This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
The distribution and chemical speciation of dissolved and particulatephosphorus in the Bering Sea and the Chukchi–Beaufort Seas
Peng Lin a, Laodong Guo b,n, Min Chen a,c, Jinlu Tong a, Feng Lin a
a College of Ocean and Earth Sciences, Xiamen University, Xiamen 361005, Chinab School of Freshwater Sciences, University of Wisconsin-Milwaukee, Milwaukee, WI 53204, USAc State Key Laboratory of Marine Environmental Science, Xiamen University, Xiamen 361005, China
a r t i c l e i n f o
Available online 16 July 2012
Keywords:
Phosphorus
Nutrient cycles
Suspended particulate matter
Chemical speciation
Bering Sea
Western Arctic Ocean
a b s t r a c t
Water samples were collected from the Bering Sea and the Chukchi–Beaufort Seas during the 4th
Chinese Arctic Research Expedition in summer 2010 to examine the abundance, spatial distribution and
phase partitioning of phosphorus (P) between dissolved and particulate, inorganic and organic phases.
Measurements of P species included dissolved inorganic P (DIP), dissolved organic P (DOP) and five
particulate P species operationally defined by sequential extraction techniques. Inorganic P was
generally the predominant form of both dissolved and particulate P, comprising on average up to
91713% of the dissolved phase and 72723% of the particulate phase in the study area. Concentrations
of DIP in the water column of the Chukchi Sea and Bering Sea were considerably higher than those in
the Beaufort Sea (43 mM vs. �1 mM), showing a strong influence of North Pacific inflow waters. The
percentage of DOP in the dissolved P pool was generally higher in surface waters than deeper waters,
and higher in the Chukchi Sea than in the Bering Sea, consistent with their difference in primary
productivity and reflecting sources of DOP from the surface waters. Within the particulate P pool, the
labile P and ferric bound P were the two predominant species, especially in the upper water column
(o1000 m), while authigenic or CaCO3 bound P and detrital P were the two least abundant particulate
P species. A positive correlation between labile particulate P and particulate organic carbon (POC) or
particulate nitrogen (PN) was observed, where labile P concentrations decreased with increasing DIP
concentrations and decreasing biological activity in the upper water column, indicating the contribu-
tion of labile particulate P from biological production. Labile P was an active component in regeneration
of particulate P in the water column. The authigenic P was relatively abundant in deeper waters,
especially in the Bering Sea, suggesting an active transformation between labile-P and authigenic P and
a possible sink for P as authigenic particulate P in the deeper water column. In contrast to labile P,
authigenic P, and organic P, the distributions of ferric-bound P and detrital P were mostly regulated by
physicochemical processes, such as lateral transport and sediment resuspension. Therefore, different
particulate P species could play a distinct role in biogeochemical cycling of P in the water column and
understanding the detailed chemical and phase speciation of P should provide a better understanding
of the dynamic cycling of P in the marine environment.
& 2012 Elsevier Ltd. All rights reserved.
1. Introduction
Carbon (C), nitrogen (N) and phosphorus (P) are major bioactiveelements and play an essential role in ecosystem functions andbiogeochemical processes in marine environments (Benitez-Nelson,2000). In some ocean ecosystems, P could become a limitingnutrient for biological productivity (e.g., Wu et al., 2000; Karlet al., 2001; Carmack et al., 2004), by regulating biogeochemicalcycling of carbon, nutrients, and other bioactive trace elements,
such as iron, in the water column (Takata et al., 2004; Boye et al.,2010). However, most studies focus on the biogeochemicalcycling of carbon and nitrogen species, including those conductedin the Arctic Ocean (e.g., Grebmeier and Harvey, 2005; Grebmeieret al., 2006; Mathis et al., 2007; McGuire et al., 2009; Cai et al.,2010) and those conducted during the Chinese Arctic ResearchExpedition (CHINARE). To the best of our knowledge, there arevery few studies on the biogeochemical cycling of P in thewestern Arctic Ocean.
In aquatic environments, the biogeochemical forms ofP include dissolved and particulate, and inorganic and organicP (Yoshimura et al., 2007; Cai and Guo, 2009). Operationally, totaldissolved P (TDP, o0.45 mm) can be divided into dissolved
Contents lists available at SciVerse ScienceDirect
journal homepage: www.elsevier.com/locate/dsr2
Deep-Sea Research II
0967-0645/$ - see front matter & 2012 Elsevier Ltd. All rights reserved.
http://dx.doi.org/10.1016/j.dsr2.2012.07.005
n Corresponding author.
E-mail address: [email protected] (L. Guo).
Deep-Sea Research II 81-84 (2012) 79–94

Author's personal copy
inorganic P (DIP) and dissolved organic P (DOP), while totalparticulate P (TPP, 40.45 mm) can be fractionated into particulateinorganic P (PIP) and particulate organic P (POP). The primaryinput of P to the ocean comes from riverine dissolved P (Froelichet al., 1982; Delaney, 1998; Faul et al., 2005). Consequently, TPP inoceanic waters is mostly produced through biological processesand recycled within the water column. In the Arctic Ocean,potential sources of particulate P may also come from terrestrialinputs, including riverine export and coastal erosion (Guo et al.,2004; Ping et al., 2011), atmospheric deposition, and sedimentsassociated with sea ice rafting (Eicken et al., 2005; Chen et al.,2012). Knowledge of the transport and transformation processesof P between dissolved and particulate phases is thus importantfor our understanding of the dynamic cycling of P in the ArcticOcean and the impact of environmental change on Arcticecosystems.
Due to its high reactivity and easy uptake by marine organ-isms, DIP and its bioavailability in marine ecosystems have beenextensively studied (e.g., Perry and Eppley, 1981; Harrison andHarris, 1986; Benitez-Nelson, 2000; Zhang and Huang, 2007,2011). Over the past decades, biogeochemical cycling of DOPhas received increasing attention since DOP could also be apotential P source for biological communities (Karl and Craven,1980; Bjorkman and Karl, 1994, 2003; Cavender-Bares et al.,2001; Yoshimura et al., 2007; Huang and Zhang, 2010, 2011;Ruttenberg and Sulak, 2011). Compared with the dissolved P pool,TPP has yet to be quantified and characterized in the watercolumn (Paytan et al., 2003; Suzumura and Ingall, 2004; Zhanget al., 2010). Studies examining both dissolved and particulate,and inorganic and organic phosphorus are even scarcer, especiallyin the Arctic marine environment. The development of TPPanalytical methods, such as the sequential extraction (SEDEX)technique (Ruttenberg, 1990, 1992), has provided a possible wayto evaluate the role of different particulate P phases in thebiogeochemical cycling of P in marine environments.
The Arctic Ocean plays an important role in ocean circulationand marine biogeochemical cycles (Smith et al., 1989; Walsh,1989). Ongoing climate and environmental changes observed inthe Arctic have stimulated increasing research on the impact andbiogeochemical consequences of these changes in Arctic ecosys-tems (e.g., Chen et al., 2002, 2003a; Grebmeier and Harvey, 2005and accompanying papers; McGuire et al., 2009; Cai et al., 2010).The Bering Sea is the only pathway for water exchange betweenthe North Pacific and the Arctic Ocean (Kawaguchi, 2001). Watermasses with high nutrients entering the Arctic through the BeringStrait support active biological activity on the Arctic shelves(Grebmeier et al., 2006). However, the western Arctic Ocean islikely a P-limited marine ecosystem (Carmack et al., 2004).Variations in the distribution and partitioning of inorganic andorganic P species among the Bering Sea, the Chukchi Sea and theBeaufort Sea are poorly understood.
We present here the spatial variations in the concentrationsand partitioning of various forms of P, including DIP, DOP, PIP, andPOP, as well as different particulate P species in the Bering Seaand the Chukchi–Beaufort Seas. We also examined the nature andcharacteristics of various operationally defined particulate Pspecies and their transformations in the water column, as wellas their role in the biogeochemical cycling of P in the westernArctic Ocean.
2. Materials and methods
2.1. Study area
Water samples were collected from the western Arctic Ocean,including the Bering Sea, the Chukchi Sea and the Beaufort Sea,during the 4th Chinese Arctic Research Expedition (CHINARE)from July to September 2010 (Fig. 1). A total of 12 stations alongtwo transects comprising 7 stations from the Bering Sea and
Fig. 1. Sampling locations from the Bering Sea and the Chukchi–Beaufort Seas during summer 2010.
P. Lin et al. / Deep-Sea Research II 81-84 (2012) 79–9480

Author's personal copy
5 stations from the Chukchi–Beaufort Seas, were occupied andsampled for measurements of dissolved and particulate P species.For stations on the Bering Sea transect, B05 and B09 were locatedin the Aleutian Basin with a water depth 43000 m, B11 in thecontinental slope region, and four stations (B15, NB01, NB03and NB05) on the Bering Shelf with a water depth o110 m(Fig. 1). For stations on the Chukchi–Beaufort Seas transect,four stations (C07, C08, Co9 and S22) located on the ChukchiShelf with a water depth o150 m and one station (S24) on theBeaufort Slope (Fig. 1). Water masses in the Aleutian Basinwere influenced by North Pacific inflow waters with high tem-peratures, salinity, and nutrients, while water masses on theBering Shelf were affected by the Bering Shelf Water (BSW) andthe Alaska Coastal Current (ACC) with relatively lower salinityand nutrients. Both BSW and ACC play an important role inaffecting the nutrient budget and distribution in the ChukchiShelves. Compared to the Bering Sea, the Chukchi–Beaufort Seascould receive terrestrial inputs and sea ice melting waters,especially in the Beaufort Slope regions (Aagaard and Carmack,1989; Chen et al., 2003a).
2.2. Sampling
Seawater samples were collected by a CTD-Rosette systemequipped with temperature, salinity, and fluorescence sensors.Detailed information about the sampling locations and theirhydrographic data, including water depth, sampling depth, tem-perature, and salinity, is given in Table 1. Immediately aftercollection, water samples were filtered through 0.45 mm Nucle-pore filters (Whatman, 47 mm). The filtrate samples for DIP andDOP were collected in acid-cleaned HDPE bottles and stored in afreezer until analysis in the laboratory. For the determination ofchemical speciation of particulate P, predetermined amounts ofseawater samples were filtered through 0.45 mm Nuclepore filtersonboard ship. Samples for particulate organic carbon (POC) andparticulate nitrogen (PN) measurements were filtered on pre-combusted GF/F filters (Whatman, 47 mm).
2.3. Measurements of dissolved inorganic and organic P species
TDP was measured by an autoclave-assisted acid persulfatemethod (Koroleff, 1983) with some modifications (Cai and Guo,2009; Lin et al., 2012a). In brief, 10 mL of seawater sample and1 mL of acidified K2S2O8 solution (50 g/L, pH¼1) were added to aTeflon vial with a Teflon screw cap. The mixed solution wasdigested for 4 h at 140 1C in an oven. After digestion, TDP wasmeasured by the classical phosphomolybdenum blue methodusing a Cary 300 UV–Visible spectrophotometer and a 5 cmquartz cuvette (Parsons et al., 1984). DIP was measured withthe same method as TDP except with no digestion. Concentrationsof DOP were calculated from the difference between TDP and DIP.Standard solutions were treated the same as samples duringsample processing and analysis to ensure data quality.The detection limit was 8–10 nM for DIP and DOP based onreplicate blank measurements, with a precision better than 2% forboth DIP and TDP (Cai and Guo, 2009; Lin et al., 2012a).
2.4. Measurements of particulate phosphorus
The SEDEX technique has been widely used in the measurementof sedimentary P species (e.g., Slomp et al., 1998; Latimer andFilippelli, 2001; Tamburini et al., 2002; van der Zee et al., 2002;Zhang et al., 2004, 2010) and suspended particulate P species inaquatic environments (e.g., Sutula et al., 2004; Lin et al., submittedfor publication). Using the SEDEX technique (Ruttenberg, 1990,1992; Zhang et al., 2010), total particulate phosphorus (TPP) was
chemically fractionated into five operationally defined particulate Pspecies: (1) loosely adsorbed (exchangeable) P; (2) ferric-bound P;(3) authigenic carbonate fluorapatiteþbiogenic apatiteþCaCO3-associated P; (4) detrital apatite P; and (5) refractory organic P.These five particulate P species are named herein as Labile-P, Fe-P,CFA-P, Detr-P and Org-P, respectively.
Briefly, sequential extraction of particulate P on filters wasperformed in 25 mL centrifuge tubes based on proceduresdescribed in Zhang et al. (2010). After each extraction, the samplewas centrifuged and the supernatant solution containing theextracted P was colorimetrically measured for DIP with a mod-ified chromogenic agent. Labile-P, CFA-P and TDP were measuredwith methods described in Section 2.3 along with measurementsof organic P. The detection limit was 0.1 nM for particulate Pmeasurements with a high concentration factor and a longpathlength quartz cuvette (5 cm).
TPP concentration was calculated from the sum of five differ-ent particulate P species determined by SEDEX. The PIP concen-tration was the sum of the labile inorganic P fraction (Labile-IP),Fe-P, inorganic fraction of CFA-P (CFAP-IP) and Detr-P, and POPequaled the sum of the labile organic P fraction (Labile-OP),organic fraction of CFA-P (CFAP-OP) and Org-P.
2.5. Measurements of particulate organic carbon and nitrogen
GF/F filter samples were treated with acid fumes overnight formeasurements of POC and PN on an elemental analyzer (Guo andSantschi, 1997; Chen et al., 2006). POC was used along with POP,as well as total reactive particulate P to determine the particulateorganic C:P ratio and the particulate organic C:reactive-P ratio(see detail below). Note that the filter pore sizes for POC and POPwere slightly different (0.7 vs. 0.45 mm) and the C:P ratios arethus operational.
Concentrations of suspended particulate matter (SPM) werecalculated by the weight difference between filter samples andblank filters. Aliquots of water samples were filtered through pre-weighed 0.45 mm Nuclepore polycarbonate membranes (Whatman)and rinsed with nanopure water 3–4 times to remove salts. Filtersamples were weighed after drying at 60 1C until achieving aconstant weight.
3. Results
3.1. Hydrographic features
Surface water temperatures varied from �1.30 to 10.47 1C,showing a general increase from the Chukchi–Beaufort Seas tothe Bering Sea (Table 1). In the Bering Sea, surface watertemperatures ranged from 4.49 to 10.47 1C with an average of7.64 1C, and it ranged from �1.30 to 6.81 1C, with an average of2.93 1C in the Chukchi–Beaufort transect. Surface water salinitywas highly variable, ranging from 25.97 to 33.14 with an averageof 30.93 (Table 1). Low salinity and relatively high temperatureswere observed near the Yukon River delta (Fig. 2a and b),showing the influence of freshwater discharge. High salinitieswere observed at stations in the Aleutian Basin and the low oneson the Bering Shelf. In the Chukchi–Beaufort Seas, salinitywas generally low except near the northwestern edge of theAlaska coastline (Fig. 3b), likely resulting from the sea-icemelting.
Distributions of temperature and salinity showed differentcharacteristics between the Beaufort Slope and the Aleutian Basin.High temperature in deeper waters was observed in the AleutianBasin and at shallow stations in the Chukchi–Beaufort Seas.A strong halocline occurred on the Beaufort Slope but not in the
P. Lin et al. / Deep-Sea Research II 81-84 (2012) 79–94 81

Author's personal copyT
ab
le1
Sa
mp
lin
glo
cati
on
s,h
yd
rog
rap
hic
da
tain
clu
din
gw
ate
rd
ep
th,t
em
pe
ratu
re(T
),sa
lin
ity
an
dsu
spe
nd
ed
pa
rtic
ula
tem
att
er
(SP
M)
con
cen
tra
tio
ns,
an
dth
eco
nce
ntr
ati
on
so
fd
isso
lve
da
nd
pa
rtic
ula
teP
spe
cie
s,a
sw
ell
as
pa
rtic
ula
te
nit
rog
en
(PN
),p
art
icu
late
org
an
icca
rbo
n(P
OC
),p
art
icu
late
rea
ctiv
ep
ho
sph
oru
s(R
ea
-P),
pa
rtic
ula
teo
rga
nic
C:P
rati
os
(PO
C/P
OP
)a
nd
pa
rtic
ula
teo
rga
nic
C:r
ea
ctiv
eP
rati
os
(PO
C/R
ea
-P)
inth
ew
est
ern
Arc
tic
Oce
an
.
Sta
tio
nLo
cati
on
sD
ep
th
(m)
T(1
C)
Sa
lin
ity
SP
M
(mg
/L)
DIP
( mM
)
DO
P
(mM
)
Lab
ile
-P(n
M)
Fe-P
(nM
)
CFA
-P(n
M)
De
tr-P
(nM
)
Org
-P
(nM
)
PIP
(nM
)
PO
P
(nM
)
PN
(mM
)
PO
C
(mM
)
Re
a-P
(nM
)
PO
C/
Re
a-P
PO
C/
PO
P
IPO
PT
PIP
OP
TP
Be
rin
gS
ea
B0
55
515
60 3
900N
17
311
10 2
800E
07
.70
33
.00
–1
.73
B.D
.1
1.5
6.1
17
.60
.3B
.D.
0.2
0.2
B.D
.2
.61
1.7
8.9
0.6
13
.04
20
.61
47
34
0
20
7.5
03
2.9
9–
1.7
00
.01
17
.66
.42
4.0
0.1
0.1
0.4
0.5
B.D
.2
.01
7.7
8.9
0.7
03
.40
26
.61
28
38
2
50
3.1
13
3.1
2–
2.0
70
.08
7.7
3.0
10
.70
.10
.10
.10
.2B
.D.
2.4
7.9
5.5
0.4
52
.25
13
.41
68
41
0
75
2.3
23
3.2
0–
2.2
8B
.D.
0.6
2.7
3.3
0.2
0.6
B.D
.0
.60
.95
.92
.38
.50
.22
1.1
89
.91
19
13
8
10
01
.72
33
.22
–1
.95
B.D
.1
.82
.23
.90
.20
.3B
.D.
0.3
0.2
3.9
2.5
6.1
0.2
51
.26
8.4
15
12
08
12
01
.67
33
.26
–2
.39
0.2
7B
.D.
0.5
0.5
0.2
0.3
B.D
.0
.3B
.D.
12
.90
.51
3.5
0.1
40
.72
14
.05
25
4
30
03
.81
33
.94
–3
.24
B.D
.1
1.2
9.7
20
.90
.10
.6B
.D.
0.6
0.9
10
6.8
12
.81
16
.50
.11
0.6
51
28
.45
6
50
03
.57
34
.14
–3
.27
B.D
.1
.2B
.D.
1.2
0.5
3.0
B.D
.3
.0B
.D.
2.2
4.7
2.2
0.1
10
.69
6.9
10
03
09
10
00
2.7
93
4.3
7–
3.4
50
.23
0.6
0.6
1.2
0.5
0.9
0.1
1.0
0.5
2.2
2.4
2.9
0.1
00
.68
4.8
14
02
35
15
00
2.2
83
4.5
0–
3.4
1B
.D.
B.D
.0
.50
.5B
.D.
0.3
B.D
.0
.30
.22
.20
.52
.70
.07
0.4
73
.01
53
17
0
20
00
1.9
03
4.5
8–
3.4
1B
.D.
B.D
.0
.50
.50
.11
.8B
.D.
1.8
B.D
.2
.21
.92
.70
.11
0.6
64
.71
40
24
0
38
00
(38
21
)
1.6
03
4.6
7–
3.1
1B
.D.
B.D
.B
.D.
B.D
.B
.D.
2.1
0.3
2.4
0.9
2.7
3.0
3.0
0.1
20
.71
5.1
13
82
37
B0
95
911
30 2
100N
17
812
80 1
900E
08
.31
32
.64
0.9
70
.61
B.D
.4
5.9
B.D
.4
5.9
32
.51
.00
.51
.61
.13
.28
0.5
3.7
1.3
57
.79
83
.19
42
10
5
20
7.5
13
2.6
22
.61
1.1
30
.10
36
.16
.94
3.1
19
.10
.1B
.D.
0.1
0.5
2.9
55
.89
.91
.38
7.6
26
5.2
11
77
73
28
4.3
23
2.7
20
.95
1.8
00
.23
45
.23
.24
8.4
50
.7B
.D.
1.0
1.0
0.7
2.2
96
.56
.51
.16
6.5
61
02
.36
41
01
7
50
2.0
03
2.9
30
.74
2.2
8B
.D.
12
.01
.81
3.8
0.3
B.D
.B
.D.
B.D
.0
.51
.71
2.8
3.5
0.4
53
.19
15
.92
01
91
4
75
2.4
83
3.0
60
.64
1.9
2B
.D.
18
.73
.82
2.5
0.9
0.1
B.D
.0
.11
.14
.12
0.8
7.9
0.8
34
.91
27
.61
78
61
8
10
02
.48
33
.12
0.6
32
.56
B.D
.5
.43
.18
.50
.70
.12
.42
.51
.42
.47
.67
.90
.25
1.9
31
4.1
13
72
45
15
02
.27
33
.20
–2
.35
B.D
.1
.90
.32
.27
.1B
.D.
B.D
.B
.D.
B.D
.3
.79
.04
.00
.22
1.2
61
3.0
97
31
7
30
02
.81
33
.44
–2
.72
B.D
.1
.4B
.D.
1.4
42
.9B
.D.
0.3
0.3
B.D
.0
.84
4.3
1.1
0.3
42
.26
45
.45
01
99
2
50
03
.75
33
.91
–3
.31
B.D
.0
.5B
.D.
0.5
14
.3B
.D.
0.6
0.6
B.D
.1
.01
4.8
1.6
0.1
30
.70
16
.44
34
26
10
00
2.9
73
4.3
21
.06
3.3
5B
.D.
0.5
0.1
0.5
57
.1B
.D.
B.D
.B
.D.
B.D
.2
.55
7.6
2.5
0.1
10
.77
60
.21
33
04
20
00
1.9
43
4.5
70
.88
3.1
4B
.D.
B.D
.B
.D.
B.D
.0
.96
.12
.88
.9B
.D.
6.6
7.1
9.4
0.1
00
.70
16
.54
37
5
34
21
(35
10
)
1.6
03
4.6
60
.64
3.2
4B
.D.
1.0
B.D
.1
.00
.92
.91
.84
.70
.46
.45
.28
.20
.13
1.2
31
3.0
95
15
0
B1
15
915
70 5
600N
17
914
80 5
900E
08
.26
32
.61
1.3
70
.95
B.D
.–
––
––
––
––
––
––
0.9
55
.80
––
–
20
5.3
63
2.7
01
.53
1.2
20
.03
98
.81
5.7
11
4.5
82
.4B
.D.
0.6
0.6
3.7
5.3
18
4.9
21
.71
.26
7.6
62
02
.93
83
53
30
2.4
03
2.8
51
.19
2.1
4B
.D.
47
.01
0.5
57
.51
0.7
0.6
0.1
0.7
1.6
2.1
60
.01
2.7
0.7
24
.01
71
.05
63
16
50
2.3
73
3.0
10
.56
2.0
1B
.D.
25
.74
.53
0.1
10
.7B
.D.
0.7
0.7
B.D
.1
.03
6.4
6.3
0.2
81
.86
42
.64
42
97
75
2.4
53
3.1
20
.47
2.1
1B
.D.
37
.16
.84
3.8
38
.61
.2B
.D.
1.2
1.2
1.8
78
.18
.60
.17
1.0
68
5.5
12
12
2
10
02
.31
33
.17
0.4
12
.52
0.2
73
.82
.76
.55
3.6
3.1
3.7
6.9
B.D
.0
.26
0.5
6.6
0.1
51
.17
67
.21
71
77
17
01
.94
33
.22
0.6
62
.46
0.2
2B
.D.
3.0
3.0
60
.7B
.D.
1.6
1.6
B.D
.2
.16
0.7
6.6
0.1
40
.94
67
.31
41
42
30
03
.78
33
.62
2.1
92
.81
B.D
.8
.03
.01
0.9
25
.00
.6B
.D.
0.6
B.D
.1
.43
3.6
4.4
0.1
40
.95
38
.02
52
16
50
03
.66
33
.96
0.8
33
.31
B.D
.B
.D.
0.5
0.5
35
.7B
.D.
0.3
0.3
B.D
.2
.13
5.7
2.9
0.2
02
.07
38
.65
47
23
10
00
2.9
13
4.3
30
.84
3.3
1B
.D.
B.D
.1
.01
.0B
.D.
0.3
B.D
.0
.3B
.D.
4.1
0.3
5.1
0.1
20
.73
5.4
13
51
43
20
00
1.8
93
4.5
8–
3.3
7B
.D.
––
––
––
––
––
–0
.13
0.7
7–
––
25
50
(26
25
)
1.6
73
4.6
30
.98
3.3
3B
.D.
––
––
––
––
––
–0
.09
0.6
9–
––
B1
56
1140 500N
-
17
614
00 2
100W
06
.86
30
.84
0.9
70
.38
0.2
56
7.8
7.9
75
.71
5.0
B.D
.3
.13
.1B
.D.
7.7
82
.81
8.7
––
10
1.5
––
10
6.8
83
0.8
21
.22
0.4
30
.22
17
.75
.02
2.7
13
.8B
.D.
1.9
1.9
B.D
.4
.63
1.5
11
.5–
–4
3.0
––
36
�1
.42
31
.81
0.9
51
.73
0.1
61
6.1
7.4
23
.52
5.6
3.8
B.D
.3
.80
.85
.44
6.3
12
.80
.59
4.1
35
8.3
71
32
3
50
�0
.65
32
.04
0.9
11
.80
0.3
01
0.2
1.0
11
.21
1.1
5.4
B.D
.5
.40
.41
3.7
27
.11
4.7
0.5
43
.81
41
.49
22
60
75
1.6
13
2.6
22
.70
2.1
7B
.D.
23
.4B
.D.
23
.42
2.1
17
.6B
.D.
17
.62
.09
.86
5.1
9.8
0.5
84
.99
72
.96
85
10
10
8(1
10
)1
.62
32
.62
3.7
52
.59
0.1
32
5.9
0.3
26
.21
6.4
30
.2B
.D.
30
.21
0.2
14
.48
2.6
14
.80
.92
8.0
98
7.2
93
54
7
NB
01
6111
40 1
200N
-
17
5140 1
600W
07
.45
30
.74
0.8
10
.48
0.1
01
1.2
3.8
15
.02
5.5
1.9
B.D
.1
.9B
.D.
2.9
38
.66
.7–
–4
5.3
––
10
7.4
53
0.7
32
.28
0.5
00
.45
2.9
B.D
.2
.92
1.4
1.1
B.D
.1
.1B
.D.
2.0
25
.42
.0–
–2
7.4
––
20
3.8
13
0.7
60
.69
0.6
30
.21
31
.25
.73
6.9
47
.44
.60
.55
.10
.42
.08
3.6
8.2
––
91
.3–
–
30
�1
.23
31
.47
0.9
90
.90
B.D
.5
2.7
6.1
58
.85
4.7
4.2
B.D
.4
.21
.21
.01
12
.87
.1–
–1
18
.7–
–
46
�1
.33
31
.64
1.9
72
.06
B.D
.1
00
.03
3.7
13
3.7
37
.27
.9B
.D.
7.9
3.3
24
.91
48
.35
8.6
1.7
71
1.8
22
03
.75
82
02
85
(92
)�
0.6
53
1.9
12
.58
1.8
80
.03
45
.59
.75
5.3
51
.79
.0B
.D.
9.0
14
.37
.11
20
.51
6.8
0.3
12
.44
12
3.1
20
14
5
P. Lin et al. / Deep-Sea Research II 81-84 (2012) 79–9482

Author's personal copy
NB
03
6113
00 1
800N
-
17
211
00 2
500W
07
.12
31
.13
1.2
50
.96
0.2
22
.92
.55
.57
.7B
.D.
B.D
.B
.D.
B.D
.2
.71
0.6
5.2
1.0
57
.62
15
.84
82
14
64
10
7.1
23
1.1
31
.19
0.9
70
.32
2.5
B.D
.2
.51
0.2
B.D
.B
.D.
B.D
.B
.D.
3.1
12
.73
.10
.36
2.2
21
5.8
14
07
06
20
6.0
53
1.1
31
.00
1.0
20
.22
2.0
B.D
.2
.09
.2B
.D.
B.D
.B
.D.
B.D
.3
.11
1.2
3.1
0.3
32
.09
14
.31
46
66
6
30
�1
.03
31
.42
1.4
41
.61
0.2
03
.4B
.D.
3.4
13
.2B
.D.
B.D
.B
.D.
B.D
.2
.71
6.7
2.7
1.1
76
.58
19
.33
41
24
77
45
�1
.40
31
.57
1.1
41
.45
0.0
39
.2B
.D.
9.2
19
.60
.4B
.D.
0.4
B.D
.4
.52
9.2
4.5
0.6
33
.84
33
.71
14
84
8
56
(61
)�
1.3
03
1.8
73
.18
1.7
30
.20
49
.1B
.D.
49
.14
1.4
25
.3B
.D.
25
.31
.42
3.9
11
7.3
23
.90
.89
6.2
41
39
.84
52
61
NB
05
6114
40 100N
-
16
911
00 5
500W
06
.94
30
.11
0.7
30
.90
B.D
.1
1.3
2.8
14
.11
0.0
B.D
.B
.D.
B.D
.B
.D.
2.4
21
.35
.20
.51
2.9
82
6.5
11
35
77
10
6.9
33
0.1
10
.65
0.8
8B
.D.
15
.71
.81
7.6
6.7
1.5
B.D
.1
.50
.71
.62
4.6
3.4
0.4
52
.74
27
.31
00
80
8
20
�0
.97
32
.12
4.4
70
.84
0.2
63
4.5
23
.75
8.2
5.8
3.6
B.D
.3
.61
.27
.24
5.1
30
.90
.63
4.4
67
4.8
60
14
4
25
�1
.00
32
.12
2.5
10
.99
0.3
67
5.6
30
.01
05
.66
1.4
5.2
B.D
.5
.21
.67
.41
43
.73
7.4
1.0
96
.65
17
9.5
37
17
8
32
(40
)�
1.0
03
2.1
23
.05
1.0
40
.07
45
.71
5.5
61
.21
84
.38
.5B
.D.
8.5
1.7
7.4
24
0.1
22
.91
.16
7.2
22
61
.42
83
15
Ch
uk
chi
Se
aC
07
7213
20 2
800N
-
16
511
90 3
000W
0�
0.4
72
9.1
30
.76
0.5
40
.31
24
.34
.02
8.3
7.3
1.9
B.D
.1
.90
.22
.03
3.8
6.0
1.4
07
.98
39
.52
02
13
26
15
�0
.81
32
.32
1.2
51
.22
B.D
.2
9.5
12
.84
2.3
53
.71
.9B
.D.
1.9
0.2
4.4
85
.41
7.2
1.2
37
.69
10
2.4
75
44
7
26
�1
.66
32
.81
0.8
61
.68
0.3
63
1.0
11
.94
2.8
9.5
0.2
0.3
0.5
0.7
4.0
41
.41
6.2
1.9
11
1.1
15
6.9
19
56
87
46
(51
)�
1.7
23
3.0
82
.25
2.1
9B
.D.
34
.45
.94
0.3
62
.62
2.2
B.D
.2
2.2
B.D
.1
3.6
11
9.2
19
.62
.84
15
.42
13
8.8
11
17
88
C0
87
2160 2
100N
-
16
212
20 3
400W
0�
0.7
52
8.2
30
.80
0.6
4B
.D.
18
.15
.42
3.5
0.9
1.7
B.D
.1
.70
.75
.82
1.4
11
.20
.51
3.2
43
1.8
10
22
90
10
�0
.65
28
.35
0.7
90
.53
0.4
02
4.3
2.9
27
.28
.37
.1B
.D.
7.1
1.7
16
.44
1.4
19
.40
.54
3.3
95
9.2
57
17
5
20
1.8
83
2.3
11
.20
0.8
1B
.D.
40
.58
.84
9.3
12
9.6
5.7
B.D
.5
.71
.75
.31
77
.51
4.1
1.1
07
.42
19
0.0
39
52
5
30
(32
)�
1.2
93
2.5
61
.53
0.8
40
.11
64
.81
5.7
80
.59
3.1
10
.4B
.D.
10
.42
.95
.61
71
.22
1.2
1.4
99
.55
18
9.6
50
44
9
Co
97
113
50 1
200N
-
15
715
00 3
100W
0�
1.2
12
9.2
4–
0.7
80
.34
6.4
B.D
.6
.48
.0B
.D.
B.D
.B
.D.
B.D
.2
.41
4.4
2.4
0.5
32
.96
16
.81
76
12
25
10
�1
.24
29
.83
–0
.82
B.D
.9
.3B
.D.
9.3
12
.3B
.D.
0.8
0.8
B.D
.2
.72
1.7
3.5
0.4
62
.50
25
.29
97
15
20
�1
.40
30
.66
–0
.85
B.D
.7
.4B
.D.
7.4
23
.6B
.D.
0.1
0.1
B.D
.6
.33
0.9
6.4
0.5
63
.17
37
.38
54
93
30
�0
.67
31
.49
–0
.88
0.3
51
6.2
B.D
.1
6.2
14
.41
.21
.72
.9B
.D.
3.1
31
.94
.90
.42
3.4
43
6.8
93
70
5
40
�1
.07
31
.92
–1
.09
B.D
.6
.4B
.D.
6.4
14
.0B
.D.
0.8
0.8
B.D
.3
.42
0.4
4.2
0.4
92
.51
24
.61
02
59
5
60
(65
)�
1.2
43
2.3
4–
1.5
30
.27
15
.27
.52
2.7
17
.34
.02
.36
.3B
.D.
4.3
36
.61
4.1
0.3
92
.72
50
.75
41
92
S2
27
114
80 5
600N
-
15
411
10 4
600W
04
.13
31
.37
1.1
80
.60
B.D
.2
2.9
B.D
.2
2.9
0.5
2.1
B.D
.2
.14
.52
.73
0.0
2.7
1.2
36
.20
28
.22
20
23
07
20
4.1
73
2.0
81
.50
.59
0.1
43
5.1
B.D
.3
5.1
4.6
8.6
B.D
.8
.62
.46
.15
0.7
6.1
0.9
85
.12
54
.39
48
38
30
3.9
23
2.1
23
.72
0.5
50
.41
28
.31
.12
9.4
14
.66
.9B
.D.
6.9
5.3
6.6
55
.17
.70
.85
4.4
85
7.5
78
58
1
50
3.5
63
2.2
11
.15
0.5
40
.03
23
.9B
.D.
23
.91
.24
.8B
.D.
4.8
2.4
4.4
32
.34
.40
.88
4.8
13
4.3
14
01
09
3
10
02
.32
32
.33
2.3
0.8
2B
.D.
26
.86
.93
3.7
0.4
0.7
1.2
1.9
0.8
3.2
28
.81
1.2
0.8
64
.76
39
.21
21
42
4
14
1
(14
7)
�0
.68
33
.27
5.1
41
.47
B.D
.4
6.2
B.D
.4
6.2
1.6
9.0
0.8
9.8
0.8
7.4
57
.78
.21
.22
7.0
86
5.1
10
98
66
Be
au
fort
Se
aS
24
7211
50 2
300N
-
15
311
40 900W
00
.38
26
.49
0.2
40
.53
0.1
71
6.3
B.D
.1
6.3
B.D
.0
.1B
.D.
0.1
B.D
.2
.91
6.4
2.9
0.1
81
.23
19
.36
44
20
10
�0
.71
27
.29
0.3
10
.68
0.2
03
.1B
.D.
3.1
B.D
.0
.1B
.D.
0.1
B.D
.1
.83
.21
.80
.17
1.1
65
.02
34
64
4
30
�1
.08
30
.14
0.7
20
.85
B.D
.3
.61
.95
.50
.30
.1B
.D.
0.1
B.D
.2
.63
.94
.50
.14
1.0
48
.41
23
23
0
50
�1
.28
30
.74
0.6
41
.00
B.D
.2
.60
.22
.7B
.D.
0.1
B.D
.0
.1B
.D.
1.4
2.6
1.6
0.2
21
.23
4.2
29
27
89
70
�0
.91
31
.48
0.3
2–
––
––
––
––
––
––
0.1
30
.79
––
–
10
0�
1.3
13
2.2
70
.33
1.7
10
.33
0.1
5.4
5.5
B.D
.0
.4B
.D.
0.4
B.D
.1
.80
.47
.20
.07
0.4
77
.66
26
6
15
0�
1.4
53
2.8
9–
1.9
20
.12
B.D
.0
.50
.5B
.D.
0.7
B.D
.0
.7B
.D.
1.4
0.7
1.9
0.1
10
.78
2.6
30
14
01
25
0�
0.1
13
4.5
2–
1.0
9B
.D.
B.D
.1
.61
.67
.10
.7B
.D.
0.7
B.D
.1
.47
.83
.00
.05
0.3
81
0.8
35
12
4
40
00
.76
34
.82
–0
.99
B.D
.0
.1B
.D.
0.1
3.6
0.4
B.D
.0
.40
.21
.44
.21
.40
.06
0.6
05
.41
11
42
8
10
00
0.0
13
4.8
8–
0.8
60
.05
0.2
B.D
.0
.2B
.D.
0.9
B.D
.0
.9B
.D.
2.4
1.1
2.4
0.0
50
.39
3.5
10
91
61
15
00
–0
.30
34
.91
–0
.93
1.5
30
.7B
.D.
0.7
7.1
0.1
1.1
1.1
B.D
.1
.47
.92
.50
.05
0.4
31
0.3
42
17
5
24
89
(25
70
)
–0
.37
34
.95
–1
.13
B.D
.0
.22
.52
.73
5.7
0.4
0.5
0.8
B.D
.0
.63
6.3
3.5
0.0
40
.41
39
.81
01
17
‘‘–’’
de
no
tes
the
da
tais
no
ta
va
ila
ble
.
‘‘B.D
.’’d
en
ote
sco
nce
ntr
ati
on
be
low
sth
ed
ete
ctio
nli
mit
.
P. Lin et al. / Deep-Sea Research II 81-84 (2012) 79–94 83
Author's personal copy
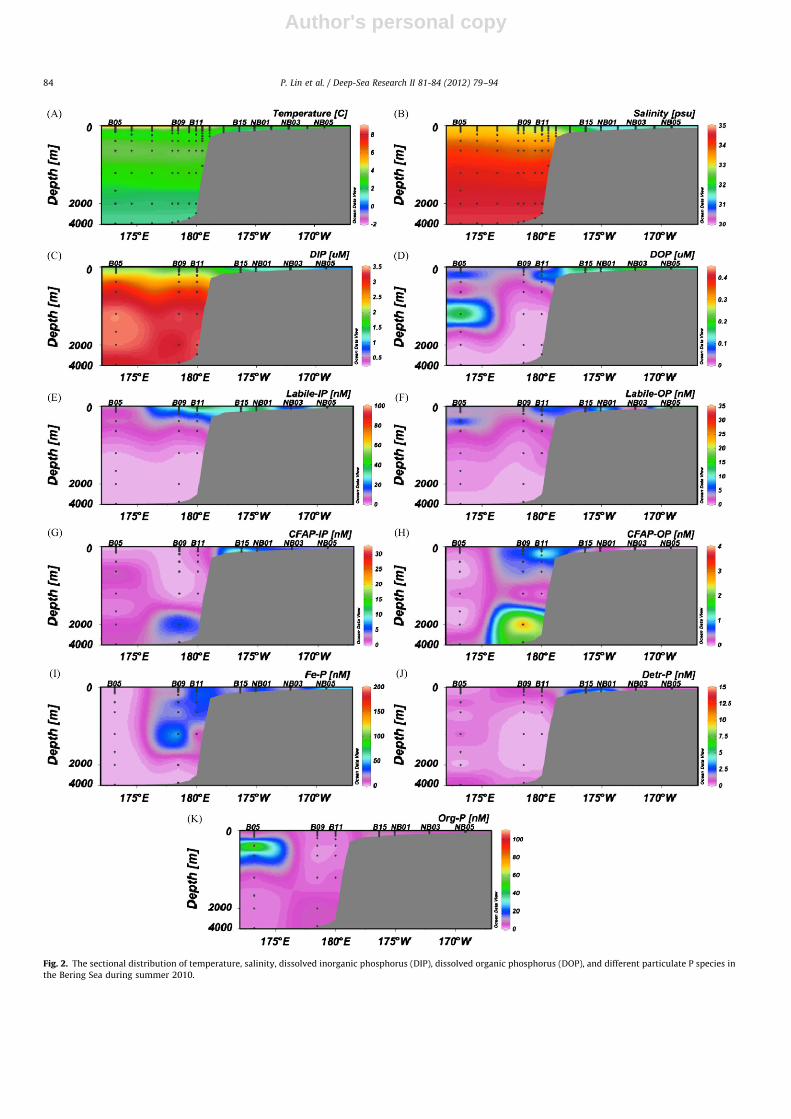
Fig. 2. The sectional distribution of temperature, salinity, dissolved inorganic phosphorus (DIP), dissolved organic phosphorus (DOP), and different particulate P species in
the Bering Sea during summer 2010.
P. Lin et al. / Deep-Sea Research II 81-84 (2012) 79–9484
Author's personal copy
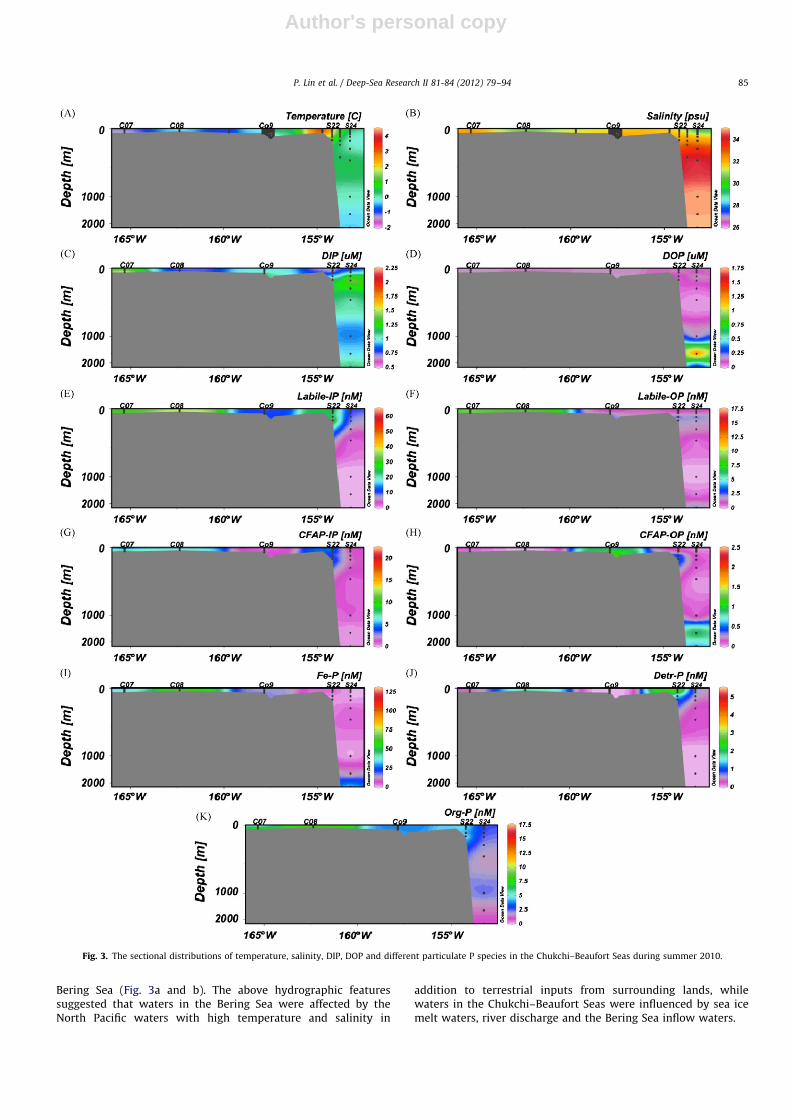
Bering Sea (Fig. 3a and b). The above hydrographic featuressuggested that waters in the Bering Sea were affected by theNorth Pacific waters with high temperature and salinity in
addition to terrestrial inputs from surrounding lands, whilewaters in the Chukchi–Beaufort Seas were influenced by sea icemelt waters, river discharge and the Bering Sea inflow waters.
Fig. 3. The sectional distributions of temperature, salinity, DIP, DOP and different particulate P species in the Chukchi–Beaufort Seas during summer 2010.
P. Lin et al. / Deep-Sea Research II 81-84 (2012) 79–94 85

Author's personal copy
3.2. Variations in dissolved inorganic and organic phosphorus
Concentrations of dissolved inorganic phosphorus (DIP) anddissolved organic phosphorus (DOP) are listed in Table 1 anddepicted as a sectional distribution in Fig. 2c and d and Fig. 3c andd for the Bering Sea and the Chukchi–Beaufort Seas, respectively.DIP concentrations ranged from 0.38 mM at station B15 to1.73 mM at station B05, with an average of 0.8670.45 mMin the Bering Sea. The DOP concentration averaged0.0870.11 mM in the Bering Sea, varying from below the detec-tion limit (B.D.) at most stations to 0.25 mM at station B15. Incontrast to the Bering Sea, concentrations of DIP in the Chukchi–Beaufort Seas were similar for surface waters, ranging from 0.53to 0.78 mM with an average of 0.6270.10 mM, while DOP con-centrations were more variable, ranging from B.D. to 0.34 mMwith an average of 0.1770.17 mM. Overall, surface DIP concen-trations in the Bering Sea were higher than those in the Chukchi–Beaufort Seas.
At deep-water stations, DIP concentrations showed a classicalvertical distribution in the water column, with low DIP in surfacewaters, elevated DIP around the halocline layer (e.g., station B05),and higher concentrations in deeper waters (below 300 m)(Fig. 2c). However, an exception to this trend was observed atS24, showing the highest concentration of DIP at 150 m but asudden decrease from 150 to 250 m (Fig. 3c), clearly demonstrat-ing the two-halocline-layer hydrological characteristic of CanadaBasin (Chen et al., 2003b). At shallow water stations, DIP showeda general increase with increasing water depth (Figs. 2c and 3c).A striking difference existed in deeper water DIP concentrationsbetween the Bering Sea and the Chukchi–Beaufort Seas, with thehighest DIP at Aleutian Basin stations (e.g., B05), the lowest atBeaufort Slope station (S24), and intermediate DIP at Chukchi Seastations (Table 1), showing the influence of North Pacific inflowwaters on the Chukchi Shelf regions adjacent to the Bering Strait,as well as the existence of Atlantic waters with relatively lownutrient concentrations below 250 m at the Beaufort Slope station(Fig. 3c).
The distribution of DOP showed a general decrease withincreasing water depth, although DOP concentrations were rela-tively low and more variable (Figs. 2d and 3d). For example, atstation B05, DOP concentrations were mostly lower than thedetection limit, especially for the deeper water column(41000 m) although an increase occurred at 1500 m at S24,similar to those observed in other oceanic regions (Thomson-Bulldis and Karl, 1998; Loh and Bauer, 2000). For shallow waterstations, DOP seemed to decrease with water depth, with thehighest concentration in subsurface waters, consistent with pre-vious reports (Ridal and Moore, 1990; Suzumura and Ingall, 2004).
3.3. Variations in particulate phosphorus species
The concentrations of five particulate P species, includingLabile-P, Fe-P, CFA-P, Detr-P and Org-P, are also listed inTable 1. Labile-P and CFA-P were further fractionated intoinorganic and organic P fractions, namely, Labile-IP, Labile-OP,CFAP-IP and CFAP-OP. Unfortunately, data for samples of surfacewater, 2000 and 2550 m at station B11, and 70 m at S24, were notavailable due to limited amounts of particulate matter.The sectional distributions of the five particulate P phases in theBering Sea and Chukchi–Beaufort Seas are shown in Figs. 2 and 3,respectively, and discussed below for each particulate phase.
3.3.1. Loosely adsorbed (exchangeable) phosphorus
The concentrations of Labile-IP in surface waters varied from2.9 to 98.8 nM with an average of 35.6736.5 nM in the Bering
Sea, but were much lower in the Chukchi–Beaufort Seas (17.677.1 nM), varying from 6.4 to 24.3 nM. Concentrations of theLabile-OP were generally low, with relatively higher concentra-tions in the Bering Sea (5.575.2 nM) and lower in the Chukchi–Beaufort Seas (1.972.6 nM), similar to the partitioning of Pbetween DIP and DOP.
Similar to the variation in DIP concentrations in the watercolumn, concentrations of Labile-P were generally higheralthough highly variable within the euphotic zone, but theydecreased sharply below the euphotic zone (Fig. 2e and f andFig. 3e and f), suggesting the influence of biological processes anddynamic transformation of P between different species.
3.3.2. Ferric bound phosphorus
Concentrations of Fe-P had a similar spatial distribution(Fig. 2i) as Labile-P in surface waters and had an averageconcentration of 24.8727.7 nM in the Bering Sea, with thehighest concentration at station B11 (82.4 nM) and the lowestat B05 (0.3 nM). Compared to the high concentrations and largevariability of Fe-P in the Bering Sea, concentrations of Fe-P insurface waters were very low in the Chukchi–Beaufort Seas andwere mostly lower than 1 nM except at stations C07 and Co9(Table 1).
Similar to the Labile-IP, Fe-P concentrations were generallylow or near the detection limit in the deeper water column(41000 m) (Figs. 2i and 3i). An exception was found at BeaufortSlope stations (e.g., S24), showing an increase in particulate Fe-Ptoward bottom waters from 1000 m. Different from the largevariability of Fe-P concentrations in the upper water column atdeep stations, Fe-P concentrations were similar at shallow sta-tions in the Bering Sea, except that an elevated concentration wasobserved in bottom waters at station NB05 (Table 1). The elevatedFe-P concentrations were also observed for middle and bottomwaters in the Chukchi Shelf, indicating a source of Fe-P in thecontinental shelf regions, likely from sediment resuspension and/orlateral transport (Chen et al., 2012).
3.3.3. Authigenic carbonate fluorapatite, biogenic apatite and
CaCO3-associated phosphorus
Compared with the concentrations of Labile-P and Fe-P, CFA-Pconcentrations in surface waters were relatively low in the BeringSea and the Chukchi–Beaufort Seas (Fig. 2g and h and 3g and h).In general, CFAP-IP concentrations (average of 1.271.0 nM) in theChukchi–Beaufort Seas were higher than those in the Bering Sea,where detectable CFAP-IP in surface waters was only found at B09(1.0 nM) and NB01 (1.9 nM). The particulate CFAP-OP concentra-tions were below the detection limit in the Chukchi–BeaufortSeas, and had an average of 0.671.1 nM in the Bering Sea.
Distributions of the CFAP-IP and the CFAP-OP showed adynamic variation in the water column at both deepwater andshallow water stations. Elevated CFA-P concentrations werefound in the middle (100 to 500 m) and deep waters(41000 m). For example, the CFAP-IP and CFAP-OP concentra-tions were as high as �6 and 3 nM, respectively, in the AleutianBasin (Table 1). Similarly, elevated concentrations were alsoobserved for Labile-P in the middle waters, suggesting an inputof CFA-P or active transformation processes. In the Bering Shelfregion, CFAP-IP concentrations increased with increasing depthand had a concentration as high as �30 nM in bottom waters atstation B15, while CFAP-OP concentrations were generally belowthe detection limit in the entire water column (e.g., NB03 andNB05) or decreased with increasing depth (e.g., B15 and NB01).Both CFAP-IP and CFAP-OP at the Chukchi Shelf stations seemedto increase with increasing depth, similar to those of CFAP-IPobserved in the Bering Sea. However, the increasing trend for
P. Lin et al. / Deep-Sea Research II 81-84 (2012) 79–9486

Author's personal copy
CFAP-OP in the Chukchi Shelf region was not observed for theBering Shelf. Particulate CFAP-IP could be an important source tothe sediments in both the Bering Sea and the Chukchi Sea, whereCFAP-OP was more variable depending on locations.
3.3.4. Detrital apatite phosphorus
In surface waters, higher concentrations of Detr-P were onlyobserved at five stations (Table 1). Detr-P concentrations were1.1 and 3.7 nM at B09 and B11 near the Bering Slope, respectively,while increased from B.D. to 0.2 nM at stations C07 and C08 in theChukchi–Beaufort Seas. The highest Detr-P concentration of4.5 nM was observed at station S22.
Detr-P showed high concentrations in both upper water columnand bottom waters at deepwater stations although it was consis-tently low at S24 (Fig. 3j). In contrast to the slope and basin regions,Detr-P concentrations increased with increasing depth in the shelfregions, especially at Bering Shelf stations (Table 1, Fig. 2j). Thehighest concentration was found in bottom waters, up to 10.2 and14.3 nM at B15 and NB01, respectively. Over the Chukchi Shelf,concentrations of Detr-P varied from uniformly low or undetectablein the entire water column at C09 to higher levels at other stations,with elevated values at middle water depths at S22, and theyincreased with increasing depth at C07 and C08 (Fig. 3j).
3.3.5. Refractory organic phosphorus
The surface distribution of Org-P was similar to those of DOPand Labile-OP, especially in the Bering Sea (Table 1). The surfaceconcentrations of Org-P in the Bering Sea varied from 2.4 nM atNB05 to 7.7 nM at B15, with an average of 3.872.0 nM. The Org-Pconcentrations were relatively low in the Chukchi–Beaufort Seas,ranging from 2.0 nM at station C07 to 5.8 nM at C08 with anaverage of 3.271.5 nM.
Org-P showed a very dynamic distribution pattern, withmaximum or elevated Org-P concentrations often occurring insubsurface or middle waters for both shelf and deep stations, anda general increase with increasing depth at shelf stations (Fig. 2kand 3k). For example, the highest Org-P concentration wasmeasured at 300 m depth at B05 in the Aleutian Basin(106.8 nM, Table 1). Elevated Org-P concentrations were alsoobserved at 30–60 m in the shelf regions.
3.4. Partitioning of P between inorganic and organic in dissolved and
particulate phases
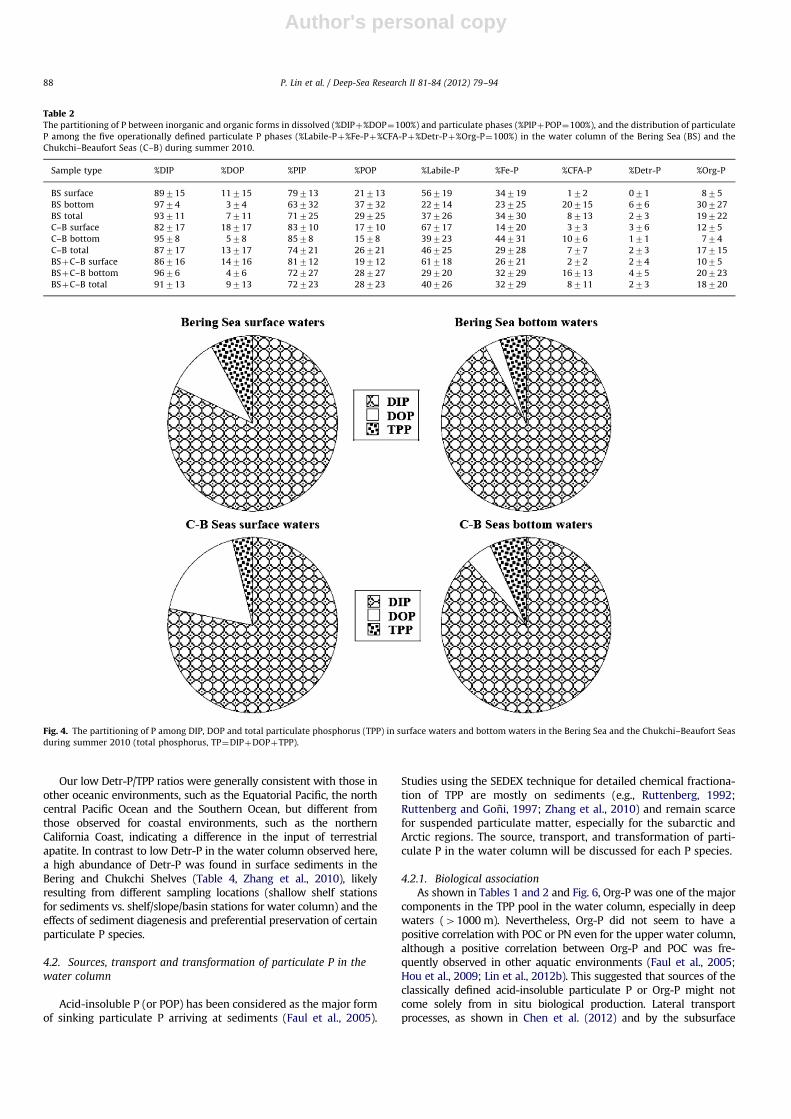
In the dissolved phase, DIP was the predominant form of theTDP pool in the water column. On average, surface DIP comprisedup to 89715% of the TDP in the Bering Sea and 82717% in theChukchi–Beaufort Seas (Table 2). As shown in Fig. 4, the percentageof DIP in TDP pool increased from surface to deep waters in boththe Bering Sea (9774%) and the Chukchi–Beaufort Seas (9578%),showing biological uptake of DIP in the upper water column.Conversely, the percentage of DOP was higher in surface watersthan that in deep waters, and higher in the Chukchi–Beaufort Seasthan in the Bering Sea, indicating biological production of DOP insurface waters. Higher DOP/TDP ratios in the Chukchi–BeaufortSeas were also consistent with these areas’ higher biologicalproductivity (Cota et al., 1996; Mathis et al., 2009).
TPP concentrations were generally much lower than TDP in thewater column (Table 2, Fig. 4). On average, TPP comprised 474%of the total P pool (TP¼TPPþTDP) in the study areas.The percentage of TPP in the TP pool decreased from 876% insurface waters to 577% in deeper waters in the Bering Sea, butincreased from 472% in surface waters to 776% in bottomwaters in the Chukchi–Beaufort Seas (Fig. 4), showing the impor-tance of TPP in bottom waters of the shelf region.
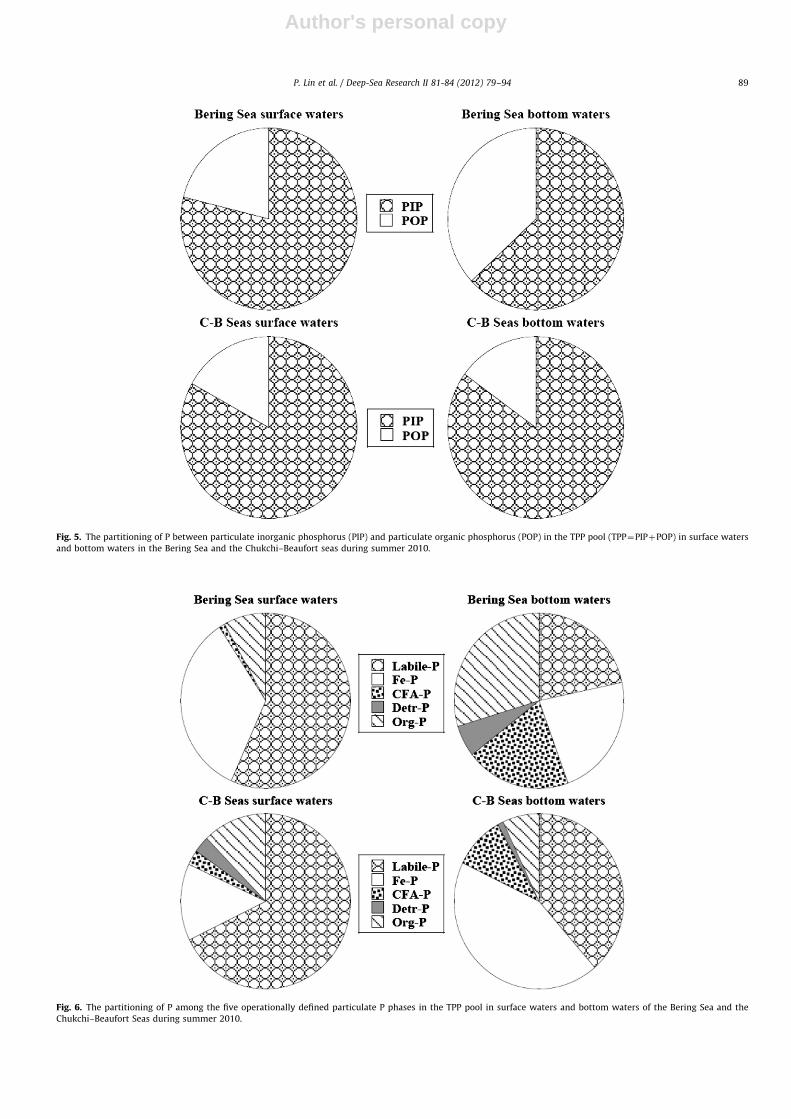
Similar to the partitioning of inorganic and organic P in thedissolved phase, PIP had a significantly higher contribution(72723%) to the TPP pool than POP (Table 2). On average, PIPmade up 71725% of the TPP pool in the Bering Sea and 74721%in the Chukchi–Beaufort Seas (Table 2). As shown in Fig. 5, PIP/TPPdecreased from the surface (79713%) to the bottom (63732%) inthe Bering Sea, indicating a considerable transformation betweendifferent particulate P species in the water column. The oppositewas true in the Chukchi–Beaufort Seas, which had a slight increasein %PIP toward the bottom from 83710 to 8578%.
Based on the results of chemical fractionation of the TPP pool,Labile-P and Fe-P were the two predominant phases, especially inthe upper water column (o1000 m), while CFA-P and Detr-Pwere the two least abundant particulate species (Table 2, Fig. 6).Percentages of Labile-P were higher in surface waters comparedto deeper waters. On average, Labile-P comprised 56719 and67717% of TPP in surface waters in the Bering Sea and theChukchi–Beaufort seas, respectively, and decreased to 22714and 39723% in bottom waters. A similar trend from surface tobottom waters was found for the Fe-P in the Bering Sea, varyingfrom 34719% at the surface to 23725% in bottom waters, whilea different trend was observed for the Chukchi–Beaufort Seasshowing a significant increase from surface (14720%) to bottom(44731%). In contrast to the decreasing trend for Labile-P and Fe-P in the Bering Sea, the other three particulate P species generallyincreased from surface to bottom waters, showing an increasefrom 172% at the surface to 20715% in bottom waters for CFA-Pand from 875 to 30727% for Org-P. The increase in Detr-P withdepth was relatively small, varying from �071% at the surface to676% in bottom waters. In the Chukchi–Beaufort Seas, CFA-Pincreased from 373% at the surface to 1076% in bottom waters,showing a potential source of CFA-P in deeper waters. Conversely,Detr-P and Org-P in surface waters comprised 376 and 1275%of TPP in the Chukchi–Beaufort Seas, respectively, and decreasedto 171% and 774% in bottom waters.
4. Discussion
4.1. Comparisons in the abundance and partitioning of P species
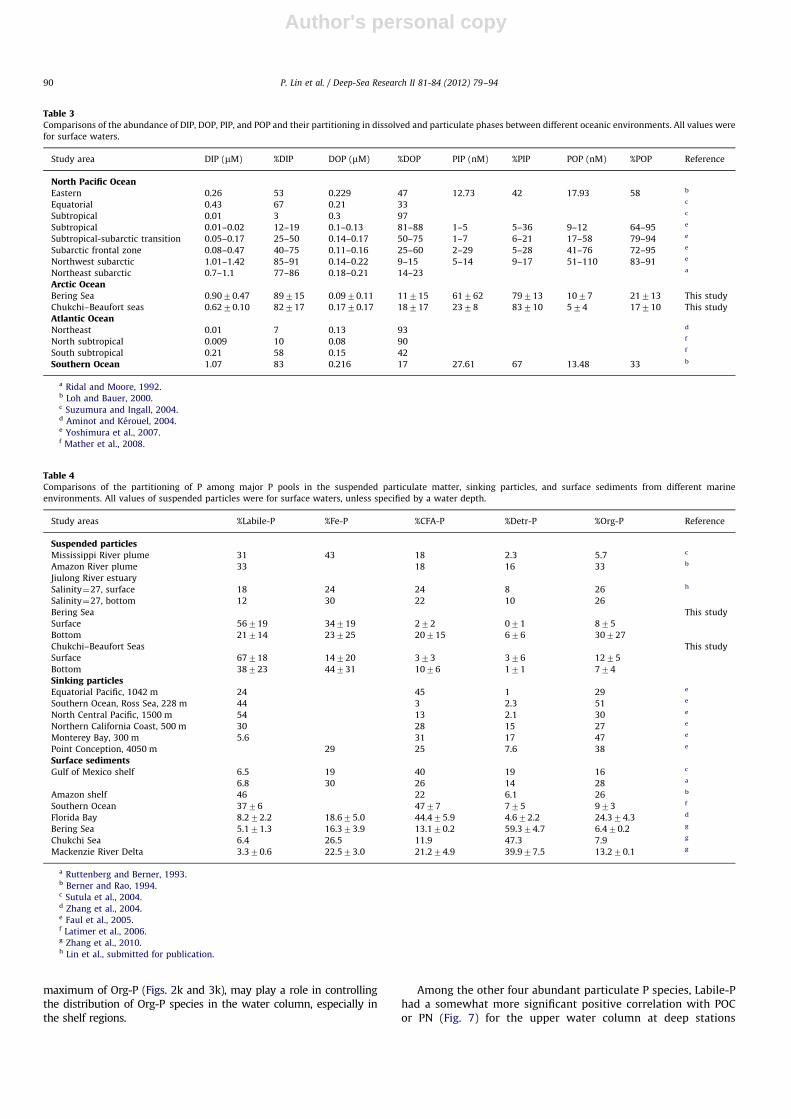
The predominance of DIP in the TDP in the study areas wasdistinct from observations made in the subtropical North PacificOcean and the North Atlantic Ocean (Table 3). On average, thesurface DIP concentration and DIP/TDP ratio in the Bering Seawere comparable with those of the subarctic North Pacific,showing an input of upwelling North Pacific deep waters. Inaddition, nutrients from the Bering Sea could also affect theChukchi Sea, resulting in a gradient of DIP concentrations withlow values on the Beaufort Slope.
The average DOP concentration in the Bering Sea was generallylower than those reported for other oceanic environments(Table 3). However, the average DOP concentration in theChukchi–Beaufort Seas was higher or comparable to that foundin the Southern Ocean.
The low abundance of CFA-P, especially in surface waters, wassimilar to the case of sinking particles in the Ross Sea, likelyrelated to low abundance of carbonate in particulate matter(Latimer et al., 2006; Dutay et al., 2009; Kretschmer et al.,2011). Low CFA-P abundance could be a unique characteristic ofhigh latitude oceanic environments. Additionally, the increase inCFA-P abundance observed in deeper waters of the Bering Sea andthe Chukchi–Beaufort Seas was also consistent with previousobservations showing an increase in authigenic P from sinkingparticles to bottom sediments (Faul et al., 2005).
P. Lin et al. / Deep-Sea Research II 81-84 (2012) 79–94 87
Author's personal copy
Our low Detr-P/TPP ratios were generally consistent with those inother oceanic environments, such as the Equatorial Pacific, the northcentral Pacific Ocean and the Southern Ocean, but different fromthose observed for coastal environments, such as the northernCalifornia Coast, indicating a difference in the input of terrestrialapatite. In contrast to low Detr-P in the water column observed here,a high abundance of Detr-P was found in surface sediments in theBering and Chukchi Shelves (Table 4, Zhang et al., 2010), likelyresulting from different sampling locations (shallow shelf stationsfor sediments vs. shelf/slope/basin stations for water column) and theeffects of sediment diagenesis and preferential preservation of certainparticulate P species.
4.2. Sources, transport and transformation of particulate P in the
water column
Acid-insoluble P (or POP) has been considered as the major formof sinking particulate P arriving at sediments (Faul et al., 2005).
Studies using the SEDEX technique for detailed chemical fractiona-tion of TPP are mostly on sediments (e.g., Ruttenberg, 1992;Ruttenberg and Goni, 1997; Zhang et al., 2010) and remain scarcefor suspended particulate matter, especially for the subarctic andArctic regions. The source, transport, and transformation of parti-culate P in the water column will be discussed for each P species.
4.2.1. Biological association
As shown in Tables 1 and 2 and Fig. 6, Org-P was one of the majorcomponents in the TPP pool in the water column, especially in deepwaters (41000 m). Nevertheless, Org-P did not seem to have apositive correlation with POC or PN even for the upper water column,although a positive correlation between Org-P and POC was fre-quently observed in other aquatic environments (Faul et al., 2005;Hou et al., 2009; Lin et al., 2012b). This suggested that sources of theclassically defined acid-insoluble particulate P or Org-P might notcome solely from in situ biological production. Lateral transportprocesses, as shown in Chen et al. (2012) and by the subsurface
Table 2The partitioning of P between inorganic and organic forms in dissolved (%DIPþ%DOP¼100%) and particulate phases (%PIPþPOP¼100%), and the distribution of particulate
P among the five operationally defined particulate P phases (%Labile-Pþ%Fe-Pþ%CFA-Pþ%Detr-Pþ%Org-P¼100%) in the water column of the Bering Sea (BS) and the
Chukchi–Beaufort Seas (C–B) during summer 2010.
Sample type %DIP %DOP %PIP %POP %Labile-P %Fe-P %CFA-P %Detr-P %Org-P
BS surface 89715 11715 79713 21713 56719 34719 172 071 875
BS bottom 9774 374 63732 37732 22714 23725 20715 676 30727
BS total 93711 7711 71725 29725 37726 34730 8713 273 19722
C–B surface 82717 18717 83710 17710 67717 14720 373 376 1275
C–B bottom 9578 578 8578 1578 39723 44731 1076 171 774
C–B total 87717 13717 74721 26721 46725 29728 777 273 17715
BSþC–B surface 86716 14716 81712 19712 61718 26721 272 274 1075
BSþC–B bottom 9676 476 72727 28727 29720 32729 16713 475 20723
BSþC–B total 91713 9713 72723 28723 40726 32729 8711 273 18720
Fig. 4. The partitioning of P among DIP, DOP and total particulate phosphorus (TPP) in surface waters and bottom waters in the Bering Sea and the Chukchi–Beaufort Seas
during summer 2010 (total phosphorus, TP¼DIPþDOPþTPP).
P. Lin et al. / Deep-Sea Research II 81-84 (2012) 79–9488
Author's personal copy
Fig. 5. The partitioning of P between particulate inorganic phosphorus (PIP) and particulate organic phosphorus (POP) in the TPP pool (TPP¼PIPþPOP) in surface waters
and bottom waters in the Bering Sea and the Chukchi–Beaufort seas during summer 2010.
Fig. 6. The partitioning of P among the five operationally defined particulate P phases in the TPP pool in surface waters and bottom waters of the Bering Sea and the
Chukchi–Beaufort Seas during summer 2010.
P. Lin et al. / Deep-Sea Research II 81-84 (2012) 79–94 89
Author's personal copy
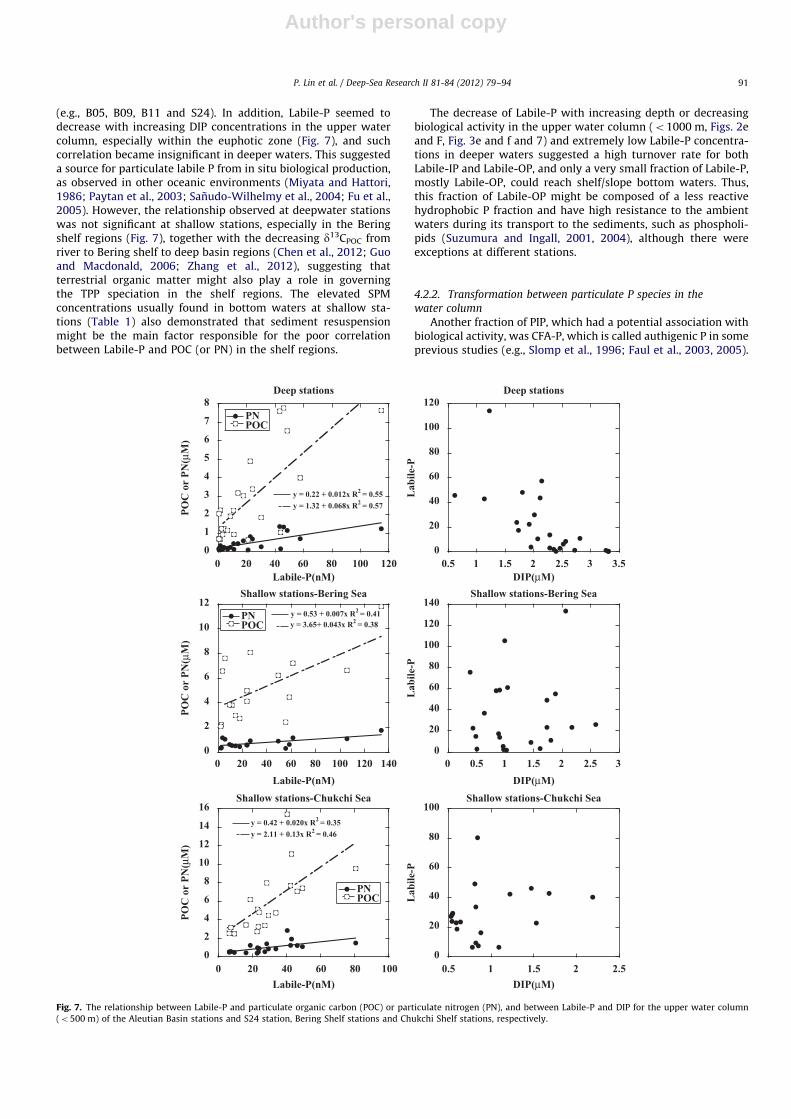
maximum of Org-P (Figs. 2k and 3k), may play a role in controllingthe distribution of Org-P species in the water column, especially inthe shelf regions.
Among the other four abundant particulate P species, Labile-Phad a somewhat more significant positive correlation with POCor PN (Fig. 7) for the upper water column at deep stations
Table 3Comparisons of the abundance of DIP, DOP, PIP, and POP and their partitioning in dissolved and particulate phases between different oceanic environments. All values were
for surface waters.
Study area DIP (mM) %DIP DOP (mM) %DOP PIP (nM) %PIP POP (nM) %POP Reference
North Pacific OceanEastern 0.26 53 0.229 47 12.73 42 17.93 58 b
Equatorial 0.43 67 0.21 33 c
Subtropical 0.01 3 0.3 97 c
Subtropical 0.01–0.02 12–19 0.1–0.13 81–88 1–5 5–36 9–12 64–95 e
Subtropical-subarctic transition 0.05–0.17 25–50 0.14–0.17 50–75 1–7 6–21 17–58 79–94 e
Subarctic frontal zone 0.08–0.47 40–75 0.11–0.16 25–60 2–29 5–28 41–76 72–95 e
Northwest subarctic 1.01–1.42 85–91 0.14–0.22 9–15 5–14 9–17 51–110 83–91 e
Northeast subarctic 0.7–1.1 77–86 0.18–0.21 14–23 a
Arctic OceanBering Sea 0.9070.47 89715 0.0970.11 11715 61762 79713 1077 21713 This study
Chukchi–Beaufort seas 0.6270.10 82717 0.1770.17 18717 2378 83710 574 17710 This study
Atlantic OceanNortheast 0.01 7 0.13 93 d
North subtropical 0.009 10 0.08 90 f
South subtropical 0.21 58 0.15 42 f
Southern Ocean 1.07 83 0.216 17 27.61 67 13.48 33 b
a Ridal and Moore, 1992.b Loh and Bauer, 2000.c Suzumura and Ingall, 2004.d Aminot and Kerouel, 2004.e Yoshimura et al., 2007.f Mather et al., 2008.
Table 4Comparisons of the partitioning of P among major P pools in the suspended particulate matter, sinking particles, and surface sediments from different marine
environments. All values of suspended particles were for surface waters, unless specified by a water depth.
Study areas %Labile-P %Fe-P %CFA-P %Detr-P %Org-P Reference
Suspended particlesMississippi River plume 31 43 18 2.3 5.7 c
Amazon River plume 33 18 16 33 b
Jiulong River estuary
Salinity¼27, surface 18 24 24 8 26 h
Salinity¼27, bottom 12 30 22 10 26
Bering Sea This study
Surface 56719 34719 272 071 875
Bottom 21714 23725 20715 676 30727
Chukchi–Beaufort Seas This study
Surface 67718 14720 373 376 1275
Bottom 38723 44731 1076 171 774
Sinking particlesEquatorial Pacific, 1042 m 24 45 1 29 e
Southern Ocean, Ross Sea, 228 m 44 3 2.3 51 e
North Central Pacific, 1500 m 54 13 2.1 30 e
Northern California Coast, 500 m 30 28 15 27 e
Monterey Bay, 300 m 5.6 31 17 47 e
Point Conception, 4050 m 29 25 7.6 38 e
Surface sedimentsGulf of Mexico shelf 6.5 19 40 19 16 c
6.8 30 26 14 28 a
Amazon shelf 46 22 6.1 26 b
Southern Ocean 3776 4777 775 973 f
Florida Bay 8.272.2 18.675.0 44.475.9 4.672.2 24.374.3 d
Bering Sea 5.171.3 16.373.9 13.170.2 59.374.7 6.470.2 g
Chukchi Sea 6.4 26.5 11.9 47.3 7.9 g
Mackenzie River Delta 3.370.6 22.573.0 21.274.9 39.977.5 13.270.1 g
a Ruttenberg and Berner, 1993.b Berner and Rao, 1994.c Sutula et al., 2004.d Zhang et al., 2004.e Faul et al., 2005.f Latimer et al., 2006.g Zhang et al., 2010.h Lin et al., submitted for publication.
P. Lin et al. / Deep-Sea Research II 81-84 (2012) 79–9490
Author's personal copy
(e.g., B05, B09, B11 and S24). In addition, Labile-P seemed todecrease with increasing DIP concentrations in the upper watercolumn, especially within the euphotic zone (Fig. 7), and suchcorrelation became insignificant in deeper waters. This suggesteda source for particulate labile P from in situ biological production,as observed in other oceanic environments (Miyata and Hattori,1986; Paytan et al., 2003; Sanudo-Wilhelmy et al., 2004; Fu et al.,2005). However, the relationship observed at deepwater stationswas not significant at shallow stations, especially in the Beringshelf regions (Fig. 7), together with the decreasing d13CPOC fromriver to Bering shelf to deep basin regions (Chen et al., 2012; Guoand Macdonald, 2006; Zhang et al., 2012), suggesting thatterrestrial organic matter might also play a role in governingthe TPP speciation in the shelf regions. The elevated SPMconcentrations usually found in bottom waters at shallow sta-tions (Table 1) also demonstrated that sediment resuspensionmight be the main factor responsible for the poor correlationbetween Labile-P and POC (or PN) in the shelf regions.
The decrease of Labile-P with increasing depth or decreasingbiological activity in the upper water column (o1000 m, Figs. 2eand F, Fig. 3e and f and 7) and extremely low Labile-P concentra-tions in deeper waters suggested a high turnover rate for bothLabile-IP and Labile-OP, and only a very small fraction of Labile-P,mostly Labile-OP, could reach shelf/slope bottom waters. Thus,this fraction of Labile-OP might be composed of a less reactivehydrophobic P fraction and have high resistance to the ambientwaters during its transport to the sediments, such as phospholi-pids (Suzumura and Ingall, 2001, 2004), although there wereexceptions at different stations.
4.2.2. Transformation between particulate P species in the
water column
Another fraction of PIP, which had a potential association withbiological activity, was CFA-P, which is called authigenic P in someprevious studies (e.g., Slomp et al., 1996; Faul et al., 2003, 2005).
0
1
2
3
4
5
6
7
8Deep stations
PNPOC
y = 0.22 + 0.012x R2 = 0.55y = 1.32 + 0.068x R2 = 0.57
POC
or
PN(µ
M)
Labile-P(nM)
0
2
4
6
8
10
12Shallow stations-Bering Sea
y = 0.53 + 0.007x R2 = 0.41y = 3.65+ 0.043x R2 = 0.38
POC
or
PN(µ
M)
Labile-P(nM)
0
20
40
60
80
100
120Deep stations
Lab
ile-P
DIP(µM)
0
20
40
60
80
100
120
140Shallow stations-Bering Sea
Lab
ile-P
DIP(µM)
0
20
40
60
80
100Shallow stations-Chukchi Sea
Lab
ile-P
DIP(µM)
0
2
4
6
8
10
12
14
16
0 20 40 60 80 100 120
0 20 40 60 80 100 120 140
0.5 1 1.5 2 2.5 3 3.5
0 0.5 1 1.5 2 2.5 3
0.5 1 1.5 2 2.50 20 40 60 80 100
Shallow stations-Chukchi Sea
y = 0.42 + 0.020x R2 = 0.35y = 2.11 + 0.13x R2 = 0.46
POC
or
PN(µ
M)
Labile-P(nM)
PNPOC
PNPOC
Fig. 7. The relationship between Labile-P and particulate organic carbon (POC) or particulate nitrogen (PN), and between Labile-P and DIP for the upper water column
(o500 m) of the Aleutian Basin stations and S24 station, Bering Shelf stations and Chukchi Shelf stations, respectively.
P. Lin et al. / Deep-Sea Research II 81-84 (2012) 79–94 91

Author's personal copy
As shown in Tables 1 and 2 and Fig. 6, the CFA-P contribution tothe TPP pool was generally low, especially in the upper watercolumn, and its correlation with POC or DIP was weak. Highconcentrations of CFA-P were usually found in deeper waters,regardless of location, suggesting a weak control on CFA-Pdistributions by biological processes, likely related to the factthat plankton communities in high latitude marine environmentswere dominated by siliceous rather than calcareous organisms(Dutay et al., 2009; Crosta, 2011). Therefore, the cycling pathwayswere likely different between CFA-P and Labile-P components.Interestingly, while the concentration of total reactive P, the sumof Labile-P, Fe-P, CFA-P and Org-P, remained relatively constant,both Labile-P and Fe-P concentrations were consistently low anddecreased with depth in the deeper water column, but CFA-Pabundance increased with increasing depth (Tables 1 and 2).We hypothesized that there existed an active transformationbetween Labile-P and CFA-P in deep waters (e.g., at stations B05and B09). Indeed, many previous studies have reported anefficient transformation from labile forms of P to authigenic P inthe water column and the sediments (Ruttenberg and Berner,1993; Filippelli and Delaney, 1995, 1996; Anderson et al., 2001;Faul et al., 2003, 2005), consistent with our conclusion that theCFA-P, i.e., authigenic P, was an important P sink in deeper waters.In contrast, at shallow water stations, it was clear that sedimentresuspension and/or lateral transport was the major cause for theincreasing CFA-P abundance in bottom waters in the Bering Seaand the Chukchi shelves (Fig. 2g, h and 3g, h), as was also evidentfrom the elevated SPM concentrations in bottom waters (Table 1).
4.2.3. Fe-P and Detr-P
While sources of particulate Labile-P, CFA-P and Org-P weremostly biologically associated, Fe-P and Detr-P were largely derivedfrom terrestrial inputs and had been shown to be the two mostabundant particulate P species in coastal waters and sediments (e.g.,Zhang et al., 2010; Lin et al., submitted for publication.). The Fe-Pspecies could be more active than the Detr-P in the biogeochemicalrecycling of P in the water column, and the Detr-P was believed tobe the most inert P species and could be used as a proxy ofterrigenous origin (van Cappellen and Berner, 1988; Ruttenberg,1992; Hou et al., 2009). Thus, the behavior of Fe-P could be similarto that of Labile-P, as also suggested by Faul et al. (2005).
Concentrations of Fe-P were somewhat similar throughout thewater column, but the concentrations of Detr-P, like Labile-P andCFA-P, increased with water depth on the Bering shelf whereevidence of sediment resuspension was observed. However, highFe-P concentrations were found at the layer between 75 and500 m at station B11 and between 300 and 1000 m at B09(Fig. 2i). Furthermore, high Detr-P concentrations were also foundin the euphotic zone but became undetectable below 100 m atboth B09 and B11 (Fig. 2j), where SPM concentrations remainedlow at these water depths (Table 1). Such unexpected distributionpatterns of Fe-P and Detr-P indicated sources for these terrestrialparticulate P species, especially in the middle water column forFe-P. Since it was less likely to have upwelling inputs or sedimentresuspension for the middle water column based on the tem-perature, salinity, DIP and SPM data (Table 1), high concentrationsof Detr-P and Fe-P in the middle water column at deep waterstations (e.g., B09 and B11) were likely derived from the lateraltransport of shelf bottom waters containing high Detr-P (e.g., B15)and the interaction between shelf and slope waters (for Fe-P).
At station S24 on the Beaufort Slope, Fe-P and Detr-P weregenerally undetectable in the upper halocline layer (o250 m,Fig. 3i and j), suggesting that inputs of Fe-P from the North Pacificwaters had faded away during their transport from the Bering Seato the Arctic region (Chen et al., 2003a). However, high concen-trations of Detr-P and Fe-P were found at S22 in the Chukchi shelf
region adjacent to the Beaufort Slope (Table 1, Fig. 3i and j),indicating little exchange between the upper halocline waters atthe Chukchi shelf station (S22) and the Beaufort Slope station(S24) during our sampling period. Significant differences intemperature and salinity (Table 1) between these two stationsalso supported our speculation here. In addition, the concurrenthigh concentrations of Fe-P and very low concentrations of Detr-Pin the lower halocline (4250 m) at S24 suggested that the Fe-Pwas likely produced from in situ physicochemical processes. Onthe other hand, the relatively higher concentrations of Detr-P,salinity and temperature were ubiquitously observed at differentwater depths among the stations over the Chukchi Shelf (e.g.,S22), indicating a possible influence of water masses from theBering Sea (Chen et al., 2012), such as the Alaska Coastal Currentthat might bring additional terrestrial apatite.
4.3. The importance of considering chemical and phase speciation
In this study, particulate Labile-IP, Fe-P, CFAP-IP and Detr-Pwere classified as PIP, while Labile-OP, CFAP-OP and Org-P(or acid-insoluble P) belonged to POP in the TPP pool. In addition,total reactive P concentrations were calculated from the sum ofthe Labile-P, Fe-P, CFA-P and Org-P species (Sutula et al., 2004;Faul et al., 2005). As shown in Tables 1 and 2 and Fig. 5, PIPcomprised a major fraction of TPP at most stations, especially inthe upper water column, but not at B05 and in bottom waters forsome stations. High PIP/TPP ratios had also been reported forsinking particles in other oceanic environments (Loh and Bauer,2000; Benitez-Nelson et al., 2004; Faul et al., 2005). The generallylow POP/TPP ratios (Table 2) also resulted in high POC/POP molarratios up to 2477 (Table 1). However, due to the low Detr-P to TPPratios and thus high reactive P abundance (490%) except inbottom waters at stations with sediment resuspension, POC toreactive P ratios (POC/Rea-P) were much lower than POC/POPratios (Table 1), and they became less than or close to the Redfieldratio (106) at some stations (e.g., B11), implying high turnoverrates of particulate phosphorus in the water column. As reportedby Faul et al. (2005) for sinking particulate matter, some of theregenerated biological P could be retained in the TPP pool ratherthan being released into the TDP pool.
As shown above, the Org-P was not correlated with POC or PN,but the Labile-P was broadly correlated with POC or DIP (Fig. 7).Thus, Labile-P seemed more active in the upper water column(o300 m) and should be among the active P components in theregeneration and cycling of particulate P in the water column.The apparent POC/POP ratios could be high (e.g., 2105 at B09) ifthe Labile-P was not included in the POP pool. Sanudo-Wilhelmyet al. (2004) addressed the effect on the Redfield ratio from thepartitioning of P into the surface-adsorbed and intracellular Ppools. Similar processes were observed for the distribution of Fe-Pspecies with considerable regeneration of Fe-P in the upper watercolumn (e.g., Fig. 2i) or additional Fe-P inputs in the middle watercolumn (e.g., B09 and B11), resulting in significantly low POC/Rea-P ratios. Furthermore, it seemed that not all regenerated P had tobe released into the dissolved pool in the water column(Faul et al., 2005), as also supported by the distribution andpartitioning of CFA-P in the study area, especially in the deeperwater column (Section 4.2.2, Fig. 2g and h).
Overall, the three TPP species, Labile-P, Fe-P and CFA-P, whichhave been usually excluded from the acid-insoluble POP pool andfrom the calculation of particulate organic C/P ratios, may allactively participate in the regeneration and dynamic cycling of Pin the water column. Therefore, the use of traditionally defined acid-insoluble organic P concentrations in the suspended particulatematter may not be sufficient for assessing the elemental stoichio-metry of particulate matter and deciphering the biogeochemical
P. Lin et al. / Deep-Sea Research II 81-84 (2012) 79–9492

Author's personal copy
processes of P species in oceanic environments, especially in regionswhere POP abundance is lower than that of PIP, like the westernArctic Ocean and the Southern Ocean (Table 3). Thus, in addition tothe quantity of total P, a detailed chemical and phase speciation ofthe TPP pool is indispensable for a better understanding of thetransport and biogeochemical cycling of P in marine environments.
5. Conclusions
Dissolved and particulate P species, including DIP, DOP, PIP andPOP, as well as five operationally defined particulate P species,were measured in the Bering Sea and Chukchi/Beaufort Seasduring July-September 2010. Both DIP and PIP were the predomi-nant species in dissolved and particulate phases with higher DIPbut lower DOP in the Bering Sea and higher DOP, but lower DIP inthe Chukchi–Beaufort Seas. Variations in spatial distributions ofDIP, DOP, PIP, and POP and their dynamic transformation werelargely related to (1) in situ primary productivity in the upperwater column, (2) water masses from the North Pacific, the AlaskaCoastal Current, and the Atlantic deep waters, and (3) sedimentresuspension and lateral transport processes.
Among the five major particulate P species, Labile-P and Fe-Pcontributed the highest percentage to the TPP pool, especially inshallow waters, while the CFA-P and Detr-P were the two leastabundant particulate P species. Although Org-P was classicallyconsidered as the most biologically associated phase in the TPP, itwas found that Labile-P, rather than Org-P, was broadly correlatedwith POC, PN or DIP, indicating its association with biologicalprocesses, and thus showing that it was an active P component inthe dynamic cycling of particulate P in the water column.Although another possible biologically associated fraction, CFA-P,had a weak relationship with POC, an active transformationbetween Labile-P and CFA-P was observed in deeper waters, andthe CFA-P could be a main sink in deeper waters. In contrast to theabove three biologically associated particulate P species, theabundance and distribution of Fe-P and Detr-P seemed to bemostly controlled by physicochemical processes, such as sedimentresuspension in the shallow water regions, lateral transport ofshelf bottom waters, and the interaction of shelf and slope waters,especially at the edge between continental slope and shelf regions.
Measurements of inorganic and organic P in dissolved andparticulate phases and their detailed chemical and phase specia-tion could provide important information to elucidate effectivelythe source, transport, and transformation of P species in the watercolumn and their interrelationship with environmental para-meters in the marine environment.
Acknowledgments
We are grateful for the assistance of the crew and scienceparty of the R/V Xuelong in sample collection during the 4thChinese Arctic Research Expedition to the Arctic. We also thanktwo anonymous reviewers for their constructive comments andSarah Cooley for technical editing on the manuscript. This workwas supported in part by a Cheung Kong Scholarship throughMinistry of Education of China, the National Natural ScienceFoundation of China (41125020), and a special scientific researchproject for public welfare supported by the State Oceanic Admin-istration of China (201105022-4).
References
Aagaard, K., Carmack, E.C., 1989. The role of sea ice and other fresh water in theArctic circulation. J. Geophys. Res. 94, 14485–14498.
Aminot, A., Kerouel, R., 2004. Dissolved organic carbon, nitrogen and phosphorus inthe N-E Atlantic and the N-W Mediterranean with particular reference to non-refractory fractions and degradation. Deep Sea Res. I 51 (12), 1975–1999.
Anderson, L.D., Delaney, M.L., Faul, K.L., 2001. Carbon to phosphorus ratios insediments: implications for nutrient cycling. Global Biogeochem. Cycles 15,65–79.
Benitez-Nelson, C.R., 2000. The biogeochemical cycling of phosphorus in marinesystems. Earth Sci. Rev. 51, 109–135.
Benitez-Nelson, C.R., O’Neill, L., Kolowith, L.C., Pellechia, P., Thunell, R., 2004.Phosphonates and particulate organic phosphorus cycling in an anoxic marinebasin. Limnol. Oceanogr. 49, 1593–1604.
Berner, R., Rao, J.-L., 1994. Phosphorus in sediments of the Amazon River andestuary: implications for the global flux of phosphorus to the sea. Geochim.Cosmochim. Acta 58, 2333–2339.
Bjorkman, K., Karl, D.M., 1994. Bioavailability of inorganic and organic phosphoruscompounds to natural assemblages of microorganisms in Hawaiian coastalwaters. Mar. Ecol. Prog. Ser. 111, 265–273.
Bjorkman, K.M., Karl, K.M., 2003. Bioavailability of dissolved organic phosphorusin the euphotic zone at Station ALOHA. North Pacific Subtrop. Gyre. Limnol.Oceanogr. 48, 1049–1057.
Boye, M., Nishioka, J., Croot, P., Laan, P., Timmermans, K.R., Strass, V.H., Takeda, S.,de Baar, H.J.W., 2010. Significant portion of dissolved organic Fe complexes infact is Fe colloids. Mar. Chem. 122, 20–27.
Cai, Y., Guo, L., 2009. Abundance and variation of colloidal organic phosphorus inriverine, estuarine and coastal waters in the northern Gulf of Mexico. LimnolOceanogr 54, 1393–1402.
Cai, W., Chen, L., Chen, B., Gao, Z., Lee, S.H., Chen, J., Pierrot, D., Sullivan, K., Wang, Y.,Hu, X., Huang, W.-J., Zhang, Y., Xu, S., Murata, A., Grebmeiser, J.M., Jones, E.P.,Zhang, H., 2010. Decrease in the CO2 uptake capacity in an ice-free Arctic Oceanbasin. Science 329, 556–559, http://dx.doi.org/10.1126/science.1189338.
Carmack, E., Macdonald, R.W., Jasper, S., 2004. Phytoplankton productivity on theCanadian Shelf of the Beaufort Sea. Mar. Ecol. Prog. Ser. 277, 37–57.
Cavender-Bares, K.K., Karl, D.M., Chisholm, S.W., 2001. Nutrient gradients in thewestern North Atlantic Ocean: relationship to microbial community structureand comparison to patterns in the Pacific Ocean. Deep Sea Res. I 48,2373–2395.
Chen, M., Huang, Y., Guo, L., Cai, P., Yang, W., Liu, G., Qiu, Y., 2002. Biologicalproductivity and carbon cycling in the Arctic Ocean. Chin. Sci. Bull. 47 (12),1037–1040.
Chen, M., Huang, Y., Cai, P., Guo, L., 2003a. Particulate organic carbon export fluxesin the Canada Basin and Bering Sea as derived from 234Th/238U disequilibria.Arctic 56 (1), 32–44.
Chen, M., Huang, Y., Jin, M., Qiu, Y., 2003b. The isotopic tracers for the formationmechanisms of the upper and lower thermocline waters in Canada Basin. Sci.China, Ser. D 33 (2), 127–138, http://dx.doi.org/10.3321/j.issn:1006.9267.2003.02.005.
Chen, M., Guo, L., Ma, Q., Qiu, Y., Zhang, R., Lv, E., Huang, Y., 2006. Zonal patterns of13C, 15N and 210Po in the tropical and subtropical North Pacific. Geophys. Res.Lett. 33, L04609, http://dx.doi.org/10.1029/2005GL025186.
Chen, M., Ma, Q., Guo, L., Qiu, Y., Li, Y., Yang, W., 2012. Importance of lateraltransport to 210Pb budget in the eastern Chukchi Sea. Deep Sea Res. II 81–84,53–62.
Cota, G.F., Pomeroy, L.R., Harrison, W.G., Jones, E.P., Peters, F., Sheldon, W.M.,Weingartner, T.R., 1996. Nutrients, primary production and microbial hetero-trophy in the southeastern Chukchi Sea: arctic summer nutrient depletion andheterotrophy. Mar. Ecol. Prog. Ser. 135, 247–258.
Crosta, X., 2011. Marine diatoms in polar and sub-polar environments and theirapplication to Late Pleistocene paleoclimate reconstruction. IOP Conf. Ser.:Earth Environ. Sci. 14, 012006, http://dx.doi.org/10.1088/1755-1315/14/1/012006.
Delaney, M.L., 1998. Phosphorus accumulation in marine sediments and theoceanic phosphorus cycle. Global Biogeochem. Cycles 12, 563–572.
Dutay, J.-C., Lacan, F., Roy-Barman, M., Bopp, L., 2009. Influence of particle size andtype on 231Pa and 230Th simulation with a global coupled biogeochemical-ocean general circulation model: a first approach. Geochem. Geophys. Geosyst.10 (1), http://dx.doi.org/10.1029/2008GC002291.
Eicken, H., Gradinger, R., Gaylord, A., Mahoney, A., Rigor, I., Melling, H., 2005.Sediment transport by sea ice in the Chukchi and Beaufort Seas: increasingimportance due to changing ice conditions? Deep-Sea Res. II 52, 3281–3302.
Faul, K.L., Anderson, L.D., Delaney, M.L., 2003. Late Cretaceous and early Paleogenenutrient and paleoproductivity records from Blake Nose, western NorthAtlantic Ocean. Paleoceanography 18 (2), 1042, http://dx.doi.org/10.1029/2001PA000722.
Faul, K.L., Paytan, A., Delaney, M.L., 2005. Phosphorus distribution in sinkingoceanic particulate matter. Mar. Chem. 97, 307–333.
Filippelli, G.M., Delaney, M.L., 1995. Phosphorus geochemistry and accumulationrates in the eastern equatorial Pacific Ocean: results from Leg 138. Proc. OceanDrill. Program Part B Sci. Results 138, 757–767.
Filippelli, G.M., Delaney, M.L., 1996. Phosphorus geochemistry of equatorial Pacificsediments. Geochim. Cosmochim. Acta 60, 1479–1495.
Froelich, P., Bender, M.L., Luedtke, N.A., Heath, G.R., Devries, T., 1982. The marinephosphorus cycle. Am. J. Sci. 282, 474–511.
Fu, F.-X., Xiang, Y., Leblanc, K., Sanudo-Wilhelmy, S.A., Hutchins, D.A., 2005. Thebiological and biogeochemical consequences of phosphate scavenging ontophytoplankton cell surfaces. Limnol. Oceanogr. 50, 1459–1472.
P. Lin et al. / Deep-Sea Research II 81-84 (2012) 79–94 93

Author's personal copy
Grebmeier, J.M., Harvey, H.R., 2005. The Western Arctic Shelf–Basin Interactions(SBI) project: an overview. Deep-Sea Res. II 52, 3109–3115.
Grebmeier, J.M., Cooper, L.W., Feder, H.,.M., Sirenko, B.I., 2006. Ecosystemdynamics of the Pacific-influenced Northern Bering and Chukchi Seas in theAmerasian Arctic. Prog. Oceanogr. 71, 331–361.
Guo, L., Santschi, P.H., 1997. Isotopic and elemental characterization of colloidalorganic matter from the Chesapeake Bay and Galveston Bay. Mar. Chem. 59,1–15.
Guo, L., Zhang, J.-Z., Gueguen, C., 2004. Speciation and fluxes of nutrients (N, P, Si)from the upper Yukon River. Global Biogeochem. Cycles 18 (1), GB1038,http://dx.doi.org/10.1029/2003GB2152.
Guo, L., Macdonald, R.W., 2006. Source and transport of terrigenous organic matterin the upper Yukon River: evidence from isotope (d13C, D14C, and d15N)composition of dissolved, colloidal, and particulate phases. Global Biogeo-chem. Cycles 20, GB2011, http://dx.doi.org/10.1029/2005GB002593.
Harrison, W.G., Harris, L.R., 1986. Isotope-dilution and its effects on measurementsof nitrogen and phosphorus uptake by oceanic microplankton. Mar. Ecol. Prog.Ser. 27, 253–261.
Hou, L., Liu, M., Yang, Y., Ou, D., Lin, X., Chen, H., Xu, S., 2009. Phosphorusspeciation and availability in intertidal sediments of the Yangtze Estuary,China. Appl. Geochem. 24, 120–128.
Huang, X.-L., Zhang, J.-Z., 2010. Spatial variation in sediment-water exchange ofphosphorus in Florida Bay: AMP as a model organic compound. Environ. Sci.Technol. 44, 7790–7795, http://dx.doi.org/10.1021/es100057r.
Huang, X.-L., Zhang, J.-Z., 2011. Phosphorus sorption on marine carbonate sedi-ment: phosphonate as model organic compounds. Chemosphere 85 (8),1227–1232, http://dx.doi.org/10.1016/j.chemosphere.2011.07.016.
Karl, D.M., Craven, D.B., 1980. Effects of alkaline phosphatase activity onnucleotide measurements in aquatic microbial communities. Appl. Environ.Microbiol. Sept, 549–561.
Karl, D.M., Bjorkman, K.M., Dore, J.E., Fujieki, L., Hebel, D.V., Houlihan, T., Letelier,R.M., Tupas, L.M., 2001. Ecological nitrogen-to-phosphorus stoichiometry atstation ALOHA. Deep-Sea Res. II 48, 1529–1566.
Kawaguchi, K., 2001. PSECS: pacific subarctic ecosystem study (preface). Oceano-graphy 57 (3), 251–252.
Koroleff, F., 1983. Determination of phosphorus. In: Grasshoff, K., Kremling, K.,Ehrhardt, M. (Eds.), Methods of Seawater Analysis. Verlag Chemie, pp. 167–173.
Kretschmer, S., Geibert, W., Rutgers van der Loeff, M.M., Schnabel, C., Xu, S.,Mollenhauer, G., 2011. Fractionation of 230Th, 231Pa, and 10Be induced byparticle size and composition within an opal-rich sediment of the AtlanticSouthern Ocean. Geochim. Cosmochim. Acta 75, 6971–6987.
Latimer, J.C., Filippelli, G.M., 2001. Terrigenous input and paleoproductivity in theSouthern Ocean. Paleoceanography 16, 627–643.
Latimer, J.C., Filippelli, G.M., Hendy, I., Newkirk, D.R., 2006. Opal-associatedparticulate phosphorus: implications for the marine P cycle. Geochim. Cos-mochim. Acta 70, 3843–3854.
Lin, P., Chen, M., Guo, L., 2012. Speciation and transformation of phosphorus andits mixing behavior in the Bay of St. Louis estuary in the northern Gulf ofMexico. Geochim Cosmochim Acta 87, 283–298, http://dx.doi.org/10.1016/j.gca.2012.03.040.
Lin, P., Chen, M., Cai, Y., Guo, L. Partitioning and mixing behavior of phosphorusspecies in the Jiulong River Estuary, China. Estuar. Coast. Shelf. Sci. Submittedfor publication.
Loh, A.N., Bauer, J.E., 2000. Distribution, partitioning and fluxes of dissolved andparticulate organic C, N and P in the eastern North Pacific and SouthernOceans. Deep-Sea Res. I 47, 2287–2316.
Mather, R.L., Reynolds, S.E., Wolff, G.A., Williams, R.G., Torres-valdes, S., Woodward,E.M., Landolfi, A., Pan, X., Sanders, R., Achterberg, E., 2008. Phosphorus cycling inthe North and South Atlantic Ocean subtropical gyres. Nat. Geosci. 1, 439–443.
Mathis, J.T., Pickart, R.S., Hansell, D.A., Kadko, D., Bates, N.R., 2007. Eddy transportof organic carbon and nutrients from the Chukchi Shelf: impact on the upperhalocline of the western Arctic Ocean. J. Geophys. Res. 112, C05011, http://dx.doi.org/10.1029/2006JC003899.
Mathis, J.T., Bates, N.R., Hansell, D.A., Babila, T., 2009. Net community productionin the northeastern Chukchi Sea. Deep-Sea Res. II 56, 1213–1222.
McGuire, A.D., Anderson, L., Christensen, T.R., Dallimore, S., Guo, L., Hayes, D.,Heimann, M., Macdonald, R., Roulet, N., 2009. Sensitivity of the Carbon Cycle inthe Arctic to Climate Change (Review). Ecol. Monograph. 79 (4), 523–555.
Miyata, K., Hattori, A., 1986. A simple fractionation method for determination ofphosphorus components in phytoplankton: application to natural populationsof phytoplankton in summer surface waters of Tokyo Bay. J. Oceanogr. Soc. Jpn.42, 255–265.
Parsons, T.R., Maita, Y., Lalli, C.M., 1984. A Manual of Chemical and BiologicalMethods for Seawater Analysis. Pergamon Press, New York.
Paytan, A., Cade-Menun, B.J., McLaughlin, K., Faul, K.L., 2003. Selective phosphorusregeneration of sinking marine particles: evidence from 31P-NMR. Mar. Chem.82, 55–70.
Perry, M.J., Eppley, R.W., 1981. Phosphate uptake by phytoplankton in the centralNorth Pacific Ocean. Deep-Sea Res. I 28, 39–49.
Ping, C.-L., Michaelson, G.J., Guo, L., Jorgenson, M.T., Kanevskiy, M., Shur, Y., Dou, F.,Liang, J., 2011. Soil carbon and material flux across the eroding coastline of the
Beaufort Sea, Alaska. J. Geophys. Res. Biogeosci. 116, G02004, http://dx.doi.org/10.129/2010JG001588.
Ridal, J.J., Moore, R.M., 1990. A re-examination of the measurement of dissolvedorganic phosphorus in seawater. Mar. Chem. 29, 19–31.
Ridal, J.J., Moore, R.M., 1992. Dissolved organic phosphorus in the northeastsubarctic Pacific Ocean. Limnol. Oceanogr. 37 (5), 1067–1075.
Ruttenberg, K.C., 1990. Diagenesis and Burial of Phosphorus in Marine Sediments,Implications for the Marine Phosphorus Budget. Ph.D. Thesis. Yale Univ., 375 pp.
Ruttenberg, K.C., 1992. Development of a sequential extraction method fordifferent forms of phosphorus in marine sediments. Limnol. Oceanogr. 37,1460–1482.
Ruttenberg, K.C., Berner, R.A., 1993. Authigenic apatite formation and burial insediments from non-upwelling, continental margin environments. Geochim.Cosmochim. Acta 57, 991–1007.
Ruttenberg, K.C., Goni, M.A., 1997. Phosphorus distribution, C:N:P ratios, andd13Coc in arctic, temperate, and tropical coastal sediments: tools for char-acterizing bulk sedimentary organic matter. Mar. Geol. 139, 123–145.
Ruttenberg, K.C., Sulak, D.J., 2011. Sorption and desorption of dissolved organicphosphorus onto iron (oxyhydr)oxides in seawater. Geochim. Cosmochim.Acta 75, 4095–4112.
Sanudo-Wilhelmy, S.A., Tovar-Sanchez, A., Fu, F.-X., Capone, D.G., Carpenter, E.J.,Hutchins, D.A., 2004. The impact of surface-adsorbed phosphorus on phyto-plankton Redfield stoichiometry. Nature 432, 897–901.
Slomp, C.P., Epping, E.H.G., Helder, W., van Raaphorst, W., 1996. A key role foriron-bound phosphorus in authigenic apatite formation in North Atlanticcontinental platform sediments. J. Mar. Res. 54, 1179–1205.
Slomp, C.P., Malschaert, J.F.P., van Raaphorst, W., 1998. The role of adsorption insediment-water exchange of phosphate in North Sea continental marginsediments. Limnol. Oceanogr. 43, 832–846.
Smith Jr., K.L., Williams, P.M., Druffel, E.R.M., 1989. Upward fluxes of particulateorganic matter in the deep North Pacific. Nature (London) 337, 724–726.
Sutula, M., Bianchi, T.S., McKee, B.A., 2004. Effect of seasonal sediment storage inthe lower Mississippi River on the flux of reactive particulate phosphorus tothe Gulf of Mexico. Limnol. Oceanogr. 49 (6), 2223–2235.
Suzumura, M., Ingall, E.D., 2001. Concentration of lipid phosphorus and itsabundance in dissolved and particulate organic phosphorus in coastal sea-water. Mar. Chem. 75, 141–149.
Suzumura, M., Ingall, E.D., 2004. Distribution and dynamics of various forms ofphosphorus in seawater: insights from field observations in the Pacific Oceanand a laboratory experiment. Deep-Sea Res. I 51, 1113–1130.
Tamburini, F., Huon, S., Steinmann, P., Grousset, F.E., Adatte, T., Follmi, K.B., 2002.Dysaerobic conditions during Heinrich events 4 and 5: evidence fromphosphorus distribution in a North Atlantic deep-sea core. Geochem. Cosmo-chim. Acta 66, 4069–4083.
Takata, H., Kuma, K., Iwade, S., Yamajyoh, Y., Yamaguchi, A., Takagi, S., Sakaoka, K.,Yamashita, Y., Tanoue, E., Midorikawa, T., Kimura, K., Nishioka, J., 2004. Spatialvariability of iron in the surface water of the northwestern North PacificOcean. Mar. Chem. 86, 139–157.
Thomson-Bulldis, A., Karl, D., 1998. Application of a novel method for phosphorusdeterminations in the oligotrophic North Pacific Ocean. Limnol. Oceanogr. 43,1565–1577.
van Cappellen, P., Berner, R.A., 1988. A mathematical model for the earlydiagenesis of phosphorus and fluorine in marine sediments: apatite precipita-tion. Am. J. Sci. 288, 289–333.
van der Zee, C., Slomp, C.P., van Raaphorst, W., 2002. Authigenic P formation andtotal reactive P burial in sediments of the Nazare canyon on the Iberian margin(NE Atlantic). Mar. Geol. 185, 379–392.
Walsh, J.J., 1989. Arctic carbon sinks: present and future. Global Biogeochem.Cycles 3 (4), 393–411.
Wu, J., Sunda, W., Boyle, E.A., Karl, D.M., 2000. Phosphate depletion in the WesternNorth Atlantic Ocean. Science 289, 759–762.
Yoshimura, T., Nishioka, J., Saito, H., Takeda, S., Tsuda, A., Wells, M.L., 2007.Distributions of particulate and dissolved organic and inorganic phosphorus inNorth Pacific surface waters. Mar. Chem. 103, 112–121.
Zhang, J.-Z., Fischer, C.J., Ortner, P.B., 2004. Potential availability of sedimentaryphosphorus to sediment resuspension in Florida Bay. Global Biogeochem.Cycles 18, GB4008, http://dx.doi.org/10.1029/2004GB002255.
Zhang, J.-Z., Huang, X.-L., 2007. Relative importance of solid -phase phosphorusand iron in sorption behavior of sediments. Int. J. Environ. Sci. Technol. 41 (8),2789–2795, http://dx.doi.org/10.1021/es061836q.
Zhang, J.-Z., Guo, L., Fischer, C., 2010. Abundance and chemical speciationof phosphorus in sediments of the Mackenzie River Delta, the Chukchi Seaand the Bering Sea: importance of Detrital Apatite. Aquat. Geochem. 16,353–371.
Zhang, J.-Z., Huang, X.-L., 2011. Effect of temperature and salinity on phosphatesorption on marine sediments. Environ. Sci. Technol. 45, 6831–6837, http://dx.doi.org/10.1021/es200867p.
Zhang, R., Chen, M., Guo, L., Gao, Z., Ma, Q., Cao, J., Qiu, Y., Li, Y., 2012. Variations inthe isotopic composition of particulate organic carbon and their relation withcarbon dynamics in the western Arctic Ocean. Deep-Sea Res. II 81–84, 72–78.
P. Lin et al. / Deep-Sea Research II 81-84 (2012) 79–9494
Related Documents











