Journal of Fish Biology (2001) 58, 305–324 doi:10.1006/jfbi.2000.1450, available online at http://www.idealibrary.com on REGULAR PAPERS Steroid hormone profiles and correlative gonadal histological changes during natural sex reversal of sobaity kept in tanks and sea-cages K. P. L*, S. A-A A. A-Y Mariculture and Fisheries Department, Kuwait Institute for Scientific Research, P.O. Box 1638, Salmiya, Kuwait 22017 (Received 17 March 2000, Accepted 1 September 2000) The gonadosomatic index of sobaity Sparidentex hasta fits well with the natural spawning season of February–March. At 12 and 24 months all the population matured as males. However, during this time, between April–September intersex gonads were seen. From October of the third year of life, females were seen for the first time. These changes in the histology of the gonads were correlated with the hormonal profile. Testosterone showed no significant correlation with sex but 11-ketotestosterone correlated very well with the mature male gonads and thus can be used for sexing. Oestradiol-17 was higher in sex reversed individuals (females) in the spawning season than in males. The sequence of events in the protandrous sex change of this fish during the first two spawning seasons is maleintersexmale. However, during the third spawning season (age 36–37 months) this sequence is maleintersexmale or female (10–17%). Key words: protandric hermaphroditism; histology; oestradiol-17; testosterone; 11- ketotestosterone; Sparidentex. INTRODUCTION Yamamoto (1969) suggested that sex steroids are the natural inducers of gonadal differentiation in teleost fish. However, very little is known about the role of sex steroids in natural sex differentiation and sex control of hermaphroditic fish. In a study on the steroids of four hermaphrodite species, Idler et al. (1976) showed that 11-hydroxytestosterone (11-HT) was the predominant steroid in the plasma, whereas 11-ketotestosterone (11-KT) was not detected. In the same study, gonochoristic species had 11-KT as the predominant steroid, and 11-HT was extremely low. In another hermaphrodite, Spicara maena (L.), 11-HT was higher than 11-KT in both testicular and ovarian tissues. In the protogynous Coris julis L., conversion of testosterone (T) to 11-HT was more active in secondary male and sex-reversing specimens. Primary males had a pattern similar to females, as in Serranus cabrilla (L.) (Reinboth, 1979). In Rhabdosargus sarba (Forsska ˚l), a protandrous hermaphrodite, the plasma steroid pattern was similar in male and intersex individuals. However, the intersex individuals had higher levels of conjugated oestradiol-17 (E 2 ), conjugated androstenedione *Author to whom correspondence should be addressed. Tel.: (+965) 5711297; fax: (+965) 5711293; email: [email protected] 305 0022–1112/01/020305+20 $35.00/0

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Fish Biology (2001) 58, 305–324doi:10.1006/jfbi.2000.1450, available online at http://www.idealibrary.com on
REGULAR PAPERS
Steroid hormone profiles and correlative gonadal histologicalchanges during natural sex reversal of sobaity kept in tanks
and sea-cages
K. P. L*, S. A-A A. A-Y
Mariculture and Fisheries Department, Kuwait Institute for Scientific Research,P.O. Box 1638, Salmiya, Kuwait 22017
(Received 17 March 2000, Accepted 1 September 2000)
The gonadosomatic index of sobaity Sparidentex hasta fits well with the natural spawningseason of February–March. At 12 and 24 months all the population matured as males.However, during this time, between April–September intersex gonads were seen. From Octoberof the third year of life, females were seen for the first time. These changes in the histology ofthe gonads were correlated with the hormonal profile. Testosterone showed no significantcorrelation with sex but 11-ketotestosterone correlated very well with the mature male gonadsand thus can be used for sexing. Oestradiol-17� was higher in sex reversed individuals (females)in the spawning season than in males. The sequence of events in the protandrous sex change ofthis fish during the first two spawning seasons is male�intersex�male. However, during thethird spawning season (age 36–37 months) this sequence is male�intersex�male or female(10–17%).
Key words: protandric hermaphroditism; histology; oestradiol-17�; testosterone; 11-ketotestosterone; Sparidentex.
*Author to whom correspondence should be addressed. Tel.: (+965) 5711297; fax: (+965) 5711293;email: [email protected]
INTRODUCTION
Yamamoto (1969) suggested that sex steroids are the natural inducers of gonadaldifferentiation in teleost fish. However, very little is known about the role of sexsteroids in natural sex differentiation and sex control of hermaphroditic fish. Ina study on the steroids of four hermaphrodite species, Idler et al. (1976) showedthat 11�-hydroxytestosterone (11�-HT) was the predominant steroid in theplasma, whereas 11-ketotestosterone (11-KT) was not detected. In the samestudy, gonochoristic species had 11-KT as the predominant steroid, and 11�-HTwas extremely low. In another hermaphrodite, Spicara maena (L.), 11�-HT washigher than 11-KT in both testicular and ovarian tissues. In the protogynousCoris julis L., conversion of testosterone (T) to 11�-HT was more active insecondary male and sex-reversing specimens. Primary males had a patternsimilar to females, as in Serranus cabrilla (L.) (Reinboth, 1979). In Rhabdosargussarba (Forsskal), a protandrous hermaphrodite, the plasma steroid pattern wassimilar in male and intersex individuals. However, the intersex individuals hadhigher levels of conjugated oestradiol-17� (E2), conjugated androstenedione
305
0022–1112/01/020305+20 $35.00/0

306 . . .
(AD) and free T than did the females, even during the spawning period. Thismay indicate that, to have a sex change from male to female, the oestrogen levelshave to be higher to suppress the antagonistic effects of androgens (Yeung &Chan, 1985, 1987a).
Some sex-changing fishes display the normal teleost pattern of steroid levels,with higher levels of serum 11-KT in males than in females (Nakamura et al.,1989, 1994; Cardwell & Liley, 1991a,b; Cochran & Grier, 1991; Kime et al., 1991;Godwin & Thomas, 1993). Also, in some fish, there is higher gonadalbiosynthesis of 11-oxygenated steroids in males than in females (Reinboth &Becker, 1984; Hourigan et al., 1991; Latz et al., 1991; Yeung et al., 1993). On theother hand, there are many sex-reversing fishes with very low or non-detectablecirculating 11-KT levels (Yeung & Chan, 1987a, b; Chan & Yeung, 1989;Guiguen et al., 1993).
In some cases, androgen administration can masculinize protogynous fishes.Testosterone derivatives induce male secondary sexual characters and at leastpartial masculinization of gonads in the protogynous wrasses C. julis, Thalas-soma bifasciatum (Bloch) and Halichoeres bivittatus (Bloch) (Reinboth, 1975).11-KT stimulated spermatogenesis and a change to male colouration whenadministered to female stoplight parrotfish Sparisoma viride (Bonnaterre)(Cardwell & Liley, 1991a). Sex change has also been induced in a protandrousporgy Acanthopagrus schlegeli Bleeker by feeding E2 (Chang et al., 1994, 1995a,b; Chang & Lin, 1998).
Sex steroids have significant effects on gonadal development. In thisconnection, E2, T and 11-KT have received most attention (Fostier et al., 1983;Borg, 1994). In sobaity Sparidentex hasta Valenciennes, Kime et al. (1991)showed that during sexual maturation E2 and 11-KT can be used to sex the fishwhile T did not relate to any sex. Very little is known about the role of sexsteroids and their relation to the histology of the natural sex change in sobaity.Therefore these topics were examined during the natural sex reversal of thisspecies in two different environments, tanks and sea cages.
MATERIALS AND METHODS
HISTOLOGY OF GONADSAround 500 sobaity from the 1996 spawning season (8 months old; weight c. 200 g)
were stocked in a net cage (2·5�2·5�2·5 m deep) at the Doha sea cage site. The samenumber of fish were also kept in tanks (30 t capacity) at the Mariculture and FisheriesDepartment’s Salmiya Fish Hatchery site of the Kuwait Institute for Scientific Research.Fish were fed to satiation with sea-bream pellets (Aqualim: 46% crude protein, 15% crudelipids) once a day at both sites. From January 1997, monthly samples of 5–10 fish weretaken randomly from both sites to study gonadal changes with time and growth of thefish. At sampling, the fish were anaesthetized deeply with quinaldine (4–5 mg l�1),weighed to the nearest 0·01 g, measured to the nearest mm and dissected to removegonads, which were weighed immediately to the nearest mg and fixed in Bouin’s fluid or10% neutral buffered formalin. Thin paraffin sections were cut at 3–5 �m and stainedwith Harris haematoxylin and eosin. At the end of the experiment, at the age of36 months in February 1999, all the fish in the cages and in the tanks were killed todetermine the final sex of the fish. The length, body weight and gonad weights were alsotaken at this time.
307
The gonadosomatic index (IG) was computed as IG=100WG . W�1 where WG isweight of the gonad and W is weight of the fish.
MEASUREMENTS OF SEX HORMONES IN BLOODFor measuring hormones in blood, 60 fish at each site (cage and tanks) were tagged
individually. Each month, when these fish were measured and weighed, blood sampleswere taken from all the marked fish. The blood (1 ml) was drawn into a syringe from thecaudal vein and kept on ice until all the required fish had been sampled. The sampleswere spun in a refrigerated centrifuge at 4000 g to separate the serum, which was frozenin Eppendorf microfuge tubes at �70� C.
Serum steroid hormones, i.e. T, 11-KT and E2 were determined by an ELISA methodusing acetylcholinesterase as a label. Normally, 5 ml of methylene chloride was added to250 �l of the thawed serum in a glass-stoppered conical base test tube. After vigorousshaking and vortexing, the lower methylene chloride layer was removed carefully. Thisprocedure was repeated once more and both extracts were mixed, poured into a glass vialand the solvent allowed to evaporate overnight. The residue was reconstituted in 400 �lof absolute ethanol, pipetted into an uncoated microtitre plate and the alcohol allowed toevaporate. The plates were sealed with a plate sealer and stored at �20� C.
The 11-KT was determined according to the method of Cuisset et al. (1994), while Tand E2 were determined according to Nash et al. (2000). The details of the validation andcross-reactivities are described in these papers. For each group of samples run on a plate,a separate standard curve was also run. IG-values were compared according to thetwo-tailed Student’s t test, with a level of significance of 0·05. The numbers of femalesencountered at the end of the experiment at cage and tank sites were compared accordingto the Pearson’s �2 test with the Yates’s continuity correction. The hormone data werecompared across sites and months according to the General Linear Model MANOVA.Site and months were used as fixed factors. Type III sums of squares were used becausesample size varied between sites, months, and sexes. Tukey’s post hoc test was used withSPSS (Norusis, 1996).
5F
50
1997
Mea
n s
ea w
ater
tem
pera
ture
(°C
)
J0
16
Day
len
gth
(h
)
14
12
10
8
6
4
2
45
40
35
30
25
20
15
10
F M A M J J A S O N D J F M A M J J A S O N D J
Month1998 1999
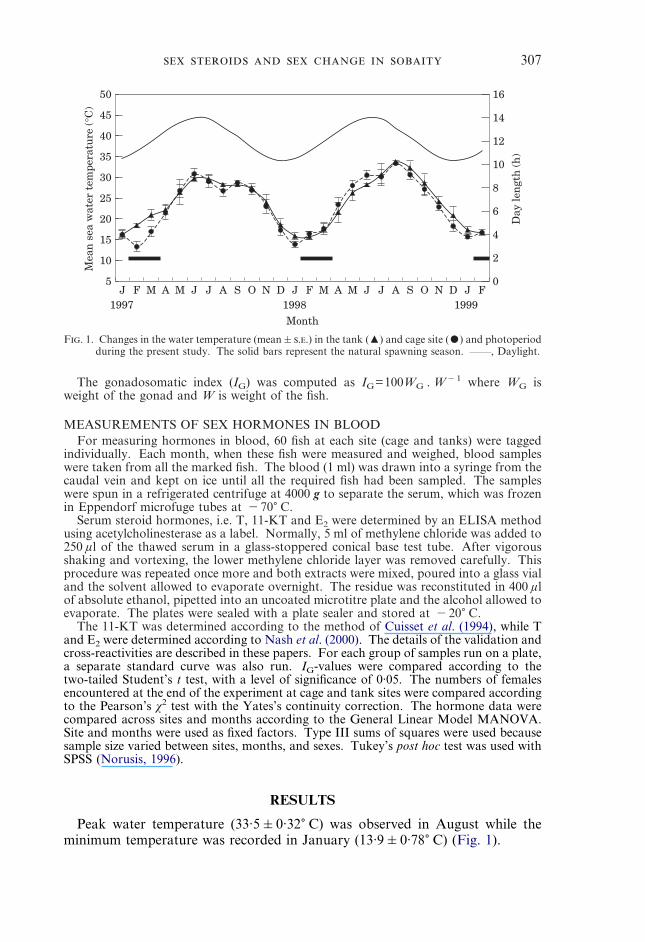
F. 1. Changes in the water temperature (mean�..) in the tank (�) and cage site (�) and photoperiodduring the present study. The solid bars represent the natural spawning season. ——, Daylight.
RESULTS
Peak water temperature (33·5�0·32� C) was observed in August while theminimum temperature was recorded in January (13·9�0·78� C) (Fig. 1).
T
I.T
otal
leng
than
dbo
dyw
eigh
t(m
ean�
.
.)of
indi
vidu
ally
tagg
edso
bait
yke
ptin
tank
san
dca
ges
duri
ngth
eag
eof
11–3
6m
onth
s.T
hem
easu
rem
ents
wer
eta
ken
ever
ym
onth
whe
nbl
ood
was
draw
nfo
rho
rmon
ean
alys
is.
For
the
sake
ofbr
evit
y,da
taar
epr
esen
ted
ona
bi-m
onth
lyba
sis.
Fro
mag
e11
to31
mon
ths
nopu
refe
mal
eco
uld
bese
enhi
stol
ogic
ally
,an
dth
eref
ore,
leng
than
dw
eigh
tof
tota
lex
peri
men
tal
popu
lati
onar
egi
ven
for
thes
em
onth
s
Age
(mon
ths)
Mon
th
Tan
kC
age
Len
gth
(cm
)W
eigh
t(g
)L
engt
h(c
m)
Wei
ght
(g)
Mal
eF
emal
eM
ale
Fem
ale
Mal
eF
emal
eM
ale
Fem
ale
11Ja
n.(1
987)
26·4
1—
338·
62—
25·4
5—
306·
89—
�1·
21�
53·7
5�
0·74
�28
·83
13M
ar.
26·5
3—
338·
97—
26·0
1—
295·
68—
�1·
19�
57·4
4�
0·75
�53
·71
15M
ay28
·51
—40
2·90
—27
·62
—34
6·51
—�
1·25
�55
·36
�1·
08�
36·4
117
Jul.
31·6
2—
540·
00—
30·0
0—
443·
00—
�1·
38�
65·4
7�
1·32
�53
·37
19Se
p.35
·53
—69
0·80
—32
·28
—56
8·57
—�
2·09
�82
·88
�1·
33�
62·8
921
Nov
.38
·09
—94
3·41
—34
·73
—72
0·87
—�
1·57
�11
4·60
�1·
39�
84·4
623
Jan.
(199
8)38
·39
—10
30·7
8—
35·5
4—
761·
20—
�2·
30�
129·
03�
1·43
�95
·11
25M
ar.
38·5
8—
1040
·18
—35
·85
—73
7·46
—�
2·01
�12
9·87
�1·
42�
84·6
427
May
40·0
3—
1224
·90
—36
·93
—80
7·25
—�
1·93
�16
3·59
�1·
73�
110·
6329
Jul.
41·8
7—
1410
·93
—38
·06
—93
6·53
—�
2·32
�23
5·72
�3·
16�
171·
3431
Sep.
43·4
0—
1651
·16
—40
·32
—11
74·1
4—
�2·
32�
243·
16�
3·17
�15
7·83
33N
ov.
47·0
946
·92
2093
·59
2087
·55
43·2
242
·51
1609
·76
1632
·71
�3·
78�
4·45
�32
5·15
�36
2·16
�2·
59�
2·71
�25
8·75
�26
7·34
35Ja
n.(1
999)
47·6
547
·84
2097
·07
2280
·20
43·7
542
·80
1509
·84
1589
·96
�2·
96�
4·95
�38
7·41
�68
0·08
�2·
69�
2·34
�27
5·17
�22
0·47
36F
eb.
47·3
247
·54
2039
·23
1953
·80
43·7
642
·90
1415
·36
1420
·60
�2·
98�
4·83
�37
6·61
�50
7·22
�2·
49�
3·11
�27
1·59
�29
4·77
309
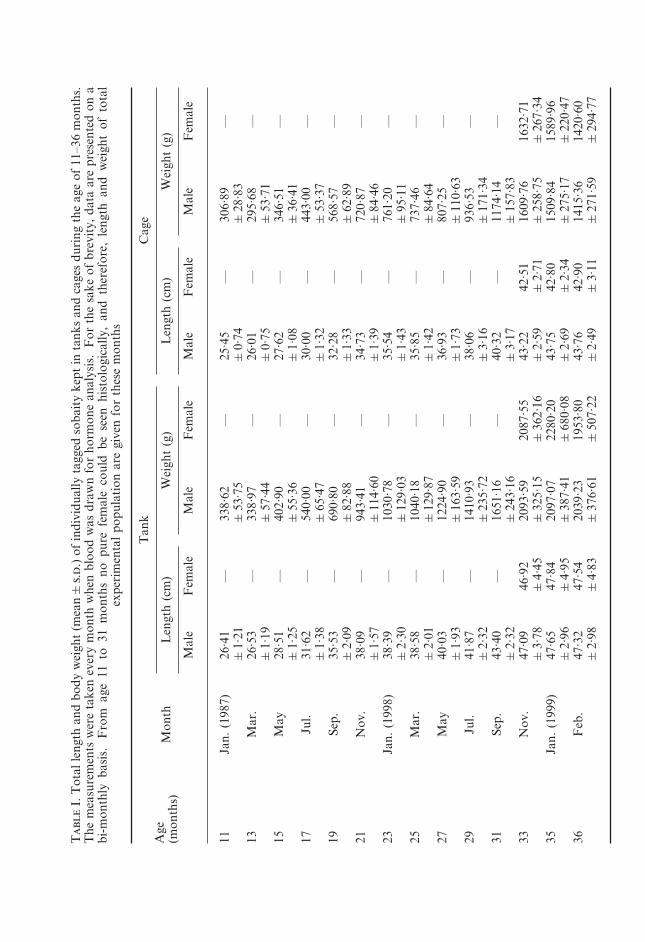
For the first 21 months of the experiment no true female was seen. Therewas no difference in weight and length of males and females for the same site(Table I). However, tank males were heavier and longer (P<0·001) than the cagemales. This difference between females was not significant (P>0·05). Thisprobably was due to the low number of females (5–7) and more variations intheir measurements.
–1.0F
9.0
1997
Gon
ados
omat
ic in
dex
(%)
J F M A M J J A S O N D J F M A M J J A S O N D J
Month1998 1999
8.0
7.0
6.0
5.0
4.0
3.0
2.0
1.0
0.0
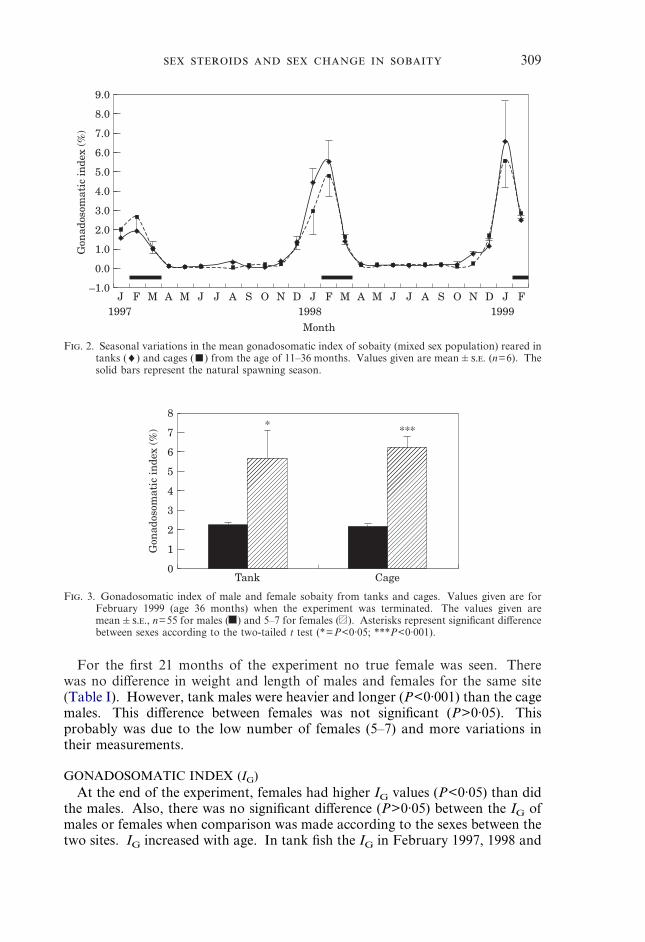
F. 2. Seasonal variations in the mean gonadosomatic index of sobaity (mixed sex population) reared intanks (♦) and cages (�) from the age of 11–36 months. Values given are mean�.. (n=6). Thesolid bars represent the natural spawning season.
0
8
Gon
ados
omat
ic in
dex
(%)
Tank
7
6
5
4
3
2
1
Cage
* ***
F. 3. Gonadosomatic index of male and female sobaity from tanks and cages. Values given are forFebruary 1999 (age 36 months) when the experiment was terminated. The values given aremean�.., n=55 for males ( ) and 5–7 for females ( ). Asterisks represent significant differencebetween sexes according to the two-tailed t test (*=P<0·05; ***P<0·001).
GONADOSOMATIC INDEX (IG)At the end of the experiment, females had higher IG values (P<0·05) than did
the males. Also, there was no significant difference (P>0·05) between the IG ofmales or females when comparison was made according to the sexes between thetwo sites. IG increased with age. In tank fish the IG in February 1997, 1998 and
310 . . .

311
January 1999 was 1·94�0·70, 5·57�1·12 and 6·63�2·08, respectively. Similardata were obtained for the caged fish. The IG dropped suddenly in April at theend of the spawning season (Fig. 2). In February 1999 IG of females wassignificantly higher than that of males (Fig. 3).
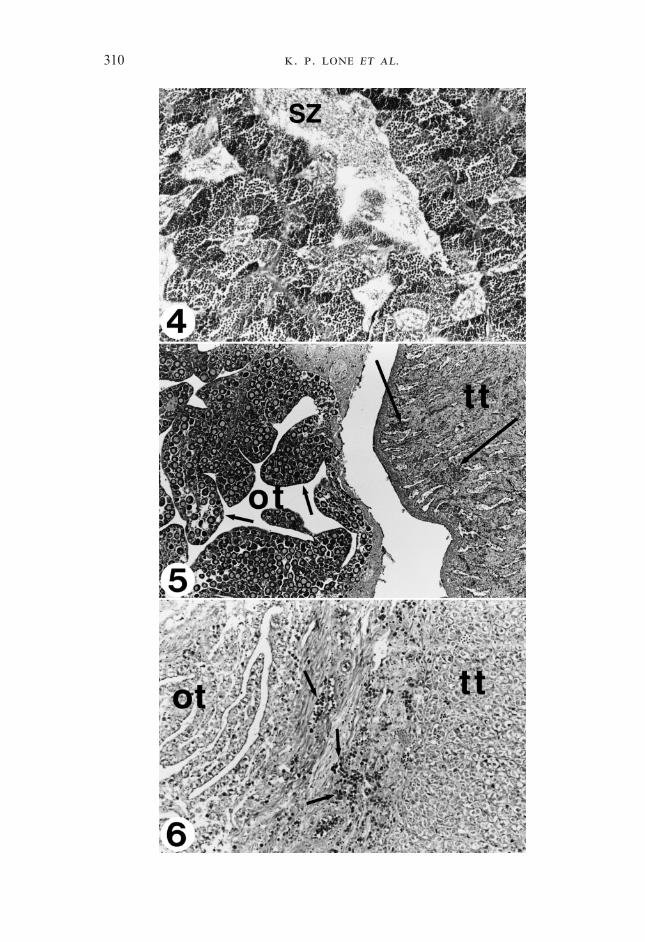
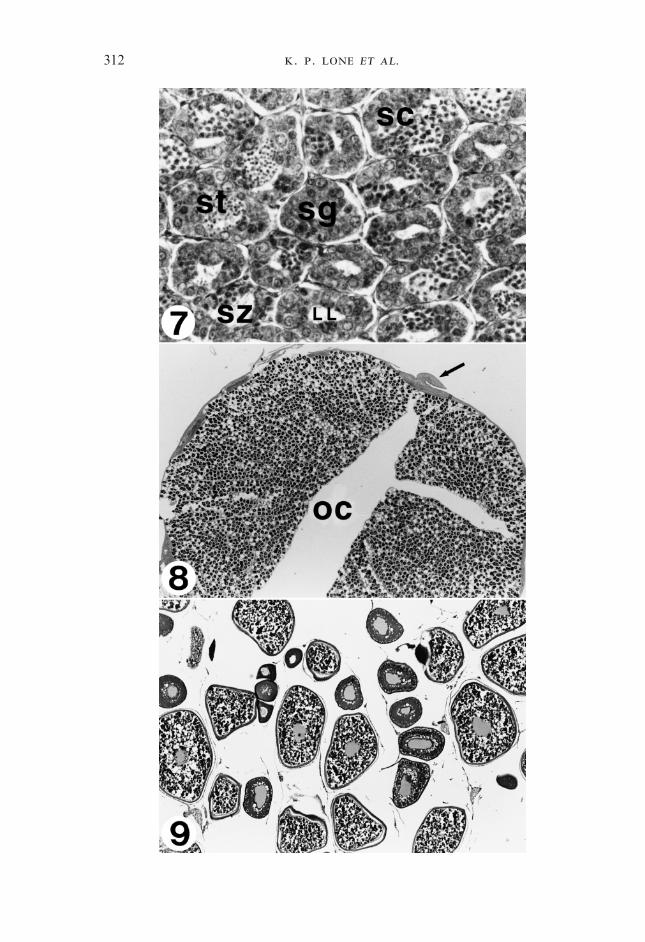
F 4–6. Fig. 4. Transverse section of a mature testis of sobaity taken in January 1997 at the age of 11months. All the stages of spermatogenesis are clearly seen. No female tissue was observed at thisage and maturity stage. SZ, Spermatozoa; haematoxylin and eosin; �120. Fig. 5. Transversesection of a bisexual gonad of sobaity at the end of May showing the developing ovarian lamellae(small arrows) occupied by previtellogenic oocytes. The testicular portion (tt) has regressed andlost its characteristic architecture and is invaded by the acidophilic granulocytes (large arrows). ot,Ovarian tissue; haematoxylin and eosin; �75. Fig. 6. Transverse section of an intersex gonad inAugust showing the actin of acidophilic granulocytes (arrows) invading the ovarian tissue forclearance and the re-establishment of the testicular tissue for the impending spawning season tomature again as males. The testicular tissue is comprised of spermatogonia but primaryspermatocytes were also seen scattered among this section of the tissue. The lobular structure ofthe testis has not been organized yet. ot, Degenerating ovarian lamellae; haematoxylin and eosin;�150.
HISTOLOGY OF THE GONADS (TANKS AND CAGES)There was no difference in the histology of gonads between the tank- and
cage-reared fish. During January–March 1997 (the spawning season of sobaity;age 11–13 months), the gonads were all mature testes with freely running milt onslight handling, although the quantity of the milt varied according to the ripenessof the individual fish. Gross morphological examination showed a typicalmature gonad had a white creamy colour and very good supply of blood throughthe testicular artery. Histologically, primary and secondary spermatocytes,spermatids and spermatozoa (Fig. 4) could be seen. In samples collected in thelast week of March, the lobular structure of the testis was more or less lostbecause of the spawning. However, some intact lobules could be seen scatteredin the tissue.
Histologically, a start towards the intersex gonad could be seen in April 1997,with the appearance of rudimentary ovarian folds devoid of any oocytes. Thegonad’s male portion regressed considerably compared with the March sample.There were portions of the testis which showed appearance of connective tissuewhich was at the expense of the male germinal tissue.
The intersex gonad was well established by May, and gonads could bedifferentiated histologically into male and female portions (Fig. 5). All oocytesin these gonads were at the peri-nucleolar stage. The intersex gonads remainedtill the end of September (Fig. 6). There was no further differentiation from theperi-nucleolar stage of the oocytes in the intersex gonad from May to September.At this time, the majority of the gonads were rearranging into typical testicularpattern. From November 1997 till March 1998, only male gonads were seen(Fig. 7). All fish were running milt by mid-January 1998. It appeared that all theindividuals reverted back towards the male type.
From April (age 26 months) to August 1998 (age 30 months), only intersexgonads were seen. In September 1998, pure males appeared again along withintersex gonads. In October 1998, three types of gonads were found, i.e. puremale, intersex gonads and pure female gonads for the first time (Fig. 8). By theage of 36 months (January 1999), some of the male fish were already ripe and
312 . . .

313
running while in the female gonad the most advanced oocyte was at the tertiaryyolk globule (granule) stage (Fig. 9). When all the fish were killed at the age of36 months, 10 and 17% of the population in tanks and cages, respectively, weretrue females (Table II).
In May 1997, when the intersex gonads were established, acidophilicgranulocytes infiltrated the testicular portion of the gonad. This infiltrationbecame quite prevalent later on. By June 1997, the testicular portion showed aheavy infestation of the acidophilic granulocytes. In September 1997, when theintersex gonads started to revert back to male, the ovarian portion wasundergoing clear resorption. Acidophilia was now confined to the female part ofthe gonad and the final clearance of the remnants of the ovarian folds, whichwere still seen in some gonads, was underway (Fig. 6). A similar pattern wasseen in 1998.
SERUM SEX HORMONES AND REPRODUCTIVE CYCLE
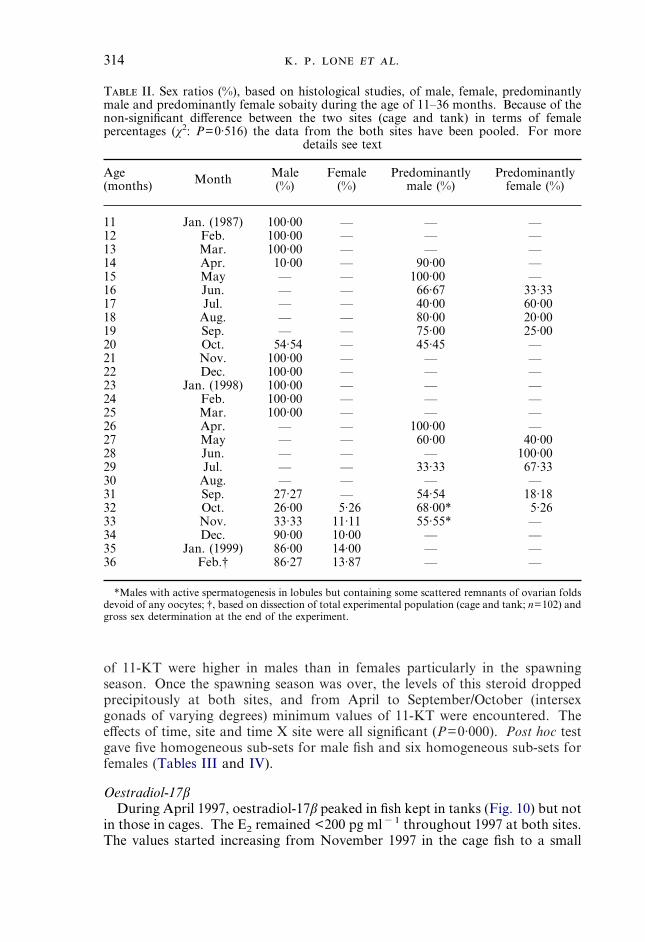
TestosteroneSerum T values showed a small peak in March in all fish examined. At thistime, T concentrations were 0·56�0·08 and 0·48�0·28 ng ml�1 for the maleand future female fish respectively. T values dropped in April and remained lowuntil September 1997. During April–September, the fish had intersex gonads inwhich the area occupied by male and female portions differed. After September1997, the testosterone levels started rising in all fish and showed a clear peak(1·7�0·14 ng ml�1) in December 1997 (male fish) or in January 1998 (potentialfuture female fish). After this peak, the T levels decreased gradually once againuntil November 1998. From November 1998, when the experimental populationhad separated into males and females, the rise in serum T was more rapid in malefish than in true female fish (Fig. 10). In cage reared fish, males had significantlyhigher (P<0·001) values than females (Fig. 11).
Although the general pattern of serum T levels in fish kept in sea cagesresembled the changes described for the tank fish (Fig. 11), the levels of T weregenerally lower in the cage fish than in the tank fish (Tables III and IV). TheTukey post hoc test gave 10 homogeneous sub-sets for both male and female fishfor this hormone.
F 7–9. Fig. 7. Start of the spermatogenesis in testis in October 1997 at the age of 20 months after anintersexual phase from May to September 1997. The typical lobular structure is now establishedagain and all the stages of spermatogenesis can be seen. LL, Lobule lumen; SC, spermatocytes; Sg,spermatogonia; St, spermatids; SZ, spermatozoa; haematoxylin and eosin; �400. Fig. 8. A finalstage towards the female gonad by sex change. Note that the testicular tissue (arrow) has beenreduced to a narrow band only. In the next stage even this area is also resorbed and a pure femalegonad (ovary) is obtained. The oocytes are at the primary growth phase. OC, Ovarian cavity;haematoxylin and eosin; �15. Fig. 9. Transverse section of a female gonad at the age of36 months in February 1999, showing various stages of vitellogenesis. The most advanced stageoocytes are in the tertiary growth stage while a second wave of less advanced oocytes is alsopresent. Some primary oocytes can be seen scattered among these vitellogenic oocytes;haematoxylin and eosin; �40.
11-Ketotestosterone11-Ketotestosterone peaked during the spawning season of the sobaity at both
sites (Figs 10 and 11), and was correlated with the IG data. The concentrations
314 . . .
of 11-KT were higher in males than in females particularly in the spawningseason. Once the spawning season was over, the levels of this steroid droppedprecipitously at both sites, and from April to September/October (intersexgonads of varying degrees) minimum values of 11-KT were encountered. Theeffects of time, site and time X site were all significant (P=0·000). Post hoc testgave five homogeneous sub-sets for male fish and six homogeneous sub-sets forfemales (Tables III and IV).
T II. Sex ratios (%), based on histological studies, of male, female, predominantlymale and predominantly female sobaity during the age of 11–36 months. Because of thenon-significant difference between the two sites (cage and tank) in terms of femalepercentages (�2: P=0·516) the data from the both sites have been pooled. For more
details see text
Age(months) Month Male
(%)Female
(%)Predominantly
male (%)Predominantly
female (%)
11 Jan. (1987) 100·00 — — —12 Feb. 100·00 — — —13 Mar. 100·00 — — —14 Apr. 10·00 — 90·00 —15 May — — 100·00 —16 Jun. — — 66·67 33·3317 Jul. — — 40·00 60·0018 Aug. — — 80·00 20·0019 Sep. — — 75·00 25·0020 Oct. 54·54 — 45·45 —21 Nov. 100·00 — — —22 Dec. 100·00 — — —23 Jan. (1998) 100·00 — — —24 Feb. 100·00 — — —25 Mar. 100·00 — — —26 Apr. — — 100·00 —27 May — — 60·00 40·0028 Jun. — — — 100·0029 Jul. — — 33·33 67·3330 Aug. — — — —31 Sep. 27·27 — 54·54 18·1832 Oct. 26·00 5·26 68·00* 5·2633 Nov. 33·33 11·11 55·55* —34 Dec. 90·00 10·00 — —35 Jan. (1999) 86·00 14·00 — —36 Feb.† 86·27 13·87 — —
*Males with active spermatogenesis in lobules but containing some scattered remnants of ovarian foldsdevoid of any oocytes; †, based on dissection of total experimental population (cage and tank; n=102) andgross sex determination at the end of the experiment.
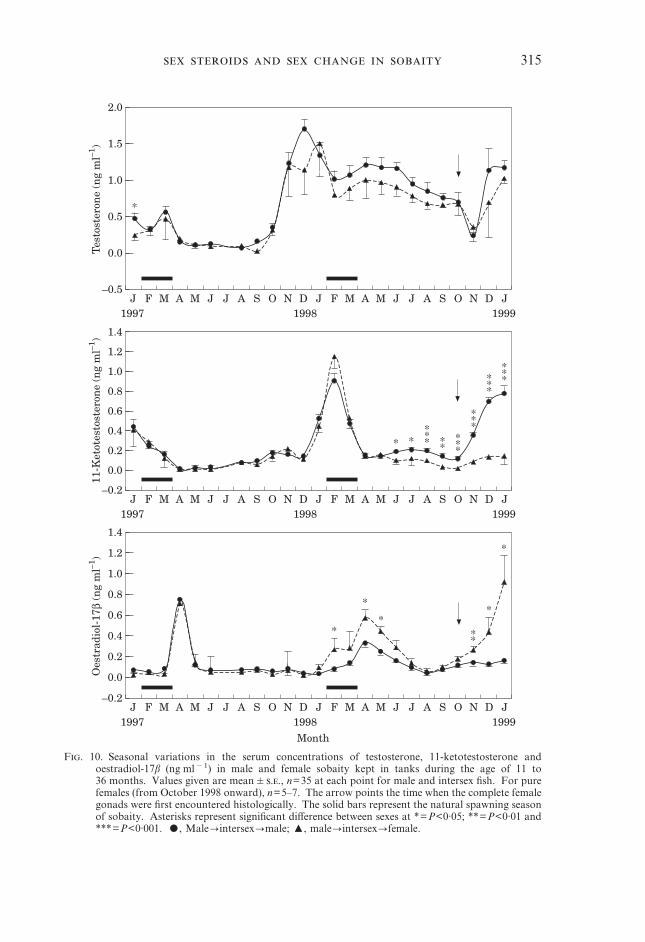
Oestradiol-17�During April 1997, oestradiol-17� peaked in fish kept in tanks (Fig. 10) but not
in those in cages. The E2 remained <200 pg ml�1 throughout 1997 at both sites.The values started increasing from November 1997 in the cage fish to a small
315
Month
J
1.4
1997
Oes
trad
iol-
17β
(ng
ml–1
)
J F M A M J J A S O N D J F M A M J J A S O N D1998 1999
1.2
1.0
0.8
0.6
0.4
0.2
0.0
–0.2
*
*
**
*
*
*
J
1.4
1997
11-K
etot
esto
ster
one
(ng
ml–1
)
J F M A M J J A S O N D J F M A M J J A S O N D1998 1999
1.2
1.0
0.8
0.6
0.4
0.2
0.0
–0.2
****
**
***
***
*** *
***
J
2.0
1997
Tes
tost
eron
e (n
g m
l–1)
J F M A M J J A S O N D J F M A M J J A S O N D1998 1999
1.5
0.0
–0.5
0.5
1.0
*
F. 10. Seasonal variations in the serum concentrations of testosterone, 11-ketotestosterone andoestradiol-17� (ng ml�1) in male and female sobaity kept in tanks during the age of 11 to36 months. Values given are mean�.., n=35 at each point for male and intersex fish. For purefemales (from October 1998 onward), n=5–7. The arrow points the time when the complete femalegonads were first encountered histologically. The solid bars represent the natural spawning seasonof sobaity. Asterisks represent significant difference between sexes at *=P<0·05; **=P<0·01 and***=P<0·001. �, Male�intersex�male; �, male�intersex�female.
316 . . .
Month
J
1.8
1997
Oes
trad
iol-
17β
(ng
ml–1
)
J F M A M J J A S O N D J F M A M J J A S O N D1998 1999
1.61.41.21.00.80.60.40.20.0
–0.2
***
***
***
J
1.4
1997
11-K
etot
esto
ster
one
(ng
ml–1
)
J F M A M J J A S O N D J F M A M J J A S O N D1998 1999
1.2
1.0
0.8
0.6
0.4
0.2
0.0
–0.2
****
**
*
J
2.0
1997
Tes
tost
eron
e(n
g m
l–1)
J F M A M J J A S O N D J F M A M J J A S O N D1998 1999
1.5
1.0
0.5
0.0
–0.5
* *
***
***
F. 11. Seasonal variations in the serum concentrations of testosterone, 11-ketotestosterone andoestradiol-17� (ng ml�1) in male and female sobaity kept in sea-cages during the age of11–36 months. For more details see Fig. 10. �, Male�intersex�male; �,male�intersex�female.
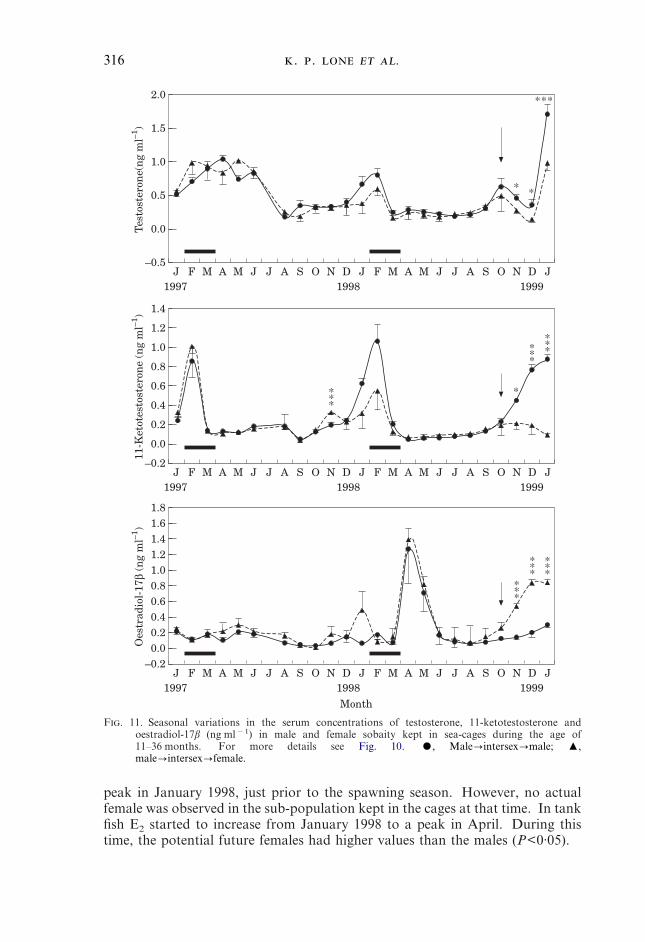
peak in January 1998, just prior to the spawning season. However, no actualfemale was observed in the sub-population kept in the cages at that time. In tankfish E2 started to increase from January 1998 to a peak in April. During thistime, the potential future females had higher values than the males (P<0·05).
317
The April (1998) peak occurred at both sites. Following that peak, which wasslightly lower in fish from tanks, E2 values started declining to a minimum inAugust 1998. From this month onward, the hormone level increased slowly atboth sites but only in true females, which had significantly higher concentrationsthan the male fish (Figs 10 and 11). Like the other two hormones oestradiolconcentrations were also highly significant when analysed by MANOVA(P=0·000). Post hoc test gave seven subsets of homogeneous groups for maleswhile this number for females was six (Tables III and IV).
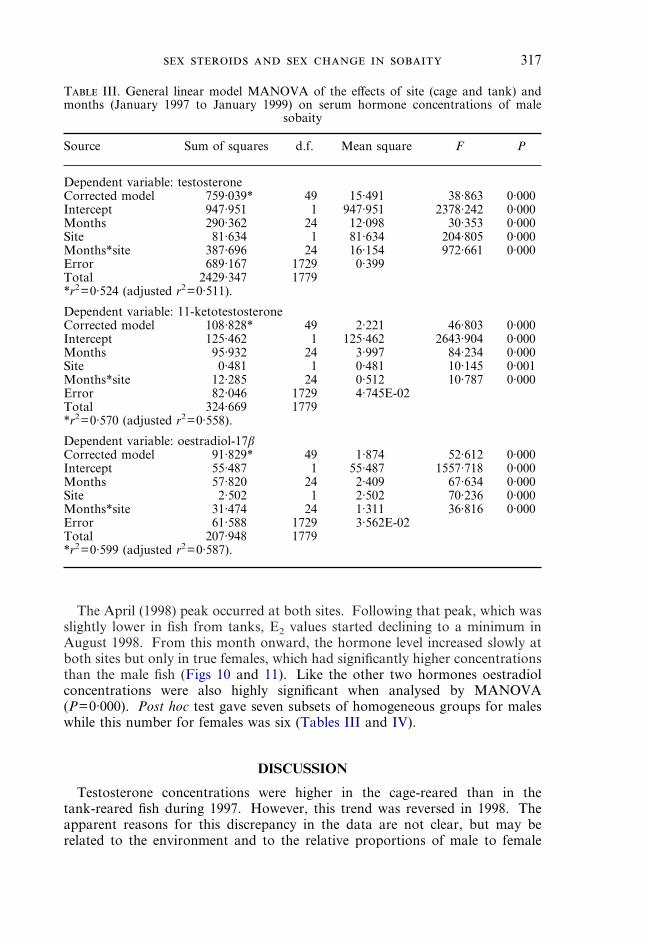
T III. General linear model MANOVA of the effects of site (cage and tank) andmonths (January 1997 to January 1999) on serum hormone concentrations of male
sobaity
Source Sum of squares d.f. Mean square F P
Dependent variable: testosteroneCorrected model 759·039* 49 15·491 38·863 0·000Intercept 947·951 1 947·951 2378·242 0·000Months 290·362 24 12·098 30·353 0·000Site 81·634 1 81·634 204·805 0·000Months*site 387·696 24 16·154 972·661 0·000Error 689·167 1729 0·399Total 2429·347 1779*r2=0·524 (adjusted r2=0·511).
Dependent variable: 11-ketotestosteroneCorrected model 108·828* 49 2·221 46·803 0·000Intercept 125·462 1 125·462 2643·904 0·000Months 95·932 24 3·997 84·234 0·000Site 0·481 1 0·481 10·145 0·001Months*site 12·285 24 0·512 10·787 0·000Error 82·046 1729 4·745E-02Total 324·669 1779*r2=0·570 (adjusted r2=0·558).
Dependent variable: oestradiol-17�Corrected model 91·829* 49 1·874 52·612 0·000Intercept 55·487 1 55·487 1557·718 0·000Months 57·820 24 2·409 67·634 0·000Site 2·502 1 2·502 70·236 0·000Months*site 31·474 24 1·311 36·816 0·000Error 61·588 1729 3·562E-02Total 207·948 1779*r2=0·599 (adjusted r2=0·587).
DISCUSSION
Testosterone concentrations were higher in the cage-reared than in thetank-reared fish during 1997. However, this trend was reversed in 1998. Theapparent reasons for this discrepancy in the data are not clear, but may berelated to the environment and to the relative proportions of male to female
318 . . .
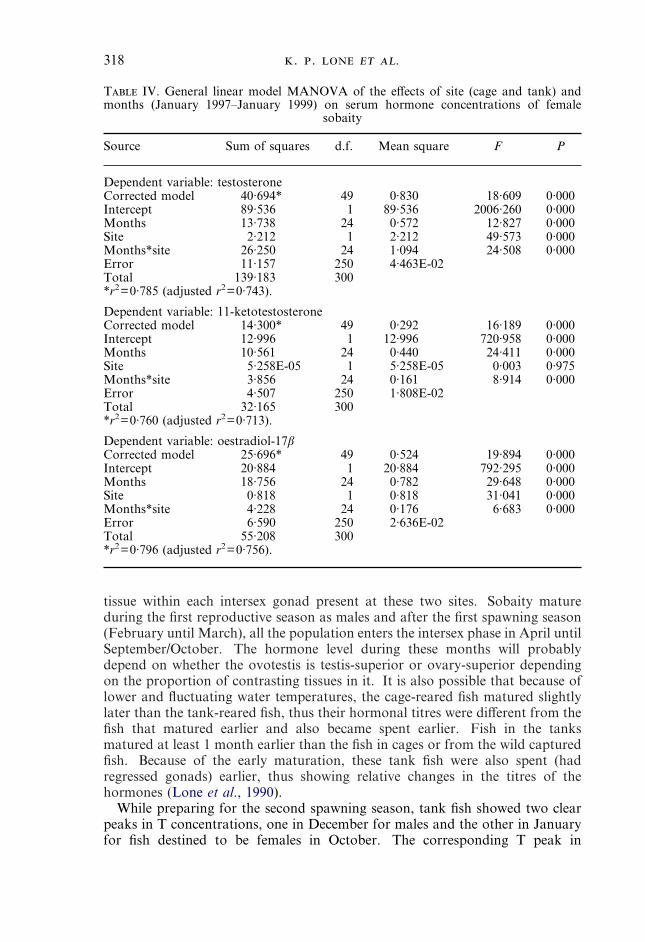
T IV. General linear model MANOVA of the effects of site (cage and tank) andmonths (January 1997–January 1999) on serum hormone concentrations of female
sobaity
Source Sum of squares d.f. Mean square F P
Dependent variable: testosteroneCorrected model 40·694* 49 0·830 18·609 0·000Intercept 89·536 1 89·536 2006·260 0·000Months 13·738 24 0·572 12·827 0·000Site 2·212 1 2·212 49·573 0·000Months*site 26·250 24 1·094 24·508 0·000Error 11·157 250 4·463E-02Total 139·183 300*r2=0·785 (adjusted r2=0·743).
Dependent variable: 11-ketotestosteroneCorrected model 14·300* 49 0·292 16·189 0·000Intercept 12·996 1 12·996 720·958 0·000Months 10·561 24 0·440 24·411 0·000Site 5·258E-05 1 5·258E-05 0·003 0·975Months*site 3·856 24 0·161 8·914 0·000Error 4·507 250 1·808E-02Total 32·165 300*r2=0·760 (adjusted r2=0·713).
Dependent variable: oestradiol-17�Corrected model 25·696* 49 0·524 19·894 0·000Intercept 20·884 1 20·884 792·295 0·000Months 18·756 24 0·782 29·648 0·000Site 0·818 1 0·818 31·041 0·000Months*site 4·228 24 0·176 6·683 0·000Error 6·590 250 2·636E-02Total 55·208 300*r2=0·796 (adjusted r2=0·756).
tissue within each intersex gonad present at these two sites. Sobaity matureduring the first reproductive season as males and after the first spawning season(February until March), all the population enters the intersex phase in April untilSeptember/October. The hormone level during these months will probablydepend on whether the ovotestis is testis-superior or ovary-superior dependingon the proportion of contrasting tissues in it. It is also possible that because oflower and fluctuating water temperatures, the cage-reared fish matured slightlylater than the tank-reared fish, thus their hormonal titres were different from thefish that matured earlier and also became spent earlier. Fish in the tanksmatured at least 1 month earlier than the fish in cages or from the wild capturedfish. Because of the early maturation, these tank fish were also spent (hadregressed gonads) earlier, thus showing relative changes in the titres of thehormones (Lone et al., 1990).
While preparing for the second spawning season, tank fish showed two clearpeaks in T concentrations, one in December for males and the other in Januaryfor fish destined to be females in October. The corresponding T peak in

319
cage-reared fish was in February 1998 for both male and future female fish.Also, the peak values were higher and declined one month earlier in the tank fishthan in the caged fish. After this time the T concentrations remained higher inthe tank fish than in the cage fish. The serum T in the tank fish reached aminimum level in November 1998, and then rose in both male and newlyconverted female fish. This increase was most pronounced in male fish. In thecage-reared fish T showed a small peak in October 1998, then declined untilDecember 1998 and then exhibited a very sharp increase in January 1999. Onceagain, this peak was higher in males than in females.
The role of T in sex reversal depends on the species and environment. In theprotogynous Sparisoma viride (Bonaterre), T did not influence sex inversion(Cardwell & Liley, 1991a, b). In T. duperrey, plasma T did not change in males,intersexes or females (Nakamura et al., 1989). Similarly, in two otherprotogynous species, Monopterous albus (Zuiew) and Centropristes striatus (L.),T levels could not be correlated with the process of sex change (Chan & Yeung,1989; Cochran & Grier, 1991). However, plasma T levels increased during thechange of sex from female to male in the protognyous grouper, Epinephelusmicrodon (Bleeker) and decreased when the change was reversed (Debas et al.,1990). In the protandrous R. sarba, A. schlegeli and Amphiprion melanopusBleeker, sex change showed no positive correlation with the blood T titres(Yeung & Chan, 1987b; Chang & Yueh, 1990; Godwin & Thomas, 1993; Changet al., 1994, 1995a, b). Therefore, these studies, indicate that T generally doesnot have a direct role in the initiation or actual process of sex change, as deducedalso from present results.
11-KT is considered as a male specific steroid in the blood of gonochoristic aswell as hermaphroditic species (Carragher & Pankhurst, 1993; Godwin &Thomas, 1993; Guiguen et al., 1993; Harmin et al., 1995; Hobby & Pankhurst,1997; Haddy & Pankhurst, 1998; Sol et al., 1998). In the present study also,11-KT was higher in the male phase of sobaity than in the female or intersexphases. Furthermore, the blood titres of 11-KT were related directly to the IG
from both tank and cage fish. These and earlier results (Lone et al., 1990; Kimeet al., 1991), indicate that 11-KT concentrations can be used reliably for sexdetermination in sobaity, a species that exhibits no phenotypic differencebetween male and female fish.
The 11-KT concentrations always peaked at the height of the spawning seasonof sobaity (February–March) and then declined precipitously, again parallel withthe IG changes. After October 1998, when male and female fish becameseparated histologically, 11-KT was increasing only in the male sobaity, whereasin the female, it declined as the spawning time approached (Figs 10 and 11). Inthe protogynous hermaphrodite, M. albus, plasma 11-KT was similar in bothsexes and in intersex individuals (Chan & Yeung, 1989). In T. duperrey, plasma11-KT was highest in males and especially in secondary males that were showingreproductive and aggressive behaviour (Nakamura et al., 1989; Hourigan et al.,1991). In the grouper, E. microdon and Lates calcarifer (Bloch) 11-KT in bloodwas similar in different sex types, and levels increased during sex inversion anddecreased during reversion (Debas et al., 1990; Guiguen et al., 1993). It appearsfrom these studies that 11-KT has some physiological role in natural sex changein hermaphrodite fish.

320 . . .
The time-course of E2 in the blood of sobaity from 11–36 months age showedthat males also had E2 in their blood, but most of the time, these concentrationswere very low. During April 1997, male tank fish showed a peak in serumconcentration of E2. This is the time when the spawning has ceased and the titresof the male hormones T and 11-KT were very low (Figs 10 and 11) and intersexgonads appeared for the first time. It is not clear whether this E2 peak wasdue to the appearance of the oocytes in the intersex gonads coupled with lowlevels of male sex steroids or was responsible for the appearance of the intersexgonads.
Some of the intersex fishes (3–5%) had excessively high concentrations of E2 intheir serum (up to 9 ng ml�1). Whether these concentrations were actual or anexperimental artifact is not clear and requires further investigation. Therefore,these data were not included in the present study. However, levels of E2 upto 7·6 ng ml�1 have been reported in males with intersex gonads (giltheadseabream Sparus auratus L. and sea bass) (Kadmon et al., 1985; Guiguen et al.,1993). This phenomenon has been described only in protandrous hermaphro-dites. It may be possible that all the potential sex changing fish needed a suddenstimulus prior to the start of the changes in gonads from male to female. Thisstimulus may be provided by the very high concentrations of E2 suppressing theantagonistic actions of male sex steroids (when these high titres of E2 wereencountered, the male hormone 11-KT had already declined).
Oocytes in the gonads of developing hermaphrodite sparid males have beendescribed many times but they do not develop beyond the early peri-nucleolarstage (Kadmon et al., 1985; Reinboth, 1988; Chang & Yueh, 1990; Guiguenet al., 1993, 1994). These transitory phases were associated with high levels ofE2. There are many reasons for this. Firstly, in vitro studies have shown thatoestrogen synthesis was high in mature female and sex-reversing male S. aurata(Colombo et al., 1972). These high levels of E2 probably give a stimulus for thechange of sex from male to female (Chang et al., 1994, 1995a, b; Chang & Lin,1998). Similar processes may be active at this time in sobaity, another sparid.
The gonad of sobaity seems to differ from the gonads of other hermaphroditespecies described so far. The gonad of a sexually mature sobaity is homo-geneous, both morphologically and histologically, in the sense that whether maleor female an individual contains only one kind of tissue, i.e. either testicular orovarian. Also, there is no colour or other zonation in the gonad belonging toeither sex. In other hermaphrodites (both protandrous or protogynous) species,some portion of the tissue belonging to the opposite sex can be foundhistologically, at least, in fully mature stages (Zohar et al., 1978, 1984; Kadmonet al., 1985; Pollock, 1985; Micale et al., 1987; Chang & Yueh, 1990; Changet al., 1994; Guiguen et al., 1994; Micale & Perdichizzi, 1994; Nakamura et al.,1994; Tobin et al., 1997). Morphologically also, in other hermaphroditic species,the two portions of the gonad can be differentiated. For example, in the twoclosely related S. auratus and A. schlegeli, the female portion is from yellow tored in colour while the testicular portion is whitish or creamy in colour (pers.obs.; Zohar et al., 1978, 1984; Chang et al., 1995b). Moreover, in most speciesthe male and female protions in an intersex gonad are either separated from eachother through a layer of connective tissue or the male and female elements areintermingled freely with each other. In the sobaity intersex gonad, the male and

321
female elements make histologically discrete sections but there is no connectivetissue layer in between them.
There are now some indications that cells having immune properties play anactive part in the elimination or clearance of the degenerating testicular orovarian tissue. This has been shown in the majority of protogynous species(Hastings, 1981; Roberts & Schlieder, 1983; Nemtzov, 1985; Reinboth &Brusle-Sicard, 1997). These cells have been termed phagocytes or melano-macrophages or granulocytes by different authors depending upon the type ofcells and the function involved in their studies. These cells are most active at thetime of degrading oocytes and residual spermatogenic cells in the gonads. In theprotogynous species, they are present during the intersexual periods where theyhelp in the phagocytosis of female tissue and allow the differentiation of maletissue (Ravaglia & Maggese, 1995; Lo Nostro & Guerrero, 1996). This processhas been described also in at least three protandrous species (Besseau & Faliex,1990; Besseau & Brusle-Sicard, 1991; Guiguen et al., 1994). In the present studythese cells were termed acidophilic granulocytes because of their stainingproperties. These cells were present in the testicular portion when the intersexgonad was developing towards the female type; similarly, they were active in theovarian folds when the intersex gonads were reverting towards the male type.This means that these cells are active in the opposite tissue of the same gonad ina hermaphrodite species at different stages of development (Fig. 6).
This work was funded partially by the Kuwait Foundation for the Advancement ofSciences. We thank the Mariculture and Fisheries Department of the Kuwait Institutefor Scientific Research for providing the required facilities and support services for thesestudies; and D. E. Kime for providing the antibodies and steroid labels used.
2001 The Government of Kuwait
References
Besseau, L. & Faliex, E. (1990). Presence de granulocytes et de ‘corps bruns’ dansl’ovotestis de margre, Lithognathus mormyrus (L.) (Teleostei, Sparide).Ichtyophysiologica Acta 13, 109–114.
Besseau, L. & Brusle-Sicard, S. (1991). Sex inversion in a protandric hermaphroditeLithognathus mormyrus (L., 1758) (Teleostei, Sparidae): Histocytological pecu-liarities. In Proceedings of the Fourth International Symposium on the ReproductivePhysiology of Fish (Scott, A. P., Sumpter, J. P., Kime, D. E. & Rolfe, M. S., eds),p. 95. Sheffield: Fish Symposium 91.
Borg, B. (1994). Androgens in teleost fishes. Comparative Biochemistry and Physiology109C, 219–245.
Cardwell, J. R. & Liley, N. R. (1991a). Hormonal control of sex and colour change in thestoplight parrotfish, Sparisoma viride. General and Comparative Endocrinology 81,7–20.
Cardwell, J. R. & Liley, N. R. (1991b). Androgen control of social status in males of awild population of stoplight parrotfish, Sparisoma viride (Scaridae). Hormones andBehavior 25, 1–18.
Carragher, J. F. & Pankhurst, N. W. (1993). Plasma levels of sex steroids during sexualmaturation of snapper, Pagrus auratus (Sparidae) caught from the wild.Aquaculture 109, 375–388.
Chan, S. T. H. & Yeung, W. S. B. (1989). Sex steroids in intersexual fishes. FishPhysiology and Biochemistry 7, 229–235.

322 . . .
Chang, C. G. & Lin, B-Y. (1998). Estradiol-17� stimulates aromatase activity andreversible sex change in protandrous black porgy Acanthopagrus schlegeli. Journalof Experimental Zoology 280, 165–173.
Chang, C. F. & Yueh, W. S. (1990). Annual cycle of gonadal histology and steroidprofile in the juvenile males and adult females of the protandrous black porgy,Acanthopagrus schlegeli. Aquaculture 91, 179–196.
Chang, C. F., Lee, M. F. & Chen, G. R. (1994). Estradiol-17� associated with thesex reversal in protandrous black porgy, Acanthopagrus schlegeli. Journal ofExperimental Zoology 268, 53–58.
Chang, C. F., Lau, E. L. & Li, B. Y. (1995a). Estradiol-17� suppresses testiculardevelopment and stimulates sex reversal in protandrous black porgy,Acanthopagrus schlegeli. Fish Physiology and Biochemistry 14, 481–488.
Chang, C. F., Lau, E. L. & Li, B. Y. (1995b). Stimulation of spermatogenesis or of sexreversal according to the dose of exogenous estradiol-17� in juvenile males ofprotandrous black porgy, Acanthopagrus schlegeli. General and ComparativeEndocrinology 100, 355–367.
Cochran, R. C. & Grier, H. J. (1991). Regulation of sexual succession in the protogynousblack sea bass Centropristis striatus (Osteichthys: Serranidae). General andComparative Endocrinology 82, 69–77.
Colombo, L., del Conte, E. & Clemenze, P. (1972). Steroid biosynthesis in vitro by thegonads of Sparus auratus L. (Teleostei) at different stages during natural sexreversal. General and Comparative Endocrinology 19, 26–36.
Cuisset, B., Pradelles, P., Kime, D. E., Kuhn, E. R., Babin, P., Davail, S. & Le Menn, F.(1994). Enzyme immunoassay for 11-ketotestosterone using acetylcholinesteraseas label: application to the measurement of 11-ketotestosterone in plasma ofSiberian sturgeon. Comparative Biochemistry and Physiology 108C, 229–241.
Debas, L., Fostier, A., Fuchs, J., Weppe, M., Nedelec, G., Benett, A., Cauty, C. &Jalabert, B. (1990). The sexuality of cultured hermaphroditic fish species: analysisof morphological and endocrinological features in a protogynous hermaphrodite,Epinephelus microdon, as a basis for further research to control reproduction in thegrouper. In Proceedings of Advances in Tropical Aquaculture, pp. 543–557. Tahiti:IFREMER.
Fostier, A., Jalabert, B., Billard, R., Breton, B. & Zohar, Y. (1983). The gonadalsteroids. In Fish Physiology, Vol. 9A (Hoar, W. S., Randall, D. J. & Donaldson,E. M., eds), pp. 277–372. New York: Academic Press.
Godwin, J. R. & Thomas, P. (1993). Sex change and steroid profiles in the protandrousanemonefish Amphiprion melanopus (Pomacentridae, Teleostei). General andComparative Endocrinology 91, 144–157.
Guiguen, Y., Jalabert, B., Thouard, E. & Fostier, A. (1993). Changes in plasma andgonadal steroidal hormones in relation to the reproductive cycle and the sexinversion process in the protandrous seabass, Lates calcarifer. General andComparative Endocrinology 92, 327–338.
Guiguen, Y., Cauty, C., Fostier, A., Fuchs, J. & Jalabert, B. (1994). Reproductive cycleand sex inversion of the seabass, Lates calcarifer, reared in sea cages in FrenchPolynesia: histological and morphometric description. Environmental Biology ofFishes 39, 231–247.
Haddy, J. A. & Pankhurst, N. W. (1998). Annual change in reproductive condition andplasma concentrations of sex steroids in black bream, Acanthopagrus butcheri(Munro) (Sparidae). Marine and Freshwater Research 49, 389–397.
Harmin, S. A., Crim, L. W. & Wiegand, M. D. (1995). Plasma sex steroids profiles andthe seasonal reproductive cycle in male and female winter flounder, Pleuronectesamericanus (Walbaum). Marine Biology 121, 601–610.
Hastings, P. A. (1981). Gonad morphology and sex succession in the protogynoushermaphrodite Hemanthias vivanus (Jordon & Swain). Journal of Fish Biology 18,443–454.
Hobby, A. C. & Pankhurst, N. W. (1997). The relationship between plasma andovarian levels of gonadal steroids in the repeat spawning marine fishes Pagrus

323
auratus (Sparidae) and Chromis dispilus (Pomacentridae). Fish Physiology andBiochemistry 16, 65–75.
Hourigan, T. F., Nakamura, M., Nagahama, Y., Yamauchi, K. & Grau, E. G. (1991).Histology, ultrastructure and in vitro steroidogenesis of the testes of two malephenotypes of the protogynous fish, Thalassoma duperrey (Labridae). General andComparative Endocrinology 83, 193–217.
Idler, D. R., Reinboth, R., Walsh, J. M. & Truscott, B. (1976). A comparison of11-hydroxytestosterone and 11-ketotestosterone in blood of ambisexual andgonochoristic teleosts. General and Comparative Endocrinology 30, 517–521.
Kadmon, G., Yaron, Z. & Gordon, H. (1985). Sequence of gonadal events and estradiollevels in Sparus auratus (L.) under two photoperiod regimes. Journal of FishBiology 26, 609–620.
Kime, D. E., Lone, K. P. & Al-Marzouk, A. (1991). Seasonal changes in serum steroidhormones in a protandrous teleost, the sobaity (Sparidentex hasta Valenciennes).Journal of Fish Biology 39, 745–753.
Latz, M., Stahlschmidt-Allner, P. & Reinboth, R. (1991). In vitro metabolism oftestosterone by gonadal tissue of a protandric anemonefish at various sexualstages. In Proceedings of the Fourth International Symposium on the ReproductivePhysiology of Fish, Norwich (Scott, A. P., Sumpter, J. P., Kime, D. E. & Rolfe,M. S., eds), pp. 89–91. Sheffield: Fish Symposium 91.
Lone, K. P., Wuan, T. O. & Al-Marzouk, A. (1990). A preliminary study on the sexualdifferentiation and sex-related growth of sobaity. Kuwait Institute for ScientificResearch, Report No. KISR 3392.
Lo Nostro, F. L. & Guerrero, G. A. (1996). Presence of primary and secondary males ina population of the protogynous Synbranchus marmoratus. Journal of Fish Biology49, 788–800.
Micale, V. & Perdichizzi, F. (1994). Further studies on the sexuality of thehermaphroditeic teleost Diplodus sargus, with particular reference to protandroussex inversion. Journal of Fish Biology 45, 661–670.
Micale, V., Perdichizzi, F. & Santangelo, G. (1987). The gonadal cycle of captive whitebream, Diplodus sargus (L.). Journal of Fish Biology 31, 435–440.
Nakamura, M., Hourigan, T. F., Yamauchi, K., Nagahama, Y. & Grau, E. G. (1989).Histological and ultrastructural evidence for the role of gonadal steroid hormonesin sex changes in the protogynous wrasse Thalassoma duperrey. EnvironmentalBiology of Fishes 2, 117–136.
Nakamura, M., Mariko, T. & Nagahama, Y. (1994). Ultrastructure and in vitrosteroidogenesis of the gonads in the protandrous anemonefish Amphiprionfrenatus. Japanese Journal of Ichthyology 41, 47–56.
Nash, J. P., Davail-Cuisset, B., Bhattacharyya, S., Suter, H. C., Le Menn, F. & Kime,D. E. (2000). An enzyme linked immunosorbant assay (ELISA) for testosterone,estradiol, and 17,20�-dihydroxy-4-pregnen-3-one using acetylcholinesterase astracer: application to measurement of diel patterns in rainbow trout(Oncorhynchus mykiss). Fish Physiology and Biochemistry 22, 355–363.
Nemtzov, S. C. (1985). Social control of sex change in the red sea razorfish Xyrichtyspentadactylus (Teleostei: Labridae). Environmental Biology of Fishes 14, 199–211.
Norusis, M. J. (1996). SPSS Advanced Statistics 7. Chicago: SPSS Inc.Pollock, B. R. (1985). The reproductive cycle of yellowfin bream Acanthopagrus australis
(Gunther), with particular reference to protandrous sex inversion. Journal of FishBiology 26, 301–311.
Ravaglia, M. A. & Maggese, M. C. (1995). Melano-macrophage centers in the gonads ofswamp eel, Synbranchus marmoratus Bloch, (Pisces, Synbranchidae), histologicaland histochemical characterization. Journal of Fish Diseases 18, 117–125.
Reinboth, R. (1975). Spontaneous and hormone induced sex-inversion in wrasses(Labridae). Pubblicazioni della Stazione Zoologica di Napoli 39(Suppl.), 550–573.
Reinboth, R. (1979). On steroidogenic pathways in ambisexual fishes. Proceedings of theIndian National Science Academy 45B, 421–428.

324 . . .
Reinboth, R. (1988). Physiological problems of teleost ambisexuality. EnvironmentalBiology of Fishes 22, 249–259.
Reinboth, R. & Becker, B. (1984). In vitro studies on steroid metabolism by gonadaltissues from ambisexual teleosts. I. Conversion of [14C]testosterone by males andfemales of the protogynous wrasse Coris julis. General and ComparativeEndocrinology 55, 245–250.
Reinboth, R. & Brusle-Sicard, S. (1997). Histological and ultrastructural studies on theeffects of hCG on sex inversion in the protogynous teleost Coris julis. Journal ofFish Biology 51, 738–749.
Roberts, D. E. & Schlieder, R. A. (1983). Induced sex inversion, maturation, spawningand embryogeny of the protogynous grouper, Mycteroperca microlepis. Journal ofWorld Mariculture Society 14, 639–649.
Sol, S. Y., Olson, O. P., Lomax, D. P. & Johnson, L. L. (1998). Gonadal developmentand associated changes in plasma reproductive steroids in English sole,Pleuronectes vetulus, from Puget Sound, Washington. Fishery Bulletin 96,859–870.
Tobin, A. J., Sheaves, M. J. & Molony, B. W. (1997). Evidence of protandroushermaphroditism in the tropical sparid Acanthopagrus berda. Journal of FishBiology 50, 22–33.
Yamamoto, T. (1969). Sex differentiation. In Fish Physiology, Vol. 3 (Hoar, W. S. &Randall, D. J., eds), pp. 117–175. New York: Academic Press.
Yeung, W. S. B. & Chan, S. T. H. (1985). The in vitro metabolism of radioactiveprogesterone and testosterone by the gonads of the protandrous Rhabdosargussarba at various sexual phases. General and Comparative Endocrinology 59,171–183.
Yeung, W. S. B. & Chan, S. T. H. (1987a). The gonadal anatomy and sexual pattern ofthe protandrous sex-reversing fish, Rhabdosargus sarba (Teleostei: Sparidae).Journal of Zoology 212, 521–532.
Yeung, W. S. B. & Chan, S. T. H. (1987b). A radioimmunoassay study of the plasmalevels of sex steroid profiles in the protandrous, sex reversing fish Rhabdosargussarba (Sparidae). General and Comparative Endocrinology 66, 353–363.
Yeung, W. S. B., Chen, H. & Chan, S. T. H. (1993). The in vitro metabolism ofradioactive androstenedione and testosterone by the gonads of the protogynousMonopterus albus at different sexual phases: a time-course and seasonal study.General and Comparative Endocrinology 89, 313–322.
Zohar, Y., Abraham, M. & Gordin, H. (1978). The gonadal cycle of the captivity-rearedhermaphroditic teleost Sparus aurata (L.) during the first two years of life. Annalsde Biologie Animale, Biochemie et Biophysique 18, 877–882.
Zohar, Y., Billard, R. & Weil, C. (1984). La reproduction de la daurade (Sparus aurata)et du bar (Dicentrarchus labrax): connaissance du cycle sexuelle et controle de lagametogenese et de la ponte. In L’Aquaculture du Bar et des Sparides (Barnabe, G.& Billard, R., eds), pp. 3–24. Paris: I.N.R.A. Publication.
Related Documents











