ELSEVIER Applied Animal BehaviourScience45 ( 1995) 151-163 APPLIED ANIMAL BEHAVIOUR SCIENCE The behavior of singly-caged, yearling rhesus monkeys is affected by the environment outside of the cage Steven J. Schapiro *, Leila M. Porter, Scott A. Suarez, Mollie A. Bloomsmith Department of Veterinary Sciences, The University of Texas, M.D. Anderson Cancer Center, Science Park, Rt. 2, Box 151-81, Bastrop, TX 78602, USA Accepted 12 April 1995 Abstract Numerous studies have demonstrated that the behavior of singly caged laboratory primates can be positively affected by inanimate enrichment within the cage. The environment outside of the cage may also influence the behavior of singly caged rhesus monkeys. To test this, we compared two cohorts of yearling rhesus (Mucaca muluttu) ; one that had only limited stimulation in the environment outside of the cage (singly caged in indoor rooms), and one that had considerable naturalistic and social stimulation in the extra-cage environment (singly caged in an outdoor building). Half the animals in each housing condition received a three-phase enrichment program and the rest served as controls. Subjects that had sensory access to social groups of conspecifics spent significantly more time drinking, feeding, and behaving abnormally, and significantly less time inactive and playing than did subjects that only had sensory access to other singly caged rhesus. Enriched subjects in both settings spent significantly more time playing and less time self-grooming than did controls. There was also a significant interaction for time spent in play, with enriched subjects living indoors playing the most. Although subjects that received considerable stimulation from outside of the cage spent more time engaged in abnormal behavior, most was pacing; usually in response to human manipu- lations of the other monkeys in the colony. Subjects housed indoors, with limited extra-cage stimu- lation spent less time pacing, but more time in potentially injurious abnormal activities. Among enriched subjects only, those housed indoors spent less time drinking and behaving abnormally and more time playing and using enrichment. Inanimate enrichment within the cage led to consistent positive changes in behavior for singly housed rhesus. Although the environment outside of the cage was shown to influence behavior, it was not clear whether exposure to numerous social and naturalistic stimuli was preferable to more limited stimulation for promoting well-being. Although the increased stimulation associated with a complex extra-cage environment is usually considered beneficial, not all such stimulation is positive. Frequent disturbances to social groups in view of singly caged subjects resulted in increased pacing, but the complex extra-cage environment also provided subjects with *Corresponding author. 0168-1591/95/$09.50 0 1995Elsevier Science B.V. All rights reserved SSDIO168-1591(95)00597-8

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ELSEVIER Applied Animal Behaviour Science 45 ( 1995) 151-163
APPLIED ANIMAL BEHAVIOUR
SCIENCE
The behavior of singly-caged, yearling rhesus monkeys is affected by the environment outside of
the cage
Steven J. Schapiro *, Leila M. Porter, Scott A. Suarez, Mollie A. Bloomsmith
Department of Veterinary Sciences, The University of Texas, M.D. Anderson Cancer Center, Science Park, Rt. 2, Box 151-81, Bastrop, TX 78602, USA
Accepted 12 April 1995
Abstract
Numerous studies have demonstrated that the behavior of singly caged laboratory primates can be positively affected by inanimate enrichment within the cage. The environment outside of the cage may also influence the behavior of singly caged rhesus monkeys. To test this, we compared two cohorts of yearling rhesus (Mucaca muluttu) ; one that had only limited stimulation in the environment outside of the cage (singly caged in indoor rooms), and one that had considerable naturalistic and social stimulation in the extra-cage environment (singly caged in an outdoor building). Half the animals in each housing condition received a three-phase enrichment program and the rest served as controls. Subjects that had sensory access to social groups of conspecifics spent significantly more time drinking, feeding, and behaving abnormally, and significantly less time inactive and playing than did subjects that only had sensory access to other singly caged rhesus. Enriched subjects in both settings spent significantly more time playing and less time self-grooming than did controls. There was also a significant interaction for time spent in play, with enriched subjects living indoors playing the most. Although subjects that received considerable stimulation from outside of the cage spent more time engaged in abnormal behavior, most was pacing; usually in response to human manipu- lations of the other monkeys in the colony. Subjects housed indoors, with limited extra-cage stimu- lation spent less time pacing, but more time in potentially injurious abnormal activities. Among enriched subjects only, those housed indoors spent less time drinking and behaving abnormally and more time playing and using enrichment. Inanimate enrichment within the cage led to consistent positive changes in behavior for singly housed rhesus. Although the environment outside of the cage was shown to influence behavior, it was not clear whether exposure to numerous social and naturalistic stimuli was preferable to more limited stimulation for promoting well-being. Although the increased stimulation associated with a complex extra-cage environment is usually considered beneficial, not all such stimulation is positive. Frequent disturbances to social groups in view of singly caged subjects resulted in increased pacing, but the complex extra-cage environment also provided subjects with
*Corresponding author.
0168-1591/95/$09.50 0 1995 Elsevier Science B.V. All rights reserved SSDIO168-1591(95)00597-8

152 S.J. Schupiro et al. /Applied Animal Behaviour Science 45 (1995) 151-163
more opportunities to express and observe components of the species-typical behavioral repertoire.
Keywords: Behavioral management; Single housing; Social restriction; Extra-cage environment; Abnormal behavior; Environmental enrichment
1. Introduction
Considerable data have accumulated in the past several years demonstrating the positive effects that inanimate environmental enrichment can have on the behavior of captive pri- mates (Bloomsmith et al., 1988; Novak et al., 1993; Schapiro and Bloomsmith, 1995). The precise effects of enrichment have varied, depending on the type of enhancements utilized (Bloomsmith et al., 1990; Bayne et al., 1991; Parks and Novak, 1993; Schapiro and Bloomsmith, 1995) ; the age, gender, species, and housing condition (physical and social) of the subjects (O’Neill et al., 1990; Coe, 1991; Line et al., 1991; Ruppenthal et al., 1991; Schapiro et al., 1993; Crockett et al., 1994; Schapiro and Bloomsmith, 1994); and the experimental design applied (Schapiro et al., 1995). Although positive enrichment effects have been measured for socially housed primates (Bloomsmith et al., 1988; Novak et al., 1993; Parks and Novak, 1993), there is more interest in studying enrichment for those housed alone (Bryant et al., 1988; Brent et al., 1989; Bayne et al., 1991, 1992; Line and Morgan, 1991; Schapiro and Bloomsmith, 1995).
Primates housed in single cages are often arranged in rows that face one another or that provide visual access to social groups (Dienske et al., 1980; Schapiro et al., 1994; Schapiro and Bloomsmith, 1995) so that animals that are physically restricted from contacting conspecifics can view, hear, and smell them (Dienske et al., 1980; Novak and Suomi, 1988; Markowitz and Line, 1989; O’Neill, 1989; Novak and Bayne, 1991). The goal of this strategy, which directly addresses regulations that require such arrangements, is that singly caged animals will receive social stimulation even when physically separated. The operative assumption is that receiving non-contact social stimulation is a positive experience, although few data are available to support this notion.
One of our earlier studies (Schapiro and Bloomsmitb, 1995) showed that inanimate enrichment, including physical, feeding, and sensory enhancements to the single cage, influenced the behavior of yearling rhesus monkeys. Specifically, enriched subjects spent more time playing and less time self-grooming than did control subjects. Several of the analyses from this study suggested that variability in the environment outside of the cage may have influenced the findings.
There has been little systematic investigation of the effects of the extra-cage environment on the behavior of captive primates. Studies have examined the effects of a total change of environment from an indoor to an outdoor setting (Nieuwenhuijsen and de Waal, 1982; Goerke et al., 1987; Chamove and Rohrhuber, 1989; O’Neill et al., 1991; Novak et al., 1992), but in these studies, there were important changes in the usable volume available to subjects between indoor and outdoor conditions.
The present investigation compares the effects of single housing with and without sensory access to social groups of conspecifics and other extra-cage stimuli. We hypothesized that singly caged subjects housed outdoors that could see other singly caged and group-housed

S.J. Schapiro et al. /Applied Animal Behaviour Science 45 (1995) 151-163 153
rhesus monkeys, and that were exposed to natural variation in environmental conditions would behave differently than would subjects that were housed in indoor rooms without visible social groups and with little variability in environmental conditions.
2. Animals, materials and methods
2. I. Subjects and housing
Two groups of rhesus monkeys (Macaca mulatta) served as subjects in this investigation. Group 1 consisted of 34 yearlings ( 18 males, 16 females) born in 1988; data from this group have been reported previously (as have data from Groups 2 and 3 (Schapiro and Bloomsmith 1995) ) . Group 4 comprised 32 yearlings ( 16 males, 16 females) born in 199 1. One half of the animals in each group received an environmental enrichment program that consisted of three separate conditions and the other half served as controls, receiving no environmental enrichment. Control subjects could see when enrichment devices were given to experimental subjects but had only limited sensory access to monkeys using enrichment. One enriched subject from Group 4 had to be replaced; thus only 15 subjects in this group were observed in all three enrichment conditions. Monkey biscuits were provided twice daily, and supplemental pieces of oranges were given three times per week to all animals. Water was available ad libitum. All subjects were housed in rack-mounted stainless steel cages (0.4 m2) with visual, auditory, and olfactory access to similarly housed monkeys. Physical contact between subjects was restricted.
All monkeys spent their first year of life in their natal group, which consisted of one adult male, four to six adult females, and their most recent offspring. All study animals were singly caged from 1 to 2 years of age. Subjects in G~pup 1 spent their year of single caging in indoor rooms and had sensory access to only singly housed conspecifics. Group 4 subjects spent their year of single caging in an outdoor building and had sensory access to both group-housed and singly housed conspecifics.
The indoor rooms were maintained on a fixed light: dark ( 12 : 12) schedule and within a narrow temperature range (73-85°F). Animals in the outdoor building were subject to the fluctuations in daylength, temperature, and environmental events that are typical for central Texas.
2.2. Enrichment
Enriched subjects in both groups received the three types of enrichment in the same order. Physical enrichment (toys, perches, etc.) was provided for 3 months immediately upon being separated from the natal group and placed in the single cage. This was followed by 6 months of feeding enrichment (devices, foods, etc.) and finally, by 3 months of sensory enrichment (videotapes of primates; for additional details on the three-phase enrichment program see Schapiro and Bloomsmith, 1995). The enrichment program was consistent across years, with only one toy and one feeding device differing between groups.
The environment outside of the single cage, which we will refer to as the extracage environment, was isolated as the factor of interest in the present study by maintaining a

154 S.J. Schapiro et al. /Applied Animal Behaviour Science 45 (1995) 151-163
Table 1 Categories of activity analyzed with component behaviors (see Schapiro and Bloomsmith, 1995 for complete operational definitions)
Activity Component behaviors
Inactivity Sexual behavior Social behavior Drinking Self-grooming Feeding Playing Vocalizing Abnormal behavior
Exploring
Sit, sleep Masturbate, thrust Touch neighbor, threaten, present, lipsmack Drink Self-groom Eat, forage Play Vocalize Drink urine, pick, suck digit, salute, twirl, rock, somersault, pace, head toss, coprophagy, self-aggression, self-clasp, abnormal (other) Investigate, look, olfactory explore
constant enclosure size (0.4 m* single cage) and the same order of enrichment for subjects singly-caged in Groups 1 and 4. Social factors, ecological factors, and the occurrence of routine husbandry procedures (or daily events as examined by Crockett and Bowden ( 1994) and by Line and colleagues (Line et al., 1989a, 1991)) were identified as important components of the extra-cage environment.
2.3. Data collection
Fifteen-minute focal animal observations (Altmann, 1974) were conducted on all sub- jects. Twelve hundred and seventy-nine hours of data were collected for subjects in Group 1 (Schapiro and Bloomsmith, 1995) and another 696 h were collected for subjects in Group 4, using a Tandy 102 portable computer and The Observer observational software package (Noldus, 1991). Data were collected between 0830 and 1730 throughout the year, were balanced for time of day, and as in the previous study (Schapiro and Bloomsmith, 1995)) included periods when enrichment was available and when it was not.
Abnormal behavior was emphasized on the ethogram employed (see Table 1 and Schapiro and Bloomsmith, 1995), and enrichment use could be recorded simultaneously with all other focal animal behavior. Interobserver reliability for cumulative durations of behaviors within observation sessions was measured monthly for a total of six observers and averaged 88.7% for behavior and 96.8% for enrichment use.
Data were analyzed using multivariate analysis of variance (MANOVA) techniques within the Systat statistical package. Individual behaviors from the ethogram were grouped into the ten mutually exclusive categories of activity presented in Table 1. The dependent measures entered into the analysis were the mean cumulative durations of each activity category for each subject during each enrichment condition, averaged across all observation sessions. A total of 198 mean scores per behavior were analyzed. Observations on control subjects were treated in this way so that we could distinguish changes in behavior due to enrichment from changes in behavior due to the length of time that subjects had spent singly caged.

S.J. Schapiro et al. /Applied Animal Behmiour Science 45 (I 995) 151-l 63 155
2.4. Analyses
The primary analysis was a 2 X 2 MANOVA with the extra-cage environment (limited stimulation in indoor rooms vs. considerable stimulation in an outdoor building) and treatment (enriched vs. control) as the independent variables.
Two other analyses were conducted for enriched subjects only: a one-way repeated measures MANOVA by enrichment condition and a mixed 2 ( X 3) ANOVA for the dependent measure ‘enrichment use’ with the extra-cage environment as the between- subjects variable and enrichment condition as the within-subjects variable. This ANOVA was done separately to prevent confounding caused by the overlap of enrichment use with the mutually exclusive categories of behavior. The two analyses of data from enriched subjects only were designed to determine whether there were differences in behavioral effects across the three enrichment conditions. As in a previous study (Schapiro and Bloom- smith, 1995)) our goal was to measure behavior as a function of the combined manipulations that comprised each enrichment condition which would include short-term ‘carryover effects’ when, for example, devices were empty or the videotapes were finished within an observation session.
3. Results
3.1. All subjects
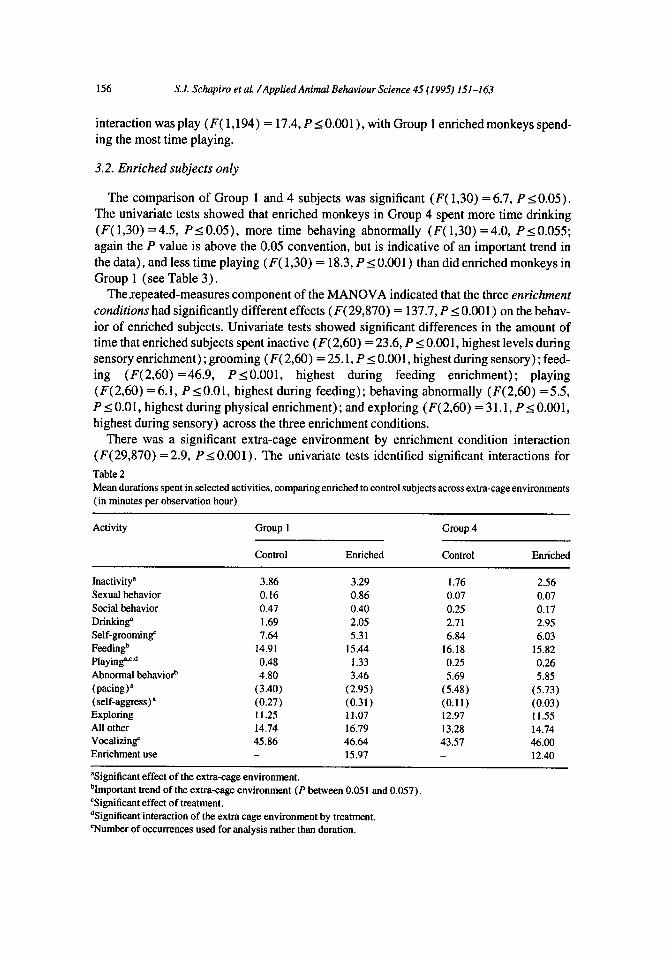
The multivariate test showed that the effect of the extra-cage environment was significant (Wilks’ lambda = 0.64, F( 10,185) = 10.4, P I 0.001) (see Table 2). Subsequent univariate tests indicated that Group 4 subjects (complex extra-cage environment; singly caged out- doors) spent significantly less time inactive (F( 1,194) = 8.5, P10.01) and playing (F( 1,194) = 46.5, PI 0.001) andspent significantly more timedrinking (F( 1,194) = 18.7, P I 0.001) , feeding (F( 1,194) = 3.9, P = 0.05 1) , and behaving abnormally (F( 1,194) = 3.7, P = 0.057) than did Group 1 animals (less stimulating extra-cage envi- ronment; singly caged in indoor rooms). The P values for feeding and abnormal behavior are slightly above the conventional 0.05 level. The obtained values (0.051 and 0.057, respectively) are close enough to the 0.05 level to indicate important trends in the data; trends which will be discussed where appropriate. Post-hoc analyses showed that within the category of abnormal behavior, Group 4 subjects spent significantly more time pacing (97% vs. 72% of abnormal behavior; F( 1,194) = 12.8, P 10.001) but significantly less time self- aggressing (0.07 vs. 0.28 min per observation hour; F( 1,194) = 17.3, P I 0.001) than did Group 1 monkeys.
The multivariate test examining treatment (enriched vs. control) was also significant (Wilks’ lambda = 0.83, F( 10,185) = 3.9, PI 0.001)) and subsequent univariate tests revealed that enriched subjects spent less time self-grooming (F( 1,194) = 13.0, P s 0.001) and more time playing (F( 1,194) = 18.2, PI 0.001) than did control subjects.
There was a significant extra-cage environment by treatment interaction (Wilks’ lambda = 0.87, F( 10,185) = 2.8, P I 0.01). The only activity with a significant univariate
156 S.J. Schapiro et al. /Applied Animal Behaviour Science 45 (1995) 151-163
interaction was play (F( 1,194) = 17.4, PI 0.001) , with Group 1 enriched monkeys spend- ing the most time playing.
3.2. Enriched subjects only
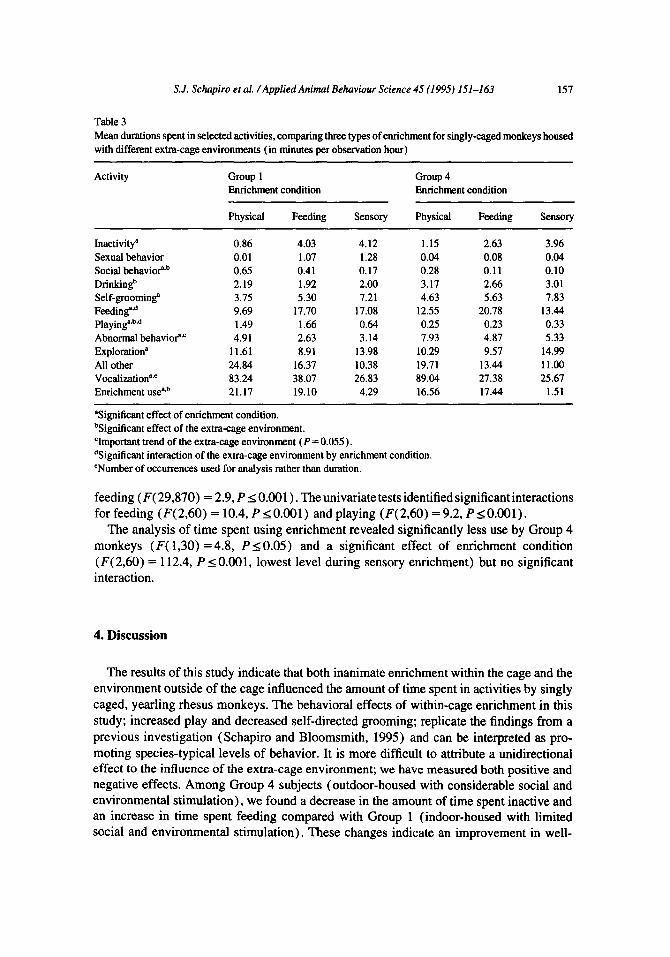
The comparison of Group 1 and 4 subjects was significant (F( 1,30) = 6.7, PI 0.05). The univariate tests showed that enriched monkeys in Group 4 spent more time drinking (F( 1,30) = 4.5, PI 0.05), more time behaving abnormally (F( 1,30) = 4.0, PI 0.055; again the P value is above the 0.05 convention, but is indicative of an important trend in the data), and less time playing (F( 1,30) = 18.3, P s 0.001) than did enriched monkeys in Group 1 (see Table 3).
Therepeated-measures component of the MANOVA indicated that the three enrichment conditions had significantly different effects (F( 29,870) = 137.7, PI 0.001) on the behav- ior of enriched subjects. Univariate tests showed significant differences in the amount of time that enriched subjects spent inactive (F( 2,60) = 23.6, PI 0.001, highest levels during sensory enrichment) ; grooming (F( 2,60) = 25.1, P I 0.001, highest during sensory) ; feed- ing (F( 2,60) = 46.9, P s 0.001, highest during feeding enrichment) ; playing (F( 2,60) = 6.1, P I 0.01, highest during feeding) ; behaving abnormally (F( 2,60) = 5.5, P I 0.01, highest during physical enrichment) ; and exploring (F( 2,60) = 3 1.1, P I 0.001, highest during sensory) across the three enrichment conditions.
There was a significant extra-cage environment by enrichment condition interaction (F( 29,870) = 2.9, P < 0.001) . The univariate tests identified significant interactions for Table 2 Mean durations spent in selected activities, comparing enriched to control subjects across extra-cage environments (in minutes per observation hour)
Activity Group 1
Control Enriched
Group 4
Control Enriched
Inactivity’ Sexual behavior Social behavior Drinking” Self-grooming” Feeding” Playing”L.d Abnormal behaviop (pacing)” (self-aggress)’ Exploring All other Vocalizing’ Enrichment use
3.86 0.16 0.47 1.69 7.64
14.91 0.48 4.80
(3.40) (0.27) 11.25 14.74 45.86
3.29 0.86 0.40 2.05 5.31
15.44 1.33 3.46
(2.95) (0.31) 11.07 16.79 46.64 15.97
1.76 0.07 0.25 2.71 6.84
16.18 0.25 5.69
(5.48) (0.11) 12.97 13.28 43.57 _
2.56 0.07 0.17 2.95 6.03
15.82 0.26 5.85
(5.73) (0.03) 11.55 14.74 46.00 12.40
“Significant effect of the extra-cage environment. bImportant trend of the extra-cage environment (P between 0.05 1 and 0.057). ‘Significant effect of treatment. dSignificaut interaction of the extra cage environment by treatment. ‘Number of occurrences used for analysis rather than duration.
S.J. Schapiro et al. /Applied Animal Behaviour Science 45 (1995) 151-163 157
Table 3 Mean durations spent in selected activities, comparing three types of enrichment for singly-caged monkeys housed with different extra-cage environments (in minutes per observation hour)
Activity Group 1 Enrichment condition
Group 4 Enrichment condition
Physical Feeding Sensory Physical Feeding Sensory
Inactivity” 0.86 4.03 4.12 1.15 2.63 3.96 Sexual behavior 0.01 1.07 1.28 0.04 0.08 0.04 Social behavioQb 0.65 0.41 0.17 0.28 0.11 0.10 Drinkingb 2.19 1.92 2.00 3.17 2.66 3.01 Self-grooming” 3.75 5.30 7.21 4.63 5.63 7.83 Feeding”.“ 9.69 17.70 17.08 12.55 20.78 13.44 Playing”.b” 1.49 1.66 0.64 0.25 0.23 0.33 Abnormal behavioe’ 4.91 2.63 3.14 7.93 4.87 5.33 Exploration” 11.61 8.91 13.98 10.29 9.57 14.99 All other 24.84 16.37 10.38 19.71 13.44 11.00 Vocalization”~ 83.24 38.07 26.83 89.04 27.38 25.67 Enrichment useab 21.17 19.10 4.29 16.56 17.44 1.51
“Significant effect of enrichment condition. bSignificant effect of the extra-cage environment. ‘Important trend of the extra-cage environment (P = 0.055). %ignificant interaction of the extra-cage environment by enrichment condition. ‘Number of occurrences used for analysis rather than duration.
feeding (F( 29,870) = 2.9, P I 0.001) . The univariate tests identified significant interactions for feeding (F( 2,60) = 10.4, PI 0.001) and playing (F( 2,60) = 9.2, P s 0.001).
The analysis of time spent using enrichment revealed significantly less use by Group 4 monkeys (F( 1,30) = 4.8, PS 0.05) and a significant effect of enrichment condition (F( 2,60) = 112.4, P I 0.001, lowest level during sensory enrichment) but no significant interaction.
4. Discussion
The results of this study indicate that both inanimate enrichment within the cage and the environment outside of the cage influenced the amount of time spent in activities by singly caged, yearling rhesus monkeys. The behavioral effects of within-cage enrichment in this study; increased play and decreased self-directed grooming; replicate the findings from a previous investigation (Schapiro and Bloomsmith, 1995) and can be interpreted as pro- moting species-typical levels of behavior. It is more difficult to attribute a unidirectional effect to the influence of the extra-cage environment; we have measured both positive and negative effects. Among Group 4 subjects (outdoor-housed with considerable social and environmental stimulation), we found a decrease in the amount of time spent inactive and an increase in time spent feeding compared with Group 1 (indoor-housed with limited social and environmental stimulation). These changes indicate an improvement in well-

158 S.J. Shapiro et al. /Applied Animal Behaviour Science 45 (1995) 151-163
being. However, we also found decreased time spent playing and increased time spent behaving abnormally, changes which indicate a regression in well-being.
Few published investigations have isolated and examined the effects of the environment outside of the primary enclosure. Studies that have reported more species-typical behavior for groups of marmosets and tamarins (Chamove and Rohrhuber, 1989), rhesus monkeys (O’Neill et al., 1991; Novak et al., 1992)) and chimpanzees (Nieuwenhuijsen and de Waal, 1982) housed outdoors vs. indoors all included a substantial increase in the volume of space available to the subjects from the indoor to the outdoor condition. Unlike such dramatic increases in available space, minimal changes in cage size for singly housed macaques lead to little if any change in putative measures of psychological well-being (Line et al., 1989b, 1990, 1991; Crockett et al., 1993; Crockett and Bowden, 1994). We maintained an equiv- alent cage size while altering the complexity of the extra-cage environment (in terms of social stimulation and ecological variability), something not achieved in any of these other studies.
Many discussions of appropriate captive housing conditions mention the potential benefits to social, reproductive, parental, and learning abilities of housing socially restricted primates with visual access to conspecifics (Dienske et al., 1980, Novak and Suomi, 1988; Markowitz and Line, 1989; Novak and Bayne, 1991). In the present study, behaviors changed as a function of the extra-cage environment, but the data are equivocal; some changes are indicative of an improvement in well-being, whereas others are indicative of diminished well being. Specifically, increased durations spent feeding and decreased time spent inactive while housed in an outdoor building with a complex extra-cage environment, are more representative of species-typical levels. However, a decrease in time spent playing and an increase in time spent behaving abnormally outdoors are less representative of species- typical levels. The effect on abnormal behavior is the most interesting.
Abnormal behavior was not significantly affected in any of our previous studies of manipulations to the intra-cage environment (inanimate enrichment within the primary enclosure (Schapiro et al., 1993; submitted Schapiro and Bloomsmith, 1994, 1995)). In the present study, almost all time spent in abnormal behaviors by Group 4 subjects was spent pacing, as compared with only 72% of time spent behaving abnormally by Group 1 subjects. Group 1 monkeys, however, spent four times the amount of time in self-directed aggression as did Group 4. Repetitive locomotor patterns (pacing) are aesthetically unde- sirable and are abnormal by a variety of definitions (Mason, 1991) , but they do not seem to be physically harmful; none of our monkeys have exhibited any pacing-related injuries. However, several monkeys have required veterinary treatment for self-induced bite wounds, highlighting the potential danger of self-aggressive behavior. Other researchers have made similar qualitative distinctions between cage associated stereotypies (e.g. pacing) and deprivation acts (e.g. self-directed aggression; Berkson, 1967; Erwin and Deni, 1979; Line, 1987; Mason, 1991).
Group 4 monkeys singly caged in an outdoor building seemed to pace in response to husbandry manipulations in the outdoor colony, particularly to the capture and initial restraint of group-housed monkeys. Similar husbandry routines have been demonstrated to disturb the behavior of macaques in other colonies (Line et al., 1989a, 1991; Crockett and Bowden, 1994). Sanitization of the outdoor buildings in our colony required that monkeys be removed from their primary enclosure and temporarily restrained in a portable tunnel.

S.J. Schapiro et al. /Applied Animal Behaviour Science 45 (1995) 151-163 1.59
Most monkeys were coerced into moving from their primary to the temporary holding enclosures by showing them a net as a negative reinforcer. Singly caged yearlings began to pace in response to the appearance or sound of the net and tunnels and to the sound of other monkeys’ alarmcalls to the net or tunnels. For portions of approximately 60% of observation days for Group 4 subjects, one building in the outdoor colony was being sanitized. In contrast, Group l’s indoor rooms were sanitized on a single day every 2 weeks; no nets were used, and no observations were conducted on those days. Therefore, the higher levels of pacing exhibited by outdoor-housed, singly caged monkeys may be attributable to the frequent manipulations of the colony by caregivers using nets and tunnels. Being outdoors and seeing social interactions in the colony positively affects some behaviors, but we have also demonstrated that potentially negative stimulation arises from routine management procedures. Although in our colony, outdoor-housed monkeys had sensory access to social groups and indoor-housed subjects did not, similar effects would be expected if indoor- housed subjects could see social groups as they were captured and restrained. While indoor vs. outdoor housing is an important factor in this study, it is probably not as important in more conventional colonies where social groups as well as singly caged animals would be housed indoors. It seems that singly caged monkeys that benefit from being able to see and hear social groups, will also have to bear the costs of observing human disturbances to the groups (sanitization procedures) on a regular, if infrequent, basis. Table 2 reveals that abnormal behavior decreased similarly for both Group 1 and Group 4 as time spent in the single cage increased (from the physical through the sensory enrichment phases). This suggests that all subjects acclimated to single caging and that Group 4 subjects did not noticeably habituate to the capture and restraint of social groups.
In this study, confinement and repeated capture and restraint of groups were coupled with an inability to escape from the threat. Subjects could run, but they could not get out of the vicinity of the net or tunnel; thus, it may be appropriate to interpret repetitive pacing as an ‘acceptable’ response to this set of conditions (Erwin and Deni, 1979). Continuing from this viewpoint, one of the defining criteria for ascertaining psychological well-being is the subject’s ability to successfully cope with a challenge (Novak and Suomi, 1988; Mason, 1991; Mendoza, 1991) . Continuous and repetitive locomotion may be a successful short- term coping strategy (G. Mason, 1991) to the threat of capture and restraint. If nothing else, it may represent an ‘ecologically valid’ (Sackett, 1968; Erwin and Deni, 1979) response to the stimulus. To stress this point, Group 4 monkeys were not self aggressing (or engaging in other abnormal activities) in response to alarm calls, nets, and/or tunnels. Situations in which alarm calls should be given and responded to, are naturalistic and thus, may not be totally inappropriate experiences; however, we still consider the high level of pacing observed for Group 4 subjects to be undesirable.
The enrichment effects from this study replicated those of an earlier investigation (Schap- iro and Bloomsmith, 1995) in which Group 1 from the current study was also included. In general, the feeding and physical enrichment techniques that we used yielded more species- typical behavior patterns (more feeding and playing; less inactivity, self-grooming, and abnormal behavior), whereas our sensory enrichment program did not. We have studied 130 control and enriched yearling rhesus monkeys in four cohorts across 4 years (Schapiro and Bloomsmith, 1995; and the present investigation), and it is clear that the three-phase enrichment program we used yields overall decreased self-directed grooming and increased

160 S.J. Schapiro et al. /Applied Animal Behaviour Science 4.5 (I 99s) 151-163
play, regardless of the order in which the enrichment conditions were presented and whether subjects were housed with sensory access to social groups or not. Behavioral changes toward levels that are more species-typical, such as those mentioned above, are considered to be indicative of an improvement in psychological well-being as a function of enrichment (Line, 1987; Novak and Suomi, 1988; Schapiro and Bloomsmith, 1995).
Enrichment was used significantly more by Group 1 monkeys, and Group 1 enriched subjects played the most of all groups. These data support previous findings (Schapiro and Bloomsmith, 1995) and strengthen the suggestion that the relevance and attractiveness of within-cage enhancements increases as the amount of stimulation outside of the cage decreases.
One of the most valuable functions of an enrichment program for young singly caged rhesus may be to provide these animals with a set of potentially relevant (i.e. simulating naturalistic) stimuli on which they can focus their behavior. When relevant stimuli are available, young monkeys may be less likely to focus appropriate behavior on inappropriate stimuli (i.e. themselves: lower levels of self-grooming among enriched subjects being the clearest example in our studies). Stimulation from the extra-cage environment may serve a similar function, acting as another set of relevant stimuli on which subjects can focus their behavior, again making them less likely to focus potentially harmful behavior on themselves (lower self-aggression levels among Group 4 subjects in this study). This may be a profitable path of inquiry, but considerably more data are needed.
Is it better for the monkeys to be singly housed with or without sensory access to social groups? The opportunity to observe normal social activity of conspecifics may be beneficial in the long run for socially restricted monkeys (Dienske et al., 1980) and so may outweigh the negative effect of increased pacing. Decreased levels of inactivity and increased time spent feeding among Group 4 subjects more closely approximated species-typical levels, again arguing for housing subjects so that they can see and hear social groups, although decreased time spent playing was a negative and confusing change. That Group 1 enriched subjects played the most and used their enrichment the most suggests that inanimate enrich- ment may be particularly valuable when young rhesus monkeys are housed with little social and ecological stimulation in the extra-cage environment.
5. Conclusions
Yearling rhesus monkeys singly caged in an outdoor building with exposure to social groups and to variable environmental conditions spent less time inactive and playing, and more time feeding, drinking, and behaving abnormally than did similar monkeys singly caged in indoor rooms without access to social groups or environmental variation. A three- phase environmental enrichment program incorporating physical, feeding, and sensory enhancements resulted in a significant decrease in time spent self-grooming and a significant increase in time spent playing. Whereas the effects of within-cage enrichment were consis- tently positive, the effects of the extra-cage environment were not unidirectional. High levels of pacing by monkeys that had sensory access to social groups may have resulted from frequent capture and restraint of groups in the outdoor colony. Repetitive locomotion is an abnormal behavior, but may be an ecologically valid response to capture threats that

S.I. Schapiro et al. /Applied Animal Behaviour Science 45 (1995) 151-163 161
cannot be escaped. Housing singly caged yearling rhesus so that they can see and hear social groups of conspecifics (outdoors or indoors) may be superior to housing them without this stimulation despite the observed increase in pacing activity. A complex extra-cage environ- ment provides socially-restricted subjects with more opportunities to express, observe, and respond to components of the species-typical behavioral repertoire.
Acknowledgements
We thank A. Kessel, M. McDonald, and D. Oubari for dedicated collection of data and D. Glasgow and C.J. Maliniemi for data management efforts. We also thank J. McMahan, S. Buchl, and the rest of the primate section staff. Animals are maintained in facilities approved by the American Association for Accreditation of Laboratory Animal Care and in accordance with current United States Department of Agriculture, Department of Health and Human Services, and National Institutes of Health regulations and standards. Financial support for this project came from the National Institutes of Health/NCRR grants U42- RR05080 and ROl-RR05092.
References
Altmann, J., 1974. Observational study of behavior: Sampling methods. Behaviour, 49: 227-267. Bayne, K., Mainzer, H., Dexter, S., Campbell, CL, Yamada, F. and Suomi, S., 1991. The reduction of abnormal
behaviors in individually housed rhesus monkeys (Macaca mulatta) with a foraging/grooming board. Am. J. Primatol., 23: 23-35.
Bayne, K., Dexter, S., Mainzer, H., McCully, C., Campbell, G. and Yamada, F., 1992. The use of artificial turf as a foraging substrate for individually housed rhesus monkeys (Macaca mulatta). Anim. Welf., 1: 39-53.
Berkson, G., 1967. Abnormal stereotyped motor acts. In: J. Zubin and H. Hunt (Editors), Comparative Psycho- pathology - Animals and Humans. Grune and Stratton, New York, pp. 76-94.
Bloomsmith, M.A., Alford, P.L. and Maple, T.L., 1988. Successful feeding enrichment for captive chimpanzees. Am. J. Primatol., 16: 155-164.
Bloomsmith, M.A., Finlay, T.W., Merhalski, J.J. and Maple, T.L., 1990. Rigid plastic balls as enrichment devices for captive chimpanzees. Lab. Anim. Sci., 40: 319-322.
Brent, L., Lee, D.R. and Eichberg, J.W., 1989. Evaluation of two environmental enrichment devices for singly caged chimpanzees (Pan troylodytes). Am. J. Primatol., Supp. 1: 65-70.
Bryant, C.E., Rupniak, N.M.J. and Iversen, S.D., 1988. Effects of different environmental enrichment devices on cage stereotypies and autoaggression in captive cynomolgus monkeys. J. Med. Primatol., 17: 257-269.
Chamove, A.S. and Rohrhuber, B., 1989. Moving Callitrichid monkeys from cages to outside areas. Zoo Biol., 8: 151-163.
Coe, CL., 1991. Is social housing ofprimates always the optimal choice? In: M.A. Novak and A.J. Petto (Editors), Through the Looking Glass: Issues of Psychological WeBbeing in Captive Nonhuman Primates. American Psychological Association, Washington, DC, pp. 78-92.
Crockett, C.M. and Bowden, D.M., 1994. Challenging conventional wisdom for housing monkeys. Lab Anim., 23: 29-33.
Crockett, C.M., Bowers, C.L., Sackett, G.P. and Bowden, D.M., 1993. Urinary cortisol responses of longtailed macaques to five cage sizes, tethering, sedation, and room change. Am. J. Primatol., 30: 55-74.
Crockett, CM., Bowers, C.L., Bowden, D.M. and Sackett, G.P., 1994. Sex differences in compatibility of pair- housed adult longtailed macaques. Am. J. Primatol.. 32: 73-94.
Dienske, H., van Vreeswijk, W. and Koning, H., 1980. Adequate mothering by partially isolated rhesus monkeys after observation of maternal care. J. Abn. Psych., 89: 489-492.

162 S.J. Schapiro et al. /Applied Animal Behaviour Science 45 (1995) 151-163
Erwin, J. and Deni, R., 1979. Strangers in a strange land: Abnormal behaviors or abnormal environments? In: J. Erwin, T.L. Maple and G. Mitchell (Editors), Captivity and Behavior: Primates in Breeding Colonies, Lab- oratories and Zoos. Van Nostrand Reinhold, New York, pp. 1-28.
Goerke, B., Fleming, L. and Creel, M., 1987. Behavioral changes of a juvenile gorilla after a transfer to a more naturalistic environment. Zoo Biol., 6: 283-295.
Line, SW., 1987. Environmental enrichment for laboratory primates. J. Am. Vet. Med. Assoc., 190: 854-859. Line, SW. and Morgan, K.N., 1991. The effects of two novel objects on the behavior of singly caged adult rhesus
macaques. Lab. Anim. Sci., 41: 365-369. Line, S.W., Morgan, K.N., Markowitz, H. and Strong, S., 1989a. Heart rate and activity of rhesus monkeys in
response to routine events. Lab. Prim. News]., 28: 9-12. Line, S.W., Morgan, K.N., Markowitz, H. and Strong, S., 1989b. Influence of cage size on heart rate and behavior
in rhesus monkeys Am. J. Vet. Res., 50: 1523-1526. Line, S.W., Morgan, K.N. and Markowitz, H., 1991. Simple toys do not alter the behavior of aged rhesus monkeys.
Zoo Biol., 10: 473-484. Line, SW., Markowitz, H., Morgan, K.N. and Strong, S., 1991. Effects of cage size and environmental enrichment
on behavioral and physiological responses of rhesus macaques to the stress of daily events. In M.A. Novak and A.J. Petto (Editors), Through the Looking Glass: Issues of Psychological Well-being in Captive Non- human Primates. American Psychological Association, Washington, DC, pp. 160-179.
Line, S.W., Morgan, K.N., Markowitz, H. and Strong, S., 1990. Increased cage size does not alter heart rate or behavior in female rhesus monkeys. Am. J. Primatol., 20: 107-l 13.
Markowitz, H. and Line, S., 1989. Primate research models and environmental enrichment. In: E. Segal (Editor), Housing, Care and Psychological Wellbeing of Captive and Laboratory Primates Noyes Publications, Park Ridge, NJ, pp. 203-212.
Mason, G.J., 1991. Stereotypies: acritical review. Anim. Behav., 41: 1015-1037. Mason, W.A., 1991. Effects of social interaction on well-being: Development aspects. Lab. Anim. Sci., 41: 323-
328. Mendoza, S.P., 1991. Sociophysiology of well-being in nonhuman primates. Lab. Anim. Sci., 41: 344-349. Nieuwenhuijsen, K. and de Waal, F.B.M., 1982. Effects of spatial crowding on social behavior in a chimpanzee
colony. Zoo Biol., 1: 5-28. Noldus, L.P.J.J., 1991. The observer: A software system for collection and analysis of observational data. Beh.
Res. Meth. Instr. Comp., 23: 415-429. Novak, M.A. and Suomi, S.J., 1988. Psychological well-being of primates in captivity. Am. Psychol., 43: 765-
773. Novak, M.A. and Bayne, K., 1991. Monkey behavior and laboratory issues: Introduction. Lab. Anim. Sci., 41:
306-307. Novak, M.A., O’Neill, P. and Suomi, S.J., 1992. Adjustments and adaptations to indoor and outdoor environments:
Continuity and change in young adult rhesus monkeys. Am. J. Primatol., 28: 125-138. Novak, M.A., Musante, A., Munroe, H., O’Neill, P.L., Price, C. and Suomi, S.J., 1993. Old, socially housed rhesus
monkeys manipulate objects. Zoo Biol., 12: 285-298. O’Neill, P., 1989. A room with a view for captive primates: Issues, goals, related research and strategies. In: E.
SegaI (Editor), Housing, Care and Psychological Wellbeing of Captive and Laboratory Primates. Noyes Publications, Park Ridge, NJ, pp. 135-160.
O’Neill, P., Price, C. and Suomi, S.J., 1990. Designing captive primate environments sensitive to age and gender related activity profiles for rhesus monkeys (Macaca mulatta). AAZPA Reg. Proceed., pp. 546-554.
G’Neill, P.L., Novak, M.A. and Suomi, S.J., 1991. Normalizing laboratory-reared rhesus macaque (Macaca mulutta) behavior with exposure to complex outdoor enclosures. Zoo Biol., 10: 237-245.
Parks, K.A. and Novak, M.A., 1993. Observations of increased activity and tool use in captive rhesus monkeys exposed to troughs of water. Am. J. Primatol., 29: 13-25.
Ruppenthal, G.C., Walker, C.G. and Sackett, G.P., 1991. Rearing infant monkeys (Macaca nemestrina) in pairs produces deficient social development compared with rearing in single cages. Am. J. Primatol., 25: 103-l 13.
Sackett, G.P., 1968. Abnormal behavior in laboratory-reared rhesus monkeys. In: M. Fox (Editor), Abnormal Behavior of Animals. Saunders, Philadelphia, pp. 293-331.
Schapiro, S.J. and Bloomsmith, M.A., 1994. Behavioral effects of enrichment on pair-housed juvenile rhesus monkeys. Am. J. Primatol., 32: 159-170.

S.J. Schapiro et al. /Applied Animal Behaviour Science 45 (1995) 151-163 163
Schapiro, S.J. and Bloomsmith, M.A., 1995. Behavioral effects of enrichment on singly-housed, yearling rhesus monkeys: An analysis including three enrichment conditions and a control group. Am. J. Primatol., 35: 89- 101.
Schapiro, S.J., Bloomsmith, M.A., Porter, L.M. and Suarez, S.A., 1993. Housing conditions and/or age more strongly affect the behavior of young rhesus monkeys than does inanimate enrichment. Am. J. Primatol., 30: 346.
Schapiro, S.J., Bloomsmith, M.A., Suarez, S.A. and Porter, L.M., 1995. The behavior of group-housed, subaduh rhesus monkeys is not affected by inanimate environmental enrichment. (Am. J. Primatol., submitted).
Schapiro, S.J., Lee-Pa&z, D.E., Taylor, L.L., Watson, L., Bloomsmith, M.A. and Petto, A., 1994. Behavioral management of specific pathogen-free (SPF) rhesus macaques: Group formation, reproduction, and parental competence. Lab. Anim. Sci., 44: 229-234.
Related Documents











