
154 NATURE | VOL 406 | 13 JULY 2000 | www.nature.com
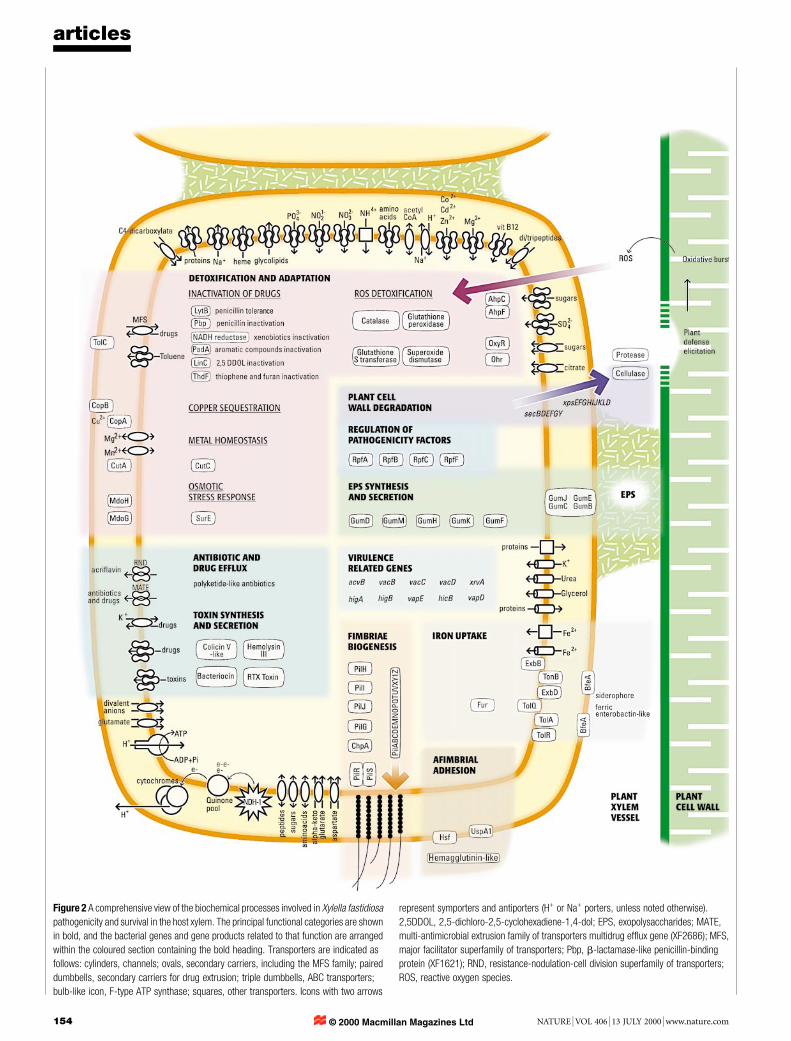
Figure 2 A comprehensive view of the biochemical processes involved in Xylella fastidiosapathogenicity and survival in the host xylem. The principal functional categories are shownin bold, and the bacterial genes and gene products related to that function are arrangedwithin the coloured section containing the bold heading. Transporters are indicated asfollows: cylinders, channels; ovals, secondary carriers, including the MFS family; paireddumbbells, secondary carriers for drug extrusion; triple dumbbells, ABC transporters;bulb-like icon, F-type ATP synthase; squares, other transporters. Icons with two arrows
represent symporters and antiporters (H+ or Na+ porters, unless noted otherwise).2,5DDOL, 2,5-dichloro-2,5-cyclohexadiene-1,4-dol; EPS, exopolysaccharides; MATE,multi-antimicrobial extrusion family of transporters multidrug efflux gene (XF2686); MFS,major facilitator superfamily of transporters; Pbp, b-lactamase-like penicillin-bindingprotein (XF1621); RND, resistance-nodulation-cell division superfamily of transporters;ROS, reactive oxygen species.
© 2000 Macmillan Magazines Ltd

include many more enzymes, which we did not find; however, someof the genes listed lie close to ORFs without significant databasematches, suggesting that at least one (as yet undiscovered) poly-ketide pathway may be functional.
ProphagesBacteriophages can mediate the evolution and transfer of virulencefactors and occasional acquisition of new traits by the bacterial host.Because as much as 7% of the X. fastidiosa genome sequencedcorresponds to double-stranded (ds) DNA phage sequences, mostlyfrom the Lambda group, we suspect that this route may have beenof particular importance for this bacterium. It is noteworthy thata very high percentage of phage-related sequences has also beendetected in a second vascular-restricted plant pathogen,Spiroplasma citri33. We identified four regions, with a high densityof ORFs homologous to phage sequences, that we considered to beprophages, in addition to isolated phage sequences dispersedthroughout the genome. Two of these prophages (each ,42 kbp,designated XfP1 and XfP2) are similar to each other, lie in oppositeorientations in distinct regions and appear to belong to the dsDNA,tailed-phage group. Both appear to contain most of the genesresponsible for particle assembly, although we know of no reportsof phage particle release from X. fastidiosa cultures. In prophageXfP1, we found two ORFs between tail genes V and W that aresimilar to ORF118 and vapA from the virulence-associated region ofthe animal pathogen Dichelobacter nodosus, which by homologyencode a killer and a suppressor protein34. Interestingly, in prophageXfP2, we found two other ORFs also between tail genes V and Wthat are similar to hypothetical ORFs of Ralstonia eutropha trans-poson Tn4371 (ref. 35). The other two identified prophages, XfP3and XfP4, are also similar in sequence to each other and to theH. influenzae cryptic prophage fflu (ref. 36). They both contain a14,317-bp exact repeat. Few particle-assembly genes were found inthese regions, suggesting that these prophages are defective. An ORFsimilar to hicB from H. influenzae, a component of the major pilusgene cluster in some isolates, was found in XfP4 (ref. 37).
The presence of virulence-associated genes from other organismswithin the prophage sequences is strong evidence for a direct role forbacteriophage-mediated horizontal gene transfer in the definitionof the bacterial phenotype.
Absence of avirulence genesPhytopathogenic bacteria generally have a limited host range, oftenconfined to members of a single species or genus. This specificity isdefined by the products of the so-called avirulence (avr) genespresent in the pathogen, which are injected directly into host cells,on infection, through a type III secretory system38–40. BLAST41
searches with all known avr and type III secretory system sequencesfailed to identify genes encoding proteins with significant simi-larities in the genome of X. fastidiosa. Although the variability of avrgenes amongst bacteria might account for this apparent lack, thehigh level of similarity of some components of the type III secretorysystem argues against this. We suspect that these genes are, in fact,not required because of the insect-mediated transmission andvascular restriction of the bacterium that obviates the necessityof host cell infection. Furthermore, if the differing host ranges ofX. fastidiosa are molecularly defined, this may be by a quite differentmechanism not involving avr proteins.
ConclusionsBefore the elucidation of its complete genome sequence, very littlewas known of the molecular mechanisms of X. fastidiosa pathogeni-city. Indeed, this bacterium was probably the least characterized of allorganisms that have been fully sequenced. Our complete geneticanalysis has determined not only the basic metabolic and replicativecharacteristics of the bacterium, but also a number of potential
pathogenicity mechanisms. Some of these have not previously beenpostulated to occur in phytopathogens, providing new insights intothe generality of these processes. Indeed, the availability of this firstcomplete plant pathogen genome sequence will now allow theinitiation of the detailed comparison of animal and plant pathogensat the whole-genome level. In addition, the information containedin the sequence should provide the basis for an accelerated andrational experimental dissection of the interactions between X.fastidiosa and its hosts that might lead to fresh insights intopotential approaches to the control of CVC. M
MethodsThe sequencing and analysis in this project were carried out by a network of 34 biologylaboratories and one bioinformatics centre. The network is called the Organization forNucleotide Sequencing and Analysis (ONSA)42, and is entirely located in the state of SaoPaulo, Brazil.
Sequencing and assemblyThe sequence was generated using a combination of ordered cosmid and shotgunstrategies43. A cosmid library was constructed, providing roughly 15-fold genome cover-age, containing 1,056 clones with average insert size of 40 kilobases (kb). High-densitycolony filters of the library were made, and a physical map of the genome was constructedusing a strategy of hybridization without replacement44. A total of 113 cosmid clones wasselected for sequencing on the basis of the hybridization map and end-sequence analysis.The cosmid sequences were assembled into 15 contigs covering 90% of the genome.Additionally, shotgun libraries with different insert sizes (0.8–2.0 kb and 2.0–4.5 kb) wereconstructed from nebulized or restricted genomic DNA cloned into plasmids, andsequenced to achieve a 3.74-fold coverage of high-quality sequence (29,140 reads). Most ofthe sequencing was performed with BigDye terminators on ABI Prism 377 DNAsequencers.
Cosmid and shotgun sequences were assembled into six contigs. We identified sequencegaps by linking information from forward and reverse reads, and closed either by primerwalking or insert subcloning. The remaining physical gaps were closed by combinatorialPCR and by lambda clones selected from a lDash library by end-sequencing. Thecollinearity between the genome and the obtained sequence was confirmed by digestion ofgenomic DNA with AscI, NotI, SfiI, SmiI and SrfI, followed by comparison of the digestionpattern with the electronic digestion of the generated sequence. In addition, sequencesfrom both ends of most cosmid clones and 236 l clones were used to confirm theorientation and integrity of the contigs. The sequence was assembled usingphred+phrap+consed45. All consensus bases have quality with Phred value of at least 20.There are no unexplained high quality discrepancies, each consensus base is confirmed byat least one read from each strand, and the overall error estimate is less than 1 in every10,000 bases.
ORF prediction and annotationORFs were determined using glimmer 2.0 (ref. 46) and the glimmer post-processorRBSfinder (S. L. Salzberg, personal communication). A few ORFs were found by handguided by BLAST41 results. Annotation was carried out in a cooperative way, mostly bycomparison with sequences in public databases, using BLAST41 and tRNAscan-SE (ref. 47)and was based on the functional categories for E. coli48. Only one tmRNA was located(K. Williams, personal communication). To help annotate transport proteins, we built acustom BLAST41 database using sequences from http://www-biology.ucsd.edu/,msaier/transport/toc.html and compared our ORFs with these sequences. Phylogenetic trees forconserved COGs12 were built using ClustalX49 for multiple alignment and Phylip50.Paralogous gene families (Table 2) were determined using BLASTX with the E-valuecut-off equal to e-5 and such that at least 60% of the query sequence and at least 30% of thesubject sequence were aligned.
Received 24 March; accepted 24 May 2000.
1. Rosseti, V. et al. Presence de bacteries dans le xyleme d’orangers atteints de chlorose variegee, une
nouvelle maladie des agrumes au Bresil. C. R. Acad. Sci. Paris serie III, 310, 345–349 (1990).
2. Chang, C. J. et al. Culture and serological detection of the xylem-limited bacterium causing citrus
variegated chlorosis and its identification as a strain of Xylella fastidiosa. Curr. Microbiol. 27, 137–142
(1993).
3. Roberto, S. R., Coutinho, A., De Lima, J. E. O., Miranda, V. S. & Carlos, E. F. Transmissao de Xylella
fastidiosa pelas cigarrinhas Dilobopterus costalimai, Acrogonia terminalis e Oncometopia facialis em
citros. Fitopatol. Bras. 21, 517–518 (1996).
4. Purcell, A. H. & Hopkins, D. L. Fastidious xylem-limited bacterial plant pathogens. Annu. Rev.
Phytopathol. 34, 131–151 (1996).
5. Li, W. B. et al. A triply cloned strain of Xylella fastidiosa multiplies and induces symptoms of citrus
variegated chlorosis in sweet orange. Curr. Microbiol. 39, 106–108 (1999).
6. Ye, F., Renaudin, J., Bove, J. M. & Laigret, F. Cloning and sequencing of the replication origin (oriC) of
the Spiroplasma citri chromosome and construction of autonomously replicating artificial plasmids.
Curr. Microbiol. 29, 23–29 (1994).
7. Francino, M. P. & Ochman, H. Strand asymmetries in DNA evolution. Trends Genet. 13, 240–245
(1997).
8. Nelson, K. E. et al. Evidence for lateral gene transfer between Archaea and bacteria from genome
sequence of Thermotoga maritima. Nature 399, 323–329 (1999).
articles
NATURE | VOL 406 | 13 JULY 2000 | www.nature.com 155© 2000 Macmillan Magazines Ltd

9. White, O. et al. Genome sequence of the radioresistant bacterium Deinococcus radiodurans R1. Science
286, 1571–1577 (1999).
10. Tettelin, H. et al. Complete genome sequence of Neisseria meningitidis serogroup B strain MC58.
Science 287, 1809–1815 (2000).
11. Katz, M. E., Strugnell, R. A. & Rood, J. I. Molecular characterization of a genomic region associated
with virulence in Dichelobacter nodosus. Infect. Immun. 60, 4586–4592 (1992).
12. Tatusov, R. L., Galperin, M. Y., Natale, D. A. & Koonin, E. V. The COG database: a tool for genome
scale analysis of protein functions and evolution. Nucleic Acids Res. 28, 33–36 (2000).
13. Preston, G. M., Haubold, B. & Rainey, P. B. Bacterial genomics and adaptation to life on plants:
implications for the evolution of pathogenicity and symbiosis. Curr. Opin. Microbiol. 1, 589–597
(1998).
14. Knoop, V., Kloska, S. & Brennicke, A. On the identification of group II introns in nucleotide sequence
data. J. Mol. Biol. 242, 389–396 (1994).
15. Ferat, J. L. & Michel, F. Group II self-splicing introns in bacteria. Nature 364, 358–361 (1993).
16. Blattner, F. R. et al. The complete genome sequence of Escherichia coli K-12. Science 277, 1453–1474
(1997).
17. Bridges, B. A. DNA repair: Polymerases for passing lesions. Curr. Biol. 9, R475–R477 (1999).
18. Brodbeck, B. V., Andersen, P. C. & Mizell, R. F. Effects of total dietary nitrogen and nitrogen form on
the development of xylophagous leafhoppers. Arch. Insect Biochem. Physiol. 42, 37–50 (1999).
19. Paulsen, I. T., Sliwinski, M. K. & Saier, M. H. J. Microbial genome analyses: global comparisons of
transport capabilities based on phylogenies, bioenergetics and substrate specificities. J. Mol. Biol. 277,
573–592 (1998).
20. Chagas, C. M., Rossetti, V. & Beretta, M. J. G. Electron-microscopy studies of a xylem-limited
bacterium in sweet orange affected with citrus variegated chlorosis disease in Brazil. J. Phytopathol.
134, 306–312 (1992).
21. Tang, J. L. et al. Genetic and molecular analysis of a cluster of rpf genes involved in positive regulation
of synthesis of extracellular enzymes and polysaccharide in Xanthomonas campestris pathovar
campestris. Mol. Gen. Genet. 226, 409–417 (1991).
22. Raju, C. B. & Wells, J. M. Diseases caused by fastidious xylem-limited bacteria. Plant Disease 70, 182–
186 (1986).
23. Brlansky, R. H., Timmer, I. W., French, W. J. & McCoy, R. E. Colonization of the sharpshooter vectors,
Oncometopia igricans and Homalodisca coagulata, by xylem-limited bacteria. Phytopathology 73, 530–
535 (1983).
24. Fernandez, L. A. & Berenguer, J. Secretion and assembly of regular surface structures in Gram-negative
bacteria. FEMS Microbiol. Rev. 24, 21–44 (2000).
25. Soto, G. E. & Hultgren, S. J. Bacterial adhesins: common themes and variations in architecture and
assembly. J. Bacteriol. 181, 1059–1071 (1999).
26. Geme, J. W. Molecular determinants of the interaction between Haemophilus influenzae and human
cells. Am. J. Respir. Crit. Care Med. 154, S192–S196 (1996).
27. Cope, L. D. et al. Characterization of the Moraxella catarrhalis uspA1 and uspA2 genes and their
encoded products. J. Bacteriol. 181, 4026–4034 (1999).
28. Henderson, I. R., Navarro-Garcia, F. & Nataro, J. P. The great escape: structure and function of the
autotransporter proteins. Trends Microbiol. 6, 370–378 (1998).
29. Rahme, L. G. et al. Common virulence factors for bacterial pathogenicity in plants and animals.
Science 268, 1899–1902 (1995).
30. Oresnik, I. J., Twelker, S. & Hynes, M. F. Cloning and characterization of a Rhizobium leguminosarum
gene encoding a bacteriocin with similarities to RTX toxins. Appl. Environ. Microbiol. 65, 2833–2840
(1999).
31. Lally, E. T., Hill, R. B., Kieba, I. R. & Korostoff, J. The interaction between RTX toxins and target cells.
Trends Microbiol. 7, 356–361 (1999).
32. Havarstein, L. S., Holo, H. & Nes, I. F. The leader peptide of colicin V shares consensus sequences with
leader peptides that are common among peptide bacteriocins produced by gram-positive bacteria.
Microbiology 140, 2383–2389 (1994).
33. Ye, F. et al. A physical and genetic map of the Spiroplasma citri genome. Nucleic Acids Res. 20, 1559–
1565 (1992).
34. Billington, S. J., Johnston, J. L. & Rood, J. I. Virulence regions and virulence factors of the ovine footrot
pathogen, Dichelobacter nodosus. FEMS Microbiol. Lett. 145, 147–156 (1996).
35. Merlin, C., Springael, D. & Toussaint, A. Tn4371: A modular structure encoding a phage-like
integrase, a Pseudomonas-like catabolic pathway, and RP4/Ti-like transfer functions. Plasmid 41, 40–
54 (1999).
36. Hendrix, R. W., Smith, M. C., Burns, R. N., Ford, M. E. & Hatfull, G. F. Evolutionary relationships
among diverse bacteriophages and prophages: All the world’s a phage. Proc. Natl Acad. Sci. USA 96,
2192–2197 (1999).
37. Mhlanga-Mutangadura, T., Morlin, G., Smith, A. L., Eisenstark, A. & Golomb, M. Evolution of the
major pilus gene cluster of Haemophilus influenzae. J. Bacteriol. 180, 4693–4703 (1998).
38. Alfano, J. R. & Collmer, A. The type III (Hrp) secretion pathway of plant pathogenic bacteria:
trafficking harpins, Avr proteins, and death. J. Bacteriol. 179, 5655–5662 (1997).
39. Galan, J. E. & Collmer, A. Type III secretion machines: bacterial devices for protein delivery into host
cells. Science 284, 1322–1328 (1999).
40. Young, G. M., Schmiel, D. H. & Miller, V. L. A new pathway for the secretion of virulence factors by
bacteria: the flagellar export apparatus functions as a protein-secretion system. Proc. Natl Acad. Sci.
USA 96, 6456–6461 (1999).
41. Altschul, S. F. et al. Gapped BLAST and PSI-BLAST: a new generation of protein database search
programs. Nucleic Acids Res. 25, 3389–3402 (1997).
42. Simpson, A. J. & Perez, J. F. ONSA, the Sao Paulo Virtual Genomics Institute. Nature Biotechnol. 16,
795–796 (1998).
43. Fleischmann, R. D. et al. Whole-genome random sequencing and assembly of Haemophilus influenzae
Rd. Science 269, 496–512 (1995).
44. Hoheisel, J. D. et al. High resolution cosmid and P1 maps spanning the 14 Mb genome of the fission
yeast S. pombe. Cell 73, 109–120 (1993).
45. Gordon, D., Abajian, C. & Green, P. Consed: a graphical tool for sequence finishing. Genome Res. 8,
195–202 (1998).
46. Delcher, A. L., Harmon, D., Kasif, S., White, O. & Salzberg, S. L. Improved microbial gene
identification with GLIMMER. Nucleic Acids Res. 27, 4636–4641 (1999).
47. Lowe, T. M. & Eddy, S. R. tRNAscan-SE: a program for improved detection of transfer RNA genes in
genomic sequences. Nucleic Acids Res. 25, 955–964 (1997).
48. Riley, M. Functions of the gene products of Escherichia coli. Microbiol. Rev. 57, 862–952 (1993).
49. Thompson, J. D., Gibson, T. J., Plewniak, F., Jeanmougin, F. & Higgins, D. G. The ClustalX windows
interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic
Acids Res. 25, 4876–4882 (1997).
50. Felsenstein, J. PHYLIP—Phylogeny Inference Package (Version 3. 2). Cladistics 5, 164–166 (1989).
Supplementary information is available on Nature’s World-Wide Web site(http://www.nature.com), or on the author’s World-Wide Web site(http://www.lbi.ic.unicamp.br/xf) or as paper copy from the London editorial office ofNature.
AcknowledgementsThe consortium is indebted to J. F. Perez, Scientific Director of Fundacao de Amparo aPesquisa do Estado de Sao Paulo (FAPESP), for his strategic vision in creating andnurturing this project as well as C. A. de Pian and Jucara Parra for their administrativecoordination. We thank our Steering Committee: S. Oliver, A. Goffeau, J. Sgouros, A. C.M. Paiva and J. L. Azevedo for their critical accompaniment of the work. We also thank R.Fulton and P. Minx for their timely contribution and advice. Project funding was fromFAPESP, the RHAE programme of the Conselho Nacional de Desenvolvimento Cientıficoe Tecnologico (CNPq) and Fundecitrus. For the full list of individuals who contributed tothe completion of this project see (http://www.lbi. ic.unicamp.br/xf).
Correspondence and requests for materials should be addressed to J.C.S. (e-mail:[email protected]). The sequence has been deposited in GenBank with accessionnumbers AE003849 (chromosome), AE003850 (pXF1.3) and AE003851 (pXF51).
articles
156 NATURE | VOL 406 | 13 JULY 2000 | www.nature.com
Authors: A. J. G. Simpson1, F.C. Reinach2, P. Arruda3, F. A. Abreu4, M. Acencio5, R. Alvarenga2, L. M. C. Alves6, J. E. Araya7, G. S. Baia2,C. S. Baptista8, M. H. Barros8, E. D. Bonaccorsi2, S. Bordin9, J. M. Bove10, M. R. S. Briones7, M. R. P. Bueno11, A. A. Camargo1, L. E. A.Camargo12, D. M. Carraro12, H. Carrer12, N. B. Colauto13, C. Colombo14, F. F. Costa9, M. C. R. Costa15, C. M. Costa-Neto16, L. L. Coutinho12,M. Cristofani17, E. Dias-Neto1, C. Docena2, H. El-Dorry2, A. P. Facincani6, A. J. S. Ferreira2, V. C. A. Ferreira18, J. A. Ferro6, J. S. Fraga4, S. C.Franca19, M. C. Franco20, M. Frohme21, L. R. Furlan22, M. Garnier10, G. H. Goldman23, M. H. S. Goldman24, S. L. Gomes2, A. Gruber4, P. L. Ho25,J. D. Hoheisel21, M. L. Junqueira26, E. L. Kemper3, J. P. Kitajima27, J. E. Krieger26, E. E. Kuramae28, F. Laigret10, M. R. Lambais12, L. C. C.Leite25, E. G. M. Lemos6, M. V. F. Lemos29, S. A. Lopes19, C. R. Lopes13, J. A. Machado30†, M. A. Machado17, A. M. B. N. Madeira4, H. M. F.Madeira12†, C. L. Marino13, M. V. Marques8, E. A. L. Martins25, E. M. F. Martins18, A. Y. Matsukuma2, C. F. M. Menck8, E. C. Miracca5,C. Y. Miyaki11, C. B. Monteiro-Vitorello12, D. H. Moon20, M. A. Nagai5, A. L. T. O. Nascimento25, L. E. S. Netto11, A. Nhani Jr6, F. G. Nobrega8†,L. R. Nunes31, M. A. Oliveira32, M. C. de Oliveira33, R. C. de Oliveira31, D. A. Palmieri13, A. Paris13, B. R. Peixoto2, G. A. G. Pereira32,H. A. Pereira Jr6, J. B. Pesquero16, R. B. Quaggio2, P. G. Roberto19, V. Rodrigues34, A. J. de M. Rosa34, V. E. de Rosa Jr28, R. G. de Sa34,R. V. Santelli2, H. E. Sawasaki14, A. C. R. da Silva2, A. M. da Silva2, F. R. da Silva3,27, W. A. Silva Jr15, J. F. da Silveira7, M. L. Z. Silvestri2,W. J. Siqueira14, A. A. de Souza17, A. P. de Souza3, M. F. Terenzi23, D. Truffi12, S. M. Tsai20, M. H. Tsuhako18, H. Vallada35, M. A. Van Sluys33,S. Verjovski-Almeida2, A. L. Vettore3, M. A. Zago15, M. Zatz11, J. Meidanis27 & J. C. Setubal27.
Addresses: 1, Instituto Ludwig dePesquisa sobre oCancer, Rua Prof. Antonio Prudente,109 – 4o andar, 01509-010, Sao Paulo - SP, Brazil;2, Departamento de Bioquımica, Instituto de Quımica, Universidade de Sao Paulo, Av. Prof. Lineu Prestes, 748, 05508-900, Sao Paulo -SP, Brazil; 3, Centro de Biologia Molecular e Engenharia Genetica, Universidade Estadual de Campinas, Caixa Postal 6010,13083-970,Campinas – SP, Brazil; 4, Laboratorio de BiologiaMolecular, Departamento de Patologia, Faculdade de MedicinaVeterinaria e Zootecnia,Universidade de Sao Paulo, Av. Prof. Dr. Orlando Marques de Paiva, 87, 05508-000, Sao Paulo – SP, Brazil; 5, Disciplina de Oncologia,Departamento de Radiologia, Faculdade de Medicina, Universidade de Sao Paulo, Av. Dr. Arnaldo, 455, 01296-903, Sao Paulo – SP,Brazil; 6, Departamento de Tecnologia, Faculdadede Ciencias Agrarias e Veterinarias de Jaboticabal, UniversidadeEstadual Paulista, Via
© 2000 Macmillan Magazines Ltd

articles
NATURE | VOL 406 | 13 JULY 2000 | www.nature.com 157
de Acesso Prof. Paulo D. Castellane s/n, Km 5,14884-900, Jaboticabal – SP, Brazil; 7, Departamento de Microbiologia, Imunologia eParasitologia, Escola Paulista de Medicina,UniversidadeFederal de Sao Paulo, Rua Botucatu, 862, 04023-062, Sao Paulo – SP, Brazil; 8,Departamento deMicrobiologia, Instituto deCienciasBiomedicas,UniversidadedeSaoPaulo,Av. Prof. LineuPrestes,1374, 05508-900,Sao Paulo – SP, Brazil; 9, Hemocentro, Faculdade de Ciencias Medicas, Universidade Estadual de Campinas,13083-97, Campinas – SP,Brazil; 10, Institut National de la Recherche Agronomique et Universite Victor Segalen Bordeaux 2, 71 Avenue Edouard Bourleaux,Laboratoire de Biologie Cellulaire et Moleculaire, 33883 Vilenave d’Ornon Cedex, France; 11, Departamento de Biologia, Instituto deBiociencias,UniversidadedeSaoPaulo,RuadoMatao, 277, 05508-900,SaoPaulo – SP,Brazil; 12, EscolaSuperiordeAgricultura Luiz deQueiroz, Universidade de Sao Paulo, Av. Padua Dias,11,13418-900, Piracicaba – SP, Brazil; 13, Departamento de Genetica, Instituto deBiociencias, Universidade Estadual Paulista, Distrito de Rubiao Junior,18618-000, Botucatu – SP, Brazil; 14, Centro de Genetica, BiologiaMolecular e Fitoquımica, Instituto Agronomico de Campinas, Av. Barao de Itapura,1481, Caixa Postal 28,13001-970, Campinas – SP,Brazil; 15, Departamento de Clınica Medica, Faculdade de Medicina de Ribeirao Preto, Universidade de Sao Paulo, Av. Bandeirantes,3900,14049-900, Ribeirao Preto – SP, Brazil; 16, Departamento de Biofısica, Escola Paulista de Medicina, Universidade Federal de SaoPaulo, Rua Botucatu, 862, 04023-062, Sao Paulo – SP, Brazil; 17, Centro de Citricultura Sylvio Moreira, Instituto Agronomico deCampinas, Caixa Postal 04,13490-970, Cordeiropolis – SP, Brazil; 18, Laboratorio de Bioquımica Fitopatologica e Laboratorio deImunologia, Instituto Biologico, Av. Cons. Rodrigues Alves,1252, 04014-002, Sao Paulo – SP, Brazil; 19, Departamento de BiotecnologiadePlantasMedicinais, UniversidadedeRibeirao Preto,Av.Costabile Romano, 2201,14096-380, Ribeirao Preto – SP,Brazil; 20,Centro deEnergia Nuclear na Agricultura, Universidade de Sao Paulo, Av. Centenario, 303, Caixa Postal 96,13400-970, Piracicaba – SP, Brazil; 21,Funktionelle Genomanalyse, Deutsches Krebsforschungszentrum, Im Neuenheimer Feld 506, D-69120, Heidelberg, Germany; 22,Departamento de Melhoramento e Nutricao Animal, Faculdade de Medicina Veterinaria e Zootecnia, Universidade Estadual Paulista,Fazenda Lageado, Caixa Postal 560,18600-000, Botucatu - SP, Brazil; 23, Departamento de Ciencias Farmaceuticas , Faculdade deCiencias Farmaceuticas de Ribeirao Preto, Universidade de Sao Paulo, Av. do Cafe s/n,14040-903, Ribeirao Preto – SP, Brazil; 24,Departamento de Biologia, Faculdade de Filosofia, Ciencias e Letras de Ribeirao Preto, Universidade de Sao Paulo, Av. Bandeirantes,3900,14040-901,Ribeirao Preto – SP,Brazil; 25,Centro deBiotecnologia, InstitutoButantan,Av. Vital Brasil,1500, 05503-900, Sao Paulo– SP, Brazil; 26, Laboratorio de Genetica e Cardiologia Molecular/LIM 13, Instituto do Coracao (InCor), Faculdade de Medicina,Universidade de Sao Paulo, Av. Dr. Eneas de Carvalho Aguiar, 44, 05403-000, Sao Paulo – SP, Brazil; 27, Instituto de Computacao,Universidade Estadual de Campinas, Caixa Postal 6176,13083-970, Campinas – SP, Brazil; 28, Departamento de Producao Vegetal,Faculdade de Ciencias Agronomicas, Universidade Estadual Paulista, Fazenda Lageado, Caixa Postal 237,18603-970, Botucatu – SP,Brazil; 29, Departamento de Biologia Aplicada a Agropecuaria, Faculdade de Ciencias Agrarias e Veterinarias de Jaboticabal,UniversidadeEstadual Paulista, Via deAcessoProf. PauloDonato Castellane,14884-900, Jaboticabal – SP, Brazil; 30, Hospital doCancer– A.C. Camargo, R. Antonio Prudente, 211, 01509-010, Sao Paulo – SP, Brazil; 31, Nucleo Integrado de Biotecnologia, Universidade deMogi dasCruzes,Av.Dr.CandidoXavierdeAlmeidaSouza, 200, 08780-911,Mogi dasCruzes – SP,Brazil; 32,Departamento deGeneticae Evolucao, Instituto de Biologia, Universidade Estadual de Campinas, Caixa Postal 6010,13083-970, Campinas– SP, Brazil; 33,Departamento de Botanica, Instituto de Biociencias, Universidade de Sao Paulo, Rua do Matao, 277, 05508-900, Sao Paulo - SP, Brazil;34, Departamento de Parasitologia, Microbiologia e Imunologia, Faculdade de Medicina de Ribeirao Preto, Universidade de Sao Paulo,Av. Bandeirantes, 3900,14049-900, Ribeirao Preto – SP, Brazil; 35, Departamento de Psiquiatria, Instituto de Psiquiatria, Faculdade deMedicina, Universidade de Sao Paulo, Rua Dr. Ovıdeo Pires de Campos s/n, Sala 4051 (3o. andar), 05403-010, Sao Paulo – SP, Brazil.† Present addresses: Novartis Seeds LTDA, Av. Prof. Vicente Rao, 90, 04706-900, Sao Paulo – SP, Brazil (J. A. Machado); Centro de Ciencias Agrarias e Ambientais,Pontifıcia Universidade Catolica do Parana, BR-376, Km 14, Caixa Postal 129, 83010-500, Sao Jose dos Pinhais – PR, Brazil (H. M. F. Madeira); Instituto de Pesquisa eDesenvolvimento, Universidade do Vale do Paraıba, Av. Shishimi Hifumi, 2911, 12244-000, Sao Jose dos Campos – SP, Brazil (F. G. Nobrega).
© 2000 Macmillan Magazines Ltd

articles
158 NATURE | VOL 406 | 13 JULY 2000 | www.nature.com
113
5000
1 2
3 4
5 6
7 8
9 1
0 1
1 1
2 13 1
4 1
5 1
6 1
7 1
8 19 2
0 21
22 2
3 24M 2
5 2
6 2
7 2
8 2
9 3
0 3
1 3
2 3
3 3
4 3
5 3
6 3
7 3
8 3
9 4
0 41
42
43
44
45
46
47
48
49
50
51
52
53
54 5
5 56 5
7 5
8 59
60
61
62
63
64
65
66
67
68
R 69
70
A I 7
1 7
2 7
3 7
4 7
5 7
6 7
7 7
8 7
9 8
0 8
1 8
2 8
3 8
4 85
86
87
88
89
90
91
92
93
94
95 9
6 9
7 98
99
100
101
102
103
104
105
106
107 10
810
911
011
111
2 113
114
115
116
117
118
119
120
121
122
123
124
125 S
126
127
128
129 13
0131
132
133
134
134
1350
0127
0000
13513
6 137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
159
161
162 16
316
416
516
616
716
816
9A I
170
171
172
173
174
175
176
177
178 17
918
018
1 182
183
184
185
186
187
188
189
190
191
192
193 19
4195
196
197
198
199
200 20
120
220
320
420
520
620
720
820
921
021
121
221
321
421
5216
217 A
218 21
922
022
122
222
322
422
522
6E
227
228
229
230
231
232
M23
323
423
523
623
723
823
924
024
124
224
324
424
5 24624
7 24824
925
025
125
225
325
425
525
625
725
8
258
2700
0140
5000
259
260
261 26
226
326
4 265
266
267
268
269 27
0271 27
2273
274
275
276
277
278
279
280
281
282 28
3284
285
286
287
288
289
290
291
292
293 F
294
295
296
297
298
299
300 30
1302
303
304L
305 30
630
730
830
931
031
131
231
331
431
631
731
831
932
032
132
232
332
432
532
632
7328 32
9330
332 33
3 33433
5 336 33
7338
339
340
34134
234
334
4 345
346
347
348 34
9 35035
135
235
335
435
535
635
735
835
9 360
361
362
363
364
365
366
367
368
369
370
371
372
373
374
375
376
377
378 37
9380
381
382 38
338
438
538
638
738
838
939
039
139
2
392
4050
0154
0000
QM39
339
4 39539
639
739
8 39940
040
140
240
4405
406
407
408
409 410
411
412
413
414 41
541
641
7 418
419
420
421
422
423
424
425
426 42
742
842
943
043
143
243
343
443
543
643
743
843
944
044
144
244
3444
445
446
447 44
844
945
045
145
245
345
445
545
645
745
845
946
046
1 46246
346
446
546
646
7 46846
947
047
147
247
347
547
6 477
478
479 N
480
481 48
2483 48
4485
486
487
488 48
9490 49
1492 49
3494 49
549
649
7 49849
950
150
3504
505
506
507
508
509
510
511
512
513 51
451
551
6 517
518
519 52
052
152
252
352
4 525
526
527
528
529
530
531
532 53
353
453
553
653
7 538
539
540 54
1542
544 54
5546 54
7548
549
550
550
5400
0167
5000
551
552
553
554 55
555
655
7 558
559 56
056
156
256
3 564
565
566
567
568
569
570 57
157
257
3 57457
557
657
7 578 57
9580
581 58
2583
584
585
586
587
588
589
590
591
592
593
594
595
596
597
598
599
600
601
602
603
604 60
5606
607
608
609
610
611
612
613
614
615
616 61
7618 61
962
062
162
362
462
562
662
862
963
063
163
263
363
463
563
6 637
638 63
964
064
1S
642 64
364
464
564
764
8 64965
065
165
265
365
465
565
665
765
865
966
066
166
2 663
664
665 66
666
766
866
967
067
167
267
367
467
567
667
7 V
678
679
680
681 68
268
368
468
568
668
768
9 69069
1 69269
3 69469
569
669
7 69869
9 70070
1 70270
370
470
570
670
7 70870
971
071
171
271
3
713
6750
0181
0000
714
715 71
6717
718 71
9720 72
1 72272
372
472
572
672
772
8 729
730
731 73
273
373
473
573
673
7 738 73974
074
174
274
3 744 P74
574
774
874
9 750
751
752
753 75
475
575
675
775
875
976
076
176
276
376
476
5 76676
776
876
977
0 771
773 77
477
577
677
777
877
978
0G
781
782
783
784
785
786 78
7788
789
790
791
792
793
794
795
796
797
798
799
800
801
802
803
804
805
806
807
808809
810
811
812 81
381
4 815
816
817
818
819
820
821
822
823
824
825
826
827
828 82
9830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
846
847
848
848
8100
0194
5000
849 85
085
185
285
385
4 P85
585
685
785
885
9 860
861
862
863
864
865
866
867
868
869
870
872
873
874
875
876
877
879
880
881
882
883
884
885
886
887
888
889
890
891
892
893
894
895
896
897
898 89
990
090
190
290
490
590
690
790
890
991
091
191
291
391
491
5 91691
791
992
092
192
2 923
924
925
926
927
928
929
930 93
1932
933
934
935
936
937
938
939
940
941
942
943
944
945
946
947
948
949
950
951
952
953
954 95
595
695
7 95895
996
096
196
296
396
496
696
7V
968
969 97
0 971
972
973
974
975
976
977
978
979
980
98198
2 983
984
985
986
986
9450
0110
8000
0
987
988
989
990 99
199
299
399
499
599
6 997
998
999
1000
1001
1002
1003
1004
1005
1006
1007
1008
100910
1010
1110
1210
131014
1015
1016 1017
1018
101910
2010
2110
2210
231024
1025
1026
1027
1028
1029
1030
1031
1032 10
33 103410
3510
3610
3710
3810
39 1040
1041
1042
1043
1045
1046
1047
1048
1049
1050
1052
1053
1054
1055
1056
1057
1058 10
591060 L
EA10
6110
6210
6310
6410
6510
6610
6710
6810
69 107010
7110
7210
7310
741075
1076
1077
1078
1079
1080
1081
1082
1083
1084
1085
1086
1087
1088
108910
9010
91 1092
1093
1094
109510
96 1
097
1098 10
9911
0011
0111
0211
0311
04 1105
1106
1107
1108
1109
1110
1111
1112
1113
1114
1115
1116
1117
1118
L11
19 1120
1120
1080
001
1215
000
1121
1122 11
2311
2411
2511
2611
2711
28 1129 11
30
1131
1132
1133 11
34 1135
1137
1138 11
3911
4011
4111
4211
4311
4411
4511
4611
4711
4811
4911
5011
51 1152
1153 11
5411
5511
561157
1158
1159 11
601161 11
621163 11
641165
116611
6711
6811
69 117011
7111
7211
73 1174
1175
1176
1177
1178
1179
1180
1181
1182
L11
8311
8411
85L
1186
1187
1188
1189
1190 V D
1191
1192
1193 11
941195
1196
1197
1198 11
9912
0012
0112
02 120312
0412
051206 12
07 1208
1209
1210
1211
1212
1213
1214 12
1512
1612
17 1218
1219
1220
1221
1222
1223
1224
1225
1226
1227
1228
1229
1230
1231
1232
1233
1234
K12
35 1236
1237 12
3812
39 1240
1241
1242
1243
1244
1245 12
461247
1248
1249
1250
1251
1252
1253
1254
1255 12
5612
5712
5812
59
1259
1215
001
1350
000
1260
1261
1262
1263
1264
1265
1266
1267
1268
1269
1270 12
7112
7212
731274 12
751276 12
7712
7812
7912
8012
8112
8212
83 1284 12
85
1286
1287
1288
1289
1290
1291
1292
1293
1294
1295
1296
1297
1298
1299
1300
1301
1302
1303
1304
1305 13
06 1307
1308
1309
1310
1311
1312
1313
1314
1315
1316
1317
1318
1319 13
2013
2113
2213
2313
2413
25 1326
1327
1328
1329
1330
1331
1332
1333
1334
1335 13
361337
1338
1339 13
4013
4113
42 1343
1344
1345
1346
1347
1348
1349
1350
1351 13
5213
5313
5413
55 1356
1357
1358
1359
1360
1361
1362
1363
1364
1365
1366 13
6713
6813
6913
7013
7113
7213
7313
7413
7513
7613
7713
7813
7913
8013
8113
8213
8313
8413
8513
86 138713
8813
8913
9013
9113
9213
9313
94 1395 1396
1397
1398
1398
1350
001
1485
000
1399 14
0014
0114
0214
03 1404
1405
1406
1407
1408
1409
1410
1411
1412
1413
1414
1415
1416 14
171418
1419
1420 14
211422
1423
1424
1425
1426
1427
1428
1429
143014
31 143214
3314
3414
35 1436
1437
1438
1439
1440
1441
1442
1443
1444 14
4514
4614
4714
4814
4914
5014
5114
5214
5314
5414
5514
5614
5714
5814
5914
6014
6114
6214
6314
6414
6514
6614
6714
6814
6914
7014
71 1472
1473
1474
1475
1476
1477
1479
1480
1481
1482
1483
1484
1485
1486
1487
1488
1489
1490
1491
1492
1493
1494
1495
1496
1497
1498
1499
1500
1501
1502
1504
1505
1506
1507
1508
1509
1510
1511 15
1215
1315
1415
1515
1615
1715
1815
19 152015
2115
2215
2315
2415
2515
2715
2815
29
1529
1485
001
1620
000
1530
1531
1532
1533
1534
1535
1536 15
3715
3815
3915
4015
411542 15
431544
1545
1546 15
4715
4815
4915
5015
5115
5215
5315
5415
5515
561557 15
581559
1560
1561
1562 15
6315
64 156515
66 156715
6815
6915
7015
7115
72 1573
1574
1575
1576
1577
1578 15
791580 15
811582
1583
1584 15
851586
1587
1588
158915
9015
9115
9215
9315
9415
951596 15
971598 15
991600
1601
1602
1603
1604
1605
1606
1607 16
0816
0916
1016
1116
12 1613
1614
1615 1616
1617
1618 16
1916
2016
2116
2216
2316
2416
2516
2616
2716
2816
2916
3016
3116
3216
3316
34 1635 16
36
1637
1638
1639
1640
1641
1642
1643 16
4416
4516
4616
4716
4816
4916
50 1651 16
521653 16
5416
5516
56 1657
1658
1659
166016
61 1662
1663
1664
1665
1666
1667
1668
166916
70 167116
72 1673
1674
1675
1676
1677
1679
1680
1681
1682
1683 16
841685 16
861687
1688
1689 16
901691
1692
1620
001
1755
000
1693 16
94 1695
1696
1697
1698
1699
1700
1701 17
021703
1704
1705
1706
1707
1708
1709
1710
1711
R17
1217
13 1714
1716
1717
G17
1817
1917
201721 17
2217
2317
2417
25 1726
1727
1728
1729
1730
1731
1732
1733
1734
1735
1736
1737
1738
1739
1740
1741
1742
1743
1744
1745
1746
1747
174
817
4917
5017
5117
5217
5317
5417
5517
57 175817
5917
6017
6117
6217
6417
6517
6617
6717
6817
6917
70
177117
7217
7317
74
1775
1776
1777 17
7817
7917
8017
8117
8217
8317
8417
8517
8617
8717
8817
891790 17
911792
1793
T17
9417
9617
9717
9817
9918
0018
0118
0218
0318
0418
05 1806
S18
0718
0818
0918
101811
1812
1813
1814
1815
1816
1817
1818
1819
1820
1821
1822
1823
1824
1825
1827
1828
1829
1830
1831
1832
1833
1834
1835
1836
1837
1838
1838
1755
001
1890
000
1839
1840
1841
1842
1843
1844
1845 18
461847
1848
1849
1850
1851
1852
1853
1854
1855
1856
1857 18
58K
1859
1860 18
61 186218
6318
6418
6518
6618
6718
681869
187
018
71 1872
1873 18
7418
7518
7618
7718
78 1879 18
80
1881
188
218
83 188418
8518
8618
8718
8818
8918
9018
9118
9218
9318
9418
9518
9618
9718
9818
9919
0019
0119
0219
0319
0419
0619
0719
0819
1019
1119
1219
1319
1419
1519
1619
1719
1919
2019
21 1922
1923
1924
1925
1926
1927
1928 19
291930
1931
1932
1933
1934
1935
1936
1937
1938 19
3919
4019
4219
4319
4419
4519
4619
4719
4819
4919
5019
5119
5219
5319
5419
5519
5619
5719
5819
5919
6019
6119
62 1963
1964
1965
1966
1967
1968
1969
1970
1971
1972
1973 19
7419
7519
7619
7719
7819
79
1979
1890
001
2025
000
1980
1981
1982
1983
1984
1985
1986
1987
1988
1989
1990
1991
1992
1993
1994
1995
1996
1997
1998
1999
2000
2001
2002 20
0320
0420
0520
06 2007
2008
2009
2010
201120
1220
1320
1420
1520
16 2017
2018
2019
2020
2021
2022
2023
2024
2025
202620
2720
30 203120
32 2033
2034
2035
2036
2037
2038
2039
2040
2041
2042
2043
2044
2045
2046
2047
2048
2049
2050
2051
2052
2053
2054 20
55 205620
57 2058
2059
2060
2061
2062 20
632064 20
652066 20
672068
2069
2070 20
71 207220
73 207420
7520
7620
7720
782079 20
802081
2082
2083
2084
2085
2086
2087
2088
2089
2090
2091
2092
2093
2094
2095
2096
2097 20
9820
9921
0021
01 2102
2103 21
042105
2106
2107
C21
08 2109
2110
2111 21
122113 21
1421
1521
16 2117
2118 21
1921
2021
2121
2221
23 2124
2125 21
2621
2721
28 2129 21
302131 21
3221
3321
34
2134
2025
001
2160
000
2135
2136
2137
2138 21
3921
4021
4121
4221
4321
4421
4521
4621
4721
4821
5021
5121
52 2153
2154
2155
S21
5621
5721
5821
5921
6021
61 2162 21
632164
2165
2166 21
67 2168
2169
2170
2171 21
722173
2174
2175
2176
2177
2178
2179
2180
2181 21
822183
2184
2185
2186
218721
88 2189
2190 21
9121
93 2194
2195
2196
2197 21
982199 22
0022
0122
0222
032204
2205
2206
2207
2208
2209
2210
2211
221222
1322
1422
1522
1622
1722
1822
1922
2022
2122
2222
2322
2422
2522
2622
2722
2822
2922
3022
3122
3222
3322
3422
3522
3622
3722
3822
3922
4122
4222
4322
4422
4522
4622
4722
4822
4922
5022
5122
52 2253 22
54
2255
2256
2257
2258
2259
2260
2261
2262 22
63 226422
6522
6622
6722
68
2268
2160
001
2295
000
2269
2270
2271
2272
2273
227422
7522
7622
7722
7822
7922
8022
8122
8222
8322
842285
2286
2287
2288
2289
2290
2291
229222
9322
9422
9522
9622
972298
2299
2300
2301
2302
2303 23
0423
0523
0623
0723
0823
09 G23
10
2311
2312
2313
2314 23
152316
2317
2318
231923
2023
2123
22 2323
2324
2325
2326
2327
2328
2329
2330
2331
2332
2333
2334 23
352336
2337
2338
2339
2340
2341
2342
2343
2344 23
452346 23
4723
4823
4923
50 235123
5223
5323
5423
5523
562357
2358
2359
2360
2361
2362
2363
2364
2365
2366
2367
2368
2369
2370
2371
2372
2373
2374
2375
2376 23
77 237823
79 238023
81 2382
2383
2384
2385
2386
2387 23
8823
8923
90 239123
9223
9323
9423
9523
9623
9723
9823
9924
00 240124
02 240324
04 240524
0624
0724
0824
0924
1024
1124
1224
1324
14
2414
2295
001
2430
000
T 2415
2416
2417
2418
2419
2420
2421
2422
2423 24
242425
2426
2427
2428
2429
2430
2431
2432
2433
2434
2435
2436 24
3724
3824
3924
4124
4224
4324
4424
4524
4624
4724
4824
4924
5024
5124
5224
5324
5424
5524
5624
57 245824
5924
6024
6124
6224
6324
6424
6524
6624
67 2468
2469
2470
2471
2472
2473
2474
2475
2476
2477
2478
2479
2480 24
8124
8224
83 2484
2485
2486
2487
2488
2489
2490 24
9124
9224
9324
94 249524
9624
9724
9824
9925
0025
0125
02 250325
0425
0525
0625
07 2508 25
09
2510
2511
2512
2513
2514
251525
1625
1725
18 2519 25
202521
2522
2523
2524
2525
2526 25
2725
2825
2925
3025
3125
3225
3325
3425
3525
3625
3725
3825
3925
40 2541
2542
2543
2544
2545
2546
2547
2548
2549
2550
2551
2552
2552
2430
001
2565
000
2553
2554
2555
2556
2557
2558
2559
256025
6125
6225
6325
64 2565 25
66
2567
2568
256925
7025
7125
7225
7325
7425
7525
7625
7725
7825
7925
80R 25
8125
8225
8325
8425
8525
8625
8725
8825
8925
9025
9125
9225
9325
9425
9525
9625
9725
9825
9926
00 2601
2602
2603
260426
0526
0626
0726
0826
0926
1026
1126
1326
1426
15 2616
2617
2618
2619
2620 2621 26
22
2623
2624
2625
2626 26
2726
2826
2926
30 2631
2632
2633
2634 26
3526
3626
3726
3826
39W26
40T GY
2641
2642
2643
2644
Q26
4526
4626
4726
4826
4926
5026
5126
52 2653 26
542655 26
5626
57 2658
2660 26
612662
2663
2664
2666
266726
68 266926
7026
7126
72 2673
2674
2675
2676
2677
2678
2679
2680
2681
2682
2683 26
8426
8526
8626
87
2687
2565
001
2679
305
2688
2689
2690
2691
2692
2693
2694
2695
2696
2697 26
9826
9927
0027
01 270227
0327
0427
0527
0627
0727
0827
0927
1027
1127
1227
1327
1427
1527
1627
17 2718 27
192720
2721
2722
2723
2724
2725
2726
2728
2729
273027
31 2732
2733
2734
2735
2736 27
37 2738
2739
2740
2741
2742
2743 27
442745
2746
2747
2748 2749
2750
2751
2752
2753
2754
2755
2756 27
572758
2759
2760
P RHK27
6127
6227
63 2764
2765
276627
67 276827
6927
70 2771
2772
2773
2774
2775
2776 27
7727
7827
7927
8027
81 2782
151
158
1 2
3 4
5 6
7 8
9 1
0 1
1 1
2 1
3 1
4 1
5 1
6 1
7 1
8 1
9 2
0 21 2
2 23
24
25 2
6 27 2
8 2
9 30
31
32
33
34 35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57 5
8 5
9 6
0 6
1 6
2 6
3 6
4
112
85
1 2
rRNA
-ope
ron
rRNA
-ope
ron
XfP1
XfP1
XfP3
XfP4
XfP4
XfP2
LR-1
LR-1
LR-2
LR-2
LR-3
LR-3
LR-4
LR-4
LR-5
LR-5
LR-6L
R-6
LR-7
LR-7
LR-7
REPE
AT-IN
-XfP
3-AN
D-Xf
P4RE
PEAT
-IN-X
fP3-
AND-
XfP4
REPE
AT-1
REPE
AT-1
REPE
AT-2
REPE
AT-2
Inte
rmed
iary
met
abol
ism
Ener
gy m
etab
olis
mRe
gula
tory
func
tions
Bios
ynth
esis
of s
mal
l mol
ecul
esAm
ino
acid
bio
synt
hesi
sN
ucle
otid
e bi
osyn
thes
is
Mac
rom
olec
ule
met
abol
ism
DNA
proc
essi
ngRN
A pr
oces
sing
Cell
stru
ctur
eUn
defin
ed c
ateg
ory
Cellu
lar p
roce
sses
Tran
spor
tM
obile
gen
etic
ele
men
tsPa
thog
enic
ity a
nd a
dapt
atio
nCo
nser
ved
hypo
thet
ical
Hypo
thet
ical
Fram
eshi
ft/Po
int m
utat
ion
(FSP
M)
Intro
n
AtR
NA (l
ette
r ind
icat
es a
min
o ac
id)
1 kb
2.44
2.96
2.58
3.58
2.65
1.38
2.89
3.72
6.13
4.41
0.83
0.79
3.27
4.92
4.61
10.95
41.87
315
403
158
160
331
474
500
502
688
543
622
627
646
871
746
878
772
903
1044
1051
918
965
1136
1715
1756
1763
1478
1795
1503
1678
1826
1526
2149
2440
2028
2029
2192
1905
1909
1918
2240
2659
2665
2727
2612
1941
tmRN
A
Mai
n Ch
rom
osom
e Ge
ne M
ap
Plas
mid
pXF
51 G
ene
Map
Plas
mid
pXF
1.3
Gene
Map
© 2000 Macmillan Magazines Ltd

articles
NATURE | VOL 406 | 13 JULY 2000 | www.nature.com 159
INTE
RMED
IARY
MET
ABOL
ISM
/ DEG
RADA
TION
/Deg
rada
tion
of s
mal
l mol
ecul
es
XF23
95ac
etyl
xyla
n es
tera
se (a
xeA)
34%
XF12
50ar
gini
ne d
eam
inas
e (ro
cF)
45%
XF14
72be
nzen
e 1,
2-di
oxyg
enas
e,fe
rredo
xin
prot
ein
(bed
B)37
%XF
0840
beta
-gal
acto
sida
se (b
ga)
71%
XF04
39be
ta-g
luco
sida
se (b
glX)
41%
XF08
46be
ta-m
anno
sida
se p
recu
rsor
35%
XF12
34ca
rbox
ypho
spho
noen
olpy
ruva
teph
osph
onom
utas
e (p
rpB)
62%
XF22
10di
oxyg
enas
e48
%XF
1743
este
rase
(est
)46
%XF
1610
fruct
okin
ase
36%
XF17
40gl
ucos
e de
hydr
ogen
ase
B (y
liI)
44%
XF10
64gl
ucos
e ki
nase
(glk
)41
%XF
1460
gluc
ose
kina
se (g
lk)
35%
XF19
65ha
loal
kane
deh
alog
enas
e (d
haA)
30%
XF26
77L-
asco
rbat
e ox
idas
e (a
ao)
29%
XF12
53lip
ase
(lipP
)46
%XF
0781
lipas
e/es
tera
se (e
stA)
25%
XF18
25N
AD(P
)H s
tero
id d
ehyd
roge
nase
(cdh
)35
%XF
1201
phos
phog
lyco
late
pho
spha
tase
(yfb
T)35
%XF
2470
phos
phog
lyco
late
pho
spha
tase
(cbb
ZC)
37%
XF22
59po
lyvi
nyla
lcoh
ol d
ehyd
roge
nase
22%
XF03
66rib
okin
ase
(rbsK
)38
%XF
0379
rubr
edox
in (r
ubA)
54%
XF18
19th
reon
ine
dehy
drat
ase
cata
bolic
(tdc
B)39
%XF
1181
triac
ylgl
ycer
ol li
pase
pre
curs
or (l
ip)
56%
XF06
10UD
P-gl
ucos
e 4-
epim
eras
e (g
alE)
27%
XF24
32UT
P-gl
ucos
e-1-
phos
phat
eur
idyl
yltra
nsfe
rase
(gta
B)82
%
INTE
RMED
IARY
MET
ABOL
ISM
/CEN
TRAL
INTE
RMED
IARY
MET
ABOL
ISM
/Am
ino
suga
rs
XF01
41gl
ucos
amin
e--fr
ucto
se-6
-pho
spha
team
inot
rans
fera
se (g
lmS)
54%
XF14
64gl
ucos
amin
e--fr
ucto
se-6
-ph
osph
atea
min
otra
nsfe
rase
47%
XF23
55N
-ace
tyl-b
eta-
gluc
osam
inid
ase
(exo
)45
%XF
1465
N-a
cety
lglu
cosa
min
e-6-
phos
phat
ede
acet
ylas
e (n
agA)
39%
INTE
RMED
IARY
MET
ABOL
ISM
/CEN
TRAL
INTE
RMED
IARY
MET
ABOL
ISM
/Ent
ner-D
oude
roff
XF10
612-
keto
-3-d
eoxy
-6-p
hosp
hogl
ucon
ate
aldo
lase
(eda
/hga
/kdg
A)41
%XF
1062
6-ph
osph
oglu
cona
te d
ehyd
rata
se (e
dd)
58%
INTE
RMED
IARY
MET
ABOL
ISM
/CEN
TRAL
INTE
RMED
IARY
MET
ABOL
ISM
/Glu
cone
ogen
esis
XF09
77m
alat
e ox
idor
educ
tase
(mae
B)57
%XF
1259
phos
phoe
nolp
yruv
ate
synt
hase
(pps
A/pp
s)64
%
INTE
RMED
IARY
MET
ABOL
ISM
/CEN
TRAL
INTE
RMED
IARY
MET
ABOL
ISM
/Non
-oxi
dativ
e br
anch
, pen
tose
pat
hway
XF02
08D-
ribul
ose-
5-ph
osph
ate
3-ep
imer
ase
(rpe/
dod)
76%
XF20
15rib
ose-
5-ph
osph
ate
isom
eras
e A
(rpiA
)57
%XF
1936
trans
keto
lase
1 (t
ktA/
tkt)
63%
INTE
RMED
IARY
MET
ABOL
ISM
/CEN
TRAL
INTE
RMED
IARY
MET
ABOL
ISM
/Nuc
leot
ide
inte
rcon
vers
ions
XF12
88CT
P sy
nthe
tase
(pyr
G)65
%XF
1058
urid
ylat
e ki
nase
(pyr
H)54
%
INTE
RMED
IARY
MET
ABOL
ISM
/CEN
TRAL
INTE
RMED
IARY
MET
ABOL
ISM
/Pho
spho
rus
com
poun
ds
XF06
57al
kalin
e ph
osph
atas
e (p
hoA)
32%
XF09
04AT
P-bi
ndin
g pr
otei
n (y
beZ)
55%
XF25
90ex
opol
ypho
spha
tase
(ppx
)42
%XF
2591
poly
phos
phat
e ki
nase
(ppk
)33
%
INTE
RMED
IARY
MET
ABOL
ISM
/CEN
TRAL
INTE
RMED
IARY
MET
ABOL
ISM
/Poo
l, m
ultip
urpo
se c
onve
rsio
ns
XF22
55ac
etyl
coe
nzym
e A
synt
heta
se (a
cs)
61%
XF10
37ad
enos
ylho
moc
yste
inas
e (a
hcY)
65%
XF08
80ca
rbon
ic a
nhyd
rase
(yad
F)46
%XF
2095
carb
onic
anh
ydra
se32
%XF
2249
deox
yxyl
ulos
e-5-
phos
phat
e sy
ntha
se (d
xs)
55%
XF18
89fe
rredo
xin-
NAD
P re
duct
ase
(fpr)
59%
XF22
68gl
ycer
ol k
inas
e (g
lpK)
59%
XF01
81gl
ycin
e cl
eava
ge H
pro
tein
(gcv
H)48
%XF
0183
glyc
ine
clea
vage
T p
rote
in (g
cvT)
55%
XF13
85gl
ycin
e de
carb
oxyl
ase
(gcv
P)57
%XF
2160
hydr
oxya
cylg
luta
thio
ne h
ydro
lase
(glo
B)45
%XF
2171
inor
gani
c py
roph
osph
atas
e (p
pa)
59%
XF13
99la
ctoy
lglu
tath
ione
lyas
e (g
loA)
63%
XF03
92m
ethi
onin
e ad
enos
yltra
nsfe
rase
(met
K)70
%XF
0913
tropi
none
redu
ctas
e43
%
INTE
RMED
IARY
MET
ABOL
ISM
/CEN
TRAL
INTE
RMED
IARY
MET
ABOL
ISM
/Sug
ar-n
ucle
otid
e bi
osyn
thes
is, c
onve
rsio
ns
XF06
09GD
P-m
anno
se 4
,6 d
ehyd
rata
se (g
md)
50%
XF22
79nu
cleo
tide
suga
r epi
mer
ase
(wbn
F)51
%XF
0260
phos
phog
luco
mut
ase/
phos
phom
anno
mut
ase
(xan
A)84
%XF
1468
phos
phom
anno
mut
ase
(mrs
A)55
%XF
0259
phos
phom
anno
se is
omer
ase-
GDP-
man
nose
pyr
opho
spho
ryla
se (x
anB)
84%
XF16
06UD
P-gl
ucos
e de
hydr
ogen
ase
(ugd
)63
%
INTE
RMED
IARY
MET
ABOL
ISM
/CEN
TRAL
INTE
RMED
IARY
MET
ABOL
ISM
/Sul
fur m
etab
olis
m
XF14
973’
-pho
spho
aden
osin
e5’
-pho
spho
sulfa
te re
duct
ase
(cys
H)57
%XF
1501
ATP
sulfu
ryla
se, l
arge
sub
unit
(nod
Q)55
%XF
1500
ATP
sulfu
ryla
se, s
mal
l sub
unit
(cys
D)67
%XF
1499
NAD
PH-s
ulfit
e re
duct
ase,
flav
opro
tein
subu
nit (
cysJ
)40
%XF
1498
NAD
PH-s
ulfit
e re
duct
ase,
iron
-sul
fur
prot
ein
(cys
I)52
%
XF01
87su
lfite
syn
thes
is p
athw
ay p
rote
in(c
ysQ/
amtA
)45
%
INTE
RMED
IARY
MET
ABOL
ISM
/EN
ERGY
MET
ABOL
ISM
,CA
RBON
/Aer
obic
resp
iratio
n
XF03
47D-
lact
ate
dehy
drog
enas
e (d
ld1)
38%
XF18
02gl
ycer
ol-3
-pho
spha
tede
hydr
ogen
ase
(gps
A)46
%XF
2266
glyc
erol
-3-p
hosp
hate
dehy
drog
enas
e (g
lpD)
54%
XF03
10N
ADH-
ubiq
uino
ne o
xido
redu
ctas
e,N
QO1
subu
nit (
nuoF
)53
%XF
0314
NAD
H-ub
iqui
none
oxi
dore
duct
ase,
NQO
10 s
ubun
it (n
uoJ)
45%
XF03
15N
ADH-
ubiq
uino
ne o
xido
redu
ctas
e,N
QO11
sub
unit
(nuo
K)69
%XF
0316
NAD
H-ub
iqui
none
oxi
dore
duct
ase,
NQO
12 s
ubun
it (n
uoL)
42%
XF03
17N
ADH-
ubiq
uino
ne o
xido
redu
ctas
e,N
QO13
sub
unit
(nuo
M)
50%
XF03
18N
ADH-
ubiq
uino
ne o
xido
redu
ctas
e,N
QO14
sub
unit
(nuo
N)
43%
XF03
09N
ADH-
ubiq
uino
ne o
xido
redu
ctas
e,N
QO2
subu
nit (
nuoE
)41
%XF
0311
NAD
H-ub
iqui
none
oxi
dore
duct
ase,
NQO
3 su
buni
t (nu
oG)
38%
XF03
08N
ADH-
ubiq
uino
ne o
xido
redu
ctas
e,N
QO4
subu
nit (
nuoD
)58
%XF
0307
NAD
H-ub
iqui
none
oxi
dore
duct
ase,
NQO
5 su
buni
t (nu
oC)
63%
XF03
06N
ADH-
ubiq
uino
ne o
xido
redu
ctas
e,N
QO6
subu
nit (
nuoB
)87
%XF
0305
NAD
H-ub
iqui
none
oxi
dore
duct
ase,
NQO
7 su
buni
t (nu
oA)
53%
XF03
12N
ADH-
ubiq
uino
ne o
xido
redu
ctas
e,N
QO8
subu
nit (
nuoH
)56
%XF
0313
NAD
H-ub
iqui
none
oxi
dore
duct
ase,
NQO
9 su
buni
t (nu
oI)
60%
INTE
RMED
IARY
MET
ABOL
ISM
/EN
ERGY
MET
ABOL
ISM
,CA
RBON
/Ana
erob
ic re
spira
tion
and
ferm
enta
tion
XF17
46al
coho
l deh
ydro
gena
se (y
ahK)
81%
XF23
89al
coho
l deh
ydro
gena
se (y
ahK)
78%
XF11
36N
ADP-
alco
hol d
ehyd
roge
nase
56%
XF17
27N
ADP-
alco
hol d
ehyd
roge
nase
(adh
)42
%XF
1734
NAD
P-al
coho
l deh
ydro
gena
se56
%
INTE
RMED
IARY
MET
ABOL
ISM
/EN
ERGY
MET
ABOL
ISM
,CA
RBON
/Ele
ctro
n tra
nspo
rt
XF24
60c-
type
cyt
ochr
ome
biog
enes
ism
embr
ane
prot
ein
(cyc
K)52
%XF
2459
c-ty
pe c
ytoc
hrom
e bi
ogen
esis
pro
tein
(cyc
J)39
%XF
2462
c-ty
pe c
ytoc
hrom
e bi
ogen
esis
pro
tein
(cyc
L)52
%XF
0620
c-ty
pe c
ytoc
hrom
e bi
ogen
esis
prot
ein
(cop
per t
oler
ance
) (ds
bD)
32%
XF24
61c-
type
cyt
ochr
ome
biog
enes
ispr
otei
n/th
iore
doxi
n (d
sbE/
ccm
G)39
%XF
1328
cyto
chro
me
B561
(yod
B)34
%XF
1360
cyto
chro
me
C ox
idas
e as
sem
bly
fact
or (c
oxD)
41%
XF13
89cy
toch
rom
e O
ubiq
uino
l oxi
dase
,su
buni
t I (c
yoB)
69%
XF13
90cy
toch
rom
e O
ubiq
uino
l oxi
dase
,su
buni
t II (
cyoA
)56
%XF
1388
cyto
chro
me
O ub
iqui
nol o
xida
se,
subu
nit I
II (c
yoC)
63%
XF13
87cy
toch
rom
e O
ubiq
uino
l oxi
dase
,su
buni
t IV
(cyo
D)41
%XF
0253
elec
tron
trans
fer f
lavo
prot
ein
alph
asu
buni
t (et
fA)
51%
XF02
54el
ectro
n tra
nsfe
r fla
vopr
otei
n be
tasu
buni
t (et
fB/e
tfS)
54%
XF12
98el
ectro
n tra
nsfe
r fla
vopr
otei
nub
iqui
none
oxi
dore
duct
ase
(etf-
QO)
46%
XF05
57el
ectro
n tra
nsfe
r pro
tein
azu
rin I
(az1
)49
%XF
0983
ferre
doxi
n56
%XF
1964
ferre
doxi
n (y
kgJ)
47%
XF26
01fe
rredo
xin
70%
XF05
47fe
rredo
xin
II (y
dgM
)54
%XF
0053
flavo
hem
opro
tein
(fhp
)40
%XF
2082
oxid
ored
ucta
se31
%XF
1990
thio
redo
xin
(yne
N)
30%
XF09
09ub
iqui
nol c
ytoc
hrom
e C
oxid
ored
ucta
se,
cyto
chro
me
B su
buni
t (pe
tB)
56%
XF09
10ub
iqui
nol c
ytoc
hrom
e C
oxid
ored
ucta
se,
cyto
chro
me
C1 s
ubun
it (p
etC)
48%
XF09
08ub
iqui
nol c
ytoc
hrom
e C
oxid
ored
ucta
se,
iron-
sulfu
r sub
unit
(pet
A)52
%
INTE
RMED
IARY
MET
ABOL
ISM
/EN
ERGY
MET
ABOL
ISM
,CA
RBON
/Gly
coly
sis
XF02
746-
phos
phof
ruct
okin
ase
(pfk
A)31
%XF
1291
enol
ase
(eno
)63
%XF
0826
fruct
ose-
bisp
hosp
hate
ald
olas
e52
%XF
0232
gluc
ose-
6-ph
osph
ate
isom
eras
e (p
gi)
79%
XF04
57gl
ycer
alde
hyde
-3-p
hosp
hate
dehy
drog
enas
e (g
apA)
68%
XF08
23ph
osph
ogly
cera
te k
inas
e (p
gk)
60%
XF18
93ph
osph
ogly
cero
mut
ase
(gpm
A/gp
m)
56%
XF08
24py
ruva
te k
inas
e ty
pe II
(pyk
A)50
%XF
0303
trios
epho
spha
te is
omer
ase
(tpiA
/tpi
)48
%
INTE
RMED
IARY
MET
ABOL
ISM
/EN
ERGY
MET
ABOL
ISM
,CA
RBON
/Oxi
dativ
e br
anch
, pen
tose
pat
hway
XF10
636-
phos
phog
luco
nola
cton
ase
(pgl
)35
%XF
1065
gluc
ose-
6-ph
osph
ate
1-de
hydr
ogen
ase
(zwf)
46%
INTE
RMED
IARY
MET
ABOL
ISM
/EN
ERGY
MET
ABOL
ISM
,CA
RBON
/Pyr
uvat
e de
hydr
ogen
ase
XF08
69di
hydr
olip
oam
ide
acet
yltra
nfer
ase
(pdh
B)52
%XF
0868
dihy
drol
ipoa
mid
e de
hydr
ogen
ase
(lpdA
/lpd)
57%
XF06
69py
ruva
te d
ehyd
roge
nase
(ace
E)56
%
INTE
RMED
IARY
MET
ABOL
ISM
/EN
ERGY
MET
ABOL
ISM
,CA
RBON
/TCA
cyc
le
XF02
92ac
onita
te h
ydra
tase
2 (a
cnB)
72%
XF15
35ci
trate
syn
thas
e (g
ltA)
61%
XF15
48di
hydr
olip
oam
ide
dehy
drog
enas
e (lp
d)58
%XF
1549
dihy
drol
ipoa
mid
e S-
succ
inyl
trans
fera
se(s
ucB)
53%
XF15
54fu
mar
ate
hydr
atas
e (fu
mC)
58%
XF18
55fu
mar
ate
hydr
atas
e (fu
mB)
30%
XF25
96is
ocitr
ate
dehy
drog
enas
e (ic
d) 4
9%XF
2700
isoc
itrat
e de
hydr
ogen
ase
(icd)
75%
XF12
11m
alat
e de
hydr
ogen
ase
(mdh
) 6
2%XF
0942
mal
ate:
quin
one
oxid
ored
ucta
se (y
ojH)
57%
XF15
50ox
oglu
tara
te d
ehyd
roge
nase
(odh
A) 5
4%XF
1073
succ
inat
e de
hydr
ogen
ase
iron-
sulfu
rpr
otei
n (s
dhB)
70%
XF10
72su
ccin
ate
dehy
drog
enas
e,fla
vopr
otei
n su
buni
t (sd
hA)
69%
XF10
70su
ccin
ate
dehy
drog
enas
e,m
embr
ane
anch
or s
ubun
it (s
dhC)
38%
XF10
71su
ccin
ate
dehy
drog
enas
e, m
embr
ane
anch
or s
ubun
it (s
dhD)
36%
XF25
48su
ccin
yl-C
oA s
ynth
etas
e,al
pha
subu
nit (
sucD
) 7
1%XF
2547
succ
inyl
-CoA
syn
thet
ase,
bet
a su
buni
t (su
cC)
58%
INTE
RMED
IARY
MET
ABOL
ISM
/EN
ERGY
MET
ABOL
ISM
,CA
RBON
/ATP
-pro
ton
mot
ive
forc
e in
terc
onve
rsio
n
XF11
49AT
P sy
ntha
se, A
cha
in (a
tpB/
uncB
/pap
D) 4
6%XF
1145
ATP
synt
hase
, alp
ha c
hain
(atp
A/un
cA)
74%
XF11
47AT
P sy
ntha
se, B
cha
in (a
tpF/
uncF
) 4
5%XF
1143
ATP
synt
hase
, bet
a ch
ain
(atp
D) 7
8%XF
1148
ATP
synt
hase
, C c
hain
(atp
E/un
cE)
56%
XF11
46AT
P sy
ntha
se, d
elta
cha
in (a
tpH/
uncH
) 3
7%XF
1142
ATP
synt
hase
, eps
ilon
chai
n (a
tpC)
46%
XF11
44AT
P sy
ntha
se, g
amm
a ch
ain
(atp
G/un
cG/p
apC)
57%
INTE
RMED
IARY
MET
ABOL
ISM
/REG
ULAT
ORY
FUN
CTIO
NS
XF12
41ac
onita
te h
ydra
tase
1 (a
cnA/
acn)
---%
XF13
16AT
P:GT
P 3’
-pyr
opho
spho
tranf
eras
e (re
lA)
42%
XF01
25ca
rbon
sto
rage
regu
lato
r (cs
rA)
84%
XF23
52co
ld s
hock
pro
tein
(sco
F) 6
9%XF
2476
extra
geni
c su
pres
sor (
suhB
/ssy
A) 4
4%XF
2342
heat
-indu
cibl
e tra
nscr
iptio
nal
repr
esso
r (hr
cA)
40%
XF01
22Le
xA re
pres
sor (
lexA
) 7
6%XF
1182
lipas
e m
odul
ator
(act
) 5
3%XF
1813
met
hano
l deh
ydro
gena
se re
gula
tory
pro
tein
53%
XF18
30ni
trile
hyd
rata
se a
ctiv
ator
46%
XF18
43ni
troge
n re
gula
tory
pro
tein
P-II
(gln
B) 8
7%XF
0352
pent
apho
spha
te g
uano
sine
-3’-
pyro
phos
phoh
ydro
lase
(spo
T) 4
7%XF
2145
phos
phat
e re
gulo
n tra
nscr
iptio
nal
regu
lato
r (ph
oU)
43%
XF12
75po
ly(h
ydro
xyal
cano
ate)
gra
nule
asso
ciat
ed p
rote
in (p
haF)
20%
XF26
91RN
A po
lym
eras
e si
gma-
32 fa
ctor
(rpo
H) 8
0%XF
1408
RNA
poly
mer
ase
sigm
a-54
fact
or (r
poN
) 6
4%XF
1350
RNA
poly
mer
ase
sigm
a-70
fact
or (r
poD)
63%
XF22
39RN
A po
lym
eras
e si
gma-
H fa
ctor
(alg
U/al
gT)
56%
XF14
07si
gma-
54 m
odul
atio
n pr
otei
n 4
4%XF
0911
strin
gent
sta
rvat
ion
prot
ein
A(s
spA/
ssp/
pog)
46%
XF09
12st
ringe
nt s
tarv
atio
n pr
otei
n B
(ssp
B) 4
2%XF
2165
trans
crip
tion-
rela
ted
prot
ein
(tex)
59%
XF09
62tra
nscr
iptio
nal r
egul
ator
(gcv
R) 2
6%XF
1749
trans
crip
tiona
l reg
ulat
or (o
pdE)
77%
XF18
58tra
nscr
iptio
nal r
egul
ator
(exs
B) 4
9%XF
2038
trans
crip
tiona
l reg
ulat
or (p
aiB)
31%
XF22
28tra
nscr
iptio
nal r
egul
ator
(alg
H) 4
5%XF
2085
trans
crip
tiona
l reg
ulat
or (A
crR
fam
ily)
23%
XF12
54tra
nscr
iptio
nal r
egul
ator
(Ara
C fa
mily
) (ar
aL)
34%
XF07
67tra
nscr
iptio
nal r
egul
ator
(Ars
R fa
mily
) (hl
yU)
34%
XF15
40tra
nscr
iptio
nal r
egul
ator
(Crp
/Fnr
fam
ily) (
clp)
85%
XF08
21tra
nscr
iptio
nal r
egul
ator
(Fur
fam
ily) (
zur)
38%
XF23
44tra
nscr
iptio
nal r
egul
ator
(Fur
fam
ily) (
fur)
85%
XF14
63tra
nscr
iptio
nal r
egul
ator
(Lac
I fam
ily)
32%
XF09
72tra
nscr
iptio
nal r
egul
ator
(Lux
R/Uh
pA fa
mily
) (ag
mR/
glpR
) 3
8%XF
2608
trans
crip
tiona
l reg
ulat
or(L
uxR/
UhpA
fam
ily) (
gacA
) 4
4%XF
0833
trans
crip
tiona
l reg
ulat
or (L
ysR
fam
ily) (
cysB
) 3
9%XF
1132
trans
crip
tiona
l reg
ulat
or (L
ysR
fam
ily) (
act)
33%
XF17
30tra
nscr
iptio
nal r
egul
ator
(Lys
R fa
mily
) (ya
fC)
35%
XF17
52tra
nscr
iptio
nal r
egul
ator
(Lys
R fa
mily
) 8
0%XF
1768
trans
crip
tiona
l reg
ulat
or (L
ysR
fam
ily) (
ycjZ
) 4
5%XF
0216
trans
crip
tiona
l reg
ulat
or (M
arR
fam
ily) (
prsX
) 22
%XF
1354
trans
crip
tiona
l reg
ulat
or (M
arR
fam
ily) (
yybA
) 28%
XF14
90tra
nscr
iptio
nal r
egul
ator
(Mar
R/Em
rR fa
mily
) 26
%XF
1996
trans
crip
tiona
l reg
ulat
or (P
bsX
fam
ily)
35%
XF00
61tra
nscr
iptio
nal r
epre
ssor
(kor
B) 2
8%XF
2062
trans
crip
tiona
l rep
ress
or (k
orC)
69%
XF19
20Tr
p op
eron
tran
scrip
tiona
l rep
ress
or (t
rpR)
36%
XF10
94try
ptop
han
repr
esso
r bin
ding
prot
ein
(wrb
A) 3
9%XF
1133
trypt
opha
n re
pres
sor b
indi
ng p
rote
in 3
3%XF
1733
trypt
opha
n re
pres
sor b
indi
ng p
rote
in 3
1%XF
1455
two-
com
pone
nt s
yste
m, h
ybrid
sens
or/r
egul
ator
y pr
otei
n 2
8%XF
0322
two-
com
pone
nt s
yste
m, r
egul
ator
ypr
otei
n (tc
tD)
45%
XF03
89tw
o-co
mpo
nent
sys
tem
, reg
ulat
ory
prot
ein
(pop
P/fe
uP/p
hoP)
54%
XF04
50tw
o-co
mpo
nent
sys
tem
, reg
ulat
ory
prot
ein
(pilH
) 5
0%XF
1113
two-
com
pone
nt s
yste
m, r
egul
ator
y pr
otei
n 3
0%XF
1626
two-
com
pone
nt s
yste
m, r
egul
ator
ypr
otei
n (a
lgR)
47%
XF18
48tw
o-co
mpo
nent
sys
tem
, reg
ulat
ory
prot
ein
(gln
G/nt
rC/g
lnT)
51%
XF23
36tw
o-co
mpo
nent
sys
tem
, reg
ulat
ory
prot
ein
(col
R)51
%XF
2534
two-
com
pone
nt s
yste
m, r
egul
ator
ypr
otei
n (c
olR)
59%
XF25
45tw
o-co
mpo
nent
sys
tem
, reg
ulat
ory
prot
ein
(pilR
)58
%XF
2578
two-
com
pone
nt s
yste
m, r
egul
ator
ypr
otei
n (a
ctR)
40%
XF25
93tw
o-co
mpo
nent
sys
tem
, reg
ulat
ory
prot
ein
(pho
B)56
%XF
0323
two-
com
pone
nt s
yste
m, s
enso
r pro
tein
(tct
E)31
%XF
0390
two-
com
pone
nt s
yste
m, s
enso
r pro
tein
(pho
Q)36
%
XF08
53tw
o-co
mpo
nent
sys
tem
, sen
sor p
rote
in (p
hs)
37%
XF09
73tw
o-co
mpo
nent
sys
tem
, sen
sor p
rote
in (f
lhS)
35%
XF16
25tw
o-co
mpo
nent
sys
tem
, sen
sor p
rote
in (a
lgZ)
40%
XF18
49tw
o-co
mpo
nent
sys
tem
, sen
sor p
rote
in (n
trB)
37%
XF25
35tw
o-co
mpo
nent
sys
tem
, sen
sor p
rote
in (c
olS)
34%
XF25
46tw
o-co
mpo
nent
sys
tem
, sen
sor p
rote
in (p
ilS)
37%
XF25
77tw
o-co
mpo
nent
sys
tem
, sen
sor p
rote
in (a
ctS)
25%
XF25
92tw
o-co
mpo
nent
sys
tem
, sen
sor p
rote
in (p
hoR)
38%
BIOS
YNTH
ESIS
OF
SMAL
L M
OLEC
ULES
/AM
INO
ACID
SBI
OSYN
THES
IS/G
luta
mat
e fa
mily
|nitr
ogen
ass
imila
tion
XF10
01ac
etyl
glut
amat
e ki
nase
(arg
B)30
%XF
1000
acet
ylor
nith
ine
deac
etyl
ase
(arg
E)28
%XF
1003
argi
nino
succ
inat
e ly
ase
(asl
)35
%XF
0999
argi
nino
succ
inat
e sy
ntha
se (a
rgG)
36%
XF10
05ga
mm
a-gl
utam
yl p
hosp
hate
redu
ctas
e (p
roA)
51%
XF10
04gl
utam
ate
5-ki
nase
46%
XF27
10gl
utam
ate
synt
hase
, alp
ha s
ubun
it(g
ltB/a
spB)
53%
XF27
09gl
utam
ate
synt
hase
, bet
a su
buni
t (gl
tD/a
spB)
55%
XF18
42gl
utam
ine
synt
heta
se (g
lnA)
64%
XF10
02N
-ace
tyl-g
amm
a-gl
utam
yl-p
hosp
hate
redu
ctas
e33
%XF
0998
orni
thin
e ca
rbam
oyltr
ansf
eras
e (a
rgF)
36%
XF27
12py
rrolin
e-5-
carb
oxyl
ate
redu
ctas
e (p
roC)
49%
XF14
27su
ccin
ylor
nith
ine
amin
otra
nsfe
rase
(arg
M/a
stC/
cstC
)48
%
BIOS
YNTH
ESIS
OF
SMAL
L M
OLEC
ULES
/AM
INO
ACID
SBI
OSYN
THES
IS/A
spar
tate
fam
ily, p
yruv
ate
fam
ily
XF01
142,
3,4,
5-te
trahy
drop
yrid
ine-
2-ca
rbox
ylat
eN
-suc
ciny
ltran
sfer
ase
(dap
D)60
%XF
1818
2-is
opro
pylm
alat
e sy
ntha
se (l
euA)
47%
XF23
753-
isop
ropy
lmal
ate
dehy
drat
ase
larg
esu
buni
t (le
uA)
63%
XF23
743-
isop
ropy
lmal
ate
dehy
drat
ase
smal
lsu
buni
t (le
uD)
79%
XF23
723-
isop
ropy
lmal
ate
dehy
drog
enas
e (le
uB)
78%
XF11
215,
10-m
ethy
lene
tetra
hydr
ofol
ate
redu
ctas
e (m
etF)
37%
XF22
725-
met
hylte
trahy
drop
tero
yltri
glut
amat
e--
hom
ocys
tein
e m
ethy
ltran
sfer
ase
(met
E)56
%XF
1821
acet
olac
tate
syn
thas
e is
ozym
e II,
larg
e su
buni
t (ilv
G)56
%XF
1473
amin
otra
nsfe
rase
(nifS
)47
%XF
2396
amin
otra
nsfe
rase
(asp
C)45
%XF
0118
aspa
ragi
ne s
ynth
ase
B (a
snB)
60%
XF13
71as
parta
te-B
-sem
iald
ehyd
ede
hydr
ogen
ase
(asd
)59
%XF
2100
aspa
rtyl/a
spar
agin
yl b
eta-
hydr
oxyl
ase
(asp
H) 2
8%XF
2443
beta
-ala
nine
syn
thet
ase
62%
XF22
25bi
func
tiona
l asp
arto
kina
se/h
omos
erin
ede
hydr
ogen
ase
I (th
rA/t
hrA1
/thr
A2)
40%
XF11
16bi
func
tiona
l dia
min
opim
elat
ede
carb
oxyl
ase/
aspa
rtate
kin
ase
(lysA
)33
%XF
1999
bran
ched
-cha
in a
min
o ac
idam
inot
rans
fera
se (i
lvE)
55%
XF08
64cy
stat
hion
ine
gam
ma-
synt
hase
(met
B)57
%XF
1481
diam
inop
imel
ate
epim
eras
e (d
apF)
45%
XF11
05di
hydr
odip
icol
inat
e re
duct
ase
(dap
B)34
%XF
0099
dihy
drox
y-ac
id d
ehyd
rata
se (i
lvD)
73%
XF09
63di
hydr
oxyd
ipic
olin
ate
synt
hase
(dap
A)42
%XF
2211
enol
ase-
phos
phat
ase
(mas
A)50
%XF
2224
hom
oser
ine
kina
se (t
hrB)
33%
XF08
63ho
mos
erin
e O-
acet
yltra
nsfe
rase
(met
2)30
%XF
2465
hom
oser
ine
O-ac
etyl
trans
fera
se34
%XF
1822
keto
l-aci
d re
duct
oiso
mer
ase
(ilvC
)59
%XF
0116
succ
inyl
-dia
min
opim
elat
ede
succ
inyl
ase
(dap
E)52
%XF
2223
thre
onin
e sy
ntha
se (t
hrC)
38%
BIOS
YNTH
ESIS
OF
SMAL
L M
OLEC
ULES
/AM
INO
ACID
SBI
OSYN
THES
IS/G
lyci
ne-s
erin
e fa
mily
|sul
fur m
etab
olis
m
XF06
03cy
stat
hion
ine
beta
-syn
thas
e (c
ysB)
31%
XF01
28cy
stei
ne s
ynth
ase
27%
XF08
31cy
stei
ne s
ynth
ase
(cys
K)51
%XF
2206
D-3-
phos
phog
lyce
rate
deh
ydro
gena
se (s
erA)
62%
XF23
26ph
osph
oser
ine
amin
otra
nsfe
rase
(ser
C)49
%XF
0946
serin
e hy
drox
ymet
hyltr
ansf
eras
e (g
lyA)
71%
BIOS
YNTH
ESIS
OF
SMAL
L M
OLEC
ULES
/AM
INO
ACID
SBI
OSYN
THES
IS/A
rom
atic
am
ino
acid
fam
ily
XF13
343-
dehy
droq
uina
te s
ynth
ase
(aro
B)47
%XF
2324
3-ph
osph
oshi
kim
ate
1-ca
rbox
yvin
yltra
nsfe
rase
(aro
E)46
%XF
0212
anth
rani
late
pho
spho
ribos
yltra
nsfe
rase
(trp
D)48
%XF
0210
anth
rani
late
syn
thas
e co
mpo
nent
I (tr
pE)
57%
XF06
74an
thra
nila
te s
ynth
ase
com
pone
nt I
(trpE
)36
%XF
1914
anth
rani
late
syn
thas
e co
mpo
nent
I (tr
pE)
37%
XF02
11an
thra
nila
te s
ynth
ase
com
pone
nt II
(trp
G)67
%XF
1915
anth
rani
late
syn
thas
e co
mpo
nent
II (t
rpG)
43%
XF00
36ar
omat
ic-a
min
o-ac
id a
min
otra
nfer
ase
(tyrB
)55
%XF
0047
cata
bolic
deh
ydro
quin
ase
(aro
Q)65
%XF
1141
chor
ism
ate
mut
ase
25%
XF23
38ch
oris
mat
e m
utas
e29
%XF
1369
chor
ism
ate
synt
hase
(aro
C)62
%XF
0213
indo
le-3
-gly
cero
l pho
spha
te s
ynth
ase
(trpC
)63
%XF
1374
N-(5
’-pho
spho
ribos
yl) a
nthr
anila
teis
omer
ase
(trpF
)43
%XF
2325
P-pr
otei
n (a
roQ/
pheA
)75
%XF
0026
phos
pho-
2-de
hydr
o-3-
deox
yhep
tona
teal
dola
se (a
roG)
61%
XF06
24sh
ikim
ate
5-de
hydr
ogen
ase
(aro
E)43
%XF
1335
shik
imat
e ki
nase
(aro
K)40
%XF
1376
trypt
opha
n sy
ntha
se a
lpha
cha
in (t
rpA)
46%
XF13
75try
ptop
han
synt
hase
bet
a ch
ain
(trpB
)65
%
BIOS
YNTH
ESIS
OF
SMAL
L M
OLEC
ULES
/AM
INO
ACID
SBI
OSYN
THES
IS/H
istid
ine
XF22
16am
idot
rans
fera
se (h
isH)
43%
XF22
20AT
P ph
osph
orib
osyl
trans
fera
se (h
isG)
43%
XF22
14cy
clas
e (h
isF)
61%
XF22
19hi
stid
inol
deh
ydro
gena
se (h
isD)
50%
XF22
18hi
stid
inol
-pho
spha
te a
min
otra
nsfe
rase
(his
C)42
%XF
2217
imid
azol
egly
cero
lpho
spha
tede
hydr
atas
e/hi
stid
inol
-pho
spha
te p
hosp
hata
sebi
func
tiona
l enz
yme
(his
B)50
%
XF22
13ph
osph
orib
osyl
-AM
Pcy
cloh
ydro
lase
/pho
spho
ribos
yl-A
TPpy
roph
osph
atas
e bi
func
tiona
len
zym
e (h
isI/h
isIE
)56
%XF
2215
phos
phor
ibos
ylfo
rmim
ino-
5-am
inoi
mid
azol
eca
rbox
amid
e rib
otid
e is
omer
ase
(his
A)44
%
BIOS
YNTH
ESIS
OF
SMAL
L M
OLEC
ULES
/NUC
LEOT
IDES
BIOS
YNTH
ESIS
/Pur
ine
ribon
ucle
otid
es
XF05
875’
-pho
spho
ribos
yl-5
-am
inoi
mid
azol
esy
nthe
tase
(pur
M/p
urG)
59%
XF05
855’
-pho
spho
ribos
ylgl
ycin
amid
etra
nsfo
rmyl
ase
(pur
N)
43%
XF02
75ad
enyl
ate
kina
se (a
dk)
47%
XF15
53ad
enyl
osuc
cina
te ly
ase
(pur
B)57
%XF
0455
aden
ylos
ucci
nate
syn
thet
ase
(pur
A)59
%XF
1949
amid
opho
spho
ribos
yltra
nsfe
rase
(pur
F)61
%XF
1975
bifu
nctio
nal p
urin
e bi
osyn
thes
ispr
otei
n (p
urH)
59%
XF24
29gl
utam
ine
amid
otra
nsfe
rase
(gua
A)68
%XF
1976
glyc
inam
ide
ribon
ucle
otid
e sy
nthe
tase
(pur
D)64
%XF
0560
GMP
synt
hase
37%
XF15
03gu
anyl
ate
kina
se (g
mk/
spoR
)52
%XF
2430
inos
ine-
5’-m
onop
hosp
hate
dehy
drog
enas
e (g
uaB)
64%
XF04
58nu
cleo
side
dip
hosp
hate
kin
ase
(ndk
)68
%XF
2644
phos
phor
ibos
yl p
yrop
hosp
hate
synt
heta
se (p
rsA/
prs)
60%
XF26
71ph
osph
orib
osyl
amin
oim
idaz
ole
carb
oxyl
ase,
ATP
ase
subu
nit (
purK
)50
%XF
2672
phos
phor
ibos
ylam
inoi
mid
azol
eca
rbox
ylas
e, c
atal
ytic
sub
unit
(pur
E)66
%XF
0205
phos
phor
ibos
ylam
inoi
mid
azol
e-su
ccin
ocar
boxa
mid
e sy
ntha
se (p
urC)
53%
XF14
23ph
osph
orib
osyl
form
ylgl
ycin
amid
ine
synt
heta
se (p
urL/
purI)
50%
BIOS
YNTH
ESIS
OF
SMAL
L M
OLEC
ULES
/NUC
LEOT
IDES
BIOS
YNTH
ESIS
/Pyr
imid
ine
ribon
ucle
otid
es
XF22
26as
parta
te c
arba
moy
ltran
sfer
ase
(pyr
B)52
%XF
1107
carb
amoy
l-pho
spha
te s
ynth
ase
larg
e ch
ain
(car
B/py
rA)
68%
XF11
06ca
rbam
oyl-p
hosp
hate
syn
thas
e sm
all
chai
n (c
arA)
68%
XF24
39cy
tidyl
ate
kina
se (c
mkA
)48
%XF
0988
dihy
droo
rota
se (p
yrC)
30%
XF25
71di
hydr
ooro
tate
deh
ydro
gena
se (p
yrD)
51%
XF01
53or
otat
e ph
osph
orib
osyl
tran
sfer
ase
(pyr
E)51
%XF
0034
orot
idin
e 5’
-pho
spha
te d
ecar
boxy
lase
(pyr
F)41
%
BIOS
YNTH
ESIS
OF
SMAL
L M
OLEC
ULES
/NUC
LEOT
IDES
BIOS
YNTH
ESIS
/2’-D
eoxy
ribon
ucle
otid
es
XF07
62de
oxyc
ytid
ine
triph
osph
ate
deam
inas
e (d
cd)
65%
XF14
41ph
osph
ohyd
rola
se (o
rfU1)
45%
XF11
96rib
onuc
leos
ide-
diph
osph
ate
redu
ctas
eal
pha
chai
n (n
rdA)
44%
XF11
97rib
onuc
leos
ide-
diph
osph
ate
redu
ctas
ebe
ta c
hain
(nrd
B)34
%XF
1448
thio
redo
xin
redu
ctas
e (tr
xB)
70%
XF05
80th
ymid
ylat
e ki
nase
39%
BIOS
YNTH
ESIS
OF
SMAL
L M
OLEC
ULES
/NUC
LEOT
IDES
BIOS
YNTH
ESIS
/Sal
vage
of n
ucle
osid
es a
nd n
ucle
otid
es
XF20
895’
-nuc
leot
idas
e42
%XF
2150
diad
enos
ine
tetra
phos
phat
ase
(apa
H)44
%XF
0150
dUTP
ase
(dut
/dna
S/so
f)59
%XF
1831
form
ylte
trahy
drof
olat
e de
form
ylas
e (p
urU)
47%
XF23
54hy
poxa
nthi
ne-g
uani
neph
osph
orib
osyl
trans
fera
se (h
pt)
27%
XF25
99ph
osph
odie
ster
ase-
nucl
eotid
epy
roph
osph
atas
e pr
ecur
sor
39%
XF23
53pu
rine
nucl
eosi
de p
hosp
hory
lase
45%
XF23
32th
ymid
ylat
e sy
ntha
se (t
hyA)
70%
BIOS
YNTH
ESIS
OF S
MAL
L MOL
ECUL
ES/S
UGAR
S AN
D SU
GAR
NUC
LEOT
IDES
BIO
SYN
THES
IS
XF12
97gl
ucon
olac
tona
se p
recu
rsor
37%
XF06
08m
anno
syltr
ansf
eras
e (m
tfA)
38%
BIOS
YNTH
ESIS
OF
SMAL
L M
OLEC
ULES
/COF
ACTO
RS,
PROS
THET
IC G
ROUP
S, C
ARRI
ERS
BIOS
YNTH
ESIS
/Bio
tin
XF13
578-
amin
o-7-
oxon
onan
oate
syn
thas
e (b
ioF)
42%
XF01
89ad
enos
ylm
ethi
onin
e-8-
amin
o-7-
oxon
onan
oate
amin
otra
nsfe
rase
(bio
A)47
%XF
1356
biot
in b
iosy
nthe
sis
prot
ein
(bio
H/bi
oB)
36%
XF00
64bi
otin
syn
thas
e (b
ioB)
60%
XF20
99bi
otin
syn
thes
is p
rote
in (b
ioC)
36%
XF03
56cy
toch
rom
e P-
450
hydr
oxyl
ase
37%
XF03
77cy
toch
rom
e P4
50-li
ke e
nzym
e (b
ioI)
37%
XF24
77de
thio
biot
in s
ynth
etas
e (b
ioD)
44%
BIOS
YNTH
ESIS
OF
SMAL
L M
OLEC
ULES
/CO
FACT
ORS,
PRO
STHE
TIC
GROU
PS,
CARR
IERS
BIO
SYN
THES
IS/F
olic
aci
d
XF02
282-
amin
o-4-
hydr
oxy-
6-hy
drox
ymet
hyld
ihyd
ropt
erid
ine
pyro
phos
phok
inas
e (fo
lK)
50%
XF14
562-
amin
o-4-
hydr
oxy-
6-hy
drox
ymet
hyld
ihyd
ropt
erid
ine
pyro
phos
phok
inas
e (fo
lK)
35%
XF24
31bi
func
tiona
l met
hyle
nete
trahy
drof
olat
ede
hydr
ogen
ase/
met
heny
ltetra
hydr
ofol
ate
cycl
ohyd
rola
se (f
olD)
58%
XF23
31di
hydr
ofol
ate
redu
ctas
e ty
pe II
I43
%XF
0436
dihy
dron
eopt
erin
ald
olas
e (fo
lB)
50%
XF00
91di
hydr
opte
roat
e sy
ntha
se (f
olP)
52%
XF19
46fo
lylp
olyg
luta
mat
e sy
ntha
se/d
ihyd
rofo
late
synt
hase
(fol
C/de
dC)
44%
XF19
83GT
P cy
cloh
ydro
lase
I (fo
lE)
52%
BIOS
YNTH
ESIS
OF
SMAL
L M
OLEC
ULES
/COF
ACTO
RS,
PROS
THET
IC G
ROUP
S, C
ARRI
ERS
BIOS
YNTH
ESIS
/Lip
oate
XF12
70lip
oate
bio
synt
hesi
s pr
otei
n B
(lipB
)55
%XF
1269
lipoi
c ac
id s
ynth
etas
e (li
pA/li
p)60
%
BIOS
YNTH
ESIS
OF
SMAL
L M
OLEC
ULES
/COF
ACTO
RS,
PROS
THET
IC G
ROUP
S, C
ARRI
ERS
BIOS
YNTH
ESIS
/Mol
ybdo
pter
in
XF04
66m
olyb
dopt
erin
bio
synt
hesi
s pr
otei
n (m
oeB)
53%
XF15
45m
olyb
dopt
erin
bio
synt
hesi
s pr
otei
n42
%
BIOS
YNTH
ESIS
OF
SMAL
L M
OLEC
ULES
/COF
ACTO
RS,
PROS
THET
IC G
ROUP
S, C
ARRI
ERS
BIOS
YNTH
ESIS
/Pan
toth
enat
e
XF02
293-
met
hyl-2
-oxo
buta
noat
ehy
drox
ymet
hyltr
ansf
eras
e (p
anB)
55%
XF02
31as
parta
te 1
-dec
arbo
xyla
se p
recu
rsor
(pan
D)53
%XF
0230
pant
oate
--bet
a-al
anin
e lig
ase
(pan
C)46
%
BIOS
YNTH
ESIS
OF
SMAL
L M
OLEC
ULES
/COF
ACTO
RS,
PROS
THET
IC G
ROUP
S, C
ARRI
ERS
BIOS
YNTH
ESIS
/Pyr
idox
ine
XF00
60py
ridox
al p
hosp
hate
bio
synt
hetic
prot
ein
(pdx
J)49
%XF
0839
pyrid
oxal
pho
spha
te b
iosy
nthe
ticpr
otei
n (p
dxA)
50%
XF13
37py
ridox
amin
e 5’
-pho
spha
te o
xida
se (f
prA)
52%
BIOS
YNTH
ESIS
OF
SMAL
L M
OLEC
ULES
/COF
ACTO
RS,
PROS
THET
IC G
ROUP
S, C
ARRI
ERS
BIOS
YNTH
ESIS
/Pyr
idin
enu
cleo
tides
XF19
24L-
aspa
rtate
oxi
dase
40%
XF19
61N
H3-d
epen
dent
NAD
syn
thet
ase
(nad
E/ad
gA)4
7%XF
1097
nico
tinat
e ph
osph
orib
osyl
trans
fera
se (p
ncB)
43%
XF19
25ni
cotin
ate-
mon
onuc
leot
ide
pyro
phos
phor
ylas
e (n
adC)
54%
XF19
23qu
inol
inat
e sy
nthe
tase
A (n
adA)
42%
BIOS
YNTH
ESIS
OF
SMAL
L M
OLEC
ULES
/COF
ACTO
RS,
PROS
THET
IC G
ROUP
S, C
ARRI
ERS
BIOS
YNTH
ESIS
/Thi
amin
XF10
481-
deox
y-D-
xylu
lose
5-p
hosp
hate
redu
ctoi
som
eras
e (d
xr)
46%
XF06
21ph
osph
omet
hylp
yrim
idin
e ki
nase
45%
XF03
78th
iam
in-p
hosp
hate
pyr
opho
spho
ryla
se (t
hiE)
41%
XF05
94th
iam
ine
bios
ynth
esis
lipo
prot
ein
ApbE
prec
urso
r (ap
bE)
33%
XF07
83th
iam
ine
bios
ynth
esis
pro
tein
(thi
G)57
%XF
1888
thia
min
e bi
osyn
thes
is p
rote
in (t
hiC/
thiA
)67
%XF
0956
thia
min
e-m
onop
hosp
hate
kin
ase
(thiL
)44
%
BIOS
YNTH
ESIS
OF
SMAL
L M
OLEC
ULES
/COF
ACTO
RS,
PROS
THET
IC G
ROUP
S, C
ARRI
ERS
BIOS
YNTH
ESIS
/Rib
ofla
vin
XF17
485-
amin
o-6-
(5-p
hosp
horib
osyl
amin
o)ur
acil
redu
ctas
e21
%XF
0954
6,7-
dim
ethy
l-8-ri
bity
llum
azin
e sy
ntha
se (r
ibH)
45%
XF09
53GT
P cy
cloh
ydro
lase
II/3
,4-d
ihyd
roxy
-2-b
utan
one
4-ph
osph
ate
synt
hase
(rib
A)52
%XF
1992
ribof
lavi
n bi
osyn
thes
is p
rote
in (r
ibA)
35%
XF24
19rib
ofla
vin
bios
ynth
esis
pro
tein
(rib
F)43
%XF
0952
ribof
lavi
n sy
ntha
se a
lpha
cha
in (r
ibE)
45%
XF09
50rib
ofla
vin-
spec
ific
deam
inas
e (ri
bD/r
ibG)
51%
BIOS
YNTH
ESIS
OF
SMAL
L M
OLEC
ULES
/COF
ACTO
RS,
PROS
THET
IC G
ROUP
S, C
ARRI
ERS
BIOS
YNTH
ESIS
/Thi
ored
oxin
,gl
utar
edox
in, g
luta
thio
ne
XF09
84ga
mm
a-gl
utam
yltra
nspe
ptid
ase
(ggt
)39
%XF
1428
glut
amat
e-cy
stei
ne li
gase
pre
curs
or (g
sh1)
57%
XF25
95gl
utar
edox
in (g
rxC)
53%
XF23
94gl
utar
edox
in-li
ke p
rote
in54
%XF
1956
glut
athi
one
synt
heta
se (g
shB/
gsh-
II)53
%XF
1199
thio
redo
xin
(trxA
)34
%XF
2174
thio
redo
xin
(ybb
N)
31%
XF26
98th
iore
doxi
n (tr
xA/t
snC/
fipA)
49%
BIOS
YNTH
ESIS
OF
SMAL
L M
OLEC
ULES
/COF
ACTO
RS,
PROS
THET
IC G
ROUP
S, C
ARRI
ERS
BIOS
YNTH
ESIS
/Men
aqui
none
, ubi
quin
one
XF08
352-
octa
pren
yl-6
-met
hoxy
phen
olhy
drox
ylas
e (u
biH/
visB
)34
%XF
2471
3-de
met
hylu
biqu
inon
e-9
3-m
ethy
ltran
sfer
ase
(ubi
G/pu
fX)
49%
XF06
61ge
rany
ltran
stra
nsfe
rase
(far
nesy
l-dip
hosp
hate
synt
hase
) (is
pA)
54%
XF00
68hy
drox
yben
zoat
e oc
tapr
enyl
trans
fera
se(u
biA/
cyr)
52%
XF13
91oc
tapr
enyl
-dip
hosp
hate
syn
thas
e (is
pB/c
el)
49%
XF15
38ub
iqui
none
bio
synt
hesi
s pr
otei
n (c
oq7)
29%
XF18
33ub
iqui
none
bio
synt
hesi
s pr
otei
n (a
arF)
43%
XF14
87ub
iqui
none
/men
aqui
none
tran
sfer
ase
(ubi
E)58
%
BIOS
YNTH
ESIS
OF
SMAL
L M
OLEC
ULES
/COF
ACTO
RS,
PROS
THET
IC G
ROUP
S, C
ARRI
ERS
BIOS
YNTH
ESIS
/Hem
e,po
rphy
rin
XF00
17co
prop
orph
yrin
ogen
III o
xida
se, a
erob
ic(h
emF)
63%
XF23
06de
lta-a
min
olev
ulin
ic a
cid
dehy
drat
ase
(hem
B)65
%XF
0566
ferro
chel
atas
e (h
emH)
48%
XF23
02gl
utam
ate-
1-se
mia
ldeh
yde
2,1-
amin
omut
ase
(hem
L)80
%XF
2648
glut
amyl
-tRN
A re
duct
ase
(hem
A)76
%XF
1627
hydr
oxym
ethy
lbila
ne s
ynth
ase
(hem
C)52
%XF
1797
porp
hyrin
bio
synt
hesi
s pr
otei
n (h
emY)
20%
XF15
12pr
otop
orph
yrin
ogen
oxi
dase
(hem
K)45
%XF
0832
siro
hem
e sy
ntha
se (c
ysG)
45%
XF13
32ur
opor
phyr
inog
en d
ecar
boxy
lase
(hem
E)54
%XF
1799
urop
orph
yrin
ogen
-III s
ynth
ase
(hem
D)27
%
BIOS
YNTH
ESIS
OF
SMAL
L M
OLEC
ULES
/COF
ACTO
RS,
PROS
THET
IC G
ROUP
S, C
ARRI
ERS
BIOS
YNTH
ESIS
/Cob
alam
in
XF18
86ph
osph
ogly
cera
te m
utas
e (p
gmA)
32%
BIOS
YNTH
ESIS
OF
SMAL
L M
OLEC
ULES
/COF
ACTO
RS,
PROS
THET
IC G
ROUP
S, C
ARRI
ERS
BIOS
YNTH
ESIS
/Bio
pter
in
XF01
936-
pyru
voyl
tetra
hydr
obio
pter
in s
ynth
ase
(ygc
M)
63%
XF14
57pt
erid
ine
redu
ctas
e 1
(ptr1
/ltdH
)37
%XF
2604
pter
in-4
-alp
ha-c
arbi
nola
min
ede
hydr
atas
e (p
hhB)
34%
BIOS
YNTH
ESIS
OF
SMAL
L M
OLEC
ULES
/COF
ACTO
RS,
PROS
THET
IC G
ROUP
S, C
ARRI
ERS
BIOS
YNTH
ESIS
/Oth
ers
XF19
16co
enzy
me
F390
syn
thet
ase
25%
BIOS
YNTH
ESIS
OF
SMAL
L M
OLEC
ULES
/FAT
TY A
CID
AND
PHOS
PHAT
IDIC
ACI
D BI
OSYN
THES
IS
XF10
44(3
r)-hy
drox
ymyr
isto
yl A
CP d
ehyd
rase
(fab
Z)49
%XF
2269
3-al
pha-
hydr
oxys
tero
id d
ehyd
roge
nase
34%
XF01
733-
oxoa
cyl-[
ACP]
redu
ctas
e42
%XF
0671
3-ox
oacy
l-[AC
P] re
duct
ase
(fabG
)66
%XF
0673
3-ox
oacy
l-[AC
P] s
ynth
ase
II (fa
bF)
60%
XF19
703-
oxoa
cyl-[
ACP]
syn
thas
e III
24%
XF03
19ac
etoa
cety
l-CoA
redu
ctas
e (p
hbB)
65%
XF02
03ac
etyl
-coe
nzym
e A
carb
oxyl
ase
carb
oxyl
trans
fera
se s
ubun
it al
pha
(acc
A)64
%XF
1467
acet
yl-c
oenz
yme
A ca
rbox
ylas
e ca
rbox
yltra
nsfe
rase
sub
unit
beta
(acc
D/de
dB/u
sg)
62%
XF06
72ac
yl c
arrie
r pro
tein
(acp
P)72
%XF
0771
acyl
car
rier p
rote
in (a
cpC)
40%
XF05
72be
ta-h
ydro
xyde
cano
yl-A
CP d
ehyd
rata
se(fa
bA)
63%
XF16
39be
ta-k
etoa
cyl-[
ACP]
syn
thas
e II
(fabF
/fab
J)34
%XF
1817
beta
-ket
oacy
l-[AC
P] s
ynth
ase
III (f
abH)
62%
XF00
48bi
otin
car
boxy
l car
rier p
rote
in o
fac
etyl
-CoA
car
boxi
lase
(acc
B/fa
bE)
53%
XF00
49bi
otin
car
boxy
lase
sub
unit
ofac
etyl
CoA
car
boxy
lase
(acc
C/fa
bG)
76%
XF10
87ca
rdio
lipin
syn
thas
e33
%XF
1209
card
iolip
in s
ynth
ase
(cls
/nov
)34
%XF
2716
keto
redu
ctas
e29
%XF
0670
mal
onyl
CoA
-ACP
tran
sacy
lase
(fab
D)51
%XF
1049
phos
phat
idat
e cy
tidyl
yltra
nsfe
rase
(cds
A/cd
s)36
%XF
1365
phos
phat
idyl
serin
e de
carb
oxyl
ase
(psd
)44
%
BIOS
YNTH
ESIS
OF
SMAL
L M
OLEC
ULES
/POL
YAM
INES
BIOS
YNTH
ESIS
XF01
44bi
osyn
thet
ic a
rgin
ine
deca
rbox
ylas
e (s
peA)
45%
XF15
39S-
aden
osyl
met
hion
ine
deca
rbox
ylas
epr
oenz
yme
(spe
D)66
%XF
0143
sper
mid
ine
synt
hase
(spe
E)41
%
MAC
ROM
OLEC
ULE
MET
ABOL
ISM
/DN
AM
ETAB
OLIS
M/R
eplic
atio
n
XF26
80AT
P-de
pend
ent D
NA
helic
ase
(rep)
43%
XF08
82AT
P-de
pend
ent h
elic
ase
(yoa
A)43
%XF
1229
ATP-
depe
nden
t hel
icas
e43
%XF
0001
chro
mos
omal
repl
icat
ion
initi
ator
(dna
A)53
%XF
2552
DNA
gyra
se s
ubun
it A
(gyr
A)57
%XF
0005
DNA
gyra
se s
ubun
it B
(gyr
B)60
%XF
2556
DNA
ligas
e (li
gA/li
g/dn
aL/p
deC/
lop)
46%
XF11
03DN
A po
lym
eras
e I (
polA
/res
A)52
%XF
0136
DNA
poly
mer
ase
III h
oloe
nzym
e ch
i sub
unit
(hol
C)34
%XF
1807
DNA
poly
mer
ase
III s
ubun
it(d
naX/
dnaZ
/dna
ZX)
40%
XF02
04DN
A po
lym
eras
e III
, alp
ha c
hain
(dna
E/po
lC)
48%
XF00
02DN
A po
lym
eras
e III
, bet
a ch
ain
(dna
N)
51%
XF06
76DN
A po
lym
eras
e III
, del
ta s
ubun
it (h
olB)
35%
XF21
78DN
A po
lym
eras
e III
, del
ta s
ubun
it (h
olA)
29%
XF21
57DN
A po
lym
eras
e III
, eps
ilon
chai
n (d
naQ)
49%
XF04
30DN
A pr
imas
e (d
naG/
dnaP
/par
B)46
%XF
2025
DNA
prim
ase
(traC
)56
%XF
0920
DNA
topo
isom
eras
e I (
topA
)41
%XF
1776
DNA
topo
isom
eras
e III
(top
B)48
%XF
0149
DNA/
pant
othe
nate
met
abol
ism
flavo
prot
ein
(dfp
)51
%XF
1935
gluc
ose
inhi
bite
d di
visi
on p
rote
in (g
idB)
45%
XF21
06gl
ucos
e in
hibi
ted
divi
sion
pro
tein
A (g
idA)
65%
XF13
83he
licas
e, A
TP d
epen
dent
(hrp
A)50
%XF
2689
prim
osom
al p
rote
in N
’ (pr
iA)
44%
XF03
61re
plic
ativ
e DN
A he
licas
e (d
naB/
groP
/grp
A)53
%XF
1127
TldD
pro
tein
(tld
D)61
%XF
1353
topo
isom
eras
e IV
sub
unit
(par
C)61
%XF
1286
topo
isom
eras
e IV
sub
unit
B (p
arE)
64%
MAC
ROM
OLEC
ULE
MET
ABOL
ISM
/DN
AM
ETAB
OLIS
M/S
truct
ural
DN
A bi
ndin
g pr
otei
ns
XF25
58ch
rom
osom
e se
greg
atio
n pr
otei
n (s
mc)
27%
XF04
46DN
A-bi
ndin
g pr
otei
n (b
bh3)
33%
XF19
98DN
A-bi
ndin
g pr
otei
n (fi
s)58
%XF
1190
hist
one-
like
prot
ein
(hbs
/hbs
U)64
%XF
1943
hist
one-
like
prot
ein
54%
XF04
82si
ngle
-stra
nded
DN
A bi
ndin
g pr
otei
n (s
sb)
49%
XF13
92si
ngle
-stra
nded
DN
A bi
ndin
g pr
otei
n (s
sb)
56%
XF15
58si
ngle
-stra
nded
DN
A bi
ndin
g pr
otei
n (s
sb)
36%
XF16
44si
ngle
-stra
nded
DN
A bi
ndin
g pr
otei
n (s
sb)
54%
XF17
78si
ngle
-stra
nded
DN
A bi
ndin
g pr
otei
n (s
sb)
25%
MAC
ROM
OLEC
ULE
MET
ABOL
ISM
/DN
AM
ETAB
OLIS
M/R
ecom
bina
tion
XF03
54AT
P-de
pend
ent D
NA
helic
ase
(recG
)55
%XF
1381
DNA
helic
ase
54%
XF22
48DN
A re
pair
prot
ein
RecO
(rec
O)35
%XF
0003
DNA
repl
icat
ion
and
repa
ir Re
cF p
rote
in(re
cF/u
vrF)
32%
XF18
92en
donu
clea
se V
(deo
xyin
osin
e3’
endo
nucl
ease
) (nf
i)53
%XF
0425
exod
eoxy
ribon
ucle
ase
V al
pha
chai
n (re
cD)
38%
XF04
23ex
odeo
xyrib
onuc
leas
e V
beta
cha
in(re
cB/r
orA)
30%
XF04
22ex
odeo
xyrib
onuc
leas
e V
gam
ma
chai
n (re
cC)
29%
XF14
25in
tegr
ase/
reco
mbi
nase
(xer
D/xp
rB)
53%
XF07
43in
tegr
atio
n ho
st fa
ctor
, alp
ha s
ubun
it (h
imA)
73%
XF24
37in
tegr
atio
n ho
st fa
ctor
, bet
a su
buni
t (hi
mD)
72%
XF18
09re
com
bina
tion
prot
ein
(recR
)54
%XF
2343
reco
mbi
natio
n pr
otei
n N
(rec
N)
38%
XF01
23re
com
bina
tion
prot
ein
RecA
(rec
A)91
%XF
1110
sing
le-s
trand
ed D
NA
exon
ucle
ase
(recJ
)45
%XF
1483
site
-spe
cific
reco
mbi
nase
(sss
/xer
C)53
%XF
2028
site
-spe
cific
reco
mbi
nase
(rin
)81
%
MAC
ROM
OLEC
ULE
MET
ABOL
ISM
/DN
A M
ETAB
OLIS
M/R
epai
r
XF25
986-
O-m
ethy
lgua
nine
-DN
A m
ethy
ltran
sfer
ase
(dat
/dat
1)46
%XF
1262
7,8-
dihy
dro-
8-ox
ogua
nine
-trip
hosp
hata
se(m
utX)
38%
XF19
09A/
G-sp
ecifi
c ad
enin
e gl
ycos
ylas
e(m
utY/
mut
B)46
%XF
0050
DNA
helic
ase
II (u
vrD/
mut
U/pd
eB/r
ad/r
ecL)
56%
XF07
60DN
A m
ism
atch
repa
ir pr
otei
n M
utL
(mut
L)42
%XF
0148
DNA
repa
ir pr
otei
n (ra
dC)
41%
XF13
78DN
A re
pair
prot
ein
(radA
/sm
s)61
%XF
1299
DNA
repa
ir sy
stem
spe
cific
for a
lkyl
ated
DNA
(alk
B)28
%XF
1326
DNA-
3-m
ethy
lade
nine
gly
cosi
dase
(mag
1)35
%XF
1806
DNA-
dam
age-
indu
cibl
e pr
otei
n (d
inD/
pcsA
)---
%XF
2081
DNA-
dam
age-
indu
cibl
e pr
otei
n (d
inJ)
57%
XF06
47en
donu
clea
se II
I (nt
h)67
%XF
2426
exci
nucl
ease
ABC
sub
unit
A (u
vrA)
70%
XF09
67ex
cinu
clea
se A
BC s
ubun
it B
(uvr
B)83
%XF
2311
exci
nucl
ease
ABC
sub
unit
C (u
vrC)
50%
XF01
64ex
odeo
xyrib
onuc
leas
e43
%XF
2022
exod
eoxy
ribon
ucle
ase
I (sb
cB/x
onA/
cpeA
)43
%XF
1933
exod
eoxy
ribon
ucle
ase
II I (
xthA
)46
%XF
0660
exod
eoxy
ribon
ucle
ase
smal
l sub
unit
(xse
B)39
%XF
0755
exod
eoxy
ribon
ucle
ase
VII l
arge
sub
unit
(xse
A)41
%XF
0071
form
amid
opyr
imid
ine
DNA
glyc
osyl
ase
(mut
M/f
pg)
51%
XF01
70fo
rmam
idop
yrim
idin
e DN
A gl
ycos
ylas
e(m
utM
/fpg
)51
%XF
1902
holli
day
junc
tion
bind
ing
prot
ein,
DNA
helic
ase
(ruvB
)69
%XF
1904
holli
day
junc
tion
bind
ing
prot
ein,
DNA
helic
ase
(ruvA
)42
%XF
1905
holli
day
junc
tion
reso
lvas
e,en
dode
oxyr
ibon
ucle
ase
(ruvC
)52
%XF
1716
mis
mat
ch re
pair
prot
ein
(mut
S)54
%XF
0045
trans
crip
tion-
repa
ir co
uplin
g fa
ctor
(mfd
)49
%XF
2692
urac
il-DN
A gl
ycos
ylas
e (u
ng)
55%
MAC
ROM
OLEC
ULE
MET
ABOL
ISM
/DN
AM
ETAB
OLIS
M/R
estri
ctio
n, m
odifi
catio
n
XF13
68ad
enin
e-sp
ecifi
c m
ethy
lase
46%
XF22
97DN
A m
ethy
lase
40%
XF06
41DN
A m
ethy
ltran
sfer
ase
(sfiI
M)
31%
XF17
74DN
A m
ethy
ltran
sfer
ase
(gen
e w
ith in
tron)
---%
XF09
24DN
A pr
oces
sing
cha
in A
(sm
f/dp
rA)
37%
XF09
35m
ethy
ltran
sfer
ase
37%
XF19
68m
ethy
ltran
sfer
ase
(kpn
IM)
33%
XF18
04si
te-s
peci
fic D
NA-
met
hyltr
ansf
eras
e (s
phIM
)39
%XF
2313
site
-spe
cific
DN
A-m
ethy
ltran
sfer
ase
41%
XF27
24ty
pe I
rest
rictio
n-m
odifi
catio
n sy
stem
(hsd
M)
25%
XF02
97ty
pe I
rest
rictio
n-m
odifi
catio
n sy
stem
DNA
met
hyla
se---
%XF
2723
type
I re
stric
tion-
mod
ifica
tion
syst
emDN
A m
ethy
lase
(hsd
M)
44%
XF27
28ty
pe I
rest
rictio
n-m
odifi
catio
n sy
stem
DNA
met
hyla
se61
%XF
2742
type
I re
stric
tion-
mod
ifica
tion
syst
emDN
A m
ethy
lase
(hsd
M)
53%
XF02
95ty
pe I
rest
rictio
n-m
odifi
catio
n sy
stem
endo
nucl
ease
---%
XF27
21ty
pe I
rest
rictio
n-m
odifi
catio
n sy
stem
endo
nucl
ease
(hsd
R1)
20%
XF27
25ty
pe I
rest
rictio
n-m
odifi
catio
n sy
stem
endo
nucl
ease
53%
XF27
39ty
pe I
rest
rictio
n-m
odifi
catio
n sy
stem
endo
nucl
ease
(hsd
R)42
%XF
0296
type
I re
stric
tion-
mod
ifica
tion
syst
emsp
ecifi
city
det
erm
inan
t27
%XF
2722
type
I re
stric
tion-
mod
ifica
tion
syst
emsp
ecifi
city
det
erm
inan
t21
%XF
2726
type
I re
stric
tion-
mod
ifica
tion
syst
emsp
ecifi
city
det
erm
inan
t (hs
dS)
24%
XF27
41ty
pe I
rest
rictio
n-m
odifi
catio
n sy
stem
spec
ifici
ty d
eter
min
ant (
hsdS
)24
%
MAC
ROM
OLEC
ULE
MET
ABOL
ISM
/RN
AM
ETAB
OLIS
M/R
ibos
omal
pro
tein
s
XF24
3830
S rib
osom
al p
rote
in S
1 (rp
sA/s
syF)
67%
XF11
5130
S rib
osom
al p
rote
in S
10 (r
psJ)
72%
XF11
7430
S rib
osom
al p
rote
in S
11 (r
psK)
86%
XF26
3130
S rib
osom
al p
rote
in S
12 (r
psL)
81%
XF11
7330
S rib
osom
al p
rote
in S
13 (r
psM
)85
%XF
1165
30S
ribos
omal
pro
tein
S14
(rps
N)
62%
XF02
3830
S rib
osom
al p
rote
in S
15 (r
psO/
secC
)62
%XF
0107
30S
ribos
omal
pro
tein
S16
(rps
P)65
%XF
1161
30S
ribos
omal
pro
tein
S17
(rps
Q)51
%XF
2560
30S
ribos
omal
pro
tein
S18
(rps
R)70
%XF
1156
30S
ribos
omal
pro
tein
S19
(rps
S)60
%XF
2580
30S
ribos
omal
pro
tein
S2
(rpsB
)60
%XF
2421
30S
ribos
omal
pro
tein
S20
(rps
T)54
%XF
0434
30S
ribos
omal
pro
tein
S21
(rps
U)67
%XF
1158
30S
ribos
omal
pro
tein
S3
(rpsC
)67
%XF
1175
30S
ribos
omal
pro
tein
S4
(rpsD
)84
%XF
1169
30S
ribos
omal
pro
tein
S5
(rpsE
/spc
)71
%XF
2561
30S
ribos
omal
pro
tein
S6
(rpsF
)67
%XF
2630
30S
ribos
omal
pro
tein
S7
(rpsG
/rps
7)60
%XF
1166
30S
ribos
omal
pro
tein
S8
(rpsH
/rps
8)56
%XF
1536
30S
ribos
omal
pro
tein
S9
(rpsI
/rps
9)68
%XF
2636
50S
ribos
omal
pro
tein
L1
(rplA
/rpl
1)64
%XF
2635
50S
ribos
omal
pro
tein
L10
(rpl
J)43
%XF
2637
50S
ribos
omal
pro
tein
L11
(rpl
K/re
lC)
69%
XF15
3750
S rib
osom
al p
rote
in L
13 (r
plM
)71
%XF
1162
50S
ribos
omal
pro
tein
L14
(rpl
N)
80%
XF11
7150
S rib
osom
al p
rote
in L
15 (r
plO)
48%
XF11
5950
S rib
osom
al p
rote
in L
16 (r
plP)
69%
XF11
7750
S rib
osom
al p
rote
in L
17 (r
plQ)
80%
XF11
6850
S rib
osom
al p
rote
in L
18 (r
plR)
53%
XF01
1050
S rib
osom
al p
rote
in L
19 (r
plS)
65%
XF11
5550
S rib
osom
al p
rote
in L
2 (rp
lB)
69%
XF07
4050
S rib
osom
al p
rote
in L
20 (r
plT)
72%
XF24
2450
S rib
osom
al p
rote
in L
21 (r
plU)
53%
XF11
5750
S rib
osom
al p
rote
in L
22 (r
plV)
63%
XF11
5450
S rib
osom
al p
rote
in L
23 (r
plW
)52
%XF
1163
50S
ribos
omal
pro
tein
L24
(rpl
X)48
%XF
2643
50S
ribos
omal
pro
tein
L25
(rpl
Y)48
%XF
2423
50S
ribos
omal
pro
tein
L27
(rpm
A)64
%XF
1206
50S
ribos
omal
pro
tein
L28
(rpm
B)71
%XF
1160
50S
ribos
omal
pro
tein
L29
(rpm
C)50
%XF
1152
50S
ribos
omal
pro
tein
L3
(rplC
/rpl
3)61
%XF
1170
50S
ribos
omal
pro
tein
L30
(rpm
D)47
%XF
1534
50S
ribos
omal
pro
tein
L31
(rpm
E)57
%XF
1816
50S
ribos
omal
pro
tein
L32
(rpm
F)75
%XF
1207
50S
ribos
omal
pro
tein
L33
(rpm
G)73
%XF
2782
50S
ribos
omal
pro
tein
L34
(rpm
H)76
%XF
0739
50S
ribos
omal
pro
tein
L35
(rpm
I)60
%XF
2440
50S
ribos
omal
pro
tein
L36
(rpm
J)65
%XF
1153
50S
ribos
omal
pro
tein
L4
(rplD
)54
%XF
1164
50S
ribos
omal
pro
tein
L5
(rplE
/rpl
5)63
%XF
1167
50S
ribos
omal
pro
tein
L6
(rplF
)52
%XF
2634
50S
ribos
omal
pro
tein
L7/
L12
(rplL
)67
%XF
2559
50S
ribos
omal
pro
tein
L9
(rplI)
52%
MAC
ROM
OLEC
ULE
MET
ABOL
ISM
/RN
AM
ETAB
OLIS
M/R
ibos
omes
- m
atur
atio
n an
d m
odifi
catio
n
XF01
0816
S rR
NA
proc
essi
ng p
rote
in (r
imM
)36
%XF
2607
ribon
ucle
ase
E (rn
e/am
s/hm
p1)
43%
XF11
25rib
onuc
leas
e G
(caf
A/rn
g)53
%XF
0939
ribos
omal
larg
e su
buni
t pse
udou
ridin
esy
ntha
se D
(rlu
D/sf
hB)
54%
XF22
01rib
osom
al p
rote
in L
11 m
ethy
ltran
sfer
ase
(prm
A)46
%XF
2532
ribos
omal
pro
tein
S6
mod
ifica
tion
prot
ein
(rim
K)---
%XF
1200
ribos
omal
sm
all s
ubun
it ps
eudo
urid
ine
synt
hase
(rsu
A)42
%XF
0236
ribos
omal
-bin
ding
fact
or A
(rbf
A)35
%XF
0441
ribos
omal
-pro
tein
-ala
nine
acet
yltra
nsfe
rase
(rim
I)38
%XF
2358
RNA
met
hyltr
ansf
eras
e (y
gcA)
38%
XF19
72tR
NA/
rRN
A m
ethy
lase
(yib
K)57
%XF
1985
tRN
A/rR
NA
met
hyltr
ansf
eras
e (y
jfH)
50%
MAC
ROM
OLEC
ULE
MET
ABOL
ISM
/RN
AM
ETAB
OLIS
M/A
min
oacy
l tRN
A sy
nthe
tase
s,tR
NA
mod
ifica
tion
XF01
24al
anyl
-tRN
A sy
nthe
tase
(ala
S/lo
vB)
56%
XF01
47ar
giny
l-tRN
A sy
nthe
tase
(arg
S)40
%XF
2563
aspa
ragi
nyl-t
RNA
synt
heta
se (a
snS/
tss)
66%
XF18
56as
party
l-tRN
A sy
nthe
tase
(asp
S)53
%XF
0995
cyst
einy
l-tRN
A sy
nthe
tase
(cys
S)46
%XF
1338
glut
amin
yl-tR
NA
synt
heta
se (g
lnS)
54%
XF08
22gl
utam
yl-tR
NA
synt
heta
se (g
ltX)
54%
XF19
60gl
ycyl
-tRN
A sy
nthe
tase
alp
ha c
hain
(gly
Q/gl
yS(A
))74
%XF
1959
glyc
yl-tR
NA
synt
heta
se b
eta
chai
n(g
lyS/
glyS
(B))
39%
XF22
22hi
stid
yl-tR
NA
synt
heta
se (h
isS)
36%
XF24
18is
oleu
cyl-t
RNA
synt
heta
se (i
leS/
ilvS)
54%
XF21
76le
ucyl
-tRN
A sy
nthe
tase
(leu
S)58
%XF
1112
lysy
l-tRN
A sy
nthe
tase
(lys
U)56
%XF
2555
lysy
l-tRN
A sy
nthe
tase
(yje
A/ge
nX)
47%
XF09
27m
ethi
onyl
-tRN
A fo
rmyl
trans
fera
se (f
mt)
56%
XF05
49m
ethi
onyl
-tRN
A sy
nthe
tase
(met
G)54
%XF
0742
phen
ylal
anyl
-tRN
A si
nthe
tase
bet
a ch
ain
(phe
T)42
%XF
0741
phen
ylal
anyl
-tRN
A sy
nthe
tase
alp
hach
ain
(phe
S)59
%XF
0445
prol
yl-tR
NA
synt
heta
se (p
roS/
drpA
)55
%XF
0751
ribon
ucle
ase
D (rn
d)31
%XF
2781
ribon
ucle
ase
P (rn
pA)
37%
XF13
14S-
aden
osyl
met
hion
ine:
tRN
Arib
osyl
trans
fera
se-is
omer
ase
(que
A)52
%XF
2286
sery
l-tRN
A sy
nthe
tase
(ser
S)59
%XF
0736
thre
onyl
-tRN
A sy
nthe
tase
(thr
S)59
%XF
0109
tRN
A (g
uani
ne-N
1-)-m
ethy
ltran
sfer
ase
(trm
D)51
%XF
1362
tRN
A ad
enyl
yltra
nsfe
rase
(cca
)52
%XF
0090
tRN
A de
lta(2
)-iso
pent
enyl
pyro
phos
phat
etra
nsfe
rase
(mia
A/trp
X)49
%XF
1440
tRN
A m
ethy
ltran
sfer
ase
(trm
U/as
uE)
55%
XF13
73tR
NA
pseu
dour
idin
e sy
ntha
se A
(truA
/his
T/as
uC/le
uK)
52%
XF02
37tR
NA
pseu
dour
idin
e sy
ntha
se B
(tru
B)42
%XF
2475
tRN
A/rR
NA
met
hyltr
ansf
eras
e41
%XF
0428
trypt
opha
nyl-t
RNA
synt
heta
se34
%XF
0169
tyro
syl-t
RNA
synt
heta
se (t
yrS)
53%
XF01
34va
lyl-t
RNA
synt
heta
se (v
alS)
51%
MAC
ROM
OLEC
ULE
MET
ABOL
ISM
/RN
A M
ETAB
OLIS
M/R
NA
synt
hesi
s, m
odifi
catio
n, D
NA
trans
crip
tion
XF01
92AT
P-de
pend
ent R
NA
helic
ase
(rhlE
)52
%XF
0252
ATP
-dep
ende
nt R
NA
helic
ase
(dea
D)55
%XF
2696
ATP-
depe
nden
t RN
A he
licas
e (rh
lB/m
mrA
)52
%XF
0234
N u
tiliza
tion
subs
tanc
e pr
otei
n A
(nus
A)56
%XF
0227
poly
nucl
eotid
e ad
enyl
trans
fera
se (p
cnB)
48%
XF26
06ps
eudo
urid
ylat
e sy
ntha
se (r
luC)
48%
XF11
76RN
A po
lym
eras
e al
pha
subu
nit (
rpoA
)87
%XF
2633
RNA
poly
mer
ase
beta
sub
unit
(rpoB
/gro
N/n
itB/r
if/ro
n)67
%XF
2632
RNA
poly
mer
ase
beta
’ sub
unit
(rpoC
/tab
B)71
%XF
1502
RNA
poly
mer
ase
omeg
a su
buni
t (rp
oZ)
43%
XF26
38tra
nscr
iptio
n an
titer
min
atio
n fa
ctor
(nus
G)60
%XF
0955
trans
crip
tion
term
inat
ion
fact
or (n
usB/
ssyB
)49
%XF
2699
trans
crip
tion
term
inat
ion
fact
or R
ho(rh
o/ni
tA/p
suA/
rnsC
/tsu
)72
%XF
1108
trans
crip
tiona
l elo
ngat
ion
fact
or (g
reA)
62%
MAC
ROM
OLEC
ULE
MET
ABOL
ISM
/RN
A M
ETAB
OLIS
M/R
NA
degr
adat
ion
XF12
57ol
igor
ibon
ucle
ase
(orn
)58
%XF
0239
poly
nucl
eotid
e ph
osph
oryl
ase
(pnp
)64
%XF
2615
ribon
ucle
ase
44%
XF21
58rib
onuc
leas
e H
(rnhA
/rnh
)60
%XF
1041
ribon
ucle
ase
HII (
rnhB
)55
%XF
2246
ribon
ucle
ase
III (r
nc)
50%
XF15
05rib
onuc
leas
e PH
(rph
)63
%XF
2146
ribon
ucle
ase
T (rn
t)47
%
MAC
ROM
OLEC
ULE
MET
ABOL
ISM
/PRO
TEIN
MET
ABOL
ISM
/Tra
nsla
tion
and
mod
ifica
tion
XF10
18ar
gini
ne-tR
NA-
prot
ein
trans
fera
se (a
te1)
25%
XF14
36di
sulfi
de o
xido
redu
ctas
e (d
sbA)
38%
XF18
34di
sulp
hide
isom
eras
e30
%XF
2629
elon
gatio
n fa
ctor
G (f
usA)
70%
XF22
03el
onga
tion
fact
or P
(yei
P)39
%XF
2473
elon
gatio
n fa
ctor
P (e
fp)
63%
XF25
79el
onga
tion
fact
or T
s (ts
f)50
%XF
2628
elon
gatio
n fa
ctor
Tu
(tufA
)82
%XF
2640
elon
gatio
n fa
ctor
Tu
(tufB
)82
%XF
2553
initi
atio
n fa
ctor
eIF
-2B,
alp
hasu
buni
t-rel
ated
50%
XF14
45in
itiat
ion
fact
or IF
-1 (i
nfA)
73%
XF02
35in
itiat
ion
fact
or IF
-2 (i
nfB)
49%
XF07
37in
itiat
ion
fact
or IF
-3 (i
nfC)
60%
XF08
57L-
isoa
spar
tate
O-m
ethy
ltran
sfer
ase
(pcm
)47
%XF
2417
lipop
rote
in s
igna
l pep
tidas
e (ls
pA)
45%
XF22
98lo
w m
olec
ular
wei
ght p
hosp
hoty
rosi
nepr
otei
n ph
osph
atas
e (s
tp1)
40%
XF05
76m
etal
lope
ptid
ase
37%
XF01
11m
ethi
onin
e am
inop
eptid
ase
(map
)60
%XF
2649
pept
ide
chai
n re
leas
e fa
ctor
1(p
rfA/s
ueB/
uar)
61%
XF11
11pe
ptid
e ch
ain
rele
ase
fact
or 2
(prfB
)66
%XF
0174
pept
ide
chai
n re
leas
e fa
ctor
3 (p
rfC)
62%
XF19
40pe
ptid
e m
ethi
onin
e su
lfoxi
de re
duct
ase
(msr
A/pm
s)59
%XF
2642
pept
idyl
tRN
A hy
drol
ase
(pth
)49
%XF
0644
pept
idyl
-pro
lyl c
is-tr
ans
isom
eras
e (m
ip)
38%
XF06
52pe
ptid
yl-p
roly
l cis
-tran
s is
omer
ase
(sly
D)43
%XF
0838
pept
idyl
-pro
lyl c
is-tr
ans
isom
eras
e (s
urA)
32%
XF11
91pe
ptid
yl-p
roly
l cis
-tran
s is
omer
ase
(ppi
D)25
%XF
1212
pept
idyl
-pro
lyl c
is-tr
ans
isom
eras
e (p
piB/
ppi)
60%
XF16
05pe
ptid
yl-p
roly
l cis
-tran
s is
omer
ase
36%
XF04
42ph
osph
atid
yltra
nsfe
rase
(ptr)
37%
XF09
26po
lype
ptid
e de
form
ylas
e (d
ef/f
ms)
55%
XF21
56pr
otei
n ph
osph
atas
e (y
loO)
35%
XF14
46pr
otei
n tra
nsfe
rase
(aat
)51
%XF
2585
prot
ein-
L-is
oasp
arta
teO-
met
hyltr
ansf
eras
e (p
cm)
34%
XF10
51rib
osom
e re
cycl
ing
fact
or (f
rr)58
%XF
2244
sign
al p
eptid
ase
I (le
pB)
40%
XF14
37th
iol:d
isul
fide
inte
rcha
nge
prot
ein
(dsb
A)39
%XF
0353
trans
latio
n in
itiat
ion
inhi
bito
r54
%
MAC
ROM
OLEC
ULE
MET
ABOL
ISM
/PRO
TEIN
MET
ABOL
ISM
/Cha
pero
nes
XF06
1610
kDa
chap
eron
in (g
roES
/htp
A)59
%XF
0615
60kD
a ch
aper
onin
(mop
A/gr
oEL)
77%
XF14
52ch
aper
one
(lolA
)32
%XF
2233
DnaJ
pro
tein
(dna
J)41
%XF
2339
DnaJ
pro
tein
(dna
J)59
%XF
2340
DnaK
pro
tein
(dna
K/gr
pF/g
roP/
seg)
72%
XF09
91Dn
aK s
upre
ssor
48%
XF09
78he
at s
hock
pro
tein
G (h
tpG)
58%
XF23
41he
at s
hock
pro
tein
Grp
E (g
rpE)
38%
XF11
86pe
ptid
yl-p
roly
l cis
-tran
s is
omer
ase
(tig)
32%
MAC
ROM
OLEC
ULE
MET
ABOL
ISM
/PRO
TEIN
MET
ABOL
ISM
/Pro
tein
deg
rada
tion
XF22
60al
anyl
dip
eptid
yl p
eptid
ase
30%
XF01
38am
inop
eptid
ase
A/I (
pepA
/xer
B/ca
rP)
47%
XF14
88am
inop
eptid
ase
N36
%XF
2009
amin
opep
tidas
e P
(pep
P)46
%XF
1188
ATP-
depe
nden
t Clp
pro
teas
e AT
Pbi
ndin
g su
buni
t Clp
x (c
lpX/
lopC
)73
%XF
1187
ATP-
depe
nden
t Clp
pro
teas
e pr
oteo
lytic
subu
nit (
clpP
/lopP
)73
%XF
0381
ATP-
depe
nden
t Clp
pro
teas
e su
buni
t(c
lpB/
htpM
)69
%XF
1443
ATP-
depe
nden
t Clp
pro
teas
e su
buni
t(c
lpA/
lopD
)61
%XF
1189
ATP-
depe
nden
t ser
ine
prot
eina
se L
a(lo
n/ca
pR/d
eg/m
uc/lo
pA)
64%
XF27
04ca
rbox
yl-te
rmin
al p
rote
ase
(ctp
A)41
%XF
1282
carb
oxyp
eptid
ase
rela
ted
prot
ein
33%
XF01
56cy
stei
ne p
rote
ase
(cpr
6)28
%XF
0015
dipe
ptid
yl-p
eptid
ase
28%
XF14
84he
at s
hock
pro
tein
(hsl
V/ht
pO)
65%
XF14
85he
at s
hock
pro
tein
(hsl
U/ht
pI)
66%
XF04
52in
tegr
al m
embr
ane
prot
ease
(hflK
/hflA
)33
%XF
0453
inte
gral
mem
bran
e pr
otei
nase
(hflC
)36
%XF
0280
leuc
ine
amin
opep
tidas
e (p
epA)
34%
XF04
35O-
sial
ogly
copr
otei
n en
dope
ptid
ase
(gcp
)63
%XF
0127
olig
opep
tidas
e A
(prlC
/opd
A/op
tA)
50%
XF14
79pe
ptid
ase
(ptrB
/tlp
)44
%XF
1944
pept
idyl
-dip
eptid
ase
(dcp
)53
%XF
2241
perip
lasm
ic p
rote
ase
(muc
D)47
%XF
0220
prol
ine
dipe
ptid
ase
(pep
Q)33
%XF
1510
prol
ine
imin
o-pe
ptid
ase
(pip
/xap
)81
%XF
2330
prot
eina
se (s
lpD)
22%
XF02
67se
rine
prot
ease
(psp
B)28
%XF
1026
serin
e pr
otea
se (p
spB)
27%
XF18
51se
rine
prot
ease
25%
XF18
23ta
il-sp
ecifi
c pr
otea
se (p
rc/t
sp)
39%
XF08
16zin
c pr
otea
se40
%
MAC
ROM
OLEC
ULE
MET
ABOL
ISM
/OTH
ER M
ACRO
MOL
ECUL
ESM
ETAB
OLIS
M/P
olys
acch
arid
es
XF27
14al
pha-
L-fu
cosi
dase
(fuc
A1)
36%
XF22
78do
licho
l-pho
spha
te m
anno
syltr
ansf
eras
e(d
pm1)
24%
XF08
87m
anno
syltr
ansf
eras
e (m
tfA)
31%
MAC
ROM
OLEC
ULE
MET
ABOL
ISM
/OTH
ER M
ACRO
MOL
ECUL
ESM
ETAB
OLIS
M/P
hosp
holip
ids
XF23
10CD
P-di
acyl
glyc
erol
-gly
cero
l-3-p
hosp
hate
3-
phos
phat
idyl
trans
fera
se (p
gsA)
49%
XF10
31gl
ycer
ol-3
-pho
spha
te a
cyltr
ansf
eras
e (p
lsB)
37%
CELL
STR
UCTU
RE/M
EMBR
ANE
COM
PON
ENTS
/Inne
rm
embr
ane
XF27
8060
kDa
inne
r-mem
bran
e pr
otei
n40
%XF
1640
anky
rin-li
ke p
rote
in (a
nk2)
26%
XF22
35bi
func
tiona
l pen
icill
in-b
indi
ngpr
otei
n 1C
pre
curs
or (p
bpC)
47%
XF00
82ch
aper
one
prot
ein
prec
urso
r (ec
pD)
40%
XF08
51D-
amin
o ac
id d
ehyd
roge
nase
sub
unit
(dad
A/da
dR)
54%
XF23
34di
acyl
glyc
erol
kin
ase
(dgk
A)54
%XF
0340
disu
lfide
bon
d fo
rmat
ion
prot
ein
B (d
sbB)
33%
XF24
33ep
imer
ase/
dehy
drat
ase
prot
ein
(cap
D)37
%XF
0256
gluc
ose-
1-ph
osph
ate
thym
idyl
yltra
nsfe
rase
(rfbA
)78
%XF
0375
inne
r mem
bran
e pr
otei
n (y
jdB)
36%
XF01
03m
embr
ane
prot
ein
25%
XF07
64m
embr
ane
prot
ein
36%
XF07
77m
embr
ane
prot
ein
(act
II-3)
22%
XF22
30pe
nici
llin-
bind
ing
prot
ein
6 pr
ecur
sor (
dacC
)47
%XF
0151
phos
phom
anno
mut
ase
(alg
C)45
%XF
1183
poly
sacc
harid
e bi
osyn
thet
ic p
rote
in(v
ipA/
tviB
)52
%XF
2333
prol
ipop
rote
in d
iacy
lgly
cery
l tra
nsfe
rase
(lgt/
umpA
)48
%XF
1310
rod
shap
e-de
term
inin
g pr
otei
n (m
reC)
37%
XF13
13ro
d sh
ape-
dete
rmin
ing
prot
ein
(mrd
B/ro
dA)
48%
XF19
79tra
nsm
embr
ane
prot
ein
(am
pE)
28%
XF11
40UD
P-N
-ace
tylg
luco
sam
ine
pyro
phos
phor
ylas
e (g
lmU)
49%
CELL
STR
UCTU
RE/M
EMBR
ANE
COM
PON
ENTS
/Out
erm
embr
ane
cons
titue
nts
XF23
92au
toly
tic ly
sozy
me
(lyc)
30%
XF08
47be
ta-h
exos
amin
idas
e pr
ecur
sor (
nahA
)37
%XF
2184
mem
bran
e-bo
und
lytic
trans
glyc
osyl
ase
(mltB
)36
%
XF10
46ou
ter m
embr
ane
antig
en (o
ma)
37%
XF03
84ou
ter m
embr
ane
hem
in re
cept
or (p
huR)
22%
XF03
43ou
ter m
embr
ane
prot
ein
(mop
B)31
%XF
0872
oute
r mem
bran
e pr
otei
n (o
mpW
)30
%XF
0873
oute
r mem
bran
e pr
otei
n43
%XF
1053
oute
r mem
bran
e pr
otei
n (o
mpP
1)28
%XF
1123
oute
r mem
bran
e pr
otei
n34
%XF
1739
oute
r mem
bran
e pr
otei
n (ro
mA)
43%
XF23
45ou
ter m
embr
ane
prot
ein
(sm
pA)
38%
XF10
24ou
ter m
embr
ane
prot
ein
H.8
prec
urso
r42
%XF
1811
oute
r mem
bran
e pr
otei
n Sl
p pr
ecur
sor (
slp)
31%
XF15
47pe
ptid
ogly
can-
asso
ciat
ed o
uter
mem
bran
elip
opro
tein
pre
curs
or (p
cp/lp
p)29
%XF
0033
PilE
pro
tein
(pilE
)37
%XF
0975
poly
phos
phat
e-se
lect
ive
porin
O (o
prO)
23%
XF03
21po
rin O
pre
curs
or (o
prO)
23%
XF00
28pr
e-pi
lin li
ke le
ader
seq
uenc
e (fi
mT)
37%
XF13
63so
lubl
e ly
tic m
urei
n tra
nsgl
ycos
ylas
epr
ecur
sor (
slt/
sltY
)25
%
CELL
STR
UCTU
RE/M
UREI
N S
ACCU
LUS,
PEP
TIDO
GLYC
AN
XF08
52al
anin
e ra
cem
ase
(alr)
45%
XF07
99D-
alan
ine-
-D-a
lani
ne li
gase
B (d
dlB/
ddl)
51%
XF08
55lip
opro
tein
(nlp
D/lp
pB)
39%
XF04
16lip
opro
tein
pre
curs
or (v
acJ)
36%
XF17
15m
onof
unct
iona
l bio
synt
hetic
pept
idog
lyca
n tra
nsgl
ycos
ylas
e (m
tgA)
50%
XF07
59N
-ace
tylm
uram
oyl-L
-ala
nine
am
idas
epr
ecur
sor (
amiC
)45
%XF
2646
oute
r mem
bran
e lip
opro
tein
pre
curs
or(lo
lB/h
emM
)29
%XF
1896
oute
r mem
bran
e pr
otei
n P6
pre
curs
or(p
al/o
mpP
6)37
%XF
1614
peni
cilli
n bi
ndin
g pr
otei
n (p
bp4)
39%
XF03
68pe
nici
llin-
bind
ing
prot
ein
1A (m
rcA/
ponA
)39
%XF
0884
peni
cilli
n-bi
ndin
g pr
otei
n 1B
(pon
B)41
%XF
1312
peni
cilli
n-bi
ndin
g pr
otei
n 2
(mrd
A/pb
p2)
38%
XF07
92pe
nici
llin-
bind
ing
prot
ein
3 (ft
sI/p
bpB)
43%
XF07
95ph
osph
o-N
-ace
tylm
uram
oyl-p
enta
pept
ide-
trans
fera
se (m
raY/
mur
X)59
%XF
2185
rare
lipo
prot
ein
A (rl
pA)
30%
XF13
09ro
d sh
ape-
dete
rmin
ing
prot
ein
(mre
B/en
vB/r
odY)
77%
XF13
11ro
d sh
ape-
dete
rmin
ing
prot
ein
(mre
D)32
%XF
0907
solu
ble
lytic
mur
ein
trans
glyc
osyl
ase
prec
urso
r (yj
bJ)
42%
XF07
97UD
P-N
-ace
tylg
luco
sam
ine-
-N-a
cety
lmur
amyl
-(p
enta
pept
ide)
pyr
opho
spho
ryl-u
ndec
apre
nol
(mur
G)47
%XF
0798
UDP-
N-a
cety
lmur
amat
e--a
lani
ne li
gase
(mur
C)53
%XF
0276
UDP-
N-a
cety
lmur
amat
e-L-
alan
ine
ligas
e(m
pl)
50%
XF11
18UD
P-N
-ace
tylm
uram
oyla
lani
ne--D
-glu
tam
ate
ligas
e (m
urD)
29%
XF07
93UD
P-N
-ace
tylm
uram
oyla
lany
l-D-g
luta
mat
e--
2,6-
diam
inop
imel
ate
ligas
e (m
urE)
43%
XF07
94UD
P-N
-ace
tylm
uram
oyla
lany
l-D-g
luta
myl
-2,6
diam
inop
imel
ate-
-D-a
lany
l-D-a
lany
llig
ase
(mur
F/m
ra)
41%
XF25
72UD
P-N
-ace
tylp
yruv
oylg
luco
sam
ine
redu
ctas
e (m
urB)
43%
XF10
50un
deca
pren
yl p
yrop
hosp
hate
syn
thet
ase
(upp
S/rth
)51
%
CELL
STR
UCTU
RE/S
URFA
CE P
OLYS
ACCH
ARID
ES,
LIPO
POLY
SACC
HARI
DES,
AN
D AN
TIGE
NS
XF12
892-
dehy
dro-
3-de
oxyp
hosp
hooc
tona
teal
dola
se (k
dsA)
44%
XF01
053-
deox
y-D-
man
no-o
ctul
oson
ic a
cid
trans
fera
se (k
dtA/
waa
A)44
%XF
2299
3-de
oxy-
man
no-o
ctul
oson
ate
cytid
ylyl
trans
fera
se (k
dsB)
50%
XF14
19ac
etyl
trans
fera
se (l
pxD/
firA/
omsA
)26
%XF
0918
acyl
-[ACP
]-UDP
-N-a
cety
lglu
cosa
min
e (lp
xA)
32%
XF19
94be
ta 1
,4 g
luco
syltr
ansf
eras
e29
%XF
0612
dolic
hol-p
hosp
hate
man
nosy
ltran
sfer
ase
(dm
t)24
%XF
1638
dolic
hyl-p
hosp
hate
man
nose
syn
thas
ere
late
d pr
otei
n34
%XF
0257
dTDP
-4-d
ehyd
rorh
amno
se 3
,5-e
pim
eras
e(rf
bD)
75%
XF02
58dT
DP-4
-ket
o-L-
rham
nose
redu
ctas
e (rf
bC)
54%
XF02
55dT
DP-g
luco
se 4
,6-d
ehyd
rata
se (r
fbB)
76%
XF06
11dT
DP-g
luco
se 4
-6-d
ehyd
rata
se (r
fbB)
63%
XF16
37gl
ycos
yl tr
ansf
eras
e (s
psQ)
29%
XF10
82lip
id A
4’-k
inas
e (lp
xK)
38%
XF01
04lip
id A
bio
synt
hesi
s la
uroy
l acy
ltran
sfer
ase
(htrB
/waa
M)
40%
XF13
48lip
id A
bio
synt
hesi
s la
uroy
l acy
ltran
sfer
ase
(htrB
/waa
M)
26%
XF10
42lip
id A
dis
acch
arid
e sy
ntha
se (l
pxB/
pgsB
)45
%XF
0879
lipop
olys
acch
arid
e bi
osyn
thes
is p
rote
in (r
fbU)
26%
XF24
34lip
opol
ysac
char
ide
core
bio
synt
hesi
s pr
otei
n37
%XF
0980
lipop
olys
acch
arid
e sy
nthe
sis
enzy
me
(kdt
B)56
%XF
0778
O-an
tigen
ace
tyla
se (o
afA)
26%
XF14
13po
lysi
alic
aci
d ca
psul
e ex
pres
sion
pro
tein
(kps
F)45
%XF
2154
sacc
harid
e bi
osyn
thes
is re
gula
tory
pro
tein
(ops
X)73
%XF
0176
suga
r tra
nsfe
rase
43%
XF10
45UD
P-3-
O-(R
-3-h
ydro
xym
yris
toyl
)-glu
cosa
min
eN
-acy
ltran
sfer
ase
(lpxD
/firA
/ssc
)41
%XF
1646
UDP-
3-O-
(R-3
-hyd
roxy
myr
isto
yl)-g
luco
sam
ine
N-a
cyltr
ansf
eras
e (lp
xD/f
irA)
28%
XF04
86UD
P-3-
O-[3
-hyd
roxy
myr
isto
yl] g
luco
sam
ine
N-
acyl
trans
fera
se (l
pxD)
31%
XF08
03UD
P-3-
O-[3
-hyd
roxy
myr
isto
yl] N
-ac
etyl
gluc
osam
ine
deac
etyl
ase
(lpxC
)59
%XF
1415
UDP-
N-a
cety
lglu
cosa
min
e 1-
carb
oxyv
inyl
trans
fera
se (m
urA/
mur
Z)62
%XF
1043
UDP-
N-a
cety
lglu
cosa
min
e ac
yltra
nsfe
rase
(lpxA
)44
%
CELL
STR
UCTU
RE/S
URFA
CE S
TRUC
TURE
S
XF00
77fim
bria
l adh
esin
pre
curs
or (m
rkD)
30%
XF00
78fim
bria
l adh
esin
pre
curs
or (m
rkD)
32%
XF00
80fim
bria
l adh
esin
pre
curs
or (f
imA/
pilA
)29
%XF
0369
fimbr
ial a
ssem
bly
mem
bran
e pr
otei
n (p
ilM)
61%
XF03
70fim
bria
l ass
embl
y m
embr
ane
prot
ein
(pilN
)44
%XF
0371
fimbr
ial a
ssem
bly
mem
bran
e pr
otei
n (p
ilO)
43%
XF03
72fim
bria
l ass
embl
y pr
otei
n (p
ilP)
38%
XF03
73fim
bria
l ass
embl
y pr
otei
n (p
ilQ)
34%
XF04
78fim
bria
l ass
embl
y pr
otei
n (p
ilY1)
33%
XF25
38fim
bria
l ass
embl
y pr
otei
n (p
ilC)
56%
XF25
39fim
bria
l pro
tein
48%
XF25
42fim
bria
l pro
tein
53%
XF00
83fim
bria
l sub
unit
prec
urso
r42
%XF
0487
fimbr
illin
37%
XF05
38fim
brill
in37
%XF
0539
fimbr
illin
(fim
A)38
%XF
1791
fimbr
illin
48%
XF00
81ou
ter m
embr
ane
ushe
r pro
tein
pre
curs
or(fi
mD)
36%
XF19
53pi
lus
biog
enes
is p
rote
in (p
ilJ)
46%
XF19
54pi
lus
biog
enes
is p
rote
in (p
ilI)
33%
XF25
44pi
lus
biog
enes
is p
rote
in (p
ilB)
55%
XF19
55pi
lus
prot
ein
(pilG
)82
%XF
0031
PilX
pro
tein
(pilX
)28
%XF
0032
PilY
1 ge
ne p
rodu
ct (p
ilY1)
32%
XF12
24Pi
lY1
gene
pro
duct
(pilY
1)30
%XF
2537
pre-
pilin
lead
er p
eptid
ase
(xps
O)76
%XF
0029
pre-
pilin
lead
er s
eque
nce
(pilV
)33
%XF
1632
twitc
hing
mot
ility
pro
tein
(pilU
)60
%XF
1633
twitc
hing
mot
ility
pro
tein
(pilT
)74
%XF
0677
type
4 fi
mbr
iae
asse
mbl
y pr
otei
n (p
ilZ)
62%
XF04
79ty
pe IV
pili
n (p
ilE)
35%
CELL
ULAR
PRO
CESS
ES/T
RAN
SPOR
T/Am
ino
acid
s, a
min
es
XF04
08am
ino
acid
tran
spor
ter (
yhdG
)43
%XF
2730
amin
o ac
id tr
ansp
orte
r (rh
tC)
28%
XF22
07ca
tioni
c am
ino
acid
tran
spor
ter
43%
XF22
08ca
tioni
c am
ino
acid
tran
spor
ter
41%
XF18
91di
-trip
eptid
e AB
C tra
nspo
rter m
embr
ane
prot
ein
(ycl
F)35
%XF
0656
glut
amat
e sy
mpo
rt pr
otei
n (g
ltT)
28%
XF19
37pr
oton
glu
tam
ate
sym
port
prot
ein
(gltP
)35
%
CELL
ULAR
PRO
CESS
ES/T
RAN
SPOR
T/An
ions
XF04
11AB
C tra
nspo
rter n
itrat
e pe
rmea
se (n
rtB)
26%
XF21
41AB
C tra
nspo
rter p
hosp
hate
bin
ding
pro
tein
(pho
X)67
%XF
2142
ABC
trans
porte
r pho
spha
te p
erm
ease
(pst
C/ph
oW)
50%
XF21
43AB
C tra
nspo
rter p
hosp
hate
per
mea
se(p
stA/
phoT
)53
%XF
1344
ABC
trans
porte
r sul
fate
bin
ding
pro
tein
(sbp
)64
%XF
1345
ABC
trans
porte
r sul
fate
per
mea
se(c
ysU/
cysT
)50
%XF
1346
ABC
trans
porte
r sul
fate
per
mea
se (c
ysW
)49
%XF
0412
nitra
te A
BC tr
ansp
orte
r ATP
-bin
ding
prot
ein
(nrtD
)46
%XF
2144
phos
phat
e AB
C tra
nspo
rter A
TP-b
indi
ngpr
otei
n (p
stB/
phoT
)65
%XF
1347
sulfa
te A
BC tr
ansp
orte
r ATP
-bin
ding
prot
ein
(cys
A)61
%
CELL
ULAR
PRO
CESS
ES/T
RAN
SPOR
T/Ca
rboh
ydra
tes,
org
anic
acid
s, a
lcoh
ols
XF24
46AB
C tra
nspo
rter s
ugar
per
mea
se40
%XF
2447
ABC
trans
porte
r sug
ar p
erm
ease
(lac
F)37
%XF
2448
ABC
trans
porte
r sug
ar-b
indi
ng p
rote
in (m
alE)
24%
XF00
87al
pha-
keto
glut
arat
e pe
rmea
se s
ympo
rter
(kgt
P/w
itA)
64%
XF09
76C4
-dic
arbo
xyla
te tr
ansp
ort p
rote
in (d
ctA)
66%
XF14
62gl
ucos
e/ga
lact
ose
trans
porte
r (gl
uP)
31%
XF16
09gl
ucos
e/ga
lact
ose
trans
porte
r (gl
uP)
48%
XF22
67gl
ycer
ol u
ptak
e fa
cilit
ator
pro
tein
(glp
F)39
%XF
1406
HPr k
inas
e/ph
osph
atas
e (p
tsK)
36%
XF03
20M
g++/
citra
te c
ompl
ex tr
ansp
orte
r(c
itN/c
itH)
60%
XF14
02ph
osph
otra
nsfe
rase
sys
tem
enz
yme
I (ph
bI)
40%
XF14
03ph
osph
otra
nsfe
rase
sys
tem
HPr
enz
yme
(phb
H)52
%XF
1067
suga
r ABC
tran
spor
ter A
TP-b
indi
ng p
rote
in53
%
CELL
ULAR
PRO
CESS
ES/T
RAN
SPOR
T/Ca
tions
XF23
28AB
C tra
nspo
rter s
odiu
m p
erm
ease
(nat
B)27
%XF
1844
amm
oniu
m tr
ansp
orte
r (am
tB)
57%
XF03
95ba
cter
iofe
rritin
(bfr)
61%
XF21
40ca
tion:
prot
on a
ntip
orte
r (yb
aL)
57%
XF21
34fe
rric
ente
roba
ctin
rece
ptor
(bfe
A)22
%XF
2137
ferri
c en
tero
bact
in re
cept
or (b
feA)
22%
XF09
32fe
rrous
iron
tran
spor
t pro
tein
47%
XF09
33fe
rrous
iron
tran
spor
t pro
tein
B (f
eoB)
52%
XF14
26io
n tra
nspo
rter
48%
XF09
00m
agne
sium
and
cob
alt t
rans
port
prot
ein
(cor
A)33
%XF
1015
man
gane
se tr
ansp
ort p
rote
in (m
ntH2
)64
%XF
1401
Mg+
+ tra
nspo
rter (
mgt
E)26
%XF
1398
Na+
/H+
exch
ange
pro
tein
24%
XF20
19N
a+:H
+ an
tipor
ter (
yjcE
)42
%XF
0324
perip
lasm
ic ir
on-b
indi
ng p
rote
in (a
fuA)
24%
XF09
01po
lar a
min
o ac
id tr
ansp
orte
r (yb
eX)
46%
XF19
03po
tass
ium
upt
ake
prot
ein
(kup
/trk
D)48
%XF
2329
sodi
um A
BC tr
ansp
orte
r ATP
-bin
ding
prot
ein
(nat
A)47
%XF
0599
TonB
-dep
ende
nt re
cept
or fo
r iro
ntra
nspo
rt (y
biL)
47%
XF14
96To
nB-d
epen
dent
rece
ptor
for i
ron
trans
port
(ync
D)23
%XF
0367
volta
ge-g
ated
pot
assi
um c
hann
el b
eta
subu
nit
50%
CELL
ULAR
PRO
CESS
ES/T
RAN
SPOR
T/Pr
otei
n, p
eptid
e se
cret
ion
XF24
55he
me
ABC
trans
porte
r ATP
-bin
ding
prot
ein
(ccm
A)41
%XF
2456
hem
e AB
C tra
nspo
rter m
embr
ane
prot
ein
(ccm
B)50
%XF
2457
hem
e AB
C tra
nspo
rter m
embr
ane
prot
ein
(ccm
C)48
%XF
2261
olig
opep
tide
trans
porte
r40
%XF
0806
prep
rote
in tr
ansl
ocas
e Se
cA s
ubun
it(s
ecA/
prlD
/azi/
pea)
60%
XF11
72pr
epro
tein
tran
sloc
ase
SecY
sub
unit
(sec
Y/pr
lA)
55%
XF26
39pr
epro
tein
tran
sloc
ase
subu
nit (
secE
)33
%XF
0224
prep
rote
in tr
ansl
ocas
e Ya
jC s
ubun
it (y
ajC)
36%
XF26
85pr
otea
se IV
(spp
A)40
%XF
0225
prot
ein-
expo
rt m
embr
ane
prot
ein
(sec
D)44
%XF
0226
prot
ein-
expo
rt m
embr
ane
prot
ein
(sec
F)38
%XF
0304
prot
ein-
expo
rt m
embr
ane
prot
ein
(sec
G)43
%XF
1801
prot
ein-
expo
rt pr
otei
n (s
ecB)
43%
XF05
62se
c-in
depe
nden
t pro
tein
tran
sloc
ase
(tatC
/mttB
)47
%XF
0073
sign
al re
cogn
ition
par
ticle
pro
tein
(ffh
)63
%
XF19
13ty
pe V
sec
reto
ry p
athw
ay p
rote
in (m
ttC)
46%
CELL
ULAR
PRO
CESS
ES/T
RAN
SPOR
T/Ot
her
XF08
75AB
C tra
nspo
rter A
TP-b
indi
ng p
rote
in (y
usC)
48%
XF09
44AB
C tra
nspo
rter A
TP-b
indi
ng p
rote
in (y
jjK)
69%
XF10
77AB
C tra
nspo
rter A
TP-b
indi
ng p
rote
in (y
cfV)
55%
XF10
81AB
C tra
nspo
rter A
TP-b
indi
ng p
rote
in (m
sbA)
41%
XF12
23AB
C tra
nspo
rter A
TP-b
indi
ng p
rote
in (y
adG)
54%
XF13
02AB
C tra
nspo
rter A
TP-b
indi
ng p
rote
in42
%XF
1409
ABC
trans
porte
r ATP
-bin
ding
pro
tein
60%
XF14
75AB
C tra
nspo
rter A
TP-b
indi
ng p
rote
in (y
nhD)
63%
XF16
02AB
C tra
nspo
rter A
TP-b
indi
ng p
rote
in31
%XF
2133
ABC
trans
porte
r ATP
-bin
ding
pro
tein
(yhe
S)45
%XF
2568
ABC
trans
porte
r ATP
-bin
ding
pro
tein
(rfb
E)45
%XF
2582
ABC
trans
porte
r ATP
-bin
ding
pro
tein
42%
XF26
17AB
C tra
nspo
rter A
TP-b
indi
ng p
rote
in (u
up)
49%
XF26
95AB
C tra
nspo
rter A
TP-b
indi
ng p
rote
in (f
tsE)
47%
XF14
74AB
C tra
nspo
rter m
embr
ane
prot
ein
(ynh
C)33
%XF
1476
ABC
trans
porte
r mem
bran
e pr
otei
n69
%XF
0874
ABC
trans
porte
r per
mea
se p
rote
in48
%XF
2567
ABC
trans
porte
r per
mea
se p
rote
in (r
fbD)
35%
XF16
04AB
C tra
nspo
rter v
itam
in B
12 u
ptak
epe
rmea
se (b
tuE)
44%
XF04
06ex
port
prot
ein
(ygj
T)54
%XF
0039
larg
e-co
nduc
tanc
e m
echa
nose
nsiti
vech
anne
l (m
scL)
52%
XF05
89pe
rmea
se28
%XF
2301
poly
sacc
harid
e ex
port
prot
ein
(mrp
)63
%XF
1258
smal
l con
duct
ance
mec
hano
sens
itive
ion
chan
nel (
yggB
)35
%XF
2251
solu
te:N
a+ s
ympo
rter (
ppa)
58%
XF02
81tra
nspo
rt pr
otei
n (y
ueF)
24%
XF06
51tra
nspo
rt pr
otei
n (y
xaH)
33%
XF17
28tra
nspo
rt pr
otei
n27
%
CELL
ULAR
PRO
CESS
ES/C
ELL
DIVI
SION
XF06
59ce
ll cy
cle
prot
ein
(mes
J)42
%XF
0286
cell
divi
sion
inhi
bito
r (su
lA)
39%
XF13
22ce
ll di
visi
on in
hibi
tor (
min
C)35
%XF
0093
cell
divi
sion
pro
tein
(hflB
/fts
H/m
rsC/
tolZ
)64
%XF
0094
cell
divi
sion
pro
tein
(fts
J/m
rsF)
52%
XF07
91ce
ll di
visi
on p
rote
in (f
tsL)
30%
XF07
96ce
ll di
visi
on p
rote
in (f
tsW
)44
%XF
0800
cell
divi
sion
pro
tein
(fts
Q)32
%XF
0801
cell
divi
sion
pro
tein
(fts
A/di
vA)
45%
XF08
02ce
ll di
visi
on p
rote
in (f
tsZ)
59%
XF14
50ce
ll di
visi
on p
rote
in (f
tsK)
50%
XF19
10ce
ll di
visi
on p
rote
in (f
tsY)
45%
XF25
57ce
ll di
visi
on p
rote
in (z
ipA)
27%
XF26
94ce
ll di
visi
on p
rote
in (f
tsX/
ftsS)
31%
XF13
20ce
ll di
visi
on to
polo
gica
l spe
cific
ity fa
ctor
(min
E)48
%XF
1657
cell
filam
enta
tion
prot
ein
(fic)
38%
XF22
81ch
rom
osom
e pa
rtitio
ning
pro
tein
43%
XF22
82ch
rom
osom
e pa
rtitio
ning
pro
tein
(par
A)49
%XF
1785
chro
mos
ome
parti
tioni
ng re
late
dpr
otei
n (s
oj)
27%
XF22
47GT
P bi
ndin
g pr
otei
n (e
ra)
51%
XF10
84pa
rtitio
n pr
otei
n (p
arA)
32%
XF13
21se
ptum
site
-det
erm
inin
g pr
otei
n (m
inD)
68%
CELL
ULAR
PRO
CESS
ES/C
HEM
OTAX
IS A
ND
MOB
ILIT
Y
XF19
52ch
emot
axis
-rela
ted
prot
ein
kina
se (c
hpA)
42%
MOB
ILE
GEN
ETIC
ELE
MEN
TS/P
HAGE
-REL
ATED
FUN
CTIO
NS
AND
PROP
HAGE
S
XF00
89ho
st fa
ctor
-I pr
otei
n (h
fq)
92%
XF07
19ph
age-
rela
ted
base
plat
e as
sem
bly
prot
ein
(gpV
)32
%XF
0723
phag
e-re
late
d ba
sepl
ate
asse
mbl
y pr
otei
n(g
pW)
40%
XF07
24ph
age-
rela
ted
base
plat
e as
sem
bly
prot
ein
(gpJ
)46
%XF
2488
phag
e-re
late
d ba
sepl
ate
asse
mbl
y pr
otei
n(g
pJ)
46%
XF24
89ph
age-
rela
ted
base
plat
e as
sem
bly
prot
ein
(gpW
)40
%XF
2492
phag
e-re
late
d ba
sepl
ate
asse
mbl
y pr
otei
n(g
pV)
32%
XF07
27ph
age-
rela
ted
cont
ract
ile ta
il sh
eath
prot
ein
(fIR2
)46
%XF
2485
phag
e-re
late
d co
ntra
ctile
tail
shea
thpr
otei
n (fI
R2)
46%
XF07
28ph
age-
rela
ted
cont
ract
ile ta
il tu
be p
rote
in(fI
IR2)
41%
XF06
83ph
age-
rela
ted
DNA
poly
mer
ase
(dpo
L)50
%XF
2525
phag
e-re
late
d DN
A po
lym
eras
e (d
poL)
49%
XF05
13ph
age-
rela
ted
endo
lysi
n (ly
cV)
46%
XF06
31ph
age-
rela
ted
inte
gras
e (in
t)28
%XF
0678
phag
e-re
late
d in
tegr
ase
(int)
27%
XF16
42ph
age-
rela
ted
inte
gras
e (in
t)28
%XF
1718
phag
e-re
late
d in
tegr
ase
(int)
91%
XF22
88ph
age-
rela
ted
inte
gras
e (in
t)26
%XF
2530
phag
e-re
late
d in
tegr
ase
(int)
28%
XF15
64ph
age-
rela
ted
lyso
zym
e (ly
cV)
43%
XF16
69ph
age-
rela
ted
lyso
zym
e (ly
cV)
43%
XF23
14ph
age-
rela
ted
lyso
zym
e (ly
cV)
42%
XF07
13ph
age-
rela
ted
porta
l pro
tein
(gp4
)25
%XF
2498
phag
e-re
late
d po
rtal p
rote
in (g
p4)
25%
XF04
85ph
age-
rela
ted
prot
ein
34%
XF06
80ph
age-
rela
ted
prot
ein
56%
XF06
82ph
age-
rela
ted
prot
ein
50%
XF06
84ph
age-
rela
ted
prot
ein
49%
XF06
85ph
age-
rela
ted
prot
ein
53%
XF06
86ph
age-
rela
ted
prot
ein
41%
XF07
04ph
age-
rela
ted
prot
ein
32%
XF07
05ph
age-
rela
ted
prot
ein
32%
XF07
07ph
age-
rela
ted
prot
ein
44%
XF07
10ph
age-
rela
ted
prot
ein
27%
XF15
55ph
age-
rela
ted
prot
ein
34%
XF15
70ph
age-
rela
ted
prot
ein
29%
XF15
73ph
age-
rela
ted
prot
ein
27%
XF16
45ph
age-
rela
ted
prot
ein
31%
XF16
63ph
age-
rela
ted
prot
ein
47%
XF16
75ph
age-
rela
ted
prot
ein
29%
XF16
78ph
age-
rela
ted
prot
ein
27%
XF17
63ph
age-
rela
ted
prot
ein
30%
XF17
86ph
age-
rela
ted
prot
ein
42%
XF18
64ph
age-
rela
ted
prot
ein
48%
XF18
69ph
age-
rela
ted
prot
ein
37%
XF18
70ph
age-
rela
ted
prot
ein
35%
XF18
75ph
age-
rela
ted
prot
ein
36%
XF18
76ph
age-
rela
ted
prot
ein
36%
XF21
27ph
age-
rela
ted
prot
ein
24%
XF21
32ph
age-
rela
ted
prot
ein
34%
XF22
90ph
age-
rela
ted
prot
ein
57%
XF22
91ph
age-
rela
ted
prot
ein
47%
XF22
92ph
age-
rela
ted
prot
ein
50%
XF22
94ph
age-
rela
ted
prot
ein
32%
XF22
95ph
age-
rela
ted
prot
ein
53%
XF24
79ph
age-
rela
ted
prot
ein
32%
XF24
82ph
age-
rela
ted
prot
ein
21%
XF24
84ph
age-
rela
ted
prot
ein
41%
XF25
01ph
age-
rela
ted
prot
ein
27%
XF25
04ph
age-
rela
ted
prot
ein
44%
XF25
05ph
age-
rela
ted
prot
ein
34%
XF25
11ph
age-
rela
ted
prot
ein
28%
XF25
22ph
age-
rela
ted
prot
ein
41%
XF25
23ph
age-
rela
ted
prot
ein
52%
XF25
24ph
age-
rela
ted
prot
ein
43%
XF25
26ph
age-
rela
ted
prot
ein
50%
XF25
27ph
age-
rela
ted
prot
ein
21%
XF25
28ph
age-
rela
ted
prot
ein
56%
XF05
00ph
age-
rela
ted
repr
esso
r pro
tein
(rac
R)50
%XF
0696
phag
e-re
late
d re
pres
sor p
rote
in (r
pcI)
29%
XF16
58ph
age-
rela
ted
repr
esso
r pro
tein
(CI)
24%
XF17
06ph
age-
rela
ted
tail
fiber
pro
tein
(gp3
7)25
%XF
0725
phag
e-re
late
d ta
il pr
otei
n (g
pI)
35%
XF07
30ph
age-
rela
ted
tail
prot
ein
(gpT
)21
%XF
0731
phag
e-re
late
d ta
il pr
otei
n (g
pU)
40%
XF07
32ph
age-
rela
ted
tail
prot
ein
(gpX
)41
%XF
0733
phag
e-re
late
d ta
il pr
otei
n (g
pD)
32%
XF24
80ph
age-
rela
ted
tail
prot
ein
(gpX
)41
%XF
2481
phag
e-re
late
d ta
il pr
otei
n (g
pU)
40%
XF24
87ph
age-
rela
ted
tail
prot
ein
(gpI
)35
%XF
0508
phag
e-re
late
d te
rmin
ase
larg
e su
buni
t (gp
2)25
%XF
0711
phag
e-re
late
d te
rmin
ase
larg
e su
buni
t (gp
2)28
%XF
2500
phag
e-re
late
d te
rmin
ase
larg
e su
buni
t (gp
2)28
%
MOB
ILE
GEN
ETIC
ELE
MEN
TS/P
LASM
ID-R
ELAT
ED F
UNCT
ION
S
XF20
45co
njug
al tr
ansf
er p
rote
in (t
rbN
)76
%XF
2046
conj
ugal
tran
sfer
pro
tein
(trb
L)45
%XF
2048
conj
ugal
tran
sfer
pro
tein
(trb
J)---
%XF
2049
conj
ugal
tran
sfer
pro
tein
(trb
I)53
%XF
2050
conj
ugal
tran
sfer
pro
tein
(trb
H)45
%XF
2051
conj
ugal
tran
sfer
pro
tein
(trb
G)65
%XF
2052
conj
ugal
tran
sfer
pro
tein
(trb
F)49
%XF
2053
conj
ugal
tran
sfer
pro
tein
(trb
E)---
%XF
2054
conj
ugal
tran
sfer
pro
tein
(trb
D)68
%XF
2055
conj
ugal
tran
sfer
pro
tein
(trb
C)52
%XF
2056
conj
ugal
tran
sfer
pro
tein
(trb
B)69
%XF
2058
conj
ugal
tran
sfer
pro
tein
(tra
F)59
%XF
2059
conj
ugal
tran
sfer
pro
tein
(tra
E)73
%XF
2060
conj
ugal
tran
sfer
pro
tein
(tra
D)43
%XF
1759
cons
erve
d pl
asm
id p
rote
in35
%XF
2066
cons
erve
d pl
asm
id p
rote
in (y
acB)
35%
XF21
61co
nser
ved
plas
mid
pro
tein
30%
XF20
61DN
A pr
imas
e (tr
aC)
44%
XF24
44ph
erom
one
shut
dow
n pr
otei
n (tr
aB)
31%
XF15
89pl
asm
id s
tabi
lizat
ion
prot
ein
50%
XF15
90pl
asm
id s
tabi
lizat
ion
prot
ein
62%
XF20
31pl
asm
id s
tabi
lizat
ion
prot
ein
(par
D)81
%XF
2032
plas
mid
sta
biliz
atio
n pr
otei
n (p
arE)
75%
XF15
74pl
asm
id-re
late
d pr
otei
n (tr
aN)
34%
XF16
79pl
asm
id-re
late
d pr
otei
n (tr
aN)
34%
XF19
45Vi
rK p
rote
in (v
irK)
35%
MOB
ILE
GEN
ETIC
ELE
MEN
TS/T
RAN
SPOS
ON- A
ND
INTR
ON-
RELA
TED
FUN
CTIO
NS
XF20
63DN
A-in
verta
se (r
in)
---%
XF19
30re
solv
ase
(tnpR
)---
%XF
1775
reve
rse
trans
crip
tase
---%
XF19
31tra
nspo
sase
(tnp
A)---
%XF
2303
trans
posa
se (t
npA)
71%
XF03
25tra
nspo
sase
OrfA
64%
XF05
35tra
nspo
sase
OrfA
64%
XF03
26tra
nspo
sase
OrfB
---%
XF05
36tra
nspo
sase
OrfB
35%
PATH
OGEN
ICIT
Y, V
IRUL
ENCE
, AN
D AD
APTA
TION
/TOX
INPR
ODUC
TION
AN
D DE
TOXI
FICA
TION
XF17
262,
5-di
chlo
ro-2
,5-c
yclo
hexa
dien
e-1,
4-di
olde
hydr
ogen
ase
(linC
)39
%XF
0243
acrif
lavi
n re
sist
ance
pro
tein
(yer
P)23
%XF
2386
acrif
lavi
n re
sist
ance
pro
tein
(yeg
N)
42%
XF23
85ac
rifla
vin
resi
stan
ce p
rote
in D
(yeg
N)
50%
XF24
07ba
cter
ioci
n27
%XF
0165
beta
-lact
amas
e in
duct
ion
sign
al tr
ansd
ucer
prot
ein
(am
pG)
38%
XF16
21be
ta-la
ctam
ase-
like
prot
ein
(pbp
)24
%XF
0010
biop
olym
er tr
ansp
ort E
xbB
prot
ein
(exb
B)81
%XF
0011
biop
olym
er tr
ansp
ort E
xbD1
pro
tein
(exb
D1)
78%
XF00
12bi
opol
ymer
tran
spor
t Exb
D2 p
rote
in (e
xbD2
)82
%XF
2232
cata
lase
/per
oxid
ase
(cpe
B)66
%XF
2083
catio
n ef
flux
syst
em p
rote
in (c
zcA)
24%
XF02
62co
licin
V p
recu
rsor
(cva
C)36
%XF
0263
colic
in V
pre
curs
or (c
vaC)
31%
XF19
48co
licin
V p
rodu
ctio
n pr
otei
n (c
vpA)
31%
XF12
20co
licin
V s
ecre
tion
ABC
trans
porte
rAT
P-bi
ndin
g pr
otei
n (c
vaB)
40%
XF12
16co
licin
V s
ecre
tion
prot
ein
(cva
A)25
%XF
2084
com
pone
nt o
f mul
tidru
g ef
flux
syst
em (m
exE)
28%
XF13
41co
pper
hom
eost
asis
pro
tein
(cut
C)47
%XF
0132
copp
er re
sist
ance
pro
tein
A p
recu
rsor
(cop
A)61
%XF
0133
copp
er re
sist
ance
pro
tein
B p
recu
rsor
(cop
B)38
%XF
1741
daun
orub
icin
C-1
3 ke
tore
duct
ase
(dnr
U)31
%XF
2148
dim
ethy
lade
nosi
ne tr
ansf
eras
e (k
sgA/
rsm
A)48
%XF
2416
drug
tole
ranc
e pr
otei
n (ly
tB)
64%
XF09
93dr
ug:p
roto
n an
tipor
ter (
tetV
)24
%XF
1765
drug
:pro
ton
antip
orte
r (m
mr)
33%
XF10
29gl
utar
yl-7
-ACA
acy
lase
pre
curs
or (g
aa)
60%
XF18
90gl
utat
hion
e pe
roxi
dase
-like
pro
tein
(gpo
)50
%XF
1210
glut
athi
one
S-tra
nsfe
rase
(gst
)38
%XF
0175
hem
olys
in II
I pro
tein
48%
XF23
98he
mol
ysin
sec
retio
n pr
otei
n D
(hly
D)39
%XF
0668
hem
olys
in-ty
pe c
alci
um b
indi
ng p
rote
in (f
rpC)
32%
XF10
11he
mol
ysin
-type
cal
cium
bin
ding
pro
tein
(frp
C)30
%XF
2759
hem
olys
in-ty
pe c
alci
um b
indi
ng p
rote
in (f
rpC)
35%
XF19
34He
tI pr
otei
n (h
etI)
35%
XF02
44m
embr
ane
fusi
on p
rote
in (m
exC)
25%
XF23
84m
embr
ane
fusi
on p
rote
in p
recu
rsor
(mtrC
)37
%XF
2686
mul
tidru
g ef
flux
prot
ein
(ydh
E)34
%XF
2094
mul
tidru
g-ef
flux
trans
porte
r (ac
rF/e
nvD)
50%
XF17
32N
AD(P
)H-d
epen
dent
2-c
yclo
hexe
n-1-
one
redu
ctas
e (n
cr)
40%
XF11
37N
onF-
rela
ted
prot
ein
(non
F)34
%
XF18
27or
gani
c hy
drop
erox
ide
resi
stan
cepr
otei
n (o
hr)
70%
XF25
86ou
ter m
embr
ane
expo
rt fa
ctor
(tolC
/mtc
B/m
ukA/
refI)
33%
XF25
50ou
ter m
embr
ane
hem
olys
in a
ctiv
ator
prot
ein
(hec
B)26
%XF
1532
oxid
ativ
e st
ress
tran
scrip
tiona
l reg
ulat
or(o
xyR)
81%
XF10
38pe
ptid
e sy
ntha
se23
%XF
2276
pept
ide
synt
hase
26%
XF06
19pe
ripla
smic
div
alen
t cat
ion
tole
ranc
epr
otei
n (c
utA/
cycY
/cut
A1)
37%
XF17
29ph
enyl
acet
alde
hyde
deh
ydro
gena
se61
%XF
1131
PmbA
pro
tein
(pm
bA/t
ldE)
45%
XF21
35po
lyke
tide
synt
hase
(PKS
) (frn
E)35
%XF
2093
prec
urso
r of d
rug
resi
stan
ce p
rote
in(a
crA/
mtc
A/lir
)42
%XF
0769
pter
idin
e-de
pend
ent d
eoxy
gena
se (r
apK)
34%
XF15
30su
buni
t C o
f alk
yl h
ydro
pero
xide
redu
ctas
e(a
hpC)
70%
XF15
31su
buni
t F o
f alk
yl h
ydro
pero
xide
redu
ctas
e(a
hpF)
77%
XF26
14su
pero
xide
dis
mut
ase
[MN
] (so
dA/s
od)
78%
XF19
21su
pero
xide
dis
mut
ase
[MN
] pre
curs
or(s
odA/
sodM
)57
%XF
2778
thio
phen
e an
d fu
ran
oxid
atio
n pr
otei
n(th
dF/t
rmE)
47%
XF18
98To
lA p
rote
in (t
olA)
21%
XF18
97To
lB p
rote
in p
recu
rsor
(tol
B)40
%XF
1900
TolQ
pro
tein
(tol
Q/fii
)48
%XF
1899
TolR
pro
tein
(tol
R)34
%XF
0009
TonB
pro
tein
(ton
B)72
%XF
1957
TonB
pro
tein
(ton
B)26
%XF
2287
TonB
pro
tein
(ton
B)29
%XF
2327
TonB
pro
tein
(ton
B)28
%XF
2605
TonB
pro
tein
(ton
B)36
%XF
2397
toxi
n se
cret
ion
ABC
trans
porte
r ATP
-bin
ding
prot
ein
(hly
B)61
%XF
1841
unde
capr
enol
kin
ase
(bac
A)38
%
PATH
OGEN
ICIT
Y, V
IRUL
ENCE
, AN
D AD
APTA
TION
/HOS
T CE
LLW
ALL
DEGR
ADAT
ION
XF12
671,
4-be
ta-c
ello
bios
idas
e (c
bhA)
46%
XF08
18en
do-1
,4-b
eta-
gluc
anas
e (e
ngXC
A)64
%XF
2708
endo
-1,4
-bet
a-gl
ucan
ase
(egl
)47
%XF
0810
extra
cellu
lar e
ndog
luca
nase
pre
curs
or(e
ngXC
A)33
%XF
0845
fam
ily 3
gly
cosi
de h
ydro
lase
(xyl
A)45
%XF
2466
poly
gala
ctur
onas
e pr
ecur
sor (
pglA
)---
%
PATH
OGEN
ICIT
Y, V
IRUL
ENCE
, AN
DAD
APTA
TION
/EXO
POLY
SACC
HARI
DES
XF23
70Gu
mB
prot
ein
(gum
B)67
%XF
2369
Gum
C pr
otei
n (g
umC)
62%
XF23
67Gu
mD
prot
ein
(gum
D)73
%XF
2366
Gum
E pr
otei
n (g
umE)
61%
XF23
65Gu
mF
prot
ein
(gum
F)44
%XF
2364
Gum
H pr
otei
n (g
umH)
64%
XF23
62Gu
mJ
prot
ein
(gum
J)65
%XF
2361
Gum
K pr
otei
n (g
umK)
69%
XF23
60Gu
mM
pro
tein
(gum
M)
69%
PATH
OGEN
ICIT
Y, V
IRUL
ENCE
, AN
D AD
APTA
TION
/SUR
FACE
PROT
EIN
S
XF08
89he
mag
glut
inin
-like
sec
rete
d pr
otei
n (p
spA)
25%
XF21
96he
mag
glut
inin
-like
sec
rete
d pr
otei
n (p
spA)
35%
XF27
75he
mag
glut
inin
-like
sec
rete
d pr
otei
n (p
spA)
35%
XF15
29su
rface
pro
tein
(hsf
)22
%XF
1981
surfa
ce p
rote
in (h
sf)
24%
XF15
16su
rface
-exp
osed
out
er m
embr
ane
prot
ein
(usp
A1)
28%
PATH
OGEN
ICIT
Y, V
IRUL
ENCE
, AN
DAD
APTA
TION
/ADA
PTAT
ION
, ATY
PICA
L CO
NDI
TION
S
XF04
32Br
kB p
rote
in (b
rk)
33%
XF08
66co
balt-
zinc-
cadm
ium
resi
stan
ce p
rote
in(c
zcD)
53%
XF02
85he
at s
hock
pro
tein
(htrA
/deg
P/pt
d)43
%XF
2625
heat
sho
ck p
rote
in (h
tpX)
53%
XF22
34lo
w m
olec
ular
wei
ght h
eat s
hock
pro
tein
(hsp
A)38
%XF
1379
luci
fera
se31
%XF
0837
orga
nic
solv
ent t
oler
ance
pre
curs
or(im
p/os
tA)
29%
XF16
23pe
ripla
smic
glu
can
bios
ynth
esis
pro
tein
(mdo
H)37
%XF
2682
perip
lasm
ic g
luca
n bi
osyn
thes
is p
rote
in(m
doG)
36%
XF07
85su
lfur d
epriv
atio
n re
spon
se re
gula
tor (
sac1
)22
%XF
0858
surv
ival
pro
tein
(sur
E)60
%XF
2622
tem
pera
ture
acc
limat
ion
prot
ein
B (ta
pB)
85%
XF04
18to
luen
e to
lera
nce
prot
ein
(ttg2
D)24
%XF
0419
tolu
ene
tole
ranc
e pr
otei
n (tt
g2C)
41%
XF04
20to
luen
e to
lera
nce
prot
ein
(ttg2
B)42
%XF
0421
tolu
ene
tole
ranc
e pr
otei
n (tt
g2A)
50%
PATH
OGEN
ICIT
Y, V
IRUL
ENCE
, AN
D AD
APTA
TION
/OTH
ER
XF02
90ac
onita
se (r
pfA)
79%
XF14
24ch
itina
se (c
hi)
43%
XF15
27ge
nera
l sec
reto
ry p
athw
ay p
rote
in D
prec
urso
r (xp
sD/p
efD)
68%
XF15
17ge
nera
l sec
reto
ry p
athw
ay p
rote
in E
(xps
E/pe
fE)
81%
XF15
18ge
nera
l sec
reto
ry p
athw
ay p
rote
in F
(xps
F)66
%XF
1519
gene
ral s
ecre
tory
pat
hway
pro
tein
Gpr
ecur
sor (
xpsG
/pef
G)78
%XF
1520
gene
ral s
ecre
tory
pat
hway
pro
tein
Hpr
ecur
sor (
xpsH
)67
%XF
1521
gene
ral s
ecre
tory
pat
hway
pro
tein
Ipr
ecur
sor (
xpsI
/pef
I)55
%XF
1522
gene
ral s
ecre
tory
pat
hway
pro
tein
Jpr
ecur
sor (
xpsJ
/pef
J)56
%XF
1523
gene
ral s
ecre
tory
pat
hway
pro
tein
K (p
efK)
60%
XF15
24ge
nera
l sec
reto
ry p
athw
ay p
rote
in L
(pef
L)60
%XF
1020
path
ogen
icity
-rela
ted
prot
ein
48%
XF07
20pr
otei
c ki
ller a
ctiv
e pr
otei
n (h
igB)
33%
XF07
21pr
otei
c ki
ller s
uppr
essi
on p
rote
in (h
igA)
40%
XF02
23qu
euin
e tR
NA-
ribos
yltra
nsfe
rase
(tgt
/vac
C)61
%XF
0287
regu
lato
r of p
atho
geni
city
fact
ors
(rpfB
)72
%XF
1114
regu
lato
r of p
atho
geni
city
fact
ors
(rpfC
)59
%XF
1115
regu
lato
r of p
atho
geni
city
fact
ors
(rpfF
)67
%XF
1987
VacB
pro
tein
(vac
B)46
%
XF05
91vi
rule
nce
fact
or48
%XF
2420
viru
lenc
e fa
ctor
(mvi
N)
39%
XF07
54vi
rule
nce
prot
ein
(acv
B)39
%XF
2679
viru
lenc
e pr
otei
n (a
cvB)
28%
XF07
49vi
rule
nce
regu
lato
r (xr
vA)
47%
XF14
93vi
rule
nce
regu
lato
r (xr
vA)
46%
XF05
06vi
rule
nce-
asso
ciat
ed p
rote
in E
(vap
E)31
%XF
2121
viru
lenc
e-as
soci
ated
pro
tein
E (v
apE)
32%
ORFS
WIT
H UN
DEFI
NED
CAT
EGOR
Y
XF21
49Ap
aG p
rote
in (a
paG/
corD
)45
%XF
1828
ATPa
se28
%XF
0961
bact
erio
ferri
tin c
omig
rato
ry p
rote
in (b
cp)
51%
XF11
20bi
func
tiona
l DGT
P-py
roph
osph
ohyd
rola
se/t
hiam
ine
phos
phat
esy
ntha
se (m
utT/
thiE
1)43
%XF
1796
bifu
nctio
nal t
rans
crip
tiona
l rep
ress
or o
fth
e bi
otin
ope
ron/
biot
in a
cety
l-CoA
-car
boxy
lase
synt
heta
se (b
irA/b
ioR/
dhbB
)40
%XF
1624
carb
oxyl
este
rase
(est
B)37
%XF
0063
com
pete
nce
prot
ein
F (c
omF)
37%
XF11
79co
mpe
tenc
e re
late
d pr
otei
n (c
omM
)52
%XF
1078
DNA
upta
ke p
rote
in (c
omA)
35%
XF22
43GT
P bi
ndin
g pr
otei
n (le
pA)
69%
XF12
13GT
P-bi
ndin
g el
onga
tion
fact
or p
rote
in (t
ypA)
69%
XF00
88GT
P-bi
ndin
g pr
otei
n (h
flX)
54%
XF04
65GT
P-bi
ndin
g pr
otei
n (y
fgK)
46%
XF14
30GT
P-bi
ndin
g pr
otei
n (y
ihA)
52%
XF24
22GT
P-bi
ndin
g pr
otei
n (y
hbZ)
55%
XF26
41GT
P-bi
ndin
g pr
otei
n68
%XF
1668
HicB
-rela
ted
prot
ein
38%
XF11
92in
tegr
al m
embr
ane
prot
ein
49%
XF19
27in
tegr
al m
embr
ane
prot
ein
---%
XF26
45is
open
teny
l mon
opho
spha
te k
inas
e (ip
k)50
%XF
0145
oxid
ored
ucta
se (y
dfG)
50%
XF17
44ox
idor
educ
tase
71%
XF17
23su
gar-p
hosp
hate
deh
ydro
gena
se (y
rpG)
55%
XF17
24su
gar-p
hosp
hate
deh
ydro
gena
se (y
rpG)
55%
pXF5
1 ge
nes:
INTE
RMED
IARY
MET
ABOL
ISM
/REG
ULAT
ORY
FUN
CTIO
NS
XFa0
001
trans
crip
tiona
l reg
ulat
or31
%XF
a005
7tra
nscr
iptio
nal r
egul
ator
(kor
A)40
%
MAC
ROM
OLEC
ULE
MET
ABOL
ISM
/DN
AM
ETAB
OLIS
M/R
eplic
atio
n
XFa0
061
sing
le-s
trand
bin
ding
pro
tein
(ssb
)54
%XF
a000
3to
pois
omer
ase
I (to
pA/s
upX)
35%
MAC
ROM
OLEC
ULE
MET
ABOL
ISM
/DN
AM
ETAB
OLIS
M/R
ecom
bina
tion
XFa0
019
site
-spe
cific
reco
mbi
nase
(rin
)91
%
MOB
ILE
GEN
ETIC
ELE
MEN
TS/P
LASM
ID-R
ELAT
ED F
UNCT
ION
S
XFa0
002
conj
ugal
tran
sfer
pro
tein
(tra
L/vi
rB1)
46%
XFa0
005
conj
ugal
tran
sfer
pro
tein
(trb
C/vi
rB2)
32%
XFa0
006
conj
ugal
tran
sfer
pro
tein
(tra
A/vi
rB3)
39%
XFa0
007
conj
ugal
tran
sfer
pro
tein
(tra
B/vi
rB4)
43%
XFa0
008
conj
ugal
tran
sfer
pro
tein
(tra
C/vi
rB5)
28%
XFa0
011
conj
ugal
tran
sfer
pro
tein
(tra
D/vi
rB6)
23%
XFa0
012
conj
ugal
tran
sfer
pro
tein
(tra
E/vi
rB8)
33%
XFa0
013
conj
ugal
tran
sfer
pro
tein
(tra
O/vi
rB9)
34%
XFa0
014
conj
ugal
tran
sfer
pro
tein
(tra
F/vi
rB10
)40
%XF
a001
5co
njug
al tr
ansf
er p
rote
in(tr
aG/v
irB11
)41
%XF
a001
6co
njug
al tr
ansf
er p
rote
in (t
raG/
virD
4)35
%XF
a003
6co
njug
al tr
ansf
er p
rote
in (t
rbN
)76
%XF
a003
7co
njug
al tr
ansf
er p
rote
in (t
rbL)
45%
XFa0
039
conj
ugal
tran
sfer
pro
tein
(trb
J)54
%XF
a004
0co
njug
al tr
ansf
er p
rote
in (t
rbI)
52%
XFa0
041
conj
ugal
tran
sfer
pro
tein
(trb
H)44
%XF
a004
2co
njug
al tr
ansf
er p
rote
in (t
rbG)
65%
XFa0
043
conj
ugal
tran
sfer
pro
tein
(trb
F)47
%XF
a004
4co
njug
al tr
ansf
er p
rote
in (t
rbE)
---%
XFa0
047
nick
ase
(taxC
)30
%XF
a002
7pl
asm
id m
aint
enan
ce p
rote
in (p
emK)
48%
XFa0
060
plas
mid
repl
icat
ion
prot
ein
(incC
)34
%XF
a005
9pl
asm
id re
plic
atio
n/pa
rtitio
n pr
otei
n32
%XF
a002
9pl
asm
id s
tabi
lizat
ion
prot
ein
(par
E)76
%
PATH
OGEN
ICIT
Y, V
IRUL
ENCE
, AN
D AD
APTA
TION
/OTH
ER
XFa0
052
viru
lenc
e-as
soci
ated
pro
tein
D (v
apD)
64%
pXF1
.3 g
enes
:
MOB
ILE
GEN
ETIC
ELE
MEN
TS/P
LASM
ID-R
ELAT
ED F
UNCT
ION
S
XFb0
001
repl
icat
ion
prot
ein
41%
Tabl
e 3
Puta
tive
iden
tific
atio
n of
Xyl
ella
fast
idio
sa g
enes
. Gen
e nu
mbe
rs c
orre
spon
d to
thos
e in
Fig
. 1. P
erce
ntag
es re
pres
ent %
iden
titie
sre
lativ
e to
the
mos
t rel
evan
t dat
abas
e hi
t (co
nser
ved
hypo
thet
ical
s an
d %
iden
titie
s of
gen
es w
ith fr
ames
hift/
stop
cod
on in
fram
e no
t sho
wn)
.
© 2000 Macmillan Magazines Ltd
Related Documents











