of July 25, 2022. This information is current as Susceptible to HIV Infection Are Cells Require FoxP3 for Suppression and Naive Precursors of Human Regulatory T and Derya Unutmaz Michelle Tseng, Christopher W. Arendt, Spyros A. Kalams Amanda K. Antons, Rui Wang, Kyra Oswald-Richter, http://www.jimmunol.org/content/180/2/764 doi: 10.4049/jimmunol.180.2.764 2008; 180:764-773; ; J Immunol References http://www.jimmunol.org/content/180/2/764.full#ref-list-1 , 27 of which you can access for free at: cites 68 articles This article average * 4 weeks from acceptance to publication Fast Publication! • Every submission reviewed by practicing scientists No Triage! • from submission to initial decision Rapid Reviews! 30 days* • Submit online. ? The JI Why Subscription http://jimmunol.org/subscription is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/About/Publications/JI/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/alerts Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2008 by The American Association of 1451 Rockville Pike, Suite 650, Rockville, MD 20852 The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on July 25, 2022 http://www.jimmunol.org/ Downloaded from by guest on July 25, 2022 http://www.jimmunol.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

of July 25, 2022.This information is current as
Susceptible to HIV InfectionAreCells Require FoxP3 for Suppression and
Naive Precursors of Human Regulatory T
and Derya UnutmazMichelle Tseng, Christopher W. Arendt, Spyros A. Kalams Amanda K. Antons, Rui Wang, Kyra Oswald-Richter,
http://www.jimmunol.org/content/180/2/764doi: 10.4049/jimmunol.180.2.764
2008; 180:764-773; ;J Immunol
Referenceshttp://www.jimmunol.org/content/180/2/764.full#ref-list-1
, 27 of which you can access for free at: cites 68 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2008 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on July 25, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on July 25, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from

Naive Precursors of Human Regulatory T Cells Require FoxP3for Suppression and Are Susceptible to HIV Infection1
Amanda K. Antons,* Rui Wang,2‡ Kyra Oswald-Richter,2* Michelle Tseng,‡
Christopher W. Arendt,§ Spyros A. Kalams,*† and Derya Unutmaz3‡
CD4�CD25� human regulatory T cells (Treg cells), which express the transcription factor FoxP3, suppress T cell activation. In thisstudy, we sought to define cellular and molecular mechanisms of human Treg cell differentiation. A subset of human naive CD4�
T cells that are CD25� express high levels of FoxP3. We show that upon activation through the TCR, these FoxP3-expressing naiveT cells (termed TNreg cells) greatly expand in vitro. Expanded TNreg cells acquire a full Treg phenotype with potent suppressiveactivity and display low IL-2 production upon TCR stimulation. TNreg cells in which FoxP3 expression was reduced through RNAinterference lost their suppressive activity, but retained their low IL-2 secretion in response to TCR stimulation. Furthermore, insupport of the notion that TNreg cells represent a separate lineage of naive cells, we found that they were more susceptible to HIVinfection as compared with naive CD4� T cells. Based on these findings, we propose that TNreg cells are precursors for human Treg
cells and that these cells require a high level of FoxP3 expression to maintain their suppressive function. Accordingly, modulationof TNreg cell numbers during infections such as HIV may disrupt human Treg cell development, and contribute to chronic immuneactivation. The Journal of Immunology, 2008, 180: 764–773.
T he adaptive immune system has evolved several mecha-nisms to prevent responsiveness to autoantigens, includ-ing elimination of self-reactive T cells during thymic se-
lection or functional inactivation in the periphery (1). It is nowclear that professional suppressor T cells called regulatory T cells(Treg cells)4 (2) represent a major component of regulating un-wanted T cell activation (2, 3). Treg cells are very potent in sup-pressing the activation and proliferation of T cells, through amechanism that is likely cell-contact dependent (4–9). Indeed, de-letion or elimination of Treg cells in vivo results in T cell hyper-activation and severe autoimmunity (10–12).
In both humans and mice, the Treg subset is defined asCD4�CD25� cells that express high levels of the transcriptionfactor FoxP3, which is required for their development in vivo (13,14). A natural mutation or targeted deletion of the mouse FoxP3gene results in CD4� T cell-mediated fatal lymphoproliferativedisease (11, 13). Mutation of the FoxP3 gene in humans also re-sults in similar T cell hyperactivation and immune dysregulation
characterized by polyendocrinopathy and enteropathy, called IPEXsyndrome (12, 15). FoxP3 has been shown to play a role in tran-scriptional repression (16–20) and induction (19–21) and appearsto be necessary for mature murine Treg cell function (22, 23). Therole of FoxP3 in human Treg cell development and function is notyet defined.
Treg cells appear to develop as a functionally distinct subsetanalogous to conventional T cell development in thymus (24). Inmice, Treg cells are selected after recognition of self-Ag in thethymus, forming a distinct subset of T cells expressing high levelsof CD25 (25). It is not yet clear whether human Treg cells undergomaturation in the thymus or whether they arise from a peripheralprecursor subset.
Human Treg cells express the memory marker CD45RO, andseveral activation markers such as glucocorticoid-induced TNFR(GITR) and HLA-DR, in addition to high levels of CD25 andFoxP3 (26). Recent studies indicate that Treg cells lack expressionof the IL-7 receptor CD127 (18, 27). However, Treg cells withmemory markers are not observed in neonatal blood, suggestingthat human Treg cells potentially develop postnatally from precur-sor cells in the periphery. A subset of human T cells that expressthe naive phenotype of CD45RO�CD45RA� has been identifiedthat expresses CD25 and possesses Treg characteristics, includingFOXP3 expression and partial suppressive function (28–30). Butdetailed characteristics and proliferative capacity of this novel sub-set of T cells remains to be elucidated. It is also not yet knownwhether sustained expression of FoxP3 is required to maintain thephenotype and function of human Treg cells and whether they de-velop from naive precursors in vivo.
Human Treg cells have been shown to be highly relevant forunderstanding HIV pathogenesis (26, 31–37). We have previ-ously shown that Treg cells are depleted in some HIV-infectedindividuals and that Treg cells are susceptible to HIV infection(26). This depletion of Treg cells in HIV-infected patients maycontribute to the excessive T cell hyperactivation observed inpatients by reducing the active suppression of conventional Tcells (26, 31, 36). It is also possible that an excessive Treg
*Department of Microbiology and Immunology and †Department of Medicine,Vanderbilt University School of Medicine, Nashville, TN 37232; ‡Department ofMicrobiology, New York School of Medicine, New York, NY 10016; and §OncologyDepartment, Sanofi-Aventis, Bridgewater, NJ 08807
Received for publication July 20, 2007. Accepted for publication November 8, 2007.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was support by National Institutes of Health Grant R01 AI065303 (toD.U.).2 R.W. and K.O.-R. contributed equally.3 Address correspondence and reprint requests to Dr. Derya Unutmaz, Department ofMicrobiology, New York School of Medicine, Smilow Research Center, 522 FirstAvenue, Smilow Building 10th Floor, New York, NY 10016. E-mail address:[email protected] Abbreviations used in this paper: Treg, regulatory T cell; GITR, glucocorticoid-induced TNFR; TN, naive CD4� T cell; TNreg, FoxP3�CD25� naive T cell; TM,memory CD4� T cell; DC, dendritic cell; SEB, staphylococcal enterotoxin B; shRNA,short hairpin RNA; CBA, cytometric bead array; VSV-G, vesicular stomatitis virus gly-coprotein; MOI, multiplicity of infection; RNAi, RNA interference; IFU, infectious unit.
Copyright © 2008 by The American Association of Immunologists, Inc. 0022-1767/08/$2.00
The Journal of Immunology
www.jimmunol.org
by guest on July 25, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

response can blunt HIV-specific immune responses or reducethe effectiveness of potential vaccines (33, 35, 37). Therefore,deciphering human Treg cell biology would benefit our under-standing of the perturbation of the immune system during HIVinfection and potentially reveal new ways to modulate inflam-mation and disease.
In this study, we show that human Treg cells develop in vitrofrom a naive precursor subset, and that FoxP3 expression is re-quired for maintenance of their suppressive function. In addition,our findings indicate that the precursor population of Treg cells,which we termed naive Treg cells (TNreg cells), are highly suscep-tible to HIV infection. These findings have important implicationsfor understanding human Treg cell development and for developingtherapeutic strategies to treat conditions associated with chronicimmune activation such as HIV infection, or to enhance the effec-tiveness of vaccination.
Materials and MethodsCell isolation and culture
The blood obtained from healthy donors for this study have been reviewedand approved by Vanderbilt and New York University School of MedicineInstitutional Review Board committees. PBMC were isolated from bloodof healthy donors through Ficoll-Hypaque (Pharmacia). Resting CD4� Tcells were purified using CD4� Dynabeads (Dynal Biotech) as previouslydescribed (38) and were at least 99.5% pure as determined by postpurifi-cation FACS analysis. To purify naive, memory, Treg, and TNreg subsets,purified CD4� cells were stained with CD25 and CD45RO Abs andCD45RO�CD25� (naive CD4� T cells (TN)), CD45RO�CD25�/low (mem-ory CD4� T cells (TM)), CD45RO�CD25high (Treg), and CD45RO�CD25�
(TNreg) subsets were sorted on a FACS Aria flow cytometer. The culturemedium used in all experiments was RPMI 1640 (Invitrogen Life Tech-nologies) and was prepared as described previously (38). All cytokineswere purchased from R&D Systems. Monocyte-derived dendritic cells(DCs) were generated as previously described (38). Superantigen, staphy-lococcal enterotoxin B (SEB; Sigma-Aldrich) was used to stimulate restingT cells in the presence of DCs (39).
FACS analysis
T cells were stained with the relevant Ab on ice for 30 min (chemokinereceptor staining performed at room temperature for 20 min to enhancestaining) in PBS buffer containing 2% FCS and 0.1% sodium azide. Cellswere then washed twice, fixed with 1% paraformaldehyde, and analyzedwith a FACSCalibur or FACSAria flow cytometer. Live cells were gatedbased on forward and side scatter properties and analysis was performedusing FlowJo software (Tree Star). The following anti-human Abs wereused for staining: CD4, CD45RA, CD45RO, HLA-DR, CD25, GITR,CTLA-4, CD127, CCR5, CXCR4 (all from BD Biosciences), and CCR4(R&D Systems). TCR V region staining was done using the IOTest � MarkTCR V� Repertoire kit (Beckman Coulter) according to the accompanyingprotocol.
FoxP3 intracellular staining
PBMC or purified CD4� T cells were stained with appropriate cell surfacemarkers, as described above. Cells were washed and resuspended in 1�eBioscience fixation/permeabilization buffer, and incubated at 4°C for 30min. Cells were washed once with PBS buffer containing 2% FCS and0.1% sodium azide, and washed twice with 1� eBioscience permeabiliza-tion buffer. Cells were stained with Ab against FoxP3 (clone PCH110;eBioscience) for 30 min at 4°C. Cells were washed twice with 1� eBio-science permeabilization buffer, and analyzed with either FACSCalibur orLSR-II flow cytometers.
T cell proliferation and cytokine assay
To assess proliferation and quantify cell divisions, purified CD4� T cellsubsets were labeled with CFSE (Molecular Probes). Purified cells werefirst washed and resuspended in (PBS). While vortexing the cells, CFSEwas added at a final concentration of 5 �M. The mixture was vortexed foran additional 15 s and incubated at 37°C for 3 min. Labeling was quenchedby addition of 50% FCS in PBS. Cells were washed once more with 50%serum PBS, followed by two washes with RPMI 1640-supplemented me-dium. CFSE-labeled T cells were stimulated with either anti-CD3 (OKT-3;American Type Culture Collection) coated plates in the presence of soluble
anti-CD28 (1 �g/ml; BD Biosciences), or with either allogeneic or autol-ogous DCs in the presence of SEB (10-0.1 ng/ml) for 18 h. IL-2 in thesupernatants were assayed using a commercially available cytometric beadarray (CBA; BD Biosciences) (40) and analyzed using CBA six-bead anal-ysis software (BD Biosciences).
T cell suppression assay
Purified CD4� T cells were FACS sorted, as described above, based onexpression of CD25 and CD45RO. Target CD25�CD45RO� TN wereCFSE labeled. Effector, target, and mature allogeneic or autologous DCswere cultured in a round-bottom well of a 96-well plate, in the presence ofSEB (10 ng/ml-0.1 pg/ml). Cells were collected, fixed with 1% parafor-maldehyde, and analyzed on day 4 postactivation. Suppression experimentswith short hairpin RNA (shRNA)-expressing T cells were performed asabove, except target cells were labeled with the red dye SNARF-1 (Mo-lecular Probes) to discriminate from GFP-expressing (shRNA�) effector Tcells. Cells were incubated with 10 �M SNARF-1 and remaining protocolwas the same as CFSE labeling described above.
Real-time PCR
RNA was isolated from T cell subsets using an RNeasy kit (Qiagen). Tosynthesize cDNA, 100 ng of RNA was treated with Superscript II ReverseTranscriptase (Invitrogen Life Technologies). TaqMan Assays-on-DemandGene Expression Primers (Applied Biosystems) were used in real-timePCR analyses: �-actin primer mix assay ID Hs99999903_ml; FoxP3primer mix assay ID Hs00203958_m1. Real-time PCR was performed us-ing the ABI 7700 apparatus (Applied Biosystems). The reaction mixtures(20 ml total volume) contained 2 �l of serially diluted cDNA, 10 �l ofTaqMan Universal PCR Master Mix (Applied Biosystems), and 1 �l ofeither FoxP3 or �-actin primer mix. The reactions were amplified as fol-lows: 50°C for 2 min and 95°C for 10 min, followed by 40 cycles of 95°Cfor 1 min and 65°C for 1 min. Expression of FoxP3 mRNA was normalizedto �-actin expression in each sample.
HIV production and infections
HIV pseudotyped with vesicular stomatitis virus glycoprotein (VSV-G)envelope (VSV-G.HIV) was generated as previously described (41). Rep-lication competent CCR5-tropic HIV (R5.HIV) was prepared similarly bytransfecting 293T cells with HIV that encodes R5-tropic (BaL) envelopeand enhanced GFP (BD Clontech) in place of the nef gene as previouslydescribed (41). Typically, viral titers ranged from 1 to 5 � 106 infectiousunits (IFU)/ml for replication-competent viruses and 10–30 � 106 IFU/mlfor VSV-G.HIV. T cells were infected at varying multiplicities of infection(MOI), and infection was quantified by GFP expression using flow cytom-etry. Viral replication in T cell cultures was determined by measuring p24levels within supernatants by an ELISA (38).
shRNA construct design and virus production
shRNAs were designed using Invitrogen Life Technologies’s RNAi De-signer online program (https://rnaidesigner.invitrogen.com/rnaiexpress/setOption.do?designOption � shrna). FoxP3 (accession number NM_014009)was used as the target sequence to design the single-stranded oligonucle-otides against the open reading frame. A custom loop sequence (AGAGCTTG) was used. shRNA sequences used were the following: 5�-GTTATTCGCGCGAATAACGTTAGAGCTTGAACGTTATTCGCGCGAATAAC-3� (control shRNA); 5�-CACCGGACCATCTTCTGGATGAGAAAGAGCTTGTTCTCATCCAGAAGATGGTCC-3� (FoxP3 sh1), and 5�-CACCGCATGTTTGCCTTCTTCAGAAAGAGCTTGTTCCTGAAGAAGGCAAACATGC-3� (FoxP3 sh2).
Double-stranded oligos were generated and cloned into pENTR/U6 us-ing the Block-iT U6 RNAi Entry vector kit (Invitrogen Life Technologies).Ligation products were transformed into One Shot TOP10 competent Esch-erichia coli. Minipreps were performed on selected colonies and shRNAfidelity was verified by nucleotide sequencing. Gateway LR clonase reac-tion (Invitrogen Life Technologies) was performed to transfer the pENTR/U6-FoxP3 shRNA constructs into the GFP-expressing lentiviral vector sys-tem (destination vector) according to the protocol that accompanies thepENTR/U6 cloning kit. Lentiviruses encoding shRNAs were generated bycotransfecting 293T cells with the lentiviral vector along with plasmidsencoding VSV-G and Gag/Pol as described (38). Viral supernatants wereconcentrated using Amicon Ultra Centrifugal Filter Devices 100 K (Mil-lipore). Typically, concentrated viral titers ranged from 5 to 30 � 106
IFU/ml.
765The Journal of Immunology
by guest on July 25, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
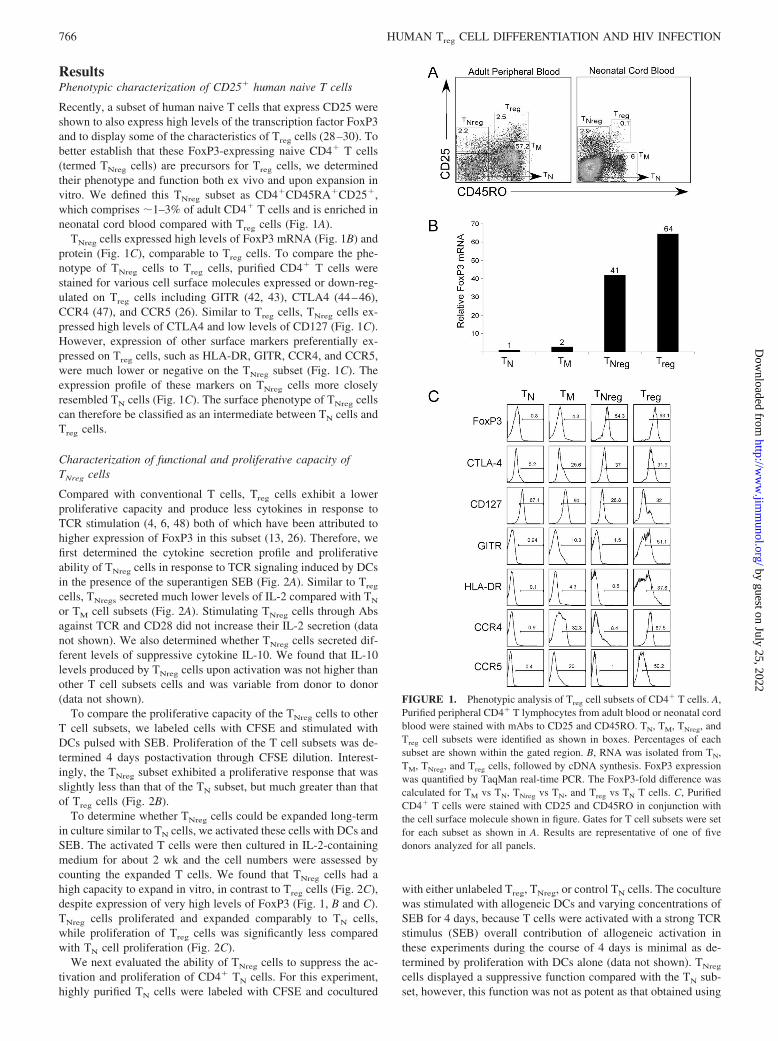
ResultsPhenotypic characterization of CD25� human naive T cells
Recently, a subset of human naive T cells that express CD25 wereshown to also express high levels of the transcription factor FoxP3and to display some of the characteristics of Treg cells (28–30). Tobetter establish that these FoxP3-expressing naive CD4� T cells(termed TNreg cells) are precursors for Treg cells, we determinedtheir phenotype and function both ex vivo and upon expansion invitro. We defined this TNreg subset as CD4�CD45RA�CD25�,which comprises �1–3% of adult CD4� T cells and is enriched inneonatal cord blood compared with Treg cells (Fig. 1A).
TNreg cells expressed high levels of FoxP3 mRNA (Fig. 1B) andprotein (Fig. 1C), comparable to Treg cells. To compare the phe-notype of TNreg cells to Treg cells, purified CD4� T cells werestained for various cell surface molecules expressed or down-reg-ulated on Treg cells including GITR (42, 43), CTLA4 (44–46),CCR4 (47), and CCR5 (26). Similar to Treg cells, TNreg cells ex-pressed high levels of CTLA4 and low levels of CD127 (Fig. 1C).However, expression of other surface markers preferentially ex-pressed on Treg cells, such as HLA-DR, GITR, CCR4, and CCR5,were much lower or negative on the TNreg subset (Fig. 1C). Theexpression profile of these markers on TNreg cells more closelyresembled TN cells (Fig. 1C). The surface phenotype of TNreg cellscan therefore be classified as an intermediate between TN cells andTreg cells.
Characterization of functional and proliferative capacity ofTNreg cells
Compared with conventional T cells, Treg cells exhibit a lowerproliferative capacity and produce less cytokines in response toTCR stimulation (4, 6, 48) both of which have been attributed tohigher expression of FoxP3 in this subset (13, 26). Therefore, wefirst determined the cytokine secretion profile and proliferativeability of TNreg cells in response to TCR signaling induced by DCsin the presence of the superantigen SEB (Fig. 2A). Similar to Treg
cells, TNregs secreted much lower levels of IL-2 compared with TN
or TM cell subsets (Fig. 2A). Stimulating TNreg cells through Absagainst TCR and CD28 did not increase their IL-2 secretion (datanot shown). We also determined whether TNreg cells secreted dif-ferent levels of suppressive cytokine IL-10. We found that IL-10levels produced by TNreg cells upon activation was not higher thanother T cell subsets cells and was variable from donor to donor(data not shown).
To compare the proliferative capacity of the TNreg cells to otherT cell subsets, we labeled cells with CFSE and stimulated withDCs pulsed with SEB. Proliferation of the T cell subsets was de-termined 4 days postactivation through CFSE dilution. Interest-ingly, the TNreg subset exhibited a proliferative response that wasslightly less than that of the TN subset, but much greater than thatof Treg cells (Fig. 2B).
To determine whether TNreg cells could be expanded long-termin culture similar to TN cells, we activated these cells with DCs andSEB. The activated T cells were then cultured in IL-2-containingmedium for about 2 wk and the cell numbers were assessed bycounting the expanded T cells. We found that TNreg cells had ahigh capacity to expand in vitro, in contrast to Treg cells (Fig. 2C),despite expression of very high levels of FoxP3 (Fig. 1, B and C).TNreg cells proliferated and expanded comparably to TN cells,while proliferation of Treg cells was significantly less comparedwith TN cell proliferation (Fig. 2C).
We next evaluated the ability of TNreg cells to suppress the ac-tivation and proliferation of CD4� TN cells. For this experiment,highly purified TN cells were labeled with CFSE and cocultured
with either unlabeled Treg, TNreg, or control TN cells. The coculturewas stimulated with allogeneic DCs and varying concentrations ofSEB for 4 days, because T cells were activated with a strong TCRstimulus (SEB) overall contribution of allogeneic activation inthese experiments during the course of 4 days is minimal as de-termined by proliferation with DCs alone (data not shown). TNreg
cells displayed a suppressive function compared with the TN sub-set, however, this function was not as potent as that obtained using
FIGURE 1. Phenotypic analysis of Treg cell subsets of CD4� T cells. A,Purified peripheral CD4� T lymphocytes from adult blood or neonatal cordblood were stained with mAbs to CD25 and CD45RO. TN, TM, TNreg, andTreg cell subsets were identified as shown in boxes. Percentages of eachsubset are shown within the gated region. B, RNA was isolated from TN,TM, TNreg, and Treg cells, followed by cDNA synthesis. FoxP3 expressionwas quantified by TaqMan real-time PCR. The FoxP3-fold difference wascalculated for TM vs TN, TNreg vs TN, and Treg vs TN T cells. C, PurifiedCD4� T cells were stained with CD25 and CD45RO in conjunction withthe cell surface molecule shown in figure. Gates for T cell subsets were setfor each subset as shown in A. Results are representative of one of fivedonors analyzed for all panels.
766 HUMAN Treg CELL DIFFERENTIATION AND HIV INFECTION
by guest on July 25, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

Treg cells (Fig. 2D). Efficiency of suppression by resting TNreg cellsranged from �50% to �10% suppression of target cells (data notshown), though Treg cells consistently suppressed 70–90% targetcells (data not shown). To confirm that cell death in the TNreg andTreg conditions was not playing a role in the lack of target cellproliferation, we assessed cell death by percent of live cells basedon forward vs side scatter plots. Approximately 85–90% of cellswere within the live cell gate in all conditions (data not shown).Thus, we conclude that lack of proliferation is not due to excessivecell death. Together, these results show that TNreg cells possess anintermediate functional profile as compared with Treg cells and,importantly, that they have the capacity to expand upon Agstimulation.
Phenotypic characterization of expanded TNreg cells
Because TNreg cells can be expanded �100-fold in IL-2 after TCRstimulation (Fig. 2C), we sought to address whether their surfacephenotype and suppressive capacity is altered by in vitro expan-sion. To ensure that TNreg cells maintained expression of FoxP3,and to exclude the possibility that the expanded cell populationwas contaminated with conventional T cell subsets, we performedsurface phenotype analysis in conjunction with intracellular stain-ing of FoxP3. Almost all of the expanded TNreg cells expressedhigh levels of FoxP3, whereas expanded TN cells remained low ornegative for FoxP3 expression (Fig. 3A).
We next analyzed the Treg cell-associated surface markers onexpanded TNreg cultures 2 wk postactivation. Expanded TNreg cellsexpressed higher levels of CD25, GITR, HLA-DR, CCR4, andCTLA-4 as compared with expanded TN cells (Fig. 3B). Similar toTN cells, TNreg cells also lose the expression of CD45RA andexpress CD45RO after activation and expansion in vitro (data notshown). This differentiated cell surface phenotype of TNreg cells isconsistent with a Treg cell profile, ex vivo (Fig. 1C). In addition,expanded TNreg cells expressed high levels of the HIV coreceptorsCCR5 and CXCR4 (Fig. 3C), similar to Treg cells (26) suggestingthat these cells could be potentially susceptible to HIV infection.
Functional characterization of expanded TNreg cells
We asked whether activation and expansion of TNreg cells resultedin full acquisition of Treg cell functions. We first determined theresponsiveness of expanded TNreg cells to TCR restimulation bymeasuring IL-2 secretion. Expanded TNreg cells stimulated withDCs and SEB continued to secrete low levels of IL-2 comparedwith the TN subset (Fig. 4A), indicating that they maintain this Treg
cell characteristic even after expansion in vitro.We then evaluated whether expanded TNreg cells acquired more
potent suppressive function compared with freshly isolated cells.We found that expanded TNreg cells had displayed more potentimmunosuppressive activity (Fig. 4B) than freshly purified TNreg
counterparts (Fig. 2D). At suboptimal TCR stimulation(0.01–0.001 ng/ml SEB) the suppression by expanded TNreg cellswas similar (�80–90% inhibition of target cell proliferation) toTreg cells (Fig. 4B). Taken together, these findings suggest thatactivation and expansion of TNreg cells induces a program leadingto their differentiation into fully competent Treg cells.
TCR V� repertoire of TNreg and Treg cells
Our data support the hypothesis that, upon their activation andexpansion, TNreg cells may contribute to the pool of Treg cells upon
FIGURE 2. Functional analysis of Treg subsets. A, Sorted Treg subsetswere activated in the presence of DCs pulsed with SEB (1–10 ng/ml).Supernatants were collected 18–24 h postactivation and analyzed for cy-tokines using the CBA assay. Results from three different donors areshown. Statistical significance was determined using the Student t test com-paring TN and TNreg conditions across donors. B, Resting, sorted Treg sub-sets were labeled with CFSE and activated with SEB (1–10 ng/ml) pulsedDCs. At day 4 or 5 postactivation, cells were fixed and analyzed for CFSEexpression by flow cytometry. Results represent one of five donors ana-lyzed. C, Sorted Treg subsets were activated with DCs � SEB (1–10 ng/ml)and expanded in the presence of IL-2. Cells were counted on day 12 andfold expansion was calculated for each subset. Two representative donorsare shown in the figure. Statistical significance was determined using theStudent t test comparing TN and TNreg cells (not significantly different) andTN and Treg cells across donors. D, Resting naive cells were labeled withCFSE and cocultured with unlabeled TNreg, Treg, or TN cells, and DCs inthe presence of SEB at a 1:1:30 ratio. Proliferation of target cells wasassessed by gating on live cells and analysis of forward scatter (cell size)
vs CFSE labeling. Number on top of rectangle boxes on right side of eachplot indicates the percentage of target TN cells that have divided based onCFSE expression. Results represent one of five donors analyzed. �, p �0.05.
767The Journal of Immunology
by guest on July 25, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

activation and expansion. As a corollary to this, we hypothesizedthat the TCR repertoire of TNreg cells would be similar to that ofTreg cells. To investigate the breadth of the TCR V� repertoire ofthe naive and memory T cell subsets, we stained purified CD4� Tcells with CD25 and CD45RO as subset markers, and costainedwith Abs directed against �80% of the mature human TCR V�repertoire of T cells (49). This analysis indicated that TCR V�repertoire of TNreg cells was very similar to that of other T cellsubsets (Fig. 5). Furthermore, we noted that the relative expressionof different V� families was very similar between Treg and TNreg
subsets (Fig. 5). This finding supports a model whereby TNreg dif-ferentiate into Treg cells upon activation with self or nonself Ags invivo, acquiring a suppressive capability while maintaining the ca-pacity to recognize similar Ags as conventional T cell subsets.
Is FoxP3 required for maintaining TNreg cell phenotype andfunction?
To determine whether sustained expression of FoxP3 is required tomaintain the capability of TNreg cells to differentiate into fullysuppressive Treg cells, we reduced expression of FoxP3 in thesesubsets by RNA interference (RNAi). Accordingly, shRNAs tar-geting the FoxP3 gene were stably expressed in activated TN orTNreg cells using a lentiviral delivery system in which the coex-pression of GFP allows for the identification of transduced cells(50).
Using this system, we specifically and stably reduced expressionof FoxP3, as shown by intracellular staining with a FoxP3 Ab (Fig.
FIGURE 3. Phenotypic analysis of Treg subsets postactivation and ex-pansion. TN and TNreg cells were sorted and expanded in the presence ofIL-2 for 10–14 days. Post expansion, the subsets were stained intracellu-larly for expression of FoxP3 (A), CD25, CTLA4, HLA-DR, CCR4, andGITR (B). One representative experiment of five different donors analyzedis shown. C, Expanded TN and TNreg cells were additionally stained forexpression of CCR5 and CXCR4. Numbers in upper right corner of allplots are gated percentage.
FIGURE 4. Functional characterization of expanded TNreg cells. A,Sorted Treg subsets were activated using DCs � SEB (1–10 ng/ml) andexpanded in IL-2-containing medium for 10–14 days. Expanded cells werewashed to remove IL-2 and restimulated using DCs and SEB (1-0.1 ng/ml)and supernatants were collected 18–24 h postactivation. IL-2 productionwas measured using a CBA assay. Shown are three representative donors.Statistical significance was determined using the Student t test across do-nors. �, p � 0.05. B, A suppression assay was performed as described inFig. 2 using expanded TNreg cells and allogeneic TN cells. Target cells werefreshly isolated resting TN. After gating on live cells, forward scatter (cellsize) vs CFSE labeling is shown for three concentration range of SEBstimulation via DCs. Number on top of boxes on right side of each plotindicates the percentage of target TN cells that have divided in the presenceof TN or TNreg cells based on lower CFSE expression. Shown is one rep-resentative experiment of three experiments.
768 HUMAN Treg CELL DIFFERENTIATION AND HIV INFECTION
by guest on July 25, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

6A). We next assessed the surface phenotype of the expandedTNreg cells in which FoxP3 expression have been reduced byRNAi. Knockdown of FoxP3 was associated with lower expres-sion of CD25, CD62L, CTLA4, CCR4, and HLA-DR as comparedwith TNreg cells transduced with control shRNA vector (Fig. 6A).Similar results were obtained by stably expressing a second dis-tinct shRNA against FoxP3 in TNreg cells (Fig. 6A).
We next evaluated the effects of FoxP3 knockdown in activatedTNreg cells on their cytokine production and suppressive function.For this experiment, we first sorted the shRNA transduced ex-panded TNreg cells, restimulated them through the TCR, and mea-sured secreted IL-2. TNreg cells with reduced FoxP3 were un-changed in their inability to produce IL-2 compared with controlTNreg cells expressing high levels of FoxP3 (Fig. 6B). However,reduction of FoxP3 levels in TNreg cells greatly diminished theirability to suppress the proliferation of conventional TN cells (Fig.6C). These results indicate that a high-level expression of FoxP3 isindispensable for suppressive activity of Treg cells.
Are TNreg cells susceptible to HIV infection?
Treg cells were shown to potentially play an important role in mod-ulating the immune response to HIV and pathogenesis associatedwith HIV infection (26, 33). We have shown that TNreg cells arecapable of significant proliferation upon TCR stimulation and ex-press high levels of HIV coreceptors CCR5 and CXCR4. Wetherefore studied the susceptibility of TNreg cells to HIV infectionin vitro. T cell subsets were purified and activated with DCs andSEB and at the time of activation, these cells were also infectedwith either replication-defective viruses pseudotyped with VSV-G.HIV or replication-competent HIV viruses, that use CCR5(R5.HIV) for entry, both of which also express GFP as a marker ofinfection (38). TNreg cells were highly infected by VSV-G.HIVand were also more susceptible to infection with R5.HIV com-pared with TN cells (Fig. 7A). To determine the level of HIV rep-lication in the TNreg cells as compared with activated TN cells,subsets were infected with R5.HIV for 3 days and washed to re-move input virus. Supernatants from infected cultures were thenmonitored for HIV replication by measuring the GFP expressionfrom infected T cells and the level of HIV p24 Ag levels using anELISA, as described (26). R5.HIV spread within TNreg cell cul-tures over a 2-wk period and replicated at levels comparable tomemory T cells, and importantly higher than TN cells during theearly stages of the infection (Fig. 7, B and C).
FIGURE 5. V� repertoire of Treg subsets from peripheral blood. CD4�
T cells were stained with mAbs to CD25 and CD45RO, and gates were setfor TN, TM, TNreg, and Treg cells. Cells were costained with the Abs againstthe V� family indicated in the figure. Percentage shown is percent of V��
cells in the given population. This is a representative profile from threeseparate donors.
FIGURE 6. shRNA mediated knockdown of FoxP3 in TNreg cells. SortedTN or TNreg cells were activated with DC and SEB (10 ng/ml), and concur-rently infected with shRNA expressing virus at an MOI of 3–5. Cells wereexpanded in IL-2-containing medium for 10–14 days. A, TNreg cells werestained for FoxP3, or cell surface marker shown. Uninfected, a control non-mammalian shRNA (control sh), and two FoxP3-specific shRNAs (FoxP3 sh1and FoxP3 sh2) transduced cells are shown. Transduced cells were identifiedby expression of GFP. Cells were gated on GFP-positive cells and furtheranalyzed for the indicated surface molecule. B, TN or TNreg cells transducedwith shRNA were sorted based on GFP expression, and restimulated with DCplus SEB (0.01 ng/ml), supernatants were collected 18–24 h postactivation.IL-2 production was measured using a CBA assay. C, TN or TNreg cells trans-duced with shRNA were sorted based on GFP expression, and a suppressionassay was performed as described in Fig. 4. Target naive T cells were labeledwith a red dye (SNARF). Live cells were gated and percentage of target cellshaving undergone at least one cell division are shown. This experiment isrepresentative of three experiments for all panels.
769The Journal of Immunology
by guest on July 25, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

Although neither resting TN nor TNreg cells expressed detectablelevels of CCR5 by Ab staining (Fig. 2C) or mRNA by quantitativereal-time PCR (data not shown), we hypothesized that TNreg cellsmore rapidly up-regulate CCR5 upon activation compared with TN
cells. To test this hypothesis, we sorted TN and TNreg cell popu-lations and stained each subset for the expression of CCR5 on days5 and 7 postactivation. We found that TNreg cells contained �8- to10-fold higher CCR5� T cells at these earlier time points (Fig.7D). This result was corroborated by �5-fold higher CCR5 mRNAexpression, by real-time PCR analysis upon activation, on TNreg
cells compared with TN cells (data not shown). CCR5 expressionwas detectable on activated TN cells by surface staining 8–10 dayspostactivation (Fig. 3C), thus lagging several days compared withactivated TNreg cells.
DiscussionIn this study, we demonstrate that a population of naive T cellscoexpressing CD25 and FoxP3, which we called TNreg cells, hasthe capacity to differentiate into fully functional Treg cells in vitro.In contrast to Treg cells, TNreg cells exhibit a high proliferativecapacity upon TCR stimulation, despite expressing high levels ofFoxP3. In our culture system, the proliferation of TNreg cells washigher compared with a prior report (28), which could be due toour activation conditions using DCs pulsed with SEB that we useto provide a more physiological cell-cell interaction. Severalgroups have reported expanding CD25� Treg cells using a multi-tude of stimulation conditions, including CD28-coated beads (51–53). Upon expansion, these cells retain their high expression ofFoxP3. However, in these studies, the CD25� T cells were notsubdivided into naive and memory subsets. Because we have notbeen able to expand mature Treg cells in vitro to any significantnumbers, our present data raise the possibility that in vitro-ex-panded Treg cells obtained by others were derived from the TNreg
cell population. This would be consistent with a recent reportshowing that in vitro expansion of only the CD45RA� populationof CD25� T cells gives rise to a pure Treg cell line (54).
Our results indicate that the TNreg cell population does not adopta full Treg phenotype or exert its function until it has undergoneactivation and expansion and reactivation through the TCR. Thisobservation agrees with FoxP3 overexpression studies performedby our group (26) and others (13, 55, 56) to gain insights into itspossible functions in naive T cells. Mere overexpression of FoxP3in naive CD4� T cells does not render them fully suppressive andhyporesponsive until FoxP3 overexpressing cells have been re-stimulated through their TCR (26). This leads to the intriguinghypothesis that the high expression of FoxP3 is not sufficient forTreg suppressive activity in the absence of other cell differentiationprograms, some of which are possibly activated by FoxP3 afterTCR stimulation.
In our experiments, we found that TNreg cells were consistentlyless suppressive relative to Treg cells with memory phenotype, butacquired equally potent suppressive activity after they are ex-panded in vitro. Prior publications (28, 29, 57), found that cellssimilar to TNreg cells were more suppressive, quantitatively com-pared with our findings. It is conceivable that these quantitativedifferences are due to the type of stimulus used in these previousstudies. Indeed, we have tried anti-CD3/28 stimulations, similar topublished reports, but the suppressions from these assays were not
FIGURE 7. TNreg cells are susceptible to HIV infection and can serve assites of replication. Sorted T cell populations were activated using DCs and10 ng/ml SEB, and concurrently infected with R5.HIV viruses expressingGFP at a range of 1–5 MOI. A, The percentage of infected cells based onGFP expression for VSV-G.HIV or R5.HIV day 6 postinfection. B, Rep-lication of R5.HIV as measured by GFP expression in TN, TM, or TNreg
sorted populations. GFP expression was assessed every other day startingon day 4 postinfection. Data shown are infection with 1 MOI. C, Super-natants from T cell populations infected with R5.HIV cultures were col-lected at different time points and the level of HIV p24 was measured byELISA. Data shown are infection with 1 MOI. Similar results were ob-tained from three different donors. Error bars represent SD of duplicatesamples within the same experiment. Statistical significance was determined
using the Student t test comparing TN and TNreg conditions. �, p � 0.05. D,TN and TNreg cells activated for 5 days under same conditions as abovewere stained with a CCR5 Ab. Data are represented by side scatter profileand CCR5 staining and percent positive cells are shown on top of boxes.
770 HUMAN Treg CELL DIFFERENTIATION AND HIV INFECTION
by guest on July 25, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

as consistent. This is not surprising because Treg cells require cell-to-cell contact and triggering only the TCR may have the caveat ofnot forming an immunological synapse with an APC and recruitingTreg cells to the site of activation. We believe our method to stim-ulate Treg cells and T cells through DCs and SEB is more physi-ological way to activate them which allows three cell (DC plus Tcell plus TNreg or Treg) clusters to form, and could be a betterreflection of the Treg-mediated suppression in vivo.
In this study, we demonstrated for the first time that reduction ofFoxP3 expression in human Treg cell precursors results in an al-tered phenotype and is required to maintain their suppressive func-tion. Similar findings were reported for murine Treg cells, whichshowed that induced ablation of FoxP3 disrupted Treg suppressivefunctions in vivo (22). In addition, it was demonstrated that atten-uated FoxP3 expression subverted Treg cell functions in vivo (23).The suppressive functions of FoxP3-attenuated mice were abro-gated, but the cells retained their hypoproliferative nature (23). Weshow that cells with reduced expression of FoxP3 are still hypo-responsive to TCR stimulation as measured by the production ofIL-2. Williams et al. (22) showed a rescue of IL-2 production uponcontrolled deletion of FoxP3 in vivo. In our shRNA system, FoxP3expression in Treg cells is greatly reduced but not completely ab-lated, leaving open the possibility that residual FoxP3 levels maybe sufficient to render T cells hyporesponsive to TCR signals, butinsufficient to maintain their suppressive function. It is also pos-sible that IL-2 secretion in Treg cells is independent of FoxP3 ex-pression, but rather due to epigenetic modifications that regulategene expression in T cells, such as in the histone tails.
The nature of the Ag recognized by Treg cells remains contro-versial. Studies conducted using transgenic mouse models suggestthat once activated through the TCR, suppressor effector functionwas completely Ag independent (58, 59). These studies, however,did not exclude the possibility that Treg cells exert their suppres-sive function in an Ag-specific manner. We show here that bothTNreg cells and Treg cells express a full repertoire of TCR V�receptors. It was shown that Ag-mediated expansions within spe-cific TCR V� families were associated with equal increases of thesame TCR V� in the Treg cell population (60). Therefore, it ispossible that Treg cells are replenished from a precursor populationin the periphery. The precursor population is likely to be the TNreg
subset as suggested by our study and others (28, 29, 54, 61). Ac-cumulating evidence in animal models indicates that Treg cells areselected by self Ag, (25, 62, 63), although it is not clear whethera naive precursor subset for Treg cells described in the presentstudy also exists in mice. It appears that, unlike the mouse system,human FoxP3 expression can be induced in human CD4�CD25�
T cells following TCR stimulation (64, 65). We propose that, atleast in the human system, acquisition of suppressive function bythe TNreg subset is similar to that of the acquisition of effectorfunctions of conventional TN cells. This suggests that during animmune response, TN cells undergo activation and differentiationto gain effector functions, and TNreg cells undergo a parallel dif-ferentiation process leading to clonal expansion. This scenariowould predict that for exogenous Ags, Treg suppression would beweaker for primary immune responses and stronger after second-ary or subsequent exposures, thereby functioning to controlchronic immune activation.
The role of the Treg compartment of T cells during HIV infec-tion and pathogenesis remains a very important question and is notyet resolved. We clearly demonstrate an enhanced ability of HIVto replicate and infect TNreg cells. Our findings also show a 10-foldhigher HIV infection rate of TNreg cells compared with TN cellswith R5-tropic viruses, which use CCR5 coreceptor for entry, dur-ing the early stages of infection (Fig. 7). We found that TNreg cells
up-regulated CCR5 upon activation several days before up-regu-lation of CCR5 in TN cells (Fig. 7D). We believe this finding couldpartly explain the difference in infection in the early days afteractivation. This finding could be important during HIV infection,because during an immune response the preferential infection ofTNreg cells early after activation could contribute to lesser controlof immune activation, and thus more cellular targets for HIV. Thisdata also suggests that CCR5 expression is differentially regulatedin TN and TNreg cells during the early stages of T cell activation.This is also consistent with the our results that HIV spreads tolarger portion of activated TN cells after day 8 (Fig. 7B), at whichtime they begin to up-regulate CCR5 expression. It has been sug-gested that the expression of FoxP3 enhances HIV long-terminalrepeat activity (21), it is therefore possible that TNreg cells, thatexpress high levels of FoxP3, also provide a more optimal intra-cellular environment for HIV gene expression, similar to cells thatoverexpress FoxP3 during early stages of infection (21). A con-tradictory report demonstrated that FoxP3 represses retroviral tran-scription (66). Differences in these two studies may be due to themodel systems to used to test HIV long-terminal repeat activity,such as transient transfection assay (66) vs a luciferase reporterHIV infection assay (21). Thus, the effect of FoxP3 on HIV in-fection remains to be further clarified in future studies.
The effect of Treg disruption during HIV infection has been as-sociated with both beneficial (26, 31, 36) and detrimental out-comes (26, 32–35). It has been suggested that HIV-immune dys-function is associated with Treg cells that may target HIV-specificeffectors, leading to an inefficient immune response against HIV(32–35, 37). We and others have proposed that Treg cells mayserve a protective function by limiting the overall immune activa-tion that is a hallmark of HIV infection (26, 31, 36). In this context,the depletion of Treg cells could potentially hasten the progressionto acquired immunodeficiency syndrome. At the same time, thespecific depletion of TNreg cells may prevent insufficient regener-ation of the Treg cell pools during primary immune responses, andcould be one of the reasons for immune hyperactivation associatedwith HIV infection. This is supported by data showing HIV-pos-itive individuals with higher relative levels of FoxP3 have lowerimmune activation as measured by expression of HLA-DR (26,31). We demonstrate that when compared with TN cells, TNreg
cells are highly susceptible to infection by R5.HIV during earlystages post-TCR activation. This result suggests that TNreg cellscould be preferentially targeted during HIV infection. How TNreg
cells are modulated in vivo during different stages of HIV infectionremains to be determined. Furthermore, it will be important todetermine whether a similar subset of TNreg cells can be identifiedin monkey models of SIV infection and whether this can be cor-related with the presence or absence of immune activation in thesemodels, which highly correlate with disease progression (67). In-terestingly, it has been reported that administration of IL-2 to HIV-positive individuals leads to the expansion of TNreg subset (30).Thus, enhancing this subset by IL-2 treatment could be addition-ally beneficial in regulating T cell homeostasis during HIV infec-tion by dampening chronic immune activation.
In summary, our results indicate that the TNreg cell populationacquires a full Treg phenotype upon activation, and that this dif-ferentiation process is dependent upon sustained, high-level FoxP3expression. This subset also appears to be more susceptible to HIVinfection, when compared with TN cells. The TNreg cell populationcould provide an exciting target cell for potential therapeutics. Theability to manipulate this population of cells could allow for thegeneration of homogenous, Ag-specific Treg cells that could beused for the prevention and treatment of T cell-mediated diseases,
771The Journal of Immunology
by guest on July 25, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

including graft-vs-host disease or reducing chronic immune acti-vation during HIV infection. It has been shown that the memorysubset of Treg cells is highly susceptible to apoptosis and has lim-ited replicative capacity in vivo (60, 68), suggesting that the mod-ulation of Treg cells for therapeutic purposes may not be long-lasting. The therapeutic manipulation of TNreg cells may provide amore tractable way to obtain specific and long-term protectionagainst aberrant T cell activation, and permit effective immunemodulation for vaccine responses.
AcknowledgmentsWe thank Dr. Luc Van Kaer, Dr. Scott VanCompernolle, Brenna Simons,Micheal Vetter, Frances Mercer, and Dr. Simge Akbulut for critical readingand valuable suggestions. We thank Dr. Chris Aiken for p24 ELISA re-agents and Aimee El Hed for help with CCR5 staining.
DisclosuresThe authors have no financial conflict of interest.
References1. Hogquist, K. A., T. A. Baldwin, and S. C. Jameson. 2005. Central tolerance:
learning self-control in the thymus. Nat. Rev. Immunol. 5: 772–782.2. Sakaguchi, S., T. Takahashi, S. Yamazaki, Y. Kuniyasu, M. Itoh, N. Sakaguchi,
and J. Shimizu. 2001. Immunologic self tolerance maintained by T-cell-mediatedcontrol of self-reactive T cells: implications for autoimmunity and tumor immu-nity. Microbes Infect. 3: 911–918.
3. Hori, S., T. Takahashi, and S. Sakaguchi. 2003. Control of autoimmunity bynaturally arising regulatory CD4� T cells. Adv. Immunol. 81: 331–371.
4. Jonuleit, H., E. Schmitt, M. Stassen, A. Tuettenberg, J. Knop, and A. H. Enk.2001. Identification and functional characterization of human CD4�CD25� Tcells with regulatory properties isolated from peripheral blood. J. Exp. Med. 193:1285–1294.
5. Baecher-Allan, C., J. A. Brown, G. J. Freeman, and D. A. Hafler. 2001.CD4�CD25high regulatory cells in human peripheral blood. J. Immunol. 167:1245–1253.
6. Ng, W. F., P. J. Duggan, F. Ponchel, G. Matarese, G. Lombardi, A. D. Edwards,J. D. Isaacs, and R. I. Lechler. 2001. Human CD4�CD25� cells: a naturallyoccurring population of regulatory T cells. Blood 98: 2736–2744.
7. Stephens, L. A., C. Mottet, D. Mason, and F. Powrie. 2001. Human CD4�CD25�
thymocytes and peripheral T cells have immune suppressive activity in vitro. Eur.J. Immunol. 31: 1247–1254.
8. Piccirillo, C. A., J. J. Letterio, A. M. Thornton, R. S. McHugh, M. Mamura,H. Mizuhara, and E. M. Shevach. 2002. CD4�CD25� regulatory T cells canmediate suppressor function in the absence of transforming growth factor �1production and responsiveness. J. Exp. Med. 196: 237–246.
9. Shevach, E. M. 2001. Certified professionals: CD4�CD25� suppressor T cells.J. Exp. Med. 193: F41–F46.
10. Ziegler, S. F. 2006. FOXP3: of mice and men. Annu. Rev. Immunol. 24: 209–226.11. Brunkow, M. E., E. W. Jeffery, K. A. Hjerrild, B. Paeper, L. B. Clark,
S. A. Yasayko, J. E. Wilkinson, D. Galas, S. F. Ziegler, and F. Ramsdell. 2001.Disruption of a new forkhead/winged-helix protein, scurfin, results in the fatallymphoproliferative disorder of the scurfy mouse. Nat. Genet. 27: 68–73.
12. Bennett, C. L., and H. D. Ochs. 2001. IPEX is a unique X-linked syndromecharacterized by immune dysfunction, polyendocrinopathy, enteropathy, and avariety of autoimmune phenomena. Curr. Opin. Pediatr. 13: 533–538.
13. Fontenot, J. D., M. A. Gavin, and A. Y. Rudensky. 2003. Foxp3 programs thedevelopment and function of CD4�CD25� regulatory T cells. Nat. Immunol. 4:330–336.
14. Hori, S., and S. Sakaguchi. 2004. Foxp3: a critical regulator of the developmentand function of regulatory T cells. Microbes Infect. 6: 745–751.
15. Bennett, C. L., J. Christie, F. Ramsdell, M. E. Brunkow, P. J. Ferguson,L. Whitesell, T. E. Kelly, F. T. Saulsbury, P. F. Chance, and H. D. Ochs. 2001.The immune dysregulation, polyendocrinopathy, enteropathy, X-linked syn-drome (IPEX) is caused by mutations of FOXP3. Nat. Genet. 27: 20–21.
16. Bettelli, E., M. Dastrange, and M. Oukka. 2005. Foxp3 interacts with nuclearfactor of activated T cells and NF-�B to repress cytokine gene expression andeffector functions of T helper cells. Proc. Natl. Acad. Sci. USA 102: 5138–5143.
17. Wu, Y., M. Borde, V. Heissmeyer, M. Feuerer, A. D. Lapan, J. C. Stroud,D. L. Bates, L. Guo, A. Han, S. F. Ziegler, et al. 2006. FOXP3 controls regulatoryT cell function through cooperation with NFAT. Cell 126: 375–387.
18. Liu, W., A. L. Putnam, Z. Xu-Yu, G. L. Szot, M. R. Lee, S. Zhu, P. A. Gottlieb,P. Kapranov, T. R. Gingeras, B. Fazekas, de St. Groth, et al. 2006. CD127 ex-pression inversely correlates with FoxP3 and suppressive function of humanCD4� T reg cells. J. Exp. Med. 203: 1701–1711.
19. Marson, A., K. Kretschmer, G. M. Frampton, E. S. Jacobsen, J. K. Polansky,K. D. MacIsaac, S. S. Levine, E. Fraenkel, H. von Boehmer, and R. A. Young.2007. Foxp3 occupancy and regulation of key target genes during T-cell stimu-lation. Nature 445: 931–935.
20. Zheng, Y., S. Z. Josefowicz, A. Kas, T. T. Chu, M. A. Gavin, andA. Y. Rudensky. 2007. Genome-wide analysis of Foxp3 target genes in devel-oping and mature regulatory T cells. Nature 445: 936–940.
21. Holmes, D., G. Knudsen, S. Mackey-Cushman, and L. Su. 2007. FoxP3 enhancesHIV-1 gene expression by modulating NF�B occupancy at the LTR in human Tcells. J. Biol. Chem. 282: 15973–15980.
22. Williams, L. M., and A. Y. Rudensky. 2007. Maintenance of the Foxp3-depen-dent developmental program in mature regulatory T cells requires continued ex-pression of Foxp3. Nat. Immunol. 8: 277–284.
23. Wan, Y. Y., and R. A. Flavell. 2007. Regulatory T-cell functions are subvertedand converted owing to attenuated Foxp3 expression. Nature 445: 766–770.
24. Shevach, E. M. 2002. CD4� CD25� suppressor T cells: more questions thananswers. Nat. Rev. Immunol. 2: 389–400.
25. Jordan, M. S., A. Boesteanu, A. J. Reed, A. L. Petrone, A. E. Holenbeck,M. A. Lerman, A. Naji, and A. J. Caton. 2001. Thymic selection of CD4�CD25�
regulatory T cells induced by an agonist self-peptide. Nat. Immunol. 2: 301–306.26. Oswald-Richter, K., S. M. Grill, N. Shariat, M. Leelawong, M. S. Sundrud,
D. W. Haas, and D. Unutmaz. 2004. HIV infection of naturally occurring andgenetically reprogrammed human regulatory T-cells. PLoS Biol. 2: E198.
27. Banham, A. H. 2006. Cell-surface IL-7 receptor expression facilitates the puri-fication of FOXP3� regulatory T cells. Trends Immunol. 27: 541–544.
28. Valmori, D., A. Merlo, N. E. Souleimanian, C. S. Hesdorffer, and M. Ayyoub.2005. A peripheral circulating compartment of natural naive CD4 Tregs. J. Clin.Invest. 115: 1953–1962.
29. Seddiki, N., B. Santner-Nanan, S. G. Tangye, S. I. Alexander, M. Solomon,S. Lee, R. Nanan, and B. F. de St. Groth. 2006. Persistence of naive CD45RA�
regulatory T cells in adult life. Blood 107: 2830–2838.30. Sereti, I., H. Imamichi, V. Natarajan, T. Imamichi, M. S. Ramchandani,
Y. Badralmaa, S. C. Berg, J. A. Metcalf, B. K. Hahn, J. M. Shen, et al. 2005. Invivo expansion of CD4CD45RO�CD25 T cells expressing foxP3 in IL-2-treatedHIV-infected patients. J. Clin. Invest. 115: 1839–1847.
31. Legrand, F. A., D. F. Nixon, C. P. Loo, E. Ono, J. M. Chapman, M. Miyamoto,R. S. Diaz, A. M. Santos, R. C. Succi, J. Abadi, et al. 2006. Strong HIV-1-specificT cell responses in HIV-1-exposed uninfected infants and neonates revealed afterregulatory T cell removal. PLoS ONE 1: e102.
32. Kinter, A., J. McNally, L. Riggin, R. Jackson, G. Roby, and A. S. Fauci. 2007.Suppression of HIV-specific T cell activity by lymph node CD25� regulatory Tcells from HIV-infected individuals. Proc. Natl. Acad. Sci. USA 104: 3390–3395.
33. Kinter, A. L., M. Hennessey, A. Bell, S. Kern, Y. Lin, M. Daucher, M. Planta,M. McGlaughlin, R. Jackson, S. F. Ziegler, and A. S. Fauci. 2004. CD25�CD4�
regulatory T cells from the peripheral blood of asymptomatic HIV-infected in-dividuals regulate CD4� and CD8� HIV-specific T cell immune responses invitro and are associated with favorable clinical markers of disease status. J. Exp.Med. 200: 331–343.
34. Kinter, A. L., R. Horak, M. Sion, L. Riggin, J. McNally, Y. Lin, R. Jackson, A.O’Shea, G. Roby, C. Kovacs, et al. 2007. CD25� regulatory T cells isolated fromHIV-infected individuals suppress the cytolytic and nonlytic antiviral activity ofHIV-specific CD8� T cells in vitro. AIDS Res. Hum. Retroviruses 23: 438–450.
35. Weiss, L., V. Donkova-Petrini, L. Caccavelli, M. Balbo, C. Carbonneil, andY. Levy. 2004. Human immunodeficiency virus-driven expansion ofCD4�CD25� regulatory T cells, which suppress HIV-specific CD4 T-cell re-sponses in HIV-infected patients. Blood 104: 3249–3256.
36. Eggena, M. P., B. Barugahare, N. Jones, M. Okello, S. Mutalya, C. Kityo,P. Mugyenyi, and H. Cao. 2005. Depletion of regulatory T cells in HIV infectionis associated with immune activation. J. Immunol. 174: 4407–4414.
37. Aandahl, E. M., J. Michaelsson, W. J. Moretto, F. M. Hecht, and D. F. Nixon.2004. Human CD4� CD25� regulatory T cells control T-cell responses to humanimmunodeficiency virus and cytomegalovirus antigens. J. Virol. 78: 2454–2459.
38. Motsinger, A., D. W. Haas, A. K. Stanic, L. Van Kaer, S. Joyce, and D. Unutmaz.2002. CD1d-restricted human natural killer T cells are highly susceptible to hu-man immunodeficiency virus 1 infection. J. Exp. Med. 195: 869–879.
39. Unutmaz, D., V. N. KewalRamani, S. Marmon, and D. R. Littman. 1999. Cyto-kine signals are sufficient for HIV-1 infection of resting human T lymphocytes.J. Exp. Med. 189: 1735–1746.
40. Cook, E. B., J. L. Stahl, L. Lowe, R. Chen, E. Morgan, J. Wilson, R. Varro,A. Chan, F. M. Graziano, and N. P. Barney. 2001. Simultaneous measurement ofsix cytokines in a single sample of human tears using microparticle-based flowcytometry: allergics vs. non-allergics. J. Immunol. Methods 254: 109–118.
41. Unutmaz, D., V. N. KewalRamani, S. Marmon, and D. R. Littman. 1999. Cyto-kine signals are sufficient for HIV-1 infection of resting human T lymphocytes.J. Exp. Med. 189: 1735–1746.
42. McHugh, R. S., M. J. Whitters, C. A. Piccirillo, D. A. Young, E. M. Shevach,M. Collins, and M. C. Byrne. 2002. CD4�CD25� immunoregulatory T cells:gene expression analysis reveals a functional role for the glucocorticoid-inducedTNF receptor. Immunity 16: 311–323.
43. Shimizu, J., S. Yamazaki, T. Takahashi, Y. Ishida, and S. Sakaguchi. 2002. Stim-ulation of CD25�CD4� regulatory T cells through GITR breaks immunologicalself-tolerance. Nat. Immunol. 3: 135–142.
44. Read, S., V. Malmstrom, and F. Powrie. 2000. Cytotoxic T lymphocyte-associ-ated antigen 4 plays an essential role in the function of CD25�CD4� regulatorycells that control intestinal inflammation. J. Exp. Med. 192: 295–302.
45. Salomon, B., D. J. Lenschow, L. Rhee, N. Ashourian, B. Singh, A. Sharpe, andJ. A. Bluestone. 2000. B7/CD28 costimulation is essential for the homeostasis ofthe CD4�CD25� immunoregulatory T cells that control autoimmune diabetes.Immunity 12: 431–440.
46. Takahashi, T., T. Tagami, S. Yamazaki, T. Uede, J. Shimizu, N. Sakaguchi,T. W. Mak, and S. Sakaguchi. 2000. Immunologic self-tolerance maintained byCD25�CD4� regulatory T cells constitutively expressing cytotoxic T lympho-cyte-associated antigen 4. J. Exp. Med. 192: 303–310.
772 HUMAN Treg CELL DIFFERENTIATION AND HIV INFECTION
by guest on July 25, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

47. Iellem, A., M. Mariani, R. Lang, H. Recalde, P. Panina-Bordignon, F. Sinigaglia,and D. D’Ambrosio. 2001. Unique chemotactic response profile and specificexpression of chemokine receptors CCR4 and CCR8 by CD4�CD25� regulatoryT cells. J. Exp. Med. 194: 847–853.
48. Dieckmann, D., H. Plottner, S. Berchtold, T. Berger, and G. Schuler. 2001. Exvivo isolation and characterization of CD4�CD25� T cells with regulatory prop-erties from human blood. J. Exp. Med. 193: 1303–1310.
49. Cohen, G. B., S. A. Islam, M. S. Noble, C. Lau, C. Brander, M. A. Altfeld,E. S. Rosenberg, J. E. Schmitz, T. O. Cameron, and S. A. Kalams. 2002. Clono-type tracking of TCR repertoires during chronic virus infections. Virology 304:474–484.
50. Arendt, C. W., G. Tang, and A. Zilberstein. 2003. Vector systems for the deliveryof small interfering RNAs: managing the RISC. Chembiochem. 4: 1129–1136.
51. Earle, K. E., Q. Tang, X. Zhou, W. Liu, S. Zhu, M. L. Bonyhadi, andJ. A. Bluestone. 2005. In vitro expanded human CD4�CD25� regulatory T cellssuppress effector T cell proliferation. Clin. Immunol. 115: 3–9.
52. Gansuvd, B., C. K. Asiedu, J. Goodwin, U. Jargal, L. A. Deckard, P. Andrades,V. Guarcello, and J. M. Thomas. 2007. Expansion of CD4�CD25� suppressiveregulatory T cells from rhesus macaque peripheral blood by FN18/antihumanCD28-coated Dynal beads. Hum. Immunol. 68: 478–490.
53. Godfrey, W. R., Y. G. Ge, D. J. Spoden, B. L. Levine, C. H. June, B. R. Blazar,and S. B. Porter. 2004. In vitro-expanded human CD4�CD25� T-regulatory cellscan markedly inhibit allogeneic dendritic cell-stimulated MLR cultures. Blood104: 453–461.
54. Hoffmann, P., R. Eder, T. J. Boeld, K. Doser, B. Piseshka, R. Andreesen, andM. Edinger. 2006. Only the CD45RA� subpopulation of CD4�CD25high T cellsgives rise to homogeneous regulatory T-cell lines upon in vitro expansion. Blood108: 4260–4267.
55. Hori, S., T. Nomura, and S. Sakaguchi. 2003. Control of regulatory T cell de-velopment by the transcription factor Foxp3. Science 299: 1057–1061.
56. Khattri, R., T. Cox, S. A. Yasayko, and F. Ramsdell. 2003. An essential role forScurfin in CD4�CD25� T regulatory cells. Nat. Immunol. 4: 337–342.
57. Takahata, Y., A. Nomura, H. Takada, S. Ohga, K. Furuno, S. Hikino,H. Nakayama, S. Sakaguchi, and T. Hara. 2004. CD25�CD4� T cells in human
cord blood: an immunoregulatory subset with naive phenotype and specific ex-pression of forkhead box p3 (Foxp3) gene. Exp. Hematol. 32: 622–629.
58. Thornton, A. M., and E. M. Shevach. 2000. Suppressor effector function ofCD4�CD25� immunoregulatory T cells is antigen nonspecific. J. Immunol. 164:183–190.
59. Li, J., M. Bracht, X. Shang, J. Radewonuk, E. Emmell, D. E. Griswold, and L. Li.2006. Ex vivo activated OVA specific and non-specific CD4�CD25� regulatoryT cells exhibit comparable suppression to OVA mediated T cell responses. Cell.Immunol. 241: 75–84.
60. Vukmanovic-Stejic, M., Y. Zhang, J. E. Cook, J. M. Fletcher, A. McQuaid,J. E. Masters, M. H. Rustin, L. S. Taams, P. C. Beverley, D. C. Macallan, andA. N. Akbar. 2006. Human CD4� CD25hi Foxp3� regulatory T cells are derivedby rapid turnover of memory populations in vivo. J. Clin. Invest. 116:2423–2433.
61. Fritzsching, B., N. Oberle, E. Pauly, R. Geffers, J. Buer, J. Poschl, P. Krammer,O. Linderkamp, and E. Suri-Payer. 2006. Naive regulatory T cells: a novel sub-population defined by resistance toward CD95L-mediated cell death. Blood 108:3371–3378.
62. Seddon, B., and D. Mason. 1999. Peripheral autoantigen induces regulatory Tcells that prevent autoimmunity. J. Exp. Med. 189: 877–882.
63. Picca, C. C., and A. J. Caton. 2005. The role of self-peptides in the developmentof CD4� CD25� regulatory T cells. Curr. Opin. Immunol. 17: 131–136.
64. Walker, M. R., D. J. Kasprowicz, V. H. Gersuk, A. Benard, M. Van Landeghen,J. H. Buckner, and S. F. Ziegler. 2003. Induction of FoxP3 and acquisition of Tregulatory activity by stimulated human CD4�CD25� T cells. J. Clin. Invest.112: 1437–1443.
65. Morgan, M. E., J. H. van Bilsen, A. M. Bakker, B. Heemskerk, M. W. Schilham,F. C. Hartgers, B. G. Elferink, L. van der Zanden, R. R. de Vries, T. W. Huizinga,et al. 2005. Expression of FOXP3 mRNA is not confined to CD4�CD25� Tregulatory cells in humans. Hum. Immunol. 66: 13–20.
66. Grant, C., U. Oh, K. Fugo, N. Takenouchi, C. Griffith, K. Yao, T. E. Newhook,L. Ratner, and S. Jacobson. 2006. Foxp3 represses retroviral transcription bytargeting both NF-�B and CREB pathways. PLoS Pathog. 2: e33.
67. Lackner, A. A., and R. S. Veazey. 2007. Current concepts in AIDS pathogenesis:insights from the SIV/macaque model. Annu. Rev. Med. 58: 461–476.
68. Taams, L. S., J. Smith, M. H. Rustin, M. Salmon, L. W. Poulter, and A. N. Akbar.2001. Human anergic/suppressive CD4�CD25� T cells: a highly differentiatedand apoptosis-prone population. Eur. J. Immunol. 31: 1122–1131.
773The Journal of Immunology
by guest on July 25, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
Related Documents











