High-performance liquid chromatography ultraviolet assay for human erythrocytic catalase activity by measuring glutathione as o-phthalaldehyde derivative Anke Böhmer, Jens Jordan, Dimitrios Tsikas ⇑ Institute of Clinical Pharmacology, Hannover Medical School, 30625 Hannover, Germany article info Article history: Received 28 September 2010 Received in revised form 4 November 2010 Accepted 11 November 2010 Available online 19 November 2010 Keywords: Azide Enzyme assay Erythrocytes Glutathione HPLC Inhibition Nitrite abstract The most frequently used catalase (CAT) activity assay is based on the spectrophotometric measurement of hydrogen peroxide (H 2 O 2 ) absorbance decrease at 240 nm. Here we report an alternative high-perfor- mance liquid chromatography (HPLC) assay for human erythrocytic CAT (heCAT) activity measurement based on glutathione (GSH) analysis as a highly stable, H 2 O 2 -insensitive o-phthalaldehyde (OPA) deriva- tive. The method was developed and validated using an isolated heCAT in phosphate-buffered saline at pH 7.4 and was applied to measure CAT activity in lysed human erythrocytes. heCAT activity was mea- sured at initial concentrations of 5 nM for heCAT, 5 mM for H 2 O 2 , and 10 mM for GSH, and the incubation time was 10 min. Nitrite (NO 2 ) was found to be an uncompetitive inhibitor of heCAT activity (IC 50 =9 lM) and of CAT activity in hemolysate (IC 50 750 lM). Nitrate (NO 3 ) at concentrations up to 100 lM did not inhibit heCAT activity. Azide (N 3 ) was found to be a very strong inhibitor of the heCAT (IC 50 = 0.2 nM) but a relatively weak CAT inhibitor (IC 50 10 lM) in human hemolysates. The novel CAT activity assay works under redox conditions that more closely resemble those prevailing in cells and allows high-throughput analysis despite the required HPLC step. Ó 2010 Elsevier Inc. All rights reserved. Catalase (CAT), 1 H 2 O 2 :H 2 O 2 oxidoreductase (EC 1.11.1.6), is ubiq- uitous in nature. This enzyme catalyzes the conversion of hydrogen peroxide (H 2 O 2 ) to molecular oxygen and water via two steps with comparable reaction rate constants k of 1.7 10 M 1 s 1 and 2.6 10 M 1 s 1 , respectively [1,2] (see Reactions (1a) and (1b) be- low). H 2 O 2 is produced by numerous reactions, including the sponta- neous and superoxide dismutase (SOD, EC 1.15.1.1)-catalyzed dismutation of the superoxide radical anion (O 2 ), which is also ubiquitous in nature. Because O 2 and H 2 O 2 are reactive oxygen spe- cies and potently damage tissue, their final conversion by SOD and CAT enzymes to harmless molecular oxygen and water represents a powerful antioxidative system of living organisms. Given this extraordinary importance, measurement of SOD and CAT activity in certain conditions is a valuable measure to assess the status of this vital defense system. Accordingly, different enzyme assays have been reported for many decades for SOD and CAT activity measurement [1]. The reaction product dioxygen (O 2 ) has been frequently used to assess activity of these enzymes in biological samples. Measurement of the CAT substrate H 2 O 2 is obviously the most frequently used CAT activity assay. The basic principle of this assay is detection of ultravi- olet (UV) absorbance decrease at 240 nm (e = 39.4 M 1 cm 1 ) due to the CAT-catalyzed removal of H 2 O 2 that is added at lower millimolar concentrations, usually at 10 mM. Despite the simplicity and rapidity of the spectrophotometric assay, UV absorbance detection at 240 nm may be fraught with problems due to interferences by endogenous and exogenous UV-absorbing components present in or added to the biological sample. In our opinion, a more important reason for alternative CAT assays is the fact that the redox conditions used in current assays are far from those prevailing in cells, including eryth- rocytes and hepatocytes that contain glutathione (GSH) at concentra- tions in the range of 1–10 mM. CAT—Fe 3þ þ H 2 O 2 ! compound I þ H 2 O ð1aÞ compound I þ H 2 O 2 ! CAT—Fe 3þ þ H 2 O þ O 2 ð1bÞ In the current article, we report a new high-performance liquid chromatography (HPLC)-based human erythrocytic CAT (heCAT) activity assay that runs in the presence of GSH at concentrations 0003-2697/$ - see front matter Ó 2010 Elsevier Inc. All rights reserved. doi:10.1016/j.ab.2010.11.026 ⇑ Corresponding author. Fax: +49 511 532 2750. E-mail address: [email protected] (D. Tsikas). 1 Abbreviations used: CAT, catalase; H 2 O 2 , hydrogen peroxide; SOD, superoxide dismutase; O 2 , superoxide radical anion; O 2 , dioxygen; UV, ultraviolet; GSH, glutathione; HPLC, high-performance liquid chromatography; heCAT, human eryth- rocytic CAT; OPA, o-phthalaldehyde; GS–OPA, GSH as OPA derivative; CAS 1609, 4-hydroxymethyl-furoxan-3-carboxyamide; ODQ, 1H-[1,2,4]oxadiazolo[4,3-a]qui- noxalin-1-one; GSNO, S-nitrosoglutathione; PBS, phosphate-buffered saline; GSSG, glutathione disulfide; GSOH, glutathione sulfenic acid; EDTA, ethylenediaminetetra- acetic acid; NO, nitric oxide; GC–MS, gas chromatography–mass spectrometry; GTN; glycerol trinitrate. Analytical Biochemistry 410 (2011) 296–303 Contents lists available at ScienceDirect Analytical Biochemistry journal homepage: www.elsevier.com/locate/yabio

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Analytical Biochemistry 410 (2011) 296–303
Contents lists available at ScienceDirect
Analytical Biochemistry
journal homepage: www.elsevier .com/locate /yabio
High-performance liquid chromatography ultraviolet assay for humanerythrocytic catalase activity by measuring glutathione as o-phthalaldehydederivative
Anke Böhmer, Jens Jordan, Dimitrios Tsikas ⇑Institute of Clinical Pharmacology, Hannover Medical School, 30625 Hannover, Germany
a r t i c l e i n f o a b s t r a c t
Article history:Received 28 September 2010Received in revised form 4 November 2010Accepted 11 November 2010Available online 19 November 2010
Keywords:AzideEnzyme assayErythrocytesGlutathioneHPLCInhibitionNitrite
0003-2697/$ - see front matter � 2010 Elsevier Inc. Adoi:10.1016/j.ab.2010.11.026
⇑ Corresponding author. Fax: +49 511 532 2750.E-mail address: [email protected] (
1 Abbreviations used: CAT, catalase; H2O2, hydrogedismutase; O2
��, superoxide radical anion; O2, dioxglutathione; HPLC, high-performance liquid chromatorocytic CAT; OPA, o-phthalaldehyde; GS–OPA, GSH a4-hydroxymethyl-furoxan-3-carboxyamide; ODQ, 1Hnoxalin-1-one; GSNO, S-nitrosoglutathione; PBS, phosglutathione disulfide; GSOH, glutathione sulfenic acid;acetic acid; NO, nitric oxide; GC–MS, gas chromatograpglycerol trinitrate.
The most frequently used catalase (CAT) activity assay is based on the spectrophotometric measurementof hydrogen peroxide (H2O2) absorbance decrease at 240 nm. Here we report an alternative high-perfor-mance liquid chromatography (HPLC) assay for human erythrocytic CAT (heCAT) activity measurementbased on glutathione (GSH) analysis as a highly stable, H2O2-insensitive o-phthalaldehyde (OPA) deriva-tive. The method was developed and validated using an isolated heCAT in phosphate-buffered saline atpH 7.4 and was applied to measure CAT activity in lysed human erythrocytes. heCAT activity was mea-sured at initial concentrations of 5 nM for heCAT, 5 mM for H2O2, and 10 mM for GSH, and the incubationtime was 10 min. Nitrite (NO2
�) was found to be an uncompetitive inhibitor of heCAT activity(IC50 = 9 lM) and of CAT activity in hemolysate (IC50 � 750 lM). Nitrate (NO3
�) at concentrations up to100 lM did not inhibit heCAT activity. Azide (N3
�) was found to be a very strong inhibitor of the heCAT(IC50 = 0.2 nM) but a relatively weak CAT inhibitor (IC50 � 10 lM) in human hemolysates. The novel CATactivity assay works under redox conditions that more closely resemble those prevailing in cells andallows high-throughput analysis despite the required HPLC step.
� 2010 Elsevier Inc. All rights reserved.
Catalase (CAT),1 H2O2:H2O2 oxidoreductase (EC 1.11.1.6), is ubiq-uitous in nature. This enzyme catalyzes the conversion of hydrogenperoxide (H2O2) to molecular oxygen and water via two steps withcomparable reaction rate constants k of 1.7 � 10 M�1 s�1 and2.6 � 10 M�1 s�1, respectively [1,2] (see Reactions (1a) and (1b) be-low). H2O2 is produced by numerous reactions, including the sponta-neous and superoxide dismutase (SOD, EC 1.15.1.1)-catalyzeddismutation of the superoxide radical anion (O2
��), which is alsoubiquitous in nature. Because O2
�� and H2O2 are reactive oxygen spe-cies and potently damage tissue, their final conversion by SOD andCAT enzymes to harmless molecular oxygen and water represents apowerful antioxidative system of living organisms. Given thisextraordinary importance, measurement of SOD and CAT activity incertain conditions is a valuable measure to assess the status of this
ll rights reserved.
D. Tsikas).n peroxide; SOD, superoxideygen; UV, ultraviolet; GSH,
graphy; heCAT, human eryth-s OPA derivative; CAS 1609,-[1,2,4]oxadiazolo[4,3-a]qui-phate-buffered saline; GSSG,EDTA, ethylenediaminetetra-hy–mass spectrometry; GTN;
vital defense system. Accordingly, different enzyme assays have beenreported for many decades for SOD and CAT activity measurement[1]. The reaction product dioxygen (O2) has been frequently used toassess activity of these enzymes in biological samples. Measurementof the CAT substrate H2O2 is obviously the most frequently used CATactivity assay. The basic principle of this assay is detection of ultravi-olet (UV) absorbance decrease at 240 nm (e = 39.4 M�1 cm�1) due tothe CAT-catalyzed removal of H2O2 that is added at lower millimolarconcentrations, usually at 10 mM. Despite the simplicity and rapidityof the spectrophotometric assay, UV absorbance detection at 240 nmmay be fraught with problems due to interferences by endogenousand exogenous UV-absorbing components present in or added tothe biological sample. In our opinion, a more important reason foralternative CAT assays is the fact that the redox conditions used incurrent assays are far from those prevailing in cells, including eryth-rocytes and hepatocytes that contain glutathione (GSH) at concentra-tions in the range of 1–10 mM.
CAT—Fe3þ þH2O2 ! compound IþH2O ð1aÞ
compound IþH2O2 ! CAT—Fe3þ þH2Oþ O2 ð1bÞ
In the current article, we report a new high-performance liquidchromatography (HPLC)-based human erythrocytic CAT (heCAT)activity assay that runs in the presence of GSH at concentrations
0 10 20 300
2
4
6
8
10
5
0 °C
25 °C
Time (min)
[GSH
] r (m
M)
HPLC–UV assay for heCAT activity / A. Böhmer et al. / Anal. Biochem. 410 (2011) 296–303 297
found in the cytosol of many types of cells. Remaining GSH (seeReactions (2a–c) below) was measured by HPLC with UV absor-bance detection as o-phthalaldehyde (OPA) derivative (GS–OPA)(see Reaction (3) below) [3,4], and its concentration was used asa measure of CAT activity. We validated this HPLC assay using he-CAT and applied it to measure CAT activity in lysed human eryth-rocytes. By this assay, the effects of sodium azide and sodiumnitrite on isolated heCAT and in hemolysate were investigated. Pre-vious findings on CAT inhibition by nitrite reported by Titov andPetrenko, who used a calorimetric CAT assay [5], were confirmed.We report here, for the first time, that nitrite is an uncompetitiveinhibitor of heCAT.
GSHþH2O2 ! GSOHþH2O ð2aÞ
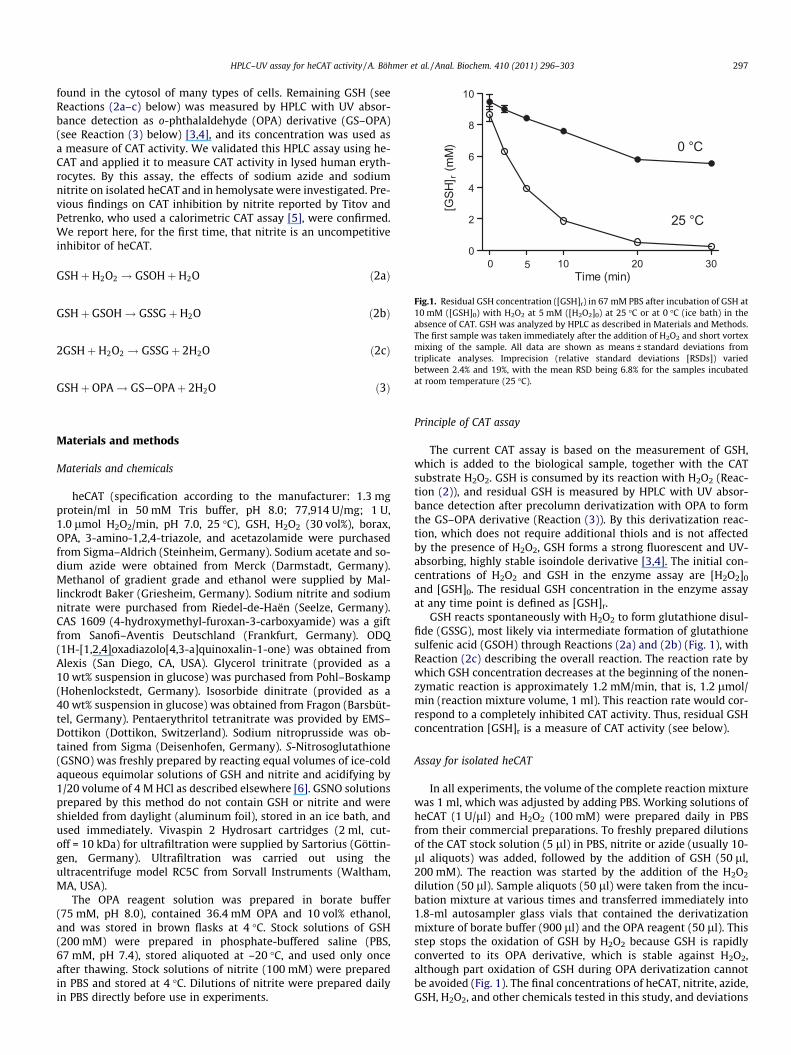
Fig.1. Residual GSH concentration ([GSH]r) in 67 mM PBS after incubation of GSH at10 mM ([GSH]0) with H2O2 at 5 mM ([H2O2]0) at 25 �C or at 0 �C (ice bath) in the
GSHþ GSOH! GSSGþH2O ð2bÞ absence of CAT. GSH was analyzed by HPLC as described in Materials and Methods.The first sample was taken immediately after the addition of H2O2 and short vortexmixing of the sample. All data are shown as means ± standard deviations from 2GSHþH2O2 ! GSSGþ 2H2O ð2cÞ triplicate analyses. Imprecision (relative standard deviations [RSDs]) variedbetween 2.4% and 19%, with the mean RSD being 6.8% for the samples incubatedat room temperature (25 �C).GSHþ OPA! GS—OPAþ 2H2O ð3Þ
Materials and methods
Materials and chemicals
heCAT (specification according to the manufacturer: 1.3 mgprotein/ml in 50 mM Tris buffer, pH 8.0; 77,914 U/mg; 1 U,1.0 lmol H2O2/min, pH 7.0, 25 �C), GSH, H2O2 (30 vol%), borax,OPA, 3-amino-1,2,4-triazole, and acetazolamide were purchasedfrom Sigma–Aldrich (Steinheim, Germany). Sodium acetate and so-dium azide were obtained from Merck (Darmstadt, Germany).Methanol of gradient grade and ethanol were supplied by Mal-linckrodt Baker (Griesheim, Germany). Sodium nitrite and sodiumnitrate were purchased from Riedel-de-Haën (Seelze, Germany).CAS 1609 (4-hydroxymethyl-furoxan-3-carboxyamide) was a giftfrom Sanofi–Aventis Deutschland (Frankfurt, Germany). ODQ(1H-[1,2,4]oxadiazolo[4,3-a]quinoxalin-1-one) was obtained fromAlexis (San Diego, CA, USA). Glycerol trinitrate (provided as a10 wt% suspension in glucose) was purchased from Pohl–Boskamp(Hohenlockstedt, Germany). Isosorbide dinitrate (provided as a40 wt% suspension in glucose) was obtained from Fragon (Barsbüt-tel, Germany). Pentaerythritol tetranitrate was provided by EMS–Dottikon (Dottikon, Switzerland). Sodium nitroprusside was ob-tained from Sigma (Deisenhofen, Germany). S-Nitrosoglutathione(GSNO) was freshly prepared by reacting equal volumes of ice-coldaqueous equimolar solutions of GSH and nitrite and acidifying by1/20 volume of 4 M HCl as described elsewhere [6]. GSNO solutionsprepared by this method do not contain GSH or nitrite and wereshielded from daylight (aluminum foil), stored in an ice bath, andused immediately. Vivaspin 2 Hydrosart cartridges (2 ml, cut-off = 10 kDa) for ultrafiltration were supplied by Sartorius (Göttin-gen, Germany). Ultrafiltration was carried out using theultracentrifuge model RC5C from Sorvall Instruments (Waltham,MA, USA).
The OPA reagent solution was prepared in borate buffer(75 mM, pH 8.0), contained 36.4 mM OPA and 10 vol% ethanol,and was stored in brown flasks at 4 �C. Stock solutions of GSH(200 mM) were prepared in phosphate-buffered saline (PBS,67 mM, pH 7.4), stored aliquoted at –20 �C, and used only onceafter thawing. Stock solutions of nitrite (100 mM) were preparedin PBS and stored at 4 �C. Dilutions of nitrite were prepared dailyin PBS directly before use in experiments.
Principle of CAT assay
The current CAT assay is based on the measurement of GSH,which is added to the biological sample, together with the CATsubstrate H2O2. GSH is consumed by its reaction with H2O2 (Reac-tion (2)), and residual GSH is measured by HPLC with UV absor-bance detection after precolumn derivatization with OPA to formthe GS–OPA derivative (Reaction (3)). By this derivatization reac-tion, which does not require additional thiols and is not affectedby the presence of H2O2, GSH forms a strong fluorescent and UV-absorbing, highly stable isoindole derivative [3,4]. The initial con-centrations of H2O2 and GSH in the enzyme assay are [H2O2]0
and [GSH]0. The residual GSH concentration in the enzyme assayat any time point is defined as [GSH]r.
GSH reacts spontaneously with H2O2 to form glutathione disul-fide (GSSG), most likely via intermediate formation of glutathionesulfenic acid (GSOH) through Reactions (2a) and (2b) (Fig. 1), withReaction (2c) describing the overall reaction. The reaction rate bywhich GSH concentration decreases at the beginning of the nonen-zymatic reaction is approximately 1.2 mM/min, that is, 1.2 lmol/min (reaction mixture volume, 1 ml). This reaction rate would cor-respond to a completely inhibited CAT activity. Thus, residual GSHconcentration [GSH]r is a measure of CAT activity (see below).
Assay for isolated heCAT
In all experiments, the volume of the complete reaction mixturewas 1 ml, which was adjusted by adding PBS. Working solutions ofheCAT (1 U/ll) and H2O2 (100 mM) were prepared daily in PBSfrom their commercial preparations. To freshly prepared dilutionsof the CAT stock solution (5 ll) in PBS, nitrite or azide (usually 10-ll aliquots) was added, followed by the addition of GSH (50 ll,200 mM). The reaction was started by the addition of the H2O2
dilution (50 ll). Sample aliquots (50 ll) were taken from the incu-bation mixture at various times and transferred immediately into1.8-ml autosampler glass vials that contained the derivatizationmixture of borate buffer (900 ll) and the OPA reagent (50 ll). Thisstep stops the oxidation of GSH by H2O2 because GSH is rapidlyconverted to its OPA derivative, which is stable against H2O2,although part oxidation of GSH during OPA derivatization cannotbe avoided (Fig. 1). The final concentrations of heCAT, nitrite, azide,GSH, H2O2, and other chemicals tested in this study, and deviations

298 HPLC–UV assay for heCAT activity / A. Böhmer et al. / Anal. Biochem. 410 (2011) 296–303
of this procedure, are described in the respective experiments. Be-cause of the very low heCAT concentration in the derivatizationmixtures (<30 nM corresponding to <700 ng/ml), no additionalsteps for protein removal are necessary prior to HPLC analysis.
Assay for CAT in lysed erythrocytes
Blood was drawn from the antecubital vein of healthy volun-teers using ethylenediaminetetraacetic acid (EDTA)-containingmonovettes (Sarstedt, Nümbrecht, Germany). Immediately aftercollection, the blood was put in an ice bath. Blood, erythrocytes,and all GSH-containing samples were stored in an ice bath and inthe dark during handling to minimize GSH oxidation. Erythrocyteswere separated from plasma by centrifugation (5 min, 4 �C, 800g)followed by complete decantation of plasma and of a small portionof erythrocytes. In all experiments, erythrocytes were used un-washed. Erythrocytes were lysed by freezing for at least 30 minat �70 �C, followed by slow thawing on ice and by vortex mixingfor 1 min with the same volume of ice-cold distilled water.
PBS-diluted (1:10, v/v) hemolysates (100 ll) were further di-luted with PBS, resulting in a dilution of 1:200 (v/v). Then nitriteor azide was added from their aqueous solutions. After 30 s of incu-bation, GSH was added at a final concentration ([GSH]0) of 10 mM.The reaction was started with H2O2 at a final concentration([H2O2]0) of 5 mM. After 10 min of incubation, the reaction mixturewas immediately put into an ice bath. Subsequently, cell frag-ments, proteins, and enzymes were removed by centrifugation(5 min, 4 �C, 8000g) using Vivaspin cartridges. These steps mini-mize chemical and enzymatic reactions and yield a protein-freecolorless ultrafiltrate. An aliquot of 50 ll is immediately subjectedto OPA derivatization (borate buffer, 900 ll; OPA reagent, 50 ll) inHPLC autosampler glass vials at room temperature to stop the non-enzymatic oxidation of GSH by H2O2, resulting in a total dilution of1:4000 (v/v). Deviations from this procedure are described in therespective experiments. Despite the high dilution factor of thehemolysate, the ultrafiltration step is necessary to remove in par-ticular hemoglobin that is present in such concentrations thatwould affect HPLC analysis and damage the HPLC column.
For GSH quantification, calibration curves were prepared usingsolutions of GSH in PBS that were derivatized exactly as describedabove and analyzed at the same time as study samples. In addition,a freshly prepared sample of 1 mM GSH in PBS was derivatized ineach experiment and served as quality control.
HPLC conditions
The GS–OPA derivative was analyzed by HPLC as reported else-where [4]. HPLC analyses were performed on an apparatus fromDionex (Sunnyvale, CA, USA) and Gynkotek (Rösrath, Germany)consisting of a pump model UltiMate 3000 (Dionex) and a detectormodel UVD 170U (Gynkotek). Injection was carried out by aGynkotek automated sample injector model GINA 160. The chro-matographic column (125 � 4.0 mm i.d., packed with 3-lm parti-cle size Nucleodur C18 gravity) from Macherey–Nagel (Düren,Germany) was used. The column was located in a Gynkotekthermostated column compartment model Column-ThermostatJetstream 2 Plus at 20 �C. The HPLC system was controlled andthe data were managed by Chromeleon software (version 6.6).The mobile phase consisting of methanol/water (30:70, v/v) con-tained 150 mM sodium acetate and had a pH value of 7.0 adjustedby the addition of acetic acid. Isocratic runs were performed at aflow rate of 0.5 ml/min. The retention time of the GS–OPA deriva-tive was approximately 3.4 min. If not otherwise specified, theinjection volume was 20 ll and the running time was 10 min.The UV–visible detector was set at 338 nm. The area of the peaks
in units of mAU �min in UV absorbance detection was used forcalculation.
Statistics
Data calculation and preparation of figures were carried outusing GraphPad Prism 5 from GraphPad Software (La Jolla, CA,USA).
Results and discussion
Assaying CAT activity by HPLC with UV absorbance detection
Recently, we found that the addition of nitrite to hemolysatesfrom human erythrocytes at millimolar concentrations induces ni-trite concentration-dependent decreases in erythrocytic GSH con-centrations and increases in methemoglobin formation [4]. Thisstudy also suggested additional nitrite-induced oxidative modifica-tions of hemoglobin, presumably formation of mixed disulfides ofhemoglobin with GSH [4], the most abundant low-molecular-massthiol in erythrocytes [2]. Because nitrite is an inhibitor of CAT [5,7],we cannot exclude that nitrite-induced oxidation of GSH andhemoglobin may also be due to inhibition of CAT activity that isabundantly present in human erythrocytes [1,2]. To address this is-sue under conditions resembling those in intact and lysed erythro-cytes, we were interested in a CAT activity assay that can run in thepresence of millimolar concentrations of GSH as they occur in thecytosol of many types of cells. Because GSH spontaneously reactswith H2O2 at neutral pH to produce GSSG and water (see Reaction(2) and Fig. 1), it was assumed that the concentration of residualGSH in the sample would be a measure of CAT activity. Therefore,we tested the possibility of developing an alternative CAT assay bymeasuring GSH instead of H2O2 or O2. There are numerous analyt-ical methods available for GSH measurement in biological samples(reviewed in Refs. [8,9]). Recently, we found that erythrocytic GSHcan easily and accurately be determined in ultrafiltrates of lysedhuman erythrocytes by HPLC with UV absorbance and/or fluores-cence detection [4]. In the current work, we used this HPLC methodto measure erythrocytic CAT activity, and we demonstrate the util-ity of this assay to investigate effects of inhibitors such as azide andnitrite on CAT activity (Fig. 2). There is no doubt that the currentCAT assay is considerably more time-consuming and laboriousthan the common spectrophotometric assay or other electrochem-ical and calorimetric CAT assays [1,2,5]. However, the assay we de-scribe here may be useful for biochemical investigations underconditions closely related to those prevailing in erythrocytes andpresumably in other types of cells that are rich in CAT activity suchas liver and kidney cells. Despite the time-consuming HPLC step,the stability of the GS–OPA derivative allows high-throughput per-formance of the current CAT activity assay (i.e., 144 samples/day).
Characterization of HPLC CAT assay
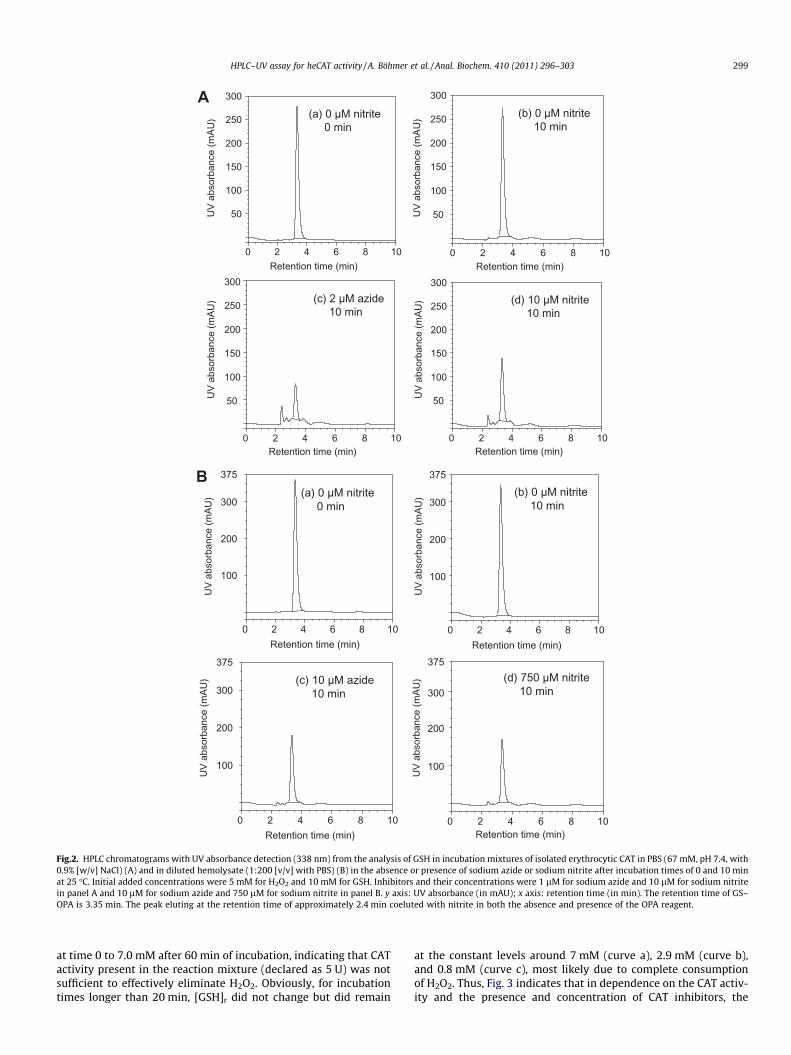
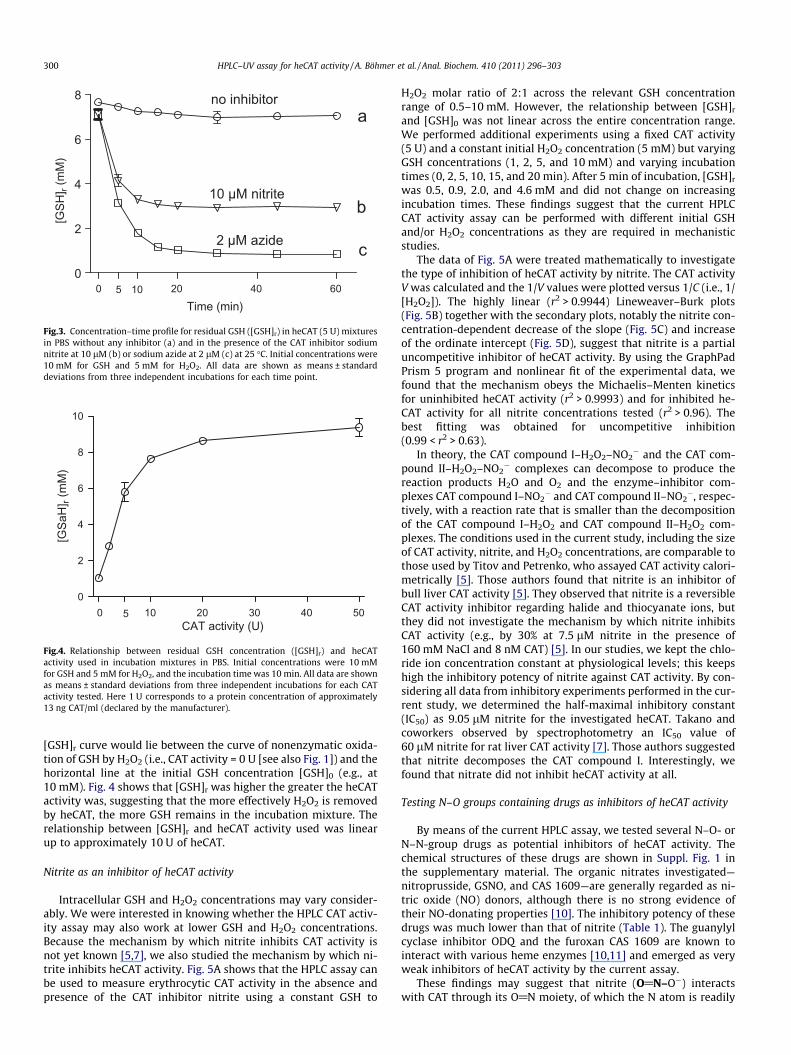
Fig. 3 shows the time course of residual GSH concentration([GSH]r) in incubation mixtures of isolated heCAT in PBS in the ab-sence of any inhibitor (curve a) and in the presence of azide (2 lM,curve c) or nitrite (10 lM, curve b). The curve for azide resemblesthat observed from spontaneous oxidation of GSH with H2O2 in theabsence of CAT activity (Fig. 1), suggesting that azide at 2 lMinhibits heCAT activity virtually completely. In curve b for nitrite,[GSH]r is higher as compared with azide (curve c) but lower incomparison with uninhibited heCAT shown by curve a. These find-ings indicate that nitrite is a weak inhibitor and azide is a stronginhibitor of heCAT (see next section). It is noteworthy that in theabsence of inhibitors, GSH concentration decreased from 7.6 mM
0 2 4 6 8 10
50
100
150
200
250
300
Retention time (min)
UV
abso
rban
ce (m
AU)
UV
abso
rban
ce (m
AU)
UV
abso
rban
ce (m
AU)
UV
abso
rban
ce (m
AU)
UV
abso
rban
ce (m
AU)
UV
abso
rban
ce (m
AU)
UV
abso
rban
ce (m
AU)
UV
abso
rban
ce (m
AU)(a) 0 µM nitrite
0 min
0 2 4 6 8 10
50
100
150
200
250
300
Retention time (min)
(b) 0 µM nitrite10 min
Retention time (min)0 2 4 6 8 10
50
100
150
200
250
300
(c) 2 µM azide10 min
0 2 4 6 8 10
50
100
150
200
250
300
Retention time (min)
(d) 10 µM nitrite10 min
0 2 4 6 8 10
100
200
300
375
Retention time (min)
(a) 0 µM nitrite0 min
0 2 4 6 8 10
100
200
300
375
Retention time (min)
(b) 0 µM nitrite10 min
0 2 4 6 8 10
100
200
300
375
Retention time (min)
(c) 10 µM azide10 min
0 2 4 6 8 10
100
200
300
375
Retention time (min)
(d) 750 µM nitrite10 min
A
B
Fig.2. HPLC chromatograms with UV absorbance detection (338 nm) from the analysis of GSH in incubation mixtures of isolated erythrocytic CAT in PBS (67 mM, pH 7.4, with0.9% [w/v] NaCl) (A) and in diluted hemolysate (1:200 [v/v] with PBS) (B) in the absence or presence of sodium azide or sodium nitrite after incubation times of 0 and 10 minat 25 �C. Initial added concentrations were 5 mM for H2O2 and 10 mM for GSH. Inhibitors and their concentrations were 1 lM for sodium azide and 10 lM for sodium nitritein panel A and 10 lM for sodium azide and 750 lM for sodium nitrite in panel B. y axis: UV absorbance (in mAU); x axis: retention time (in min). The retention time of GS–OPA is 3.35 min. The peak eluting at the retention time of approximately 2.4 min coeluted with nitrite in both the absence and presence of the OPA reagent.
HPLC–UV assay for heCAT activity / A. Böhmer et al. / Anal. Biochem. 410 (2011) 296–303 299
at time 0 to 7.0 mM after 60 min of incubation, indicating that CATactivity present in the reaction mixture (declared as 5 U) was notsufficient to effectively eliminate H2O2. Obviously, for incubationtimes longer than 20 min, [GSH]r did not change but did remain
at the constant levels around 7 mM (curve a), 2.9 mM (curve b),and 0.8 mM (curve c), most likely due to complete consumptionof H2O2. Thus, Fig. 3 indicates that in dependence on the CAT activ-ity and the presence and concentration of CAT inhibitors, the
0 20 40 600
2
4
6
8 no inhibitor
10 µM nitrite
2 µM azide
a
b
c
5 10Time (min)
[GSH
] r (m
M)
Fig.3. Concentration–time profile for residual GSH ([GSH]r) in heCAT (5 U) mixturesin PBS without any inhibitor (a) and in the presence of the CAT inhibitor sodiumnitrite at 10 lM (b) or sodium azide at 2 lM (c) at 25 �C. Initial concentrations were10 mM for GSH and 5 mM for H2O2. All data are shown as means ± standarddeviations from three independent incubations for each time point.
0 10 20 30 40 500
2
4
6
8
10
5CAT activity (U)
[GSa
H] r
(mM
)
Fig.4. Relationship between residual GSH concentration ([GSH]r) and heCATactivity used in incubation mixtures in PBS. Initial concentrations were 10 mMfor GSH and 5 mM for H2O2, and the incubation time was 10 min. All data are shownas means ± standard deviations from three independent incubations for each CATactivity tested. Here 1 U corresponds to a protein concentration of approximately13 ng CAT/ml (declared by the manufacturer).
300 HPLC–UV assay for heCAT activity / A. Böhmer et al. / Anal. Biochem. 410 (2011) 296–303
[GSH]r curve would lie between the curve of nonenzymatic oxida-tion of GSH by H2O2 (i.e., CAT activity = 0 U [see also Fig. 1]) and thehorizontal line at the initial GSH concentration [GSH]0 (e.g., at10 mM). Fig. 4 shows that [GSH]r was higher the greater the heCATactivity was, suggesting that the more effectively H2O2 is removedby heCAT, the more GSH remains in the incubation mixture. Therelationship between [GSH]r and heCAT activity used was linearup to approximately 10 U of heCAT.
Nitrite as an inhibitor of heCAT activity
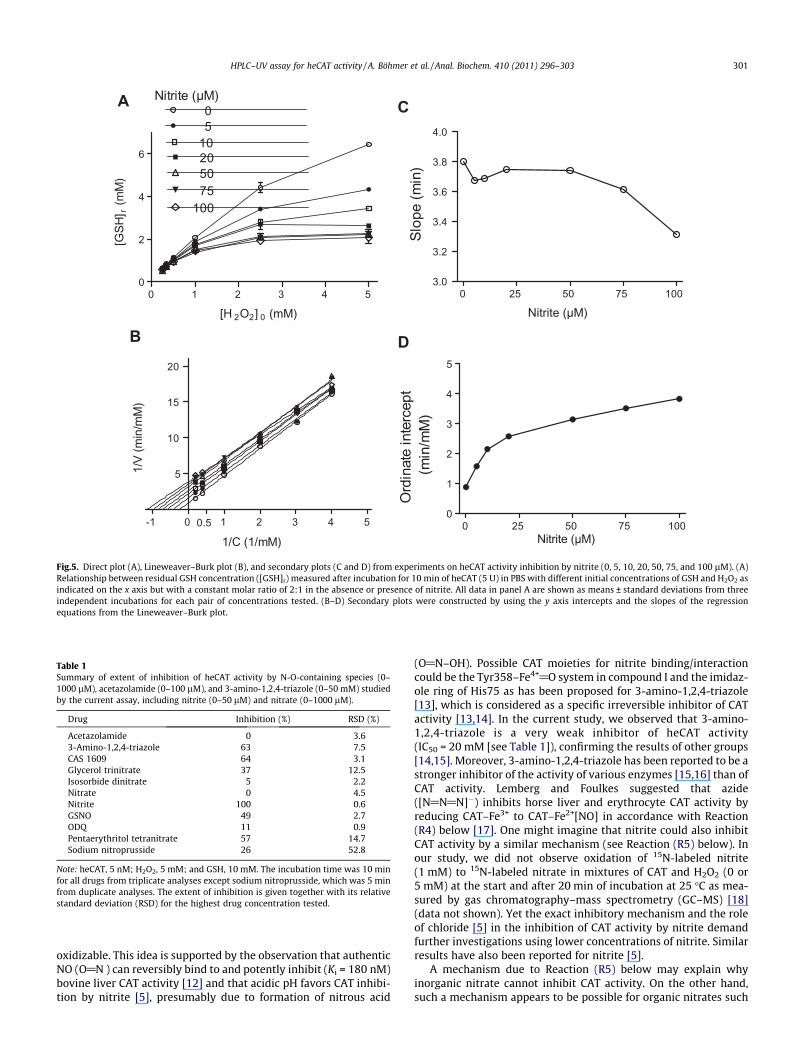
Intracellular GSH and H2O2 concentrations may vary consider-ably. We were interested in knowing whether the HPLC CAT activ-ity assay may also work at lower GSH and H2O2 concentrations.Because the mechanism by which nitrite inhibits CAT activity isnot yet known [5,7], we also studied the mechanism by which ni-trite inhibits heCAT activity. Fig. 5A shows that the HPLC assay canbe used to measure erythrocytic CAT activity in the absence andpresence of the CAT inhibitor nitrite using a constant GSH to
H2O2 molar ratio of 2:1 across the relevant GSH concentrationrange of 0.5–10 mM. However, the relationship between [GSH]r
and [GSH]0 was not linear across the entire concentration range.We performed additional experiments using a fixed CAT activity(5 U) and a constant initial H2O2 concentration (5 mM) but varyingGSH concentrations (1, 2, 5, and 10 mM) and varying incubationtimes (0, 2, 5, 10, 15, and 20 min). After 5 min of incubation, [GSH]r
was 0.5, 0.9, 2.0, and 4.6 mM and did not change on increasingincubation times. These findings suggest that the current HPLCCAT activity assay can be performed with different initial GSHand/or H2O2 concentrations as they are required in mechanisticstudies.
The data of Fig. 5A were treated mathematically to investigatethe type of inhibition of heCAT activity by nitrite. The CAT activityV was calculated and the 1/V values were plotted versus 1/C (i.e., 1/[H2O2]). The highly linear (r2 > 0.9944) Lineweaver–Burk plots(Fig. 5B) together with the secondary plots, notably the nitrite con-centration-dependent decrease of the slope (Fig. 5C) and increaseof the ordinate intercept (Fig. 5D), suggest that nitrite is a partialuncompetitive inhibitor of heCAT activity. By using the GraphPadPrism 5 program and nonlinear fit of the experimental data, wefound that the mechanism obeys the Michaelis–Menten kineticsfor uninhibited heCAT activity (r2 > 0.9993) and for inhibited he-CAT activity for all nitrite concentrations tested (r2 > 0.96). Thebest fitting was obtained for uncompetitive inhibition(0.99 < r2 > 0.63).
In theory, the CAT compound I–H2O2–NO2� and the CAT com-
pound II–H2O2–NO2� complexes can decompose to produce the
reaction products H2O and O2 and the enzyme–inhibitor com-plexes CAT compound I–NO2
� and CAT compound II–NO2�, respec-
tively, with a reaction rate that is smaller than the decompositionof the CAT compound I–H2O2 and CAT compound II–H2O2 com-plexes. The conditions used in the current study, including the sizeof CAT activity, nitrite, and H2O2 concentrations, are comparable tothose used by Titov and Petrenko, who assayed CAT activity calori-metrically [5]. Those authors found that nitrite is an inhibitor ofbull liver CAT activity [5]. They observed that nitrite is a reversibleCAT activity inhibitor regarding halide and thiocyanate ions, butthey did not investigate the mechanism by which nitrite inhibitsCAT activity (e.g., by 30% at 7.5 lM nitrite in the presence of160 mM NaCl and 8 nM CAT) [5]. In our studies, we kept the chlo-ride ion concentration constant at physiological levels; this keepshigh the inhibitory potency of nitrite against CAT activity. By con-sidering all data from inhibitory experiments performed in the cur-rent study, we determined the half-maximal inhibitory constant(IC50) as 9.05 lM nitrite for the investigated heCAT. Takano andcoworkers observed by spectrophotometry an IC50 value of60 lM nitrite for rat liver CAT activity [7]. Those authors suggestedthat nitrite decomposes the CAT compound I. Interestingly, wefound that nitrate did not inhibit heCAT activity at all.
Testing N–O groups containing drugs as inhibitors of heCAT activity
By means of the current HPLC assay, we tested several N–O- orN–N-group drugs as potential inhibitors of heCAT activity. Thechemical structures of these drugs are shown in Suppl. Fig. 1 inthe supplementary material. The organic nitrates investigated—nitroprusside, GSNO, and CAS 1609—are generally regarded as ni-tric oxide (NO) donors, although there is no strong evidence oftheir NO-donating properties [10]. The inhibitory potency of thesedrugs was much lower than that of nitrite (Table 1). The guanylylcyclase inhibitor ODQ and the furoxan CAS 1609 are known tointeract with various heme enzymes [10,11] and emerged as veryweak inhibitors of heCAT activity by the current assay.
These findings may suggest that nitrite (O@N–O�) interactswith CAT through its O@N moiety, of which the N atom is readily
0 1 2 3 4 50
2
4
6
0
20 50 75100
Nitrite (µM)A 5 10
[H 2O2] 0 (mM)
[GSH
] r (m
M)
0 25 50 75 1003.0
3.2
3.4
3.6
3.8
4.0
C
Nitrite (µM)
Slop
e (m
in)
0 25 50 75 1000
1
2
3
4
5
D
Nitrite (µM)
Ord
inat
e in
terc
ept
(min
/mM
)
-1 0 1 2 3 4 5
5
10
15
20
0.5
B
1/C (1/mM)
1/V
(min
/mM
)
Fig.5. Direct plot (A), Lineweaver–Burk plot (B), and secondary plots (C and D) from experiments on heCAT activity inhibition by nitrite (0, 5, 10, 20, 50, 75, and 100 lM). (A)Relationship between residual GSH concentration ([GSH]r) measured after incubation for 10 min of heCAT (5 U) in PBS with different initial concentrations of GSH and H2O2 asindicated on the x axis but with a constant molar ratio of 2:1 in the absence or presence of nitrite. All data in panel A are shown as means ± standard deviations from threeindependent incubations for each pair of concentrations tested. (B–D) Secondary plots were constructed by using the y axis intercepts and the slopes of the regressionequations from the Lineweaver–Burk plot.
Table 1Summary of extent of inhibition of heCAT activity by N-O-containing species (0–1000 lM), acetazolamide (0–100 lM), and 3-amino-1,2,4-triazole (0–50 mM) studiedby the current assay, including nitrite (0–50 lM) and nitrate (0–1000 lM).
Drug Inhibition (%) RSD (%)
Acetazolamide 0 3.63-Amino-1,2,4-triazole 63 7.5CAS 1609 64 3.1Glycerol trinitrate 37 12.5Isosorbide dinitrate 5 2.2Nitrate 0 4.5Nitrite 100 0.6GSNO 49 2.7ODQ 11 0.9Pentaerythritol tetranitrate 57 14.7Sodium nitroprusside 26 52.8
Note: heCAT, 5 nM; H2O2, 5 mM; and GSH, 10 mM. The incubation time was 10 minfor all drugs from triplicate analyses except sodium nitroprusside, which was 5 minfrom duplicate analyses. The extent of inhibition is given together with its relativestandard deviation (RSD) for the highest drug concentration tested.
HPLC–UV assay for heCAT activity / A. Böhmer et al. / Anal. Biochem. 410 (2011) 296–303 301
oxidizable. This idea is supported by the observation that authenticNO (O@N�) can reversibly bind to and potently inhibit (Ki = 180 nM)bovine liver CAT activity [12] and that acidic pH favors CAT inhibi-tion by nitrite [5], presumably due to formation of nitrous acid
(O@N–OH). Possible CAT moieties for nitrite binding/interactioncould be the Tyr358–Fe4+@O system in compound I and the imidaz-ole ring of His75 as has been proposed for 3-amino-1,2,4-triazole[13], which is considered as a specific irreversible inhibitor of CATactivity [13,14]. In the current study, we observed that 3-amino-1,2,4-triazole is a very weak inhibitor of heCAT activity(IC50 = 20 mM [see Table 1]), confirming the results of other groups[14,15]. Moreover, 3-amino-1,2,4-triazole has been reported to be astronger inhibitor of the activity of various enzymes [15,16] than ofCAT activity. Lemberg and Foulkes suggested that azide([N@N@N]�) inhibits horse liver and erythrocyte CAT activity byreducing CAT–Fe3+ to CAT–Fe2+[NO] in accordance with Reaction(R4) below [17]. One might imagine that nitrite could also inhibitCAT activity by a similar mechanism (see Reaction (R5) below). Inour study, we did not observe oxidation of 15N-labeled nitrite(1 mM) to 15N-labeled nitrate in mixtures of CAT and H2O2 (0 or5 mM) at the start and after 20 min of incubation at 25 �C as mea-sured by gas chromatography–mass spectrometry (GC–MS) [18](data not shown). Yet the exact inhibitory mechanism and the roleof chloride [5] in the inhibition of CAT activity by nitrite demandfurther investigations using lower concentrations of nitrite. Similarresults have also been reported for nitrite [5].
A mechanism due to Reaction (R5) below may explain whyinorganic nitrate cannot inhibit CAT activity. On the other hand,such a mechanism appears to be possible for organic nitrates such
0 250 500 750 10000
25
50
75
100A
100Nitrite (µM)
CAT
act
ivity
(%)
0 20 40 60 80 1000
25
50
75
100
10
B
Azide (µM)
CAT
act
ivity
(%)
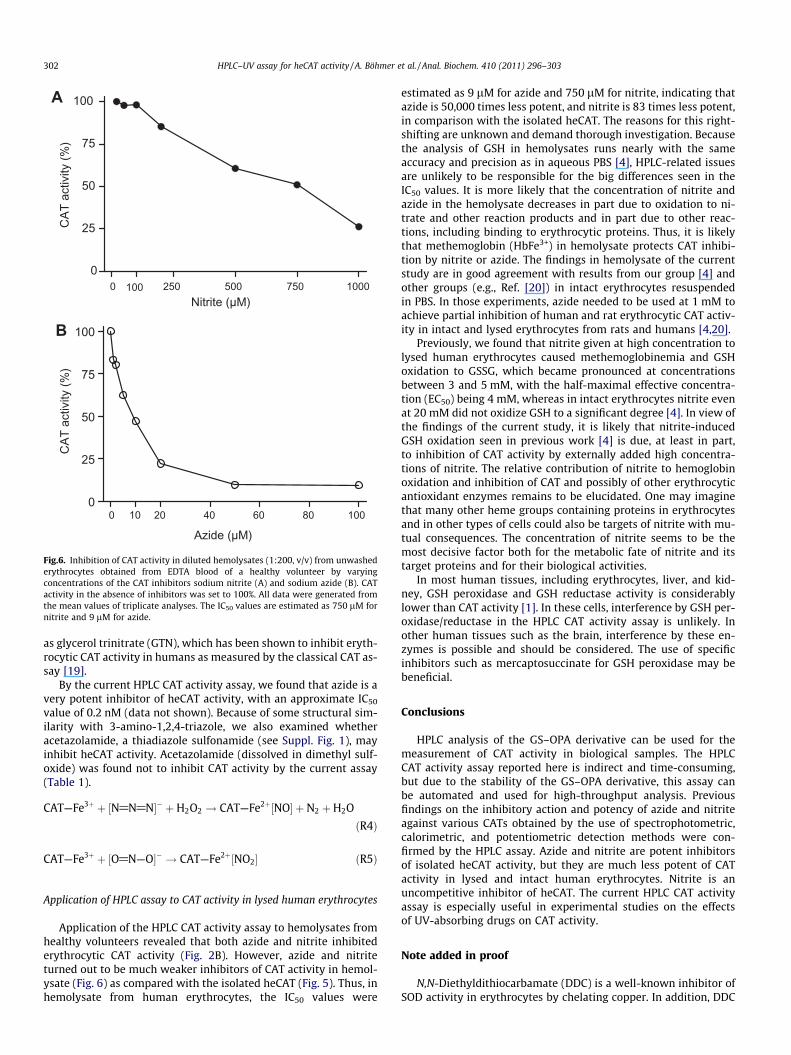
Fig.6. Inhibition of CAT activity in diluted hemolysates (1:200, v/v) from unwashederythrocytes obtained from EDTA blood of a healthy volunteer by varyingconcentrations of the CAT inhibitors sodium nitrite (A) and sodium azide (B). CATactivity in the absence of inhibitors was set to 100%. All data were generated fromthe mean values of triplicate analyses. The IC50 values are estimated as 750 lM fornitrite and 9 lM for azide.
302 HPLC–UV assay for heCAT activity / A. Böhmer et al. / Anal. Biochem. 410 (2011) 296–303
as glycerol trinitrate (GTN), which has been shown to inhibit eryth-rocytic CAT activity in humans as measured by the classical CAT as-say [19].
By the current HPLC CAT activity assay, we found that azide is avery potent inhibitor of heCAT activity, with an approximate IC50
value of 0.2 nM (data not shown). Because of some structural sim-ilarity with 3-amino-1,2,4-triazole, we also examined whetheracetazolamide, a thiadiazole sulfonamide (see Suppl. Fig. 1), mayinhibit heCAT activity. Acetazolamide (dissolved in dimethyl sulf-oxide) was found not to inhibit CAT activity by the current assay(Table 1).
CAT—Fe3þ þ ½N@N@N�� þH2O2 ! CAT—Fe2þ½NO� þ N2 þH2O
ðR4Þ
CAT—Fe3þ þ ½O@N—O�� ! CAT—Fe2þ½NO2� ðR5Þ
Application of HPLC assay to CAT activity in lysed human erythrocytes
Application of the HPLC CAT activity assay to hemolysates fromhealthy volunteers revealed that both azide and nitrite inhibitederythrocytic CAT activity (Fig. 2B). However, azide and nitriteturned out to be much weaker inhibitors of CAT activity in hemol-ysate (Fig. 6) as compared with the isolated heCAT (Fig. 5). Thus, inhemolysate from human erythrocytes, the IC50 values were
estimated as 9 lM for azide and 750 lM for nitrite, indicating thatazide is 50,000 times less potent, and nitrite is 83 times less potent,in comparison with the isolated heCAT. The reasons for this right-shifting are unknown and demand thorough investigation. Becausethe analysis of GSH in hemolysates runs nearly with the sameaccuracy and precision as in aqueous PBS [4], HPLC-related issuesare unlikely to be responsible for the big differences seen in theIC50 values. It is more likely that the concentration of nitrite andazide in the hemolysate decreases in part due to oxidation to ni-trate and other reaction products and in part due to other reac-tions, including binding to erythrocytic proteins. Thus, it is likelythat methemoglobin (HbFe3+) in hemolysate protects CAT inhibi-tion by nitrite or azide. The findings in hemolysate of the currentstudy are in good agreement with results from our group [4] andother groups (e.g., Ref. [20]) in intact erythrocytes resuspendedin PBS. In those experiments, azide needed to be used at 1 mM toachieve partial inhibition of human and rat erythrocytic CAT activ-ity in intact and lysed erythrocytes from rats and humans [4,20].
Previously, we found that nitrite given at high concentration tolysed human erythrocytes caused methemoglobinemia and GSHoxidation to GSSG, which became pronounced at concentrationsbetween 3 and 5 mM, with the half-maximal effective concentra-tion (EC50) being 4 mM, whereas in intact erythrocytes nitrite evenat 20 mM did not oxidize GSH to a significant degree [4]. In view ofthe findings of the current study, it is likely that nitrite-inducedGSH oxidation seen in previous work [4] is due, at least in part,to inhibition of CAT activity by externally added high concentra-tions of nitrite. The relative contribution of nitrite to hemoglobinoxidation and inhibition of CAT and possibly of other erythrocyticantioxidant enzymes remains to be elucidated. One may imaginethat many other heme groups containing proteins in erythrocytesand in other types of cells could also be targets of nitrite with mu-tual consequences. The concentration of nitrite seems to be themost decisive factor both for the metabolic fate of nitrite and itstarget proteins and for their biological activities.
In most human tissues, including erythrocytes, liver, and kid-ney, GSH peroxidase and GSH reductase activity is considerablylower than CAT activity [1]. In these cells, interference by GSH per-oxidase/reductase in the HPLC CAT activity assay is unlikely. Inother human tissues such as the brain, interference by these en-zymes is possible and should be considered. The use of specificinhibitors such as mercaptosuccinate for GSH peroxidase may bebeneficial.
Conclusions
HPLC analysis of the GS–OPA derivative can be used for themeasurement of CAT activity in biological samples. The HPLCCAT activity assay reported here is indirect and time-consuming,but due to the stability of the GS–OPA derivative, this assay canbe automated and used for high-throughput analysis. Previousfindings on the inhibitory action and potency of azide and nitriteagainst various CATs obtained by the use of spectrophotometric,calorimetric, and potentiometric detection methods were con-firmed by the HPLC assay. Azide and nitrite are potent inhibitorsof isolated heCAT activity, but they are much less potent of CATactivity in lysed and intact human erythrocytes. Nitrite is anuncompetitive inhibitor of heCAT. The current HPLC CAT activityassay is especially useful in experimental studies on the effectsof UV-absorbing drugs on CAT activity.
Note added in proof
N,N-Diethyldithiocarbamate (DDC) is a well-known inhibitor ofSOD activity in erythrocytes by chelating copper. In addition, DDC

HPLC–UV assay for heCAT activity / A. Böhmer et al. / Anal. Biochem. 410 (2011) 296–303 303
has been reported to result in oxyhemoglobin-catalyzed GSHdepletion and methemoglobin production [21]. By using the pres-ent CAT activity assay we found that DDC (Sigma-Aldrich; Stein-heim, Germany) inhibited heCAT activity with an IC50 value of630 lM. In the absence of CAT and H2O2, we found that DDC (0–10 mM) did not change the GSH concentration (10 mM) after anincubation time of 10 min.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.ab.2010.11.026.
References
[1] B. Halliwell, J.M.C. Gutteridge, Free Radicals in Biology and Medicine, third ed.,Oxford Science, Oxford, UK, 1999.
[2] D. Giustarini, I. Dalle-Donne, D. Tsikas, R. Rossi, Oxidative stress and humandiseases: origin, link, measurement, mechanisms, and biomarkers, Crit. Rev.Clin. Lab. Sci. 46 (2009) 241–281.
[3] B.A. Neuschwander-Tetri, F.J. Roll, Glutathione measurement by high-performance liquid chromatography separation and fluorometric detection ofthe glutathione–orthophthalaldehyde adduct, Anal. Biochem. 179 (1989) 236–241.
[4] J.T. Michaelsen, S. Dehnert, D. Giustarini, B. Beckmann, D. Tsikas, HPLC analysisof human erythrocytic glutathione forms using OPA N-acetyl-cysteine ethylester: evidence for nitrite-induced GSH oxidation to GSSG, J. Chromatogr. B877 (2009) 3405–3417.
[5] V.Y. Titov, Y.M. Petrenko, Nitrite–catalase interaction as an important elementof nitrite toxicity, Biochemistry 68 (2003) 627–633.
[6] D. Tsikas, J. Sandmann, S. Rossa, F.M. Gutzki, J.C. Frölich, Investigations ofS-transnitrosylation reactions between low- and high-molecular-weightS-nitroso compounds and their thiols by high-performance liquidchromatography and gas chromatography–mass spectrometry, Anal.Biochem. 270 (1999) 231–241.
[7] T. Takano, Y. Miyazaki, K. Nakata, Interaction of nitrite with catalase in theperfused rat liver, Food Chem. Toxicol. 26 (1988) 837–839.
[8] Y. Iwasaki, Y. Saito, Y. Nakano, K. Mochizuki, O. Sakata, R. Ito, K. Saito, H.Nakazawa, Chromatographic and mass spectrometric analysis of glutathione inbiological samples, J. Chromatogr. B 877 (2009) 3309–3317.
[9] P. Monostori, D. Wittmann, E. Karg, S. Túri, Determination of glutathione andglutathione disulfide in biological samples, J. Chromatogr. B 877 (2009) 3331–3346.
[10] O. Krause, E. Wätzig, H. Acil, S. König, K. Unthan-Fechner, D. Tsikas, I. Probst,Role of carbon monoxide and nitric oxide in adult rat hepatocytes proliferatingin vitro: effects of CAS 1609, Nitric oxide 23 (2010) 220–226.
[11] M. Feelisch, P. Kotsonis, J. Siebe, B. Clement, H.H.W. Schmidt, The solubleguanylyl cyclase inhibitor 1H-[1,2,4]oxadiazolo-[4, 3-a]quinoxalin-1-one is anonselective heme protein inhibitor of nitric oxide synthase and othercytochrome P-450 enzymes involved in nitric oxide donor bioactivation,Mol. Pharmacol. 56 (1999) 243–253.
[12] G.C. Brown, Reversible binding and inhibition of catalase by nitric oxide, Eur. J.Biochem. 232 (1995) 188–191.
[13] C.D. Putnam, A.S. Arvai, Y. Bourne, Active and inhibited human catalasestructures: ligand and NADPH binding and catalytic mechanism, J. Mol. Biol.296 (2000) 295–309.
[14] D. Darr, I. Fridovic, Irreversible inactivation of catalase by 3-amino-1, 2, 4-triazole, Biochem. Pharmacol. 35 (1986) 3642.
[15] R.M.C. Dawson, D.C. Elliott, W.H. Elliot, K.M. Jones, Data for BiochemicalResearch, 3rd ed., Oxford University Press, New York, 1993. pp. 302–303.
[16] H. Zollner, Handbook of Enzyme Inhibitors, Part B, VCH, Weinheim, Germany,1993. p. 563.
[17] R. Lemberg, E.C. Foulkes, Reaction between catalase and hydrogen peroxide,Nature 161 (1948) 131–132.
[18] D. Tsikas, Simultaneous derivatization and quantification of the nitric oxidemetabolites nitrite and nitrate in biological fluids by gas chromatography/mass spectrometry, Anal. Chem. 72 (2000) 4064–4072.
[19] Y. Alicigüzel, S. Aktas�, H. Bozan, M. Aslan, Effect of intravenous nitroglycerintherapy on erythrocytic antioxidant enzymes, J. Enzyme Inhib. Med. Chem. 20(2005) 293–296.
[20] R. Rossi, A. Milzani, I. Dalle-Donne, F. Giannerini, D. Giustarini, L. Lusini, R.Colombo, P. Di Simplicio, Different metabolizing ability of thiol reactants inhuman and rat blood: biochemical and pharmacological implications, J. Biol.Chem. 276 (2001) 7004–7010.
[21] M.J. Kelner, N.M. Alexander, Inhibition of erythrocyte superoxide dismutase bydiethyldithiocarbamate also results in oxyhemoglobin-catalyzed glutathionedepletion and methemoglobin production, J. Biol. Chem. 261 (1986) 1636–1641.
Related Documents











