Arch. Gerontol. Geriatr., 5 (1986) 333-342 333 Elsevier AGG 00162 The change with age in biogenic amines and their metabolites in the striatum of the rat * A. Machado, J. Cano and M. Santiago Department of Biochemist~, Faculty of Pharmacy, University of Sevilla, Sevilla, Spain (Received 10 July 1986; revised version received 16 September 1986; accepted 18 September 1986) Summa~ The changes in the content of the biogenic amines and their metabolites in the striatum of the rat during the aging period (3-30 months) have been studied. The maximum levels of dopamine (DA) have been found at 6 months of age and this concentration is maintained until 24 months. Between 24 to 30 months there is a decrease in the concentration of this compound. At that time, there is a slight increase in 3,4-dihydroxyphenyl acetic acid (DOPAC) and homovanillic acid (HVA) concentration, the main metabolites of DA, which suggests a slight increase in DA metabolism. The 3-methoxytyramine (3-MT) concentration also increases at this time. The maximum concentration of noradrenaline (NA) was also found at 6 months of age. Tryptophan has the maximum concentration after DA and this is maintained over the life-span of the rat. The concentration of serotonin (5-HT) is high and does not change during this period. However, the concentration of 5-HT, as occurred with DA, decreased between 24 and 30 months. Also the DA/5-HT ratio does not change during the period studied. biogenic amines; dopamine, serotonin, noradrenaline, aging; brain monoamine contents during aging Introduction Neural control plays a crucial role in regulating all body functions -- particularly homeostasis -- and this is of prime importance for understanding the major events of the life-span, including aging. The central nervous system (CNS) undergoes alterations in structure, biochemistry and function during development and with advancing age. However, the actual evidence is only sufficient to construct several theories of aging which await further verification (Timiras and Bignami, 1976; Timiras, 1978; Timiras et al., 1979; Walker and Timiras, 1981). Some of the reasons * Presented in the program of the Verzhr Memorial Symposium on Cell Aging, organized in the frame of the 2nd European Congress of Cell Biology, Budapest, 6-11 July, 1986. Correspondence should be addressed to Dr. A. Machado, Departamento de Bioquimica, Facultad de Farmacia, Universidad de Sevilla, Sevilla, Spain. 0167-4943/86/$03.50 © 1986 Elsevier Science Publishers B.V.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Arch. Gerontol. Geriatr., 5 (1986) 333-342 333 Elsevier
AGG 00162
The change with age in biogenic amines and their metabolites in the striatum of the rat *
A. Machado , J. Cano and M. Santiago Department of Biochemist~, Faculty of Pharmacy, University of Sevilla, Sevilla, Spain
(Received 10 July 1986; revised version received 16 September 1986; accepted 18 September 1986)
Summa~
The changes in the content of the biogenic amines and their metabolites in the striatum of the rat during the aging period (3-30 months) have been studied. The maximum levels of dopamine (DA) have been found at 6 months of age and this concentration is maintained until 24 months. Between 24 to 30 months there is a decrease in the concentration of this compound. At that time, there is a slight increase in 3,4-dihydroxyphenyl acetic acid (DOPAC) and homovanillic acid (HVA) concentration, the main metabolites of DA, which suggests a slight increase in DA metabolism. The 3-methoxytyramine (3-MT) concentration also increases at this time. The maximum concentration of noradrenaline (NA) was also found at 6 months of age. Tryptophan has the maximum concentration after DA and this is maintained over the life-span of the rat. The concentration of serotonin (5-HT) is high and does not change during this period. However, the concentration of 5-HT, as occurred with DA, decreased between 24 and 30 months. Also the DA/5-HT ratio does not change during the period studied.
biogenic amines; dopamine, serotonin, noradrenaline, aging; brain monoamine contents during aging
Introduction
Neural control plays a crucial role in regulating all body functions - - particularly homeostasis - - and this is of prime importance for understanding the major events of the life-span, including aging. The central nervous system (CNS) undergoes alterations in structure, biochemistry and function during development and with advancing age. However, the actual evidence is only sufficient to construct several theories of aging which await further verification (Timiras and Bignami, 1976; Timiras, 1978; Timiras et al., 1979; Walker and Timiras, 1981). Some of the reasons
* Presented in the program of the Verzhr Memorial Symposium on Cell Aging, organized in the frame of the 2nd European Congress of Cell Biology, Budapest, 6-11 July, 1986.
Correspondence should be addressed to Dr. A. Machado, Departamento de Bioquimica, Facultad de Farmacia, Universidad de Sevilla, Sevilla, Spain.
0167-4943/86/$03.50 © 1986 Elsevier Science Publishers B.V.

334
for the failure to establish both the central role of the CNS and that of neurotrans- mitters in these processes, reflect the practical and theoretical difficulties inherent in neurotransmitter identification and measurement of their brain levels, turnover and functional implication.
In this paper, our primary interest is directed to the monoaminergic system. The monoamines dopamine (DA), noradrenaline (NA) and serotonin (5-HT) have many possible roles. The 5-HT has a role in neuroendocrine regulation and has often been implicated as a 'pacemaker' of growth, development and aging (Walker and Timiras, 1981). The early appearance of the monoamine neurosystem in the CNS of mammals has led to the speculation that it has a regulatory function in brain development (Kasamatsu and Pettigrew, 1979; Lauder et al., 1983). Furthermore, pharmacologic and behavioral studies suggest that central dopaminergic and noradrenergic neurons are, to a certain extent, functional within a few days of birth.
Through the study of the content of biogenic amines and their metabolites, in some of the important periods of the life of the rat, some of their possible roles could be pointed out.
For this study we have measured the biogenic amines, dopamine (DA), noradrenaline (NA), and serotonin (5-HT) and their main metabolites (3-methoxy- tyramine, 3-MT; 3,4-dihydroxyphenyl acetic acid, DOPAC; homovanillic acid, HVA; normetanephrine, NMN; metanephrine, MN; tryptophan, Tryp; 5-hydroxy- tryptophan, 5-HTP; and 5-hydroxy-3-indoleacetic acid, 5-HIAA) during aging of the striatum of the rat, using high performance liquid chromatography with electro- chemical detection (HPLC-ED).
The striatum is an important area of the CNS and has an extremely rich content of known neurotransmitter-related compounds. This study is of gerontological relevance since changes have been described in biogenic amines in some diseases, some of them age-related, e.g. in Huntington's disease a decrease in DA and HVA levels in the caudate nucleus has been found (Bernheimer et al., 1973) and, in Parkinson's disease, a decrease in the three monoamines: DA, NA and 5-HT.
Material and Methods
Chromatography
All experiments were performed using a model Perkin-Elmer Series 4 pump in conjunction with an electrochemical detector (model LC-4B, Bioanalytical System) with a glassy carbon and a Perkin-Elmer HS-3 C18 column (100 x 4.6 mm). The potential was set at 0.8 V (vs. Ag/AgC1 reference electrode). To protect the analytical column, we used a small, easily replaceable pre-column.
Chemical
Dopamine (DA), 3-methoxytyramine (3-MT), 3,4-dihydroxyphenyl acetic acid (DOPAC), homovanillic acid (HVA), noradrenaline (NA), normetanephrine (NMN),

335
metanephrine (MN), 5-hydroxytryptamine (5-HT), tryptophan (Tryp), 5-hydroxy- tryptophan (5-HTP), and 5-hydroxy-3-indoleacetic acid (5-HIAA), Were purchased from Sigma Chemical Co. All other chemicals and solvents were of analytical or reagent grade and were used without further purification. All aqueous solutions were prepared from deionized distilled water.
Reagents
The mobile phase was prepared with water, acetonitrile and formic acid and reagent grade sodium octyl sulfate, diethylamine, EDTA, citric acid and potassium hydroxide. The solvent mixture was filtered in vacuo through a 0.45-/~m filter, and degassed by vacuum prior to ultrasound bath to eliminate air bubbles, which might interfere in the electrochemical assay.
~ample preparation
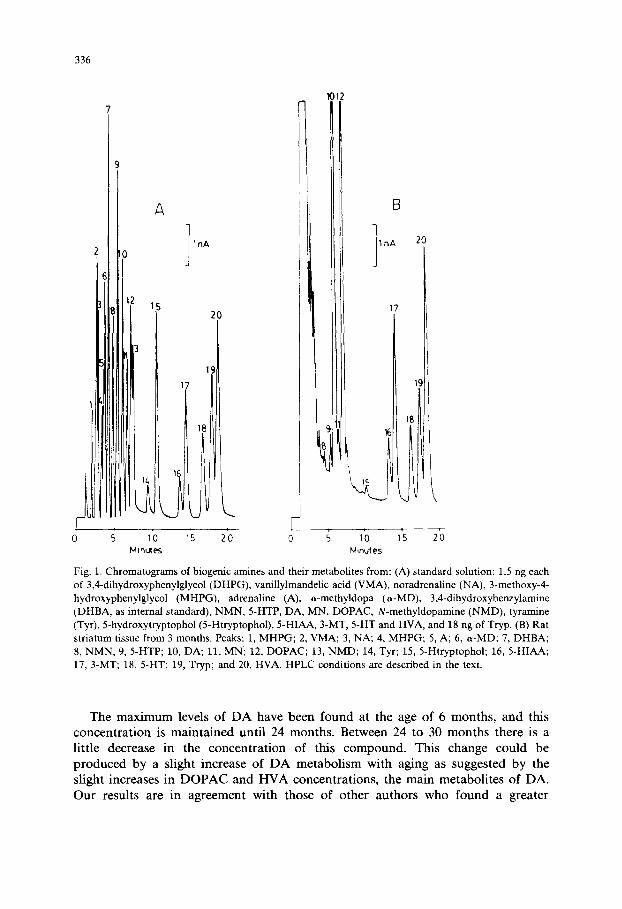
Stock standards were prepared by accurately weighing 4 mg of each of the standards and dissolving them in 4 ml of 0.1 M perchloric acid and 1 mM sodium bisulfite. The stock was then frozen at - 2 0 ° C in 0.5-ml aliquots. Fig. 1 shows the chromatogram of the standards.
Male rats of the Wistar strain were used. Animals were decapitated between 10.00 and 11.00 a.m. and the whole brain was quickly removed. Then the caudo- putamen was dissected out and weighed to the nearest milligram.
All tissues were homogenized in 0.2 ml of 0.1 M perchloric acid containing 1 mM sodium bisulfite by ultrasonic desintegration over ice using a Labsonic 1510. Samples were centrifuged at 30000 x g for 15 min at 4°C, and the supernatant was then filtered through a 0.2-#m filter.
Material was injected using a high-pressure injection valve (Rheodine) with a 6-20/~1 sample loop. Analyses were performed using a flow rate of 1.0 ml /min at room temperature. Concentrations in brain samples were calculated with the aid of (linear) calibration curves obtained after the injection of pure standards. Fig. 1 shows the chromatogram of the caudo-putamen nucleus from 3-month-old rats.
Results and Discussion
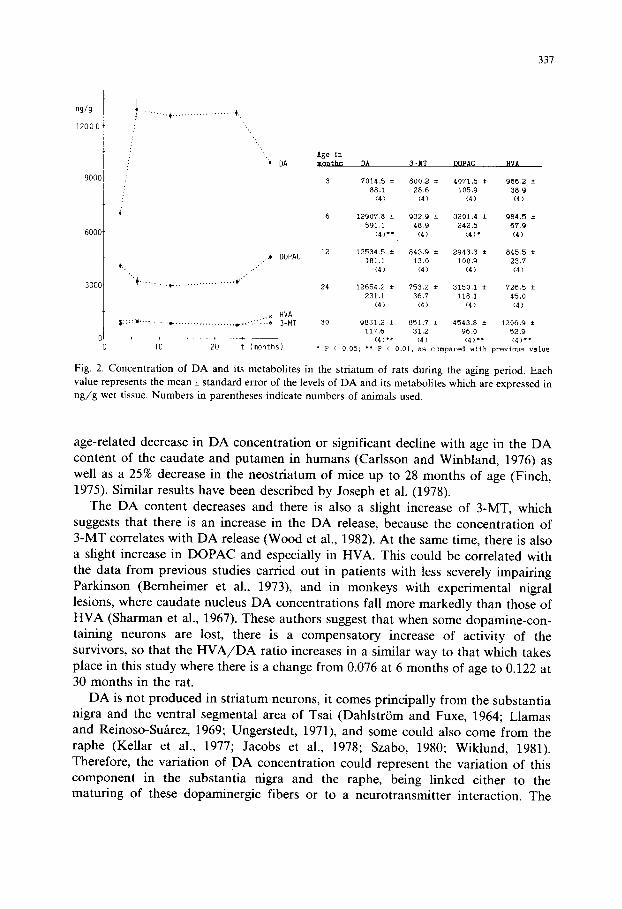
Fig. 2 shows the levels of concentration of DA and its metabolites in the striatum of the rat at different ages. As can be seen, the data are in agreement with the concept that the striatum contains a very large amount of DA. As we will see later, DA is the biogenic amine found in the highest concentration, since in adulthood its levels are about 18 times higher than those of 5-HT and 28 times higher than those of NA.
This DA concentration is in the same range as those recently found in the rat striatum with HPLC by Pileblad et al. (985). DA has an important functional role because an increase (Bird et al., 1979), or a decrease (Hornykiewicz, 1976) in this neurotransmitter are associated with dysfunction of the nervous system.
336
l lnA
20
1 1
1
1012
InA 20
17
19
o s ~o ~5 20 ~ ~ ;s 2'0 Minutes Minutes
Fig. 1. Chromatograms of biogenic amines and their metabolites from: (A) standard solution: 1.5 ng each of 3,4-dihydroxyphenylglycol (DHPG), vanillylmandelic acid (VMA), noradrenaline (NA), 3-methoxy-4- hydroxyphenylglycol (MHPG), adrenaline (A), a-methyldopa (a-MD), 3,4-dihydroxybenzylamine (DHBA, as internal standard), NMN, 5-HTP, DA, MN, DOPAC, N-methyldopamine (NMD), tyramine (Tyr), 5-hydroxytryptophol (5-Htryptophol), 5-HIAA, 3-MT, 5-HT and HVA, and 18 ng of Tryp. (B) Rat striatum tissue from 3 months. Peaks: 1, MHPG; 2, VMA; 3, NA; 4, MHPG; 5, A; 6, a-MD; 7, DHBA; 8, NMN, 9, 5-HTP; 10, DA; 11, MN; 12, DOPAC; 13, NMD; 14, Tyr; 15, 5-Htryptophol; 16, 5-HIAA; 17, 3-MT; 18, 5-HT; 19, Tryp; and 20, HVA. HPLC conditions are described in the text.
The maximum levels of D A have been found at the age of 6 months, and this concentration is maintained until 24 months. Between 24 to 30 months there is a little decrease in the concentration of this compound. This change could be produced by a slight increase of DA metabolism with aging as suggested by the slight increases in DOPAC and HVA concentrations, the main metabolites of DA. Our results are in agreement with those of other authors who found a greater
337
ng/g
1 2 0 0 0
9000
6000
3000
:' ...
.: '•,
/
i
* DA
('.•
"+ . . . . . . . . . . t- . . . . . . . . . . . . . . . . . . . . . (¢"
h DOPAC
. . , HVA
10 20 t ( m o n t h s )
Age in months DA 3-MT DOPAC HVA
3 7014.5 ± 800.2 ± 4071.5 ~ 966,2 ±
88 ,1 28,6 105.9 3 8 , 9 (4) (4) (4) (4)
8 12907,8 ± 9 3 2 . 9 ± 3 2 0 1 . 4 ± 984,5 ±
591 ,I 48,9 242,5 67.9
(4)** (4) (4)* (4)
12 1 2 5 3 4 . 5 ± 8 4 3 . 9 ± 2 9 4 3 . 3 ± 8 4 5 . 5 ± 181.1 13.0 100,9 23.7
(4) (4) (4) (4)
24 12654.2 ± 753.2 ± 3150.1 ± 726,5 ±
231.1 36,7 118.1 45,0
(4) (4) (4) (4)
30 9831.2 ± 851,7 ± 4543.8 ± 1206,9 ± 117 ,6 3 1 . 2 96 .0 5 2 . 9
(4) " * (4 ) ( 4 ) * * ( 4 ) * ~ * P < 0.05; ~* P < 0.01, as compared with previous value
Fig. 2. Concentration of D A and its metabolites in the striatum of rats during the aging period• Each value represents the mean_+ standard error of the levels of DA and its metabolites which are expressed in n g / g wet tissue. Numbers in parentheses indicate numbers of animals used.
age-related decrease in DA concentration or significant decline with age in the DA content of the caudate and putamen in humans (Carlsson and Winbland, 1976) as well as a 25% decrease in the neostriatum of mice up to 28 months of age (Finch, 1975). Similar results have been described by Joseph et al. (1978).
The DA content decreases and there is also a slight increase of 3-MT, which suggests that there is an increase in the DA release, because the concentration of 3-MT correlates with DA release (Wood et al., 1982). At the same time, there is also a slight increase in DOPAC and especially in HVA. This could be correlated with the data from previous studies carried out in patients with less severely impairing Parkinson (Bernheimer et al., 1973), and in monkeys with experimental nigral lesions, where caudate nucleus DA concentrations fall more markedly than those of HVA (Sharman et al., 1967). These authors suggest that when some dopamine-con- taining neurons are lost, there is a compensatory increase of activity of the survivors, so that the H V A / D A ratio increases in a similar way to that which takes place in this study where there is a change from 0.076 at 6 months of age to 0.122 at 30 months in the rat.
DA is not produced in striatum neurons, it comes principally from the substantia nigra and the ventral segmental area of Tsai (DahlstriSm and Fuxe, 1964; Llamas and Reinoso-Su~rez, 1969; Ungerstedt, 1971), and some could also come from the raphe (Kellar et al., 1977; Jacobs et al., 1978; Szabo, 1980; Wiklund, 1981). Therefore, the variation of DA concentration could represent the variation of this component in the substantia nigra and the raphe, being linked either to the maturing of these dopaminergic fibers or to a neurotransmitter interaction. The
3 3 8
ng/g
400
300
200
l O 0
.t, .: -.
/
.. . " '., ..
NA
.... + ..................... + .......... + . ,
" . . . . . . . . * NMN
i
l'O - - ' 2 ' 0 t ( m o n t h s )
Age in months NA R'RN M~
3 306.5 ± 68,2 ± 60,6 ±
10.7 i .7 0.9
(4) ( 4 ) (41
6 4 4 9 . 3 ± 5 8 . 4 ± 8 1 . 4 ± 17.7 2 ,8 6 .5
(41 ' ~ ( 4 ) ~ ( 4 ) *
12 244 .1 ± 23 .1 ± 130 .3 ± 9.6 i ,4 12.7
(4 ) ~" ( 4 1 ' ( 4 1 '
24 3 9 7 . 9 ± 28 .2 ± 136 ,0 ± 42 ,1 2 .0 13.9 (41' (41, (4 )
30 434.6 ± 43.2 ± 128.7 ± 5 9 . 3 2 ,6 11,7 (4) (4) ~ (41
I p < 0.05; ~* P < 0,01. as compared with previous value
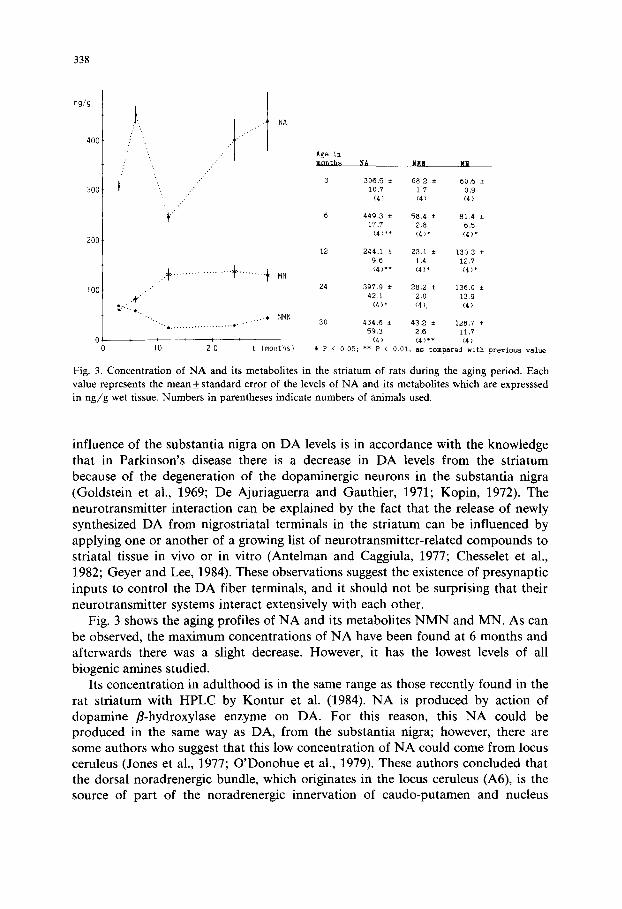
Fig. 3. Concentration of N A and its metabolites in the striatum of rats during the aging period. Each value represents the mean_+ standard error of the levels of N A and its metabolites which are expresssed in n g / g wet tissue. Numbers in parentheses indicate numbers of animals used.
influence of the substantia nigra on DA levels is in accordance with the knowledge that in Parkinson's disease there is a decrease in DA levels from the striatum because of the degeneration of the dopaminergic neurons in the substantia nigra (Goldstein et al., 1969; De Ajuriaguerra and Gauthier, 1971; Kopin, 1972). The neurotransmitter interaction can be explained by the fact that the release of newly synthesized DA from nigrostriatal terminals in the striatum can be influenced by applying one or another of a growing list of neurotransmitter-related compounds to striatal tissue in vivo or in vitro (Antelman and Caggiula, 1977; Chesselet et al., 1982; Geyer and Lee, 1984). These observations suggest the existence of presynaptic inputs to control the DA fiber terminals, and it should not be surprising that their neurotransmitter systems interact extensively with each other.
Fig. 3 shows the aging profiles of NA and its metabolites N M N and MN. As can be observed, the maximum concentrations of NA have been found at 6 months and afterwards there was a slight decrease. However, it has the lowest levels of all biogenic amines studied.
Its concentration in adulthood is in the same range as those recently found in the rat striatum with HPLC by Kontur et al. (1984). NA is produced by action of dopamine fl-hydroxylase enzyme on DA. For this reason, this NA could be produced in the same way as DA, from the substantia nigra; however, there are some authors who suggest that this low concentration of NA could come from locus ceruleus (Jones et al., 1977; O'Donohue et al., 1979). These authors concluded that the dorsal noradrenergic bundle, which originates in the locus ceruleus (A6), is the source of part of the noradrenergic innervation of caudo-putamen and nucleus
339
2000
lO00
..""t
Tryp
* " " * " . . . ..i '. '. '. '---I 5-HIAA " " - . . ~ . . 5-HT
. . . -, ® . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . * 5-HTP , , i q ,
lO 20 t (months)
Age In
mo]l~hs Tryp..__ 5-HTP 5-HT 5-HIAA
3 2905,9 ± i06,3 ± 6209 ± 484,6 ±
83.7 ]3,3 ~,9 37.1
(4) (4) (4) <4)
6 3035.6 ± 115.2 ± 702.2 ± 520.6 ±
208.5 2.0 45,6 35.0
(4) (4) (4) (4)
12 2825.6 ± 94,6 ± 6 2 1 , 2 ± 287,4 ±
173.6 4,6 8,5 11.7
(4) (4) ° (4) (4) ~
24 2655,5 ± 97.6 ± 620,3 ± 381,4 ± 101.3 2.0 18,5 8,5
(4) <4) (4) <4)**
30 3484,8 ± I~5,8 ± 411.8 ± 447.5 ±
258.5 8.2 16.3 32.1
(4)* (4) (4) *~ (4)
* P < 0.05; ~*P < 0.01. as compared with prevlous value
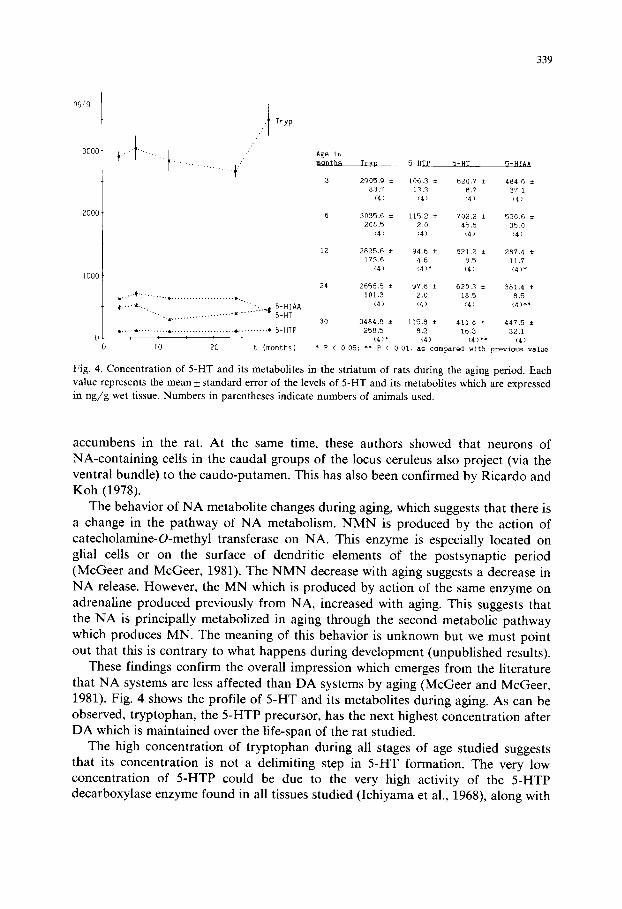
Fig. 4. Concentration of 5-HT and its metabolites in the striatum of rats during the aging period. Each value represents the mean_+ standard error of the levels of 5-HT and its metabolites which are expressed in n g / g wet tissue. Numbers in parentheses indicate numbers of animals used.
accumbens in the rat. At the same time, these authors showed that neurons of NA-containing cells in the caudal groups of the locus ceruleus also project (via the ventral bundle) to the caudo-putamen. This has also been confirmed by Ricardo and Koh (1978).
The behavior of NA metabolite changes during aging, which suggests that there is a change in the pathway of NA metabolism. NMN is produced by the action of catecholamine-O-methyl transferase on NA. This enzyme is especially located on glial cells or on the surface of dendritic elements of the postsynaptic period (McGeer and McGeer, 1981). The NMN decrease with aging suggests a decrease in NA release. However, the MN which is produced by action of the same enzyme on adrenaline produced previously from NA, increased with aging. This suggests that the NA is principally metabolized in aging through the second metabolic pathway which produces MN. The meaning of this behavior is unknown but we must point out that this is contrary to what happens during development (unpublished results).
These findings confirm the overall impression which emerges from the literature that NA systems are less affected than DA systems by aging (McGeer and McGeer, 1981). Fig. 4 shows the profile of 5-HT and its metabolites during aging. As can be observed, tryptophan, the 5-HTP precursor, has the next highest concentration after DA which is maintained over the life-span of the rat studied.
The high concentration of tryptophan during all stages of age studied suggests that its concentration is not a delimiting step in 5-HT formation. The very low concentration of 5-HTP could be due to the very high activity of the 5-HTP decarboxylase enzyme found in all tissues studied (Ichiyama et al., 1968), along with

340
the low activity of the tryptophan hydroxylase enzyme that has been suggested to be the limiting and regulatory step of 5-HT production (Lovenberg et al., 1968; Kaufman, 1974). This 5-HT may not be produced in striatum neurons, since it comes from serotoninergic fibers of the raphe nuclei (Anden et al., 1966; Ungers- tedt, 1971) and also from the substantia nigra (Streit, 1980; Steinbusch, 1981). The variation of its concentration must be related to these nuclei, where it is produced.
5-HTP-like 5-HIAA does not change during this period. However, 5-HT, as DA, decreased between 24 and 30 months.
There are some authors who have pointed out the importance of the ratio of DA to 5-HT in the striatum since motor disturbances in experimental animals may result from a modification of this ratio. Similar effects appear with either a relative increase in DA or a decrease in 5-HT (Hassler and Bak, 1969). In our results the D A / 5 - H T ratio does not change during the period studied.
The literature about the 5-HT concentration during aging is controversial, since Robinson et al. (1972) reported a small but statistically significant decline with age of 5-HT in the caudate nucleus. But the same group (Nies et al., 1973) later reported no change in concentration level of 5-HT from 25 to over 70 years of age. The decrease of 5-HT levels with age could merely reflect increased MAO activity, as corresponds to the enzyme which metabolizes serotonin.
All these results suggest that in aging the change in biogenic amine levels in the striatum of the rat is not significant. This could be in agreement with a recent paper that reported only a minor change in the numbers of neurons during aging in some areas of the CNS, e.g. the geniculate body (Satorre et al., 1985). The change seems to be very significant in some diseases related with aging, but not in 'normal ' aging. However, in spite of this lack of change in the levels of biogenic amines, during aging there could be a change in other important related parameters that are under investigation, in e.g. turnover rates, receptor action capacity, etc.
Acknowledgements
M. Santiago is a recipient of a predoctoral fellowship from I.I.E. del F.I.S.S. This work was supported by Ayuda a la Investigaci6n de la Junta de Andalucia.
References
Anden, N.E., DahlstrSm, A., Fuxe, K., Larsson, K., Olson, L. and Ungerstedt, U. (1966): Ascending monoamine neurons to the telencephalon and diencephalon. Acta Physiol. Scand., 67, 313-326.
Antelman, S.M. and Caggiula, A.R. (1977): Norepinephrine-dopamine interactions and behaviour: a new hypothesis of stress-related interactions between norepinephrine and dopamine is proposed. Science, 195, 646-653.
Bernheimer, H., Birkmayer, W., Hornykiewicz, O., Jellinger, K. and Settelberger, F. (1973): Brain dopamine and the syndrome of Parkinson and Huntington: clinical, morphological and neurochem- ical correlations. J. Neurol. Sci., 20, 415-425.

341
Bird, E.D., Spokes, E.G.S. and Iversen, L.L. (1979): Increased dopamine concentration in limbic area of brain from patients dying with schizophrenia. Brain, 102, 347-360.
Carlsson, A. and Winbland, B. (1976): Influence of age and time interval death and autopsy on dopamine and 3-methoxytyramine levels in human basal ganglia. J. Neural Transmission, 38, 271-276.
Chesselet, M.F., Cheramy, A., Reisine, T., Lubetzki, C. and Glowinski, J. (1982): Presynaptic regulation of striatal dopamine release. In vivo and in vitro studies. J. Physiol., 78, 420-425.
Dahlstr~Sm, A. and Fuxe, K. (1964): Evidence for the existence of monoamine containing neurons in the central nervous system. I. Demonstration of monoamines in the cell bodies of brain stem neurons. Acta Physiol. Scand., 62, 1-55.
De Ajuriaguerra, J. and Gauthier, G. eds. (1971): Monoamines. noyaux gris centraux, et syndrome de Parkinson. Georg, Geneva; and Masson, Paris.
Finch, C.E. (1975): Ageing and the regulation of hormones: a view in October 1874. In: Advances in Experimental Medicine and Biology, Vol. 61, Suppl. Explorations in Ageing, pp. 229-238. Editors: V.J. Cristofalo, J. Roberts and R.C. Adelman. Plenum Press, New York, London.
Geyer, M.A. and Lee, E.H.Y. (1984): Effects of clonidine, piperoxane and locus coeruleus lesion on the serotonergic and dopaminergic system in raphe and caudate nucleus. Biochem. Pharmacol., 3399-3404.
Goldstein, M., Anagnoste, B., Battista, A.F., Owen, W.S. and Nakatani, S. (1969): Studies of amines in the striatum in monkeys with nigral lesions. J. Neurochem., 16, 645.
Hassler, R. and Bak, I.J. (1969): Unbalanced rations of strial dopamine and serotonin after experimental interruption of strionigral connections. In: Third Symposium on Parkinson's disease, pp. 29-37. Editors: F.J. Gillingham and I.M.L. Donaldson. Livingston, Edinburgh, London.
Hornykiewicz, H. (1976): Neurohumoral interactions and basal function and dysfunction. In: The Basal Ganglia, pp. 269-280. Editor: M.D. Yahr, Raven Press, New York.
Ichiyama, A., Nakamura, S., Nishizuka, Y. and Hayaishi, O. (1968): Tryptophan-5-hydroxylase in mammalian brain. In: Advances in Pharmacology, Vol. 6A, pp. 5-17. Editors: S. Garattini and P.A. Shore. Academic Press, New York.
Jacobs, B.L., Foote, S.L. and Bloom, F.E. (1978): Differential projections of neurons within the dorsal raphe nucleus of the rat: a horseradish peroxidase (HRP) study. Brain Res., 147, 149-153.
Jones, B.E., Halaris, A.E., Mclljany, M. and Moore, R.Y. (1977): Ascending projections of the locus coeruleus in the rat. I. Axonal transport in central noradrenaline neurons. Brain Res., 127, 1-21.
Joseph, J.A., Berger, R.E., Engel, B.T. and Roth, G.S. (1978): Age-related changes in the nigrostriatum: a behavioral and biochemical analysis. J. Gerontol., 33,643-649.
Kasamatsu, T. and Pettigrew, J.D. (1979): Preservation of binocularly after monocular deprivation in the striate cortex of kittens treated with 6-hydroxydopamine. J. Comp. Neurol., 185, 139-162.
Kaufman, S. (1974): Properties of pterin-dependent aromatic amino acid hydroxylases. In: Aromatic Amino Acids in the Brain, pp. 85-108. Editors: G.E.W. Wolstenholme and D.W. Fitzsimons. Elsevier, Amsterdam.
Kellar, K.J., Brown, P.A., Madrid, J., Berstein, M., Vernikos-Danellis, J. and Mehler, W.R. (1977): Origins of serotonin of forebrain structures. Exp. Neurol., 56, 52-62.
Kontur, P., Dawson, R. and Monjan, A. (1984): Manipulation of mobile phase parameters for the HPLC separation of endogenous monoamines in the rat brain tissue. J. Neurosci. Meth., 11, 5-18.
Kopin, I.J. (1972): Neurotransmitters, Vol. 50. Research Publications of the Association for Research in Nervous and Mental Diseases. A.R.N.M.D., New York.
Lander, J., Wallace, J., Krebs, H., Petrusz, P. and McCarthy, K. (1983): In vivo and in vitro development of serotoninergic neurons. Brain Res. Bull., 9, 605-625.
Llamas, A. and Reinoso-Suhrez, F. (1969): Projections of substantia nigra and ventral segmental mesencephalic area. In: Third Symposium on Parkinson's disease, pp. 82-87. Editors: F.J. Gil- lingham and I.M.L. Donaldson. Livingston, Edinburgh, London.
Lovenberg, W., Jequier, E. and Sjoerdsma, A. (1968): Tryptophan hydroxylation in mammalian systems. In: Advances in Pharmacology, Vol. 6A, pp. 21-36. Editors: S. Garattini and P.A. Shore. Academic Press, New York.
McGeer, P.L. and McGeer, E.Q. (1981): Neurotransmitter in the ageing brain. In: The Molecular Basis of Neuropathology, pp. 631-648. Editors: A.N. Davinson and R.H.S. Thompson. Edward Arnold Ltd., London.

342
Nies, A., Robinson, D.S., Davis, J.M. and Ravaris, L. (1973): Changes in monoamine oxidase with aging. In: Psychopharmacology with Aging, pp. 41-54. Editors: C. Eisdorfer and W.E. Fann. Plenum Press, New York.
O'Donohue, T.L., Crowley, W.R. and Jacobowitz, D.M. (1979): Biochemical mapping of noradrenergic ventral bundle projection sites: evidence for a noradrenergic-dopaminergic interaction. Brain Res., 172, 87-100.
Pileblad, E., Fornstedt, B., Clark, O. and Carlsson, A. (1985): Acute effects of 1-methyl-4-phenyl-l,2,3,6- tetrahydropyridine on dopamine metabolism in mouse and rat striatum. J. Pharm. Pharmacol., 37, 707-712.
Ricardo, J.A. and Koh, E.T. (1978): Anatomical evidence of direct projections from the nucleus of the solitary tract to the hypothalamus, amygdala, and other forebrain structures in the rat. Brain Res., 153, 1-26.
Robinson, D.S, Nies, A., Davis, J.M., Bunney, W.E., Colburn, R.W., Bourne, H.R., Shaw, D.M. and Coppen, A.J. (1972): Aging monoamines and monoamine oxidase levels. Lancet 290-291.
Satorre, J., Cano, J. and Reinoso-Su~trez, F. (1985): Stability of the neuronal population of the dorsal lateral geniculate nucleus (LGNd) of aged rats. Brain Res., 339, 375-377.
Sharman, D.F., Poirier, L.J., Murphy, Q.F. and Sourkes, T.L. (1967): Homovanillic acid and dihydroxy- phenylacetic acid of the striatum of monkeys with brain lesions. Can. J. Physiol., 45, 57-62.
Steinbusch, H.W.M. (1981): Distribution of serotonin-immunoreactivity in the central nervous system of the rat cell bodies and terminals. Neuroscience, 6, 557-618.
Streit, P. (1980): Selective retrograde labeling indicating the transmitter of neuronal pathways. J. Comp. Neurol., 191,429-463.
Szabo, J. (1980): Distribution of striatal afferents from the mesencephalon in the cat. Brain Res., 188, 3-21.
Timiras, P.S. (1978): Biological perspectives on aging: in research of a masterplan. Ann. Sci., 66, 605-613.
Timiras, P.S. and Bignami, A. (1976): Pathophysiology of the aging brain. In: Special Review of Experimental Aging Research, Progress in Biology, pp. 351-378. Editors: M.F. Elias, B.E. Eleftherion and P.K. Elias, EAR, Ina., Bar Harbour.
Timiras, P.S., Segall, P.E. and Walker, R.F. (1979): Physiological aging in the central nervous system: perspectives on 'interventive' gerontology. In: Aging - - its Chemistry, Proceeding of the Third Arnold O. Bukman Conference in Clinical Chemistry, pp. 46-63. Editor A.A. Dietz. American Association of Clinical Chemistry, Inc. Washington, D.C.
Ungerstedt, U. (1971): Stereotaxic mapping of the monoamine pathways in the rat brain. Acta Physiol. Scand., 197, 1-48.
Walker, R.F. and Timiras, P.S. (1981): Pacemaker insufficiency and the onset of aging. In: Cellular Pacemaker II, pp. 396-425. Editor: D. Carpenter. Wiley Interscience, New York.
Wiklund, L., Leger, L. and Persson, M. (1981): Monoamine cell distribution in the cat brainstem: a fluorescence histochemical study with quantification of indolaminergic and locus coeruleus cell groups. J. Comp. Neurol., 203, 613-648.
Wood, P.L., Nair, N.P.V. and Bozarth, W. (1982): Striatal 3-methoxytyramine as an index of dopamine release: effects of electrical stimulation. Neurosci. Lett., 32, 291-294.
Related Documents











