Molecular Cell, Vol. 7, 981–991, May, 2001, Copyright 2001 by Cell Press The 19S Regulatory Particle of the Proteasome Is Required for Efficient Transcription Elongation by RNA Polymerase II that was first purified by Reinberg and coworkers based upon its ability to stimulate elongation on chromatin templates (LeRoy et al., 1998). Subsequent biochemical studies by Handa and coworkers (Wada et al., 2000) revealed that FACT also stimulates elongation, even on Anwarul Ferdous, Fernando Gonzalez, Liping Sun, Thomas Kodadek, and Stephen Albert Johnston 1 Departments of Internal Medicine and Biochemistry Ryburn Center for Molecular Cardiology nonchromatin templates, by at least one other mecha- University of Texas nism. FACT operates in concert with another factor, Southwestern Medical Center P-TEFb (positive transcription elongation factor) (Mar- 5323 Harry Hines Boulevard shall et al., 1996) to counteract inhibition of elongation Dallas, Texas 75390-8573 by two negative factors DSIF (DRB sensitivity-inducing factor) (Wada et al., 1998) and NELF (negative elongation factor) (Yamaguchi et al., 1999). Summary Analysis of the two polypeptides that comprise mam- malian FACT (Orphanides et al., 1999) revealed that they It is generally thought that the primary or even sole are the homologs of the yeast Cdc68/Spt16 (Malone et activity of the 19S regulatory particle of the 26S protea- al., 1991; Rowley et al., 1991) and Pob3 (Wittmeyer and some is to facilitate the degradation of polyubiquiti- Formosa, 1997) proteins. Cdc68 and Pob3 are known nated proteins by the 20S-core subunit. However, we to form a stable, nuclear-localized heterodimer in yeast present evidence that the 19S complex is required for (Wittmeyer et al., 1999). Genetic experiments in yeast efficient elongation of RNA polymerase II (RNAP II) in support the idea that FACT (Cdc68/Pob3) functions in vitro and in vivo. First, yeast strains carrying alleles of transcription elongation in vivo. Suppressor studies SUG1 and SUG2, encoding 19S components, exhibit demonstrated a genetic interaction between the Cdc68 phenotypes indicative of elongation defects. Second, and the elongation factors TFIIS and Spt4 (Orphanides in vitro transcription is inhibited by antibodies raised et al., 1999). The latter protein and an associated factor, against Sug1, or by heat-inactivating temperature- Spt5, are the yeast analogs of human DSIF (Hartzog et sensitive Sug1 mutants with restoration of elongation al., 1996) and regulate transcription elongation in vivo by addition of immunopurified 19S complex. Finally, (Hartzog et al., 1998). Thus, the genetic and biochemical Cdc68, a known elongation factor, coimmunoprecipi- data present a consistent picture of these positive and tates with the 19S complex, indicating a physical inter- negative regulatory factors modulating elongation. action. Inhibition of the 20S proteolytic core of the The recent realization that Cdc68 is a transcription proteasome has no effect on elongation. This work elongation factor suggests another layer of complexity defines a nonproteolytic role for the 19S complex in in elongation. Xu et al. (1995) had demonstrated that the RNAP II transcription. phenotype of a temperature-sensitive allele of CDC68 (cdc68-1) is suppressed by particular alleles of SUG1 Introduction (sug1-20 and sug1-25). However, both were unable to rescue a cdc68 null allele, indicating that the sug1-20 Eukaryotic transcription is a complex process that can and sug1-25 mutations result in restoration of Cdc68 be divided mechanistically into different steps, including activity. Given that Cdc68 is now recognized as an elon- preinitiation complex (PIC) formation, initiation, elonga- gation factor, it implies that Sug1 may also have a role tion, and termination (Roeder, 1996, and references in elongation. therein). Binding of several general transcription factors On the surface, this is a surprising hypothesis because (GTFs) IIA, IIB, IID, IIE, IIF, and IIH, as well as the RNA Sug1 (also known as Rpt6) is a component of the 26S polymerase II (RNAP II) holoenzyme to a core promoter proteasome, the large multicatalytic protease that de- element is necessary for PIC formation and the initiation grades polyubiquitinated and damaged proteins in all of transcription (Buratowski, 1994; Conaway and Cona- eukaryotic cells (Baumeister et al., 1998; Coux et al., way, 1993). This is followed by promoter escape of 1996). Specifically, Sug1 is one of six AAA (ATPases RNAP II along with some fraction of the proteins in the associated with a variety of cellular activity) ATPases PIC, elongation, and, finally, termination. While it has (Confalonieri and Duguet, 1995) located in the 19S regu- long been appreciated that initiation is subject to com- latory particle of the proteasome (Glickman et al., 1998; plex networks of positive and negative regulation, it has Rubin et al., 1996; Swaffield et al., 1996). The 19S com- only recently become clear that this is also the case for plex sits on each side of a barrel-like catalytic 20S core the elongation phase of the transcription cycle (Cona- particle and is thought to function by binding polyubiqui- way et al., 2000; Reines et al., 1996) tinated factors, unwinding them, and feeding the resul- Recently, several novel factors have been identified tant unstructured chain into the catalytic maw of the 20S and characterized which can regulate RNAP II elonga- core subunit. The six ATPases in the 19S are apparently tion both positively and negatively. Of particular interest critical for this function (Rubin et al., 1998). relative to this study is FACT (facilitates chromatin tran- Given the proteolytic function of Sug1, an obvious scription), a mammalian heterodimeric elongation factor alternative interpretation of the suppression data is that sug1-20 rescues cdc68-1 by decreasing proteasome- mediated decomposition of a structurally labile Cdc68 1 Correspondence: [email protected]

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Molecular Cell, Vol. 7, 981–991, May, 2001, Copyright 2001 by Cell Press
The 19S Regulatory Particle of the ProteasomeIs Required for Efficient Transcription Elongationby RNA Polymerase II
that was first purified by Reinberg and coworkers basedupon its ability to stimulate elongation on chromatintemplates (LeRoy et al., 1998). Subsequent biochemicalstudies by Handa and coworkers (Wada et al., 2000)revealed that FACT also stimulates elongation, even on
Anwarul Ferdous, Fernando Gonzalez,Liping Sun, Thomas Kodadek,and Stephen Albert Johnston1
Departments of Internal Medicine and BiochemistryRyburn Center for Molecular Cardiology
nonchromatin templates, by at least one other mecha-University of Texasnism. FACT operates in concert with another factor,Southwestern Medical CenterP-TEFb (positive transcription elongation factor) (Mar-5323 Harry Hines Boulevardshall et al., 1996) to counteract inhibition of elongationDallas, Texas 75390-8573by two negative factors DSIF (DRB sensitivity-inducingfactor) (Wada et al., 1998) and NELF (negative elongationfactor) (Yamaguchi et al., 1999).Summary
Analysis of the two polypeptides that comprise mam-malian FACT (Orphanides et al., 1999) revealed that theyIt is generally thought that the primary or even soleare the homologs of the yeast Cdc68/Spt16 (Malone etactivity of the 19S regulatory particle of the 26S protea-al., 1991; Rowley et al., 1991) and Pob3 (Wittmeyer andsome is to facilitate the degradation of polyubiquiti-Formosa, 1997) proteins. Cdc68 and Pob3 are knownnated proteins by the 20S-core subunit. However, weto form a stable, nuclear-localized heterodimer in yeastpresent evidence that the 19S complex is required for(Wittmeyer et al., 1999). Genetic experiments in yeastefficient elongation of RNA polymerase II (RNAP II) insupport the idea that FACT (Cdc68/Pob3) functions invitro and in vivo. First, yeast strains carrying alleles oftranscription elongation in vivo. Suppressor studiesSUG1 and SUG2, encoding 19S components, exhibitdemonstrated a genetic interaction between the Cdc68phenotypes indicative of elongation defects. Second,and the elongation factors TFIIS and Spt4 (Orphanidesin vitro transcription is inhibited by antibodies raisedet al., 1999). The latter protein and an associated factor,against Sug1, or by heat-inactivating temperature-Spt5, are the yeast analogs of human DSIF (Hartzog etsensitive Sug1 mutants with restoration of elongational., 1996) and regulate transcription elongation in vivoby addition of immunopurified 19S complex. Finally,(Hartzog et al., 1998). Thus, the genetic and biochemicalCdc68, a known elongation factor, coimmunoprecipi-data present a consistent picture of these positive andtates with the 19S complex, indicating a physical inter-negative regulatory factors modulating elongation.action. Inhibition of the 20S proteolytic core of the
The recent realization that Cdc68 is a transcriptionproteasome has no effect on elongation. This workelongation factor suggests another layer of complexitydefines a nonproteolytic role for the 19S complex inin elongation. Xu et al. (1995) had demonstrated that theRNAP II transcription.phenotype of a temperature-sensitive allele of CDC68(cdc68-1) is suppressed by particular alleles of SUG1Introduction(sug1-20 and sug1-25). However, both were unable torescue a cdc68 null allele, indicating that the sug1-20Eukaryotic transcription is a complex process that canand sug1-25 mutations result in restoration of Cdc68be divided mechanistically into different steps, includingactivity. Given that Cdc68 is now recognized as an elon-preinitiation complex (PIC) formation, initiation, elonga-gation factor, it implies that Sug1 may also have a roletion, and termination (Roeder, 1996, and referencesin elongation.
therein). Binding of several general transcription factorsOn the surface, this is a surprising hypothesis because
(GTFs) IIA, IIB, IID, IIE, IIF, and IIH, as well as the RNASug1 (also known as Rpt6) is a component of the 26S
polymerase II (RNAP II) holoenzyme to a core promoter proteasome, the large multicatalytic protease that de-element is necessary for PIC formation and the initiation grades polyubiquitinated and damaged proteins in allof transcription (Buratowski, 1994; Conaway and Cona- eukaryotic cells (Baumeister et al., 1998; Coux et al.,way, 1993). This is followed by promoter escape of 1996). Specifically, Sug1 is one of six AAA (ATPasesRNAP II along with some fraction of the proteins in the associated with a variety of cellular activity) ATPasesPIC, elongation, and, finally, termination. While it has (Confalonieri and Duguet, 1995) located in the 19S regu-long been appreciated that initiation is subject to com- latory particle of the proteasome (Glickman et al., 1998;plex networks of positive and negative regulation, it has Rubin et al., 1996; Swaffield et al., 1996). The 19S com-only recently become clear that this is also the case for plex sits on each side of a barrel-like catalytic 20S corethe elongation phase of the transcription cycle (Cona- particle and is thought to function by binding polyubiqui-way et al., 2000; Reines et al., 1996) tinated factors, unwinding them, and feeding the resul-
Recently, several novel factors have been identified tant unstructured chain into the catalytic maw of the 20Sand characterized which can regulate RNAP II elonga- core subunit. The six ATPases in the 19S are apparentlytion both positively and negatively. Of particular interest critical for this function (Rubin et al., 1998).relative to this study is FACT (facilitates chromatin tran- Given the proteolytic function of Sug1, an obviousscription), a mammalian heterodimeric elongation factor alternative interpretation of the suppression data is that
sug1-20 rescues cdc68-1 by decreasing proteasome-mediated decomposition of a structurally labile Cdc681Correspondence: [email protected]

Molecular Cell982
mutant. However, the data of Xu et al. (1995) argueagainst this idea, indicating that a more direct role ofthe 19S complex in transcription. Furthermore, thereare other reports of a connection between Sug1 andtranscription. For example, a different allele of sug1 wasshown to suppress a defect caused by truncation ofmost of the activation domain of the Gal4 transcriptionfactor (Swaffield et al., 1992). A second suppressor iso-lated in this study, sug2-1, also encodes a 19S compo-nent (Swaffield et al., 1992, Russell et al., 1996). Sug1has also been identified in two-hybrid experiments asa binding partner of the vitamin D receptor (Masuyamaand MacDonald, 1998). Moreover, biochemical studieshave revealed a potential physical association betweenthe XPB subunit of TFIIH and murine Sug1 (Weeda etal., 1997).
In this study, we employ both in vivo and in vitroapproaches to probe directly the issue of whether the19S complex is involved in RNAP II-mediated transcrip-tion. We show that yeast strains carrying specific SUG1alleles (sug1-20 and sug1-25) are extremely sensitiveto 6-azauracil and poorly induce PUR5 transcription inresponse to nucleotide depletion, phenotypes that arelinked specifically to transcription elongation defects invivo (Exinger and Lacroute, 1992; Powell and Reines,1996; Shaw and Reines, 2000). Activated transcriptionin a yeast whole-cell extract (WCE) is severely inhibitedby either the addition of anti-Sug1 antibodies or heattreatment of extracts made from a temperature-sensi-tive sug1-20 strain. The heat-treated extract can be re-activated upon the addition of immunoaffinity-purified19S or 26S complexes. Experiments using lactacystin,a potent and selective inhibitor of 20S-mediated proteol-ysis (Fenteany et al., 1995), or extracts containing tem-perature-sensitive 20S subunits, argue that the Sug1requirement in transcription does not reflect a critical
Figure 1. The 19S Is Involved in Transcription Elongation In Vivoproteolytic event. Finally, pulse-chase experiments im-Specific alleles of SUG1, which encodes a 19S ATPase, confer sensi-ply that the effect of the 19S complex in transcriptiontivity to 6-azauracil (6-AU) and result in poor induction of PUR5appears to be largely in the elongation phase of the inexpression in response to GTP depletion; phenotypes correlatedvitro reaction. These findings, combined with the geneticwith defects in RNAP II elongation. Mutations in PRE1 and PRE4,
and biochemical work described above, present a com- which encode two of the 20S proteases, do not confer these pheno-pelling argument that the 19S regulatory particle of the types.proteasome plays an important proteolysis-indepen- (A and B) Specific sug1 alleles are 6-AU sensitive, indicating a defect
in transcription elongation. Yeast strains with the indicated geno-dent role in mediating elongation of RNAP II.types were washed and suspended in synthetic complete (SC) me-dium. After 10-fold serial dilutions, 5 ml from each sample was spot-Resultsted onto SC-medium plates either lacking or containing theindicated final concentration of 6-AU.
Evidence that Mutation of SUG1 Inhibits (C) Analysis of IMPDH (PUR5 gene product) expression in wild-typeTranscription Elongation In Vivo and mutant strains. Yeast strains were diluted in SC medium andTo test whether Sug1 is involved in transcription elonga- grown to an OD600 of 0.5–0.6 and treated with 6-AU (100 mg/ml) for
1 hr at 308C. RNA was harvested and subjected to Northern blotting.tion, we compared the sensitivity of congenic SUG1 andFold-induction of PUR5 transcription is shown. Expression ofsug1-20 strains to 6-azauracil (6-AU). 6-AU sensitivitycyclophilin (Cycl) was analyzed as a loading control.correlates well with mutations that cause defects in
RNAP II elongation (Exinger and Lacroute, 1992; Or-phanides et al., 1999; Powell and Reines, 1996). Figures
The correlation between 6-AU sensitivity and a role1A and 1B show that the sug1-20 strain is extremelyin elongation is not perfect. For example, mutations insensitive to 6-AU while the SUG1 strain is relatively in-PPR1 also confer 6-AU sensitivity but have no elonga-sensitive. In contrast to the sug1-20 strain, a strain car-tion phenotype, presumably because PPR1 is directlyrying mutations in the 20S-subunit encoding genesinvolved in uracil metabolism (Exinger and Lacroute,PRE1 and PRE4 is not unusually sensitive to 6-AU (Fig-1992; Shaw and Reines, 2000). Recently, it has beenure 1A). These data suggest that Sug1 and the 19Sshown that, in response to nucleotide depletion, thecomplex are important for transcription elongation inability to induce the PUR5 gene strictly correlates withvivo, but that this effect does not require proteolysis by
the 20S-core subunit of the proteasome. an elongation phenotype (Padilla et al., 1998; Shaw and

The 19S Complex in Transcription Elongation983
Reines, 2000). This gene encodes IMP dehydrogenase Of interest is whether Sug1 protein is the only factor(IMPDH), the rate-limiting enzyme in the de novo GMP in the 19S that affects elongation. Another logical candi-synthesis pathway. To further probe the possible role date would be Sug2, for which there is genetic andof Sug1 in elongation in vivo, we compared PUR5 induc- physical evidence for its involvement in transcription. Intion in wt and mutant strains in the presence of the drug. Figure 2D, we demonstrate that the sug2-13 allele alsoWhen 6-AU was added to a SUG1 strain and PUR5 conditions sensitivity to 6AU. In addition, as for Sug1,induction was assayed by Northern blotting at different antibodies to Sug2 inhibit the in vitro transcription reac-times, maximum induction (.10-fold) was observed tion (Figure 2D). These results support the idea that atafter 1 hr (data not shown; Figure 1C, lanes 2 and 3). least two of the 19S components affect transcription andAs reported previously, disruption of the gene encoding make it unlikely that the effects of Sug1 manipulationelongation-factor SII (Dppr2) confers a 6-AUS phenotype (shown above) are due to free Sug1 protein.(Figure 1B) and reduced induction (only 3.5-fold) of thePUR5 transcription (Figure 1C, lanes 8 and 9) (Or- Addition of Purified 19S Restoresphanides et al., 1999; Shaw and Reines, 2000). Interest- Activated Transcriptioningly, like Dppr2, the sug1-20 strain induces PUR5 tran- To verify that the effect reported in the Figure 2 reflectsscription poorly (lanes 5 and 6). These results corroborate a loss of the Sug1 activity and is not due to an indirectthe 6-AU data and argue that Sug1 and the 19S complex effect, an add-back experiment was performed. Bothare involved in transcription elongation in vivo. the 19S regulatory particle and the intact 26S protea-
To determine if this phenotype is peculiar to the sug1- some were prepared by immunoaffinity chromatography20 allele, we performed the same tests on the sug1-25 from a strain expressing an epitope-tagged Pre1 proteinallele. This mutation occurs in a different domain of (a 20S constituent) (gift of R.J. Deshaies). The 26S prepa-SUG1 than sug1-20 but also suppresses the cdc68-1 ration was obtained by eluting the complex from themutation. As evident in Figures 1B and 1C, the sug1-25 immobilized antibody by competition with excess pep-allele is also 6-AU sensitive and poorly induces PUR5. tide corresponding to the epitope (Verma et al., 2000).Therefore, two different mutations confer the elongation The 19S-complex preparation was obtained by incubat-defective phenotype in vivo. ing the immobilized 26S complex in a buffer that pro-
motes dissociation of the 19S and 20S subunits, thenSug1 Is Important for Efficient Transcription In Vitro eluting the former with high salt. Each preparation wasTo further test for a role of Sug1 in transcription, we analyzed by SDS-polyacrylamide gel electrophoresisturned to in vitro transcription assays. An aliquot of yeast (Figure 3A; note that the two preparations were analyzedWCE prepared from a SUG1 strain and a DNA template, on different gels), Western blotting (Figure 3B), and elec-pGAL4CG- (Woontner and Jaehning, 1990) were incu- trospray mass spectrometry. The predicted 19S proteinsbated with or without the Gal4-VP16 activator for 40 min were detected in both preparations, and bands corre-at room temperature. This was followed by an initiation/ sponding to the 20S constituents were observed onlyelongation step for 15 min in the presence of A/C/UTP, in the 26S preparation (Figures 3A and 3B). That thea time sufficient for most of the G-less transcripts to be 19S preparation was devoid of detectable 20S was alsocompleted (data not shown). Addition of anti-Sug1/Rpt6
evidenced by the lack of any detectable peptidase activ-antibodies in the transcription reactions inhibited acti-
ity (data not shown). Peptides from all of the known 26Svated transcription (Figure 2A, lanes 2 and 4). Control
proteins could be detected by reverse-phase HPLC-antibodies (anti-Gal6) did not inhibit transcription (com-electrospray mass spectrometry after trypsinolysis ofpare lanes 2 and 6). To address the specificity of thethe peptide-eluted sample (data not shown). However,effect of the anti-Sug1 antibody on transcription, puri-peaks corresponding to peptides from 20S proteins orfied 19S complex (Figure 3) was preincubated with anti-known transcription factors were not detected when theSug1 antibodies and then added to the transcription19S sample was analyzed in the same way (data notassay. Figure 2B shows that pre-incubation of the 19Sshown). Some proteins, such as Rad23, that have beenwith anti-Sug1 antibodies indeed significantly alleviatedreported to bind the proteasome were also detected.antibody inhibition in a dose-dependent manner (lanes(Russell et al., 1999a; Schauber et al., 1998).1, 5 and 6). Heat denaturation of the anti-Sug1 antibodies
The results of the add-back experiments using thesedid not inhibit transcription (Figure 2B, lane 3).purified complexes are shown in Figure 4. Addition ofTo further test the involvement of the 19S regulatorythe purified 19S to heat-treated sug1-20 extract largelycomplex in transcription, we took advantage of the factrestored Gal4-VP16-mediated transcription activationthat the sug1-20 allele, isolated as a suppressor of a(Figure 4A, compare lane 2 with lanes 5 and 6). Thistranscriptionally defective mutant of CDC68 (Xu, et al.,activity was not observed when an equivalent amount1995), is temperature sensitive. Yeast WCEs were pre-of heat-denatured 19S was added (lane 7). On the otherpared from congenic wild-type (wt) and sug1-20 strainshand, addition of the same amount of 19S complex hadand tested for in vitro transcription activity with or with-no effect on transcription reactions lacking the chimericout heat treatment. As evident in Figure 2C, the wt ex-activator (compare lanes 3 and 4). Addition of the 19Stract showed only a slight decrease in Gal4-VP16-medi-complex to a heat-treated wt extract (lanes 8–11) or toated transcription activation when it was heat treateduntreated wt and sug1-20 extracts (data not shown) didfor 15 min at 378C (lanes 1–4). On the other hand, tran-not affect transcription levels. The stimulatory effect ofscription was reduced severely in the sug1-20 extractadded 19S complex was dose dependent (Figure 4B,under identical conditions (lanes 5–8). This experimentlanes 6–8).could not be repeated with the sug1-25 strain since the
resultant protein is not temperature sensitive. When these experiments were conducted with the
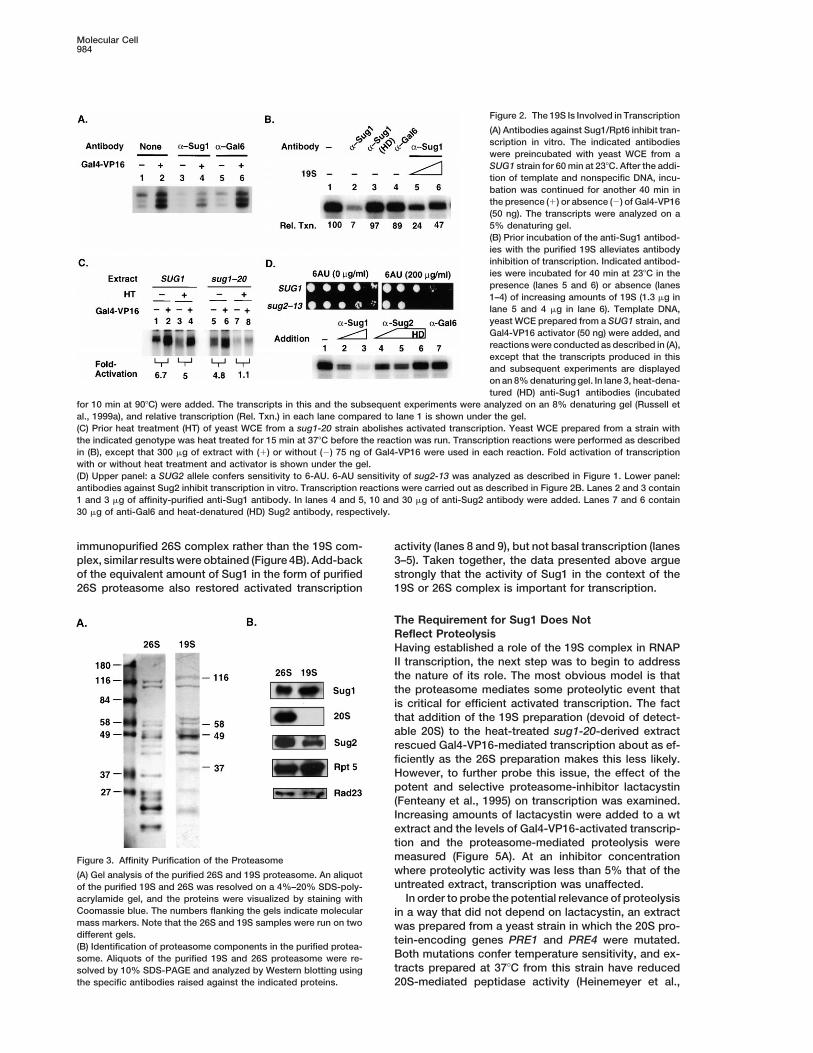
Molecular Cell984
Figure 2. The 19S Is Involved in Transcription
(A) Antibodies against Sug1/Rpt6 inhibit tran-scription in vitro. The indicated antibodieswere preincubated with yeast WCE from aSUG1 strain for 60 min at 238C. After the addi-tion of template and nonspecific DNA, incu-bation was continued for another 40 min inthe presence (1) or absence (2) of Gal4-VP16(50 ng). The transcripts were analyzed on a5% denaturing gel.(B) Prior incubation of the anti-Sug1 antibod-ies with the purified 19S alleviates antibodyinhibition of transcription. Indicated antibod-ies were incubated for 40 min at 238C in thepresence (lanes 5 and 6) or absence (lanes1–4) of increasing amounts of 19S (1.3 mg inlane 5 and 4 mg in lane 6). Template DNA,yeast WCE prepared from a SUG1 strain, andGal4-VP16 activator (50 ng) were added, andreactions were conducted as described in (A),except that the transcripts produced in thisand subsequent experiments are displayedon an 8% denaturing gel. In lane 3, heat-dena-tured (HD) anti-Sug1 antibodies (incubated
for 10 min at 908C) were added. The transcripts in this and the subsequent experiments were analyzed on an 8% denaturing gel (Russell etal., 1999a), and relative transcription (Rel. Txn.) in each lane compared to lane 1 is shown under the gel.(C) Prior heat treatment (HT) of yeast WCE from a sug1-20 strain abolishes activated transcription. Yeast WCE prepared from a strain withthe indicated genotype was heat treated for 15 min at 378C before the reaction was run. Transcription reactions were performed as describedin (B), except that 300 mg of extract with (1) or without (2) 75 ng of Gal4-VP16 were used in each reaction. Fold activation of transcriptionwith or without heat treatment and activator is shown under the gel.(D) Upper panel: a SUG2 allele confers sensitivity to 6-AU. 6-AU sensitivity of sug2-13 was analyzed as described in Figure 1. Lower panel:antibodies against Sug2 inhibit transcription in vitro. Transcription reactions were carried out as described in Figure 2B. Lanes 2 and 3 contain1 and 3 mg of affinity-purified anti-Sug1 antibody. In lanes 4 and 5, 10 and 30 mg of anti-Sug2 antibody were added. Lanes 7 and 6 contain30 mg of anti-Gal6 and heat-denatured (HD) Sug2 antibody, respectively.
immunopurified 26S complex rather than the 19S com- activity (lanes 8 and 9), but not basal transcription (lanes3–5). Taken together, the data presented above argueplex, similar results were obtained (Figure 4B). Add-back
of the equivalent amount of Sug1 in the form of purified strongly that the activity of Sug1 in the context of the19S or 26S complex is important for transcription.26S proteasome also restored activated transcription
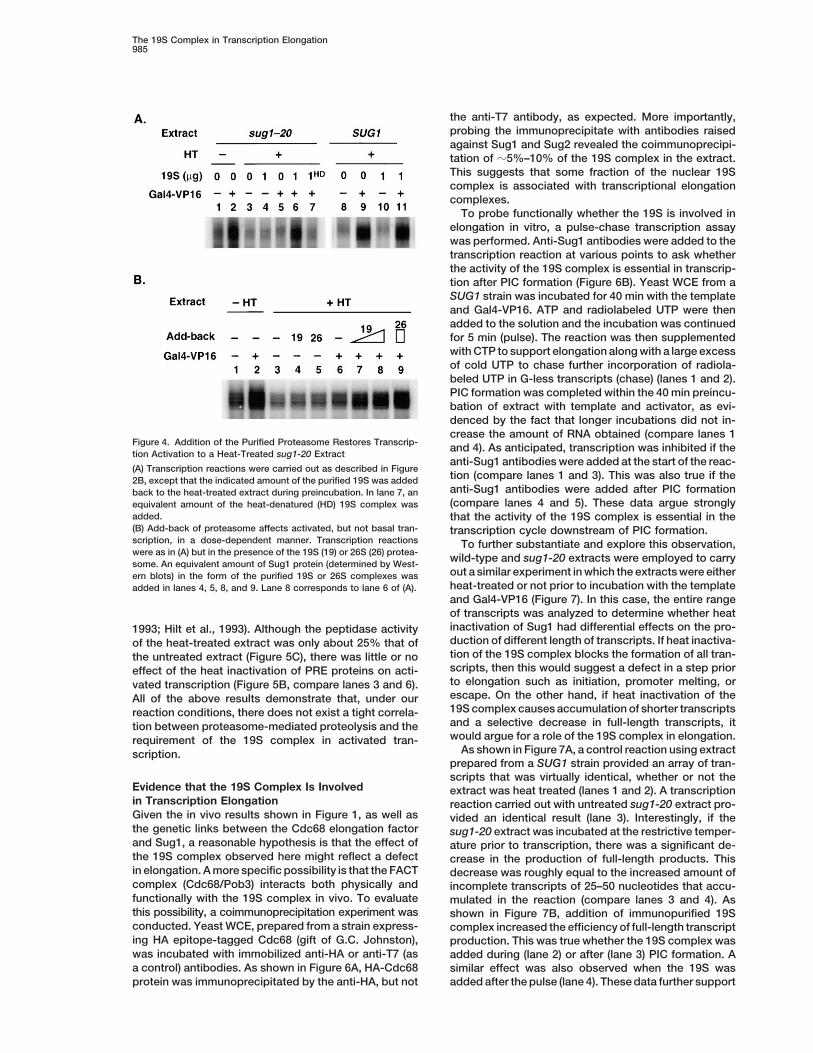
The Requirement for Sug1 Does NotReflect ProteolysisHaving established a role of the 19S complex in RNAPII transcription, the next step was to begin to addressthe nature of its role. The most obvious model is thatthe proteasome mediates some proteolytic event thatis critical for efficient activated transcription. The factthat addition of the 19S preparation (devoid of detect-able 20S) to the heat-treated sug1-20-derived extractrescued Gal4-VP16-mediated transcription about as ef-ficiently as the 26S preparation makes this less likely.However, to further probe this issue, the effect of thepotent and selective proteasome-inhibitor lactacystin(Fenteany et al., 1995) on transcription was examined.Increasing amounts of lactacystin were added to a wtextract and the levels of Gal4-VP16-activated transcrip-tion and the proteasome-mediated proteolysis weremeasured (Figure 5A). At an inhibitor concentrationFigure 3. Affinity Purification of the Proteasomewhere proteolytic activity was less than 5% that of the(A) Gel analysis of the purified 26S and 19S proteasome. An aliquotuntreated extract, transcription was unaffected.of the purified 19S and 26S was resolved on a 4%–20% SDS-poly-
acrylamide gel, and the proteins were visualized by staining with In order to probe the potential relevance of proteolysisCoomassie blue. The numbers flanking the gels indicate molecular in a way that did not depend on lactacystin, an extractmass markers. Note that the 26S and 19S samples were run on two was prepared from a yeast strain in which the 20S pro-different gels. tein-encoding genes PRE1 and PRE4 were mutated.(B) Identification of proteasome components in the purified protea-
Both mutations confer temperature sensitivity, and ex-some. Aliquots of the purified 19S and 26S proteasome were re-tracts prepared at 378C from this strain have reducedsolved by 10% SDS-PAGE and analyzed by Western blotting using
the specific antibodies raised against the indicated proteins. 20S-mediated peptidase activity (Heinemeyer et al.,
The 19S Complex in Transcription Elongation985
the anti-T7 antibody, as expected. More importantly,probing the immunoprecipitate with antibodies raisedagainst Sug1 and Sug2 revealed the coimmunoprecipi-tation of z5%–10% of the 19S complex in the extract.This suggests that some fraction of the nuclear 19Scomplex is associated with transcriptional elongationcomplexes.
To probe functionally whether the 19S is involved inelongation in vitro, a pulse-chase transcription assaywas performed. Anti-Sug1 antibodies were added to thetranscription reaction at various points to ask whetherthe activity of the 19S complex is essential in transcrip-tion after PIC formation (Figure 6B). Yeast WCE from aSUG1 strain was incubated for 40 min with the templateand Gal4-VP16. ATP and radiolabeled UTP were thenadded to the solution and the incubation was continuedfor 5 min (pulse). The reaction was then supplementedwith CTP to support elongation along with a large excessof cold UTP to chase further incorporation of radiola-beled UTP in G-less transcripts (chase) (lanes 1 and 2).PIC formation was completed within the 40 min preincu-bation of extract with template and activator, as evi-denced by the fact that longer incubations did not in-crease the amount of RNA obtained (compare lanes 1
Figure 4. Addition of the Purified Proteasome Restores Transcrip- and 4). As anticipated, transcription was inhibited if thetion Activation to a Heat-Treated sug1-20 Extract
anti-Sug1 antibodies were added at the start of the reac-(A) Transcription reactions were carried out as described in Figure
tion (compare lanes 1 and 3). This was also true if the2B, except that the indicated amount of the purified 19S was addedanti-Sug1 antibodies were added after PIC formationback to the heat-treated extract during preincubation. In lane 7, an(compare lanes 4 and 5). These data argue stronglyequivalent amount of the heat-denatured (HD) 19S complex was
added. that the activity of the 19S complex is essential in the(B) Add-back of proteasome affects activated, but not basal tran- transcription cycle downstream of PIC formation.scription, in a dose-dependent manner. Transcription reactions To further substantiate and explore this observation,were as in (A) but in the presence of the 19S (19) or 26S (26) protea- wild-type and sug1-20 extracts were employed to carrysome. An equivalent amount of Sug1 protein (determined by West-
out a similar experiment in which the extracts were eitherern blots) in the form of the purified 19S or 26S complexes washeat-treated or not prior to incubation with the templateadded in lanes 4, 5, 8, and 9. Lane 8 corresponds to lane 6 of (A).and Gal4-VP16 (Figure 7). In this case, the entire rangeof transcripts was analyzed to determine whether heatinactivation of Sug1 had differential effects on the pro-1993; Hilt et al., 1993). Although the peptidase activityduction of different length of transcripts. If heat inactiva-of the heat-treated extract was only about 25% that oftion of the 19S complex blocks the formation of all tran-the untreated extract (Figure 5C), there was little or noscripts, then this would suggest a defect in a step prioreffect of the heat inactivation of PRE proteins on acti-to elongation such as initiation, promoter melting, orvated transcription (Figure 5B, compare lanes 3 and 6).escape. On the other hand, if heat inactivation of theAll of the above results demonstrate that, under our19S complex causes accumulation of shorter transcriptsreaction conditions, there does not exist a tight correla-and a selective decrease in full-length transcripts, ittion between proteasome-mediated proteolysis and thewould argue for a role of the 19S complex in elongation.requirement of the 19S complex in activated tran-
As shown in Figure 7A, a control reaction using extractscription.prepared from a SUG1 strain provided an array of tran-scripts that was virtually identical, whether or not the
Evidence that the 19S Complex Is Involved extract was heat treated (lanes 1 and 2). A transcriptionin Transcription Elongation reaction carried out with untreated sug1-20 extract pro-Given the in vivo results shown in Figure 1, as well as vided an identical result (lane 3). Interestingly, if thethe genetic links between the Cdc68 elongation factor sug1-20 extract was incubated at the restrictive temper-and Sug1, a reasonable hypothesis is that the effect of ature prior to transcription, there was a significant de-the 19S complex observed here might reflect a defect crease in the production of full-length products. Thisin elongation. A more specific possibility is that the FACT decrease was roughly equal to the increased amount ofcomplex (Cdc68/Pob3) interacts both physically and incomplete transcripts of 25–50 nucleotides that accu-functionally with the 19S complex in vivo. To evaluate mulated in the reaction (compare lanes 3 and 4). Asthis possibility, a coimmunoprecipitation experiment was shown in Figure 7B, addition of immunopurified 19Sconducted. Yeast WCE, prepared from a strain express- complex increased the efficiency of full-length transcripting HA epitope-tagged Cdc68 (gift of G.C. Johnston), production. This was true whether the 19S complex waswas incubated with immobilized anti-HA or anti-T7 (as added during (lane 2) or after (lane 3) PIC formation. Aa control) antibodies. As shown in Figure 6A, HA-Cdc68 similar effect was also observed when the 19S was
added after the pulse (lane 4). These data further supportprotein was immunoprecipitated by the anti-HA, but not
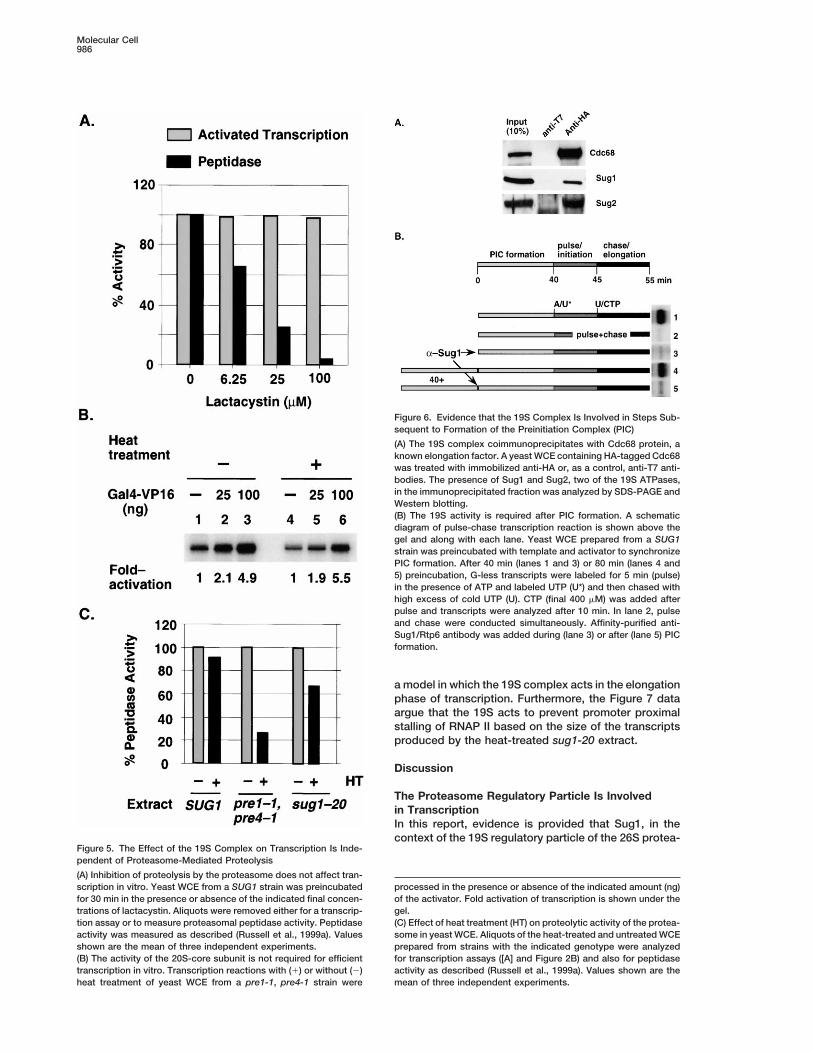
Molecular Cell986
Figure 6. Evidence that the 19S Complex Is Involved in Steps Sub-sequent to Formation of the Preinitiation Complex (PIC)
(A) The 19S complex coimmunoprecipitates with Cdc68 protein, aknown elongation factor. A yeast WCE containing HA-tagged Cdc68was treated with immobilized anti-HA or, as a control, anti-T7 anti-bodies. The presence of Sug1 and Sug2, two of the 19S ATPases,in the immunoprecipitated fraction was analyzed by SDS-PAGE andWestern blotting.(B) The 19S activity is required after PIC formation. A schematicdiagram of pulse-chase transcription reaction is shown above thegel and along with each lane. Yeast WCE prepared from a SUG1strain was preincubated with template and activator to synchronizePIC formation. After 40 min (lanes 1 and 3) or 80 min (lanes 4 and5) preincubation, G-less transcripts were labeled for 5 min (pulse)in the presence of ATP and labeled UTP (U*) and then chased withhigh excess of cold UTP (U). CTP (final 400 mM) was added afterpulse and transcripts were analyzed after 10 min. In lane 2, pulseand chase were conducted simultaneously. Affinity-purified anti-Sug1/Rtp6 antibody was added during (lane 3) or after (lane 5) PICformation.
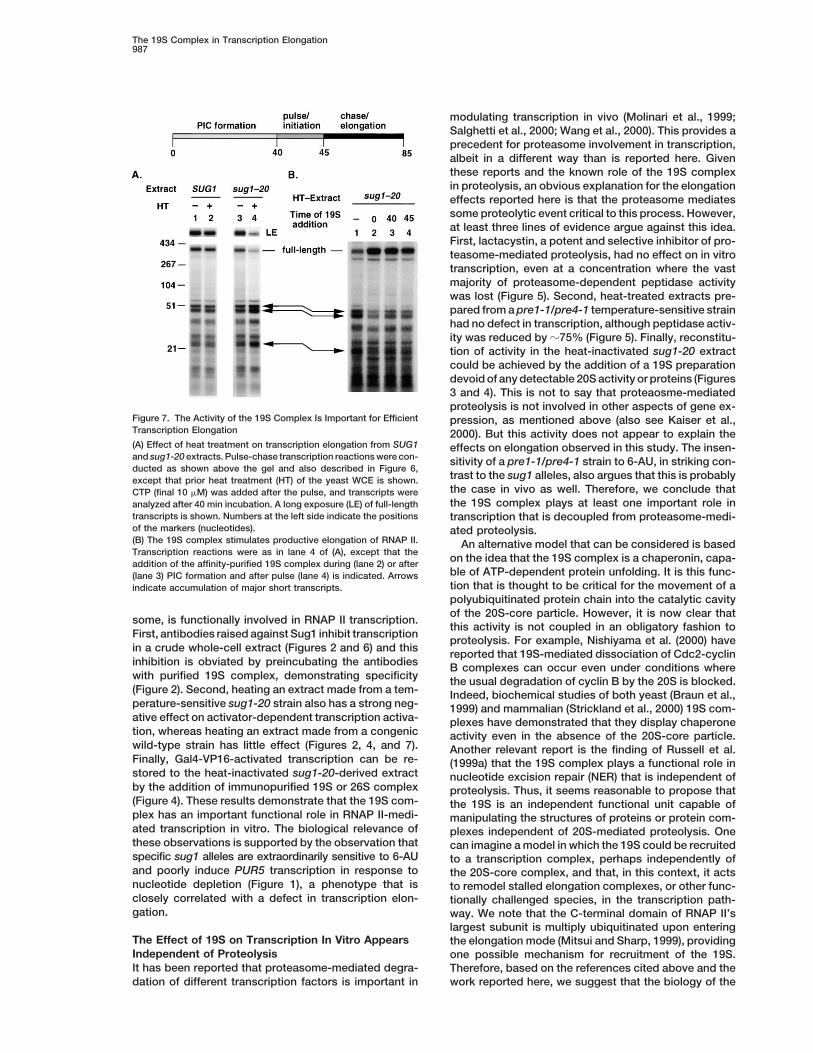
a model in which the 19S complex acts in the elongationphase of transcription. Furthermore, the Figure 7 dataargue that the 19S acts to prevent promoter proximalstalling of RNAP II based on the size of the transcriptsproduced by the heat-treated sug1-20 extract.
Discussion
The Proteasome Regulatory Particle Is Involvedin TranscriptionIn this report, evidence is provided that Sug1, in thecontext of the 19S regulatory particle of the 26S protea-
Figure 5. The Effect of the 19S Complex on Transcription Is Inde-pendent of Proteasome-Mediated Proteolysis
(A) Inhibition of proteolysis by the proteasome does not affect tran-processed in the presence or absence of the indicated amount (ng)scription in vitro. Yeast WCE from a SUG1 strain was preincubated
for 30 min in the presence or absence of the indicated final concen- of the activator. Fold activation of transcription is shown under thegel.trations of lactacystin. Aliquots were removed either for a transcrip-
tion assay or to measure proteasomal peptidase activity. Peptidase (C) Effect of heat treatment (HT) on proteolytic activity of the protea-some in yeast WCE. Aliquots of the heat-treated and untreated WCEactivity was measured as described (Russell et al., 1999a). Values
shown are the mean of three independent experiments. prepared from strains with the indicated genotype were analyzedfor transcription assays ([A] and Figure 2B) and also for peptidase(B) The activity of the 20S-core subunit is not required for efficient
transcription in vitro. Transcription reactions with (1) or without (2) activity as described (Russell et al., 1999a). Values shown are themean of three independent experiments.heat treatment of yeast WCE from a pre1-1, pre4-1 strain were
The 19S Complex in Transcription Elongation987
modulating transcription in vivo (Molinari et al., 1999;Salghetti et al., 2000; Wang et al., 2000). This provides aprecedent for proteasome involvement in transcription,albeit in a different way than is reported here. Giventhese reports and the known role of the 19S complexin proteolysis, an obvious explanation for the elongationeffects reported here is that the proteasome mediatessome proteolytic event critical to this process. However,at least three lines of evidence argue against this idea.First, lactacystin, a potent and selective inhibitor of pro-teasome-mediated proteolysis, had no effect on in vitrotranscription, even at a concentration where the vastmajority of proteasome-dependent peptidase activitywas lost (Figure 5). Second, heat-treated extracts pre-pared from a pre1-1/pre4-1 temperature-sensitive strainhad no defect in transcription, although peptidase activ-ity was reduced by z75% (Figure 5). Finally, reconstitu-tion of activity in the heat-inactivated sug1-20 extractcould be achieved by the addition of a 19S preparationdevoid of any detectable 20S activity or proteins (Figures3 and 4). This is not to say that proteaosme-mediatedproteolysis is not involved in other aspects of gene ex-
Figure 7. The Activity of the 19S Complex Is Important for Efficient pression, as mentioned above (also see Kaiser et al.,Transcription Elongation 2000). But this activity does not appear to explain the(A) Effect of heat treatment on transcription elongation from SUG1 effects on elongation observed in this study. The insen-and sug1-20 extracts. Pulse-chase transcription reactions were con- sitivity of a pre1-1/pre4-1 strain to 6-AU, in striking con-ducted as shown above the gel and also described in Figure 6,
trast to the sug1 alleles, also argues that this is probablyexcept that prior heat treatment (HT) of the yeast WCE is shown.the case in vivo as well. Therefore, we conclude thatCTP (final 10 mM) was added after the pulse, and transcripts werethe 19S complex plays at least one important role inanalyzed after 40 min incubation. A long exposure (LE) of full-length
transcripts is shown. Numbers at the left side indicate the positions transcription that is decoupled from proteasome-medi-of the markers (nucleotides). ated proteolysis.(B) The 19S complex stimulates productive elongation of RNAP II. An alternative model that can be considered is basedTranscription reactions were as in lane 4 of (A), except that the
on the idea that the 19S complex is a chaperonin, capa-addition of the affinity-purified 19S complex during (lane 2) or afterble of ATP-dependent protein unfolding. It is this func-(lane 3) PIC formation and after pulse (lane 4) is indicated. Arrowstion that is thought to be critical for the movement of aindicate accumulation of major short transcripts.polyubiquitinated protein chain into the catalytic cavityof the 20S-core particle. However, it is now clear that
some, is functionally involved in RNAP II transcription.this activity is not coupled in an obligatory fashion to
First, antibodies raised against Sug1 inhibit transcriptionproteolysis. For example, Nishiyama et al. (2000) have
in a crude whole-cell extract (Figures 2 and 6) and this reported that 19S-mediated dissociation of Cdc2-cyclininhibition is obviated by preincubating the antibodies B complexes can occur even under conditions wherewith purified 19S complex, demonstrating specificity the usual degradation of cyclin B by the 20S is blocked.(Figure 2). Second, heating an extract made from a tem- Indeed, biochemical studies of both yeast (Braun et al.,perature-sensitive sug1-20 strain also has a strong neg- 1999) and mammalian (Strickland et al., 2000) 19S com-ative effect on activator-dependent transcription activa- plexes have demonstrated that they display chaperonetion, whereas heating an extract made from a congenic activity even in the absence of the 20S-core particle.wild-type strain has little effect (Figures 2, 4, and 7). Another relevant report is the finding of Russell et al.Finally, Gal4-VP16-activated transcription can be re- (1999a) that the 19S complex plays a functional role instored to the heat-inactivated sug1-20-derived extract nucleotide excision repair (NER) that is independent ofby the addition of immunopurified 19S or 26S complex proteolysis. Thus, it seems reasonable to propose that(Figure 4). These results demonstrate that the 19S com- the 19S is an independent functional unit capable ofplex has an important functional role in RNAP II-medi- manipulating the structures of proteins or protein com-ated transcription in vitro. The biological relevance of plexes independent of 20S-mediated proteolysis. Onethese observations is supported by the observation that can imagine a model in which the 19S could be recruitedspecific sug1 alleles are extraordinarily sensitive to 6-AU to a transcription complex, perhaps independently ofand poorly induce PUR5 transcription in response to the 20S-core complex, and that, in this context, it actsnucleotide depletion (Figure 1), a phenotype that is to remodel stalled elongation complexes, or other func-closely correlated with a defect in transcription elon- tionally challenged species, in the transcription path-gation. way. We note that the C-terminal domain of RNAP II’s
largest subunit is multiply ubiquitinated upon enteringThe Effect of 19S on Transcription In Vitro Appears the elongation mode (Mitsui and Sharp, 1999), providingIndependent of Proteolysis one possible mechanism for recruitment of the 19S.It has been reported that proteasome-mediated degra- Therefore, based on the references cited above and the
work reported here, we suggest that the biology of thedation of different transcription factors is important in

Molecular Cell988
19S complex is much richer than was anticipated pre- cycle. It is true that under the conditions of our in vitroassays, there does not appear to be a major effect on theviously. In fact, the functions of the 19S complex in
cellular metabolism may reach beyond even proteolysis, synthesis of short transcripts when Sug1 is inactivated(Figure 7). However, the experimental protocol allows atranscription, and NER. Wittmeyer et al. (1999) have
found that some fraction of yeast FACT (the Cdc68/ 40 min incubation for PIC formation, and naked DNAtemplates were employed here. This may not reflect thePob3 heterodimer) copurifies with DNA polymerase a.
Given the aforementioned genetic link between sug1- more challenging conditions of promoter occupancy invivo, particularly on chromatin templates. More work20 and cdc68-1, it is therefore possible that the 19S
complex could function in replication and perhaps other will be required to probe a possible role of the 19Scomplex in a phase of the transcription cycle that pre-processes.cedes elongation.
The 19S Complex Modulates Transcription Elongationof RNAP II In Vitro and In Vivo Genetic Data Suggest Other 19S EffectsOur data argue strongly that the Sug1 protein in the on Transcriptioncontext of 19S complex is involved in transcription elon- Mutations other than the sug1-20 allele also suggest agation. Specifically, inactivation of the Sug1 protein link between the 19S and transcription. The SUG1 genedoes not alter the synthesis of short transcripts substan- was first discovered on the basis of an allele, sug1-1,tially, but results in a significant inhibition of full-length that suppressed a transcriptional defect in GAL4 (Swaf-products. Addition of immunopurified 19S complex to field et al., 1992). A mutation in the SUG2 gene (sug2-1),these stalled reactions reconstitutes synthesis of full- which encodes another 19S subunit, also suppressedlength RNAs (Figure 7). Again, the biochemistry seems the GAL4 defect. The sug1-1 and sug2-1 mutations areto correlate with the genetics. The sensitivity of the sug1- recessive, indicating that they result in the loss of some20 and sug1-25 strains to 6-AU (Figure 1) is indicative activity of the wild-type proteins that inhibits Gal4-medi-of a defect in elongation in vivo. This is further supported ated activation. This apparent negative effect is not eas-by the fact that in response to nucleotide depletion the ily explained by the requirement for Sug1 and the 19Ssug1-20 strain shows a defect in induction of PUR5 complex in transcription elongation that we have dem-transcription similar to that reported for a SII-disrupted onstrated. Therefore, we predict that the 19S complex(Dppr2) strain (Figure 1) (Shaw and Reines, 2000). We plays one or more roles in transcription in addition toalso showed that at least one other SUG1 allele, sug1- stimulating elongation, at least one of which is inhibitory.25 has an identical phenotype (Figure 1), suggesting This view is consistent with the fact that the suppressionthat the elongation defect is not specific to the sug1- of cdc68-1 by sug1-20 is allele specific; sug1-1 has no20 mutation. Coimmunoprecipitation of Cdc68 and the effect on cdc68-1, nor does sug1-20 rescue GAL4 de-19S (Figure 6A) further supports the genetic link between fects (Russell and Johnston, 2001), even though bothCDC68, which encodes a component of a known elonga- mutations confer similar effects on proteasome-medi-tion factor, and SUG1. ated proteolysis (Xu et al., 1995). Furthermore, the sug1-1
We do not yet understand the precise mechanism by strain does not exhibit the 6-AU sensitivity characteristicwhich the 19S complex acts in transcription elongation. of the sug1-20 and sug1-25 strains (A.F. and S.A.J.,As mentioned above, a reasonable working model is unpublished observations), suggesting that it affects an-that the chaperonin activity of the 19S complex is impor- other step in transcription. All these data point to atant in remodeling protein-protein or protein-DNA com- potentially extensive and complex role for the 19S inplexes that are involved in pausing of RNAP II (Conaway transcription.et al., 2000). A precedent for this type of mechanismis provided by the role of bacterial ClpX in phage Mu Experimental Procedurestransposition. ClpX is thought to be functionally and
Yeast Strainsstructurally related to the six AAA ATPases in the eukary-The Saccharomyces cerevisiae strain W303a (MATa ade2-1 ura3-1otic 19S complex. Like Sug1, it plays a role in bacterialhis3-11, 15 trp1-1 leu2-3, 112 can1-100) was used as a source of
proteolysis (Glover and Lindquist, 1998; Hoskins et al., wild-type Sug1/Rpt6. Sc658 (sug1-20) and Sc660 (sug1-25) are con-2000; Weber-Ban et al., 1999), but it has also been shown genic to W303a and were generated as described previously (Russellto be capable of unfolding proteins and destabilizing et al., 1999b) The same procedure was followed to generate the
Sc677 (sug2-13) strain. Strain Sc804 (Dppr2::KAN) was obtainedDNA-protein interactions independent of the protease,from Research Genetics (Huntsville, AL). Strain yH129/14 (pre1-1ClpP (Kim et al., 2000; Singh et al., 2000). However, whilepre 4-1) is congenic to wild-type WCG4a (MATa ura3 leu2-3, 11219S-mediated remodeling of elongation intermediateshis3-11, 15 Cans Gal 1) (Heinemeyer et. al., 1993; Hilt et al., 1993)
is reasonable to propose, biochemical studies of the was provided by D. Wolf. Strain RJD1144 (MATa his3D200 leu2-19S are in their infancy. Other currently unrecognized 3,112 lys2-801 trpD63 PRE1 FLAG::YIplac211[URA3]) was gener-or poorly characterized activities of the complex may ously provided by Ray Deshaies of Cal Tech (Pasadena, CA) and
QX6861, expressing HA epitope-tagged Cdc68 was generously pro-also play a role in transcription elongation. For example,vided by G.C. Johnston of Dalhousie University (Halifax, Nova Sco-it has been reported that recombinant murine Sug1 istia). 6-azauricil sensitivity was assayed as described previously (Ex-a DNA helicase (Fraser et al., 1997), though this activityinger and Lacroute, 1992).
remains to be demonstrated in the context of the 19Scomplex.
RNA AnalysisFinally, it is important to point out that while the data Yeast strains in synthetic complete (SC) medium were diluted and
for an elongation effect are compelling, we cannot rule grown to an OD600 of 0.5–0.6 at 308C. Total RNA isolation, quantifica-tion, and Northern analysis for IMPDH expression were performedout effects of the 19S in other phases of the transcription

The 19S Complex in Transcription Elongation989
as described (Shaw and Reines, 2000), except that cells were treated beads and incubated for 60 min at 48C in MTB buffer (50 mM HEPES-KOH [pH 7.5], 100 mM potassium acetate, 5 mM magnesium acetate,with 100 mg/ml of 6-AU. In addition, the nylon membrane (Amersham,
Piscataway, NJ)) was rinsed in 53 SSC, prehybridized and hybrid- 1 mM EGTA, 0.1 mM DTT, 10% glycerol, and 0.1% NP-40). Thebeads were isolated and washed three times with a large excessized in PerfectHyb Plus (Sigma, St. Louis, MO) and washed twice
in 23 SSC-0.1% SDS and twice in 0.23 SSC-0.1% SDS for 15 min of MTB buffer. Finally the beads were resuspended in protein dyeand boiled, and the supernatant was analyzed by SDS-PAGE andeach at 658C. IMPDH expression was analyzed and quantitated by
phosphorimager as described previously (Russell et al., 1999a). Western blotting.
Preparation of Yeast Whole-Cell Extract In Vitro TranscriptionYeast cells were grown in YEPD medium at room temperature A kinetically synchronized transcription reaction was divided into a(z238C) to an OD600 to 1, harvested by centrifugation, and washed preincubation and an initiation/elongation step. Yeast WCE (250–once with cold distilled water. WCEs were prepared according to 300 mg) with or without heat treatment (HT) for 15 min at 378C wasthe protocol of Woontner and Jaehning (1990), except that the cell mixed with 500 ng of supercoiled nonspecific and template DNApellet was resuspended in 1.3 ml/g of 200 mM Tris base (pH 7.9), (pGAL4CG– [Woontner and Jaehning, 1990]) containing a single0.39 M (NH4)2SO4, 1 mM DTT, and 13 complete protease inhibitors Gal4 binding site upstream of the CYC1 core promoter fused to a(Boehringer, Indianapolis, IN). Protein concentration was deter- G-less cassette. The reaction mixture (15 ml) in transcription (TXR)mined by Bradford assay using bovine-serum albumin as a standard. buffer (20 mM HEPES-KOH [pH 7.5], 10% glycerol, 5 mM MgSO4,
5 mM EGTA, 2.5 mM DTT, and 0.2 mM PMSF) was preincubatedAffinity Purification of 19S and 26S Proteasome for 40 min at 238C either in the presence or absence of purifiedand Measurement of Peptidase Activity Gal4-VP16 protein and lactacystin. This was followed by the additionAffinity-purified 19S and 26S complexes were prepared from S. of 15 ml of TXR buffer containing 0.5 mCi of a-32P-UTP (3000 Ci/cerevisiae strain RJD1144 (MATa his3D200 leu2-3,112 lys2-801 mmol, Amersham), the A/C/UTP mixture (final concentration: 400trpD63 PRE1FLAG::YIplac211(URA3) as described previously (Verma mM of ATP and CTP, 16 mM of UTP) and 20 mM magnesium acetate,et al., 2000), but with the following modifications. First, the 19S was 300 mM creatine phosphate, 0.5 U creatine phosphokinase, and 20eluted for 3 hr at 48C by resuspension of the immunopurified 26S U RNasin (Sigma), and incubation was continued for another 15 min.on beads in an elution buffer (25 mM Tris [pH 7.5], 5 mM MgCl2, RNase T1 (50 U) (Boehringer) in EDTA was then added to terminate15% glycerol) devoid of ATP and supplemented with 500 mM NaCl. the transcription reaction. After an additional 10 min incubation, 100Second, eluted 19S and 26S protein complexes were concentrated ml of 10 mM Tris-HCl [pH 7.5], 300 mM NaCl, 1% SDS and 7 M ureaby centrifugation through a 100 kDa exclusion filter (Amicon, Bev- was added, and G-less transcripts (350–400-mer) were isolated anderly, MA) and extensively dialyzed against the yeast-extract buffer. analyzed by urea-polyacrylamide gel electrophoresis. RadiolabeledConcentrated samples were analyzed by silver-stained SDS-PAGE transcripts in dried gels were analyzed and quantitated as describedgels, or by Western blot analysis using various anti-proteasome previously (Russell et al., 1999a).subunit antibodies. Eluted 26S fractions were further analyzed by Add-back assays were performed essentially as above, exceptproteasome peptidase activity assays as described previously (Rus- that the affinity-purified 19S or 26S proteasome, with or withoutsell et al., 1999b). incubation for 10 min at 908C, was added with the heat-treated or
untreated WCEs during preincubation.Transcription reactions in which antibody inhibition was analyzedAntibodies and Western Blotting
were carried out as above with slight modifications. Equal amountRabbit anti-Sug1/Rpt6 antibodies were raised against either full-(4 mg) of anti-Sug1/Rpt6 antibodies raised against the full-lengthlength or the N-terminal 156 amino acids of Sug1/Rpt6, as describedSug1, or anti-Gal6 antibodies (as control), was added in yeast WCEpreviously (Russell et al., 1999a; Russell et al., 1999b). Production(240 mg) in TXR buffer containing 0.1 mM DTT and preincubated forand immunopurification of antibodies against Gal6 has been de-60 min at 238C before adding the template DNA and activator. Inscribed (Zheng and Johnston, 1998; Zheng et al., 1998). Preparationsome reactions anti-Sug1/Rpt6 antibodies were heat-denatured (10of anti-Sug2 antibodies has been described (Russell et al., 1996).min at 908C) or preincubated with purified 19S before adding them toAnti-20S proteasome antibodies were a gift from Keiji Tanaka. Anti-the yeast WCE, template DNA, and activator. Transcription reactionsbodies directed against Rpt protein were a gift from Jonathan C.with or without anti-Sug2 antibodies were carried out as above,Swaffield. Horseradish peroxidase-conjugated goat anti-rabbit orexcept that 160 mg of yeast WCE was used.anti-mouse IgG (Bio-Rad, Hercules, CA) were used for chemifluores-
Pulse-chase reactions were carried out, described as follows.cent detection of each protein. Western blotting was conducted asHeat-treated or untreated yeast WCE from indicated SUG1 strain,previously described (Russell et al., 1999b).activator (75 ng), and template were mixed and preincubated for 40min (unless otherwise indicated) at 238C to synchronize the forma-Protein Expression and Polyclonal Antibody Purificationtion of PIC. The PIC were incubated for 5 min in the presence ofPlasmids expressing either Gal4 (1–141 aa)-VP16 fused to glutathi-ATP and labeled UTP to initiate transcription and to label the G-lessone-S-transferase (GST) (Woontner and Jaehning., 1990) or His-transcripts (pulse). Labeling of G-less transcripts were then chasedtagged full-length Sug1/Rpt6 proteins (Russell et al., 1999b) wereby adding a high excess of cold UTP (final 1.0 mM) and incubationdescribed. GST fusion protein was expressed and purified from E.was continued for a different time period with indicated final concen-coli with glutathione-Sepharose according to the manufacturer’stration of CTP. Addition of anti-Sug1/Rpt6 antibodies and purifiedinstruction (Pharmacia, Piscataway, NJ). His-tagged full-length19S complex at specific time point of the transcription cycle isSug1/Rpt6 protein produced in inclusion bodies in E. coli was cou-indicated. Transcripts were purified and processed as described inpled to Ni beads. After extensive washing, rabbit serum directedExperimental Procedures.against full-length Sug1/Rpt6 (Russell et al., 1999b) was loaded on
the beads. Anti-Sug1/Rpt6 antibodies were eluted with 4 M MgCl2,dialyzed against PBS, and then against yeast-extract buffer without AcknowledgmentsDTT and concentrated before use. Sera from Sug2 immunized rab-bits was purified on a sepharose A column as described by the We thank Steven J. Russell for constructing the sug1-20 and sug1-manufacturer and concentrated. Concentrations were determined 25 strains and for producing antibodies against Sug1 and Sug2.as described above. Sincere thanks to Dieter Wolf, Ray J. Deshaies, and Gerald C. John-
ston for strains, and Keiji Tanaka (anti-20S), Tim Formosa (anti-Cdc68), and Jonathan C. Swaffield (anti-Rtp5) for antibodies. WeImmunoprecipitation
Anti-HA antibodies (Boehringer, Indianapolis, IN) and anti-T7 anti- also thank Prof. Deshaies for providing us with a protocol, prior topublication, for immunoaffinity purification of epitope-tagged pro-bodies (Novagen, Madison, WI) were immobilized on Dynabeads
according to the manufacturer’s instruction and washed to remove teasome. Modification of this protocol resulted in the method em-ployed to prepare the 26S and 19S complexes used in this study.unbound protein. Yeast WCE (200 mg), prepared from a yeast strain
expressing HA epitope-tagged Cdc68, was mixed with 100 mg of We also thank Mark Combs for mass spectrometric analysis of the

Molecular Cell990
19S and 26S proteasome, Xiang Chen, Eunice Webb, and Zakir SCFMet30-mediated inactivation of the transcription factor Met4. Cell102, 303–314.Siddiquee for their technical expertise, and members of S.A.J.’s and
T.K.’s laboratories for helpful comments on the manuscript. This Kim, Y.-I., Burton, R.E., Burton, B.M., Sauer, R.T., and Baker, T.A.work was supported by a grant from the American Cancer Society (2000). Dynamics of substrate denaturation and translocation by theto T.K. and unrestricted funds to S.A.J. ClpXP degradation machine. Mol. Cell. 5, 639–648.
LeRoy, G., Orphanides, G., Lane, W.S., and Reinberg, D. (1998).Received September 18, 2000; revised March 16, 2001. Requirement of RSF and FACT for transcription of chromatin tem-
plates in vitro. Science 282, 1900–1904.References Malone, E.A., Clarke, C.D., Chiang, A., and Winston, F. (1991). Muta-
tions in SPT16/CDC68 suppress cis- and trans-acting mutationsBaumeister, W., Walz, J., Zuhl, F., and Seemuller, E. (1998). The that affect promoter function in Saccharomyces cerevisiae. Mol.proteasome: paradigm of a self-compartmentalizing protease. Cell Cell. Biol. 11, 5710–5717.92, 367–380. Marshall, N.F., Peng, J., Xie, Z., and Price, D.H. (1996). Control ofBraun, B.C., Glickman, M., Kraft, R., Dahlmann, B., Kloetzel, P.-M., RNA polymerase II elogation potential by a novel carboxyl-terminalFinley, D., and Schmidt, M. (1999). The base of the proteasome domain kinase. J. Biol. Chem. 271, 27176–27183.regulatory particle exhibits chaperone-like activity. Nat. Cell Biol. 1, Masuyama, H., and MacDonald, P.N. (1998). Proteasome-mediated221–226. degradation of the vitamin D receptor (VDR) and a putative role forBuratowski, S. (1994). The basics of basal transcription by RNA SUG1 interaction with the AF-2 domain of VDR. J. Cell. Biochem.polymerase II. Cell 77, 1–3. 71, 429–440.
Conaway, J.W., Shilatifard, A., Dvir, A., and Conaway, R.C. (2000). Molinari, E., Gilman, M., and Natesan, S. (1999). Proteasome-medi-Control of elongation by RNA polymerase II. Trends Biochem. Sci. ated degradation of transcriptional activators correlates with activa-25, 375–380. tion domain potency in vivo. EMBO J. 18, 6439–6447.
Conaway, R.C., and Conaway, J.W. (1993). General initiation factors Mitsui, A., and Sharp, P.A. (1999). Ubiquitination of RNA polymerasefor RNA polymerase II. Annu. Rev. Biochem. 62, 161–190. II large subunit signaled by phosphorylation of carboxyl-terminal
domain. Proc. Natl. Acad. Sci. USA 96, 6054–6059.Confalonieri, F., and Duguet, M. (1995). A 200 amino acid ATPasemodule in search of a basic function. Bioessays 17, 639–650. Nishiyama, A., Tachibana, K., Igarashi, Y., Yasuda, H., Tanahashi, N.,
Tanaka, K., Ohsumi, K., and Kishimoto, T.A. (2000). NonproteolyticCoux, O., Tanaka, K., and Goldberg, A.L. (1996). Structure and func-function of the proteasome is required for the dissociation of Cdc2tion of the 20S and 26S proteasomes. Ann. Rev. Biochem. 65,and cyclin B at the end of M phase. Genes Dev. 14, 2344–2357.801–847.Orphanides, G., Wu, W.-H., Lane, W.S., Hampsey, M., and Reinberg,Exinger, F., and Lacroute, F. (1992). 6-Azauricil inhibition of GTPD. (1999). The chromatin-specific transcription elongation factorbiosynthesis in Saccharomyces cerevisiae. Curr. Genet. 22, 9–11.FACT comprises human SPT16 and SSRP1 proteins. Nature 400,
Fenteany, G., Standaert, R.F., Lane, W.S., Choi, S., Corey, E.J.,284–288.
and Schreiber, S.L. (1995). Inhibition of proteasome activities andPadilla, P.A., Fuge, E.K., Crawford, M.E., Errett, A., and Werner-subunit-specific amino-terminal threonine modification by lactacys-Washburne, M. (1998). The highly conserved, coregulated SNO andtin. Science 268, 726–731.SNZ gene families in Saccharomyces cerevisiae respond to nutrient
Fraser, R.A., Rossingnol, M., Heard, D.J., Egly, J.-M., and Chambon,limitation. J. Bacteriol. 180, 5718–5726.
P. (1997). SUG1, a putative transcriptional mediator and subunit ofPowell, W., and Reines, D. (1996). Mutations in the second largestthe PA700 proteasome regulatory complex, is a DNA helicase. J.subunit of RNA polymerase II cause 6-azauricil sensitivity in yeastBiol. Chem. 272, 7122–7126.and increased transcriptional arrest in vitro. J. Biol. Chem. 271,
Glickman, M.H., Rubin, D.M., Fried, V.A., and Finley, D. (1998). The6866–6873.
regulatory particle of the Saccharomyces cerevisiae proteasome.Reines, D., Conaway, J.W., and Conaway, R.C. (1996). The RNAMol. Cell. Biol. 18, 3149–3162.polymerase II general elongation factors. Trends Biochem. Sci. 21,
Glover, J.R., and Lindquist, S. (1998). Hsp104, Hsp70 and Hsp40: a351–355.
novel chaperone system that rescues previously aggregated pro-Roeder, R.G. (1996). The role of general initiation factors in transcrip-teins. Cell 94, 73–82.tion by RNA polymerase II. Trends Biochem. Sci. 9, 327–335.
Hartzog, G.A., Basral, M.A., Ricupero-Hovasse, S.L., Hieter, P., andRowley, A., Singer, R.A., and Johnston, G.C. (1991). CDC68, a yeastWinston, F. (1996). Identification and analysis of a functional humangene that affects regulation of cell proliferation and transcription,homolog of the SPT4 gene of Saccharomyces cerevisiae. Mol. Cell.encodes a protein with a highly acidic carboxyl terminus. Mol. Cell.Biol. 16, 2848–2856.Biol. 11, 5718–5726.
Hartzog, G.A., Wada, T., Handa, H., and Winston, F. (1998). EvidenceRubin, D., Coux, O., Wefes, I., Hengartner, C., Young, R.A., Goldberg,that Spt4, Spt5 and Spt6 control transcription elongation by RNAA.L., and Finley, D. (1996). Identification of the gal4 suppressor Sug1polymerase II in Saccharomyces cerevisiae. Genes Dev. 12,as a subunit of the yeast 26S proteasome. Nature 379, 655–657.357–369.Rubin, D.M., Glickman, M.H., Larsen, C.N., Dhruvakumar, S., andHeinemeyer, W., Gruhler, A., Mohrle, V., Mahe, Y., and Wolf, D.Finley, D. (1998). Active site mutants in the six regulatory particle(1993). PRE2, highly homologous to the human major histocompata-ATPases reveal multiple roles for ATP in the proteasome. EMBO J.bility complex-linked RING10 gene, codes for a yeast proteasome17, 4086–4091.subunit necessary for chymotryptic activity and degradation of ubi-
quitinated proteins. J. Biol. Chem. 268, 5115–5120. Russell, S.J., and Johnston, S.A. (2001). Evidence that proteolysisof Gal4 can not explain the transcriptional effects of proteasomeHilt, W., Enekel, C., Gruhler, A., Singer, T., and Wolf, D.H. (1993).ATPase mutations. J. Biol. Chem., 276, 9825–9831.The PRE4 gene codes for a subunit of the yeast proteasome neces-
sary for peptidylglutamyl-peptide-hydrolyzing activity. J. Biol. Russell, S.J., Reed, S.H., Huang, W., Friedberg, E.C., and Johnston,Chem. 268, 3479–3486. S.A. (1999a). The 19S regulatory complex of the proteasome func-
tions independently of proteolysis in nucleotide excision repair. Mol.Hoskins, J.R., Singh, S.K., Maurizi, M.R., and Wickner, S. (2000).Cell 5, 687–696.Protein binding and unfolding by the chaperone ClpA and degrada-
tion by the protease ClpAP. Proc. Natl. Acad. Sci. USA 97, 8892– Russell, S.J., Sathyanarayana, U.G., and Johnston, S.A. (1996). Isola-8897. tion and characterization of SUG2. A novel ATPase family compo-
nent of the yeast 26S proteasome. J. Biol. Chem. 271, 32810–32817.Kaiser, P., Flick, K., Wittenberg, C., and Reed, S.I. (2000). Regulationof transcription by ubiquitination without proteolysis: Cdc34/ Russell, S.J., Steger, K.A., and Johnston, S.A. (1999b). Subcellular

The 19S Complex in Transcription Elongation991
localization, stoichiometry, and protein levels of 26S proteasome containing RD, cooperates with DSIF to repress RNA polymerase IIelongation. Cell 97, 41–51.subunits in yeast. J. Biol. Chem. 274, 21943–21952.
Zheng, W., and Johnston, S.A. (1998). The nucleic acid bindingSalghetti, S.E., Muratani, M., Wijnen, H., Futcher, B., and Tansey,activity of bleomycin hydrolase is involved in bleomycin detoxifica-W.P. (2000). Functional overlap of sequences that activate transcrip-tion. Mol. Cell. Biol. 18, 13776–13780.tion and signal ubiquitin-mediated proteolysis. Proc. Natl. Acad. Sci.
USA 97, 3118–3123. Zheng, W., Johnston, S.A., and Joshua-Tor, L. (1998). The unusualactive site of Gal6/bleomycin hydrolase can act as a carboxypepti-Schauber, C., Chen, L., Tongaonkar, P., Vega, I., Lambertson, D.,dase, aminopeptidase, and peptide ligase. Cell 93, 103–109.Potts, W., and Madura, K. (1998). Rad23 links DNA repair to the
ubiquitin/proteasome pathway. Nature 391, 715–718.
Shaw, R.J., and Reines, D. (2000). Saccharomyces cerevisiae tran-scription elongation mutants are defective in PUR5 induction inresponse to nucleotide depletion. Mol. Cell. Biol. 20, 7427–7437.
Singh, S.K., Grimaud, R., Hoskins, J.R., Wickner, S., and Maurizi,M.R. (2000). Unfolding and internalization of proteins by the ATP-dependent proteases ClpXP and ClpAP. Proc. Natl. Acad. Sci. USA97, 8898–8903.
Strickland, E., Hakala, K., Thomas, P.J., and DeMartino, G.N. (2000).Recognition of misfolding proteins by PA700, the regulatory sub-complex of the 26S proteasome. J. Biol. Chem. 275, 5565–5572.
Swaffield, J.C., Bromberg, J., and Johnston, S.A. (1992). Alterationsin a yeast protein resembling HIV Tat-binding protein relieve therequirement for an acidic activation domain of GAL4. Nature 357,698–700.
Swaffield, J.C., Melcher, K., and Johnston, S.A. (1996). Highly con-served ATPase protein as mediator between acidic activation do-mains and the TATA-binding protein. Nature 379, 658.
Verma, R., Chen, S., Feldman, R., Schieltz, D., Yates, J., Dohmen,J., and Deshaies, R.J. (2000). Proteasomal proteomics: identificationof nucleotide-sensitive proteasome-interacting proteins by massspectrometric analysis of affinity-purified proteasomes. Mol. Biol.Cell 11, 3425–3439.
Wada, T., Orphanides, G., Hasegawa, J., Kim, D.-K., Shima, D.,Yamaguchi, Y., Fukuda, A., Hisatake, K., Oh, S., Reinberg, D., andHanda, H. (2000). FACT relieves DSIF/NELF-mediated inhibition oftranscriptional elongation and reveals functional differences be-tween P-TEFb and TFIIH. Mol. Cell 5, 1067–1072.
Wada, T., Takagi, T., Yamaguchi, Y., Ferdous, A., Imai, T., Hirose,S., Sugimoto, S., Yano, K., Hartzog, G.A., Winston, F., et al. (1998).DSIF, a novel transcription elongation factor that regulates RNApolymerase II processivity, is composed of human Spt4 and Spt5homologs. Genes Dev. 12, 343–356.
Wang, D., Moriggl, R., Stravopodis, D., Carpino, N., Marine, J.-C.,Teglund, S., Feng, J., and Ihle, J.N. (2000). A small amphipathica-helical region is required for transcriptional activities and protea-some-dependent turnover of the tyrosine-phosphorylated Stat5.EMBO J. 19, 392–399.
Weber-Ban, E.U., Reid, B.G., Miranker, A.D., and Horwich, A.L.(1999). Global unfolding of a substrate protein by the Hsp100 chap-erone ClpA. Nature 401, 90–93.
Weeda, G., Rossignol, M., Fraser, R.A., Winkler, G.S., Vermeulen,W. van’t Veer, L.J., Ma, L., Hoeijmakers, J.H.J., and Egly, J.-M.(1997). The XPB subunit of repair/transcription factor TFIIH directlyinteracts with SUG1, a subunit of the 26S proteasome and putativetranscription factor. Nucleic Acids Res. 25, 2274–2283.
Wittmeyer, J., and Formosa, T. (1997). The Saccharomyces cerevis-iae DNA polymerase a catalytic subunit interacts with Cdc68/Spt16and with Pob3, a protein similar to an HMG-1-like protein. Mol. Cell.Biol. 17, 4178–4190.
Wittmeyer, J., Joss, L., and Formosa, T. (1999). Spt16 and Pob3 ofSaccharomyces cerevisiae form an essential, abundant heterodimerthat is nuclear, chromatin-associated, and copurifies with DNA poly-merase a. Biochemistry 38, 8961–8971.
Woontner, M., and Jaehning, J.A. (1990). Accurate initiation by RNApolymerase II in a whole cell extract from Saccharomyces cerevisiae.J. Biol. Chem. 265, 8979–8982.
Xu, Q., Singer, R.A., and Johnston, G.C. (1995). Sug1 modulatesyeast transcription by Cdc68. Mol. Cell. Biol. 15, 6025–6035.
Yamaguchi, Y., Takagi, T., Wada, T., Furuya, A., Sugimoto, S., Ha-segawa, J., and Handa, H. (1999). NELF, a multisubunit complex
Related Documents











