The Auditory System (Lectures 7 and 8) Harry R. Erwin, PhD COMM2E University of Sunderland

The Auditory System (Lectures 7 and 8) Harry R. Erwin, PhD COMM2E University of Sunderland.
Dec 16, 2015
Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

The Auditory System (Lectures 7 and 8)
Harry R. Erwin, PhD
COMM2E
University of Sunderland

Background
• This is my speciality.
• I currently have three relevant grant proposals active, in development or in review.
• I supervise research students in this area.

Organization of the Lecture
• Outside to inside.• Issues associated with specific nuclei will be discussed.• How the auditory system works will be addressed, to
the extent that it is currently known.• The goal is to give you insight into how biologically-
inspired robots might hear.• Perhaps you might want to build a system to localize
gunshots. Here’s an approach.

Resources• Webster, Popper, and Fay, 1992, The Mammalian Auditory Pathway:
Neuroanatomy, Springer Handbook of Auditory Research, volume 1.
• Popper and Fay, 1992, The Mammalian Auditory Pathway: Neurophysiology, Springer Handbook of Auditory Research, volume 2.
• Popper and Fay, 1995, Hearing by Bats, Springer Handbook of Auditory Research, volume 5.
• Hawkins, McMullen, Popper and Fay, 1996, Auditory Computation, Springer Handbook of Auditory Research, volume 6.
• Blauert, 1997, Spatial Hearing, revised edition, MIT Press.
• Nolte, 1993, The Human Brain, 3rd edition, Mosby Yearbook.
• Oertel, Fay, and Popper, 2002, Integrative Functions in the Mammalian Auditory Pathway, Springer Handbook of Auditory Research, volume 15.

The auditory system is a typical mammalian sensory system
• The auditory signal is processed by brainstem modules before the information arrives at the cortex.
• Extensive cortical and somatic reafference is used to tune the brainstem processing.
• Supports a series of functions:– Reflexive movements (e.g., startle reflex)– Orientation towards stimuli (attention)– Localization (where is it?)– Classification (what is it?)– Multisensory integration (especially with vision and touch)

Illusions are a basic tool in understanding sensory processing
• Illusions occur when the perceived stimulus does not accurately reflect the actual stimulus.
• Usually reflect specific implementation in the sensory processing.
• Examples of auditory illusions:– The precedence effect
– Elevation illusions produced by filtered sounds(based on the discussion in Middlebrooks, et al., in Oertel, Fay and Popper, 2002)

The Precedence Effect• You can usually localize clicks accurately in the horizontal
dimension. However, when the clicks are separated by a brief delay, you experience an illusion.
• If the interval is < 5 milliseconds (msec), you experience a single sound.
• If the interval is 1-5 msec, the perceived location is determined by the leading click.
• If the interval is < 1 msec, the perceived location is intermediate between the actual locations.
• Earlier and louder clicks influence the perceived location.

Elevation Illusions
• Your external ear (pinna) filters broadband sounds to produce peaks and notches in the spectrum.
• These serve as cues to location, particularly in elevation and resolving the front/back dimension.
• You can apply filters to a sound to confuse this localization.
• The types of confusion that occur give insight into how the cues are processed.

Implications
• The auditory system seems to have a minimum resolution of 1-5 msec.
• There seems to be a trade-off between sound intensity and timing.
• Different cues play different roles in localization.
• Learning is probably important in calibrating (and recalibrating) the auditory system.

Possible Localization Cues
• Azimuth (Jeffress, 1948)– Interaural intensity difference
(IID)
– Interaural phase difference
– Interaural (onset) time difference (ITD)
• Elevation– Spectral shape
– Spectral notch movement
– Interaural line rotation
• Range– Echo delay (biosonar)– Target motion analysis
(TMA)– Near-field stereophonic
audition (triangulation)
• Motion– Doppler shift– Phase shift– Intensity shift

Components of the auditory system
• Neurotransmitters and receptors
• Cell Types
• Neural Circuits
• Overall organization

Neurotransmitters
• Glutamate (Glu)– AMPA receptors—excitatory, fast – NMDA receptors—excitatory, learning, much slower
• Aspartate—excitatory, fast, found in the cochlea.• GABA—standard inhibitory*, very slow.• Glycine—inhibitory*, fast, common in audition• Acetylcholine—excitatory• Various neuromodulators
*Remember the Cl- reversal potential!

Some basic cell types of the auditory brainstem
• Primary-like (PL)
• Primary-like, notch (PL-N)
• Phase-lock (onset)
• Onset, lock (O-L)
• Chopper

Primary-like (PL)
• Their output is similar to the output of auditory neurons, hence the name. Only a few afferents, resulting in some jitter.
• Low threshold current (LTC) K+ channels open quickly and with a low threshold (10-15 mV depolarization from resting).
• A second high threshold K+ current (HTC) then activates at 20+ mV depolarization.
• These cells do not spike repetitively.• Moderate time constant unless previously depolarized.• Function as transducers

Primary-like, notch (PL-N)
• PL with many more afferents, which must sum to threshold. The presence of the notch reflects the very accurate initial spike timing and the following refractory period. Very little jitter.
• Principle cells of the MNTB are PL-N because they are tightly locked to their globular bushy cells.
• Edge detectors

Phase-lock (Onset) and Onset, lock (O-L) cells
• Octopus cells of the PVCN provide an initial well-timed spike (like PL) cells, followed by a low level of activity. However, they phase-lock to low frequency sounds (up to 800 Hz!, higher in some mammals and much higher in some birds). Thick axons = short latencies.
• May function as pitch or coherence detectors. • Sample many (>60) auditory neurons over a 200sec integration
window. LTC and IH (hyperpolarization-activated) potassium channels. Extremely short membrane time constant (~200sec) near their resting potential. Very low input resistance, so they need lots of input to depolarise.

Chopper Cells
• Stellate cells that spike repetitively.
• Have a high-threshold potassium channel, producing a classical Hodgkin-Huxley-like cycle.
• As long as the depolarizing current is sustained, will spike regularly.

Auditory Midbrain Rules of Organization
• Many specialized nuclei, organized into parallel paths.• Convergence at the inferior colliculus (IC), much of it inhibitory or
shunting. Left-to-right reversal at the IC (like vision). Does the IC function like the basal ganglia? We may know in 3 yrs.
• Glycine (Gly) is the most common inhibitory neurotransmitter, probably due to a faster time constant (~1 msec) than GABA (~5 msec). Inhibitory rebound is extensively exploited to produce delayed responses—a cell depolarizing enough to spike after being hyperpolarized.
• Glutamate (Glu) is the usual excitatory neurotransmitter. AMPA receptors are fast subtypes, so a time constant of 200 sec (200x10-6 sec!) is typical. (Brand et al., 2002, in Nature indicate 100 sec for both Gly and Glu, which is probably too low.)

Duration Tuning Mechanisms
• Duration selective neurons seem to use inhibition and inhibitory rebound.
• Involves inhibitory circuits. Can be modulated.• Initially, a duration-selective neuron is inhibited from
firing in response to a sound.• When the inhibition is released by the end of the
sound, the neuron depolarizes for a short interval.• If delayed excitation arrives while the neuron is
depolarizes, it spikes. Otherwise it remains silent.

Break

Stages in Mammalian Audition(Lecture 8)
• External Ear (Pinnae)
• Middle Ear
• Inner Ear (Cochlea)
• Inner Hair Cells
• Type I Spiral Ganglion Cells
• Cochlear Nucleus (dorsal CN and ventral CN)
• Medial Nucleus of the Trapezoidal Body
• Lateral Superior Olivary Nucleus
• Medial Superior Olivary Nucleus
• Lateral Lemniscus
• Central Nucleus of the Inferior Colliculus
• Medial Geniculate Nucleus
• Auditory Cortex
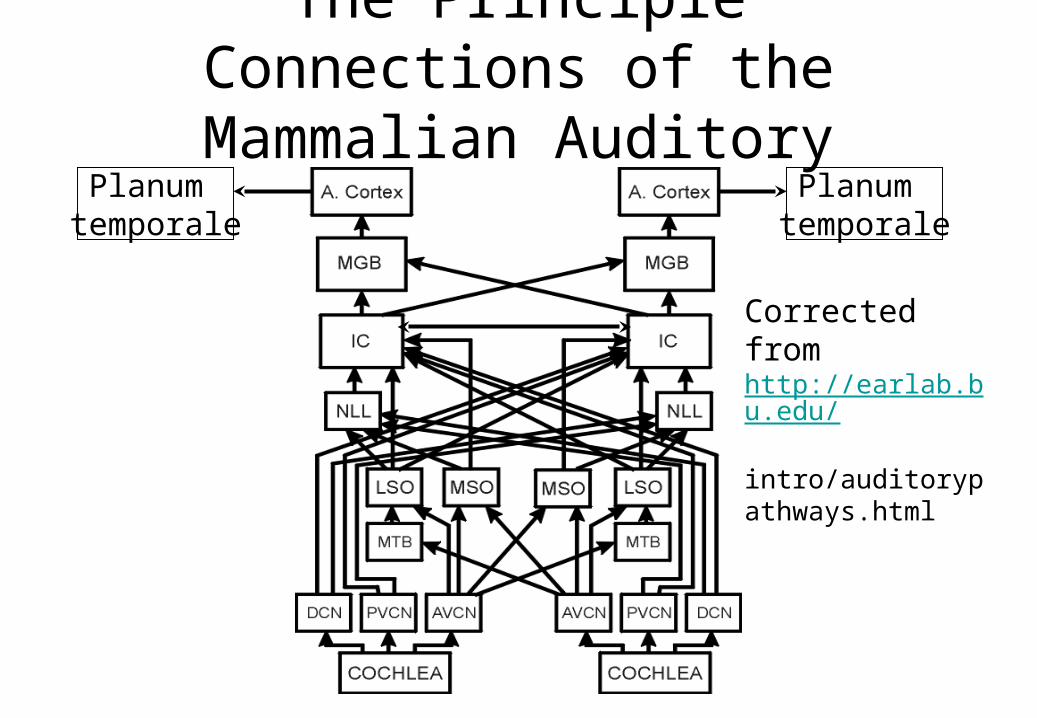
The Principle Connections of the Mammalian Auditory System
Corrected fromhttp://earlab.bu.edu/ intro/auditorypathways.html
Planum temporale
Planum temporale

External Ears (Pinnae)
• Directional receivers, steerable in many mammals.• The transfer function between the free-field sound
(with head not present) and the sound at the ear drum is called the HRTF (head-related transfer function).
• Multipath interference occurs and seems to play a role in generating elevation cues.
• Intensity, onset time, and phase differences between the two ears seem to play a role in estimating azimuth

The Middle Ear• Contains the stapes, incus, and
malleus.• Translates the motion of the ear
drum into pressure waves in the cochlea (inner ear).
• In the bat, muscles of the middle ear contract or relax to mute the sound of its cry and possibly to normalize the intensity of the echo based on distance to the target.
• Figure from http://oto.wustl.edu/cochlea/ intro1.htm
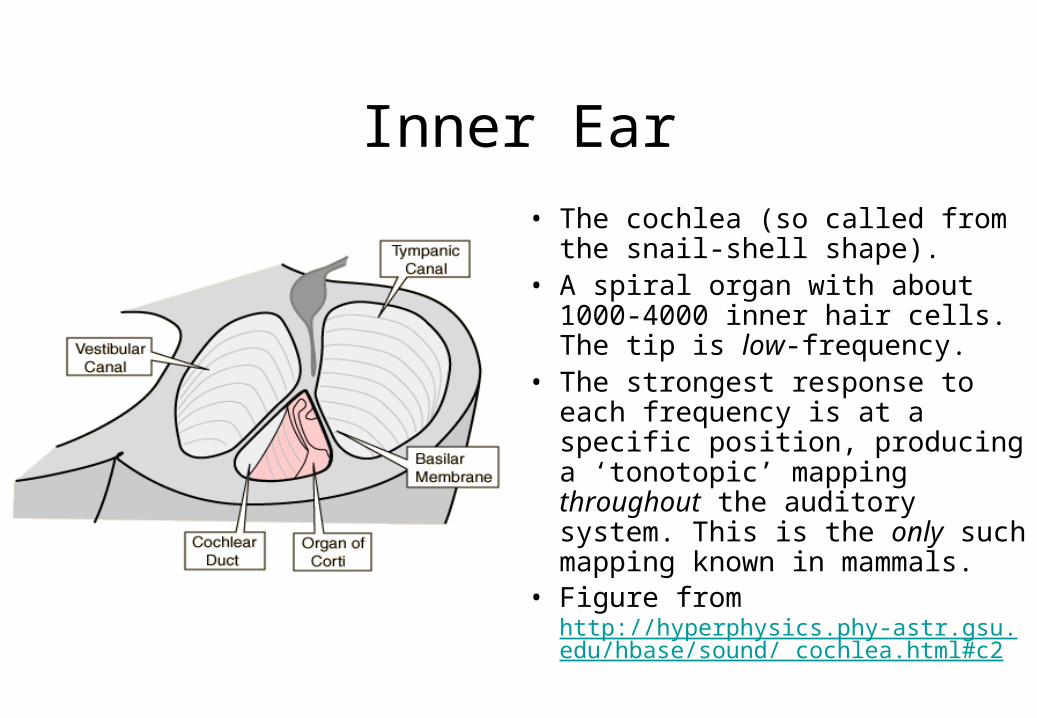
Inner Ear
• The cochlea (so called from the snail-shell shape).
• A spiral organ with about 1000-4000 inner hair cells. The tip is low-frequency.
• The strongest response to each frequency is at a specific position, producing a ‘tonotopic’ mapping throughout the auditory system. This is the only such mapping known in mammals.
• Figure from http://hyperphysics.phy-astr.gsu.edu/hbase/sound/ cochlea.html#c2

Inner Hair Cell
• Uses an excitatory neurotransmitter (Glu or Asp)
• Vesicle release in response to movement of stereocilia on the apex. Logarithmic response to pressure.
• Fast time constants. Bats can sense time intervals less than 100 nsec, probably by detecting interference.
• Figure from http://www.neurophys.wisc.edu/www/aud/johc.html

How the Inner Hair Cell Works
• Vesicle release appears to reflect Ca++ entry into the cell (Ray Meddis). Motion of the stereocilia modulates K+ influx, which causes Ca++ influx, but there is also background Ca++ leakage, so vesicles are released even without sound input. The release rate varies among synaptic terminals, resulting in variation in sensitivity.
• The auditory neurons that synapse on the inner hair cell use AMPA receptors and have a very short time constant (~200 sec).
• The cochlea functions as a biological FFT.

Outer Hair Cells
• May adjust the motion of the basilar membrane so that a specific 30 dB interval is chosen within the 120 dB range of sounds that can be detected.
• An active cochlear amplifier is likely but not fully proven.
• Would be controlled by reafference from the superior olivary complex (later).
• Figure from http://www.neurophys.wisc.edu/www/aud/johc.html

“Type I Spiral Ganglion Cells of the Eighth Nerve”
• The auditory neurons (ANs), forming the “spiral ganglion”.
• 10 to 70 synapse on each inner hair cell. Bipolar, consisting of a dendritic element, a somatic compartment, and a usually myelinated (non-myelinated in humans, so slower) axonal element that divides in the cochlear nucleus. Can synapse on multiple inner hair cells. Excitatory (Glu). Extremely sharp best frequencies.
• Cover a range of 30 dB in sensitivity.• Show spontaneous activity (up to 140 Hz, Gulick).

How the Spiral Ganglion Cells Work
• Multiple vesicles are often released at the inner hair cell synapses, although one is enough to cause firing.
• Variable vesicle release rates by synapse seem to produce the range of sensitivities seen. Vesicle release reflects Ca++ entry into the cell. Integrate and fire dynamics (Meddis), and spontaneous firing rates reflect this. Some new results.
• Fast time constant ~200-300 µsec. • Collectively can phase lock to a sinusoid up to 3-4
KHz (9 KHz in owls, Carr).

The Cochlear Nucleus (CN)
• The first stage of auditory processing after the cochlea.• At the CN, the auditory neurons divide into two
branches, one dorsal and one ventral. Each branch may terminate on multiple neurons.
• The cochlear nucleus is divided into the dorsal CN, anterior ventral CN, and posterior ventral CN, apparently with different functions. Attention plays a role in the AVCN (Covey).

Dorsal Cochlear Nucleus (DCN)
• Laminar or layered structure. ‘Cerebellar-like’ per Curtis Bell. Seems to play a role in estimating sound elevation. Lesions have subtle effects.
• Somatosensory reafference is received from the thalamic reticular nucleus (TRN), reporting on pinna muscle activity. Issues here. Startle reflex.
• Glycinergic primary cells in the DCN appear to respond to lines and notches centered on their best frequencies, reporting to the IC.
• Complex inhibitory circuits in the DCN involving sensory profiles produce this response.

DCN Circuits
• DCN cells participate in circuits that integrate somatosensory data with sound.
• Also detect spectral notches in the signal with moderate width.
• Output chopper, onset, and build-up patterns.

Ventral Cochlear Nucleus (VCN)
• Bushy cells (in the AVCN) are ‘primary-like’ cells that track the spiking of auditory nerve cells directly. These have a dendritic element, a soma, and a myelinated axon that passes to the superior olivary complex, the lateral lemniscus, and to the inferior colliculus, with excitatory signalling.
• Multipolar or stellate cells (in the DCN, AVCN, and PVCN) project to the pontine tegmentum (SOC and LL). These are ‘chopper’ cells that periodically modulate the input signal.
• Octopus cells (PVCN) appear to be broadly tuned onset detectors. Insensitive to intensity. Project to the pontine tegmentum and then to the lateral lemniscus.

Superior Olivary Complex (SOC)
• Consists of the– Lateral superior olivary nucleus (LSO)– Medial superior olivary nucleus (MSO)– Medial nucleus of the trapezoid body (MNTB)
• Size of the complex varies greatly among species as do the sizes of the individual nuclei.
• In bats, cell counts of about 20000 (Ellen Covey, personal communication)
• Secondary nuclei present as well.• Plays a role in the stapedius reflex which protects the middle
ear from loud sounds.

Trapezoidal Body
• Large multipolar principal cells. Synaptic input is via very large calyceal endings (end-bodies of Held). PL-N dynamics.
• Input from globular bushy cells in the contralateral AVCN.
• Reverses the sign of the signal. Inhibitory output.• Projects to the ipsilateral LSO and LL. Glycinergic.• High-frequency sensitive.• Not considered important in humans.

Lateral Superior Olivary Nucleus
• Ipsilateral—excitatory, spherical bushy cells (PL)• Contralateral—inhibitory input via the trapezoidal body
—globular bushy cells (PL-N)• Outputs bilaterally to the lateral lemnisci and to the IC.
Glycine with glutamate or possibly aspartate. Mostly chopper cells.
• Sensitive to high frequency sounds and used for comparing the signal intensities at each ear.
• Small multipolar principal cells.• Codes for auditory localization in azimuth.

Medial Superior Olivary Nucleus• Excitatory input from both sides into separate dendrites.
Source: spherical bushy cells that track the afferent signal. • Feeds forward to the inferior colliculus, mostly ipsilateral.• Generates one or two spikes at sound onset. Other roles
possibly present.• Important in large mammals with good low-frequency
hearing (sounds are diffracted, so intensity is not a good cue for azimuth). Note that phase ambiguity disappears over multiple frequencies.

Nucleus of the Central Acoustic Tract
• Small, importance unknown
• Directly projects to SC and MGB, bypassing IC.
• Large multipolar neurons
• Bilateral input from the AVCN

Lateral Lemniscus
• Major auditory tract. Contains 2nd, 3rd, and 4th order axons. Seems to perform spectral analysis (e.g., vowel detection, line spectra tracking) and detection of transients, and to have a role in measuring the timing of echoes. (Like the Basal Ganglia…?)
• Octopus cell axons end in the ventral nucleus of the lateral lemniscus (VNLL), with large calyceal endings. Part of the short latency acoustic startle reflex pathway to the reticular formation. Monaural. Transient detection.
• Stellate, bushy (excitatory) and MNTB, DCN (minor, glycinergic) cells also project to the VNLL.
• VNLL is glycinergic. Choppers and PL. On-going research area.• DNLL inputs binaural. Projects to the IC. Functional role unknown.

Central Nucleus of the Inferior Colliculus (Mesencephalon)
• Largest auditory structure of the brainstem on the roof of the midbrain. A tectal structure behind the superior colliculus (SC). There is a spatial mapping from the IC to the SC (that triggers visual orientation to sounds in barn owl and possibly in mammals).
• Primary point of convergence in the auditory brainstem. Sounds arrive here 2-5 msec after the inner hair cells are activated.
• Bidirectional connectivity with the auditory cortex. Excitatory inputs are received from the part of the AC (layer V) that then receives the outputs. This is fast enough to support cortically-controlled analysis of current sound afference.

IC components
• Small multipolar fusiform cells with tufted dendrites. Cochleotopic = tonotopic laminar organization, uniting inputs from all lower nuclei and the contralateral IC.
• The anterior portion of the laminae receive cortical inputs, while the posterior portion receives brainstem and IC inputs.
• Stellate cells also present that cross the laminae.• Recently it has been found that the signal at the IC is normalized
in intensity. Several possible mechanisms.• Partly ‘cerebellar-like’ (Curtis Bell). • Match/mismatch processing? Sparsification? Motion processing?• My current grant is in this area.

Medial Geniculate Nucleus (or Body)
• AKA the auditory thalamus. Similar to the LGN (vision).
• Transduces the output of the colliculi for the auditory cortex. Tonotopically organized. In bats, may encode distance.
• Ventral, dorsal, and medial (or magnocellular) divisions.
• Ventral division—about half the structure, projects to primary auditory cortex (A1). Excitatory output.
• Dorsal division—projects to association auditory cortex (A2). Auditory attention? Both excitatory and inhibitory output.
• Medial division—large multipolar neurons. Multisensory arousal system? Both excitatory and inhibitory output.

Cerebellum
• Receives auditory data from the auditory cortex and the pontine nucleus.
• Possible roles include coordinate transformation, motor timing, and localization.

Primary Auditory Cortex (A1)
• Transverse gyri of Heschl• True primary auditory cortex or koniocortex. Called
Area 41, A1, TC, or Kam/Kat depending on the author.• Six-layered. Layer III functions differently from visual
cortex. Strong contralateral connectivity from III, V and VI. Corticalfugal connectivity from V.
• Tonotopically organized with alternating bands responding to a difference signal from the ears (+/-). Sharp tuning and short latencies.
• Some visual sensitivity (from SC and late visual areas)

Secondary Auditory Cortex (A2)
• Parakoniocortex (Area 42, TB, or PB) in this area.• Visual sensitivity.• Multiple tonotopic maps, some complete. Longer latencies,
broader tuning, less sensitive to tones.• In bats, the secondary tonotopic maps are quickly sensitive to
complex sounds.• In mustached bat, there is a secondary area with a
bicoordinate frequency representation over a very narrow frequency interval centered on the second harmonic of the cry.
• Additional fields in bat are ‘delay tuned’.

Planum Temporale
• Smoother portion of the superior surface of the temporal lobe (Area 22 or Tpt)
• Area 22 tends to extend somewhat onto the parietal operculum and inferior parietal lobule in humans.
• On the left side, this is Wernicke’s area.• Areas 39 and 40, the left inferior parietal lobule, is
probably a higher association area.• Now suspected of being the point at which sounds are
correlated to auditory streams. Complex auditory computation. Motion sensitivity? Visual sensitivity.

Other Language Cortices
• An association pathway (arcuate fasciculus) connects Area 22, the inferior parietal lobule (Areas 39 and 40, a complex multimodal integration area), and the area triangularis of the inferior frontal gyrus (Areas 44 and 45, Broca’s area).

Where do things happen?
• Azimuth—binaural, measured in the SOC (MSO, LSO, and MNTB).
• Elevation—monaural, probably based on DCN notch detection.
• Range, timing, and intervals—monaural, measured by the LL, using inhibitory mechanisms.
• Line spectrum—monaural, measured by the LL.
• Sensory integration—for individual sounds, binaurally in the IC, using evidence developed by lower nuclei.
• Comparisons between sounds—auditory cortex.

Reconstructing the acoustic scene
• How separate sound sources are distinguished, assigned to sound streams, and localized is not understood.
• Attention probably chooses sounds out of background. Otherwise, the first sound has preference. Ray Meddis thinks sounds are disambiguated by ignoring ambiguous cues.
• Intervals between sounds are very important in disambiguating them. Auditory neuroscientists are dubious about the ‘binding problem.’
• Distinct sound characteristics are also important in assignment to sound streams. Harmonics important as are spectral segments of about 1 kHz.
• There are a number of interesting auditory illusions that we can explore.

Some lessons to draw
• Dense representations are found throughout the auditory brainstem. The sparse representations needed for associative learning and retrieval seem to be cortical.
• The auditory brainstem has solved the problem of handling (and modulating) duration tuning. This is currently a hard problem in cortical modeling, probably because the role of inhibition and inhibitory rebound is not well-understood. Recent results on persistent activity are important.
• There is no evidence for a spatial map anywhere in the auditory brainstem. This probably means space is represented in spectral form. (Think spatial Fourier transform and Gabor functions.)
• Timing, not synchronization, probably solves the binding problem in the auditory system.
Related Documents











