The endocannabinoid system mediates aerobic exercise-induced antinociception in rats Giovane Galdino a, * , Thiago R.L. Romero a , José Felipe P. Silva b , Daniele C. Aguiar a , Ana Maria de Paula c , Jader S. Cruz d , Cosimo Parrella e , Fabiana Piscitelli e , Igor D. Duarte a , Vincenzo Di Marzo e , Andrea C. Perez a a Department of Pharmacology, Institute of Biological Sciences, Federal University of Minas Gerais, Ave. Antônio Carlos 6627, 31270-100 Belo Horizonte, Brazil b Department of Physiology, Institute of Biological Sciences, Federal University of Minas Gerais, Ave. Antônio Carlos 6627, 31270-100 Belo Horizonte, Brazil c Biophotonics Lab, Department of Physics, Federal University of Minas Gerais, Ave. Antônio Carlos 6627, 31270-901 Belo Horizonte, Brazil d Department of Biochemistry, Institute of Biological Sciences, Federal University of Minas Gerais, Ave. Antônio Carlos 6627, 31270-100 Belo Horizonte, Brazil e Endocannabinoid Research Group, Institute of Biomolecular Chemistry, National Research Council, via Campi Flegrei 34, Compresorio Olivetti, 80078 Pozzuoli, Napoli, Italy article info Article history: Received 18 April 2013 Received in revised form 6 September 2013 Accepted 20 September 2013 Keywords: Endocannabinoids Cannabinoid receptors Aerobic exercise Pain Antinociception abstract Exercise-induced antinociception is widely described in the literature, but the mechanisms involved in this phenomenon are poorly understood. Systemic (s.c.) and central (i.t., i.c.v.) pretreatment with CB 1 and CB 2 cannabinoid receptor antagonists (AM251 and AM630) blocked the antinociception induced by an aerobic exercise (AE) protocol in both mechanical and thermal nociceptive tests. Western blot analysis revealed an increase and activation of CB 1 receptors in the rat brain, and immunofluorescence analysis demonstrated an increase of activation and expression of CB1 receptors in neurons of the periaqueductal gray matter (PAG) after exercise. Additionally, pretreatment (s.c., i.t. and i.c.v.) with endocannabinoid metabolizing enzyme inhibitors (MAFP and JZL184) and an anandamide reuptake inhibitor (VDM11) prolonged and intensified this antinociceptive effect. These results indicate that exercise could activate the endocannabinoid system, producing antinociception. Supporting this hypothesis, liquid- chromatography/mass-spectrometry measurements demonstrated that plasma levels of endocannabi- noids (anandamide and 2-arachidonoylglycerol) and of anandamide-related mediators (palmitoyletha- nolamide and oleoylethanolamide) were increased after AE. Therefore, these results suggest that the endocannabinoid system mediates aerobic exercise-induced antinociception at peripheral and central levels. Ó 2013 Elsevier Ltd. All rights reserved. 1. Introduction Several studies have demonstrated the involvement of the endocannabinoid system in pain relief (Pertwee, 2001). This system comprises two G protein-coupled membrane receptors (CB 1 and CB 2 ) negatively coupled to adenylate cyclase and positively coupled to mitogen-activated protein kinase (Mu et al., 1999; Pertwee, 2001). CB 1 receptors have been found in the central nervous sys- tem (CNS), including structures that participate in the descending control of pain, such as the periaqueductal gray matter (PAG), the rostroventromedial medulla (RVM) and the dorsal horn of the spinal cord (Herkenham et al., 1991; Tsou et al., 1998). These re- ceptors are also found in the peripheral nervous system in the dorsal root ganglia (DRG) (Fan et al., 2011). In contrast, CB 2 re- ceptors are mostly restricted to immune cells (Pertwee and Ross, 2002). However, recent studies have described these receptors in the central nervous system (Onaivi et al., 2012). CB 1 receptors, when activated by their endogenous ligands (endocannabinoids), promote hyperpolarization and a reduction in the rate of firing of excitable cells, suppressing neurotransmitter releasing, as well as a reduction in the nociceptive impulse (Di Marzo et al., 2004). The endocannabinoid arachidonoylethanolamide (anandamide, or AEA) is hydrolyzed by the enzyme fatty acid amide hydrolase (FAAH), whereas the other endogenous cannabinoid receptor ligand, 2-arachidonoylglycerol (2-AG), is degraded by the enzyme monoacylglycerol lipase (MGL) (Di Marzo et al., 2004). FAAH also * Corresponding author. Ave. Jovino Fernandes Sales, 2600, 37130-000 Alfenas, Brazil. Tel.: þ55 35 3299 1000. E-mail address: [email protected] (G. Galdino). Contents lists available at ScienceDirect Neuropharmacology journal homepage: www.elsevier.com/locate/neuropharm 0028-3908/$ e see front matter Ó 2013 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.neuropharm.2013.09.022 Neuropharmacology 77 (2014) 313e324

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

lable at ScienceDirect
Neuropharmacology 77 (2014) 313e324
Contents lists avai
Neuropharmacology
journal homepage: www.elsevier .com/locate/neuropharm
The endocannabinoid system mediates aerobic exercise-inducedantinociception in rats
Giovane Galdino a,*, Thiago R.L. Romero a, José Felipe P. Silva b, Daniele C. Aguiar a,Ana Maria de Paula c, Jader S. Cruz d, Cosimo Parrella e, Fabiana Piscitelli e, Igor D. Duarte a,Vincenzo Di Marzo e, Andrea C. Perez a
aDepartment of Pharmacology, Institute of Biological Sciences, Federal University of Minas Gerais, Ave. Antônio Carlos 6627,31270-100 Belo Horizonte, BrazilbDepartment of Physiology, Institute of Biological Sciences, Federal University of Minas Gerais, Ave. Antônio Carlos 6627, 31270-100 Belo Horizonte, BrazilcBiophotonics Lab, Department of Physics, Federal University of Minas Gerais, Ave. Antônio Carlos 6627, 31270-901 Belo Horizonte, BrazildDepartment of Biochemistry, Institute of Biological Sciences, Federal University of Minas Gerais, Ave. Antônio Carlos 6627, 31270-100 Belo Horizonte, Brazile Endocannabinoid Research Group, Institute of Biomolecular Chemistry, National Research Council, via Campi Flegrei 34, Compresorio Olivetti,80078 Pozzuoli, Napoli, Italy
a r t i c l e i n f o
Article history:Received 18 April 2013Received in revised form6 September 2013Accepted 20 September 2013
Keywords:EndocannabinoidsCannabinoid receptorsAerobic exercisePainAntinociception
* Corresponding author. Ave. Jovino Fernandes SalBrazil. Tel.: þ55 35 3299 1000.
E-mail address: [email protected] (G.
0028-3908/$ e see front matter � 2013 Elsevier Ltd.http://dx.doi.org/10.1016/j.neuropharm.2013.09.022
a b s t r a c t
Exercise-induced antinociception is widely described in the literature, but the mechanisms involved inthis phenomenon are poorly understood. Systemic (s.c.) and central (i.t., i.c.v.) pretreatment with CB1 andCB2 cannabinoid receptor antagonists (AM251 and AM630) blocked the antinociception induced by anaerobic exercise (AE) protocol in both mechanical and thermal nociceptive tests. Western blot analysisrevealed an increase and activation of CB1 receptors in the rat brain, and immunofluorescence analysisdemonstrated an increase of activation and expression of CB1 receptors in neurons of the periaqueductalgray matter (PAG) after exercise. Additionally, pretreatment (s.c., i.t. and i.c.v.) with endocannabinoidmetabolizing enzyme inhibitors (MAFP and JZL184) and an anandamide reuptake inhibitor (VDM11)prolonged and intensified this antinociceptive effect. These results indicate that exercise couldactivate the endocannabinoid system, producing antinociception. Supporting this hypothesis, liquid-chromatography/mass-spectrometry measurements demonstrated that plasma levels of endocannabi-noids (anandamide and 2-arachidonoylglycerol) and of anandamide-related mediators (palmitoyletha-nolamide and oleoylethanolamide) were increased after AE. Therefore, these results suggest that theendocannabinoid system mediates aerobic exercise-induced antinociception at peripheral and centrallevels.
� 2013 Elsevier Ltd. All rights reserved.
1. Introduction
Several studies have demonstrated the involvement of theendocannabinoid system in pain relief (Pertwee, 2001). This systemcomprises two G protein-coupled membrane receptors (CB1 andCB2) negatively coupled to adenylate cyclase and positively coupledto mitogen-activated protein kinase (Mu et al., 1999; Pertwee,2001). CB1 receptors have been found in the central nervous sys-tem (CNS), including structures that participate in the descendingcontrol of pain, such as the periaqueductal gray matter (PAG), therostroventromedial medulla (RVM) and the dorsal horn of the
es, 2600, 37130-000 Alfenas,
Galdino).
All rights reserved.
spinal cord (Herkenham et al., 1991; Tsou et al., 1998). These re-ceptors are also found in the peripheral nervous system in thedorsal root ganglia (DRG) (Fan et al., 2011). In contrast, CB2 re-ceptors are mostly restricted to immune cells (Pertwee and Ross,2002). However, recent studies have described these receptors inthe central nervous system (Onaivi et al., 2012). CB1 receptors,when activated by their endogenous ligands (endocannabinoids),promote hyperpolarization and a reduction in the rate of firing ofexcitable cells, suppressing neurotransmitter releasing, as well as areduction in the nociceptive impulse (Di Marzo et al., 2004). Theendocannabinoid arachidonoylethanolamide (anandamide, orAEA) is hydrolyzed by the enzyme fatty acid amide hydrolase(FAAH), whereas the other endogenous cannabinoid receptorligand, 2-arachidonoylglycerol (2-AG), is degraded by the enzymemonoacylglycerol lipase (MGL) (Di Marzo et al., 2004). FAAH also

G. Galdino et al. / Neuropharmacology 77 (2014) 313e324314
partly regulates the levels of N-acylethanolamine, i.e., palmitoyle-thanolamide (PEA) and oleoylethanolamide (OEA), which haveAEA-like molecular structures and metabolic pathways, but do notpotently activate cannabinoid receptors (Matias et al., 2007). BothPEA and OEA can, however, interact with several targets, includingthe peroxisome proliferator-activated receptor-a (PPAR-a) and thetransient receptor potential vanilloid channel-1 (TRPV1) (Fu et al.,2003; Piomelli et al., 2005). However, some studies have shownthat some effects of PEA are inhibited by cannabinoid receptorantagonists, thus confirming the hypothesis that this compoundcan also act as an “entourage” compound for endocannabinoids byenhancing their activity at their receptors or by inhibiting theirdegradation (Calignano et al., 1998; Onaivi et al., 2008; Petrosinoet al., 2010; Re et al., 2007).
In addition, the antinociceptive activity of AEA, 2-AG, PEA, andOEA has been confirmed in experiments using central or peripheraladministration into mice and rats, in different acute pain models(Costa et al., 2008; Pertwee, 2001; Suardiaz et al., 2007). Finally,FAAH and MGL inhibitors reduce hyperalgesia in mice and rats andincrease stress-induced analgesia in rats (Connell et al., 2006; Costaet al., 2008; Guindon et al., 2011; Haller et al., 2006; Naidu et al.,2009). These effects are reversed by cannabinoid receptorantagonists.
The endocannabinoid system is activated by exercise. Sparlinget al. (2003) found an increase in anandamide plasma levels inhumans after running and cycling. Voluntary exercise producedincreased endocannabinoid signaling within the hippocampus ofrats and cannabinoid receptor sensitization in the striatum of mice(De Chiara et al., 2010; Hill et al., 2010). More recently, it was alsoshown that intense exercise increases circulating endocannabinoidlevels in humans (Feuerecker et al., 2011; Heyman et al., 2011).
Exercise-induced analgesia has been reported in the literaturesince the early 1980s. Aerobic exercises aremore frequently studiedin rats and mice subjected to treadmills or swimming and inhumans subjected to running and cycling (Koltyn, 2002). However,after 30 years of research, few mechanisms have been shown tounderlie this effect, including, for example, those involvingendogenous opioid, serotonin and nitric oxide/cGMP/Kþ
ATP path-ways (Galdino et al., 2010; Koltyn, 2000). Thus, to elucidate themechanisms involved in the exercise-induced antinociception, weinvestigated the possible role of the endocannabinoid system inthis phenomenon.
2. Material and methods
2.1. Animals
The study was conducted in concordance with the International Association forthe Study of Pain guidelines on the use of laboratory animals (Zimmermann, 1983),and all the experiments were approved by the Ethics Committee for AnimalExperimentation of the Federal University of Minas Gerais (UFMG). The experimentswere performed using male Wistar rats that weighed 180e200 g obtained fromUFMG Brazil. The animals were divided in groups and housed in individual cagesthat were kept at a constant temperature 23 �C with a 12 h lightedark cycle, andthey had free access to food and water. Animals were kept under these conditionsuntil they were taken to the testing room, at least 1 h before the experiments.
2.2. Drugs
The following drugs were used in this study: N-(Piperidin-1-yl)-5-(4-iodophenyl)-1-(2,4-dichlorophenyl)-4-methyl-1H-pyrazole-3-carboxamide(AM251), a CB1 cannabinoid receptor antagonist; [6-Iodo-2-methyl-1-[2-(4-morpholinyl)ethyl]-1H-indol-3-yl](4-methoxyphenyl)methanone (AM630), a CB2
cannabinoid receptor antagonist; (5Z,8Z,11Z,14Z)-5,8,11,14-eicosatetraenyl-methyl-ester phosphonofluoridic acid (MAFP), an irreversible FAAH inhibitor; 4-[Bis(1,3-benzodioxol-5-yl)hydroxymethyl]-1-piperidinecarboxylic acid 4-nitrophenyl ester(JZL184), a potent and selective MGL inhibitor; (5Z,8Z,11Z,14Z) N (4Hydrox-y2methylphenyl) 5,8,11,14 eicosatetraenamide (VDM11), a selective inhibitor of theanandamide membrane transporter, all purchased from Tocris Bioscience (Ellisville,MO). AM251, AM630 and JZL184 were dissolved in physiological saline and dimethylsulphoxide (20%, Sigma, MO, USA) vehicle. MAFP was dissolved in physiological
saline and ethanol (2.0%, Merck, NJ, USA) vehicle, and VDM11 was dissolved inTocrisolve (Tocris, MN, USA) vehicle. All the drugs were administered in a volume of1 ml/kg (s.c.), 10 ml (i.t.) and 5 ml (i.c.v.).
2.3. Injections
2.3.1. Subcutaneous injectionThe subcutaneous injections were into the dorsal nuchal area of rats in a volume
of 1 ml/kg, 10 min before exercise in the exercised groups.
2.3.2. Intrathecal injectionThe intrathecal injections (i.t.) were performed in a volume of 10 ml in the
subarachnoid space between L5 and L6 using a 30 G � 1/2-inch needle and a 50 mlHamilton syringe as described by Mestre et al. (1994). Intrathecal injection isstressful for rats and, according to IASP recommendation, requires anesthesia. Thus,before injection, rats were slightly anesthetized with volatile isoflurane (3.5%) andrecovered 5 min after the removal from the anesthesia chamber. Correct i.t. posi-tioning of the needle tip was confirmed by a characteristic tail-flick response in theanimal. Lidocaine 4% (10 ml), was administered to a group of test animals, usingtemporary paralysis of the hind limbs as an end point to confirm the effectiveness ofthe injection technique. Intrathecal injections were administered immediatelybefore to exercise.
2.3.3. Intracerebroventricular injectionInitially before intracerebroventricular injections, each rat was anaesthetized
with a mixture of ketamine (80 mg/kg) and xylazine (10 mg/kg) injected i.p., andthen placed in a stereotaxic apparatus (Stoelting, IL, USA). The scalp was incised, andthe skull was leveled off around the bregma. A 22 gauge,12mm stainless-steel guidecannula was inserted in the right lateral ventricle of the brain. The type of thecannula aimed at the following coordinates: 1.5 mm posterior to the bregma,2.5 mm lateral to the midline, and 3.3 mm below the top of the skull (Paxinos andWatson, 1986). The cannula was then fixed to the skull using three screw anddental acrylic. 12.5 mm stylet was inserted in the cannula to keep in patent prior toinjection. Animals were allowed 5-day recovery period before experiments wereinitiated. For i.c.v. drugs injections, a gauge, 12.5 mm injection needle was attachedto a 30 cm polyethylene tube fitted to a 10 ml Hamilton syringe. Then, the rats wererestrained by hand, the stylet was withdrawn, and the injection needle inserted intothe guide cannula. The volume of the solutions to be injected into lateral ventriclewas 5 ml, and the injection was made over a period of 120 s. Intracerebroventricularinjections were administered immediately before the exercise.
2.4. Exercise
Acute aerobic exercise (AE) was performed using a rodent treadmill. Animals ranwith a progressive speed at 20 m/min and 0% inclination, an average time of 49.06(�3) min, until fatigue (Ji et al., 1992). Fatigue was defined as the point at which theanimals were unable to keep pace with treadmill (Soares et al., 2003). The back ofthe treadmill has an electrical stimulator (3 v) in order to encourage the animals torun. To familiarize the rats to exercise, reducing the effects of stress, they ran daily onthe treadmill at 5 m/min for 5 min/day during 3 consecutive days prior to theexperiments.
The groups were as follows (n ¼ 6 per group): Control (Co): animals that did notperform exercise and received saline; Acute aerobic exercise (AE): rats that ran andreceived saline; AE þ AM251: animals pretreated with CB1 cannabinoid receptorantagonist and exercised; AE þ AM630: animals pretreated with CB2 cannabinoidreceptor antagonist and exercised; AE þMAFP: animals pretreated with irreversibleFAAH inhibitor and exercised; AE þ JZL184: animals pretreated with MGL inhibitorand exercised; AE þ VDM11: animals pretreated with inhibitor of the anandamidemembrane transporter and exercised. Furthermore, were used different groupscomposed of animals that received the drugs previously described via s.c., i.t. andi.c.v. administration. A group received the same numbers of electrical stimulus of AEgroup, and there was no change on the nociceptive threshold.
2.5. Assessment of mechanical nociceptive threshold
Mechanical nociceptive threshold was assessed by measuring the response to apaw pressure test described by Randall and Selitto (1957). In a quiet room, rats wereplaced in acrylic cages (12 � 20 � 17 cm) that had wire grid floors 1 h before thetesting began. An analgesimeter (Ugo Basile, Comerio, Italy) with a cone-shapedpaw-presser that had a rounded tip (9 mm base diameter) was used to apply alinearly increasing force to the hind paw. The pressure intensity in grams (g) thatcaused an escape reaction was defined as the nociceptive threshold. A maximumintensity of 300 g was used to reduce the possibility of damage to the paws. Thenociceptive thresholdwasmeasured in the right paw and determined as the averageof three consecutive trials.
2.6. Assessment of thermal nociceptive threshold
Thermal nociceptive threshold was assessed by measuring the latency to tailwithdrawal from a noxious heat source, as described by D’amour and Smith (1941).Rats were placed in acrylic cages (12 � 20 � 17 cm) that had wire grid floors in a

G. Galdino et al. / Neuropharmacology 77 (2014) 313e324 315
quiet room 1 h before the testing began. In this test, the animal’s tails were placedon the tail flick apparatus and smoothed into a groove that contained a photocell. Alight source was used as the thermal stimulus. The light was activated andremained focused on the tail until the rat moved its tail (a spinal reflex), thusswitching the light off. The intensity of the light was adjusted to obtain a baselinetail-flick latency of 2e4 s and a maximum duration of 9 s was chosen to preventtissue damage.
2.7. Research design
First, the baseline latency of nociceptive threshold of each rat was measured ineach nociceptive test, starting from the test paw withdrawal, and after 3 min by thetail-flick test. In exercised groups, it was measured before the onset of exercise.Immediately after exercise, a new measurement was performed and then each15 min during the duration of the antinociceptive effect. The control group under-went similar measurements. The drugs were injected after the measurement of thenociceptive threshold and before the onset of exercise.
2.8. Western blot analysis
A group composed of non-exercised animals (control, n ¼ 5) and a groupcomposed of exercised animals (n ¼ 5) were killed by decapitation and immediatelyafter brains were homogenized in RIPA buffer and in cocktail of protease inhibitors(Sigma, MO, USA). Then 200 mg of brain tissue was used for each 500 ml lysis bufferenriched. Afterwards, the samples were centrifuged at 10,000 rpm for 10 min andproteins amount measured using the Lowry method (Lowry et al., 1951). Proteinswere separated by SDSepolyacrylamide gel electrophoresis (SDSePAGE-12%) andtransblotted onto nitrocellulose membranes (Amersham Pharmacia Biotech, LittleChalfont, UK). The membranes were blocked overnight with 2.5% dry milk and thenincubated overnight at 6 �C with rabbit polyclonal antibody against CB1 receptors(1:200; AbCam, MA, USA). The membranes were thenwashed and incubated for 2 hat room temperaturewith HRP-conjugated secondary antibody (1:2000; Santa Cruz,CA, USA). The blots were visualized in ECL solution (Amersham Pharmacia Biotech,Little Chalfont, UK) for 2 min and exposed onto hyperfilms (Amersham PharmaciaBiotech, Little Chalfont, UK) for 2e20 min. aeTubulin (1:2000; AbCam, MA, USA)antibody was used as a loading control. A total of 8 western blot trials were con-ducted and transferred to nitrocellulose membranes containing proteins fromantisense and mismatch samples. Films were scanned into Image J for analysis. Acomputer-based imaging system (Gel-Pro Analyzer) was used to measure the in-tensity of the optical density of 60 kDa bands that represented the molecular weightof the CB1 protein.
2.9. CB1 receptors and c-FOS double-immunofluorescence
To show the presence of co-localization of CB1 receptors and c-FOS in the PAGwe performed a double label immunofluorescence in sections from the dorsolateralPAG (dlPAG) and ventrolateral PAG (vlPAG) as previously described by Casarottoet al. (2012). Briefly, a group of runner rats (n ¼ 5) and a group composed ofnon-runner rats (n ¼ 5) were anaesthetized two hours after aerobic exercise (AE)with deep urethane anesthesia and perfused transcardially with saline followed by4% paraformaldehyde (PFA 4%) in 0.1 M phosphate buffer (PBS, pH 7.4). The brainswere removed and post fixed over 2 h in PFA 4% and stored for at least 30 h in 30%sucrose for cryoprotection. Coronal sections (25 mm) were obtained in a cryostatand were washed in PBS 0.01 M. After this, the sections were incubated in glicine0.1 M, washed again and incubated for 1 h in 5% bovine serum (BSA, Sigma, USA) inPBS 0.01 M, pH 7.4 containing 0.3% Triton X-100. Following, the sections wereincubated for 2 days at room temperature in a mixture of anti-CB1 receptor N-terminus (diluted 1:200, Santa Cruz raised in goat) to c-FOS N-terminus (diluted1:2000, Santa Cruz raised in rabbit) diluted in PBS 0.01 M containing BSA 5% and0.3% Triton X-100. After incubation in the primary antiserum, the tissue sectionswere washed in PBS and sequentially incubated for 1 h in a mixture of Alexa 594donkey anti-rabbit IgGs (1:1000; Molecular Probes, Eugene, OR, USA) and Alexa488 donkey anti-goat IgGs (1:1000; Molecular Probes, Eugene, OR, USA). Slideswere then rinsed in PBS and coverslipped with Fluormount-G (Electron MicroscopySciences, Hatfield, PA, USA). Images from sections were first obtained using anOlympus BX50 microscope through computerized image system (Image Pro-Plus4.0, Media Cybernetics). For a more detailed analysis of the double-stained cells,the slides were re-examined using an Olympus FV300 confocal system in an up-right configuration (BX61WI microscope), and an UPlan FLN10X/0.3 objective.The immunostained slides (Alexa Fluor 488-coupled anti-CB1 cannabinoid re-ceptors; Alexa Fluor 546-coupled anti-c-FOS) were excited by an argon laser at thewavelength of 488 nm and a HeeNe laser at 543 nm, respectively. The fluorescenceemission were separated by a dichroic mirror at 570 nm and collected usingbandpass filters at 510e540 nm for the Alexa 488 (green channel) and at 560e600 nm for the Alexa 546 (red channel). The green and red channels were mergeddigitally by the Olymous Fluoview software and the images were exported in TIFformat.
As immunofluorescence control for non-specific labeling, the brain sectionswere incubated without primary antibodies.
2.9.1. Analysis of immunofluorescence dataThe identification method of double-stained cells was similar to that described
in previous works (Aguiar and Guimaraes, 2009; Beijamini and Guimaraes, 2006).The number of c-FOS-like immunoreactive cells (FLI), and CB1 immunoreactive cells(CB1IR) and doubled-stained cells were manually counted with the help of acomputerized image analysis system Image J 1.42q (Wayne Rasband, National In-stitutes of Health, USA). An observer blind to group assignment performed theanalysis. For each group one section from each animal was evaluated. All stainedcells in the whole area of each brain region of interested were recorded. Neuroan-atomical sites were identified with the help of the Paxinos and Watson’s atlas(Paxinos and Watson, 1986). The anterioreposterior (AP) localization from bregmaof the analyzed regions were as follows: rostral portion containing the dlPAG andvlPAG was represented by level �7.4 mm from bregma.
2.10. Plasma collection
Rats submitted to aerobic exercise (n ¼ 5) and a group composed of non-exercised rats (control, n ¼ 5), were previously anesthetized with ketamine andxylazine. Blood collection was by cardiac punction using a pre-heparinized needle(80 � 10 inches), centrifuged for 10 min (2000 rpm, 313.6 G force). Plasma wascollected and stored in polyethylene tubes in a �80 �C freezer. In both groups theplasma was collected immediately after exercise.
2.11. Liquid chromatography mass spectrometry analysis of rat plasma
After collection, freezing at �80 �C and defrosting on ice, the plasma wastreated with 5 vol of chloroform:methanol (2:1) containing 10 pmol of d8-AEA, d4-palmitoylethanolamide (PEA), d5-2-arachidonoylglycerol (2-AG), d4-palmitoylethanolamide (PEA) and d4-oleoylethanolamide (OEA) and sonicated. Theextract was centrifuged at 13,000 � g for 16 min (4 �C), the aqueous phase plus debriswere collected and extracted again twice with 1 vol of chloroform. The organic phasesfrom the three extractions were pooled and the organic solvents evaporated in arotating evaporator. Lyophilized extracts were resuspended in chloroform:methanol(99:1, v v�1). The solutionswere thenpurified by open bed chromatography on silica aspreviously described (Bisogno et al.,1997). Fractions elutedwith chloroform:methanol(9:1, v v�1) and containing AEA, PEA, 2-AG and OEA, were collected and the excesssolvent evaporated with a rotating evaporator, and aliquots analyzed by isotopedilution-liquid chromatography/atmospheric pressure chemical ionization/massspectrometry (MS) carried out under conditions described previously and allowing theseparationsof 2-AG, PEA, AEAandOEA. Results are expressed aspmol perml of plasma.
2.12. Statistical analysis
The date are presented as the mean � S.E.M of the evaluated parameter andwere analyzed for statistical significance by two-way (ANOVA) followed by Bon-ferroni post-hoc test for multiple comparisons. t test (comparisons between twogroups) was used for results obtained bywestern blot analysis, immunofluorescenceand gas chromatography mass spectrometry. The minimum level of significanceconsidered was when p < 0.05. The data are reported as the mean (95% two-tailedCI). Statistical analyses and preparation of figures were performed using GraphPadPrism software, Version 4 (GraphPad Software, La Jolla, CA).
3. Results
3.1. Systemic involvement of endocannabinoid system in theaerobic exercise-induced antinociception
First, we evaluated the influence of an acute aerobic exerciseprotocol (AE) on the nociceptive threshold of rats. In both pawpressure (Fig. 1A) and tail-flick (Fig. 1B) tests, AE produced an in-crease in the nociceptive threshold. This increase was found after1 min (p < 0.01) and lasted for more than 15 min (p < 0.05) in thepaw pressure test and p < 0.01 in the tail-flick test. To evaluate thesystemic participation of cannabinoid receptors, we pre-treatedanother group of rats subcutaneously with AM251 (2 and 4 mg/kg), a CB1 receptor antagonist, and AM630 (4mg/kg), a CB2 receptorantagonist, 10 min before starting the AE protocol. SubcutaneousAM251 and AM630 totally reversed the antinociceptive effectfound 1 min (p < 0.001) and 15 min (p < 0.001) after AE in the pawpressure test (Fig. 1A) and the tail-flick test (Fig. 1B). However,2 mg/kg AM251 only partially reversed this effect after the firstminute (p < 0.05) of AE in both nociceptive tests (Fig. 1A and B). Inthese experiments, the rats ran for a mean of 41.0 (�2.25) min.
To determine the systemic involvement of endocannabinoids inexercise-induced antinociception, 10 min before AE, different groups
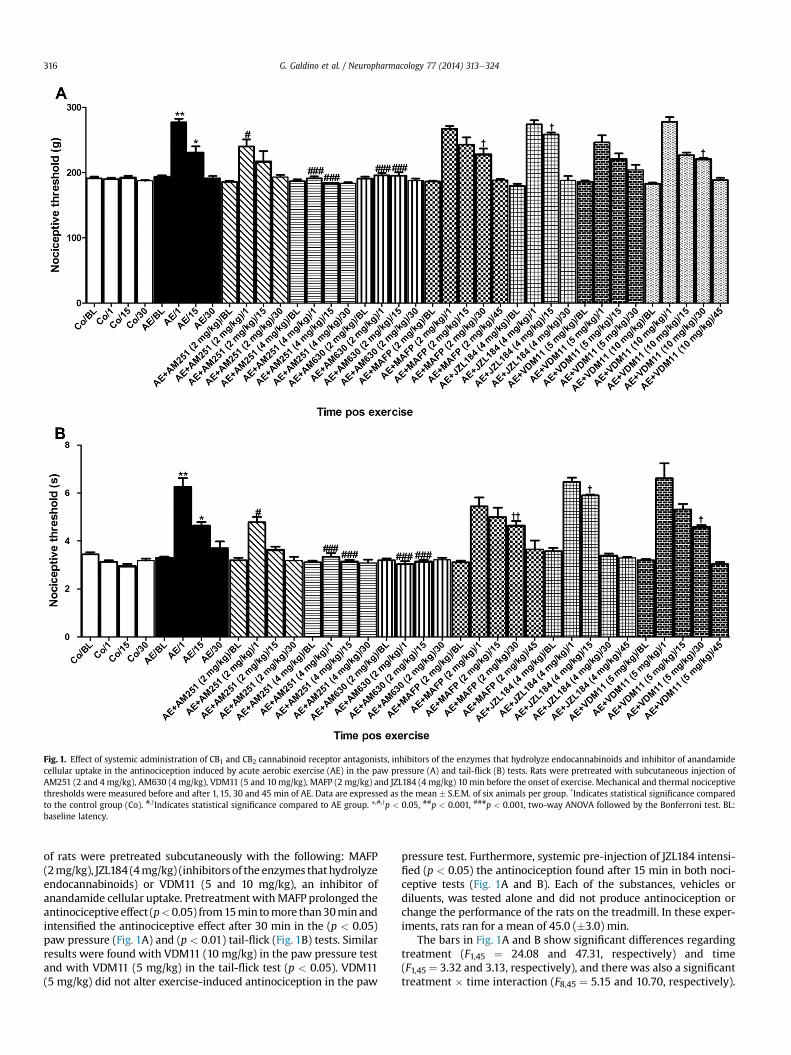
Fig. 1. Effect of systemic administration of CB1 and CB2 cannabinoid receptor antagonists, inhibitors of the enzymes that hydrolyze endocannabinoids and inhibitor of anandamidecellular uptake in the antinociception induced by acute aerobic exercise (AE) in the paw pressure (A) and tail-flick (B) tests. Rats were pretreated with subcutaneous injection ofAM251 (2 and 4 mg/kg), AM630 (4 mg/kg), VDM11 (5 and 10 mg/kg), MAFP (2 mg/kg) and JZL184 (4 mg/kg) 10 min before the onset of exercise. Mechanical and thermal nociceptivethresholds were measured before and after 1, 15, 30 and 45 min of AE. Data are expressed as the mean � S.E.M. of six animals per group. *Indicates statistical significance comparedto the control group (Co). #,yIndicates statistical significance compared to AE group. *,#,yp < 0.05, ##p < 0.001, ###p < 0.001, two-way ANOVA followed by the Bonferroni test. BL:baseline latency.
G. Galdino et al. / Neuropharmacology 77 (2014) 313e324316
of rats were pretreated subcutaneously with the following: MAFP(2mg/kg), JZL184 (4mg/kg) (inhibitors of the enzymes thathydrolyzeendocannabinoids) or VDM11 (5 and 10 mg/kg), an inhibitor ofanandamide cellular uptake. Pretreatment withMAFP prolonged theantinociceptive effect (p<0.05) from15min tomore than30min andintensified the antinociceptive effect after 30 min in the (p < 0.05)paw pressure (Fig. 1A) and (p < 0.01) tail-flick (Fig. 1B) tests. Similarresults were found with VDM11 (10 mg/kg) in the paw pressure testand with VDM11 (5 mg/kg) in the tail-flick test (p < 0.05). VDM11(5 mg/kg) did not alter exercise-induced antinociception in the paw
pressure test. Furthermore, systemic pre-injection of JZL184 intensi-fied (p < 0.05) the antinociception found after 15 min in both noci-ceptive tests (Fig. 1A and B). Each of the substances, vehicles ordiluents, was tested alone and did not produce antinociception orchange the performance of the rats on the treadmill. In these exper-iments, rats ran for a mean of 45.0 (�3.0) min.
The bars in Fig. 1A and B show significant differences regardingtreatment (F1,45 ¼ 24.08 and 47.31, respectively) and time(F1,45 ¼ 3.32 and 3.13, respectively), and there was also a significanttreatment � time interaction (F8,45 ¼ 5.15 and 10.70, respectively).
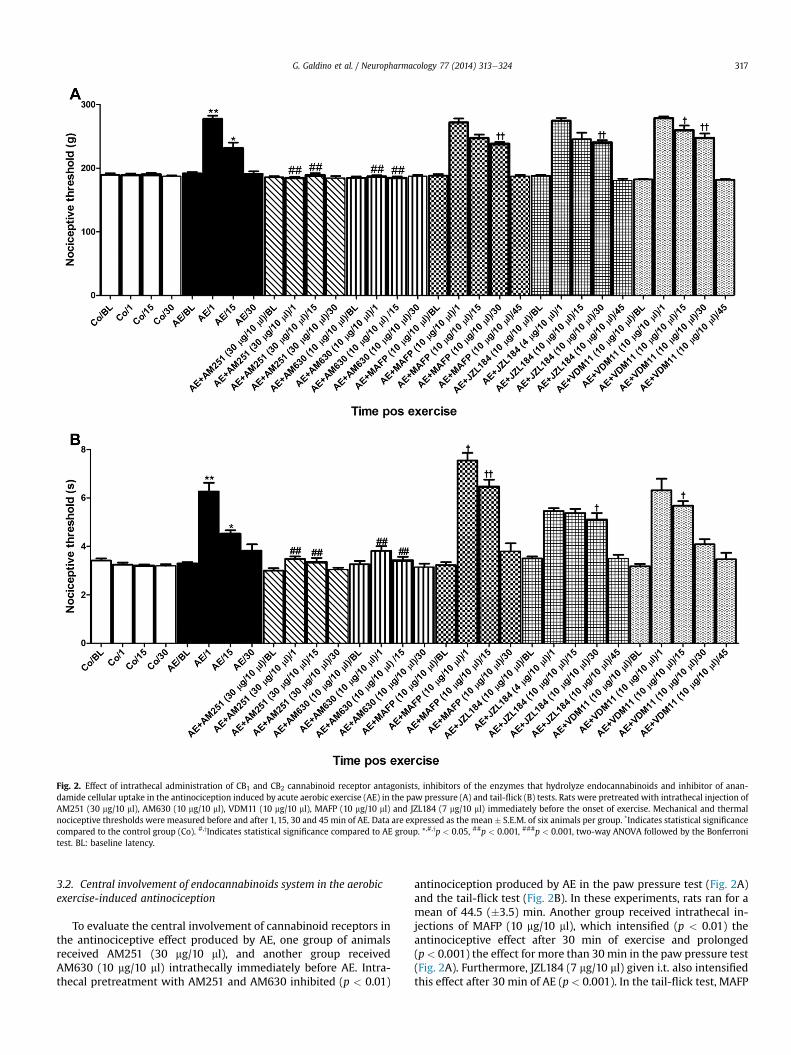
Fig. 2. Effect of intrathecal administration of CB1 and CB2 cannabinoid receptor antagonists, inhibitors of the enzymes that hydrolyze endocannabinoids and inhibitor of anan-damide cellular uptake in the antinociception induced by acute aerobic exercise (AE) in the paw pressure (A) and tail-flick (B) tests. Rats were pretreated with intrathecal injection ofAM251 (30 mg/10 ml), AM630 (10 mg/10 ml), VDM11 (10 mg/10 ml), MAFP (10 mg/10 ml) and JZL184 (7 mg/10 ml) immediately before the onset of exercise. Mechanical and thermalnociceptive thresholds were measured before and after 1, 15, 30 and 45 min of AE. Data are expressed as the mean � S.E.M. of six animals per group. *Indicates statistical significancecompared to the control group (Co). #,yIndicates statistical significance compared to AE group. *,#,yp < 0.05, ##p < 0.001, ###p < 0.001, two-way ANOVA followed by the Bonferronitest. BL: baseline latency.
G. Galdino et al. / Neuropharmacology 77 (2014) 313e324 317
3.2. Central involvement of endocannabinoids system in the aerobicexercise-induced antinociception
To evaluate the central involvement of cannabinoid receptors inthe antinociceptive effect produced by AE, one group of animalsreceived AM251 (30 mg/10 ml), and another group receivedAM630 (10 mg/10 ml) intrathecally immediately before AE. Intra-thecal pretreatment with AM251 and AM630 inhibited (p < 0.01)
antinociception produced by AE in the paw pressure test (Fig. 2A)and the tail-flick test (Fig. 2B). In these experiments, rats ran for amean of 44.5 (�3.5) min. Another group received intrathecal in-jections of MAFP (10 mg/10 ml), which intensified (p < 0.01) theantinociceptive effect after 30 min of exercise and prolonged(p< 0.001) the effect for more than 30 min in the paw pressure test(Fig. 2A). Furthermore, JZL184 (7 mg/10 ml) given i.t. also intensifiedthis effect after 30 min of AE (p < 0.001). In the tail-flick test, MAFP
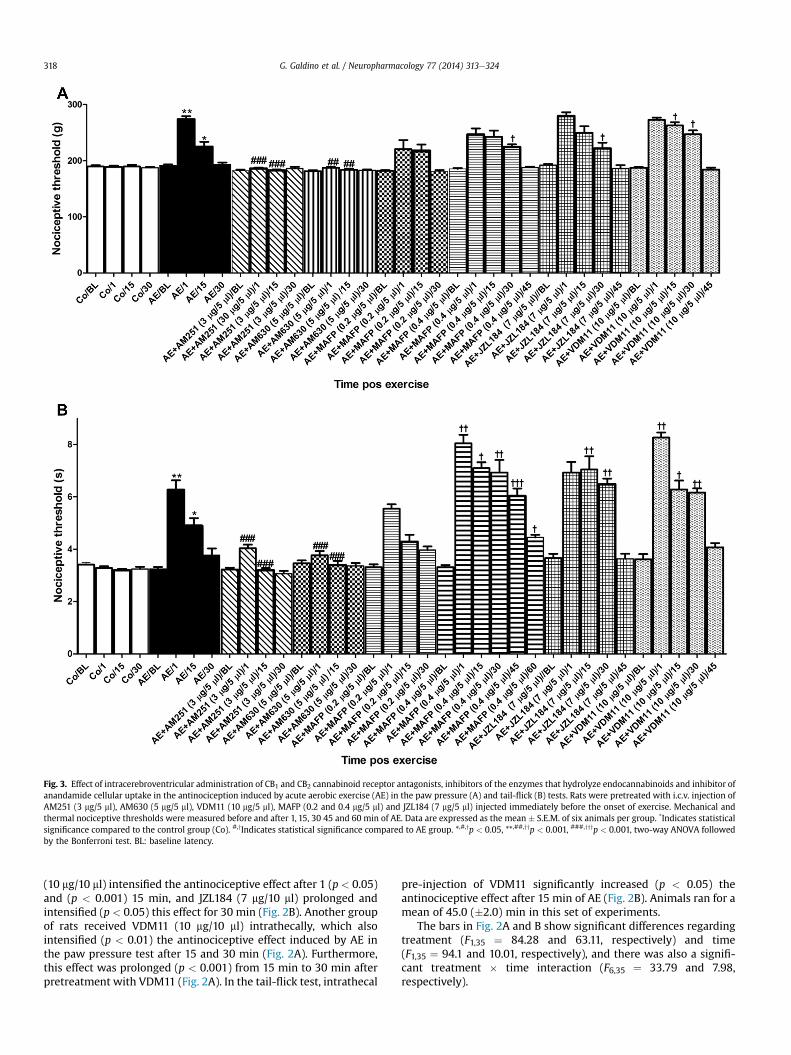
Fig. 3. Effect of intracerebroventricular administration of CB1 and CB2 cannabinoid receptor antagonists, inhibitors of the enzymes that hydrolyze endocannabinoids and inhibitor ofanandamide cellular uptake in the antinociception induced by acute aerobic exercise (AE) in the paw pressure (A) and tail-flick (B) tests. Rats were pretreated with i.c.v. injection ofAM251 (3 mg/5 ml), AM630 (5 mg/5 ml), VDM11 (10 mg/5 ml), MAFP (0.2 and 0.4 mg/5 ml) and JZL184 (7 mg/5 ml) injected immediately before the onset of exercise. Mechanical andthermal nociceptive thresholds were measured before and after 1, 15, 30 45 and 60 min of AE. Data are expressed as the mean � S.E.M. of six animals per group. *Indicates statisticalsignificance compared to the control group (Co). #,yIndicates statistical significance compared to AE group. *,#,yp < 0.05, **,##,yyp < 0.001, ###,yyyp < 0.001, two-way ANOVA followedby the Bonferroni test. BL: baseline latency.
G. Galdino et al. / Neuropharmacology 77 (2014) 313e324318
(10 mg/10 ml) intensified the antinociceptive effect after 1 (p < 0.05)and (p < 0.001) 15 min, and JZL184 (7 mg/10 ml) prolonged andintensified (p < 0.05) this effect for 30 min (Fig. 2B). Another groupof rats received VDM11 (10 mg/10 ml) intrathecally, which alsointensified (p < 0.01) the antinociceptive effect induced by AE inthe paw pressure test after 15 and 30 min (Fig. 2A). Furthermore,this effect was prolonged (p < 0.001) from 15 min to 30 min afterpretreatment with VDM11 (Fig. 2A). In the tail-flick test, intrathecal
pre-injection of VDM11 significantly increased (p < 0.05) theantinociceptive effect after 15 min of AE (Fig. 2B). Animals ran for amean of 45.0 (�2.0) min in this set of experiments.
The bars in Fig. 2A and B show significant differences regardingtreatment (F1,35 ¼ 84.28 and 63.11, respectively) and time(F1,35 ¼ 94.1 and 10.01, respectively), and there was also a signifi-cant treatment � time interaction (F6,35 ¼ 33.79 and 7.98,respectively).

Fig. 4. CB1 cannabinoid receptor expression in rat brain after acute aerobic exercise(AE) (A). Representative immunoblots for CB1 cannabinoid receptor expression withcorresponding b-actin controls are shown (B). Date expressed as the mean � SE.M. offive animals per group. *Indicates statistical significance compared to control group(Co, rats non-exercised). p < 0.05, t test.
G. Galdino et al. / Neuropharmacology 77 (2014) 313e324 319
When pre-administered intracerebroventricularly immediatelybefore AE, AM251 (3 mg/5 ml) or AM630 (5 mg/5 ml) reversed theincrease of mechanical or thermal nociceptive thresholds producedby AE (p< 0.001) (Fig. 3A and B). Themean duration of exercisewas42.75 (�2.75) min. The antinociceptive effect produced by exercisewas prolonged (p< 0.05) from 15 to 30min in the paw pressure test(Fig. 3A) and was prolonged (p< 0.05) from 15 to 60min in the tail-flick test (Fig. 3B) in the group that received intracerebroventricularpre-injection of MAFP (0.4 mg/5 ml). MAFP also intensified (p< 0.05)the antinociceptive effect after 30 min in the paw pressure test(Fig. 3A) and after 1 (p < 0.01), 15 (p < 0.05), 30 (p < 0.01), 45(p < 0.001) and 60 (p < 0.05) min in the tail-flick test (Fig. 3B).MAFP at a dose of 0.2 mg/5 ml (i.c.v.) did not alter this effect(p > 0.05) (Fig. 3A and B). In both nociceptive tests (paw pressureand tail-flick), the antinociceptive effect found after exercise wasprolonged (p < 0.05) from 15 to 30 min in the group that received apre-injection i.c.v. of JZL184 (7 mg/5 ml) (Fig. 3A and B). JZL184 (7 mg/5 ml, i.c.v.) also intensified (p < 0.05) the antinociceptive effect after30 min in the paw pressure test (Fig. 3A) and after 15 and 30 min(p < 0.001) in the tail-flick test (Fig. 3B). I.c.v. pre-injection ofVDM11 (10 mg/5 ml) intensified the antinociceptive effect producedby AE after 15 min (p < 0.05) and 30 min (p < 0.001) in the pawpressure test (Fig. 3A) and after 1 (p< 0.001), 15 min (p < 0.05) and30 min (p < 0.001) in the tail-flick test (Fig. 3B). Exercise-inducedantinociception was also prolonged (p < 0.001) from 15 to30 min after i.c.v. pre-injection of VDM11 (10 mg/5 ml) in bothnociceptive tests (Fig. 3A and B). In these experiments, rats ran for amean of 46.0 (�3.25) min. Each of the substances, vehicles or dil-uents, when tested alone, did not produce antinociception and didnot change the performance of the rats on the treadmill.
The bars in Fig. 3A and B show significant differences regardingtreatment (F1,40 ¼ 48.72 and 120.90, respectively) and time(F1,40 ¼ 26.95 and 98.95, respectively), and there was also a sig-nificant treatment � time interaction (F7,40 ¼ 13.05 and 32.21,respectively).
3.3. Aerobic exercise induces increase of CB1 receptors expression
To confirm the involvement of cannabinoid receptors inexercise-induced antinociception, we usedWestern blot analysis toshow a significant increase (p< 0.05) of CB1 receptors expression inthe brains of rats submitted to AEwhen compared to non-exercisedrats (control) (Fig. 4). In this experiment, animals ran for 40.5(�1.5) min.
3.4. Aerobic exercise activates CB1 receptors in PAG
We evaluated the expression, activation and localization of theCB1 receptor within the dorsolateral and ventrolateral PAG regions(regions important for pain modulation) by double immunofluo-rescence staining with c-FOS (a marker of neuronal activity, red)and CB1 (green).
CB1 receptors were significantly increased in neurons within thedorsolateral and ventrolateral PAG regions (p < 0.05) in the groupsubmitted to AE (n ¼ 5) when compared to the control group (non-exercised animals, n ¼ 5) (Figs. 5 and 6). Double immunofluores-cence staining for the CB1 receptor and c-FOS also demonstratedthat CB1 cannabinoid receptors were activated, upregulated andcolocalized with c-FOS (arrowheads) in both PAG regions 2 h afterexercise, which lasted for 44.0 (�1.5) min. These results indicatethat CB1 receptor expression within the dlPAG and vlPAG wasincreased by aerobic exercise and that the neurons that expressedthese receptors were activated, supporting the hypothesis thatthese receptors may participate in the antinociception induced byexercise.
3.5. Endocannabinoids levels in plasma
To determine whether exercise elevates endocannabinoidlevels, we measured AEA, 2-AG, PEA and OEA in the plasma afterthe AE protocol. Fig. 7 shows that AEA, 2-AG, PEA and OEA plasmalevels were significantly increased (df ¼ 5, t ¼ 3.67, p < 0.05) in theanimals exercised (AE, n¼ 5) when compared to non-exercised (Co,n ¼ 5). OEA baseline levels measured in the control and exercisedgroups were higher than 2-AG, PEA and AEA. Rats ran for a mean of44.0 (�2.5) min in this experiment.
4. Discussion
In the present study, moderate intensity aerobic exercise pro-duced an increase in nociceptive thresholds, as measured by ther-mal and mechanical stimuli, highlighting and confirming theimportance of exercise in reducing pain (Koltyn, 2000). In addition,Bement and Sluka (2005) demonstrated that low intensity aerobicexercise reduced muscle hyperalgesia in rats. An increase in thedental pain threshold was also found in humans, evaluated by anoxious stimulus applied to the dental pulp, followed by a cyclingexercise at moderate or high intensities (Kemppainen et al., 1990,1986). In a review, Koltyn (2002) concluded that exercise-inducedanalgesia occurs most frequently after high-intensity exercise.Our results demonstrated that moderate intensity exercise iseffective at inducing antinociception, which can be of greatimportance for the treatment of several clinical pain conditions.
This study also demonstrated that the endocannabinoid systemis involved in exercise-induced antinociception, similar to otheranalgesic systems (Koltyn, 2000). In addition, both cannabinoidreceptors (CB1 and CB2) may be involved because this anti-nociceptive effect was reversed by systemic and central pretreat-ment with AM251 and AM630. Several studies have demonstratedthe involvement of CB1 receptors in peripheral antinociception af-ter blockagewith AM251 (Hama and Sagen, 2007; Richardson et al.,1998). In addition, CB1 receptors have been identified in the DRGand peripheral neurons (Pertwee, 1997). Hohmann and Herkenham

Fig. 5. The co-expression of CB1 cannabinoid receptors and c-Fos in the dorsolateral periaqueductal gray region (dlPAG) after acute aerobic exercise. (A) Representative doubleimmunofluorescence staining of CB1 receptor (green) and c-Fos (red) in the dlPAG. Scale bars, 50 mm. (B) Number of neurons in dlPAG after aerobic exercise (AE). Data are expressedas the mean � S.E.M. of five animals per group. *Indicates statistical significance compared to the control group (CB1/Co). #Indicates statistical significance compared to controlgroup (c-Fos/Co). yIndicates statistical significance compared to control group (CB1þc-Fos/Co). p < 0.05, t-test. (C) Schematic representation of coronal section of the dlPAG (ar-rowheads on dark circle) of the rat brain (based on Paxinos and Watson, 1986). (For interpretation of the references to color in this figure legend, the reader is referred to the webversion of this article.)
G. Galdino et al. / Neuropharmacology 77 (2014) 313e324320
(1999) reported that the receptors in the DRG undergo axonaltransport to the periphery, suggesting a functional role for CB1 re-ceptors in the peripheral terminals of primary afferent neurons.These receptors are also found in high concentrations within theCNS, mainly in the important descending pathway of anti-nociception (Pertwee, 1997; Svizenska et al., 2008). In addition,studies verified that microinjection of AM251 reverses anti-nociception induced by epilepsy and by WIN55,212-2, a cannabi-noid agonist, in different rat brain sites that participate in the
Fig. 6. The co-expression of CB1 cannabinoid receptor and c-FOS in the ventrolateral periaimmunofluorescence staining of CB1 receptor (green) and c-Fos (red) in the vlPAG. Scale bexpressed as the mean � S.E.M. of five animals per group. *Indicates statistical significancecontrol group (c-Fos/Co). yIndicates statistical significance compared to control group (CB1þ(arrowheads on dark circle) of the rat brain (based on Paxinos and Watson, 1986). (For interpversion of this article.)
descending control of pain (Ebrahimzadeh and Haghparast, 2011;Samineni et al., 2011). At the spinal level, CB1 receptors have beenfound in interneurons and astrocytes in the dorsal horn (Farquhar-Smith et al., 2000; Salio et al., 2002). Further, AM251 inhibits thesuppressive effect of WIN55,212-2 on the spontaneous firing ofspinal neurons evoked by chronic constriction of the sciatic nervein rats (Liu andWalker, 2006). Thus, the reports cited above supportthe involvement of CB1 receptors in antinociception. We found thatthese receptors are involved in exercise-induced antinociception,
queductal gray region (vlPAG) after acute aerobic exercise. (A) Representative doublears, 50 mm. (B) Number of neurons in the vlPAG after aerobic exercise (AE). Data arecompared to the control group (CB1/Co). #Indicates statistical significance compared toc-Fos/Co). p < 0.05, t-test. (C) Schematic representation of coronal section of the vlPAGretation of the references to color in this figure legend, the reader is referred to the web
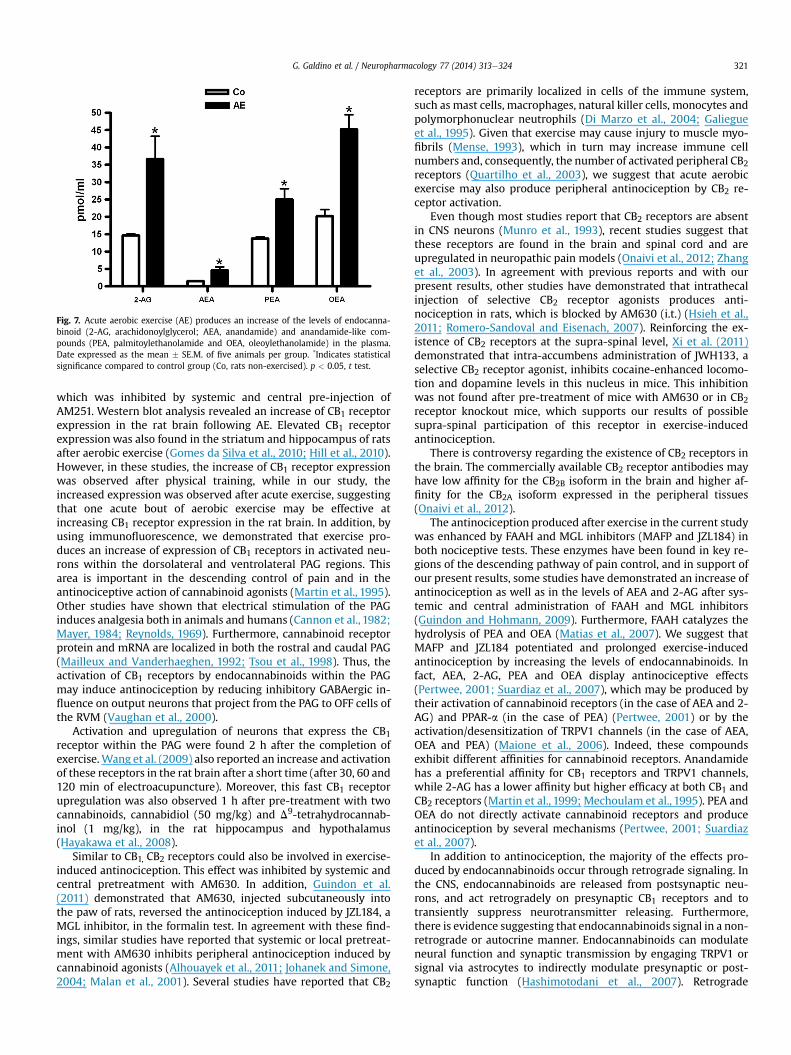
Fig. 7. Acute aerobic exercise (AE) produces an increase of the levels of endocanna-binoid (2-AG, arachidonoylglycerol; AEA, anandamide) and anandamide-like com-pounds (PEA, palmitoylethanolamide and OEA, oleoylethanolamide) in the plasma.Date expressed as the mean � SE.M. of five animals per group. *Indicates statisticalsignificance compared to control group (Co, rats non-exercised). p < 0.05, t test.
G. Galdino et al. / Neuropharmacology 77 (2014) 313e324 321
which was inhibited by systemic and central pre-injection ofAM251. Western blot analysis revealed an increase of CB1 receptorexpression in the rat brain following AE. Elevated CB1 receptorexpression was also found in the striatum and hippocampus of ratsafter aerobic exercise (Gomes da Silva et al., 2010; Hill et al., 2010).However, in these studies, the increase of CB1 receptor expressionwas observed after physical training, while in our study, theincreased expression was observed after acute exercise, suggestingthat one acute bout of aerobic exercise may be effective atincreasing CB1 receptor expression in the rat brain. In addition, byusing immunofluorescence, we demonstrated that exercise pro-duces an increase of expression of CB1 receptors in activated neu-rons within the dorsolateral and ventrolateral PAG regions. Thisarea is important in the descending control of pain and in theantinociceptive action of cannabinoid agonists (Martin et al., 1995).Other studies have shown that electrical stimulation of the PAGinduces analgesia both in animals and humans (Cannon et al., 1982;Mayer, 1984; Reynolds, 1969). Furthermore, cannabinoid receptorprotein and mRNA are localized in both the rostral and caudal PAG(Mailleux and Vanderhaeghen, 1992; Tsou et al., 1998). Thus, theactivation of CB1 receptors by endocannabinoids within the PAGmay induce antinociception by reducing inhibitory GABAergic in-fluence on output neurons that project from the PAG to OFF cells ofthe RVM (Vaughan et al., 2000).
Activation and upregulation of neurons that express the CB1receptor within the PAG were found 2 h after the completion ofexercise.Wang et al. (2009) also reported an increase and activationof these receptors in the rat brain after a short time (after 30, 60 and120 min of electroacupuncture). Moreover, this fast CB1 receptorupregulation was also observed 1 h after pre-treatment with twocannabinoids, cannabidiol (50 mg/kg) and D9-tetrahydrocannab-inol (1 mg/kg), in the rat hippocampus and hypothalamus(Hayakawa et al., 2008).
Similar to CB1, CB2 receptors could also be involved in exercise-induced antinociception. This effect was inhibited by systemic andcentral pretreatment with AM630. In addition, Guindon et al.(2011) demonstrated that AM630, injected subcutaneously intothe paw of rats, reversed the antinociception induced by JZL184, aMGL inhibitor, in the formalin test. In agreement with these find-ings, similar studies have reported that systemic or local pretreat-ment with AM630 inhibits peripheral antinociception induced bycannabinoid agonists (Alhouayek et al., 2011; Johanek and Simone,2004; Malan et al., 2001). Several studies have reported that CB2
receptors are primarily localized in cells of the immune system,such as mast cells, macrophages, natural killer cells, monocytes andpolymorphonuclear neutrophils (Di Marzo et al., 2004; Galiegueet al., 1995). Given that exercise may cause injury to muscle myo-fibrils (Mense, 1993), which in turn may increase immune cellnumbers and, consequently, the number of activated peripheral CB2receptors (Quartilho et al., 2003), we suggest that acute aerobicexercise may also produce peripheral antinociception by CB2 re-ceptor activation.
Even though most studies report that CB2 receptors are absentin CNS neurons (Munro et al., 1993), recent studies suggest thatthese receptors are found in the brain and spinal cord and areupregulated in neuropathic pain models (Onaivi et al., 2012; Zhanget al., 2003). In agreement with previous reports and with ourpresent results, other studies have demonstrated that intrathecalinjection of selective CB2 receptor agonists produces anti-nociception in rats, which is blocked by AM630 (i.t.) (Hsieh et al.,2011; Romero-Sandoval and Eisenach, 2007). Reinforcing the ex-istence of CB2 receptors at the supra-spinal level, Xi et al. (2011)demonstrated that intra-accumbens administration of JWH133, aselective CB2 receptor agonist, inhibits cocaine-enhanced locomo-tion and dopamine levels in this nucleus in mice. This inhibitionwas not found after pre-treatment of mice with AM630 or in CB2receptor knockout mice, which supports our results of possiblesupra-spinal participation of this receptor in exercise-inducedantinociception.
There is controversy regarding the existence of CB2 receptors inthe brain. The commercially available CB2 receptor antibodies mayhave low affinity for the CB2B isoform in the brain and higher af-finity for the CB2A isoform expressed in the peripheral tissues(Onaivi et al., 2012).
The antinociception produced after exercise in the current studywas enhanced by FAAH and MGL inhibitors (MAFP and JZL184) inboth nociceptive tests. These enzymes have been found in key re-gions of the descending pathway of pain control, and in support ofour present results, some studies have demonstrated an increase ofantinociception as well as in the levels of AEA and 2-AG after sys-temic and central administration of FAAH and MGL inhibitors(Guindon and Hohmann, 2009). Furthermore, FAAH catalyzes thehydrolysis of PEA and OEA (Matias et al., 2007). We suggest thatMAFP and JZL184 potentiated and prolonged exercise-inducedantinociception by increasing the levels of endocannabinoids. Infact, AEA, 2-AG, PEA and OEA display antinociceptive effects(Pertwee, 2001; Suardiaz et al., 2007), which may be produced bytheir activation of cannabinoid receptors (in the case of AEA and 2-AG) and PPAR-a (in the case of PEA) (Pertwee, 2001) or by theactivation/desensitization of TRPV1 channels (in the case of AEA,OEA and PEA) (Maione et al., 2006). Indeed, these compoundsexhibit different affinities for cannabinoid receptors. Anandamidehas a preferential affinity for CB1 receptors and TRPV1 channels,while 2-AG has a lower affinity but higher efficacy at both CB1 andCB2 receptors (Martin et al., 1999; Mechoulam et al., 1995). PEA andOEA do not directly activate cannabinoid receptors and produceantinociception by several mechanisms (Pertwee, 2001; Suardiazet al., 2007).
In addition to antinociception, the majority of the effects pro-duced by endocannabinoids occur through retrograde signaling. Inthe CNS, endocannabinoids are released from postsynaptic neu-rons, and act retrogradely on presynaptic CB1 receptors and totransiently suppress neurotransmitter releasing. Furthermore,there is evidence suggesting that endocannabinoids signal in a non-retrograde or autocrine manner. Endocannabinoids can modulateneural function and synaptic transmission by engaging TRPV1 orsignal via astrocytes to indirectly modulate presynaptic or post-synaptic function (Hashimotodani et al., 2007). Retrograde

G. Galdino et al. / Neuropharmacology 77 (2014) 313e324322
endocannabinoid signaling mediates two forms of short-termsynaptic plasticity known as depolarization-induced suppressionof inhibition (DSI) and depolarization-induced suppression ofexcitation (DSE). In addition, endocannabinoids also mediate pre-synaptic forms of long-term depression at excitatory synapses(Hashimotodani et al., 2007). Thus, once released frompostsynapticneurons, endocannabinoids may suppress neurotransmitterrelease, including neurotransmitters involved in nociceptivetransmission (i.e., GABA and glutamate).
With respect to the mechanisms, the involvement of a voltagegated Ca2þ channel has been proposed for the suppression of GABArelease in the hippocampus and glutamate release at corticostriatalsynapses. In addition, Vaughan et al. (2000) have postulated that,like opioids, cannabinoids may induce antinociception by reducinginhibitory GABAergic influences on output neurons that projectfrom the periaqueductal gray and rostral ventromedial medulla.
Moreover, the regulation of glutamatergic neurotransmissionmay also contribute to the antinociceptive activity of cannabinoidagonists. In the dorsal horn of the spinal cord, these agents sup-press glutamate release from primary sensory afferents (Ohno-Shosaku et al., 2012).
In this context, the present study demonstrated that after aer-obic exercise, there is an increase in AEA, 2-AG, PEA and OEAplasma levels. Previous studies have also reported an increase inAEA plasma levels in humans after running and cycling (Feuereckeret al., 2011; Heyman et al., 2011; Sparling et al., 2003). Furthermore,other studies have demonstrated an increase of some of theseendocannabinoids within paw tissue, the dorsal horn and the PAGduring antinociception in rats (Guindon and Hohmann, 2009).Thus, our results strengthen the hypothesis that aerobic exerciseactivates the endocannabinoid system, which in turn may inducecentral and peripheral antinociception.
Some studies have also used inhibitors of anandamide cellularuptake, which occurs via an as-yet undefined molecular mecha-nism (Hohmann and Suplita, 2006). Reuptake of endocannabinoids,mainly of anandamide in the synaptic space, may be facilitated by aputative membrane transporter that has yet to be molecularlycloned (Guindon and Hohmann, 2009). Here, we demonstrated anincrease in the intensity and duration of the antinociceptive effectproduced by exercise in animals systemically and centrally pre-treated with VDM11, thus reinforcing the hypothesis of a role ofperipheral and central anandamide cellular uptake in this effect.
Studies also have demonstrated the participation of the endo-cannabinoid system in important responses during exercise such asmuscle vasodilatation, euphoria and bronchodilatation (Sparlinget al., 2003). The endocannabinoid system may also be involvedin movement control because CB1 and CB2 receptors are found inthe basal ganglia and cerebellum (Onaivi et al., 2012; Pertwee,2001). Cannabinoid receptors have also been found within thehypothalamus and may participate in the control of thermoregu-lation during exercise (Rawls and Benamar, 2011). Furthermore,another study suggested the involvement of this system in aerobiccapacity by demonstrating that pretreatment with Rimonabant(a CB1 receptor antagonist) reduces the running speed and distancein female high runner lines (Keeney et al., 2008). In addition, arecent study demonstrated that the wheel-running performance ofmice lacking CB1 receptors in GABAergic neurons (GABA-CB1
�/�
mice) was reduced compared to that of wildtype mice, suggestingthat running performance is mediated by CB1 receptors onGABAergic neurons (Dubreucq et al., 2013).
5. Conclusion
The present study indicates that a single session of moderateintensity aerobic exercise produces antinociception that is
mediated by endocannabinoids acting at CB1 and CB2 receptors atthe peripheral, medullar, and supra-medullar levels. This observa-tion may lead to future studies in athletes and patients withdifferent pain conditions.
Conflicts of interests
The authors declare no competing financial interests.
Acknowledgments
This work was supported by grants from the Conselho Nacionalde Desenvolvimento Científico e Tecnológico (CNPq), Brazil, and bythe NIH, National Institute of Drug Abuse (DA-009789 to VD).
References
Aguiar, D.C., Guimaraes, F.S., 2009. Blockade of NMDA receptors and nitric oxidesynthesis in the dorsolateral periaqueductal gray attenuates behavioral andcellular responses of rats exposed to a live predator. J. Neurosci. Res. 87, 2418e2429.
Alhouayek, M., Lambert, D.M., Delzenne, N.M., Cani, P.D., Muccioli, G.G., 2011.Increasing endogenous 2-arachidonoylglycerol levels counteracts colitis andrelated systemic inflammation. FASEB J. 25, 2711e2721.
Beijamini, V., Guimaraes, F.S., 2006. c-Fos expression increase in NADPH-diaphorasepositive neurons after exposure to a live cat. Behav. Brain Res. 170, 52e61.
Bement, M.K., Sluka, K.A., 2005. Low-intensity exercise reverses chronic musclepain in the rat in a naloxone-dependent manner. Arch. Phys. Med. Rehabil. 86,1736e1740.
Bisogno, T., Maurelli, S., Melck, D., De Petrocellis, L., Di Marzo, V., 1997. Biosynthesis,uptake, and degradation of anandamide and palmitoylethanolamide in leuko-cytes. J. Biol. Chem. 272, 3315e3323.
Calignano, A., La Rana, G., Giuffrida, A., Piomelli, D., 1998. Control of pain initiationby endogenous cannabinoids. Nature 394, 277e281.
Cannon, J.T., Prieto, G.J., Lee, A., Liebeskind, J.C., 1982. Evidence for opioid and non-opioid forms of stimulation-produced analgesia in the rat. Brain Res. 243, 315e321.
Casarotto, P.C., Terzian, A.L., Aguiar, D.C., Zangrossi, H., Guimaraes, F.S., Wotjak, C.T.,Moreira, F.A., 2012. Opposing roles for cannabinoid receptor type-1 (CB(1)) andtransient receptor potential vanilloid type-1 channel (TRPV1) on the modula-tion of panic-like responses in rats. Neuropsychopharmacology 37, 478e486.
Connell, K., Bolton, N., Olsen, D., Piomelli, D., Hohmann, A.G., 2006. Role of thebasolateral nucleus of the amygdala in endocannabinoid-mediated stress-induced analgesia. Neurosci. Lett. 397, 180e184.
Costa, B., Comelli, F., Bettoni, I., Colleoni, M., Giagnoni, G., 2008. The endogenousfatty acid amide, palmitoylethanolamide, has anti-allodynic and anti-hyperalgesic effects in a murine model of neuropathic pain: involvement ofCB(1), TRPV1 and PPARgamma receptors and neurotrophic factors. Pain 139,541e550.
D’amour, E.F., Smith, D., 1941. A method for determining loss of pain sensation.J. Pharmacol. Exp. Ther. 72, 74e79.
De Chiara, V., Errico, F., Musella, A., Rossi, S., Mataluni, G., Sacchetti, L.,Siracusano, A., Castelli, M., Cavasinni, F., Bernardi, G., Usiello, A., Centonze, D.,2010. Voluntary exercise and sucrose consumption enhance cannabinoid CB1receptor sensitivity in the striatum. Neuropsychopharmacology 35, 374e387.
Di Marzo, V., Bifulco, M., De Petrocellis, L., 2004. The endocannabinoid system andits therapeutic exploitation. Nat. Rev. Drug Discov. 3, 771e784.
Dubreucq, S., Durand, A., Matias, I., Bénard, G., Richard, E., Soria-Gomez, E.,Glangetas, C., Groc, L., Wadleigh, A., Massa, F., Bartsch, D., Marsicano, G.,Georges, F., Chaouloff, F., 2013. Ventral tegmental area cannabinoid type-1 re-ceptors control voluntary exercise performance. Biol. Psychiatry 73, 895e903.
Ebrahimzadeh, M., Haghparast, A., 2011. Analgesic effects of cannabinoid receptoragonist WIN55,212-2 in the nucleus cuneiformis in animal models of acute andinflammatory pain in rats. Brain Res. 1420, 19e28.
Fan, Y., Hooker, B.A., Garrison, T.R., El-Kouhen, O.F., Idler, K.B., Holley-Shanks, R.R.,Meyer, M.D., Yao, B.B., 2011. Pharmacological and molecular characterization ofa dorsal root ganglion cell line expressing cannabinoid CB(1) and CB(2) re-ceptors. Eur. J. Pharmacol. 659, 161e168.
Farquhar-Smith, W.P., Egertova, M., Bradbury, E.J., McMahon, S.B., Rice, A.S.,Elphick, M.R., 2000. Cannabinoid CB(1) receptor expression in rat spinal cord.Mol. Cell. Neurosci. 15, 510e521.
Feuerecker, M., Hauer, D., Toth, R., Demetz, F., Holzl, J., Thiel, M., Kaufmann, I.,Schelling, G., Chouker, A., 2011. Effects of exercise stress on the endocannabi-noid system in humans under field conditions. Eur. J. Appl. Physiol. 112, 2777e2781.
Fu, J., Gaetani, S., Oveisi, F., Lo Verme, J., Serrano, A., Rodriguez De Fonseca, F.,Rosengarth, A., Luecke, H., Di Giacomo, B., Tarzia, G., Piomelli, D., 2003. Oley-lethanolamide regulates feeding and body weight through activation of thenuclear receptor PPAR-alpha. Nature 425, 90e93.

G. Galdino et al. / Neuropharmacology 77 (2014) 313e324 323
Galdino, G.S., Cortes, S.F., Duarte, I.D., Perez, A.C., 2010. Involvement of the nitricoxide/(C)GMP/K(ATP) pathway in antinociception induced by exercise in rats.Life Sci. 86, 505e509.
Galiegue, S., Mary, S., Marchand, J., Dussossoy, D., Carriere, D., Carayon, P.,Bouaboula, M., Shire, D., Le Fur, G., Casellas, P., 1995. Expression of central andperipheral cannabinoid receptors in human immune tissues and leukocytesubpopulations. Eur. J. Biochem. 232, 54e61.
Gomes da Silva, S., Araujo, B.H., Cossa, A.C., Scorza, F.A., Cavalheiro, E.A., Naffah-Mazzacoratti Mda, G., Arida, R.M., 2010. Physical exercise in adolescencechanges CB1 cannabinoid receptor expression in the rat brain. Neurochem. Int.57, 492e496.
Guindon, J., Guijarro, A., Piomelli, D., Hohmann, A.G., 2011. Peripheral anti-nociceptive effects of inhibitors of monoacylglycerol lipase in a rat model ofinflammatory pain. Br. J. Pharmacol. 163, 1464e1478.
Guindon, J., Hohmann, A.G., 2009. The endocannabinoid system and pain. CNSNeurol. Disord. Drug Targets 8, 403e421.
Haller, V.L., Cichewicz, D.L., Welch, S.P., 2006. Non-cannabinoid CB1, non-cannabinoid CB2 antinociceptive effects of several novel compounds in thePPQ stretch test in mice. Eur. J. Pharmacol. 546, 60e68.
Hama, A., Sagen, J., 2007. Antinociceptive effect of cannabinoid agonist WIN 55,212-2 in rats with a spinal cord injury. Exp. Neurol. 204, 454e457.
Hashimotodani, Y., Ohno-Shosaku, T., Kano, M., 2007. Endocannabinoids and syn-aptic function in the CNS. Neuroscientist 13, 127e137.
Hayakawa, K., Mishima, K., Hazekawa, M., Sano, K., Irie, K., Orito, K., Egawa, T.,Kitamura, Y., Uchida, N., Nishimura, R., Egashira, N., Iwasaki, K., Fujiwara, M.,2008. Cannabidiol potentiates pharmacological effects of Delta(9)-tetrahydrocannabinol via CB(1) receptor-dependent mechanism. Brain Res.1188, 157e164.
Herkenham, M., Lynn, A.B., Johnson, M.R., Melvin, L.S., de Costa, B.R., Rice, K.C., 1991.Characterization and localization of cannabinoid receptors in rat brain: aquantitative in vitro autoradiographic study. J. Neurosci. 11, 563e583.
Heyman, E., Gamelin, F.X., Goekint, M., Piscitelli, F., Roelands, B., Leclair, E., DiMarzo, V., Meeusen, R., 2011. Intense exercise increases circulating endo-cannabinoid and BDNF levels in humans e possible implications for reward anddepression. Psychoneuroendocrinology 37, 844e851.
Hill, M.N., Titterness, A.K., Morrish, A.C., Carrier, E.J., Lee, T.T., Gil-Mohapel, J.,Gorzalka, B.B., Hillard, C.J., Christie, B.R., 2010. Endogenous cannabinoidsignaling is required for voluntary exercise-induced enhancement ofprogenitor cell proliferation in the hippocampus. Hippocampus 20, 513e523.
Hohmann, A.G., Herkenham, M., 1999. Localization of central cannabinoid CB1 re-ceptor messenger RNA in neuronal subpopulations of rat dorsal root ganglia: adouble-label in situ hybridization study. Neuroscience 90, 923e931.
Hohmann, A.G., Suplita 2nd, R.L., 2006. Endocannabinoid mechanisms of painmodulation. AAPS J. 8, E693eE708.
Hsieh, G.C., Pai, M., Chandran, P., Hooker, B.A., Zhu, C.Z., Salyers, A.K., Wensink, E.J.,Zhan, C., Carroll, W.A., Dart, M.J., Yao, B.B., Honore, P., Meyer, M.D., 2011. Centraland peripheral sites of action for CB receptor mediated analgesic activity inchronic inflammatory and neuropathic pain models in rats. Br. J. Pharmacol.162, 428e440.
Ji, L.L., Fu, R., Mitchell, E.W., 1992. Glutathione and antioxidant enzymes in skeletalmuscle: effects of fiber type and exercise intensity. J. Appl. Physiol. 73, 1854e1859.
Johanek, L.M., Simone, D.A., 2004. Activation of peripheral cannabinoid receptorsattenuates cutaneous hyperalgesia produced by a heat injury. Pain 109, 432e442.
Keeney, B.K., Raichlen, D.A., Meek, T.H., Wijeratne, R.S., Middleton, K.M.,Gerdeman, G.L., Garland Jr., T., 2008. Differential response to a selectivecannabinoid receptor antagonist (SR141716: rimonabant) in female mice fromlines selectively bred for high voluntary wheel-running behaviour. Behav.Pharmacol. 19, 812e820.
Kemppainen, P., Paalasmaa, P., Pertovaara, A., Alila, A., Johansson, G., 1990. Dexa-methasone attenuates exercise-induced dental analgesia in man. Brain Res. 519,329e332.
Kemppainen, P., Pertovaara, A., Huopaniemi, T., Johansson, G., 1986. Elevation ofdental pain threshold induced in man by physical exercise is not reversed bycyproheptadine-mediated suppression of growth hormone release. Neurosci.Lett. 70, 388e392.
Koltyn, K.F., 2000. Analgesia following exercise: a review. Sports Med. 29, 85e98.Koltyn, K.F., 2002. Exercise-induced hypoalgesia and intensity of exercise. Sports
Med. 32, 477e487.Liu, C., Walker, J.M., 2006. Effects of a cannabinoid agonist on spinal nociceptive
neurons in a rodent model of neuropathic pain. J. Neurophysiol. 96, 2984e2994.Lowry, O.H., Rosebrough, N.J., Farr, A.L., Randall, R.J., 1951. Protein measurement
with the Folin phenol reagent. J. Biol. Chem. 193, 265e275.Mailleux, P., Vanderhaeghen, J.J., 1992. Distribution of neuronal cannabinoid re-
ceptor in the adult rat brain: a comparative receptor binding radioautographyand in situ hybridization histochemistry. Neuroscience 48, 655e668.
Maione, S., Bisogno, T., de Novellis, V., Palazzo, E., Cristino, L., Valenti, M.,Petrosino, S., Guglielmotti, V., Rossi, F., Di Marzo, V., 2006. Elevation of endo-cannabinoid levels in the ventrolateral periaqueductal grey through inhibitionof fatty acid amide hydrolase affects descending nociceptive pathways via bothcannabinoid receptor type 1 and transient receptor potential vanilloid type-1receptors. J. Pharmacol. Exp. Ther. 316, 969e982.
Malan Jr., T.P., Ibrahim, M.M., Deng, H., Liu, Q., Mata, H.P., Vanderah, T., Porreca, F.,Makriyannis, A., 2001. CB2 cannabinoid receptor-mediated peripheral anti-nociception. Pain 93, 239e245.
Martin, B.R., Mechoulam, R., Razdan, R.K., 1999. Discovery and characterization ofendogenous cannabinoids. Life Sci. 65, 573e595.
Martin, W.J., Patrick, S.L., Coffin, P.O., Tsou, K., Walker, J.M., 1995. An examination ofthe central sites of action of cannabinoid-induced antinociception in the rat.Life Sci. 56, 2103e2109.
Matias, I., Gonthier, M.P., Petrosino, S., Docimo, L., Capasso, R., Hoareau, L.,Monteleone, P., Roche, R., Izzo, A.A., Di Marzo, V., 2007. Role and regulation ofacylethanolamides in energy balance: focus on adipocytes and beta-cells. Br. J.Pharmacol. 152, 676e690.
Mayer, D.J., 1984. Analgesia produced by electrical stimulation of the brain. Prog.Neuropsychopharmacol. Biol. Psychiatry 8, 557e564.
Mechoulam, R., Ben-Shabat, S., Hanus, L., Ligumsky, M., Kaminski, N.E., Schatz, A.R.,Gopher, A., Almog, S., Martin, B.R., Compton, D.R., et al., 1995. Identification ofan endogenous 2-monoglyceride, present in canine gut, that binds to canna-binoid receptors. Biochem. Pharmacol. 50, 83e90.
Mense, S., 1993. Peripheral mechanisms of muscle nociception in local muscle pain.J. Musculoskelet. Pain 1, 133e170.
Mestre, C., Pelissier, T., Fialip, J., Wilcox, G., Eschalier, A., 1994. A method to performdirect transcutaneous intrathecal injection in rats. J. Pharmacol. Toxicol.Methods 32, 197e200.
Mu, J., Zhuang, S.Y., Kirby, M.T., Hampson, R.E., Deadwyler, S.A., 1999. Cannabinoidreceptors differentially modulate potassium A and D currents in hippocampalneurons in culture. J. Pharmacol. Exp. Ther. 291, 893e902.
Munro, S., Thomas, K.L., Abu-Shaar, M., 1993. Molecular characterization of a pe-ripheral receptor for cannabinoids. Nature 365, 61e65.
Naidu, P.S., Booker, L., Cravatt, B.F., Lichtman, A.H., 2009. Synergy between enzymeinhibitors of fatty acid amide hydrolase and cyclooxygenase in visceral noci-ception. J. Pharmacol. Exp. Ther. 329, 48e56.
Ohno-Shosaku, T., Tanimura, A., Hashimotodani, Y., Kano, M., 2012. Endocannabi-noids and retrograde modulation of synaptic transmission. Neuroscientist 18,119e132.
Onaivi, E.S., Carpio, O., Ishiguro, H., Schanz, N., Uhl, G.R., Benno, R., 2008. Behavioraleffects of CB2 cannabinoid receptor activation and its influence on food andalcohol consumption. Ann. N. Y. Acad. Sci. 1139, 426e433.
Onaivi, E.S., Ishiguro, H., Gu, S., Liu, Q.R., 2012. CNS effects of CB2 cannabinoid re-ceptors: beyond neuro-immuno-cannabinoid activity. J. Psychopharmacol. 26,92e103.
Paxinos, G., Watson, C., 1986. The Rat Brain in Stereotaxic Coordinates, second ed.Academic Press, New York.
Pertwee, R.G., 1997. Pharmacology of cannabinoid CB1 and CB2 receptors. Phar-macol. Ther. 74, 129e180.
Pertwee, R.G., 2001. Cannabinoid receptors and pain. Prog. Neurobiol. 63, 569e611.Pertwee, R.G., Ross, R.A., 2002. Cannabinoid receptors and their ligands. Prosta-
glandins Leukot. Essent. Fatty Acids 66, 101e121.Petrosino, S., Iuvone, T., Di Marzo, V., 2010. N-palmitoyl-ethanolamine: biochem-
istry and new therapeutic opportunities. Biochimie 92, 724e727.Piomelli, D., Lo Verme, J., Fu, J., Astarita, G., La Rana, G., Russo, R., Calignano, A.,
2005. The nuclear receptor peroxisome proliferator-activated receptor-alphamediates the anti-inflammatory actions of palmitoylethanolamide. Mol. Phar-macol. 67, 15e19.
Quartilho, A., Mata, H.P., Ibrahim, M.M., Vanderah, T.W., Porreca, F., Makriyannis, A.,Malan Jr., T.P., 2003. Inhibition of inflammatory hyperalgesia by activation ofperipheral CB2 cannabinoid receptors. Anesthesiology 99, 955e960.
Randall, L.O., Selitto, J.J., 1957. A method for measurement of analgesic activity oninflamed tissue. Arch. Int. Pharmacodyn. Ther. 111, 409e419.
Rawls, S.M., Benamar, K., 2011. Effects of opioids, cannabinoids, and vanilloids onbody temperature. Front. Biosci. 3, 822e845.
Re, G., Barbero, R., Miolo, A., Di Marzo, V., 2007. Palmitoylethanolamide, endo-cannabinoids and related cannabimimetic compounds in protection againsttissue inflammation and pain: potential use in companion animals. Vet. J. 173,21e30.
Reynolds, D.V., 1969. Surgery in the rat during electrical analgesia induced by focalbrain stimulation. Science 164, 444e445.
Richardson, J.D., Kilo, S., Hargreaves, K.M., 1998. Cannabinoids reduce hyperalgesiaand inflammation via interaction with peripheral CB1 receptors. Pain 75, 111e119.
Romero-Sandoval, A., Eisenach, J.C., 2007. Spinal cannabinoid receptor type 2activation reduces hypersensitivity and spinal cord glial activation after pawincision. Anesthesiology 106, 787e794.
Salio, C., Doly, S., Fischer, J., Franzoni, M.F., Conrath, M., 2002. Neuronal and astro-cytic localization of the cannabinoid receptor-1 in the dorsal horn of the ratspinal cord. Neurosci. Lett. 329, 13e16.
Samineni, V.K., Premkumar, L.S., Faingold, C.L., 2011. Post-ictal analgesia ingenetically epilepsy-prone rats is induced by audiogenic seizures and in-volves cannabinoid receptors in the periaqueductal gray. Brain Res. 1389,177e182.
Soares, D.D., Lima, N.R., Coimbra, C.C., Marubayashi, U., 2003. Evidence that tryp-tophan reduces mechanical efficiency and running performance in rats. Phar-macol. Biochem. Behav. 74, 357e362.
Sparling, P.B., Giuffrida, A., Piomelli, D., Rosskopf, L., Dietrich, A., 2003. Exerciseactivates the endocannabinoid system. Neuroreport 14, 2209e2211.

G. Galdino et al. / Neuropharmacology 77 (2014) 313e324324
Suardiaz, M., Estivill-Torrus, G., Goicoechea, C., Bilbao, A., Rodriguez de Fonseca, F.,2007. Analgesic properties of oleoylethanolamide (OEA) in visceral and in-flammatory pain. Pain 133, 99e110.
Svizenska, I., Dubovy, P., Sulcova, A., 2008. Cannabinoid receptors 1 and 2 (CB1 andCB2), their distribution, ligands and functional involvement in nervous systemstructures e a short review. Pharmacol. Biochem. Behav. 90, 501e511.
Tsou, K., Brown, S., Sanudo-Pena, M.C., Mackie, K., Walker, J.M., 1998. Immunohis-tochemical distribution of cannabinoid CB1 receptors in the rat central nervoussystem. Neuroscience 83, 393e411.
Vaughan, C.W., Connor, M., Bagley, E.E., Christie, M.J., 2000. Actions of cannabinoidson membrane properties and synaptic transmission in rat periaqueductal grayneurons in vitro. Mol. Pharmacol. 57, 288e295.
Wang, Q., Peng, Y., Chen, S., Gou, X., Hu, B., Du, J., Lu, Y., Xiong, L., 2009. Pre-treatment with electroacupuncture induces rapid tolerance to focal cerebralischemia through regulation of endocannabinoid system. Stroke 40, 2157e2164.
Xi, Z.X., Peng, X.Q., Li, X., Song, R., Zhang, H.Y., Liu, Q.R., Yang, H.J., Bi, G.H., Li, J.,Gardner, E.L., 2011. Brain cannabinoid CB receptors modulate cocaine’s actionsin mice. Nat. Neurosci. 14, 1160e1166.
Zhang, J., Hoffert, C., Vu, H.K., Groblewski, T., Ahmad, S., O’Donnell, D., 2003. In-duction of CB2 receptor expression in the rat spinal cord of neuropathic but notinflammatory chronic pain models. Eur. J. Neurosci. 17, 2750e2754.
Zimmermann, M., 1983. Ethical guidelines for investigations of experimental pain inconscious animals. Pain 16, 109e110.
Related Documents