Temporal Dynamics of Parietal Activity During Word-Location Binding Pablo Campo, Fernando Maestu ´, and Almudena Capilla Universidad Complutense de Madrid Manuel Morales Universidad de Sevilla Santiago Ferna ´ndez, David del Rı ´o, and Toma ´s Ortiz Universidad Complutense de Madrid Most of human cognitive activity involves, to a greater or lesser extent, the integration of information from different modalities, a process also referred to as ‘binding’. Although the neural basis of several forms of binding has been extensively investigated, the neurobiological mechanisms of the encoding phase of integration of words and their spatial location have not been previously investigated. This process is at the core of what Baddeley proposed in his revised model as episodic buffer. In the current experiment, the authors used magnetoencephalography to investigate the spatiotemporal patterns of brain activity related to encoding words, locations, and the integration of both types of information using a working memory paradigm. The spatiotemporal analysis showed a preferential activation of superior parietal lobe (SPL) during the integration of information, which was modulated by performance in the task. These findings are in agreement with proposals suggesting that SPL participates in binding processes by encoding and maintaining a detailed, complex integrated representation in working memory (WM). Considering Baddeley’s episodic buffer, it appears that the same mechanisms involved in integrating information within one subsystem (i.e., visuospatial buffer) also support the integration of information between previously considered independent subsystems (i.e., verbal and visuospatial buff- ers). Keywords: working memory, encoding, binding, episodic buffer, magnetoencephalography WM is considered a supporting process for high-level cognitive abilities and refers to a dynamic cognitive system that permits the comprehension and mental representation of the immediate envi- ronment (Miyake & Shah, 1999). It is widely assumed that WM can be divided into two separate components for processing verbal and visuospatial information (Baddeley, 1996; Baddeley & Hitch, 1974; Logie, 1995; Miyake & Shah, 1999). Processing of infor- mation within WM subsystems is restricted by duration and ca- pacity. Despite this limited capacity, which appears to be about four items (Cowan, 2001), people are able to process larger amounts of information. This ability involves “the reorganization of bits of information to create more complex but unified repre- sentations of previously distributed information” (Wheeler & Tre- isman, 2002, p. 60). In the context of verbal material, the capability of processing additional information beyond the highly restricted capacity of WM is explained by the organization of independent pieces of information into meaningful items, also referred to as chunking (Baddeley, 2000; Ericsson, Chase, & Faloon, 1980; Miller, 1956; Simon, 1974). In the visuospatial domain, several models have been proposed to explain the process of combining stimulus features to form a more complex representation, also refereed to as binding (Gray, 1999). The feature-based model considers that WM capacity is limited by the number of features (Wheeler & Treisman, 2002; Wolfe et al., 1990). The object-based model suggests that the capacity of visuospatial WM is limited by the number of objects that can be represented (Duncan, 1984; Luck & Vogel, 1997; O’Craven, Downing, & Kanwisher, 1999). An alternative model postulates that the number of locations occupied by visual stimuli can be a capacity constraint, referred to as the space model (Eriksen & Yeh, 1985). Other authors (Alvarez & Cavanagh, 2004; Eng, Chen, & Jiang, 2005; Olson & Jiang, 2002) consider that stimulus complexity can limit the capacity of WM. Finally, in order to account for the evidence of integrated process- ing of information, the last update of Baddeley’s WM model included a new component, the episodic buffer, “that is capable of integrating information from a variety of sources” (p. 421, Bad- deley, 2000), although the mechanisms have not been well ex- plained. The fractionation of WM into verbal and visuospatial sub- systems has been supported by neuropsychological and neuroim- aging studies providing evidence about the functional dissociation of these components in the brain (Becker, MacAndrew, & Fiez, Pablo Campo and David del Rı ´o, Centro de Magnetoencefalografı ´a Dr. Pe ´rez-Modrego, Universidad Complutense de Madrid, Madrid, Spain; Fer- nando Maestu ´, Almudena Capilla, and Santiago Ferna ´ndez, Centro de Magnetoencefalografı ´a Dr. Pe ´rez-Modrego, Universidad Complutense de Madrid, Madrid, Spain, and Departamento de Psicologı ´a Ba ´sica II (Proce- sos Cognitivos) Facultad de Psicologı ´a, Universidad Complutense de Ma- drid, Madrid, Spain; Manuel Morales, Departmento de Psicologı ´a Exper- imental, Universidad de Sevilla, Seville, Spain; Toma ´s Ortiz, Centro de Magnetoencefalografı ´a Dr. Pe ´rez-Modrego, Universidad Complutense de Madrid, Madrid, Spain and Departamento de Psiquiatrı ´a, Facultad de Medicina, Universidad Complutense de Madrid, Madrid, Spain. This research was supported by the Ministerio de Educacion y Ciencia Grant SEJ2006-14571. We also acknowledge B. A. Strange for helpful comments on an earlier version of the manuscript. Correspondence concerning this article should be addressed to Pablo Campo, Centro de Magnetoencefalografı ´a Dr. Pe ´rez-Modrego, Pabello ´n 8, Facultad de Medicina, Universidad Complutense de Madrid, 28040, Ma- drid, Spain. E-mail: [email protected] Neuropsychology Copyright 2008 by the American Psychological Association 2008, Vol. 22, No. 1, 85–99 0894-4105/08/$12.00 DOI: 10.1037/0894-4105.22.1.85 85

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Temporal Dynamics of Parietal Activity During Word-Location Binding
Pablo Campo, Fernando Maestu, andAlmudena Capilla
Universidad Complutense de Madrid
Manuel MoralesUniversidad de Sevilla
Santiago Fernandez, David del Rıo, and Tomas OrtizUniversidad Complutense de Madrid
Most of human cognitive activity involves, to a greater or lesser extent, the integration of informationfrom different modalities, a process also referred to as ‘binding’. Although the neural basis of severalforms of binding has been extensively investigated, the neurobiological mechanisms of the encodingphase of integration of words and their spatial location have not been previously investigated. Thisprocess is at the core of what Baddeley proposed in his revised model as episodic buffer. In the currentexperiment, the authors used magnetoencephalography to investigate the spatiotemporal patterns of brainactivity related to encoding words, locations, and the integration of both types of information using aworking memory paradigm. The spatiotemporal analysis showed a preferential activation of superiorparietal lobe (SPL) during the integration of information, which was modulated by performance in thetask. These findings are in agreement with proposals suggesting that SPL participates in bindingprocesses by encoding and maintaining a detailed, complex integrated representation in working memory(WM). Considering Baddeley’s episodic buffer, it appears that the same mechanisms involved inintegrating information within one subsystem (i.e., visuospatial buffer) also support the integration ofinformation between previously considered independent subsystems (i.e., verbal and visuospatial buff-ers).
Keywords: working memory, encoding, binding, episodic buffer, magnetoencephalography
WM is considered a supporting process for high-level cognitiveabilities and refers to a dynamic cognitive system that permits thecomprehension and mental representation of the immediate envi-ronment (Miyake & Shah, 1999). It is widely assumed that WMcan be divided into two separate components for processing verbaland visuospatial information (Baddeley, 1996; Baddeley & Hitch,1974; Logie, 1995; Miyake & Shah, 1999). Processing of infor-mation within WM subsystems is restricted by duration and ca-pacity. Despite this limited capacity, which appears to be aboutfour items (Cowan, 2001), people are able to process largeramounts of information. This ability involves “the reorganizationof bits of information to create more complex but unified repre-
sentations of previously distributed information” (Wheeler & Tre-isman, 2002, p. 60). In the context of verbal material, the capabilityof processing additional information beyond the highly restrictedcapacity of WM is explained by the organization of independentpieces of information into meaningful items, also referred to aschunking (Baddeley, 2000; Ericsson, Chase, & Faloon, 1980;Miller, 1956; Simon, 1974). In the visuospatial domain, severalmodels have been proposed to explain the process of combiningstimulus features to form a more complex representation, alsorefereed to as binding (Gray, 1999). The feature-based modelconsiders that WM capacity is limited by the number of features(Wheeler & Treisman, 2002; Wolfe et al., 1990). The object-basedmodel suggests that the capacity of visuospatial WM is limited bythe number of objects that can be represented (Duncan, 1984; Luck& Vogel, 1997; O’Craven, Downing, & Kanwisher, 1999). Analternative model postulates that the number of locations occupiedby visual stimuli can be a capacity constraint, referred to as thespace model (Eriksen & Yeh, 1985). Other authors (Alvarez &Cavanagh, 2004; Eng, Chen, & Jiang, 2005; Olson & Jiang, 2002)consider that stimulus complexity can limit the capacity of WM.Finally, in order to account for the evidence of integrated process-ing of information, the last update of Baddeley’s WM modelincluded a new component, the episodic buffer, “that is capable ofintegrating information from a variety of sources” (p. 421, Bad-deley, 2000), although the mechanisms have not been well ex-plained.
The fractionation of WM into verbal and visuospatial sub-systems has been supported by neuropsychological and neuroim-aging studies providing evidence about the functional dissociationof these components in the brain (Becker, MacAndrew, & Fiez,
Pablo Campo and David del Rıo, Centro de Magnetoencefalografıa Dr.Perez-Modrego, Universidad Complutense de Madrid, Madrid, Spain; Fer-nando Maestu, Almudena Capilla, and Santiago Fernandez, Centro deMagnetoencefalografıa Dr. Perez-Modrego, Universidad Complutense deMadrid, Madrid, Spain, and Departamento de Psicologıa Basica II (Proce-sos Cognitivos) Facultad de Psicologıa, Universidad Complutense de Ma-drid, Madrid, Spain; Manuel Morales, Departmento de Psicologıa Exper-imental, Universidad de Sevilla, Seville, Spain; Tomas Ortiz, Centro deMagnetoencefalografıa Dr. Perez-Modrego, Universidad Complutense deMadrid, Madrid, Spain and Departamento de Psiquiatrıa, Facultad deMedicina, Universidad Complutense de Madrid, Madrid, Spain.
This research was supported by the Ministerio de Educacion y CienciaGrant SEJ2006-14571. We also acknowledge B. A. Strange for helpfulcomments on an earlier version of the manuscript.
Correspondence concerning this article should be addressed to PabloCampo, Centro de Magnetoencefalografıa Dr. Perez-Modrego, Pabellon 8,Facultad de Medicina, Universidad Complutense de Madrid, 28040, Ma-drid, Spain. E-mail: [email protected]
Neuropsychology Copyright 2008 by the American Psychological Association2008, Vol. 22, No. 1, 85–99 0894-4105/08/$12.00 DOI: 10.1037/0894-4105.22.1.85
85

1999; D’Esposito, Postle, Ballard, & Lease, 1999; Haxby, Petit,Ungerleider, & Courtney, 2000; Smith, Jonides, Marshuetz, &Koeppe, 1998; Zurowski et al., 2002). Concerning the neural basisof information integration process, a great deal of evidence fromneuropsychological, psychophysical, functional neuroimaging,and transcranial magnetic stimulation studies strongly support thecrucial engagement of posterior parietal cortex in integration pro-cesses (Corbetta, Shulman, Miezin, & Petersen, 1995; Donner etal., 2002; Lee & Chun, 2001; Shafritz, Gore, & Marois, 2002;Simon-Thomas, Brodsky, Willing, Sinha, & Knight, 2003; Todd &Marois, 2004, 2005; Vogel & Machizawa, 2004; Xu & Chun,2006). Considered together, the findings of these studies suggest,as recently stated by Marois and Ivanoff (2005), that because of itsrelevant role in visuospatial attention and working memory, “theposterior parietal cortex is well positioned to build and integratemental representation of the visual scene” (p. 299).
Despite the increasing interest in recent years in studying theneural basis of integration of information in the visual domain,surprisingly the neural mechanisms of integration of verbal andspatial information have received less attention. However, in real-world scenes, we daily face this kind of integrated information inthe form of traffic signals, logotypes, and so forth. A potentialcandidate could be the episodic buffer, a component defined as “aninterface between a range of systems, each involving a different setof codes” (Baddeley, 2000, p. 421). In fact, the inclusion of theepisodic buffer was mainly due to the need to explain bindingbetween visual and verbal subsystems of WM and between long-term language knowledge and WM (Allen et al., 2006). To the bestof our knowledge, this issue has been considered in only twoprevious experiments (Campo et al., 2005; Prabhakaran, Naray-anan, Zhao, & Gabrieli, 2000). In one of them (Prabhakaran et al.,2000), participants were asked to maintain spatial (locations) andverbal (letters) information in an integrated or an unintegratedmanner. The authors observed a greater activation of prefrontalcortex during the maintenance of integrated representations. In theother study (Campo et al., 2005), the authors found a greateractivation of the so-called superior or dorsal network, includingfrontal and parietal regions, when participants maintained inte-grated representations of verbal (words) and spatial (locations)information. This enhancement of activation was observed mainlyduring the first 600 ms of the maintenance period.
With the exception of Campo et al.’s (2005) study, no furtherattempts have been made to study the neural patterns related toword-location integration during WM. Although these authorshave previously investigated the maintenance of integrated verbal–spatial information (Campo et al., 2005), the main purpose of thecurrent experiment was to investigate how the perceptual repre-sentations of the integrated information are identified and trans-ferred into stable WM representations (Vogel, Woodman, & Luck,2001) or how binding is encoded during WM (Treisman, 1999).We were interested in investigating where and when this processoccurs. Accurate information about the timing and duration ofbrain activity constitutes a unique contribution to the understand-ing of how activated structures are involved in cognitive processes.Magnetoencephalography (MEG) can provide real-time measuresof brain activity, thus allowing detailed tracking of the time-courseof this activity, along with a good spatial resolution. Accordingly,
we used MEG to compare the spatiotemporal patterns related toencoding words, locations, or integrated words and locations. Acomparison between integrated and single tasks could then indi-cate the brain structures specifically engaged during the functionalintegration of information from different modalities (Saito et al.,2005; Shafritz et al., 2002).
We hypothesized that the brain structures engaged during themaintenance of integrated mental representations (Campo et al.,2005) also will be recruited during the encoding period (Awh &Jonides, 2001; Awh, Vogel, & Oh, 2006; Lepsien & Nobre, 2006;Ruchkin, Grafman, Cameron, & Berndt, 2003; Xu & Chun, 2005)and that this process will occur in an advanced stage of the flow ofinformation processing (Wolfe & Cave, 1999).
Method
Participants
The sample consisted of 12 Spanish-speaking healthy volunteers(6 men and 6 women). Participants were right-handed, without anyhistory of neurological or psychiatric illness, ranging in agefrom 23 to 38 years (M � 28.58; SD � 3.75). All participants wereasked to sign a consent form, in accordance with the Declarationof Helsinki, after the nature of the procedures involved had beenexplained to them.
Procedure
MEG scans were obtained during three experimental tasks:verbal, spatial, and integrated tasks. A total of 120 trials werepresented in each task. For counterbalancing purposes the order ofpresentation of the tasks was controlled. Three different orderswere established. Experimental conditions were completed in dif-ferent sessions lasting 25 min approximately, with resting intervalsbetween them of 10 to 15 min.
According to neuropsychological studies (Robertson, 2003) andfunctional neuroimaging experiments (Shafritz et al., 2002), theintegration-related activation should primarily occur when thereare multiple objects simultaneously present in the visual display.Thus, stimuli were presented simultaneously in all experimentalconditions in the current experiment.
The stimuli were projected through an LCD video-projector(SONY VPL-X600E), situated outside the shielded room onto aseries of in-room mirrors, the last of which was suspended approx-imately 50 cm above the participant’s face and subtended a visualangle of 1° to 3° horizontally and 0.5° vertically.
Stimuli and Tasks
In the spatial task, the stimulus set consisted of four ellipses.The ellipses were located randomly in the display and were firstshown for 3 s. Participants were asked to remember the locationsof the ellipses (encoding phase). This phase was followed by aretention interval of 2,500 ms (maintenance phase). After thisdelay, participants were presented with three consecutive probedisplays, with a duration of 500 ms, consisting of one ellipse each.
86 CAMPO ET AL.

Participants had to decide whether the ellipsis was in the samelocation as one of the ellipses of the stimulus display (recognitionphase). There was an interval between probe displays of 500 to 750ms (Figure 1).
In the verbal task, stimulus set consisted of four words. Concretewords were used (5.8 � 2.23), four to seven letters in length(5.62 � 1.57) and of moderate frequency (35.62 � 4.67; Algara-bel, 1996). At the start of each trial, participants first saw astimulus array comprising four words, located centrally in thedisplay, which was projected for a duration of 3,000 ms (encodingphase). Participants were informed that, later on, three consecutiveprobe displays comprising a semantic category name would bepresented and that they would be asked to decide whether one ofthe words in the stimulus display belonged to the semantic cate-gory represented by one of the words in the probe displays. Eachprobe was presented during a 500-ms period (recognition phase).There was an interval between probe displays of 500 to 750 ms. Adelay interval of 2,500 ms during which participants viewed ablack screen (maintenance phase) was presented between the off-set of the stimulus display and the onset of the probe displays(Figure 2).
In the integrated task the stimulus set comprised four words andfour ellipses. Words were matched in concreteness (5.5 � 2.76),length (5.34 � 1.77), and frequency (32.62 � 5.18) with those ofthe verbal task. Words were placed within the ellipses, which werelocated randomly as in the spatial task. At the start of each trial,participants first saw a stimulus array consisting of four wordswithin four ellipses that was presented for 3 s and were asked toremember the words and the locations of the ellipses. As in thepreviously described tasks, participants had to maintain both typesof information for 2,500 ms. Subsequently, they were presented
with three consecutive probe displays consisting of a word, whichwas a semantic category, within a ellipse. Each probe was pre-sented during a 500-ms period, and participants had to decidewhether one of the words in the stimulus display belonged to thesemantic category represented by the word in the probe displayand whether the ellipsis was in the same location as one of theellipses of the stimulus display (Figure 3). Participants were toanswer affirmatively only when both the word and its locationwere presented in the stimulus display. There was an intervalbetween probe displays of 500 to 750 ms.
Data Collection and Analysis
All MEG recordings were done with a whole-head neuromag-netometer containing an array of 148 magnetometers (4-D 2500®,4-D Neuroimaging, San Diego, CA) housed in a magneticallyshielded room. Because the purpose of the present study was toinvestigate the brain activity during the encoding process of bind-ing, we focused our analyses on this phase. Consequently, pro-cesses of maintenance and recognition are not reported in thisarticle.
The magnetic flux measurements were filtered with a band-passfilter between 0.1 and 50 Hz and digitized at 254 Hz. MEG datawere submitted to an interactive noise reduction procedure thataided in reducing environmental noise and is part of the signalanalysis package. Single-trial event-related fields (ERFs) in re-sponse to 120 stimulus presentations were averaged separately foreach sensor after excluding those containing eye movement (asindicated by peak-to-peak amplitudes in the electro-oculogramchannel in excess of 50 �V) or other myogenic or mechanical
Figure 1. Shown are the schematic diagrams and time line of the spatial task trial type. Each box representsa stimulus display event. The arrows represent the duration of each of these events. ISI � insterstimulus interval.
87VERBAL–SPATIAL BINDING

artifacts. Finally, the averaged epochs (3,000 ms duration) weredigitally filtered with a low-pass 20-Hz filter to improve signalquality and adjusted relative to the mean amplitude in the 150-msprestimulus baseline. A minimum of 80 ERF epochs was collectedto calculate each averaged waveform.
The sources of magnetic fields were modeled as equivalentcurrent dipoles (ECDs). The location, orientation, and amplitude ofthe best fitting single ECD were estimated with a spherical volumeconductor model (Sarvas, 1987). Source estimation was attemptedonly when the surface distribution of magnetic flux was dipolar, inthat it consisted of a single region of magnetic outflux and a singleregion of magnetic influx. For a given point in time, the ECDfitting algorithm (Levenberg–Marquardt; More, 1977) was appliedto the magnetic flux measurements obtained from a group of34–38 magnetometers, always including both magnetic flux ex-tremes. An ECD was calculated at successive 4-ms intervals. TheECD solutions were considered satisfactory after meeting thefollowing criteria: (a) correlation coefficient of at least 0.9 betweenthe observed and the “best” predicted magnetic field distribution,(b) a goodness of fit of at least 0.9 or higher, and (c) a confidencevolume � 1 cm3. Occasionally, two distinct dipolar distributionswere discerned at a single time point (in one or both hemispheres),and source estimation was performed for both distributions inde-pendently. In order to avoid localization errors produced by smear-ing of the magnetic flux from one source by the flux induced by theother source, two simultaneous source solutions were retained onlyif the corresponding dipoles were found to be at least 5 cm apart.We elected this method because its concurrent validity has been
successfully tested in comparison with the results of invasivebrain-mapping techniques in a large patient series (Papanicolaou etal., 2004).
Brain response sources represented as ECDs were superimposedon the individual T1-weighted (magnetic resonance imaging[MRI]) images (repetition time [TR] � 13.6 ms; echo time [TE]� 4.8 ms; recording matrix � 256 � 256 pixels, 1 excitation, 240mm field of view, and 1.4 mm slice thickness) to delineate theanatomical location. Anatomical landmarks (nasion, common ver-tex [cz], inion, and the two auricular points corresponding to theexternal meati) were localized in each individual and used for theco-registration of the magnetic resonance imaging (MRI) andMEG scans. MRI scans were obtained for 9 of the participants. Forthe other participants, their activity sources were overlaid ontoMRIs that were matched to each participant according to thecriteria outlined by Papanicolaou et al. (2002).
The degree of activation of a particular area, or the total durationof its activation, was estimated by the total number of successivedipoles that accounted for the ERF components. There is nowconsiderable data supporting the validity of the procedure forconstructing brain activation used in the present study, which issufficiently accurate for lateralizing neurophysiological activityassociated with language function (Castillo et al., 2004; Maestu etal., 2002; Papanicolaou et al., 1999), and for identifying specificregions that are crucial for particular processes such as decodingand encoding of auditory and printed words (Maestu et al., 2002;Simos et al., 2002). The rationale for using the number of sequen-tial activity sources as a dependent measure is based on the
Figure 2. Schematic diagrams and time line of the verbal task. Each box represents a stimulus display event.The arrows represent the duration of each of these events. The Spanish words mean ‘gold’ (oro), ‘dagger’(punol), ‘lemon’ (limon), and ‘radio’ (radio) (stimulus display); ‘weapon’ (arma) (1st probe display); ‘fish’ (pez)(2nd probe display); and ‘metal’ (metal) (3rd probe display). ISI � interstimulus interval.
88 CAMPO ET AL.

premise that processing of an incoming stimulus requires a tran-sient increase in neural signaling in one or more neural popula-tions. This, in turn, produces a time-limited increase in intracellu-lar currents, which, once integrated, can be represented as anelectrical dipole. The strength of the dipole at each point in time isdirectly related to the strength of the resulting magnetic flux.Assuming the presence of a dipolar source (this can be ascertainedfrom the dipolar appearance of the surface distribution of recordedmagnetic flux), the degree of confidence with which the underlyingsource is estimated by the dipole fitting program directly reflectsthe strength of the recorded magnetic flux at that point in time. Thenumber of activity source solutions that exceed an empiricallydefined criterion reflects the amount of time that the rate ofneuronal signaling in a particular patch of cortex produces adetectable extracranial measure. Hence, the duration of focal cor-tical activity may be used as a measure of the degree of activationof a particular cortical region in a given task.
The identification of the cerebral areas where activity sourceswere observed was determined with a standard MRIcro softwareatlas of the human brain (Rorden & Brett, 2000) as a guide.Regions of interest are not established a priori in MEG studies.Modeling of activity sources is performed solely on the basis of thesurface distribution of magnetic flux without making hypotheses orplacing constraints regarding the anatomical location of the under-lying intracranial sources. When activity source locations are co-registered on individual MRI scans, the resulting individual acti-vation profiles are inspected visually (and blindly with respect toexperimental condition), to identify brain areas where activitysources are localized consistently across participants. Visual in-
spection of the resulting activation profiles showed that activitysources were computed consistently across participants in thefollowing areas: anterior ventrolateral prefrontal cortex (VLPFC);inferior frontal gyrus (IFG and anterior insula); dorsolateral pre-frontal cortex (DLPFC); premotor cortex and supplementary motorarea (PM/SMA); superior parietal lobe, including activity in theintraparietal sulcus (SPL); inferior parietal lobe (IPL); superiortemporal gyrus (STG); inferior temporal gyrus (ITG); posteriortemporal lobe (pTL); and occipital lobe. Because of the loweraccuracy of MEG localization in deep structures, we decided tocollapse all the sources identified on different medial temporallobe structures into one anatomical variable referred as MTL,including hippocampal gyrus, parahippocampal gyrus, entorhinalcortex, and subiculum, as described by Amaral and Insausti(1990).
For statistical purposes, activity sources observed in each of theabovementioned brain regions were grouped for non-overlappingsuccessive latency windows of 100 ms duration from 200 to 800ms (for a similar approach, see Papanicolaou et al., 2002; Stares-ina, Bauer, Deecke, & Walla, 2005; Tendolkar et al., 2000). Weonly considered physiological effects occurring between 200 msand 800 ms, as we found the most robust signals in this timeperiod.
Following other authors’ criteria (Mitchell, Johnson, Raye, &D’Esposito, 2000; Saito et al., 2005; Shafritz et al., 2002), weconsidered that a brain area was specifically related to integrationif it showed greater activity in the integrated condition than in boththe verbal and the spatial condition.
Figure 3. Schematic diagrams and time line of the integrated task trial type. Each box represents a stimulusdisplay event. The arrows represent the duration of each of these events. The Spanish words within the ellipsesmeans: ‘parrot’ (loro), ‘soup’ (sopa), ‘cross’ (cruz), and ‘silver’ (plata) (stimulus display); ‘fruit’ (ave) (1st probedisplay); ‘metal’ (metal) (2nd probe display); and ‘tree’ (arbor) (3rd probe display).
89VERBAL–SPATIAL BINDING
90 CAMPO ET AL.

Results
Behavioral Performance
Performance was assessed in terms of corrected hits for eachtask. Overall hit rate varied from 63.33% to 90.16% in the spatialtask (M � 78.26%; SD � 8.56); from 65.88% to 80.55% in theverbal task (M � 74.67%; SD � 3.79); and from 55.93%to 93.82% in the integrated task (M � 68.49%; SD � 12.69). Arepeated measures analysis of variance (ANOVA) on task aswithin-subject variable (verbal, spatial, integrated) was significant,F(2, 22) � 6.81, p � .05. Follow-up analyses showed that partic-ipants were more accurate in the spatial task than in the integratedtask, t(11) � 3.84, p � .05. Neither the spatial task nor theintegrated task differed from the verbal task in terms of accuracy,t(11) � 1.64, p � .05, and t(11) � 1.93, p � .05, respectively.Similar results have been observed in other studies of informationintegration (Olson & Jiang, 2002; Shafritz et al., 2002; Song &Jiang, 2005).
Brain Imaging Data
The mean number of sources of activation obtained for each ofthe abovementioned brain regions was submitted to repeated mea-sures ANOVA with task (verbal, spatial, integrated), hemisphere(left, right), and latency (200–300 ms, 300–400 ms, 400–500 ms,500–600 ms, 600–700 ms, 700–800 ms) as within-subject vari-ables. All ANOVA tests were evaluated with the Huynh-Feldtmethod as a precaution against the threat posed by inhomogene-ities of variance among cell means. Because of the large number ofcomparisons, results were Bonferroni corrected. Effects were con-sidered statistically significant when p remained less than .05 aftercorrection.
Task Differences in Spatiotemporal Activation Profiles
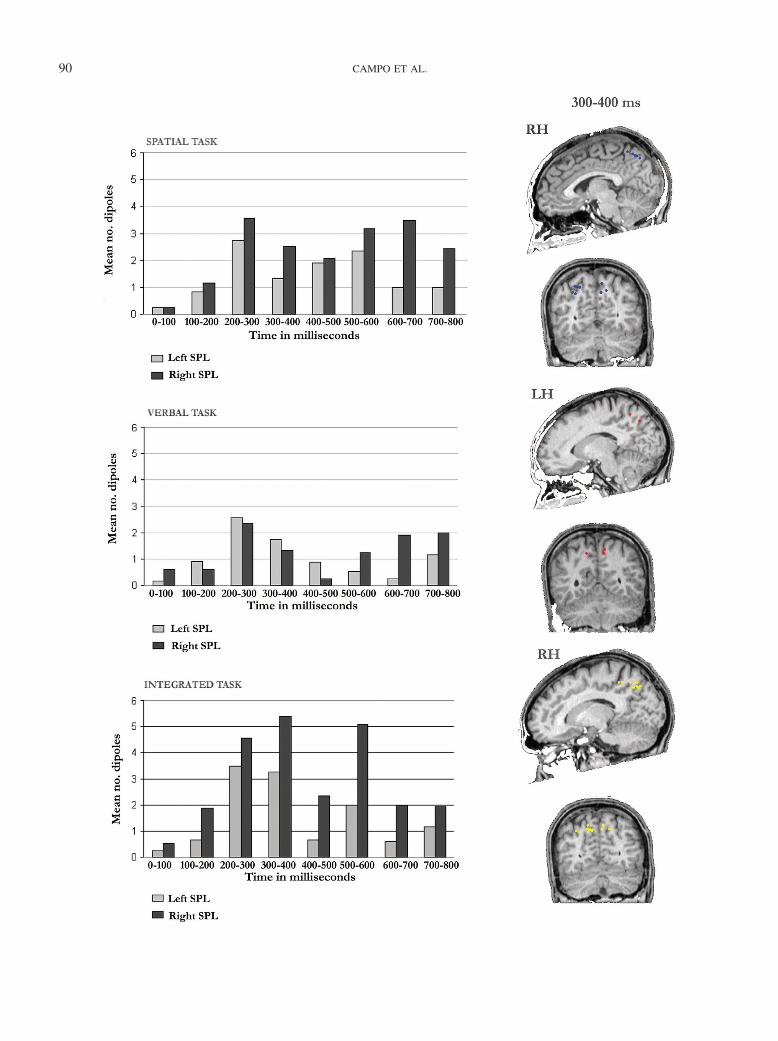
There was a main effect of task for SPL, F(2, 22) � 7.17, p �.05, �2 � .39. Further analyses revealed that SPL was moreactivated in the spatial and integrated tasks than in the verbal tasks,t(11) � 4.99, p � .01, �2 � .69, and, t(11) � 3.24, p � .01, �2 �.49, respectively. A significant task by latency interaction was alsofound for SPL, F(10, 110) � 2.90, p � .01, �2 � .21. Pairwisecomparisons indicated that SPL showed a greater number of ac-tivity sources between 300 and 400 ms in the integrated task ascompared with both the spatial, t(11) � 2.66, p � .05, �2 � .39,and the verbal tasks, t(11) � 4.96, p � .01, �2 � .69. Between 400and 500 ms, the spatial task engaged the SPL more than the verbaltask, t(11) � 2.99, p � .01, �2 � .45. Between 500 and 600 ms,both spatial and integrated tasks showed more activation than didthe verbal task, t(11) � 2.77, p � .05, �2 � .41, and t(11) � 3.59,p � .01, �2 � .54, respectively (see Figure 4). The differences in
the degree of activation between tasks were highly consistentacross participants: Between 300 and 400 ms, 9 out of 12 partic-ipants showed greater activation of SPL during the integrated taskthan in the spatial task, and 11 out of 12 showed greater activationin the integrated–verbal comparison. Between 500 and 600 ms, 7out of 12 participants showed greater activation of SPL during thespatial task than in the verbal task; 10 out of 12 showed moreactivation in the integrated task as compared with the verbal task.
The ANOVA also yielded a task by hemisphere interaction forDLPFC, F(2, 22) � 6.75, p � .01, �2 � .38. Results showed agreater number of activity sources in right DLPFC in the spatialtask than in the integrated task, t(11) � 2.91, p � .05, �2 � .43.This effect was found in 10 out of 12 participants.
A task by hemisphere interaction was also found for MTL, F(2,22) � 7.11, p � .01, �2 � .41. Further analyses revealed that leftMTL showed more activity sources during the verbal task ascompared with the spatial task, t(11) � 3.16, p � .01, �2 � .49,and the integrated task, t(11) � 2.71, p � .05, �2 � .40. Theverbal � spatial effect was found in 10 out of 12 participants, andthe verbal � integrated effect was found in 9 out of 12 participants.A review of statistical analyses can be seen in Table 1.
Task Difficulty
Increased task difficulty can affect the strength of neural re-sponses (Boudreau, Williford, & Maunsell, 2006; Donner et al.,2002; Shafritz et al., 2002). Thus, we aimed to tease apart thecontributions of task difficulty versus information integration toSPL activation during the integrated task. Accordingly, we dividedour sample into two groups according to a criterion of equivalencein performance (matched and unmatched) between the spatial taskand the integrated task. The threshold of group membership wasthe median of the differences between both tasks. Two groups of 6participants each were obtained. Using paired t tests, we thencompared performance between tasks. In the unmatched groupperformance in the spatial task (M � 77.83%, SD � 5.82) wassignificantly better than in the integrated task (M � 61.78%,SD � 7.30), t(5) � 8.09, p � .05. In the matched group, nodifferences were found between the spatial task (M � 78.70%,SD � 11.28) and the integrated task (M � 75.87%, SD � 12.39),t(5) � 1.51, p � .05. Although we had not found differences inperformance between the verbal and the integrated task in theoriginal sample, we found that in the unmatched group, perfor-mance in the verbal task (M � 73.68%, SD � 5.82) was signifi-cantly better than in the integrated task, t(5) � 3.77, p � .05. In thematched group, no differences were found between the verbal task(M � 75.65%, SD � 4.32) and the integrated task, t(5) � 0.55,p � .05. No significant differences were found between the spatialand the verbal tasks in the unmatched group, t(5) � 1.30, p � .05,or in the matched group, t(5) � 0.92, p � .05.
Figure 4 (opposite). Time courses of activation of the superior parietal lobe (SPL) of the left hemisphere (LH) and the right hemisphere (RH) during thespatial task (upper panel), the verbal task (middle panel), and the integrated task (lower panel), as indicated by the mean number of activity sources foundat successive 100-ms time windows (0–800 ms). Light shaded vertical areas represent left SPL activity and dark shaded vertical bars represent right SPL.Activity sources for a representative participant projected on the participant’s magnetic resonance image (MRI) are also depicted. Transverse slices andcoronal slices through parietal cortex show the activation in the latency during which activity was significantly greater in the integrated task as comparedwith the other two tasks are shown.
91VERBAL–SPATIAL BINDING

Tab
le1
Gro
upA
naly
ses
ofB
rain
Reg
ions
Act
ivat
edD
urin
gE
ncod
ing
Per
iod
Are
aM
ain
effe
ct:
Tas
kF
(2,
22)
Inte
ract
ion:
Tas
k�
Hem
isph
ere
F(2
,22
)E
ffec
tsi
ze:
�2
Inte
ract
ion:
Tas
k�
Lat
ency
F(1
0,11
0)E
ffec
tsi
ze:
�2
Inte
ract
ion:
Tas
k�
Hem
isph
ere
�L
aten
cyF
(10,
110)
Hem
isph
ere
Lat
ency
(ms)
Eff
ect
t(11
)E
ffec
tsi
ze:
�2
DL
PFC
1.43
6.75
*.3
81.
511.
56R
ight
Spat
ial
�In
tegr
ated
2.91
*.4
3V
LPF
C1.
491.
030.
921.
78IF
G1.
961.
810.
701.
92PM
/SM
A1.
601.
360.
940.
90SP
L7.
17*
1.08
2.90
*.3
90.
5730
0–40
0In
tegr
ated
�Sp
atia
l2.
66*
.39
4.96
**
.69
400–
500
Inte
grat
ed�
Ver
bal
2.99
*.4
53.
59**
.41
500–
600
Spat
ial
�V
erba
l2.
77*
.54
Inte
grat
ed�
Ver
bal
Spat
ial
�V
erba
lIP
L2.
172.
491.
151.
16ST
G1.
721.
201.
11.8
4IT
G0.
550.
341.
59.3
6PT
L0.
791.
990.
89.3
7M
TL
4.39
*7.
11**
.41
0.74
1.31
Lef
tV
erba
l�
Inte
grat
ed2.
71*
.40
3.16
*.4
9V
erba
l�
Spat
ial
Not
e.D
LPF
C�
dors
olat
eral
pref
ront
alco
rtex
;V
LPF
C�
vent
rola
tera
lpr
efro
ntal
cort
ex;
IFG
�in
feri
orfr
onta
lgy
rus;
PM/S
MA
�pr
emot
orco
rtex
and
supp
lem
enta
rym
otor
area
;SP
L�
supe
rior
pari
etal
lobe
;IP
L�
infe
rior
pari
etal
lobe
;ST
G�
supe
rior
tem
pora
lgy
rus;
ITG
�in
feri
orte
mpo
ral
gyru
s;PT
L�
post
erio
rte
mpo
ral
lobe
;M
TL
�m
edia
lte
mpo
ral
lobe
.*
p�
.05.
**
p�
.01.
92 CAMPO ET AL.
Because of the small size of the subsamples, we compared themean number of sources of activation on SPL for spatial task andintegrated task and for verbal task and integrated task of equal andunequal difficulty using a series of repeated measures ANOVAswith task (apatial vs. integrated; verbal vs. integrated), hemisphere(left, right), and latency (200–300 ms, 300–400 ms, 400–500 ms,500–600 ms, 600–700 ms, 700–800 ms) as within-subject vari-ables. All ANOVA tests were evaluated with the Huynh-Feldtmethod as a precaution against the threat posed by inhomogene-ities of variance among cell means. Because of the large number ofcomparisons, results were Bonferroni corrected. Effects were con-sidered statistically significant when p remained less than .05 aftercorrection.
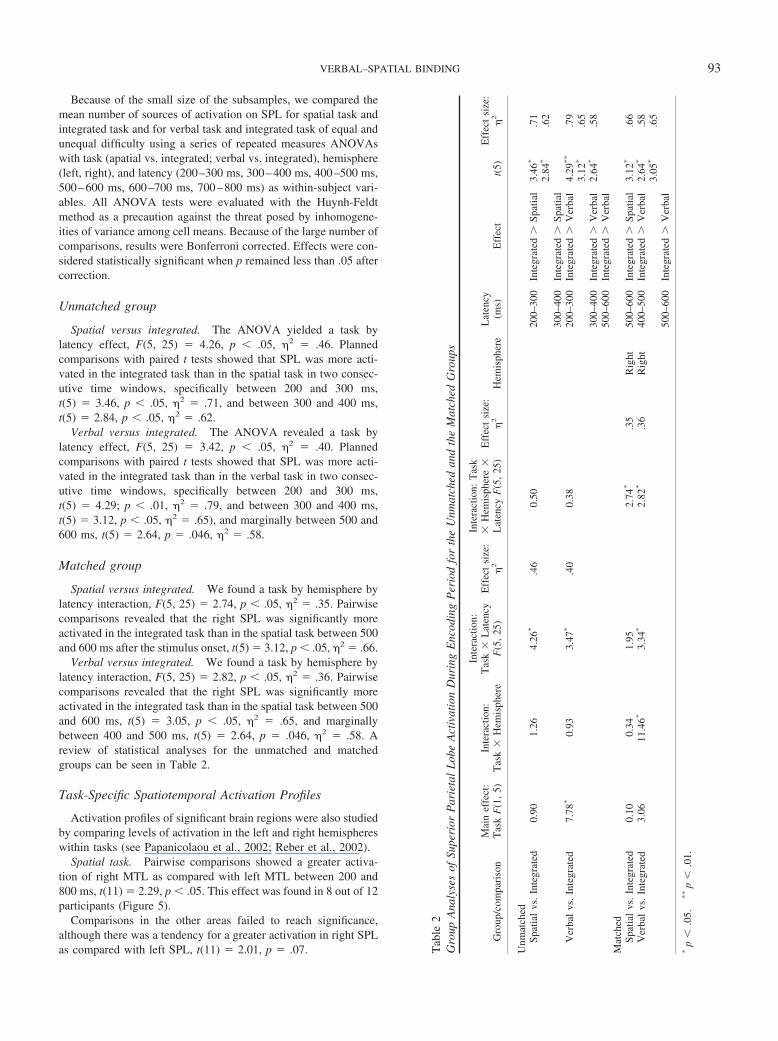
Unmatched group
Spatial versus integrated. The ANOVA yielded a task bylatency effect, F(5, 25) � 4.26, p � .05, �2 � .46. Plannedcomparisons with paired t tests showed that SPL was more acti-vated in the integrated task than in the spatial task in two consec-utive time windows, specifically between 200 and 300 ms,t(5) � 3.46, p � .05, �2 � .71, and between 300 and 400 ms,t(5) � 2.84, p � .05, �2 � .62.
Verbal versus integrated. The ANOVA revealed a task bylatency effect, F(5, 25) � 3.42, p � .05, �2 � .40. Plannedcomparisons with paired t tests showed that SPL was more acti-vated in the integrated task than in the verbal task in two consec-utive time windows, specifically between 200 and 300 ms,t(5) � 4.29; p � .01, �2 � .79, and between 300 and 400 ms,t(5) � 3.12, p � .05, �2 � .65), and marginally between 500 and600 ms, t(5) � 2.64, p � .046, �2 � .58.
Matched group
Spatial versus integrated. We found a task by hemisphere bylatency interaction, F(5, 25) � 2.74, p � .05, �2 � .35. Pairwisecomparisons revealed that the right SPL was significantly moreactivated in the integrated task than in the spatial task between 500and 600 ms after the stimulus onset, t(5) � 3.12, p � .05, �2 � .66.
Verbal versus integrated. We found a task by hemisphere bylatency interaction, F(5, 25) � 2.82, p � .05, �2 � .36. Pairwisecomparisons revealed that the right SPL was significantly moreactivated in the integrated task than in the spatial task between 500and 600 ms, t(5) � 3.05, p � .05, �2 � .65, and marginallybetween 400 and 500 ms, t(5) � 2.64, p � .046, �2 � .58. Areview of statistical analyses for the unmatched and matchedgroups can be seen in Table 2.
Task-Specific Spatiotemporal Activation Profiles
Activation profiles of significant brain regions were also studiedby comparing levels of activation in the left and right hemisphereswithin tasks (see Papanicolaou et al., 2002; Reber et al., 2002).
Spatial task. Pairwise comparisons showed a greater activa-tion of right MTL as compared with left MTL between 200 and800 ms, t(11) � 2.29, p � .05. This effect was found in 8 out of 12participants (Figure 5).
Comparisons in the other areas failed to reach significance,although there was a tendency for a greater activation in right SPLas compared with left SPL, t(11) � 2.01, p � .07. T
able
2G
roup
Ana
lyse
sof
Supe
rior
Par
ieta
lL
obe
Act
ivat
ion
Dur
ing
Enc
odin
gP
erio
dfo
rth
eU
nmat
ched
and
the
Mat
ched
Gro
ups
Gro
up/c
ompa
riso
nM
ain
effe
ct:
Tas
kF
(1,
5)In
tera
ctio
n:T
ask
�H
emis
pher
e
Inte
ract
ion:
Tas
k�
Lat
ency
F(5
,25
)E
ffec
tsi
ze:
�2
Inte
ract
ion:
Tas
k�
Hem
isph
ere
�L
aten
cyF
(5,
25)
Eff
ect
size
:�
2H
emis
pher
eL
aten
cy(m
s)E
ffec
tt(
5)E
ffec
tsi
ze:
�2
Unm
atch
edSp
atia
lvs
.In
tegr
ated
0.90
1.26
4.26
*.4
60.
5020
0–30
0In
tegr
ated
�Sp
atia
l3.
46*
.71
2.84
*.6
230
0–40
0In
tegr
ated
�Sp
atia
lV
erba
lvs
.In
tegr
ated
7.78
*0.
933.
47*
.40
0.38
200–
300
Inte
grat
ed�
Ver
bal
4.29
**
.79
3.12
*.6
530
0–40
0In
tegr
ated
�V
erba
l2.
64*
.58
500–
600
Inte
grat
ed�
Ver
bal
Mat
ched
Spat
ial
vs.
Inte
grat
ed0.
100.
341.
952.
74*
.35
Rig
ht50
0–60
0In
tegr
ated
�Sp
atia
l3.
12*
.66
Ver
bal
vs.
Inte
grat
ed3.
0611
.46*
3.34
*2.
82*
.36
Rig
ht40
0–50
0In
tegr
ated
�V
erba
l2.
64*
.58
3.05
*.6
550
0–60
0In
tegr
ated
�V
erba
l
*p
�.0
5.**
p�
.01.
93VERBAL–SPATIAL BINDING
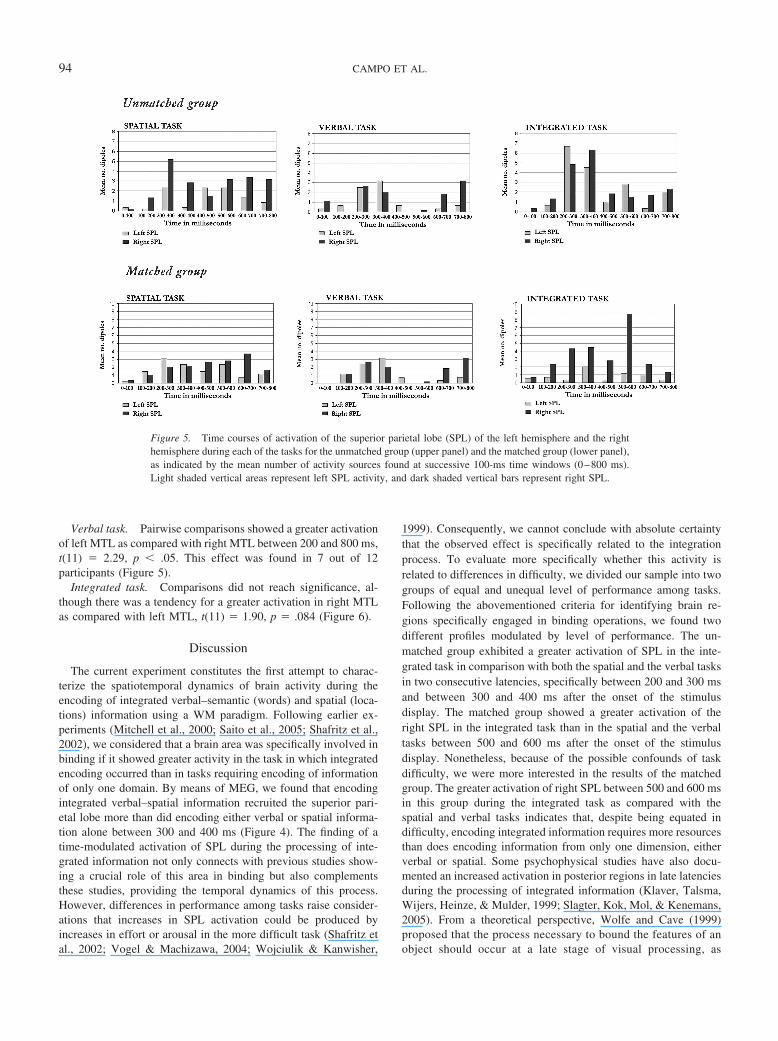
Verbal task. Pairwise comparisons showed a greater activationof left MTL as compared with right MTL between 200 and 800 ms,t(11) � 2.29, p � .05. This effect was found in 7 out of 12participants (Figure 5).
Integrated task. Comparisons did not reach significance, al-though there was a tendency for a greater activation in right MTLas compared with left MTL, t(11) � 1.90, p � .084 (Figure 6).
Discussion
The current experiment constitutes the first attempt to charac-terize the spatiotemporal dynamics of brain activity during theencoding of integrated verbal–semantic (words) and spatial (loca-tions) information using a WM paradigm. Following earlier ex-periments (Mitchell et al., 2000; Saito et al., 2005; Shafritz et al.,2002), we considered that a brain area was specifically involved inbinding if it showed greater activity in the task in which integratedencoding occurred than in tasks requiring encoding of informationof only one domain. By means of MEG, we found that encodingintegrated verbal–spatial information recruited the superior pari-etal lobe more than did encoding either verbal or spatial informa-tion alone between 300 and 400 ms (Figure 4). The finding of atime-modulated activation of SPL during the processing of inte-grated information not only connects with previous studies show-ing a crucial role of this area in binding but also complementsthese studies, providing the temporal dynamics of this process.However, differences in performance among tasks raise consider-ations that increases in SPL activation could be produced byincreases in effort or arousal in the more difficult task (Shafritz etal., 2002; Vogel & Machizawa, 2004; Wojciulik & Kanwisher,
1999). Consequently, we cannot conclude with absolute certaintythat the observed effect is specifically related to the integrationprocess. To evaluate more specifically whether this activity isrelated to differences in difficulty, we divided our sample into twogroups of equal and unequal level of performance among tasks.Following the abovementioned criteria for identifying brain re-gions specifically engaged in binding operations, we found twodifferent profiles modulated by level of performance. The un-matched group exhibited a greater activation of SPL in the inte-grated task in comparison with both the spatial and the verbal tasksin two consecutive latencies, specifically between 200 and 300 msand between 300 and 400 ms after the onset of the stimulusdisplay. The matched group showed a greater activation of theright SPL in the integrated task than in the spatial and the verbaltasks between 500 and 600 ms after the onset of the stimulusdisplay. Nonetheless, because of the possible confounds of taskdifficulty, we were more interested in the results of the matchedgroup. The greater activation of right SPL between 500 and 600 msin this group during the integrated task as compared with thespatial and verbal tasks indicates that, despite being equated indifficulty, encoding integrated information requires more resourcesthan does encoding information from only one dimension, eitherverbal or spatial. Some psychophysical studies have also docu-mented an increased activation in posterior regions in late latenciesduring the processing of integrated information (Klaver, Talsma,Wijers, Heinze, & Mulder, 1999; Slagter, Kok, Mol, & Kenemans,2005). From a theoretical perspective, Wolfe and Cave (1999)proposed that the process necessary to bound the features of anobject should occur at a late stage of visual processing, as
Figure 5. Time courses of activation of the superior parietal lobe (SPL) of the left hemisphere and the righthemisphere during each of the tasks for the unmatched group (upper panel) and the matched group (lower panel),as indicated by the mean number of activity sources found at successive 100-ms time windows (0–800 ms).Light shaded vertical areas represent left SPL activity, and dark shaded vertical bars represent right SPL.
94 CAMPO ET AL.
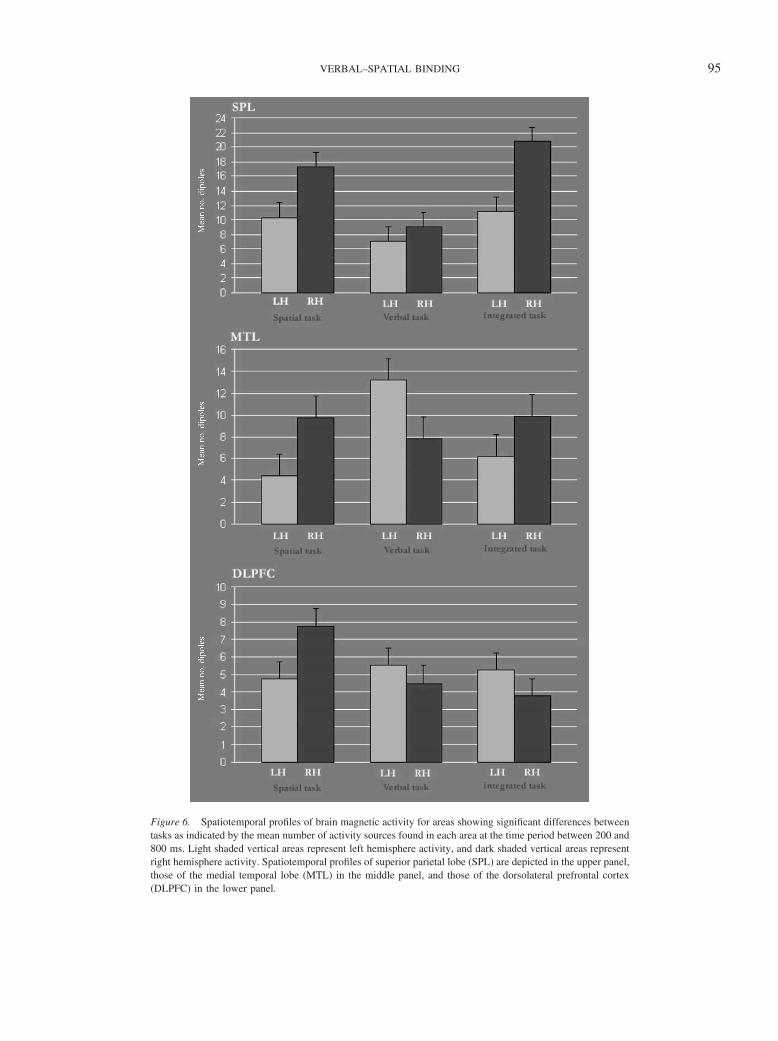
Figure 6. Spatiotemporal profiles of brain magnetic activity for areas showing significant differences betweentasks as indicated by the mean number of activity sources found in each area at the time period between 200 and800 ms. Light shaded vertical areas represent left hemisphere activity, and dark shaded vertical areas representright hemisphere activity. Spatiotemporal profiles of superior parietal lobe (SPL) are depicted in the upper panel,those of the medial temporal lobe (MTL) in the middle panel, and those of the dorsolateral prefrontal cortex(DLPFC) in the lower panel.
95VERBAL–SPATIAL BINDING

binding is only possible for objects selected by attention. It hasbeen shown that attentional control over information processingcan occur in early as well as late stages of processing (Hillyard& Anllo-Vento, 1998). This process occurring late in the infor-mation-processing pathway (Marois, Yi, & Chun, 2004) couldbe associated with the establishment of convergent associations,which enable the integration of different types of informationinto a multimodal representation (Saito et al., 2005; Vogel etal., 2001; Xu & Chun, 2006). Several studies have shown thatSPL is the neural basis responsible for representing these inte-grated representations, not only in the visual domain (Donner etal., 2002; Klaver et al., 1999; Lee & Chun, 2001; Luck &Vogel, 1997; Shomstein & Behrmann, 2006; Slagter et al.,2005; Todd & Marois, 2004; Xu & Chun, 2006) but also incross-modal representations (Macaluso & Driver, 2005; Shom-stein & Yantis, 2004). The time window in which we observedthe effect matches well with the time it takes to consciouslyidentify and consolidate a visual stimulus in WM (Jolicoeur,Sessa, Dell’Acqua, & Robitaille, 2006; Klaver et al., 1999;Marois & Ivanoff, 2005; Sergent, Baillet, & Dehaene, 2005).This probably signals a second stage of processing that appearsto be crucial for conscious processing of information (Chun &Potter, 1995; Duncan, 1980), as revealed by studies using theattentional blink paradigm (Raymond, Shapiro, & Arnell,1992). The fact that this period was the same as that in whichwe found the differences between the spatial task and the verbaltask could also point to a crucial role of spatial WM/attentionalmechanisms. It is important to take into account that it is verydifficult to distinguish whether these processes are attentionalbased or WM based, as several lines of research have shown agreat overlap between the neural substrates of both types ofprocesses (Awh & Jonides, 2001; LaBar, Gitelman, Parrish, &Mesulam, 1999; Lee & Chun, 2001; Lepsien & Nobre, 2006).Nonetheless, the differential contribution of these systemsshould be elucidated in future studies. One possibility couldinvolve a spatial cueing experiment in which the locus ofselection is manipulated by varying the cue–target stimulusonset asynchrony (SOA) as described by Vogel, Woodman, andLuck (2005; see also Bodelon, Fallah, & Reynolds, 2007;Lepsien & Nobre, 2006; Vogel, McCollough, & Machizawa,2005).
The fact that during the integrated task, participants encodemore items (eight) than in either the verbal or the spatial task (fouritems in each) could lead to alternative explanations of the increaseof activation observed in the SPL by a load effect. However, if weconsidered a strong line of research proposing that what peoplerepresent in WM are integrated object percepts (Lee & Chun,2001; Luck & Vogel, 1997; Vogel et al., 2001; Woodman & Luck,2004), we could accept that the number of objects represented inWM across tasks in the current experiment is the same, varyingonly in complexity. Several studies have shown that activity inSPL does not reflect the perceptual representation of objects in thevisual scene but rather the objects represented in WM (Song &Jiang, 2006; Todd & Marois, 2004, 2005; Vogel & Machizawa,2004). That is, although there is an increase in the activation ofposterior parietal cortex as a function of load, this increment doesnot extend to supracapacity set sizes (beyond four objects). Shaf-ritz et al. (2002) also demonstrated that the greater parietal acti-vation observed during a binding task could not be explained in
terms of load. Taken together, these findings do not support thatthe greater activation of the SPL observed in our study is due to aload effect.
Although previous neuroimaging studies have showed that theMTL and the DLPFC are involved in location binding (Mitchell etal., 2000; Prabhakaran et al., 2000; Simon-Thomas et al., 2003;Sommer, Rose, Glascher, Wolbers, & Buchel, 2005; Sommer,Rose, Weiller, & Buchel, 2005), we did not find a specific activa-tion of any of these areas during the integration of words andlocations. It is interesting, however, to indicate that the pattern ofactivation of MTL was very similar in the spatial and the inte-grated task, showing a right hemisphere asymmetry, whereas thisactivity was clearly left lateralized in the verbal task (Figure 5). Asrecently reported by Sommer et al. (2005), the connectivity of theSPL and the MTL during object location could be reflecting thelevel of processing of the spatial information. Whereas this idea ofa spatial processing network matches with the aforementionedstudies proposing a crucial role of spatial selective attention inbinding, we did not find a specific latency effect that could link thisactivation to any specific process, either attentional based or work-ing memory based.
Finally, the greater activation of right DLPFC observed in thespatial task as compared with the integrated task was unexpected,as several experiments have reported an increased activation ofthis area during binding (Mitchell et al., 2000; Simon-Thomas etal., 2003). Nonetheless, from a behavioral perspective, Allen et al.(2006) have suggested that encoding feature configurations intoWM as integrated representations might not be particularly de-manding of executive processes. Other authors have also failed tofind a specific contribution of DLPFC during tasks requiringintegration of information (Donner et al., 2002; Owen, 2004; Song& Jiang, 2006; Todd & Marois, 2004, 2005). With regard tocurrent data, we do not have a plausible explanation for thisdifference between tasks.
In summary, similarly to finding of other researchers (Olson& Jiang, 2002; Song & Jiang, 2005; Xu & Chun, 2005), ourresults suggest that the neural mechanisms supporting the en-coding of integrated information are modulated not only by thenumber of objects but also by the complexity of these objects(Alvarez & Cavanagh, 2004; Olson & Jiang, 2002). The currentfindings are partially consistent with models proposing thatspatial selective attention is crucial for integrating informationand that this process occurs in a relatively late stage in thestream of information processing, leading to the creation of anintegrated object or percept (Macaluso & Driver, 2005). How-ever, we considered that the latencies in cases where the spe-cific effect was observed are in greater agreement with propos-als suggesting that SPL participates in binding processes byencoding and maintaining a detailed complex-integrated repre-sentation in WM (O’Craven et al., 1999; Saito et al., 2005;Slagter et al., 2005; Todd & Marois, 2004; Vogel et al., 2001;Xu & Chun, 2006; see also Shafritz et al., 2002). Nonetheless,the difficulty of distinguishing attention-based processes fromprocesses of WM encoding has been highlighted by otherstudies addressing this issue (Awh & Jonides, 2001; Lee &Chun, 2001; Lepsien & Nobre, 2006). In consideration ofBaddeley’s (2000) episodic buffer, it appears that the samemechanisms involved in integrating information within onesubsystem (i.e., visuospatial buffer) also support the integration
96 CAMPO ET AL.

of information between previously considered independent sub-systems (i.e., verbal and visuospatial buffers). Therefore, thesefindings constitute new evidence of the engagement of posteriorparietal regions in functional integration of information fromdifferent domains (Calvert, Spence, & Stein, 2004; Macaluso &Driver, 2005). It also appears that SPL participates in integrat-ing information during encoding and maintenance stages(Campo et al., 2005; Lepsien & Nobre, 2006; Todd & Marois,2004; Xu & Chun, 2006).
References
Algarabel, S. (1996). Indices de interes psicolinguıstico de 1917 palabrascastellanas [Indexes of psycholinguistic interest for 1917 Spanishwords]. Cognitiva, 8, 43–88.
Allen, R. J., Baddeley, A. D., & Hitch, G. J. (2006). Is the binding of visualfeatures in working memory resource-demanding? Journal of Experi-mental Psychology: General, 135, 298–313.
Alvarez, G. A., & Cavanagh, P. (2004). The capacity of visual short-termmemory is set both by visual information load and by number of objects.Psychological Science, 15, 106–111.
Amaral, D., & Insausti, R. (1990). Hippocampal formation. In G. Paxinos(Ed.), The human nervous system (pp. 711–755). San Diego, CA: Aca-demic Press.
Awh, E., & Jonides, J. (2001). Overlapping mechanisms of attention andspatial working memory. Trends in Cognitive Science, 5, 119–126.
Awh, E., Vogel, E. K., & Oh, S. H. (2006). Interactions between attentionand working memory. Neuroscience, 139, 201–208.
Baddeley, A. (1996). The fractionation of working memory. Proceedingsof the National Academy of Sciences of the United States of America, 93,13468–13472.
Baddeley, A. (2000). The episodic buffer: A new component of workingmemory. Trends in Cognitive Neuroscience, 4, 417–423.
Baddeley, A., & Hitch, G. (1974). Working memory. In G. A. Bower (Ed.),The psychology of learning and motivation (Vol. 8, pp. 48–79.). NewYork: Academic Press.
Becker, J. T., MacAndrew, D. K., & Fiez, J. A. (1999). A comment on thefunctional localization of the phonological storage subsystem of workingmemory. Brain and Cognition, 41, 27–38.
Bodelon, C., Fallah, M., & Reynolds, J. H. (2007). Temporal resolution forthe perception of features and conjunctions. Journal of Neuro-science, 27, 725–730.
Boudreau, C. E., Williford, T. H., & Maunsell, J. H. R. (2006). Effects oftask difficulty and target likelihood in area V4 of macaque monkeys.Journal of Neurophysiology, 96, 2377–2387.
Calvert, G. A., Spence, C., & Stein, B., E. (2004). The handbook ofmultisensory processes. Cambridge, MA: MIT Press.
Campo, P., Maestu, F., Ortiz, T., Capilla, A., Santiuste, M., Fernandez, A.,et al. (2005). Time modulated prefrontal and parietal activity during themaintenance of integrated information as revealed by magnetoencepha-lography. Cerebral Cortex, 15, 123–130.
Castillo, E. M., Simos, P. G., Wheless, J. W., Baumgartner, J. E., Breier,J. I., Billingsley, R. L., et al. (2004). Integrating sensory and motormapping in a comprehensive MEG protocol: Clinical validity and rep-licability. Neuroimage, 21, 973–983.
Chun, M. M., & Potter, M. C. (1995). A two-stage model for multipletarget detection in rapid serial visual presentation. Journal of Experi-mental Psychology: Human Perception and Performance, 21, 109–127.
Corbetta, M., Shulman, G. L., Miezin, F. M., & Petersen, S. E. (1995).Superior parietal cortex activation during spatial attention shifts andvisual feature conjunction. Science, 270(5237), 802–805.
Cowan, N. (2001). The magical number 4 in short-term memory: Areconsideration of mental storage capacity. Behavioral and Brain Sci-ences, 24, 87–114; discussion, pp. 114–185.
D’Esposito, M., Postle, B. R., Ballard, D., & Lease, J. (1999). Maintenanceversus manipulation of information held in working memory: An event-related fMRI study. Brain and Cognition, 41, 66–86.
Donner, T. H., Kettermann, A., Diesch, E., Ostendorf, F., Villringer, A., &Brandt, S. A. (2002). Visual feature and conjunction searches of equaldifficulty engage only partially overlapping frontoparietal networks.Neuroimage, 15, 16–25.
Duncan, J. (1980). The locus of interference in the perception of simulta-neous stimuli. Psychological Review, 87, 272–300.
Duncan, J. (1984). Selective attention and the organization of visualinformation. Journal of Experimental Psychology: General, 113, 501–517.
Eng, H. Y., Chen, D., & Jiang, Y. (2005). Visual working memory forsimple and complex visual stimuli. Psychonomic Bulletin and Re-view, 12, 1127–1133.
Ericsson, K. A., Chase, W. G., & Faloon, S. (1980). Acquisition of amemory skill. Science, 208, 1181–1182.
Eriksen, C. W., & Yeh, Y. Y. (1985). Allocation of attention in the visualfield. Journal of Experimental Psychology: Human Perception andPerformance, 11, 583–597.
Gray, C. M. (1999). The temporal correlation hypothesis of visual featureintegration: Still alive and well. Neuron, 24, 31–47, 111–125.
Haxby, J. V., Petit, L., Ungerleider, L. G., & Courtney, S. M. (2000).Distinguishing the functional roles of multiple regions in distributedneural systems for visual working memory. Neuroimage, 11, 380 –391.
Hillyard, S. A., & Anllo-Vento, L. (1998). Event-related brain potentials inthe study of visual selective attention. Proceedings of the NationalAcademy of Sciences of the United States of America, 95, 781–787.
Jolicoeur, P., Sessa, P., Dell’Acqua, R., & Robitaille, N. (2006). Atten-tional control and capture in the attentional blink paradigm: Evidencefrom human electrophysiology. European Journal of Cognitive Psychol-ogy, 18, 560–578.
Klaver, P., Talsma, D., Wijers, A. A., Heinze, H. J., & Mulder, G. (1999).An event-related brain potential correlate of visual short-term memory.Neuroreport, 10, 2001–2005.
LaBar, K. S., Gitelman, D. R., Parrish, T. B., & Mesulam, M. (1999).Neuroanatomic overlap of working memory and spatial attention net-works: A functional MRI comparison within subjects. Neuroimage, 10,695–704.
Lee, D., & Chun, M. M. (2001). What are the units of visual short-termmemory, objects or spatial locations? Perception and Psychophysics, 63,253–257.
Lepsien, J., & Nobre, A. C. (2006). Cognitive control of attention in thehuman brain: Insights from orienting attention to mental representations.Brain Research, 1105, 20–31.
Logie, R. H. (1995). Visuo-spatial working memory. Hove, United King-dom: Erlbaum.
Luck, S. J., & Vogel, E. K. (1997). The capacity of visual working memoryfor features and conjunctions. Nature, 390(6657), 279–281.
Macaluso, E., & Driver, J. (2005). Multisensory spatial interactions: Awindow onto functional integration in the human brain. Trends inNeurosciences, 28, 264–271.
Maestu, F., Ortiz, T., Fernandez, A., Amo, C., Martin, P., Fernandez, S., etal. (2002). Spanish language mapping using MEG: A validation study.Neuroimage, 17, 1579–1586.
Marois, R., & Ivanoff, J. (2005). Capacity limits of information processingin the brain. Trends in Cognitive Science, 9, 296–305.
Marois, R., Yi, D. J., & Chun, M. M. (2004). The neural fate of consciouslyperceived and missed events in the attentional blink. Neuron, 41, 465–472.
97VERBAL–SPATIAL BINDING

Miller, G. A. (1956). The magical number seven plus or minus two: Somelimits on our capacity for processing information. Psychological Re-view, 63, 81–97.
Mitchell, K. J., Johnson, M. K., Raye, C. L., & D’Esposito, M. (2000).fMRI evidence of age-related hippocampal dysfunction in feature bind-ing in working memory. Cognitive Brain Research, 10, 197–206.
Miyake, A., & Shah, P. (1999). Models of working memory. Cambridge,England: Cambridge University Press.
More, J. (1977). The Levenberg–Marquardt algorith: Implementation andtheory. In G. Watson (Ed.), Numerical analysis (pp. 105–116): NewYork: Springer Verlag.
O’Craven, K. M., Downing, P. E., & Kanwisher, N. (1999). fMRI evidencefor objects as the units of attentional selection. Nature, 401, 584–587.
Olson, I. R., & Jiang, Y. (2002). Is visual short-term memory object based?Rejection of the “strong-object” hypothesis. Perception and Psycho-physics, 64, 1055–1067.
Owen, A. M. (2004). Working memory: Imaging the magic number four.Current Biology, 14, R573–R574.
Papanicolaou, A. C., Simos, P. G., Breier, J. I., Zouridakis, G., Willmore,L. J., Wheless, J. W., et al. (1999). Magnetoencephalographic mappingof the language-specific cortex. Journal of Neurosurgery, 90, 85–93.
Papanicolaou, A. C., Simos, P. G., Castillo, E. M., Breier, J. I., Katz, J. S.,& Wright, A. A. (2002). The hippocampus and memory of verbal andpictorial material. Learning and Memory, 9, 99–104.
Papanicolaou, A. C., Simos, P. G., Castillo, E. M., Breier, J. I., Sarkari,S., Pataraia, E., et al. (2004). Magnetocephalography: A noninvasivealternative to the Wada procedure. Journal of Neurosurgery, 100,867– 876.
Prabhakaran, V., Narayanan, K., Zhao, Z., & Gabrieli, J. D. (2000).Integration of diverse information in working memory within the frontallobe. Nature Neuroscience, 3, 85–90.
Raymond, J. E., Shapiro, K. L., & Arnell, K. M. (1992). Temporarysuppression of visual processing in an RSVP task: An attentional blink?Journal of Experimental Psychology: Human Perception and Perfor-mance, 18, 849–860.
Reber, P. J., Siwiec, R. M., Gitelman, D. R., Parrish, T. B., Mesulam,M. M., Paller, K. A., et al. (2002). Neural correlates of successfulencoding identified using functional magnetic resonance imaging. Jour-nal of Neuroscience, 22, 9541–9548.
Robertson, L. C. (2003). Binding, spatial attention and perceptual aware-ness. Nature Reviews Neuroscience, 4, 93–102.
Rorden, C., & Brett, M. (2000). Stereotaxic display of brain lesions.Behavioral Neurology, 12, 191–200.
Ruchkin, D. S., Grafman, J., Cameron, K., & Berndt, R. S. (2003).Working memory retention systems: A state of activated long-termmemory. Behavioral and Brain Sciences, 26, 709–728; discussion 728–777.
Saito, D. N., Yoshimura, K., Kochiyama, T., Okada, T., Honda, M., &Sadato, N. (2005). Cross-modal binding and activated attentional net-works during audio-visual speech integration: A functional MRI study.Cerebral Cortex, 15, 1750–1760.
Sarvas, J. (1987). Basic mathematical and electromagnetic concepts of thebiomagnetic inverse problem. Physics in Medicine and Biology, 32,11–22.
Sergent, C., Baillet, S., & Dehaene, S. (2005). Timing of the brain eventsunderlying access to consciousness during the attentional blink. NatureNeuroscience, 8, 1391–1400.
Shafritz, K. M., Gore, J. C., & Marois, R. (2002). The role of the parietalcortex in visual feature binding. Proceedings of the National Academy ofSciences of the United States of America, 99, 10917–10922.
Shomstein, S., & Behrmann, M. (2006). Cortical systems mediating visualattention to both objects and spatial locations. Proceedings of the Na-tional Academy of Sciences of the United States of America, 103,11387–11392.
Shomstein, S., & Yantis, S. (2004). Control of attention shifts betweenvision and audition in human cortex. Journal of Neuroscience, 24,10702–10706.
Simon, H. A. (1974). How big is a chunk? Science, 183, 482–488.Simon-Thomas, E. R., Brodsky, K., Willing, C., Sinha, R., & Knight,
R. T. (2003). Distributed neural activity during object, spatial andintegrated processing in humans. Cognitive Brain Research, 16,457– 467.
Simos, P. G., Breier, J. I., Fletcher, J. M., Foorman, B. R., Castillo,E. M., & Papanicolaou, A. C. (2002). Brain mechanisms for readingwords and pseudowords: An integrated approach. Cerebral Cor-tex, 12, 297–305.
Slagter, H. A., Kok, A., Mol, N., & Kenemans, J. L. (2005). Spatio-temporal dynamics of top-down control: Directing attention to locationand/or color as revealed by ERPs and source modeling. Cognitive BrainResearch, 22, 333–348.
Smith, E. E., Jonides, J., Marshuetz, C., & Koeppe, R. A. (1998). Com-ponents of verbal working memory: Evidence from neuroimaging. Pro-ceedings of the National Academy of Sciences of the United States ofAmerica, 95, 876–882.
Sommer, T., Rose, M., Glascher, J., Wolbers, T., & Buchel, C. (2005).Dissociable contributions within the medial temporal lobe to encoding ofobject–location associations. Learning and Memory, 12, 343–351.
Sommer, T., Rose, M., Weiller, C., & Buchel, C. (2005). Contributions ofoccipital, parietal and parahippocampal cortex to encoding of object–location associations. Neuropsychologia, 43, 732–743.
Song, J. H., & Jiang, Y. (2006). Visual working memory for simple andcomplex features: An fMRI study. Neuroimage, 30, 963–972.
Staresina, B. P., Bauer, H., Deecke, L., & Walla, P. (2005). Neurocognitivecorrelates of incidental verbal memory encoding: A magnetoencephalo-graphic (MEG) study. Neuroimage, 25, 430–443.
Tendolkar, I., Rugg, M., Fell, J., Vogt, H., Scholz, M., Hinrichs, H., et al.(2000). A magnetoencephalographic study of brain activity related torecognition memory in healthy young human subjects. NeuroscienceLetters, 280, 69–72.
Todd, J. J., & Marois, R. (2004). Capacity limit of visual short-termmemory in human posterior parietal cortex. Nature, 428, 751–754.
Todd, J. J., & Marois, R. (2005). Posterior parietal cortex activity predictsindividual differences in visual short-term memory capacity. Cognitive,Affective, and Behavioral Neuroscience, 5, 144–155.
Treisman, A. (1999). Solutions to the binding problem: Progress throughcontroversy and convergence. Neuron, 24, 105–110, 111–125.
Vogel, E. K., & Machizawa, M. G. (2004). Neural activity predicts indi-vidual differences in visual working memory capacity. Nature,428(6984), 748–751.
Vogel, E. K., McCollough, A. W., & Machizawa, M. G. (2005). Neuralmeasures reveal individual differences in controlling access to workingmemory. Nature, 438(7067), 500–503.
Vogel, E. K., Woodman, G. F., & Luck, S. J. (2001). Storage offeatures, conjunctions and objects in visual working memory. Jour-nal of Experimental Psychology: Human Perception and Perfor-mance, 27, 92–114.
Vogel, E. K., Woodman, G. F., & Luck, S. J. (2005). Pushing around thelocus of selection: Evidence for the flexible-selection hypothesis. Jour-nal of Cognitive Neuroscience, 17, 1907–1922.
Wheeler, M. E., & Treisman, A. M. (2002). Binding in short-term visualmemory. Journal of Experimental Psychology: General, 131, 48–64.
Wojciulik, E., & Kanwisher, N. (1999). The generality of parietal involve-ment in visual attention. Neuron, 23, 747–764.
Wolfe, J. M., & Cave, K. R. (1999). The psychophysical evidence for abinding problem in human vision. Neuron, 24, 11–17, 111–125.
Wolfe, J. M., Yu, K. P., Stewart, M. I., Shorter, A. D., Friedman-Hill, S. R.,& Cave, K. R. (1990). Limitations on the parallel guidance of visual
98 CAMPO ET AL.

search: Color � color and orientation � orientation conjunctions. Jour-nal of Experimental Psychology: Human Perception and Perfor-mance, 16, 879–892.
Woodman, G. F., & Luck, S. J. (2004). Visual search is slowed whenvisuospatial working memory is occupied. Psychonomic Bulletin andReview, 11, 269–274.
Xu, Y., & Chun, M. M. (2005). Dissociable neural mechanisms supportingvisual short-term memory for objects. Nature, 440, 91–95.
Xu, Y., & Chun, M. M. (2006). Dissociable neural mechanisms supportingvisual short-term memory for objects. Nature, 440(7080), 91–95.
Zurowski, B., Gostomzyk, J., Gron, G., Weller, R., Schirrmeister, H.,Neumeier, B., et al. (2002). Dissociating a common working memorynetwork from different neural substrates of phonological and spatialstimulus processing. Neuroimage, 15, 45–57.
Received November 27, 2006Revision received June 14, 2007
Accepted June 25, 2007 �
99VERBAL–SPATIAL BINDING
Related Documents