Structure and dynamics in solution of the complex of Lactobacillus casei dihydrofolate reductase with the new lipophilic antifolate drug trimetrexate VLADIMIR I. POLSHAKOV, 1,2 BERRY BIRDSALL, 1 THOMAS A. FRENKIEL, 3 ANGELO R. GARGARO, 1 and JAMES FEENEY 1 1 Division of Molecular Structure, National Institute for Medical Research, The Ridgeway, Mill Hill, London NW7 1AA, United Kingdom 2 Center for Drug Chemistry, Moscow 119815, Russia 3 MRC Biomedical NMR Centre, National Institute for Medical Research, The Ridgeway, Mill Hill, London NW7 1AA, United Kingdom ~Received July 29, 1998; Accepted November 3, 1998! Abstract We have determined the three-dimensional solution structure of the complex of Lactobacillus casei dihydrofolate reductase and the anticancer drug trimetrexate. Two thousand seventy distance, 345 dihedral angle, and 144 hydrogen bond restraints were obtained from analysis of multidimensional NMR spectra recorded for complexes containing 15 N-labeled protein. Simulated annealing calculations produced a family of 22 structures fully consistent with the constraints. Several intermolecular protein-ligand NOEs were obtained by using a novel approach monitoring temper- ature effects of NOE signals resulting from dynamic processes in the bound ligand. At low temperature ~5 8C! the trimethoxy ring of bound trimetrexate is flipping sufficiently slowly to give narrow signals in slow exchange, which give good NOE cross peaks. At higher temperature these broaden and their NOE cross peaks disappear thus allowing the signals in the lower-temperature spectrum to be identified as NOEs involving ligand protons. The binding site for trimetrexate is well defined and this was compared with the binding sites in related complexes formed with methotrexate and trimethoprim. No major conformational differences were detected between the different complexes. The 2,4- diaminopyrimidine-containing moieties in the three drugs bind essentially in the same binding pocket and the remaining parts of their molecules adapt their conformations such that they can make effective van der Waals interactions with essentially the same set of hydrophobic amino acids, the side-chain orientations and local conformations of which are not greatly changed in the different complexes ~similar x 1 and x 2 values!. Keywords: dihydrofolate reductase; dynamics; 15 N relaxation; NMR; protein-ligand interactions; ring flipping; structure determination; trimetrexate Dihydrofolate reductase ~ EC 1.5.1.3! catalyzes the reduction of 7,8-dihydrofolate to 5,6,7,8-tetrahydrofolate using NADPH as co- enzyme. This reaction maintains the intracellular pool of tetra- hydrofolate required by several folate-dependent enzyme systems involved in important steps in DNA synthesis, such as the biosyn- thesis of purines and the conversion of deoxyuridine monophos- phate to deoxythymidine monophosphate ~ Blakley, 1985!. The important cellular role of dihydrofolate reductase has led to it being used as the target for several clinically useful antifolate drugs. Agents, such as trimethoprim ~antibacterial!, pyrimethamine ~antimalarial!, and methotrexate ~anticancer!, act by inhibiting the enzyme in parasitic or malignant cells. The relatively small size of the enzyme ~18–20 kDa! makes it amenable to detailed structural studies ~see reviews by Roberts, 1983; Freisheim & Matthews, 1984; Blakley, 1985; Feeney, 1986, 1990, 1996; Feeney & Bird- sall, 1993!. These have been carried out on complexes of the enzyme using both X-ray ~ Bolin et al., 1982; Matthews et al., 1985a, 1985b; Sawaya & Kraut, 1997; and references in Gargaro et al., 1998! and NMR methods ~ Martorell et al., 1994; Morgan et al., 1995; Johnson et al., 1997; Gargaro et al., 1998!. These studies have provided information about structures, interactions, Reprint requests to: J. Feeney, Division of Molecular Structure, National Institute for Medical Research, Mill Hill, London NW7 1AA, United King- dom; e-mail: [email protected]. Abbreviations: COSY, correlated spectroscopy; DANTE, delays alter- nating with nutation for tailored excitation; DHFR, dihydrofolate reduc- tase; DSS, sodium 2,2-dimethyl-2-silapentane-5-sulfonate; DQF, double quantum filtered; GARP-1, a broadband decoupling sequence; HNHA, 3D experiment correlating amide NH and a-CH protons; HNHB, 3D experi- ment correlating amide NH and b-CH protons; HSQC, heteronuclear single quantum coherence spectroscopy; MTX, methotrexate; MLEV-17, isotro- pic mixing sequence; NMR, nuclear magnetic resonance; NOE, nuclear Overhauser effect; NOESY, nuclear Overhauser effect spectroscopy; RMSD, root-mean-square deviation; ROESY, rotating frame Overhauser effect spec- troscopy; SCUBA, stimulated cross peaks under bleached alphas; S rep , representative structure; TMP, trimethoprim; TMQ, trimetrexate; TOCSY, total correlation time spectroscopy; vdW, van der Waals; 1D, one- dimensional; 2D, two-dimensional; 3D, three-dimensional; 4D, four- dimensional. Protein Science ~1999!, 8:467–481. Cambridge University Press. Printed in the USA. Copyright © 1999 The Protein Society 467

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Structure and dynamics in solution of the complexof Lactobacillus caseidihydrofolate reductase withthe new lipophilic antifolate drug trimetrexate
VLADIMIR I. POLSHAKOV, 1,2 BERRY BIRDSALL,1 THOMAS A. FRENKIEL,3
ANGELO R. GARGARO,1 and JAMES FEENEY1
1Division of Molecular Structure, National Institute for Medical Research, The Ridgeway, Mill Hill,London NW7 1AA, United Kingdom
2Center for Drug Chemistry, Moscow 119815, Russia3MRC Biomedical NMR Centre, National Institute for Medical Research, The Ridgeway, Mill Hill,London NW7 1AA, United Kingdom
~Received July 29, 1998;Accepted November 3, 1998!
Abstract
We have determined the three-dimensional solution structure of the complex ofLactobacillus caseidihydrofolatereductase and the anticancer drug trimetrexate. Two thousand seventy distance, 345 dihedral angle, and 144 hydrogenbond restraints were obtained from analysis of multidimensional NMR spectra recorded for complexes containing15N-labeled protein. Simulated annealing calculations produced a family of 22 structures fully consistent with theconstraints. Several intermolecular protein-ligand NOEs were obtained by using a novel approach monitoring temper-ature effects of NOE signals resulting from dynamic processes in the bound ligand. At low temperature~5 8C! thetrimethoxy ring of bound trimetrexate is flipping sufficiently slowly to give narrow signals in slow exchange, which givegood NOE cross peaks. At higher temperature these broaden and their NOE cross peaks disappear thus allowing thesignals in the lower-temperature spectrum to be identified as NOEs involving ligand protons. The binding site fortrimetrexate is well defined and this was compared with the binding sites in related complexes formed with methotrexateand trimethoprim. No major conformational differences were detected between the different complexes. The 2,4-diaminopyrimidine-containing moieties in the three drugs bind essentially in the same binding pocket and the remainingparts of their molecules adapt their conformations such that they can make effective van der Waals interactions withessentially the same set of hydrophobic amino acids, the side-chain orientations and local conformations of which arenot greatly changed in the different complexes~similar x1 andx2 values!.
Keywords: dihydrofolate reductase; dynamics;15N relaxation; NMR; protein-ligand interactions; ring flipping;structure determination; trimetrexate
Dihydrofolate reductase~EC 1.5.1.3! catalyzes the reduction of7,8-dihydrofolate to 5,6,7,8-tetrahydrofolate using NADPH as co-
enzyme. This reaction maintains the intracellular pool of tetra-hydrofolate required by several folate-dependent enzyme systemsinvolved in important steps in DNA synthesis, such as the biosyn-thesis of purines and the conversion of deoxyuridine monophos-phate to deoxythymidine monophosphate~Blakley, 1985!. Theimportant cellular role of dihydrofolate reductase has led to itbeing used as the target for several clinically useful antifolatedrugs. Agents, such as trimethoprim~antibacterial!, pyrimethamine~antimalarial!, and methotrexate~anticancer!, act by inhibiting theenzyme in parasitic or malignant cells. The relatively small size ofthe enzyme~18–20 kDa! makes it amenable to detailed structuralstudies~see reviews by Roberts, 1983; Freisheim & Matthews,1984; Blakley, 1985; Feeney, 1986, 1990, 1996; Feeney & Bird-sall, 1993!. These have been carried out on complexes of theenzyme using both X-ray~Bolin et al., 1982; Matthews et al.,1985a, 1985b; Sawaya & Kraut, 1997; and references in Gargaroet al., 1998! and NMR methods~Martorell et al., 1994; Morganet al., 1995; Johnson et al., 1997; Gargaro et al., 1998!. Thesestudies have provided information about structures, interactions,
Reprint requests to: J. Feeney, Division of Molecular Structure, NationalInstitute for Medical Research, Mill Hill, London NW7 1AA, United King-dom; e-mail: [email protected].
Abbreviations:COSY, correlated spectroscopy; DANTE, delays alter-nating with nutation for tailored excitation; DHFR, dihydrofolate reduc-tase; DSS, sodium 2,2-dimethyl-2-silapentane-5-sulfonate; DQF, doublequantum filtered; GARP-1, a broadband decoupling sequence; HNHA, 3Dexperiment correlating amide NH anda-CH protons; HNHB, 3D experi-ment correlating amide NH andb-CH protons; HSQC, heteronuclear singlequantum coherence spectroscopy; MTX, methotrexate; MLEV-17, isotro-pic mixing sequence; NMR, nuclear magnetic resonance; NOE, nuclearOverhauser effect; NOESY, nuclear Overhauser effect spectroscopy; RMSD,root-mean-square deviation; ROESY, rotating frame Overhauser effect spec-troscopy; SCUBA, stimulated cross peaks under bleached alphas;Srep,representative structure; TMP, trimethoprim; TMQ, trimetrexate; TOCSY,total correlation time spectroscopy; vdW, van der Waals; 1D, one-dimensional; 2D, two-dimensional; 3D, three-dimensional; 4D, four-dimensional.
Protein Science~1999!, 8:467–481. Cambridge University Press. Printed in the USA.Copyright © 1999 The Protein Society
467

and ionization states in several complexes of dihydrofolate reduc-tase with substrates and inhibitors~Birdsall et al., 1981, 1982,1984, 1989a, 1989b, 1990, 1997; Cocco et al., 1981; Gronenbornet al., 1981; Antonjuk et al., 1984; Clore et al., 1984; Bevan et al.,1985; Hammond et al., 1986; Searle et al., 1988; Carr et al., 1991;Cheung et al., 1993; Falzone et al., 1994; Epstein et al., 1995;Polshakov et al., 1995a; Gargaro et al., 1996; Gerothanassis et al.,1996; Nieto et al., 1997!.
In the search for improved inhibitors of dihydrofolate reductase,numerous antifolate analogues have been synthesized and investi-gated~Roth & Cheng, 1982; Baccanari & Kuyper, 1993!. One prom-ising antifolate analogue is trimetrexate~2,4-diamino-5-methyl-6-@~3,4,5-trimethoxyanilino!methyl#quinazoline! ~Fig. 1A! ~Bertino etal., 1979!. Trimetrexate was first prepared by Elslager and co-workers~Elslager et al., 1983! and was investigated initially as a potentialantimalarial agent. Subsequently, it was found to have antineoplas-tic activity in relation to breast, head, and neck cancers~Robert, 1988!and later it was used as an antibacterial in the treatment ofPneu-mocystis cariniipneumonia~PCP! in AIDS patients~Lin & Ber-tino, 1987!. In 1993 trimetrexate~neutrexin! was approved by theU.S. Food and Drug Administration for the treatment of moderate-to-severe PCP in people with compromised immune systems.
The structure of trimetrexate~Fig. 1A! combines some of thefeatures of trimethoprim~Fig. 1B! and methotrexate~Fig. 1C!.Because trimetrexate lacks the terminal glutamic acid moiety ofmethotrexate, it is more lipophilic and does not need active trans-port for penetration into the cell~Lin & Bertino, 1991!. This sig-nificantly increases its efficacy against cells resistant to methotrexate.
Inhibition studies with the human enzyme show that althoughtrimetrexate is about an order of magnitude less potent than meth-
otrexate, it is still an extremely tightly-binding inhibitor~Jacksonet al., 1984!.
Recent studies on the antipneumocystis activity of trimetrexatehave stimulated the synthesis and biological testing of several newnonclassical antifolates~Gangjee et al., 1996!. Gangjee and co-workers have used computer assisted molecular modeling of com-plexes of dihydrofolate reductase to help in developing novelantifolates~Gangjee et al., 1995!. Such rational drug design studiescould be improved if more structural information were availablefor complexes of dihydrofolate reductases and trimetrexate.
In the present work, we report the high resolution solution struc-ture of the binary complex ofLactobacillus caseidihydrofolatereductase with trimetrexate. Detailed information about the con-formation of the bound drug and the structure of its binding sitehas been obtained. This information allows us to compare thebinding site with those of methotrexate~Gargaro et al., 1998! andtrimethoprim~Martorell et al., 1994! and to examine any differ-ences in their modes of binding. The dynamics of the protein andthe bound drug and the specific interactions between the enzymewith the drug have also been characterized.
Results
Signal assignments for the DHFR{trimetrexate complex
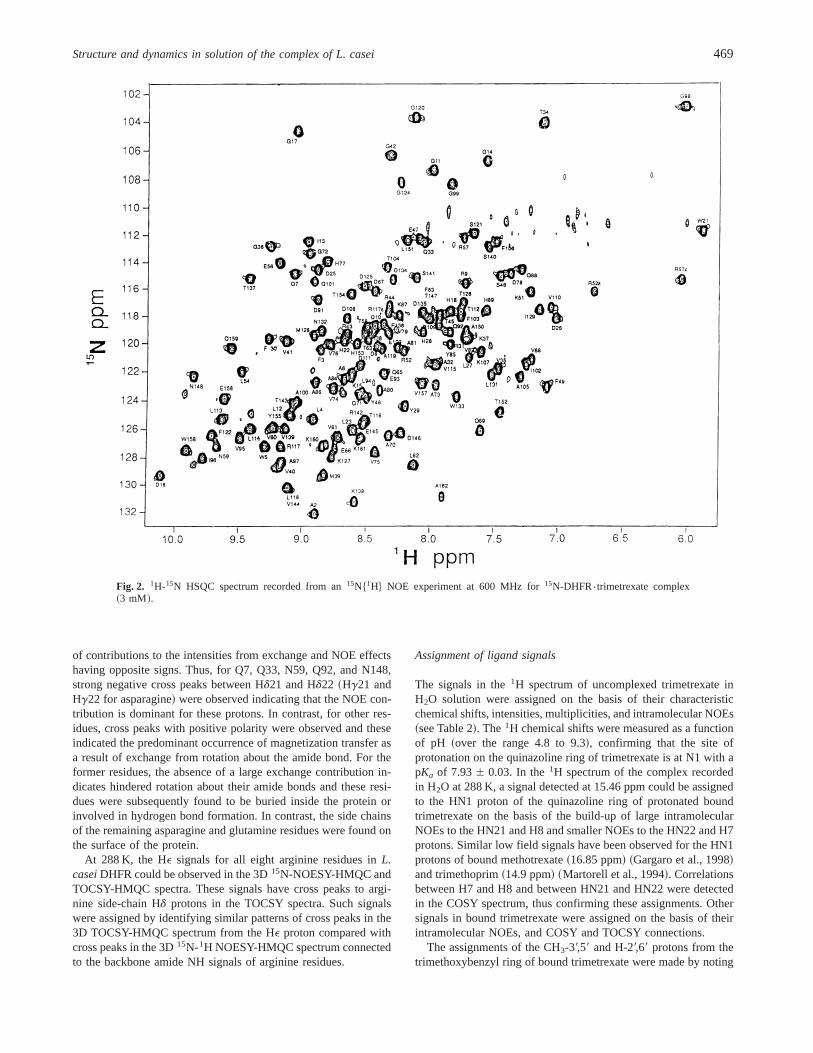
These assignments for the complex were made by analyzing 2D1H NMR spectra of the DHFR{trimetrexate complex obtainedfrom COSY, TOCSY, NOESY, and ROESY experiments and 3D15N-1H NMR spectra from TOCSY-HMQC, NOESY-HMQC,ROESY-HMQC, and HNHB experiments carried out on uniformly15N-labeled DHFR samples using procedures similar to those de-scribed earlier for the DHFR{methotrexate complex~Carr et al.,1991; Gargaro et al., 1998!. The 15N-1H HSQC spectrum of theDHFR{trimetrexate complex with all the resonances labeled isshown in Figure 2 and a table containing the1H and 15N signalassignments is provided in Electronic supplementary material.
In earlier studies on the DHFR{methotrexate complex, almostcomplete assignments of the proton NMR signals were obtained byanalyzing the 3D and 4D spectra from experiments carried out onuniformly 15N and13C015N labeled enzyme~Soteriou et al., 1993;Gargaro et al., 1998!. These assignments for the DHFR{methotrexatecomplex assisted in the assignment of signals from some of thelong-chain amino acids in the spectra of the DHFR{trimetrexatecomplex, because many signals have similar chemical shifts in thetwo complexes and show similar patterns of NOEs.
Coupling constant and distance-related NMR data were used toderive the stereospecific assignments ofb-methylene protons andof valine g-methyl groups using the program AngleSearch as de-scribed in Materials and methods~Polshakov et al., 1995b!.
Signals from all the side-chain amide protons of asparagine andglutamine residues could easily be assigned in 2D15N-1H TOCSY-HMQC and the 3D15N-1H TOCSY-HMQC and NOESY-HMQCspectra by identifying characteristic NOE patterns involving theNH2 amide side-chain protons. The side-chain amide protons Hg22in asparagine and Hd22 in glutamine were determined as thosecloser to Hb protons~or Hg protons in glutamine! than are Hg21or Hd21, respectively, and the stereospecific assignments of theseprotons could thus be obtained directly from the intensity infor-mation in ROESY spectra. In the 3D ROESY-HMQC spectra, cor-relations between amide proton of asparagine and glutamine residuessometimes had different polarities for different residues as a result
A
B
C
Fig. 1. Molecular structures of~A! trimetrexate,~B! trimethorim, and~C!methotrexate.
468 V.I. Polshakov et al.
of contributions to the intensities from exchange and NOE effectshaving opposite signs. Thus, for Q7, Q33, N59, Q92, and N148,strong negative cross peaks between Hd21 and Hd22 ~Hg21 andHg22 for asparagine! were observed indicating that the NOE con-tribution is dominant for these protons. In contrast, for other res-idues, cross peaks with positive polarity were observed and theseindicated the predominant occurrence of magnetization transfer asa result of exchange from rotation about the amide bond. For theformer residues, the absence of a large exchange contribution in-dicates hindered rotation about their amide bonds and these resi-dues were subsequently found to be buried inside the protein orinvolved in hydrogen bond formation. In contrast, the side chainsof the remaining asparagine and glutamine residues were found onthe surface of the protein.
At 288 K, the HE signals for all eight arginine residues inL.caseiDHFR could be observed in the 3D15N-NOESY-HMQC andTOCSY-HMQC spectra. These signals have cross peaks to argi-nine side-chain Hd protons in the TOCSY spectra. Such signalswere assigned by identifying similar patterns of cross peaks in the3D TOCSY-HMQC spectrum from the HE proton compared withcross peaks in the 3D15N-1H NOESY-HMQC spectrum connectedto the backbone amide NH signals of arginine residues.
Assignment of ligand signals
The signals in the1H spectrum of uncomplexed trimetrexate inH2O solution were assigned on the basis of their characteristicchemical shifts, intensities, multiplicities, and intramolecular NOEs~see Table 2!. The1H chemical shifts were measured as a functionof pH ~over the range 4.8 to 9.3!, confirming that the site ofprotonation on the quinazoline ring of trimetrexate is at N1 with apKa of 7.936 0.03. In the1H spectrum of the complex recordedin H2O at 288 K, a signal detected at 15.46 ppm could be assignedto the HN1 proton of the quinazoline ring of protonated boundtrimetrexate on the basis of the build-up of large intramolecularNOEs to the HN21 and H8 and smaller NOEs to the HN22 and H7protons. Similar low field signals have been observed for the HN1protons of bound methotrexate~16.85 ppm! ~Gargaro et al., 1998!and trimethoprim~14.9 ppm! ~Martorell et al., 1994!. Correlationsbetween H7 and H8 and between HN21 and HN22 were detectedin the COSY spectrum, thus confirming these assignments. Othersignals in bound trimetrexate were assigned on the basis of theirintramolecular NOEs, and COSY and TOCSY connections.
The assignments of the CH3-39,59 and H-29,69 protons from thetrimethoxybenzyl ring of bound trimetrexate were made by noting
Fig. 2. 1H-15N HSQC spectrum recorded from an15N$1H% NOE experiment at 600 MHz for15N-DHFR{trimetrexate complex~3 mM!.
Structure and dynamics in solution of the complex of L. casei 469
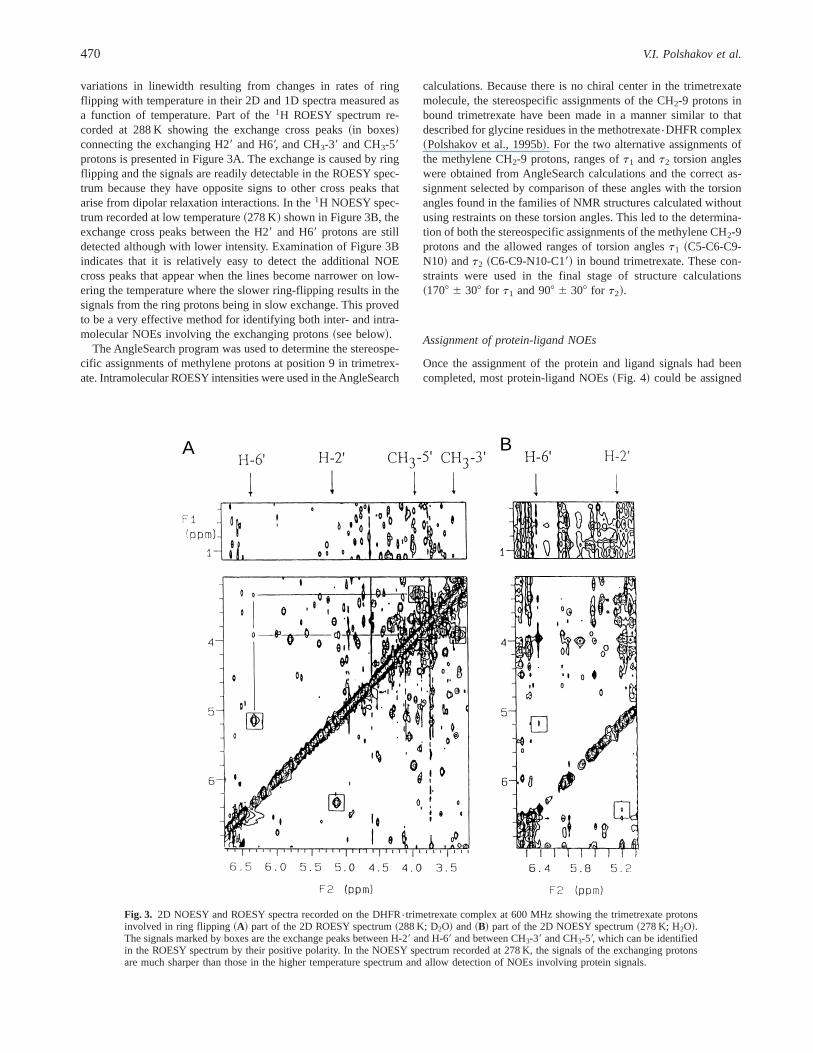
variations in linewidth resulting from changes in rates of ringflipping with temperature in their 2D and 1D spectra measured asa function of temperature. Part of the1H ROESY spectrum re-corded at 288 K showing the exchange cross peaks~in boxes!connecting the exchanging H29 and H69, and CH3-39 and CH3-59protons is presented in Figure 3A. The exchange is caused by ringflipping and the signals are readily detectable in the ROESY spec-trum because they have opposite signs to other cross peaks thatarise from dipolar relaxation interactions. In the1H NOESY spec-trum recorded at low temperature~278 K! shown in Figure 3B, theexchange cross peaks between the H29 and H69 protons are stilldetected although with lower intensity. Examination of Figure 3Bindicates that it is relatively easy to detect the additional NOEcross peaks that appear when the lines become narrower on low-ering the temperature where the slower ring-flipping results in thesignals from the ring protons being in slow exchange. This provedto be a very effective method for identifying both inter- and intra-molecular NOEs involving the exchanging protons~see below!.
The AngleSearch program was used to determine the stereospe-cific assignments of methylene protons at position 9 in trimetrex-ate. Intramolecular ROESY intensities were used in the AngleSearch
calculations. Because there is no chiral center in the trimetrexatemolecule, the stereospecific assignments of the CH2-9 protons inbound trimetrexate have been made in a manner similar to thatdescribed for glycine residues in the methotrexate{DHFR complex~Polshakov et al., 1995b!. For the two alternative assignments ofthe methylene CH2-9 protons, ranges oft1 andt2 torsion angleswere obtained from AngleSearch calculations and the correct as-signment selected by comparison of these angles with the torsionangles found in the families of NMR structures calculated withoutusing restraints on these torsion angles. This led to the determina-tion of both the stereospecific assignments of the methylene CH2-9protons and the allowed ranges of torsion anglest1 ~C5-C6-C9-N10! andt2 ~C6-C9-N10-C19! in bound trimetrexate. These con-straints were used in the final stage of structure calculations~1708 6 308 for t1 and 908 6 308 for t2!.
Assignment of protein-ligand NOEs
Once the assignment of the protein and ligand signals had beencompleted, most protein-ligand NOEs~Fig. 4! could be assigned
A B
Fig. 3. 2D NOESY and ROESY spectra recorded on the DHFR{trimetrexate complex at 600 MHz showing the trimetrexate protonsinvolved in ring flipping~A! part of the 2D ROESY spectrum~288 K; D2O! and~B! part of the 2D NOESY spectrum~278 K; H2O!.The signals marked by boxes are the exchange peaks between H-29 and H-69 and between CH3-39 and CH3-59, which can be identifiedin the ROESY spectrum by their positive polarity. In the NOESY spectrum recorded at 278 K, the signals of the exchanging protonsare much sharper than those in the higher temperature spectrum and allow detection of NOEs involving protein signals.
470 V.I. Polshakov et al.

from 2D NOESY spectra recorded with 50 and 100 ms mixingtime at 308, 288 and 278 K.
In most cases ligand NOE partners could be assigned unambig-uously because of their unique chemical shifts or characteristic lineshapes. Signals from the 29,69-ring protons and the 39,59-methoxyprotons of the trimethoxyphenyl ring could be uniquely identifiedbecause of their changes in line shape with temperature: decreaseof temperature caused these signals to become sharper due toslower ring flipping whereas most of the protein signals becamebroader. This ability to manipulate the detection of NOEs by rais-ing or lowering the temperature greatly facilitated the detection ofthe protein-ligand NOEs. Furthermore, the availability of NOEsinvolving resolved protons from opposite sides of the ring in-creases the total number of detected NOEs. The method has gen-eral applicability in cases where there is an intramolecular rateprocess that can influence the line width of the ligand signalsdifferently as the temperature is changed. In some cases, proteinpartners in protein-ligand NOEs were also assigned by identifyingtheir characteristic set of NOEs~for example, the trimetrexate H8proton has NOEs to several protons of L27—Ha, Hg, Hd1, andHd2!. For assignment of the remaining NOEs, the ambiguitieswere resolved in an iterative manner during the refinement stage ofthe structure calculation. A summary of the extensive number ofintermolecular NOEs between trimetrexate and protein protons isgiven in Figure 4.
Structure of the DHFR{trimetrexate complex
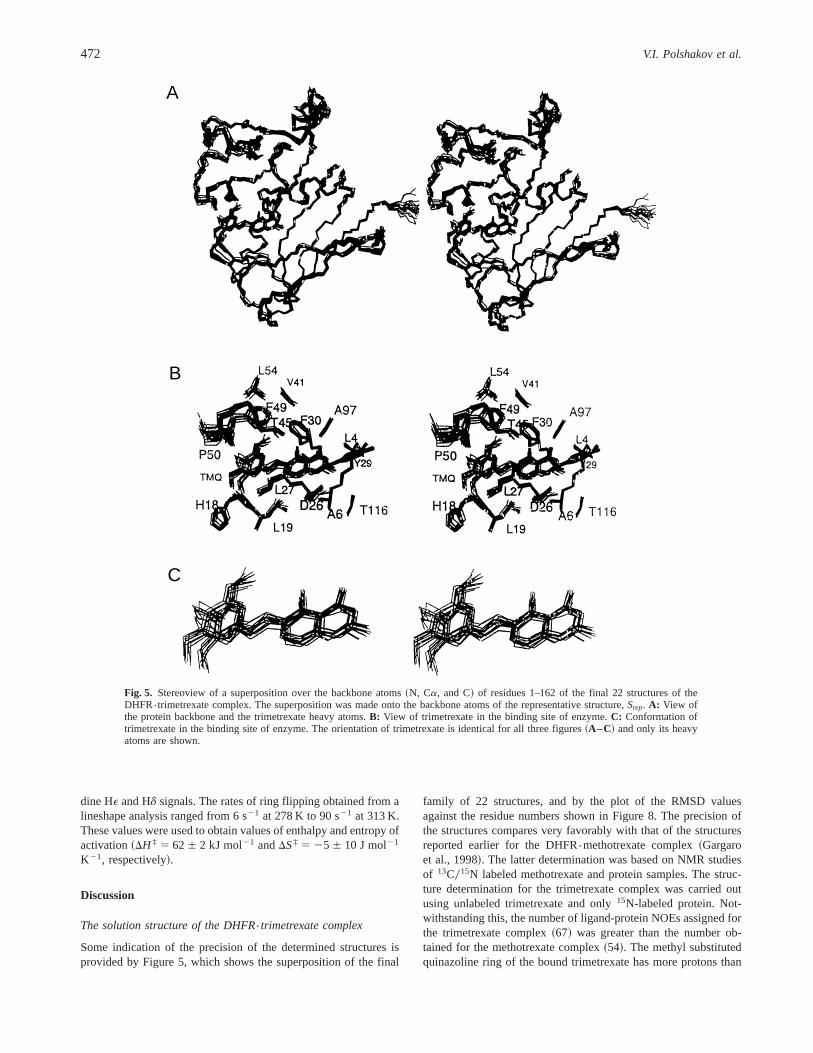
The structures shown in Figure 5 are from the final 22 calculatedstructures superimposed on the backbone of the representative struc-
ture Srep. Figure 5A shows a stereoview of the protein backboneand ligand heavy atoms for all 22 structures. Figure 5B showssome of the important protein side chains and the ligand in itsbinding site. The structures of bound trimetrexate are shown inFigure 5C. All the structures shown in Figures 5A–C are presentedin the same orientation. Some indication of the precision of thestructures can be realized by noting how well the different calcu-lated structures are superimposed~pairwise RMSD is 0.5 Å forbackbone atoms and 1.0 Å for all heavy atoms!. The absence ofdihedral angle and NOE violations and the small energies for theexperimental constraints~see Table 1! provide further evidence forthe precision of the structures. From an examination of the struc-ture ~Fig. 5A! and the PROCHECK analysis~Laskowski et al.,1993, 1996! of the data, the general features of the structure areseen to be essentially identical to the overall structural featuresfound in the solution structure of DHFR{methotrexate~Gargaroet al., 1998! and in the crystal structures of DHFR{methotrexate.NADPH complex~Bolin et al., 1982!.
Backbone dynamics of the DHFR{trimetrexate complex
The overall rotational correlation timetc for the complex of DHFRwith trimetrexate was calculated from the15N T1 andT2 data usingprocedures developed by Kay et al.~1989!. T1 andT2 values wereobtained from a total of 128 residues. Local values of the overallcorrelation time were calculated from theT10T2 ratios of 54 of theresidues~see Materials and methods!; the average of these valuesgives an overall correlation time of 8.756 0.1 ns at 308 K.
The experimental values of the relaxation parameters were in-terpreted using the model-free formalism~Lipari & Szabo, 1982!,with extensions to include slower internal motions~Clore et al.,1990b! and chemical exchange contributionsRex to the transverserelaxation ~Clore et al., 1990a! ~see Electronic supplementarymaterial!.
Values of the model-free parametersS2, te, andRex have beenobtained for all 128 residues for which experimental relaxationdata were obtained.
Figure 6 shows the measured values of the relaxation parametersR1, R2, and NOE~Ri 5 Ti
21! and the calculated generalized orderparameters,S2, plotted against the corresponding residue number.TheteandRexvalues are given in Electronic supplementary material.
Ring flipping in bound trimetrexate
Ring flipping of the 39,49,59-trimethoxyphenyl ring of bound tri-metrexate has been studied using 2D ROESY and 1D NMR-1Hspectra of the complex recorded over the temperature range 278 to313 K. In the 2D ROESY~and 2D NOESY! spectra, one canobserve strong exchange cross peaks between H29 and H69 andbetween CH3-39 and CH3-59 ~Fig. 3!. Decreasing the temperaturereduces the intensities of these cross peaks and at the same timesharpens the signals. This is typical for slow exchange behaviorwhere separate signals are detected for the exchanging sites andthe line widths of the signals have contributions from the ex-change process. This dynamic process has been investigated byline shape analysis of the H69 signal in the 1D spectrum of theDHFR{trimetrexate complex. This signal is observed as a well-resolved signal in a noncrowded region of the1H spectrum~Fig. 7!and this facilitates measurement of its lineshape. An approximateestimate of the nonexchange contribution to the linewidth wasobtained from the mean values of the linewidths of several histi-
Fig. 4. The intermolecular NOEs between protons of trimetrexate and pro-tein in the DHFR{trimetrexate complex.
Structure and dynamics in solution of the complex of L. casei 471
dine HE and Hd signals. The rates of ring flipping obtained from alineshape analysis ranged from 6 s21 at 278 K to 90 s21 at 313 K.These values were used to obtain values of enthalpy and entropy ofactivation~DH ‡ 5 626 2 kJ mol21 andDS‡ 5 25 6 10 J mol21
K21, respectively!.
Discussion
The solution structure of the DHFR{trimetrexate complex
Some indication of the precision of the determined structures isprovided by Figure 5, which shows the superposition of the final
family of 22 structures, and by the plot of the RMSD valuesagainst the residue numbers shown in Figure 8. The precision ofthe structures compares very favorably with that of the structuresreported earlier for the DHFR{methotrexate complex~Gargaroet al., 1998!. The latter determination was based on NMR studiesof 13C015N labeled methotrexate and protein samples. The struc-ture determination for the trimetrexate complex was carried outusing unlabeled trimetrexate and only15N-labeled protein. Not-withstanding this, the number of ligand-protein NOEs assigned forthe trimetrexate complex~67! was greater than the number ob-tained for the methotrexate complex~54!. The methyl substitutedquinazoline ring of the bound trimetrexate has more protons than
A
B
C
Fig. 5. Stereoview of a superposition over the backbone atoms~N, Ca, and C! of residues 1–162 of the final 22 structures of theDHFR{trimetrexate complex. The superposition was made onto the backbone atoms of the representative structure,Srep. A: View ofthe protein backbone and the trimetrexate heavy atoms.B: View of trimetrexate in the binding site of enzyme.C: Conformation oftrimetrexate in the binding site of enzyme. The orientation of trimetrexate is identical for all three figures~A–C! and only its heavyatoms are shown.
472 V.I. Polshakov et al.
the corresponding pteridine ring of methotrexate and these protonsprovide increased opportunities for measuring protein-ligand NOEs.Additionally, the favorable dynamic behavior of the trimethoxyring of trimetrexate allows the measurement of unambiguous NOEs
from protons on each side of the ring at low temperature~278 K!.At this temperature the ring flipping is sufficiently slow to allowthe H29, H69 and CH3-39,CH3-59 protons to give separate signals inslow exchange and to allow detection of protein-ligand NOEs inthe NOESY spectra. These NOEs can be assigned to protein-ligandNOEs by noting their disappearance on raising the temperaturewhere the increased rate of ring flipping broadens the ligand sig-nals. In contrast, the flipping rates of thep-aminobenzoyl ring ofmethotrexate causes line broadening of its aromatic proton signalssuch that no signals could be detected at any of the temperaturesstudied. The low temperature spectra of the DHFR{trimetrexatecomplex also allowed us to make specific NOE assignments forthe separate protons in the trimetrexate NH2 groups. A furtherimprovement in the structure determination of the trimetrexatecomplex resulted from the use of many more NOEs involvingstereospecifically assigned protons and more hydrogen bonds thanwere used in the methotrexate structure calculations. Although18% fewer intramolecular NOEs were used in the calculations forthe trimetrexate complex~resulting from the absence of some Hbs0Hbs, Hgs0Hbs assignments, which could only be obtained from13C data!, the compensating factors mentioned above result in thedetermined structures of the two complexes being of similar pre-cision. The availability of the spectral and structural data for themethotrexate complex assisted in the structural determination ofthe trimetrexate complex. High precision structures for proteins ofthis size can thus be obtained without using13C-labeled protein ifone has a reference structure to assist in obtaining the initial set ofNOE values~see Material and methods!.
Comparisons with structures of related DHFR complexes
Figure 9C shows the structures of the three bound inhibitors~Fig. 1A–C! obtained by superimposing the backbone atoms of therepresentative structures~Srep! of their DHFR complexes~Kine-
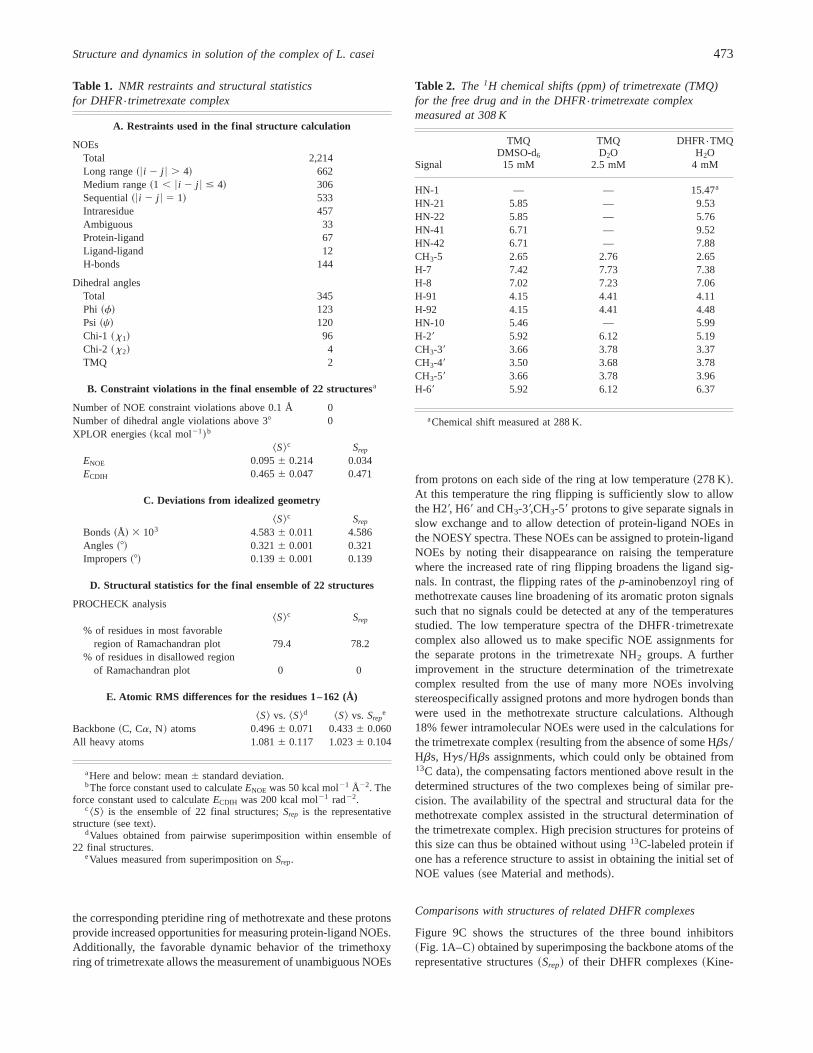
Table 1. NMR restraints and structural statisticsfor DHFR{trimetrexate complex
A. Restraints used in the final structure calculation
NOEsTotal 2,214Long range~6i 2 j 6 . 4! 662Medium range~1 , 6i 2 j 6 # 4! 306Sequential~6i 2 j 6 5 1! 533Intraresidue 457Ambiguous 33Protein-ligand 67Ligand-ligand 12H-bonds 144
Dihedral anglesTotal 345Phi ~f! 123Psi ~c! 120Chi-1 ~x1! 96Chi-2 ~x2! 4TMQ 2
B. Constraint violations in the final ensemble of 22 structuresa
Number of NOE constraint violations above 0.1 Å 0Number of dihedral angle violations above 38 0XPLOR energies~kcal mol21!b
^S&c Srep
ENOE 0.0956 0.214 0.034ECDIH 0.4656 0.047 0.471
C. Deviations from idealized geometry
^S&c Srep
Bonds~Å! 3 103 4.5836 0.011 4.586Angles ~8! 0.3216 0.001 0.321Impropers~8! 0.1396 0.001 0.139
D. Structural statistics for the final ensemble of 22 structures
PROCHECK analysis^S&c Srep
% of residues in most favorableregion of Ramachandran plot 79.4 78.2
% of residues in disallowed regionof Ramachandran plot 0 0
E. Atomic RMS differences for the residues 1–162 (Å)
^S& vs. ^S&d ^S& vs. Srepe
Backbone~C, Ca, N! atoms 0.4966 0.071 0.4336 0.060All heavy atoms 1.0816 0.117 1.0236 0.104
aHere and below: mean6 standard deviation.bThe force constant used to calculateENOE was 50 kcal mol21 Å22. The
force constant used to calculateECDIH was 200 kcal mol21 rad22.c ^S& is the ensemble of 22 final structures;Srep is the representative
structure~see text!.dValues obtained from pairwise superimposition within ensemble of
22 final structures.eValues measured from superimposition onSrep.
Table 2. The 1H chemical shifts (ppm) of trimetrexate (TMQ)for the free drug and in the DHFR{trimetrexate complexmeasured at 308 K
Signal
TMQDMSO-d6
15 mM
TMQD2O
2.5 mM
DHFR{TMQH2O
4 mM
HN-1 — — 15.47a
HN-21 5.85 — 9.53HN-22 5.85 — 5.76HN-41 6.71 — 9.52HN-42 6.71 — 7.88CH3-5 2.65 2.76 2.65H-7 7.42 7.73 7.38H-8 7.02 7.23 7.06H-91 4.15 4.41 4.11H-92 4.15 4.41 4.48HN-10 5.46 — 5.99H-29 5.92 6.12 5.19CH3-39 3.66 3.78 3.37CH3-49 3.50 3.68 3.78CH3-59 3.66 3.78 3.96H-69 5.92 6.12 6.37
aChemical shift measured at 288 K.
Structure and dynamics in solution of the complex of L. casei 473
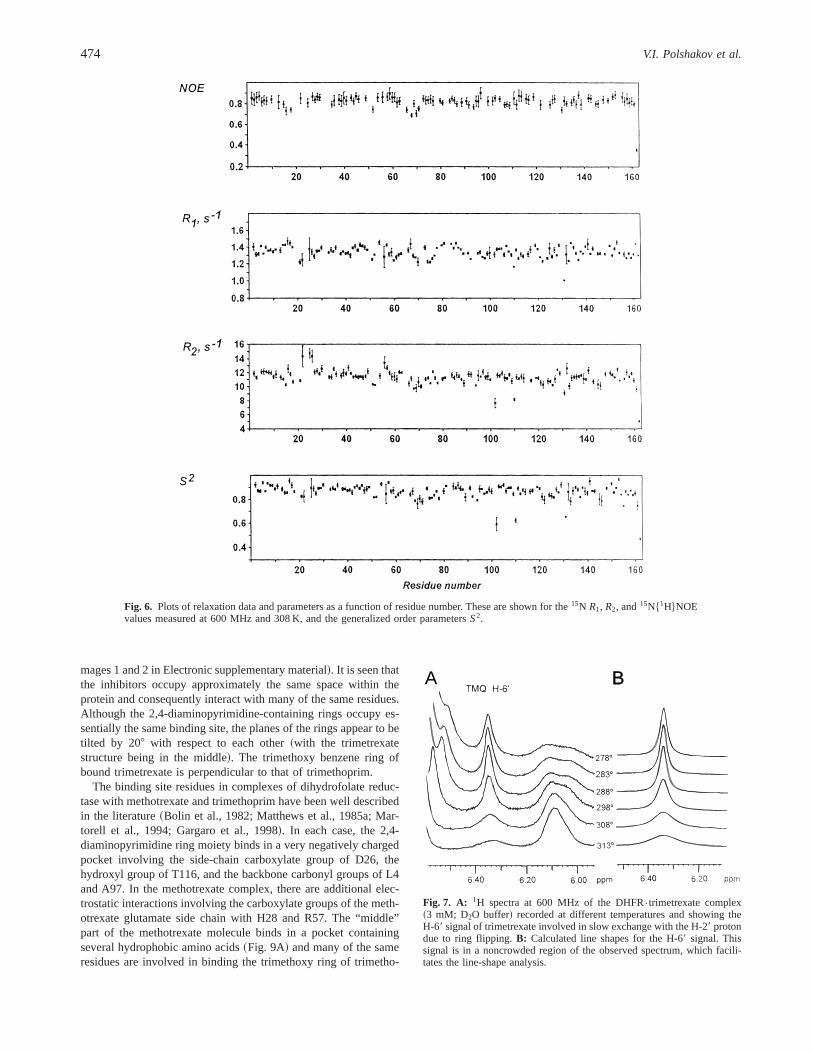
mages 1 and 2 in Electronic supplementary material!. It is seen thatthe inhibitors occupy approximately the same space within theprotein and consequently interact with many of the same residues.Although the 2,4-diaminopyrimidine-containing rings occupy es-sentially the same binding site, the planes of the rings appear to betilted by 208 with respect to each other~with the trimetrexatestructure being in the middle!. The trimethoxy benzene ring ofbound trimetrexate is perpendicular to that of trimethoprim.
The binding site residues in complexes of dihydrofolate reduc-tase with methotrexate and trimethoprim have been well describedin the literature~Bolin et al., 1982; Matthews et al., 1985a; Mar-torell et al., 1994; Gargaro et al., 1998!. In each case, the 2,4-diaminopyrimidine ring moiety binds in a very negatively chargedpocket involving the side-chain carboxylate group of D26, thehydroxyl group of T116, and the backbone carbonyl groups of L4and A97. In the methotrexate complex, there are additional elec-trostatic interactions involving the carboxylate groups of the meth-otrexate glutamate side chain with H28 and R57. The “middle”part of the methotrexate molecule binds in a pocket containingseveral hydrophobic amino acids~Fig. 9A! and many of the sameresidues are involved in binding the trimethoxy ring of trimetho-
Fig. 6. Plots of relaxation data and parameters as a function of residue number. These are shown for the15N R1, R2, and15N$1H%NOEvalues measured at 600 MHz and 308 K, and the generalized order parametersS2.
A BB
Fig. 7. A: 1H spectra at 600 MHz of the DHFR{trimetrexate complex~3 mM; D2O buffer! recorded at different temperatures and showing theH-69 signal of trimetrexate involved in slow exchange with the H-29 protondue to ring flipping.B: Calculated line shapes for the H-69 signal. Thissignal is in a noncrowded region of the observed spectrum, which facili-tates the line-shape analysis.
474 V.I. Polshakov et al.
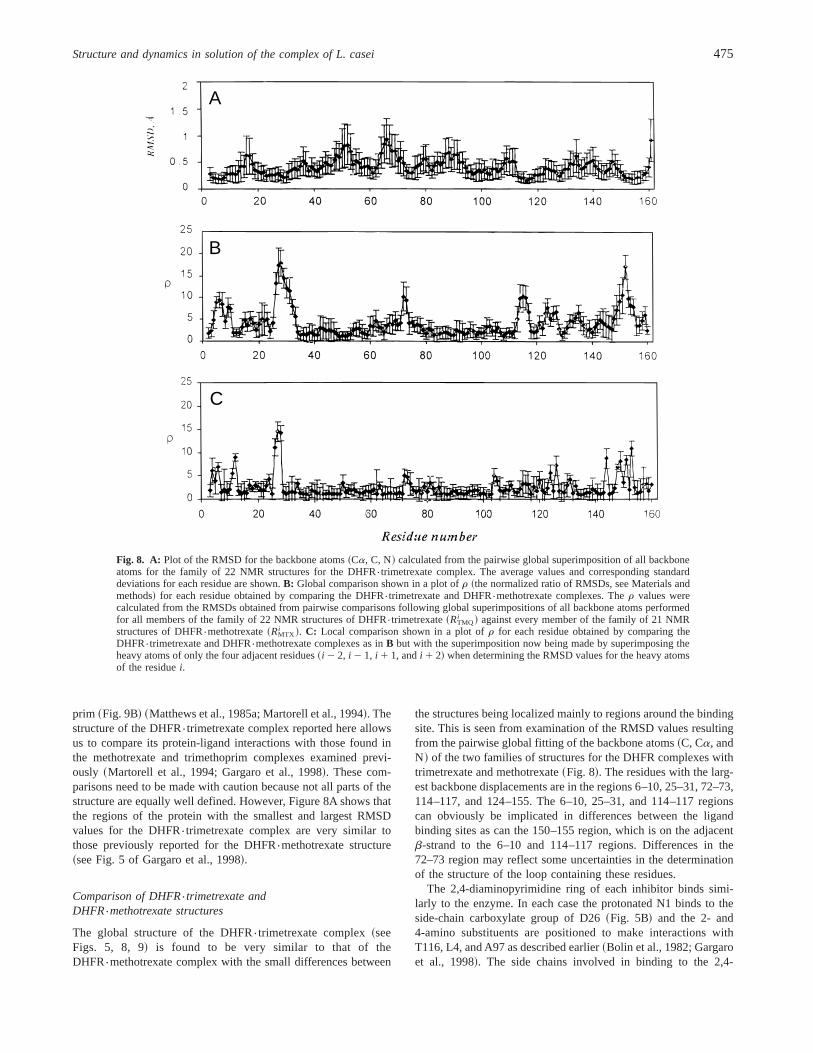
prim ~Fig. 9B! ~Matthews et al., 1985a; Martorell et al., 1994!. Thestructure of the DHFR{trimetrexate complex reported here allowsus to compare its protein-ligand interactions with those found inthe methotrexate and trimethoprim complexes examined previ-ously ~Martorell et al., 1994; Gargaro et al., 1998!. These com-parisons need to be made with caution because not all parts of thestructure are equally well defined. However, Figure 8A shows thatthe regions of the protein with the smallest and largest RMSDvalues for the DHFR{trimetrexate complex are very similar tothose previously reported for the DHFR{methotrexate structure~see Fig. 5 of Gargaro et al., 1998!.
Comparison of DHFR{trimetrexate andDHFR{methotrexate structures
The global structure of the DHFR{trimetrexate complex~seeFigs. 5, 8, 9! is found to be very similar to that of theDHFR{methotrexate complex with the small differences between
the structures being localized mainly to regions around the bindingsite. This is seen from examination of the RMSD values resultingfrom the pairwise global fitting of the backbone atoms~C, Ca, andN! of the two families of structures for the DHFR complexes withtrimetrexate and methotrexate~Fig. 8!. The residues with the larg-est backbone displacements are in the regions 6–10, 25–31, 72–73,114–117, and 124–155. The 6–10, 25–31, and 114–117 regionscan obviously be implicated in differences between the ligandbinding sites as can the 150–155 region, which is on the adjacentb-strand to the 6–10 and 114–117 regions. Differences in the72–73 region may reflect some uncertainties in the determinationof the structure of the loop containing these residues.
The 2,4-diaminopyrimidine ring of each inhibitor binds simi-larly to the enzyme. In each case the protonated N1 binds to theside-chain carboxylate group of D26~Fig. 5B! and the 2- and4-amino substituents are positioned to make interactions withT116, L4, and A97 as described earlier~Bolin et al., 1982; Gargaroet al., 1998!. The side chains involved in binding to the 2,4-
A
B
C
Fig. 8. A: Plot of the RMSD for the backbone atoms~Ca, C, N! calculated from the pairwise global superimposition of all backboneatoms for the family of 22 NMR structures for the DHFR{trimetrexate complex. The average values and corresponding standarddeviations for each residue are shown.B: Global comparison shown in a plot ofr ~the normalized ratio of RMSDs, see Materials andmethods! for each residue obtained by comparing the DHFR{trimetrexate and DHFR{methotrexate complexes. Ther values werecalculated from the RMSDs obtained from pairwise comparisons following global superimpositions of all backbone atoms performedfor all members of the family of 22 NMR structures of DHFR{trimetrexate~RTMQ
i ! against every member of the family of 21 NMRstructures of DHFR{methotrexate~RMTX
i !. C: Local comparison shown in a plot ofr for each residue obtained by comparing theDHFR{trimetrexate and DHFR{methotrexate complexes as inB but with the superimposition now being made by superimposing theheavy atoms of only the four adjacent residues~i 2 2, i 2 1, i 1 1, andi 1 2! when determining the RMSD values for the heavy atomsof the residuei.
Structure and dynamics in solution of the complex of L. casei 475
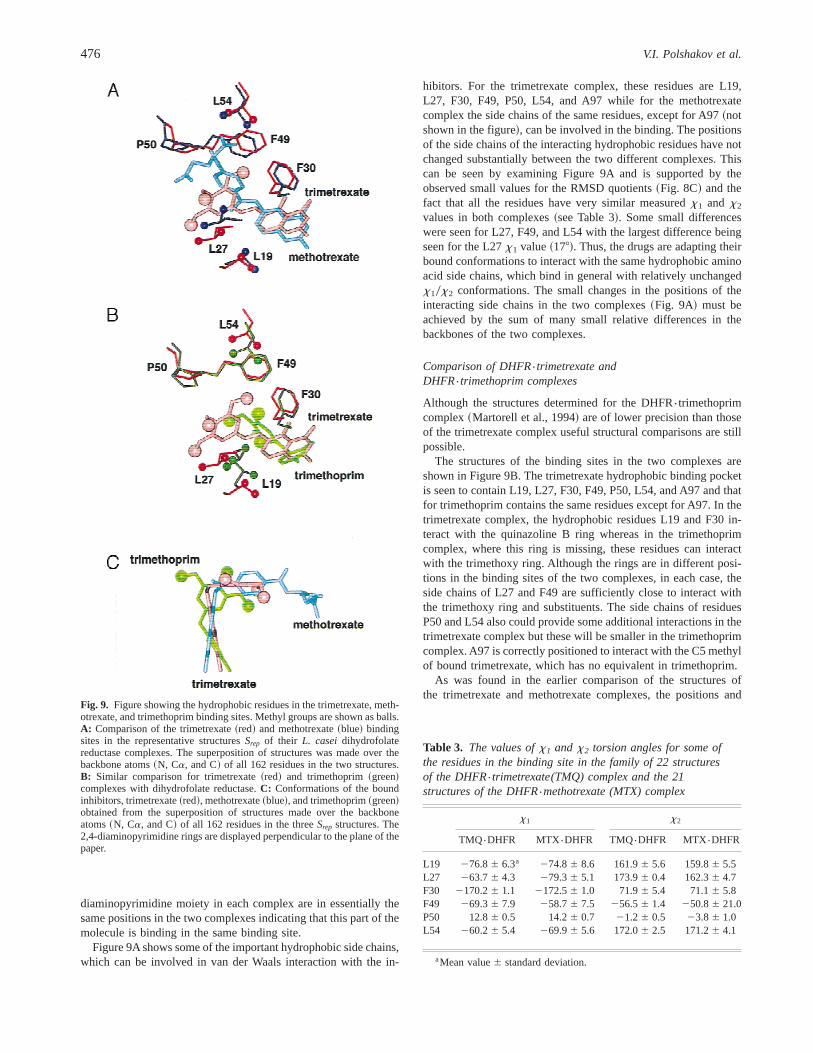
diaminopyrimidine moiety in each complex are in essentially thesame positions in the two complexes indicating that this part of themolecule is binding in the same binding site.
Figure 9A shows some of the important hydrophobic side chains,which can be involved in van der Waals interaction with the in-
hibitors. For the trimetrexate complex, these residues are L19,L27, F30, F49, P50, L54, and A97 while for the methotrexatecomplex the side chains of the same residues, except for A97~notshown in the figure!, can be involved in the binding. The positionsof the side chains of the interacting hydrophobic residues have notchanged substantially between the two different complexes. Thiscan be seen by examining Figure 9A and is supported by theobserved small values for the RMSD quotients~Fig. 8C! and thefact that all the residues have very similar measuredx1 and x2
values in both complexes~see Table 3!. Some small differenceswere seen for L27, F49, and L54 with the largest difference beingseen for the L27x1 value~178!. Thus, the drugs are adapting theirbound conformations to interact with the same hydrophobic aminoacid side chains, which bind in general with relatively unchangedx10x2 conformations. The small changes in the positions of theinteracting side chains in the two complexes~Fig. 9A! must beachieved by the sum of many small relative differences in thebackbones of the two complexes.
Comparison of DHFR{trimetrexate andDHFR{trimethoprim complexes
Although the structures determined for the DHFR{trimethoprimcomplex~Martorell et al., 1994! are of lower precision than thoseof the trimetrexate complex useful structural comparisons are stillpossible.
The structures of the binding sites in the two complexes areshown in Figure 9B. The trimetrexate hydrophobic binding pocketis seen to contain L19, L27, F30, F49, P50, L54, and A97 and thatfor trimethoprim contains the same residues except for A97. In thetrimetrexate complex, the hydrophobic residues L19 and F30 in-teract with the quinazoline B ring whereas in the trimethoprimcomplex, where this ring is missing, these residues can interactwith the trimethoxy ring. Although the rings are in different posi-tions in the binding sites of the two complexes, in each case, theside chains of L27 and F49 are sufficiently close to interact withthe trimethoxy ring and substituents. The side chains of residuesP50 and L54 also could provide some additional interactions in thetrimetrexate complex but these will be smaller in the trimethoprimcomplex. A97 is correctly positioned to interact with the C5 methylof bound trimetrexate, which has no equivalent in trimethoprim.
As was found in the earlier comparison of the structures ofthe trimetrexate and methotrexate complexes, the positions and
Fig. 9. Figure showing the hydrophobic residues in the trimetrexate, meth-otrexate, and trimethoprim binding sites. Methyl groups are shown as balls.A: Comparison of the trimetrexate~red! and methotrexate~blue! bindingsites in the representative structuresSrep of their L. caseidihydrofolatereductase complexes. The superposition of structures was made over thebackbone atoms~N, Ca, and C! of all 162 residues in the two structures.B: Similar comparison for trimetrexate~red! and trimethoprim~green!complexes with dihydrofolate reductase.C: Conformations of the boundinhibitors, trimetrexate~red!, methotrexate~blue!, and trimethoprim~green!obtained from the superposition of structures made over the backboneatoms~N, Ca, and C! of all 162 residues in the threeSrep structures. The2,4-diaminopyrimidine rings are displayed perpendicular to the plane of thepaper.
Table 3. The values ofx1 and x2 torsion angles for some ofthe residues in the binding site in the family of 22 structuresof the DHFR{trimetrexate(TMQ) complex and the 21structures of the DHFR{methotrexate (MTX) complex
x1 x2
TMQ{DHFR MTX{DHFR TMQ{DHFR MTX{DHFR
L19 276.86 6.3a 274.86 8.6 161.96 5.6 159.86 5.5L27 263.76 4.3 279.36 5.1 173.96 0.4 162.36 4.7F30 2170.26 1.1 2172.56 1.0 71.96 5.4 71.16 5.8F49 269.36 7.9 258.76 7.5 256.56 1.4 250.86 21.0P50 12.86 0.5 14.26 0.7 21.26 0.5 23.86 1.0L54 260.26 5.4 269.96 5.6 172.06 2.5 171.26 4.1
aMean value6 standard deviation.
476 V.I. Polshakov et al.

conformations of the interacting hydrophobic side chains in theprotein are not very different. Thus, once again, effective protein-ligand interactions are formed by the drug adopting a bound con-formation which takes advantage of the existing conformations ofthe interacting side chains of the protein.
Relaxation parameters
The measured values of the15N relaxation parameters~T1, T2, andNOE! and the calculated generalized order parameter~S2! plottedagainst the corresponding residue number are shown in Figure 6.The protein in the DHFR{trimetrexate complex appears to have noreally flexible parts: the only residue showing high flexibility is theC-terminal A162. Thus, the data indicate that DHFR{trimetrexatecomplex is globular~averagetc value of 8.756 0.1 ns!, which isin good agreement with the determined structure. The mean valueobtained forS2 is 0.86 and the standard deviation is 0.05. There areseveral residues withS2 values below the mean value minus onestandard deviation, namely, residues 68–71, 102, 110, 122, 131,133, 145, 146, 155, 161, and 162. Most of these are near to theunoccupied binding site of the coenzyme or are situated in loopregions.
Ring flipping in bound trimetrexate
The nonlinear fitting of the experimental kinetic data for the tri-metrexate ring flipping provides the values:DH ‡ 5 62.06 2.0 kJmol21 andDS‡ 5 25 6 10 J mol21 K21. Interestingly, the valueof DS‡ is rather close to the values obtained for ring flipping in39,59-difluoromethotrexate bound toL. caseiDHFR ~215 J mol21
K21 for binary and229 J mol21 K21 for the ternary complex withNADPH! ~Clore et al., 1984!. However, the enthalpy value for ringflipping of trimetrexate is much higher than the values found for39,59-difluoromethotrexate~DH ‡ 5 48.1 kJ mol21 for the binaryand 41.4 kJ mol21 for the ternary complex!. There are two com-ponents in the value ofDH ‡: the intrinsic barrier to rotation aboutthe N10-C19 bond and an additional barrier due to interactions ofthe benzoyl ring with protein residues including any conforma-tional rearrangements required to relieve steric interactions hin-dering the ring flipping. For trimetrexate, the intrinsic barrier torotation would be expected to be less than that for methotrexate,where the intrinsic barrier includes the effects of hindered rotationabout the amide bond to the glutamate moiety. Therefore, a sub-stantial contribution to the value ofDH ‡ ~62 kJ mol21! in thetrimetrexate complex arises from the additional barrier. Becausethe structures of the ligand binding sites are fairly similar in thecomplexes formed with each of the drugs, the difference in theDH ‡ values for complexes of DHFR with trimetrexate and with39,59-diflouromethotrexate probably reflects additional hydropho-bic interactions of the 39,49,59-trimethoxyphenyl ring of trimetrex-ate with residues in the active site. These interactions would needto be broken before ring flipping can take place, thus contribut-ing to the activation barrier~estimated contribution of 10 to 15kJ mol21, that is, the difference 622 48 kJ mol21!.
Materials and methods
Sample preparation
L. caseidihydrofolate reductase was prepared as described previ-ously from anEscherichia colistrain in which the structural gene
for the L. caseienzyme had been cloned~Dann et al., 1976; An-drews et al., 1985!. Trimetrexate was a gift from Warner-Lambert0Parke-Davis~Pontypool, Gwent!. Just over one equivalent oftrimetrexate was added as a solid to the solution of dihydrofolatereductase: after 3–10 h standing, undissolved trimetrexate wasseparated by centrifugation and the final solution dialyzed to re-move uncomplexed trimetrexate.
Binding data
Methotrexate and trimetrexate bind very tightly to DHFR, whichmakes direct measurement of the binding constant difficult.However, the ratios of the binding constantsKa
MTX0KaTMQ and
KaTMQ0Ka
TMP can be estimated from competition experiments. Asample containing a 1:1:1 mixture of DHFR, MTX, and TMQ wasmade at pH 6.5 in 500 mM KCl and 50 mM potassium phosphateand examined after 2 and 7 days to confirm that complete equi-librium had been reached. At equilibrium, the ratioKa
MTX0KaTMQ
equals$@DHFR{MTX #0@DHFR{TMQ#%{$@TMQ#0@MTX #%. Defin-ing h as the ratio@DHFR{MTX #0@DHFR{TMQ#, and assumingthat @DHFRfree# ; 0, which is a good assumption for tightly-binding ligands, it can be shown that for 1:1:1 mixture of DHFR,MTX, and TMQ, ratio@TMQ#0@MTX # can be approximated toh,and thereforeKa
MTX0KaTMQ 5 h2. The ratioh was obtained from
the integrated areas of well-resolved complex-specific signals inthe 1D1H spectra of the complexes. The ratio of binding constantsKa
MTX0KaTMQ is 20 6 10. A similar experiment carried out on a
sample containing a 1:1:1 mixture of DHFR, TMP, and TMQ gavethe ratio of binding constants,Ka
TMQ0KaTMP 5 10 6 5. Ka
TMQ canthen be estimated as;2 3 108 M21 using the previously reportedvalues forKa
MTX ~2 3 109 M 21! and KaTMP ~2 3 107 M 21!
~Birdsall et al., 1980!.
NMR experiments
The NMR experiments were carried out on Varian UNITY andUNITY plus spectrometers operating at proton frequencies of600 and 500 MHz, respectively. The spectra were recorded at278, 288, or 308 K on either unlabeled or15N-labeled proteinsamples with volumes of 0.6 mL and at concentrations 1 or4 mM in 500 mM potassium chloride and 50 mM potassiumphosphate buffer, pH* 6.5, made up either in D2O or in 90%H2O010% D2O ~pH* values are pH meter readings uncorrectedfor deuterium isotope effects!.
Full details of the NMR experiments are provided in Electronicsupplementary material. 2D DQF-COSY, TOCSY, NOESY, andROESY spectra were recorded at temperatures between 278 and308 K for both H2O and D2O samples of DHFR{trimetrexate. 2Dand 3D experiments using uniformly15N labeled protein, HSQC,TOCSY-HMQC, NOESY-HMQC, ROESY-HMQC, and HNHBwere also carried out. Longitudinal and transverse15N relaxationtimes~T1 andT2! and15N$1H%NOE values were measured at 308 Kusing the pulse sequences described by Kay and co-workers~1992!.The 1H chemical shifts were measured from dioxane as an internalreference and then referenced to sodium 2,2-dimethyl-2-silapentane-5-sulfonate~DSS!, relative to which the chemical shift of dioxaneis 3.75 ppm. The15N chemical shifts were referenced indirectly toliquid NH3 using theg ratios method~Live et al., 1984; Wishartet al., 1996!.
Structure and dynamics in solution of the complex of L. casei 477

NMR determined constraints
Distance constraints
Interproton distance constraints were obtained from NOEs mea-sured in 2D and 3D NOESY and ROESY spectra recorded in D2Oand H2O. The set of distance constraints used for generation ofinitial structures contained unambiguously assigned NOEs, andalso a limited set of NOEs for which the ambiguity was resolvedusing the NMR structures of the DHFR{MTX complex ~Gargaroet al., 1998!. The total number of NOEs used for initial structuregeneration was about 900. When the initial structures were beinggenerated, assignments of additional NOEs were carried out in theusual iterative manner. Cross-peak intensities were classified asstrong~0.0–2.5 Å!, medium~0.0–3.5 Å!, weak ~0.0–4.5 Å!, andvery weak~0.0–5.5 Å!. Pseudoatom center averaging was appliedfor methyl groups and all distance constraints based on NOEsinvolving methyl groups had their upper limits increased by 1 Å.An r26 sum averaging, where ther26 distance was weighted bythe number of ambiguous NOEs~Nilges, 1995!, was applied for allnonstereospecifically assigned pairs of methyl groups, all nonste-reospecifically assigned protons in methylene groups and for theHd and HE protons in phenylalanine and tyrosine rings.
The number and distribution of NOEs used in the structuralcalculations are given in Table 1~and illustrated in Fig. 2 of Elec-tronic supplementary material!. The final restraint list contained2,070 NOEs. No intraresidue NOEs between atoms separated bythree bonds~e.g., HN-Ha, Ha-Hb! were included in the X-PLORcalculations because these~about 430 NOEs! had already beenused for defining the torsion angles in the AngleSearch calculations.
Torsion angle constraints and stereospecific assignments
The program AngleSearch~Polshakov et al., 1995b! was usedto determine thef, c, x1, andx2 torsional angle constraints andthe stereospecific assignments for all theb-methylene protonsand for valineg-methyl groups as described previously for theDHFR{methotrexate complex~Gargaro et al., 1998!. The errorlimits obtained from AngleSearch for the dihedral angles wereincreased to6308 for f, 6408 for c, and6308 and6408 for x1
andx2, respectively, to take into account local mobilities and theuncertainties in the Karplus equations used in these studies.
The ranges for the trimetrexate torsion anglest1 ~C5-C6-C9-N10! and t2 ~C6-C9-N10-C19! were determined from the An-gleSearch analysis of ROESY effects between protons of the drug.The final restraint list contained 345 dihedral angles~f, 123; c,120; x1, 96; x2, 4; trimetrexate, 2!.
Hydrogen bond constraints
Hydrogen bond constraints involving backbone NH protonswere included in the calculation only at the last stage of structurerefinement after they had been identified in the family of structuresobtained without their use. Donors of hydrogen bonds corre-sponding to slowly exchanging NH protons were identified fromanalysis of the rates of Hr D exchange for backbone amideprotons ~measured at 288 K from a series of HSQC spectra ofDHFR{trimetrexate complexes recorded between 5 and 880 minafter dissolving the sample in D2O as described in Electronic sup-plementary material!. Acceptors of hydrogen bonds were selectedif there were only one possible oxygen atom within 3 Å distanceof the corresponding amide proton in the family of structures and
each hydrogen bond was described with two restraints. The finalcalculations included 142 hydrogen bond restraints involving back-bone NH protons~corresponding to 71 hydrogen bonds!.
A hydrogen bond restraint corresponding to the T34g-hydroxylproton interacting with the carbonyl oxygen of F30 was includedin the structure calculation. A signal for the T34g-hydroxyl protonwas observed in all NMR spectra recorded in H2O and was alsoobserved for the DHFR{methotrexate complex~Gargaro et al.,1998!.
To specify the geometry of the interactions of the trimetrexateHN1 and HN21 protons with the D26b-carboxylate oxygens~Polshakov et al., 1995a!, two distance restraints and four addi-tional torsion angles were included in the final stage of structurecalculation to maintain the planarity of the 2-NH2 group and thepyrimidine ring.
The final constraint list contained a total of 144 hydrogen bondconstraints.
Structure calculations
The structure calculations were performed in an iterative fashionusing X-PLOR 3.1~Brünger, 1992!. Initially, a set of 100 struc-tures was generated using a combined distance geometry0simulatedannealing approach~Nilges et al., 1988, 1991!. The distance ge-ometry used subembedding based on the Ca, Ha, N, HN, O, C,and Cb atoms. The ligand was included from the beginning of thecalculations. Structures produced by this method were then regu-larized using simulated annealing consisting of 6 ps of dynamics at2,000 K with the force constants determining covalent geometryscaled during this stage to their final values, followed by coolingto 100 K with the vdW repulsive term increased from 0.003 to 0.4and the vdW radii decreased from 0.9 to 0.8 of their standardvalues and then subjected to 1,000 steps of conjugate gradientminimization. Fifty-four of the structures with essentially the cor-rect fold as judged by comparison with NMR structures of theDHFR{methotrexate complex~Gargaro et al., 1998! were acceptedfor further refinement. These structures were used to resolve am-biguities in the NOE data set and then subjected to a slow coolingsimulated annealing refinement~from 1000 to 100 K during a20 ps period!. This stage of the process was repeated several timesincluding progressively more NOE and dihedral angle constraints.Twenty-four structures with no NOE violations greater than 0.2 Åand no dihedral angle violations greater than 58 were acceptedfor the final refinement. At this stage, the hydrogen bonds andthe constraints modeling specific interactions between 2,4-diaminopyrimidyl moiety of trimetrexate and the carboxy group ofD26 were introduced. The final refinement protocol was essen-tially the same as the previous one followed by 1,000 steps ofconjugate gradient minimization and yielded 22 structures with nodistance violation greater then 0.1 Å and no dihedral angle viola-tions greater than 38. Calculations of the final family of 22 struc-tures ~shown in Fig. 5! were carried out with double the earliercooling MD period~40 ps!. Figure 8A shows the RMSD valuescalculated for the backbone atoms~Ca, C, N! using pairwise glo-bal fitting of these 22 structures. Bond lengths were constrainedusing SHAKE ~Ryckaert et al., 1977!. Parameter and topologyfiles for trimetrexate were constructed using coordinates from thecrystal structure of trimetrexate~Hempel & Camerman, 1988!.
Calculations were performed on Silicon Graphics~Power Indi-go2! and Sun~Sparc 10! computers. Coordinates for the 22 refinedstructures have been deposited in the Brookhaven Protein Data
478 V.I. Polshakov et al.

Bank~coordinates ID code1bzf; assignments and restraints ID coder1bzfmr!.
Analysis of calculated structures
Structures were visualized using INSIGHT II~Molecular Simula-tions Inc., Burlington, Massachusetts! and GRASP~Nicholls et al.,1991! and analyzed using X-PLOR 3.1~Brünger, 1992! andPROCHECK-NMR0AQUA ~Laskowski et al., 1993, 1996!. Theanalysis showed that there are no residues in the disallowed re-gions of the Ramachandran plot~Fig. 3 in Electronic supplemen-tary material!. A representative structureSrep was selected from theensemble of calculated structures as being the one that is closest toall the other structures and thus gives the lowest sum of pairwiseRMSD for the remainder of the structures in the family. RMSDparameters~RTMQ
i ! for both side chain and all heavy atoms of eachresiduei have been calculated together with their standard devia-tions. Similar calculations were performed for the family of 21calculated structures of the complex of MTX~Gargaro et al., 1998!.For comparison of solution structures of MTX and TMQ, similarparameters were also obtained using cross-pairwise superimposi-tion of each structure of MTX on each structure of TMQ in theirfamilies ~462 pairs!. For comparison of two families, a parameterr i 5 ~RTMQ0MTX
i !20~RTMQi {RMTX
i ! was calculated. This parameteris especially useful when one is comparing results for superimpo-sition on all heavy atoms. For example, residues with ill-definedside-chain conformations have large values ofRTMQ0MTX
i , RTMQi ,
and RMTXi , and the values ofr i will be closer to 1. However,
residues that have well-defined side chains within families but thathave large differences between two complexes will have largevalues ofr i ~see Fig. 8!. In Figure 8C, onlya-helix B ~betweenresidues 26 and 28! shows a significant local displacement be-tween the two DHFR complexes probably due to the interactionbetween H28 and the glutamate moiety of methotrexate, which isabsent in the trimetrexate complex.
T1, T2 and 15N{1H} NOE data analysis
The relaxation data were analyzed using the “model-free” ap-proach of Lipari and Szabo~1982! with the assumption of isotropictumbling. The spectra used in theT1, T2, and 15N$1H% NOE ex-periments were processed and analyzed with the FELIX program~Molecular Simulations Inc.!. The time domain data were zero-filled to 1K and 2K complex points in thet1 andt2 dimensions, anda Gaussian window function was applied in both dimensions be-fore transformation. TheT1 andT2 values~giving the ratesR1 andR2, respectively! and their standard deviations were obtained fromnonlinear fits of both integral peak volumes and the peak intensi-ties. Standard deviations of15N$1H% NOE values were calculatedusing the RMS noise of background regions~Farrow et al., 1994!.For most of the residues, the results were identical, but for theresidues with signals situated in more crowded regions the resultsobtained from peak intensities~rather than from volumes! hadsmaller standard deviations, and therefore only these were used forfurther analysis~results shown in Fig. 6!. T1 and T2 values wereobtained from a total of 128 residues.
The overall correlation timetc for the complex of DHFR withtrimetrexate was calculated from theT10T2 ratios~Kay et al., 1989!.An initial selection was made of residues that hadT1 andT2 valueswithin one standard deviation from the mean values over all res-idues~93 of 128 residues!. Of these, the 54 residues that hadT10T2
ratios within one standard deviation of the mean of the subset of 93were then selected for further analysis. The calculations yield anaveragetc value of 8.756 0.1 ns. The overall correlation time wastreated as a fixed parameter in the analysis of the relaxation data interms of the model-free formalism.
Calculation of the model-free parameters from the measuredrelaxation rate constants and the NOE enhancements was per-formed by Powell minimization~Press et al., 1992! of a globaltarget functionx2 summed over all residues using software writtenin-house~see Electronic supplementary material!. In this analysis,15N-1H dipolar relaxation and15N CSA relaxation were examinedusing equations in the literature~Kay et al., 1989; Farrow et al.,1994!. Values of 1.02 Å and 160 ppm were assumed for the N-Hbond-length and15N CSA, respectively. For some residues, anexchange termRex was included to represent line broadening dueto exchange processes on a timescale much slower than the overallrotational correlation time.
The relaxation data were fitted to three functional forms of themodel free spectral density functions given in the literature~Lipari& Szabo, 1982; Clore et al., 1990a, 1990b! and described in Elec-tronic supplementary material.
Supplementary material in Electronic Appendix
File: tmqsupp1.doc~The tables are in Word 6.0~RTF! with Cou-rier font; the text is in Word 6.0~RTF! Times New Roman font.!
File: Fig_tmq.zip contains Figure1.bmp~Fig. 1 Supplementary!,Figure2.bmp~Fig. 2 Supplementary!, and Figure3.bmp~Fig. 3Supplementary!.
File: Dhfr_tmq.kin contains two kinemage figures
NMR experimental details
Relaxation data analysis and parameters
Table 1.~Supplementary!. 1H and15N chemical shift assignmentsfor the DHFR{trimetrexate complex at 308 K.
Table 2.~Supplementary!. NOE, dihedral angle, and hydrogen bondrestraints used for the final X-PLOR structure calculations.
Table 3.~Supplementary!. Relaxation data and calculated param-eters for the complex of DHFR and trimetrexate.
Fig. 1.~Supplementary!. Plots of relaxation data and parameters asa function of residue number.
Fig. 2. ~Supplementary!. Plot of the number and distribution ofNOEs against the amino acid sequence.
Fig. 3. ~Supplementary!. A Ramachandran plot of the torsion an-gles for all residues of the 22 final structures.
Kinemage 1. Ribbon representation of DHFR in its complex withtrimetrexate.
Kinemage 2. Comparison of three representative structures of bi-nary complexes of DHFR-with trimetrexate, methotrexate, andtrimethoprim.
Acknowledgments
V.I.P. acknowledges the award of a Howard Hughes Medical InstituteInternational Scholarship and a grant from the Russian Foundation for
Structure and dynamics in solution of the complex of L. casei 479

Basic Research~97-04-48245!. We thank Warner-Lambert0Parke-Davis~Pontypool, Gwent! for their gift of trimetrexate and are grateful to J.E.McCormick and G. Ostler for expert technical assistance and to A. Lane,M.A. Williams, and W.D. Morgan for helpful discussions.
References
Andrews J, Clore GM, Davies RW, Gronenborn AM, Gronenborn B, KalderonD, Papadopoulos PC, Schafer S, Sims PFG, Stancombe R. 1985. Nucleotidesequence of the dihydrofolate reductase gene of methotrexate-resistantLactobacillus casei. Gene 35:217–222.
Antonjuk DJ, Birdsall B, Burgen ASV, Cheung HTA, Clore GM, Feeney J,Gronenborn A, Roberts GCK, Tran W. 1984. A1H NMR study of the roleof the glutamate moiety in the binding of methotrexate to dihydrofolatereductase.Brit J Pharmacol 81:309–315.
Baccanari DP, Kuyper LF. 1993. Basis of selectivity of antibacterial diaminopy-rimidines.J Chemother 5:393–399.
Bertino JR, Sawicki WL, Moroson BA, Cashmore AR, Elslager EF. 1979.24-Diamino-5-methyl-6-@3,4,5#-trimethoxyanilino-methyl#quinazoline tri-metrexate a potent non-classical folate antagonist inhibitor: I. Effect ofdihydrofolate reductase and growth of rodent tumoursin vitro and in vivo.Biochemical Pharmacol 28:1983–1987.
Bevan AW, Roberts GCK, Feeney J, Kuyper L. 1985.1H and15N NMR studiesof protonation and hydrogen-bonding in the binding of trimethoprim todihydrofolate reductase.Eur Biophys J 11:211–218.
Birdsall B, Andrews J, Ostler G, Tendler SJB, Feeney J, Roberts GCK, DaviesRW, Cheung HTA. 1989b. NMR studies of the differences in the confor-mations and dynamics of ligand complexes formed with mutant dihydrofo-late reductases.Biochemistry 28:1353–1362.
Birdsall B, Burgen ASV, Hyde EI, Roberts GCK, Feeney J. 1981. Negativecooperativity between folinic acid and coenzyme in their binding toL. caseidihydrofolate reductase.Biochemistry 20:186–7195.
Birdsall B, Burgen ASV, Roberts GCK. 1980. Binding of coenzyme analoguesto L. caseidihydrofolate reductase: Binary and ternary complexes.Biochem-istry 19:3723–3731.
Birdsall B, Cassarotto MG, Cheung HTA, Basran J, Roberts GCK, Feeney J.1997. The influence of aspartate 26 on the tautomeric forms of folate boundto Lactobacillus caseidihydrofolate reductase.FEBS Lett 402:157–161.
Birdsall B, Feeney J, Pascual C, Roberts GCK, Kompis I, Then RL, Muller K,Kroehn A. 1984. A1H study of the interactions and conformations of ra-tionally designed brodimoprim analogues in complexes withLactobacilluscaseidihydrofolate reductase.J Med Chem 23:1672–1676.
Birdsall B, Feeney J, Tendler SJB, Hammond SJ, Roberts GCK. 1989a. Dihy-drofolate reductase: Multiple conformations and alternative modes of sub-strate binding.Biochemistry 28:2297–2305.
Birdsall B, Gronenborn AM, Hyde EI, Clore GM, Roberts GCK, Feeney J,Burgen ASV. 1982. Hydrogen-1, carbon-13 and phosphorus-31 nuclear mag-netic resonance studies of the dihydrofolate reductase-nicotinamide adeninedinucleotide phosphate-folate complex: Characterization of three coexistingconformational states.Biochemistry 21:5831–5838.
Birdsall B, Tendler SJB, Arnold JRP, Feeney J, Griffin RJ, Carr MD, ThomasJA, Roberts GCK, Stevens MFG. 1990. NMR studies of multiple confor-mations in complexes ofL. caseidihydrofolate reductase with analogues ofpyrimethamine.Biochemistry 29:9660–9667.
Blakley RL. 1985.Dihydrofolate reductase in folates and pterins, Vol 1, Chapter5. In: Blakley RL, Benkovic SJ, eds. New York, NY: J. Wiley. pp 191–253.
Bolin JT, Filman DJ, Matthews DA, Hamlin RC, Kraut J. 1982. Crystal struc-tures ofE. coli andL. caseidihydrofolate reductase refined to 1.7 Å reso-lution: I. General features and binding of methotrexate.J Biol Chem257:13650–13662.
Brünger AT. 1992. X-PLOR 3.1: A system for X-ray crystallography and NMR.New Haven, CT: Yale University Press.
Carr MD, Birdsall B, Frenkiel TA, Bauer CJ, Jimenez-Barbero J, Polshakov VI,McCormick JE, Feeney J, Roberts GCK. 1991. Dihydrofolate reductase:Sequential resonance assignments using 2D and 3D NMR and secondarystructure determination in solution.Biochemistry 30:6330–6341.
Cheung HTA, Birdsall B, Frenkiel TA, Chau DD, Feeney J. 1993.13C NMRdetermination of the tautomeric and ionization states of folate in its com-plexes with Lactobacillus caseidihydrofolate reductase.Biochemistry32:6846–6854.
Clore GM, Driscoll PC, Wingfield PT, Gronenborn AM. 1990a. Analysis of thebackbone dynamics of interleukin-1b using inverse detected heteronuclear15N-1H NMR spectroscopy.Biochemistry 29:7387–7401.
Clore GM, Gronenborn AM, Birdsall B, Feeney J, Roberts GCK. 1984.19F-NMR studies of 39,59-difluoromethotrexate binding toLactobacillus caseidihydrofolate reductase. Molecular motion and coenzyme-induced confor-mational changes.Biochem J 217:659–666.
Clore GM, Szabo A, Bax A, Kay LE, Driscoll PC, Gronenborn AM. 1990b.Deviations from the simple two-parameter model-free approach to the in-terpretation of nitrogen-15 nuclear magnetic relaxation in proteins.J AmChem Soc 112:4989–4991.
Cocco L, Roth B, Temple C, Montgomery JA, London RE, Blakley RL. 1981.Protonated state of methotrexate trimethoprim and pyrimethamine bound todihydrofolate reductase.Arch Biochem Biophys 226:567–577.
Dann JG, Ostler G, Bjur RA, King RW, Scudder P, Turner PC, Roberts GCK,Burgen ASV, Harding NGL. 1976. Large scale purification and character-ization of dihydrofolate reductase from a methotrexate-resistant strain ofLactobacillus casei. Biochem J 157:559–571.
Elslager EF, Johnson JL, Werbel LM. 1983. Folate antagonists. 20. Synthesisand anti-tumor and anti-malarial properties of trimetrexate and related6-@~phenylamino!methyl#-2,4-quinazolinediamines.J Med Chem 26:1753–1760.
Epstein DM, Benkovic SJ, Wright PE. 1995. Dynamics of the dihydrofolatereductase folate complex—Catalytic sites and regions known to undergoconformational change exhibit diverse dynamical features.Biochemistry34:11037–11048.
Falzone CJ, Cavanaugh J, Cowart M, Palmer AG III, Matthews CR, BenkovicSJ, Wright PE. 1994.1H, 15N and 13C resonance assignments secondarystructure and the conformation of substrate in the binary folate complex ofEscherichia colidihydrofolate reductase.J Biomol NMR 4:349–366.
Farrow FA, Muhandiram R, Singer AU, Pascal SM, Kay CM, Gish G, ShoelsonSE, Pawson T, Forman-Kay JD, Kay LE. 1994. Backbone dynamics of afree and phosphopeptide-complexed Src homology 2 domain studied by15NNMR relaxation.Biochemistry 33:5984–6003.
Feeney J. 1986. NMR studies of drug receptor complexes: Antifolate drugsbinding to dihydrofolate reductase.NATO Advanced Study Inst Ser C 164:347–366.
Feeney J. 1990. NMR studies of interactions of ligands with dihydrofolatereductase. Biochem Pharm 40:141–152.
Feeney J. 1996. NMR studies of ligand binding to dihydrofolate reductase andtheir application in drug design. In: Craik D, ed.NMR in drug design. BocaRaton, New York, London, Tokyo: CRC Press.
Feeney J, Birdsall B. 1993. NMR studies of protein-ligand interactions. In:Roberts GCK, ed.NMR of biological macromolecules: A practical ap-proach. Oxford, New York, Tokyo: Oxford University Press.
Freisheim JH, Matthews DA. 1984. In: Sirotnak FM, Burchill JJ, EnsmingerWD, Montgomery JA, eds.Folate antagonists as therapeutic agents, Vol 1.New York: Academic Press Inc. pp 69–131.
Gangjee A, Elzain E, Kothare M, Vasudevan A. 1996. Classical and nonclassicalantifolates as a potential antitumor antipneumocystis and antitoxoplasmaagents.Current Pharmaceutical Design 2:263–280.
Gangjee A, Zaveri N, Kothare M, Queener SF. 1995. Nonclassical 2,4-diamino-6-aminomethyl-5,6,7,8-tetrahydroquinazoline antifolates: Syntheses and bio-logical activities.J Med Chem 38:3660–3668.
Gargaro AR, Frenkiel TA, Nieto PM, Birdsall B, Polshakov VI, Morgan WD,Feeney J. 1996. NMR detection of arginine-ligand interactions in complexesof Lactobacillus caseidihydrofolate reductase.Euro J Biochem 238:435–439.
Gargaro AR, Soteriou A, Frenkiel TA, Bauer CJ, Birdsall B, Polshakov VI,Barsukov IL, Roberts GCK, Feeney J. 1998. The solution structure of thecomplex ofLactobacillus caseidihydrofolate reductase with methotrexate.J Mol Biol 277:119–134.
Gerothanassis IP, Barrie PJ, Birdsall B, Feeney J. 1996.31P solid-state NMRmeasurements used to detect interactions between NADPH and water and todetermine the ionization state of NADPH in a protein-ligand complex sub-jected to low-level hydration.Euro J Biochem 235:262–266.
Gronenborn A, Birdsall B, Hyde EI, Roberts GCK, Feeney J, Burgen ASV.1981. Direct observation by NMR of two coexisting conformations of anenzyme-ligand complex in solution.Nature 290:273–274.
Hammond SJ, Birdsall B, Searle MS, Roberts GCK, Feeney J. 1986. Dihydro-folate reductase1H resonance assignments and coenzyme-induced confor-mational changes.J Mol Biol 188:81–97.
Hempel A, Camerman N. 1988. Trimetrexate: Molecular structures and confor-mational similarities in two crystal forms.Cancer Biochem Biophys 10:25–30.
Jackson RC, Fry DW, Boritzki TJ, Besserer JA, Leopold WR, Sloan BJ, ElslagerEF. 1984. Biochemical pharmacology of the lipophilic antifolate trimetrex-ate.Advances in Enzyme Regulation 22:187–206.
Johnson JM, Meiering EM, Wright JE, Pardo J, Rosowsky A, Wagner G. 1997.NMR solution structure of the anti tumor compound PT523 and NADPH inthe ternary complex with human dihydrofolate reductase.Biochemistry36:4399–4411.
Kay LE, Nicholson LK, Delaglio F, Bax A, Torchia DA. 1992. The effects ofcross-correlation between dipolar and chemical shift anisotropy relaxationmechanism on the measurement of T1 and T2 values in proteins: Pulsesequences for the removal of such effects.J Magn Reson 97:359–375.
480 V.I. Polshakov et al.

Kay LE, Torchia DA, Bax A. 1989. Backbone dynamics of proteins as studiedby 15N inverse detected heteronuclear NMR spectroscopy: Application tostaphylococcal nuclease.Biochemistry 28:8972–8979.
Laskowski RA, MacArthur MW, Moss DS, Thornton JM. 1993. PROCHECK:A program to check the stereochemical quality of protein structures.J ApplCryst 26:283–291.
Laskowski RA, Rullmann JAC, MacArthur MW, Kaptein R, Thornton JM.1996. AQUA and PROCHECK-NMR: Programs for checking the quality ofprotein structures solved by NMR.J Biomol NMR 8:477–486.
Lin JT, Bertino JR. 1987. Trimetrexate—A 2nd generation folate antagonist inclinical-trial. J Clin Oncol 5:2032–2040.
Lin JT, Bertino JR. 1991. Update on trimetrexate a folate antagonist with anti-neoplastic and antiprotozal properties.Cancer Invest 9:159–172.
Lipari G, Szabo A. 1982. Model-free approach to the interpretation of nuclearmagnetic resonance relaxation in macromolecules: 1. Theory and range ofvalidity. J Am Chem Soc 104:4546–4559.
Live DH, Davis DG, Agosta WC, Cowburn D. 1984. Observation of 1000-foldenhancement of15N NMR via proton detected multiquantum coherences:Studies of large peptides.J Am Chem Soc 106:1939–1941.
Martorell G, Gradwell MJ, Birdsall B, Bauer CJ, Frenkiel TA, Cheung HTA,Polshakov VI, Kuyper L, Feeney J. 1994. Solution structure of bound tri-methoprim in its complex withLactobacillus caseidihydrofolate reductase.Biochemistry 33:12416–12426.
Matthews DA, Bolin JT, Burridge JM, Filman DJ, Volz KW, Kaufman BT,Beddell CR, Champness JN, Stammers DK, Kraut J. 1985a. Refined crystalstructures ofEscherichia coliand chicken liver dihydrofolate reductasecontaining bound trimethoprim.J Biol Chem 260:381–391.
Matthews DA, Bolin JT, Burridge JM, Filman DJ, Volz KW, Kraut J. 1985b.Dihydrofolate reductase—The stereochemistry of inhibitor selectivity.J BiolChem 260:392–399.
Morgan WD, Birdsall B, Polshakov VI, Sali D, Kompis I, Feeney J. 1995.Solution structure of a brodimoprim analogue in its complex withLacto-bacillus caseidihydrofolate reductase.Biochemistry 34:11690–11702.
Nicholls A, Sharp KA, Honig B. 1991. Protein folding and association: Insightsfrom the interfacial and thermodynamic properties of hydrocarbons.Pro-teins Struct Funct Genet 11:281–296.
Nieto PM, Birdsall B, Morgan WD, Frenkiel TA, Gargaro AR, Feeney J. 1997.Correlated bond rotations in interactions of arginine residues with ligandcarboxylate groups in protein ligand complexes.FEBS Lett 405:16–20.
Nilges M. 1995. Calculation of protein structures with ambiguous distanceconstraints: Automated assignment of ambiguous NOE crosspeaks and di-sulphide connectivities. J Mol Biol 245:645–660.
Nilges M, Gronenborn AM, Brünger AT, Clore GM. 1988. The determination ofthree-dimensional structures of proteins by simulated annealing with inter-proton distance restraints—Application to crambin potato carboxypeptidaseinhibitor and barley serine proteinase inhibitor-2.Protein Eng 2:27–38.
Nilges M, Kuszewski J, Brünger AT. 1991. In: Hoch JC, ed.Computationalaspects of the study of biological macromolecules by NMR. New York:Plenum Press.
Polshakov VI, Birdsall B, Gradwell MJ, Feeney J. 1995a. The use of PM3 SCFMO quantum mechanical calculations to refine NMR-determined structuresof complexes of antifolate drugs with dihydrofolate reductase in solution.JMol Struct (Theochem) 357:207–216.
Polshakov VI, Frenkiel TA, Birdsall B, Soteriou A, Feeney J. 1995b. Determi-nation of stereospecific assignments torsion-angle constraints and rotamerpopulations in proteins using the program AngleSearch.J Magn ResonSeries B 108:31–43.
Press WH, Teukolsky SA, Vetterling WT, Flannery BP. 1992.Numerical recipesin FORTRAN. The art of scientific computing, Second ed. Cambridge, UK:Cambridge University Press. pp 409–413; 564–570.
Robert F. 1988. Trimetrexate as a single agent in patients with advanced headand neck-cancer.Seminars in Oncol 15:22–26.
Roberts GCK. 1983. The interaction of substrates and inhibitors with dihydro-folate reductase. In: Blair JA, ed.Chemistry and biology of pteridines.Berlin: W. de Gruyter. pp 197–214.
Roth B, Cheng CC. 1982. Recent progress in the medicinal chemistry of 2,4-diaminopyrimidines.Prog Med Chem 19:1–58.
Ryckaert JP, Ciccotti G, Berendsen HJC. 1977. Numerical-integration of carte-sian equations of motion of a system with constraints: Molecular dynamicsof N-alkanes.J Comput Phys 23:327–341.
Sawaya MR, Kraut J. 1997. Loop and subdomain movements in the mechanismof E. coli dihydrofolate reductase: Crystallographic evidence.Biochemistry36:586–603.
Searle MS, Forster MJ, Birdsall B, Roberts GCK, Feeney J, Cheung HTA,Kompis I, Geddes AJ. 1988. Dynamics of trimethoprim bound to dihydro-folate reductase.Proc Nat Acad Sci USA 85:3787–3791.
Soteriou A, Carr MD, Frenkiel TA, McCormick JE, Bauer CJ, Sali D, BirdsallB, Feeney J. 1993. 3D13C01H NMR-based assignments for side-chainresonances ofLactobacillus caseidihydrofolate reductase. Evidence forsimilarities between the solution and crystal structures of the enzyme.J Biomol NMR 3:535–546.
Wishart DS, Bigam CG, Yao J, Abildgaard F, Dyson HJ, Oldfield E, Markley JL,Sykes BD. 1996.1H, 13C and15N chemical shift referencing in biomolec-ular NMR. J Biomol NMR 6:135–140.
Structure and dynamics in solution of the complex of L. casei 481
Related Documents











