Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters Citation Bakshi, R., A. Yeste, B. Patel, S. Tauhid, S. Tummala, R. Rahbari, R. Chu, et al. 2016. Serum Lipid Antibodies Are Associated with Cerebral Tissue Damage in Multiple Sclerosis. Neurology: Neuroimmunology & Neuroinflammation 3, no. 2: e200–e200. doi:10.1212/nxi.0000000000000200. Published Version doi:10.1212/nxi.0000000000000200 Citable link http://nrs.harvard.edu/urn-3:HUL.InstRepos:25918568 Terms of Use This article was downloaded from Harvard University’s DASH repository, and is made available under the terms and conditions applicable to Other Posted Material, as set forth at http:// nrs.harvard.edu/urn-3:HUL.InstRepos:dash.current.terms-of- use#LAA

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Serum lipid antibodies areassociated with cerebral tissue
damage in multiple sclerosisThe Harvard community has made this
article openly available Please share howthis access benefits you Your story matters
Citation Bakshi R A Yeste B Patel S Tauhid S Tummala R RahbariR Chu et al 2016 Serum Lipid Antibodies Are Associatedwith Cerebral Tissue Damage in Multiple Sclerosis NeurologyNeuroimmunology amp Neuroinflammation 3 no 2 e200ndashe200doi101212nxi0000000000000200
Published Version doi101212nxi0000000000000200
Citable link httpnrsharvardeduurn-3HULInstRepos25918568
Terms of Use This article was downloaded from Harvard Universityrsquos DASHrepository and is made available under the terms and conditionsapplicable to Other Posted Material as set forth at httpnrsharvardeduurn-3HULInstReposdashcurrentterms-of-useLAA
Rohit Bakshi MDAda Yeste PhDBonny Patel MScShahamat Tauhid MDSubhash Tummala MDRoya Rahbari PhDRenxin Chu MDKeren Regev MDPia Kivisaumlkk MD PhDHoward L Weiner MDFrancisco J Quintana
PhD
Correspondence toDr Quintanafquintanaricsbwhharvardedu
Supplemental dataat Neurologyorgnn
Serum lipid antibodies are associated withcerebral tissue damage in multiple sclerosis
ABSTRACT
Objective To determine whether peripheral immune responses as measured by serum antigenarrays are linked to cerebral MRI measures of disease severity in multiple sclerosis (MS)
Methods In this cross-sectional study serum samples were obtained from patients withrelapsing-remitting MS (n 5 21) and assayed using antigen arrays that contained 420 antigensincluding CNS-related autoantigens lipids and heat shock proteins Normalized compartment-specific global brain volumes were obtained from 3-tesla MRI as surrogates of atrophy includinggray matter fraction (GMF) white matter fraction (WMF) and total brain parenchymal fraction(BPF) Total brain T2 hyperintense lesion volume (T2LV) was quantified from fluid-attenuatedinversion recovery images
Results We found serum antibody patterns uniquely correlated with BPF GMF WMF and T2LVFurthermore we identified immune signatures linked to MRI markers of neurodegeneration (BPFGMF WMF) that differentiated those linked to T2LV Each MRI measure was correlated with aspecific set of antibodies Strikingly immunoglobulin G (IgG) antibodies to lipids were linked tobrain MRI measures Based on the association between IgG antibody reactivity and each uniqueMRI measure we developed a lipid index This comprised the reactivity directed against all of thelipids associated with each specific MRI measure We validated these findings in an additionalindependent set of patients with MS (n 5 14) and detected a similar trend for the correlationsbetween BPF GMF and T2LV vs their respective lipid indexes
Conclusions We propose serum antibody repertoires that are associated with MRI measuresof cerebral MS involvement Such antibodies may serve as biomarkers for monitoring diseasepathology and progression Neurol Neuroimmunol Neuroinflamm 20163e200 doi 101212
NXI0000000000000200
GLOSSARYBPF 5 brain parenchymal fraction GM 5 gray matter GMF 5 gray matter fraction IgG 5 immunoglobulin G MS 5 multiplesclerosis T2LV 5 T2 hyperintense lesion volume WM 5 white matter WMF 5 white matter fraction
Multiple sclerosis (MS) is characterized by immune dysfunction and inflammation leading tofocal lesions brain and spinal cord atrophy and progressive neurologic dysfunction The knownheterogeneity likely reflects myriad and complex underlying pathogenic mechanisms that makespecific and unique contributions to MS1
MRI-defined T2 hyperintense brain lesions are key to diagnosis and therapeutic monitoringHowever such lesions are nonspecific for the underlying pathology and have limited clinicalpredictive value23 Measurement of brain atrophy provides the potential to detect destructivedisease effects and show better associations with clinical status than can be obtained with lesionmeasures2 Atrophy begins early in MS and can be monitored by MRI segmentation2ndash4 Graymatter (GM) atrophy is more closely linked to clinical status than white matter (WM) atrophy
From the Partners Multiple Sclerosis Center (RB S Tauhid S Tummala RC HLW) and Ann Romney Center for Neurologic Diseases (RBAY BP RR KR PK HLW FJQ) Neurology (RB AY BP S Tauhid S Tummala RR RC KR PK HLW FJQ) andRadiology (RB) Brigham and Womenrsquos Hospital Harvard Medical School Boston MA
Funding information and disclosures are provided at the end of the article Go to Neurologyorgnn for full disclosure forms The Article ProcessingCharge was paid by the authors
This is an open access article distributed under the terms of the Creative Commons Attribution-NonCommercial-NoDerivatives License 40 (CCBY-NC-ND) which permits downloading and sharing the work provided it is properly cited The work cannot be changed in any way or usedcommercially
Neurologyorgnn copy 2016 American Academy of Neurology 1
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
whole brain atrophy or conventional lesionassessments56 This likely reflects the func-tional importance of GM and the contentionthat pseudoatrophy confounds the use ofwhole brain or WM atrophy to monitor pro-gressive neurodegeneration7
Immune processes have a central role inboth the pathogenesis and treatment ofMS8ndash12 The ability to link such changes toMRI presents the opportunity to providenew biomarkers and better understanding ofdisease pathophysiology13ndash19
Antigen microarrays are newly developedtools for the high-throughput characterizationof the immune response2021 that have beenused to identify biomarkers and mechanismsof disease pathogenesis in several autoimmunedisorders including MS22ndash31 In the presentstudy we investigated the relationship betweenantigen arrays and both GM and WM cerebralMRI involvement in MS
METHODS Patients Table 1 summarizes the patientsrsquo
demographic and clinical characteristics of the discovery and val-
idation sets All serum samples were collected from the ongoing
cohort of patients being followed in the CLIMB study (Compre-
hensive Longitudinal Investigation of MS at Brigham and
Womenrsquos Hospital32) in which participants are followed with
comprehensive clinical and imaging assessments to monitor
disease progression and response to therapy on a yearly basis
Samples were collected within (mean 6 SD) 50 6 32 months
of MRI acquisition Patients were free of relapses or changes in
disease-modifying therapy during the interval between blood
collection and MRI This was a consecutive sample meeting the
following criteria (1) age 18 to 55 years (2) diagnosis of
relapsing-remitting MS33 (3) absence of other major medical
neurologic or neuropsychiatric disorders (4) lack of any relapse
or corticosteroid use in the 4 weeks before MRI or start of disease-
modifying therapy 6 months before MRI (to reduce confounding
effects on MRI) and (5) no history of smoking or substance
abuse The majority of patients were receiving disease-
modifying treatment at the time of MRI Within 3 months of
MRI each patient received an examination by an MS specialist-
neurologist including evaluation of neurologic disability on the
Expanded Disability Status Scale and a timed 25-foot walk
Standard protocol approvals registrations and patientconsents Our study received approval from the ethical standards
committee on human experimentation at our institution
(The Partners Health Care Institutional Review Board) All par-
ticipants gave written informed consent for their participation in
the study
MRI acquisition and analysis All participants in the discoveryset underwentMRI on the same scanner (3T Signa General Electric
Healthcare Milwaukee WI) using a receive-only phase array head
coil with the same MRI protocol The scan acquisition protocol
has been detailed previously34 Contiguous slices covering the
whole brain were acquired in high-resolution protocols using
3-dimensional modified driven equilibrium Fourier transform and
T2-weighted fast fluid-attenuated inversion recovery sequences
Patients in the validation set underwent brain MRI on a 15T
scanner (GE Signa) including a 2-dimensional axial conventional
spin-echo dual-echo T2-weighted series (voxel sizes 0943 0943
3 mm) Analysis of these scans was performed by operators who
were unaware of clinical and biomarker information In the
discovery set we obtained normalized compartment-specific
global brain volumes as surrogates of atrophy including GM
fraction (GMF) WM fraction (WMF) and total brain
parenchymal fraction (BPF) using statistical parametric mapping
version 8 (WellcomeDepartment of Cognitive Neurology London
UK httpwwwfilionuclacukspm) after manual correction of
(1) misclassifications of tissue compartments due to MS lesion and
(2) ineffective contouring of the deep GM structures34 In the
validation set BPF and GMF were obtained in statistical
parametric mapping version 8 from the dual-echo images
Because the source images did not show effective contrast for
segmentation of the deep gray structures we performed manual
masking to derive only the cerebral cortical GMF Quantification
of total brain T2 hyperintense lesion volume (T2LV) was performed
using Jim (Xinapse Systems Ltd West Bergholt UK httpwww
xinapsecom) by the consensus of 2 experienced observers from the
fluid-attenuated inversion recovery (discovery set) or dual-echo
(validation set) images using a semiautomated technique For the
measurement of these atrophy and lesion surrogates from MRI
scans our methods are well established regarding their operational
procedures validity and reliability34ndash39
Antigens Peptides were synthesized at the Biopolymers Facility
of the Department of Biological Chemistry and Molecular Phar-
macology of Harvard Medical School Recombinant proteins and
lipids were purchased from Sigma (St Louis MO) Abnova
(Taipei City Taiwan) Matreya LLC (Pleasant Gap PA) Avanti
Polar Lipids (Alabaster AL) Calbiochem (San Diego CA)
Chemicon (Temecula CA) GeneTex (San Antonio TX) Novus
Table 1 Demographic clinical and brain MRI data
Discovery seta Validation setb
No of patients with relapsing-remitting MS 21 14
Age y 406 6 80 471 6 81
Women n () 14 (67) 13 (93)
Disease duration y 68 6 50 127 6 81
EDSS score 14 6 12 17 6 19
Timed 25-ft walk s 47 6 06 83 6 102
BPFc 083 6 004 076 6 005
GMFc 049 6 004d 033 6 003e
WMF 033 6 002 NP
T2LVc mL 143 6 169 49 6 79
Abbreviations BPF 5 whole brain parenchymal fraction EDSS 5 Expanded Disability Sta-tus Scale GMF 5 cerebral gray matter fraction MS 5 multiple sclerosis NP 5 not per-formed T2LV 5 cerebral T2 hyperintense lesion volume WMF 5 global cerebral whitematter fractionValues represent mean 6 SD unless otherwise indicateda Fluid-attenuated inversion recovery 3T high resolutionbDual-echo 15T low resolutionc The 2 groups had different MRI acquisitionsource images (3T high resolution vs 15T lowresolution) and different software analysis pipelines leading to a difference in scalingbetween the MRI output metrics (see the methods section for more details)dWhole brain GMFeCortical GMF
2 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
Table 2 Serum immunoglobulin Gs associated with MRI measures of disease severity
Antigens
BPF GMF WMF T2LV
r p FDR r p FDR r p FDR r p FDR
1-P-2-5-o-V-sn-G-3-PC 2038 0087 0161 2034 0131 0197 2014 0540 1000 056 0008a 0056
15-ketocholestene 2030 0190 0370 2044 0047b 0066 024 0302 0854 030 0189 0247
9 HODE 2040 0073 0097 2051 0019b 0026b 016 0499 1000 045 0043a 0057
9 S-HODE 2042 0057 0067 2048 0027b 0026b 006 0797 1000 061 0003a 0033a
a-MSH 050 0021a 0040a 040 0070 0112 028 0214 0854 2004 0858 1000
ABPF 1-12 2023 0320 0610 2040 0070 0112 032 0155 0804 046 0035a 0057
ABPP 227 2030 0179 0305 2056 0009b 0025b 047 0033a 0156 049 0024a 0056
Ac phosphatase 035 0119 0194 047 0033a 0027a 2018 0443 1000 2020 0374 0516
ADPF 1-34 2020 0393 0823 2038 0089 0133 034 0126 0804 046 0035a 0057
ANP 013 0587 0969 037 0100 0197 2048 0027b 0142 2027 0242 0516
Asialoganglioside-GM1 2052 0016b 0040b 2060 0004b 0015b 007 0749 1000 032 0152 0209
Asialoganglioside-GM2 2040 0074 0097 2048 0027b 0026b 011 0648 1000 024 0292 0516
b-MSH 2047 0033b 0058 2038 0085 0133 2025 0277 0854 020 0376 0516
b-Amyloid 2021 0350 0610 2036 0107 0197 026 0247 0854 049 0025a 0056
b-Synuclein 001 0954 0969 2022 0343 0996 048 0029a 0142 042 0055 0094
bNGF 029 0204 0413 006 0783 0996 051 0018a 0122 2006 0807 1000
Brain D-erythrosphingosine 2047 0030b 0058 2056 0009b 0025b 010 0679 1000 037 0096 0146
Brain extract VII 2052 0016b 0040b 2059 0005b 0015b 006 0789 1000 053 0013a 0056
Brain L-a-phosphatidyl-ethanolamine 2044 0044b 0067 2046 0037b 0027b 2005 0837 1000 019 0407 0537
Brain polar lipid extract 013 0573 0969 2010 0657 0996 051 0019a 0122 018 0434 0537
Chorionic G 005 0826 0969 028 0223 0303 2046 0034b 0156 2010 0656 1000
CNPase aa 106ndash125 039 0080 0161 049 0026a 0026a 2014 0557 1000 2051 0017b 0056
CNPase aa 16ndash35 042 0056 0067 046 0037a 0027a 2002 0941 1000 2037 0103 0157
CNPase aa 195ndash214 031 0173 0305 050 0020a 0026a 2035 0119 0804 2035 0118 0157
CNPase aa 286ndash305 028 0227 0413 045 0039a 0027a 2033 0144 0804 2004 0873 1000
CNPase aa 376ndash395 019 0416 0823 043 0049a 0066 2049 0024b 0142 2038 0089 0146
Collagen II 025 0278 0610 046 0034a 0027a 2040 0072 0788 2032 0151 0209
Collagen IX 2067 0001b 0025b 2075 0001b 0002b 008 0741 1000 066 0001a 0033a
Collagen VIII 2027 0242 0521 2026 0251 0605 2005 0831 1000 044 0044a 0057
Collagen X 2036 0114 0194 2012 0595 0996 2052 0016b 0122 2006 0792 1000
Disialoganglioside-GD2 2040 0073 0097 2049 0023b 0026b 013 0589 1000 043 0055 0094
Disialogang-GD1B 2038 0088 0161 2050 0022b 0026b 018 0441 1000 031 0174 0247
Disialoganglioside GD1a 2051 0017b 0040b 2053 0013b 0026b 2005 0824 1000 042 0060 0146
Disialoganglioside GD3 2040 0069 0097 2054 0012b 0026b 020 0382 1000 051 0018a 0056
Galactocerebrosides 001 0952 0969 2027 0229 0303 059 0005a 0108 025 0265 0516
Ganglioside mixture 2053 0014b 0040b 2062 0003b 0015b 011 0644 1000 053 0014a 0056
Ganglioside-GM4 002 0929 0969 029 0203 0303 2056 0009b 0109 2018 0423 0537
Gelsolin 053 0013a 0040a 057 0008a 0025a 002 0934 1000 2048 0029b 0056
GNRH 2053 0014b 0040b 2043 0054 0072 2029 0196 0854 036 0113 0157
gpMBP 017 0449 0928 2004 0857 1000 048 0029a 0142 000 0988 1000
GT1a 2043 0053 0067 2048 0028b 0026b 003 0912 1000 032 0161 0247
Hemoglobin 015 0527 0969 2006 0784 0996 046 0037a 0203 028 0212 0516
HSP60 aa 195ndash214 023 0311 0610 004 0879 1000 044 0045a 0206 000 0989 1000
HSP60 aa 210ndash229 053 0015a 0040a 049 0024a 0026a 016 0493 1000 2013 0577 1000
ContinuedNeurology Neuroimmunology amp Neuroinflammation 3
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
Table 2 Continued
Antigens
BPF GMF WMF T2LV
r p FDR r p FDR r p FDR r p FDR
HSP60 aa 240ndash259 048 0028a 0058 052 0017a 0026a 000 0990 1000 2047 0030b 0056
HSP60 aa 421ndash440 010 0655 0969 039 0080 0133 2058 0006b 0108 2022 0334 0516
HSP60 aa 436ndash455 042 0061 0067 060 0004a 0015a 2031 0173 0804 2038 0094 0146
HSP70 aa 121ndash140 044 0046a 0067 036 0111 0197 024 0289 0854 2016 0496 0728
HSP70 aa 181ndash199 051 0017a 0040a 048 0027a 0026a 015 0525 1000 2020 0378 0516
HSP70 aa 210ndash229 006 0788 0969 027 0228 0303 2044 0045b 0206 2011 0640 1000
HSP70 aa 316ndash335 2028 0216 0413 2029 0209 0303 2004 0879 1000 044 0048a 0072
HSP70 aa 376ndash395 2042 0059 0067 2046 0035b 0027b 002 0942 1000 035 0121 0157
HSP70 aa 421ndash440 048 0030a 0058 065 0002a 0015a 2027 0229 0854 2034 0137 0209
HSP70 aa 61ndash80 2002 0941 0969 2029 0198 0303 056 0008a 0109 018 0441 0537
IL-12 2050 0021b 0040b 2045 0040b 0027b 2018 0429 1000 039 0083 0146
IL-3 2041 0063 0067 2051 0019b 0026b 013 0575 1000 052 0017a 0056
Insula 046 0038a 0067 033 0143 0303 034 0133 0804 2017 0473 0550
Intrinsic factor 036 0108 0194 044 0048a 0066 2009 0702 1000 000 0986 1000
Isoprostane F2a-I 2053 0014b 0040b 2055 0010b 0025b 2004 0854 1000 035 0115 0157
Lactosylceramide 2021 0368 0630 2041 0062 0112 039 0077 0788 060 0004a 0033a
LIF 2065 0002b 0025b 2059 0005b 0015b 2022 0336 0854 041 0064 0146
MMP2 042 0055 0067 061 0003a 0015a 2032 0153 0804 2039 0082 0146
MMP9 2055 0009b 0040b 2062 0003b 0015b 006 0804 1000 045 0038a 0057
MOG peptide aa 196ndash215 2005 0837 0969 2008 0736 0996 005 0836 1000 044 0045a 0057
Monosialogang GM2 2056 0008b 0040b 2048 0028b 0026b 2026 0260 0854 017 0460 0550
Myelin protein 2 peptide aa 31ndash50 035 0124 0194 048 0029a 0027a 2022 0337 0854 2033 0141 0209
Myelin protein 2 peptide aa 91ndash110 2009 0694 0969 014 0533 0996 2051 0019b 0122 2004 0878 1000
Neurofilament 160 kDa 032 0162 0305 039 0082 0133 2009 0688 1000 2049 0026b 0056
Neurofilament 200 kDa 043 0050 0067 045 0040a 0027a 003 0881 1000 2015 0516 0728
Neurofilament 68 kDa 000 0999 1000 2021 0359 0996 044 0044a 0206 045 0043a 0057
NOGO 016 0493 0928 2012 0617 0996 060 0004a 0108 021 0352 0516
Nonhydroxy fatty acid ceramide 2053 0014b 0040b 2059 0005b 0015b 005 0826 1000 044 0044a 0057
Occipital lobe 2045 0039b 0067 2043 0054 0072 2012 0599 1000 039 0083 0146
ORF26 protein 053 0013a 0040a 052 0015a 0026a 010 0653 1000 2030 0182 0247
Parietal lobe 049 0023a 0040a 058 0005a 0015a 2011 0641 1000 2025 0283 0516
PDGF-Ra 2007 0752 0969 017 0461 0996 2052 0016b 0122 2009 0687 1000
Pepstatin 2035 0116 0194 2046 0036b 0027b 016 0483 1000 025 0277 0516
Postcentral gyrus AD 044 0044a 0067 034 0132 0197 028 0214 0854 2012 0603 1000
Proteolipid protein peptide aa 125ndash141 2009 0685 0969 010 0678 0996 2041 0064 0788 2048 0029b 0056
Proteolipid protein peptide aa 150ndash163 044 0043a 0067 051 0019a 0026a 2006 0809 1000 2050 0020b 0056
Proteolipid protein peptide aa 161ndash180 022 0343 0610 031 0166 0303 2017 0475 1000 2044 0045b 0057
Proteolipid protein peptide aa 250ndash269 037 0103 0194 050 0020a 0026 2023 0321 0854 2037 0098 0146
RBP 025 0274 0610 029 0201 0303 2005 0844 1000 2057 0007b 0056
Ribonuclease 2002 0937 0969 2017 0472 0996 031 0175 0804 054 0011a 0056
S100b 033 0141 0258 011 0649 0996 052 0015a 0122 2022 0344 0516
Spectrin 048 0026a 0058 035 0122 0197 036 0105 0804 2001 0966 1000
Sulfatides 2047 0033b 0058 2049 0023b 0026b 2002 0922 1000 034 0133 0209
Tetrasialoganglioside-GQ1B 2043 0054 0067 2057 0007b 0025b 023 0319 0854 048 0027a 0056
Continued4 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
Biologicals (Littleton CO) Assay Designs (Ann Arbor MI) Pro-
Sci Inc (Poway CA) EMD Biosciences (San Diego CA) Cay-
man Chemical (Ann Arbor MI) HyTest (Turku Finland)
Meridian Life Science (Memphis TN) and Biodesign Interna-
tional (Saco ME) The antigens used in the construction of anti-
gen microarrays are listed in table e-1 at Neurologyorgnn
Antigen microarray production development and dataanalysis The antigens listed in table e-1 were spotted in replicatesof 6 on SuperEpoxy 2 slides (TeleChem Sunnyvale CA) using
an Arrayit NanoPrint 2 LM210 microarray printer (Arrayit Corpo-
ration Sunnyvale CA) and optimized spotting conditions as
described28303140 The microarrays were hybridized using an HS
4800Pro Hybridization Station (Tecan Maumlnnedorf Switzerland)
in which they were blocked with 1 bovine serum albumin for
1 hour and incubated for 2 hours at 37degC with the test serum at
a 1100 dilution in blocking buffer The arrays were then washed
and incubated for 45 minutes with a 1500 dilution of goat
anti-human immunoglobulin G (IgG) Cy3-conjugated and
goat anti-human IgM Cy5-conjugated detection antibodies (Jackson
ImmunoResearch Labs West Grove PA) The arrays were scanned
with a Tecan PowerScanner Repeated measurements indicate that
our antigen microarray technique is reproducible exhibiting
a coefficient of variation of 1336 1231
Background signal was subtracted and raw data were normal-
ized and analyzed using the GeneSpring software (Silicon Genetics
Redwood City CA) Antigen reactivity was defined by the mean
intensity of binding to the replicates of that antigen on the micro-
array and expressed as relative fluorescence units Scatter plots were
generated using linear regression models in the R statistical package
Pearson product-moment correlation coefficients were calculated
between the MRI measures (BPF GMF WMF T2LV) and the
weighted average of a group of statistically significant lipids The
weighted average of significant lipids was calculated using the for-
mula (SWiAiSWi) (SWi 5 1 i 5 1 2 3 ) where Wi is the
proportion of intensity of antigen Ai Ai is the observed intensity of
an antigen and SWi is the sum of the weights
RESULTS Serum IgG antibodies correlate with brain
MRI measures of disease severity To study the relation-ship between the peripheral immune response andMRI measures of disease severity we analyzed serum
antibody reactivity in those MS samples We analyzedIgG serum antibodies using a panel of antigensincluding CNS antigens heat shock proteins and lip-ids The association between the antibody reactivityagainst each antigen and 4 MRI measures of disease(T2LV BPF WMF and GMF) was investigatedusing Spearman correlation tests
We found significant associations between eachMRI measure of disease and different sets of IgG anti-body reactivity which are shown in table 2 Similarpatterns of antibody reactivity were linked to BPFand GMF consistent with the known dominant con-tribution of GM atrophy to whole brain atrophymeasures3741 Strikingly there was little overlapbetween the antibody reactivity associated withGMF and WMF suggesting that different immuno-pathologic processes contribute to tissue degenerationin these areas of the brain Similarly these profiles ofantibody reactivity were also different from thoseassociated with T2LV
Serum lipid-reactive IgGs are associated with increased
disease pathogenesis determined by MRI In evaluationof MRI measures and their link to disease BPFGMF and WMF decrease with disease progressionwhile T2LV increases with disease progressionAccordingly we analyzed the linkage between eachMRI measure and the significant antibody reactivitiesshown in table 2 We identified IgG antibody reac-tivities associated with increased tissue destruction asevidenced by decreased BPF values (figure 1) Strik-ingly we found that antibodies linked to diseasepathology as measured by BPF were enriched for reac-tivity against lipids Furthermore a selective increasein lipid-reactive antibodies linked to disease severitywas also observed when we analyzed all available MRImeasures (figure 2) Of note we did not detect an
Table 2 Continued
Antigens
BPF GMF WMF T2LV
r p FDR r p FDR r p FDR r p FDR
Thyrocalcitonin 2033 0143 0258 2008 0746 0996 2059 0005b 0108 004 0878 1000
TNPAL-galactocerebroside 007 0763 0969 2015 0506 0996 047 0030a 0142 005 0823 1000
Total cerebroside 2016 0498 0928 2041 0064 0112 050 0020a 0122 062 0003a 0033
Trisialogang GT1b 2034 0128 0194 2044 0043b 0066 015 0513 1000 030 0185 0247
Troponin I 2051 0020b 0040b 2053 0013b 0026b 2003 0900 1000 043 0050 0072
Ubiquitin 031 0166 0305 045 0039a 0027 2025 0282 0854 2029 0196 0247
VEGF 2050 0023b 0040b 2047 0031b 0027b 2013 0570 1000 040 0070 0146
Abbreviations BPF 5 whole brain parenchymal fraction FDR 5 false discovery ratendashadjusted significance value GMF 5 global cerebral gray matterfraction T2LV 5 cerebral T2 (fluid-attenuated inversion recovery) hyperintense lesion volume WMF 5 global cerebral white matter fractionDiscovery setap 005 (positive association)bp 005 (negative association)
Neurology Neuroimmunology amp Neuroinflammation 5
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
enrichment in lipid-reactive IgM in the group ofantibodies associated with increased diseasepathogenesis (not shown)
An index of serum IgG reactivity to lipids is associated
with MRI measures of disease severity To further inves-tigate the association between serum IgG reactivityto lipids and MRI measures of disease severity wecalculated an IgG anti-lipid index for each patientcorresponding to the information on each lipid-specific antibody listed in table 2 normalized by thestrength of its correlation with that MRI measureunder investigation The antibody reactivities tolipids used to calculate the lipid index are shown in
figure 3A With this approach we calculated one indexfor each of the MRI measures analyzed in this studyWe found a significant correlation between each IgGlipid antibody index and BPF GMF and T2LV(figure 3B) no significant correlation was found withWMF Moreover no significant correlations werefound between BPF WMF GMF and T2LVmeasures and anti-lipid antibody indexes based onIgM reactivity (not shown)
Finally we evaluated the performance of the lipidantibody indexes linked to BPF GMF and T2LV onan additional set of independent MS samples (valida-tion set table 1) We detected a similar trend to theone detected in the discovery set with regard to the
Figure 1 Antibody reactivity in serum is associated with decreased brain volume
Heatmap in which each column represents the mean immunoglobulin G antibody reactivity in a serum sample from a patient with multiple sclerosis sortedaccording to the whole brain parenchymal fraction (BPF) (indicated at the topmdasha lower BPF indicates more brain atrophy) and each row represents theantibody reactivity to an antigen according to the colorimetric scale shown The antibody reactivities included in this heatmap are listed in table 2
6 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
Figure 2 Association of serum IgG reactivity with MRI measures of disease severity
(A) Heatmap in which each column represents an MRI measure of either atrophy (BPF GMF WMF) or lesions (T2LV) andeach row represents the correlation to IgG serum antibody reactivity according to a colorimetric scale (B) Frequency oflipid-reactive antibodies linked to higher or lower MRI disease severity BPF 5 whole brain parenchymal fraction GMF 5
global cerebral gray matter fraction IgG 5 immunoglobulin G T2LV 5 cerebral T2 (fluid-attenuated inversion recovery)hyperintense lesion volume WMF 5 global cerebral white matter fraction
Neurology Neuroimmunology amp Neuroinflammation 7
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
correlation between BPF and GMF and their respec-tive lipid indexes and we validated the correlationbetween T2LV and its lipid index (figure 3C)
DISCUSSION Previous studies have investigated theassociation of MRI with immune activity in MS13ndash19
These studies have included cellular immune measures
Figure 3 Correlation of lipid indexes to MRI measures of disease severity in multiple sclerosis
(A) Heatmap inwhich each column represents anMRImeasure and each row represents the correlation to immunoglobulin G serumantibody reactivity to lipids accord-ing to a colorimetric scale (B) Scatter plots depicting the correlation between each lipid index andMRImeasures (BPF GMF and T2LV) in the discovery set (C) Scatterplots depicting the correlation between each lipid index andMRImeasures (BPF GMF and T2LV) in the validation set BPF5whole brain parenchymal fraction GMF5
cerebral gray matter fraction (see methods section for details) T2LV 5 cerebral T2 hyperintense lesion volume WMF 5 global cerebral white matter fraction
8 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
as well as oligoclonal bands or neurofilament-specificantibodies in CSF42ndash44 In the present study weanalyzed the association between serum antibodyprofiles detected with antigen microarrays and MRImeasures of disease including cerebral atrophy andT2 hyperintense lesions We found that specificantibody patterns are associated with different aspectsof MRI-defined disease pathology Our major findingwas that increased reactivity to lipids is associated withdifferent aspects of brain MRI measures of diseaseseverity Of note the specific set of lipids associatedwith atrophy differed from those associated withlesions Taken together these data suggest that anti-lipid antibodies in serum are related to MRI measuresof disease Our findings highlight the important role oflipid and lipid-specific immunity in the pathogenesis ofMS
Although a potential limitation of these studies isthe relatively small number of samples analyzed wehave validated our findings in an independent valida-tion cohort strengthening their significance In addi-tion further studies in large cohorts from patientsaffected by non-MS neurodegenerative diseases andhealthy controls may indicate whether the antibodyreactivities and lipid indexes identified in these stud-ies are exclusive to MS or are associated with addi-tional biological processes Indeed some of theseantibody reactivities contributing to the immune sig-natures described in this work have been found innewborns45
Different sets of antibody reactivities were associ-ated with GMF WMF BPF and T2LV The differ-ent antibody link between lesions and atrophy is inkeeping with the long-held view that brain atrophyin patients with MS is multifactorial and is onlyweakly related to lesions41 This has been underscoredin recent studies showing the complementary infor-mation obtained by combining lesion and atrophymeasures4647
Among the cerebral atrophy measures there was astriking overlap between those antibodies associatedwith GMF and BPF which separated them fromWMF results These findings are in keeping with pre-vious observations that whole brain atrophy is domi-nated by GM loss3741 In the early stages of MS thisGM atrophy selectively involves the deep GM nuclei34
Thus the similarities observed in the autoantibodieslinked to GMF and BPF might reflect the dominantcontribution and colinearity of GM atrophy to totalbrain atrophy The set of antibodies linked to GMFmay be the most relevant given that MRI studies haveshown that GM volume is more closely linked to phys-ical disability37 and cognitive impairment38 than areWM volume or lesion measures
Using antigen microarrays antibody reactivity tolipids has been detected in the CSF and serum of
patients with MS at different stages of the diseaseand such antibodies in the CSF have been linkedto disease progression and MRI involvement44
Although several mechanisms are thought to drivebrain inflammation and atrophy in MS46 the connec-tion between these mechanisms and the antibodiesdetected with our antigen microarrays is yet unknownOne possibility is that the serum autoantibodies iden-tified in our studies are not pathogenic and reflect theresult of immunization against self-antigens releasedfrom the CNS during the course of the disease Indeedneurofilament is released from damaged axons duringthe course of MS and both neurofilament lightchains48 and antibodies against it42 have been linkedto MRI measures of disease Moreover heat shockproteins are upregulated in different cell types duringinflammation and have been shown to have an impor-tant role as immunomodulators when released to theextracellular medium4950 Thus it is possible that heatshock proteinndashreactive antibodies reflect changes in theproduction and release of these immunomodulatorsduring the course of disease pathogenesis Similarlybioactive lipids and their products are released fromdamaged myelin and lipid-specific antibodies havebeen detected in patients with MS27293051 Lipids havean important role in the immune response both asbioactive molecules with immunomodulatory proper-ties and also as targets of the adaptive immuneresponse52 Moreover lipids have significant effectson the murine model of MS experimental autoim-mune encephalomyelitis2729305153 Indeed we recentlyfound that the glycolipid lactosylceramide activates abroad set of biological processes in astrocytes promot-ing neurodegeneration and inflammation53 Thus thelipid-reactive antibodies detected in this work mayreflect the release of myelin lipids in the context ofMS pathogenesis andor may directly contribute toimmune-mediated damage in CNS tissue Of notelipid-reactive antibodies are highly cross-reactive52 Inaddition the lipid-reactive antibodies detected in thiswork as associated with MRI measures of disease activ-ity were of the IgG class These are important points toconsider when evaluating a potential role of lipid-reactive antibodies inMS pathogenesis Further studiesare warranted to investigate whether bioactive lipidsoffer new therapeutic targets in MS
We have found that unique patterns of immunereactivity determined with antibody arrays are associ-ated with specific MRI measures of disease severityThese patterns agree with the interpretation that dif-ferent pathogenic mechanisms drive diverse diseaseprocesses reflected by these MRI measures These pat-terns also suggest a predominant role for lipids andlipid-specific immunity in MS pathology Furtherstudies are warranted to determine whether the earlyappearance of anti-lipid antibodies predicts the
Neurology Neuroimmunology amp Neuroinflammation 9
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
subsequent development of clinical and MRI-defineddisease worsening
AUTHOR CONTRIBUTIONSRohit Bakshi draftingrevising the manuscript study concept or design
analysis or interpretation of data acquisition of data study supervision
obtaining funding Ada Yeste study concept or design acquisition of
data Bonny Patel analysis or interpretation of data statistical analysis
Shahamat Tauhid analysis or interpretation of data Subhash Tummala
analysis or interpretation of data Roya Rahbari analysis or interpretation
of data statistical analysis Renxin Chu analysis or interpretation of data
Keren Regev analysis or interpretation of data acquisition of data
Pia Kivisaumlkk draftingrevising the manuscript study concept or design
contribution of vital reagentstoolspatients acquisition of data study
supervision Howard L Weiner study concept or design obtaining fund-
ing Francisco J Quintana draftingrevising the manuscript study con-
cept or design contribution of vital reagentstoolspatients study
supervision obtaining funding
ACKNOWLEDGMENTThe authors thank Dr Mohit Neema Dr Antonella Ceccarelli and
Dr Ashish Arora for valuable assistance at the early stages of this project
STUDY FUNDINGThis work was supported in part by grants to FJQ and HLW from
EMD Serono RG4111A1 and JF2161-A-5 from the National Multiple
Sclerosis Society PA0069 from the International Progressive MS Alli-
ance and by grants from the NIH (5R01NS055083-04) and NMSS
(RG3798A2) to RB
DISCLOSUREFJ Quintana serves on the editorial board for Systems Biomedicine
Inmunologia American Journal of Clinical and Experimental Immunology
is an associate editor for Immunology (UK) is an advisory board member
for Seminars in Immunopathology received research support from Harvard
Medical School BADERC NMSS A Yeste and B Patel report no
disclosures S Tauhid is managing editor for Journal of Neuroimaging
S Tummala R Rahbari R Chu and K Regev report no disclosures
P Kivisaumlkk received research support from EMD Serono R Bakshi is
editor-in-chief for Journal of Neuroimaging received consulting fees from
AbbVie Alkermes Biogen Novartis Questcor received research support
from Biogen EMD-Serono Novartis Sanofi-Genzyme Teva his spouse
holds stock in Biogen Inc H Weiner served on the advisory board
for The Guthy-Jackson Charitable Foundation Teva Pharmaceuticals
Industries Ltd Biogen Idec Novartis Sanofi-Aventis has consulted for
Therapix Bioven Novartis Serono Teva Sanofi received research support
from National Multiple Sclerosis Society Go to Neurologyorgnn for full
disclosure forms
Received July 27 2015 Accepted in final form December 8 2015
REFERENCES1 Weiner HL The challenge of multiple sclerosis how do
we cure a chronic heterogeneous disease Ann Neurol
200965239ndash248
2 Bakshi R Thompson AJ Rocca MA et al MRI in mul-
tiple sclerosis current status and future prospects Lancet
Neurol 20087615ndash625
3 Ceccarelli A Bakshi R Neema M MRI in multiple scle-
rosis a review of the current literature Curr Opin Neurol
201225402ndash409
4 Filippi M Rocca MA Barkhof F et al Association
between pathological and MRI findings in multiple scle-
rosis Lancet Neurol 201211349ndash360
5 Geurts JJ Calabrese M Fisher E Rudick RA Measure-
ment and clinical effect of grey matter pathology in mul-
tiple sclerosis Lancet Neurol 2012111082ndash1092
6 Klaver R De Vries HE Schenk GJ Geurts JJ Grey mat-
ter damage in multiple sclerosis a pathology perspective
Prion 2013766ndash75
7 Khoury S Bakshi R Cerebral pseudoatrophy or real atro-
phy after therapy in multiple sclerosis Ann Neurol 2010
68778ndash779
8 Hauser SL Chan JR Oksenberg JR Multiple sclerosis
prospects and promise Ann Neurol 201374317ndash327
9 McFarland HF Martin R Multiple sclerosis a complicated
picture of autoimmunity Nat Immunol 20078913ndash919
10 Nylander A Hafler DA Multiple sclerosis J Clin Invest
20121221180ndash1188
11 Sospedra M Martin R Immunology of multiple sclerosis
Ann Rev Immunol 200523683ndash747
12 Steinman L Immunology of relapse and remission in mul-
tiple sclerosis Ann Rev Immunol 201432257ndash281
13 Khoury SJ Guttmann CR Orav EJ Kikinis R Jolesz FA
Weiner HL Changes in activated T cells in the blood
correlate with disease activity in multiple sclerosis Arch
Neurol 2000571183ndash1189
14 Khoury SJ Orav EJ Guttmann CR Kikinis R Jolesz FA
Weiner HL Changes in serum levels of ICAM and TNF-
R correlate with disease activity in multiple sclerosis Neu-
rology 199953758ndash764
15 Laplaud DA Berthelot L Miqueu P et al Serial blood T
cell repertoire alterations in multiple sclerosis patients cor-
relation with clinical and MRI parameters J Neuroimmunol
2006177151ndash160
16 Makhlouf K Weiner HL Khoury SJ Increased percentage
of IL-121 monocytes in the blood correlates with the
presence of active MRI lesions in MS J Neuroimmunol
2001119145ndash149
17 Prat A Biernacki K Saroli T et al Kinin B1 receptor expres-
sion on multiple sclerosis mononuclear cells correlation with
magnetic resonance imaging T2-weighted lesion volume and
clinical disability Arch Neurol 200562795ndash800
18 Rinaldi L Gallo P Calabrese M et al Longitudinal anal-
ysis of immune cell phenotypes in early stage multiple
sclerosis distinctive patterns characterize MRI-active pa-
tients Brain 20061291993ndash2007
19 Tortorella P Lagana MM Saresella M et al Determinants
of disability in multiple sclerosis an immunological and
MRI study Biomed Res Int 20142014875768
20 Robinson WH DiGennaro C Hueber W et al Autoan-
tigen microarrays for multiplex characterization of autoan-
tibody responses Nat Med 20028295ndash301
21 Yeste A Quintana FJ Antigen microarrays for the study of
autoimmune diseases Clin Chem 2013591036ndash1044
22 Li QZ Xie C Wu T et al Identification of autoantibody
clusters that best predict lupus disease activity using glomer-
ular proteome arrays J Clin Invest 20051153428ndash3439
23 Hueber W Kidd BA Tomooka BH et al Antigen micro-
array profiling of autoantibodies in rheumatoid arthritis
Arthritis Rheum 2005522645ndash2655
24 Goldschmidt Y Sharon E Quintana FJ Cohen IR
Brandt A Adaptive methods for classification of biological
microarray data from multiple experiments Report no
MCS03-07 Rehovot Israel The Arthur and Rochelle Bel-
fer Institute of Mathematics and Computer Science 2003
25 Quintana FJ Getz G Hed G Domany E Cohen IR
Cluster analysis of human autoantibody reactivities in
health and in type 1 diabetes mellitus a bio-informatic
approach to immune complexity J Autoimmun 2003
2165ndash75
10 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
26 Lalive PH Menge T Barman I Cree BA Genain CP
Identification of new serum autoantibodies in neuromye-
litis optica using protein microarrays Neurology 200667
176ndash177
27 Kanter JL Narayana S Ho PP et al Lipid microarrays
identify key mediators of autoimmune brain inflamma-
tion Nat Med 200612138ndash143
28 Quintana FJ Farez MF Izquierdo G Lucas M Cohen IR
Weiner HL Antigen microarrays identify CNS-produced
autoantibodies in RRMS Neurology 201278532ndash539
29 Farez MF Quintana FJ Gandhi R Izquierdo G Lucas M
Weiner HL Toll-like receptor 2 and poly(ADP-ribose)
polymerase 1 promote central nervous system neuroin-
flammation in progressive EAE Nat Immunol 200910
958ndash964
30 Quintana FJ Farez MF Viglietta V et al Antigen micro-
arrays identify unique serum autoantibody signatures in
clinical and pathologic subtypes of multiple sclerosis Proc
Natl Acad Sci USA 200810518889ndash18894
31 Quintana FJ Patel B Yeste A et al Epitope spreading as
an early pathogenic event in pediatric multiple sclerosis
Neurology 2014832219ndash2226
32 Gauthier SA Glanz BI Mandel M Weiner HL A model
for the comprehensive investigation of a chronic autoim-
mune disease the multiple sclerosis CLIMB study Auto-
immun Rev 20065532ndash536
33 Polman CH Reingold SC Edan G et al Diagnostic criteria
for multiple sclerosis 2005 revisions to the ldquoMcDonald
Criteriardquo Ann Neurol 200558840ndash846
34 DellrsquoOglio E Ceccarelli A Glanz BI et al Quantification
of global cerebral atrophy in multiple sclerosis from 3T
MRI using SPM the role of misclassification errors
J Neuroimaging 201525191ndash199
35 Ceccarelli A Jackson JS Tauhid S et al The impact of
lesion in-painting and registration methods on voxel-based
morphometry in detecting regional cerebral gray matter
atrophy in multiple sclerosis AJNR Am J Neuroradiol
2012331579ndash1585
36 Cohen AB Neema M Arora A et al The relationships
among MRI-defined spinal cord involvement brain involve-
ment and disability in multiple sclerosis J Neuroimaging
201222122ndash128
37 Sanfilipo MP Benedict RH Sharma J Weinstock-
Guttman B Bakshi R The relationship between whole
brain volume and disability in multiple sclerosis a com-
parison of normalized gray vs white matter with misclas-
sification correction Neuroimage 2005261068ndash1077
38 Sanfilipo MP Benedict RH Weinstock-Guttman B
Bakshi R Gray and white matter brain atrophy and neuro-
psychological impairment in multiple sclerosis Neurology
200666685ndash692
39 Stankiewicz JM Glanz BI Healy BC et al Brain MRI
lesion load at 15T and 3T versus clinical status in multi-
ple sclerosis J Neuroimaging 201121e50ndashe56
40 Quintana FJ Hagedorn PH Elizur G Merbl Y
Domany E Cohen IR Functional immunomics micro-
array analysis of IgG autoantibody repertoires predicts the
future response of mice to induced diabetes Proc Natl
Acad Sci USA 2004101(suppl 2)14615ndash14621
41 Bermel RA Bakshi R The measurement and clinical
relevance of brain atrophy in multiple sclerosis Lancet
Neurol 20065158ndash170
42 Eikelenboom MJ Petzold A Lazeron RH et al Multiple
sclerosis neurofilament light chain antibodies are corre-
lated to cerebral atrophy Neurology 200360219ndash223
43 Magraner MJ Bosca I Simo-Castello M et al Brain atro-
phy and lesion load are related to CSF lipid-specific IgM
oligoclonal bands in clinically isolated syndromes Neuro-
radiol 2012545ndash12
44 Villar LM Sadaba MC Roldan E et al Intrathecal syn-
thesis of oligoclonal IgM against myelin lipids predicts an
aggressive disease course in MS J Clin Invest 2005115
187ndash194
45 Merbl Y Zucker-Toledano M Quintana FJ Cohen IR
Newborn humans manifest autoantibodies to defined self
molecules detected by antigen microarray informatics
J Clin Invest 2007117712ndash718
46 Sormani MP Arnold DL De Stefano N Treatment effect
on brain atrophy correlates with treatment effect on disa-
bility in multiple sclerosis Ann Neurol 20147543ndash49
47 Tauhid S Neema M Healy BC Weiner HL Bakshi R
MRI phenotypes based on cerebral lesions and atrophy in
patients with multiple sclerosis J Neurol Sci 2014346
250ndash254
48 Gnanapavan S Grant D Morant S et al Biomarker
report from the phase II lamotrigine trial in secondary
progressive MS neurofilament as a surrogate of disease
progression PLoS One 20138e70019
49 Quintana FJ Cohen IR Heat shock proteins as endoge-
nous adjuvants in sterile and septic inflammation
J Immunol 20051752777ndash2782
50 Quintana FJ Cohen IR The HSP60 immune system net-
work Trends Immunol 20113289ndash95
51 Ho PP Kanter JL Johnson AM et al Identification of
naturally occurring fatty acids of the myelin sheath that
resolve neuroinflammation Sci Transl Med 20124137ra73
52 Quintana FJ Yeste A Weiner HL Covacu R Lipids and
lipid-reactive antibodies as biomarkers for multiple sclero-
sis J Neuroimmunol 201224853ndash57
53 Mayo L Trauger SA Blain M et al Regulation of astro-
cyte activation by glycolipids drives chronic CNS inflam-
mation Nat Med 2014201147ndash1156
Neurology Neuroimmunology amp Neuroinflammation 11
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
1
Table e-1 Antigens used
Group Antigen Source
27 kDa Heat Shock Protein Stressgen
HSP 32 kDa Heat Shock Protein Stressgen
40 kDa Heat Shock Protein Stressgen
47 kDa Heat Shock Protein Stressgen
60 kDa Heat Shock Protein Stressgen
60 kDa Heat Shock Protein peptide aa 106ndash125 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 121ndash140 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 136ndash155 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 151ndash170 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 16ndash35 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 166ndash185 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
2
60 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 556ndash573 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
65 kDa Heat Shock Protein M tuberculosis Stressgen
70 kDa Heat Shock Protein Stressgen
70 kDa Heat Shock Protein peptide aa 106ndash125 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 121ndash140 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 136ndash155 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 151ndash170 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 16ndash35 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 166ndash185 Biopolymers Facility Harvard Medical School
3
70 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
4
70 kDa Heat Shock Protein peptide aa 556ndash575 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 571ndash590 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 586ndash605 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 601ndash620 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 616ndash635 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 631ndash640 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
71 kDa Heat Shock Protein M tuberculosis Biopolymers Facility Harvard Medical School
90 kDa Heat Shock Protein Stressgen
GroEL Stressgen
23-cyclic nucleotide 3-phosphodiesterase
peptide aa106-125 Biopolymers Facility Harvard Medical School
CNS
23-cyclic nucleotide 3-phosphodiesterase
peptide aa1-20 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa121-140 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa136-155 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa151-170 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa16-35 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa166-185 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase Biopolymers Facility Harvard Medical School
5
peptide aa181-200
23-cyclic nucleotide 3-phosphodiesterase
peptide aa196-215 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa211-230 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa226-245 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa241-260 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa256-275 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa271-290 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa286-305 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa301-320 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa31-50 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa316-335 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa331-350 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa346-365 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa361-380 Biopolymers Facility Harvard Medical School
6
23-cyclic nucleotide 3-phosphodiesterase
peptide aa376-395 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa391-410 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa406-421 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa46-65 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa61-80 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa76-95 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa91-110 Biopolymers Facility Harvard Medical School
Acetyl Cholinesterase Sigma Aldrich
ADAM-10 Sigma Aldrich
alpha-Cristallin Stressgen
beta-Cristallin Sigma Aldrich
bovine Myelin Basic Protein Sigma Aldrich
Brain Extract I Sigma Aldrich
Brain Extract II Sigma Aldrich
Brain Extract III Sigma Aldrich
Glial Filament Acidic Protein Research Diagnostic
guinea pig Myelin Basic Protein Sigma Aldrich
human Myelin Basic Protein Sigma Aldrich
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 106-125 Biopolymers Facility Harvard Medical School
7
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 121-140 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 136-155 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 151-170 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 166-185 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 181-200 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 91-110 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
106-125 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa 1- Biopolymers Facility Harvard Medical School
8
20
Myelinoligodendrocyte glycoprotein peptide aa
121-140 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
136-155 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
151-170 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
16-35 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
166-185 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
181-200 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
196-215 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
211-230 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
226-247 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
31-50 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
35-55 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
46-65 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
61-80 Biopolymers Facility Harvard Medical School
9
Myelinoligodendrocyte glycoprotein peptide aa
76-95 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
91-110 Biopolymers Facility Harvard Medical School
murine Myelin Basic Protein Sigma Aldrich
Myelin Associated Glycoprotein Sigma Aldrich
Myelin Basic Protein peptide aa 104-123 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 11-30 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 113-132 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 121-138 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 124-142 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 138-147 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 141-161 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 143-168 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 155-178 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 26-35 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 41-60 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 51-70 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 71-92 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 84-94 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 89-101 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 173-186 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 93-112 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 106-125 Biopolymers Facility Harvard Medical School
10
Myelin Protein 2 peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 121-132 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 91-110 Biopolymers Facility Harvard Medical School
Neurofilament 160kd Chemicon
Neurofilament 200kd Chemicon
Neurofilament 68kd Chemicon
Neuronal Enolase Calbiochem
Nicastrin GeneTex
NMDA receptor Novus Biologicals
NOGO Sigma Aldrich
Olygodendrocyte-Specific Protein peptide aa 106ndash
125 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 121ndash
140 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 136ndash
155 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 151ndash
170 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 16ndash
35 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 166ndash Biopolymers Facility Harvard Medical School
11
185
Olygodendrocyte-Specific Protein peptide aa 181ndash
199 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 195ndash
217 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 31ndash
50 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 46ndash
65 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 61ndash
80 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 76ndash
95 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 91ndash
110 Biopolymers Facility Harvard Medical School
Proteolipid Protein Abnova
Proteolipid Protein peptide aa 100-119 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 10-29 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 110-129 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 1-19 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 125-141 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-150 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-154 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 150-163 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 151-173 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 158-166 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 161-180 Biopolymers Facility Harvard Medical School
12
Proteolipid Protein peptide aa 178-191 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 180-199 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 190-209 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 20-39 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 205-220 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 215-232 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-239 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-249 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 248-259 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 250-269 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 265-277 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 35-50 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 40-59 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 50-69 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 65-84 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 80-99 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 91-110 Biopolymers Facility Harvard Medical School
Retinol Binding Protein Sigma Aldrich
S100beta protein Assay Designs
Super Oxide Dismutase Sigma Aldrich
Synuclein beta Sigma Aldrich
Synuclein gamma Sigma Aldrich
Amydgala ProSci Inc
Tissue Amydgala AD ProSci Inc
Brain lysate ProSci Inc
Brain Tissue Membrane ProSci Inc
Cerebellar pedunculus ProSci Inc
13
Cerebral meninges ProSci Inc
Corpus Callosum ProSci Inc
Corpus Callosum AD ProSci Inc
Diencephalon ProSci Inc
Fetal brain ProSci Inc
Frontal lobe ProSci Inc
Frontal lobe AD ProSci Inc
Hippocampus ProSci Inc
Hippocampus AD ProSci Inc
Insula ProSci Inc
Occipital lobe ProSci Inc
Occipital lobe AD ProSci Inc
Olfactory region ProSci Inc
Optic Nerve ProSci Inc
Parietal lobe ProSci Inc
Parietal lobe AD ProSci Inc
Pons ProSci Inc
Pons AD ProSci Inc
Postcentral gyrus ProSci Inc
Postcentral gyrus AD ProSci Inc
Precentral gyrus ProSci Inc
Precentral gyrus AD ProSci Inc
Spinal cord ProSci Inc
Temporal lobe ProSci Inc
Temporal lobe AD ProSci Inc
Thalamus ProSci Inc
Thalamus AD ProSci Inc
14
Amyloid beta Sigma Aldrich
AD
related Amyloid beta 10-20 Sigma Aldrich
Amyloid beta 1-12 Sigma Aldrich
Amyloid beta 12-28 Sigma Aldrich
Amyloid beta 1-23 Sigma Aldrich
Amyloid beta 1-38 Sigma Aldrich
Amyloid beta 17-40 Sigma Aldrich
Amyloid beta 25-35 Sigma Aldrich
Amyloid beta 34-42 Sigma Aldrich
Amyloid bri protein precursor 227 Sigma Aldrich
Amyloid DAN Protein Fragment 1-34 Sigma Aldrich
Amyloid Precursor Protein Sigma Aldrich
Amyloid protein no AB component Sigma Aldrich
Secreted amyloid precursor protein (SAP) beta Sigma Aldrich
Tau isoform variant 0N3R Sigma Aldrich
Tau isoform variant 1N3R Sigma Aldrich
Tau isoform variant 0N4R Sigma Aldrich
Tau isoform variant 2N3R Sigma Aldrich
Tau phospho Ser412 Sigma Aldrich
Tau phospho Ser441 Sigma Aldrich
Tau phospho Thr181 Sigma Aldrich
Tau Protein human EMD Biosciences
(plusmn)9-HODE Cayman Chemical
Lipids
1 Palmitoyl-2-(5oxo-Valeroyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
15a-hydroxycholestene Avanti Polar Lipids
15
15-ketocholestane Avanti Polar Lipids
15-ketocholestene Avanti Polar Lipids
1-Palmitoil-2-(9oxo-Nonanoyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Azelaoyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Glutaroyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
5 α-cholestane-3 β15 α-diol Avanti Polar Lipids
9(S)-HODE Cayman Chemical
Asialoganglioside-GM1 Sigma Aldrich
Asialoganglioside-GM2 Sigma Aldrich
Brain ceramides Avanti Polar Lipids
Brain D-erythrosphingosine Avanti Polar Lipids
Brain lysophosphatidylethanolamine Avanti Polar Lipids
Brain L-α-lysophosphatidylserine Avanti Polar Lipids
Brain L-α-phosphatidylcholine Avanti Polar Lipids
Brain L-α-phosphatidyl-ethanolamine Avanti Polar Lipids
Brain L-α-phosphatidylserine Avanti Polar Lipids
Brain polar lipid extract Avanti Polar Lipids
Brain sphingomyelin Avanti Polar Lipids
Brain sulfatide Avanti Polar Lipids
Brain total lipid extract Avanti Polar Lipids
Cardiolipin Sigma Aldrich
Ceramide Sigma Aldrich
Ceramide 1-phosphate Sigma Aldrich
Cholesterol Sigma Aldrich
16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-

Rohit Bakshi MDAda Yeste PhDBonny Patel MScShahamat Tauhid MDSubhash Tummala MDRoya Rahbari PhDRenxin Chu MDKeren Regev MDPia Kivisaumlkk MD PhDHoward L Weiner MDFrancisco J Quintana
PhD
Correspondence toDr Quintanafquintanaricsbwhharvardedu
Supplemental dataat Neurologyorgnn
Serum lipid antibodies are associated withcerebral tissue damage in multiple sclerosis
ABSTRACT
Objective To determine whether peripheral immune responses as measured by serum antigenarrays are linked to cerebral MRI measures of disease severity in multiple sclerosis (MS)
Methods In this cross-sectional study serum samples were obtained from patients withrelapsing-remitting MS (n 5 21) and assayed using antigen arrays that contained 420 antigensincluding CNS-related autoantigens lipids and heat shock proteins Normalized compartment-specific global brain volumes were obtained from 3-tesla MRI as surrogates of atrophy includinggray matter fraction (GMF) white matter fraction (WMF) and total brain parenchymal fraction(BPF) Total brain T2 hyperintense lesion volume (T2LV) was quantified from fluid-attenuatedinversion recovery images
Results We found serum antibody patterns uniquely correlated with BPF GMF WMF and T2LVFurthermore we identified immune signatures linked to MRI markers of neurodegeneration (BPFGMF WMF) that differentiated those linked to T2LV Each MRI measure was correlated with aspecific set of antibodies Strikingly immunoglobulin G (IgG) antibodies to lipids were linked tobrain MRI measures Based on the association between IgG antibody reactivity and each uniqueMRI measure we developed a lipid index This comprised the reactivity directed against all of thelipids associated with each specific MRI measure We validated these findings in an additionalindependent set of patients with MS (n 5 14) and detected a similar trend for the correlationsbetween BPF GMF and T2LV vs their respective lipid indexes
Conclusions We propose serum antibody repertoires that are associated with MRI measuresof cerebral MS involvement Such antibodies may serve as biomarkers for monitoring diseasepathology and progression Neurol Neuroimmunol Neuroinflamm 20163e200 doi 101212
NXI0000000000000200
GLOSSARYBPF 5 brain parenchymal fraction GM 5 gray matter GMF 5 gray matter fraction IgG 5 immunoglobulin G MS 5 multiplesclerosis T2LV 5 T2 hyperintense lesion volume WM 5 white matter WMF 5 white matter fraction
Multiple sclerosis (MS) is characterized by immune dysfunction and inflammation leading tofocal lesions brain and spinal cord atrophy and progressive neurologic dysfunction The knownheterogeneity likely reflects myriad and complex underlying pathogenic mechanisms that makespecific and unique contributions to MS1
MRI-defined T2 hyperintense brain lesions are key to diagnosis and therapeutic monitoringHowever such lesions are nonspecific for the underlying pathology and have limited clinicalpredictive value23 Measurement of brain atrophy provides the potential to detect destructivedisease effects and show better associations with clinical status than can be obtained with lesionmeasures2 Atrophy begins early in MS and can be monitored by MRI segmentation2ndash4 Graymatter (GM) atrophy is more closely linked to clinical status than white matter (WM) atrophy
From the Partners Multiple Sclerosis Center (RB S Tauhid S Tummala RC HLW) and Ann Romney Center for Neurologic Diseases (RBAY BP RR KR PK HLW FJQ) Neurology (RB AY BP S Tauhid S Tummala RR RC KR PK HLW FJQ) andRadiology (RB) Brigham and Womenrsquos Hospital Harvard Medical School Boston MA
Funding information and disclosures are provided at the end of the article Go to Neurologyorgnn for full disclosure forms The Article ProcessingCharge was paid by the authors
This is an open access article distributed under the terms of the Creative Commons Attribution-NonCommercial-NoDerivatives License 40 (CCBY-NC-ND) which permits downloading and sharing the work provided it is properly cited The work cannot be changed in any way or usedcommercially
Neurologyorgnn copy 2016 American Academy of Neurology 1
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
whole brain atrophy or conventional lesionassessments56 This likely reflects the func-tional importance of GM and the contentionthat pseudoatrophy confounds the use ofwhole brain or WM atrophy to monitor pro-gressive neurodegeneration7
Immune processes have a central role inboth the pathogenesis and treatment ofMS8ndash12 The ability to link such changes toMRI presents the opportunity to providenew biomarkers and better understanding ofdisease pathophysiology13ndash19
Antigen microarrays are newly developedtools for the high-throughput characterizationof the immune response2021 that have beenused to identify biomarkers and mechanismsof disease pathogenesis in several autoimmunedisorders including MS22ndash31 In the presentstudy we investigated the relationship betweenantigen arrays and both GM and WM cerebralMRI involvement in MS
METHODS Patients Table 1 summarizes the patientsrsquo
demographic and clinical characteristics of the discovery and val-
idation sets All serum samples were collected from the ongoing
cohort of patients being followed in the CLIMB study (Compre-
hensive Longitudinal Investigation of MS at Brigham and
Womenrsquos Hospital32) in which participants are followed with
comprehensive clinical and imaging assessments to monitor
disease progression and response to therapy on a yearly basis
Samples were collected within (mean 6 SD) 50 6 32 months
of MRI acquisition Patients were free of relapses or changes in
disease-modifying therapy during the interval between blood
collection and MRI This was a consecutive sample meeting the
following criteria (1) age 18 to 55 years (2) diagnosis of
relapsing-remitting MS33 (3) absence of other major medical
neurologic or neuropsychiatric disorders (4) lack of any relapse
or corticosteroid use in the 4 weeks before MRI or start of disease-
modifying therapy 6 months before MRI (to reduce confounding
effects on MRI) and (5) no history of smoking or substance
abuse The majority of patients were receiving disease-
modifying treatment at the time of MRI Within 3 months of
MRI each patient received an examination by an MS specialist-
neurologist including evaluation of neurologic disability on the
Expanded Disability Status Scale and a timed 25-foot walk
Standard protocol approvals registrations and patientconsents Our study received approval from the ethical standards
committee on human experimentation at our institution
(The Partners Health Care Institutional Review Board) All par-
ticipants gave written informed consent for their participation in
the study
MRI acquisition and analysis All participants in the discoveryset underwentMRI on the same scanner (3T Signa General Electric
Healthcare Milwaukee WI) using a receive-only phase array head
coil with the same MRI protocol The scan acquisition protocol
has been detailed previously34 Contiguous slices covering the
whole brain were acquired in high-resolution protocols using
3-dimensional modified driven equilibrium Fourier transform and
T2-weighted fast fluid-attenuated inversion recovery sequences
Patients in the validation set underwent brain MRI on a 15T
scanner (GE Signa) including a 2-dimensional axial conventional
spin-echo dual-echo T2-weighted series (voxel sizes 0943 0943
3 mm) Analysis of these scans was performed by operators who
were unaware of clinical and biomarker information In the
discovery set we obtained normalized compartment-specific
global brain volumes as surrogates of atrophy including GM
fraction (GMF) WM fraction (WMF) and total brain
parenchymal fraction (BPF) using statistical parametric mapping
version 8 (WellcomeDepartment of Cognitive Neurology London
UK httpwwwfilionuclacukspm) after manual correction of
(1) misclassifications of tissue compartments due to MS lesion and
(2) ineffective contouring of the deep GM structures34 In the
validation set BPF and GMF were obtained in statistical
parametric mapping version 8 from the dual-echo images
Because the source images did not show effective contrast for
segmentation of the deep gray structures we performed manual
masking to derive only the cerebral cortical GMF Quantification
of total brain T2 hyperintense lesion volume (T2LV) was performed
using Jim (Xinapse Systems Ltd West Bergholt UK httpwww
xinapsecom) by the consensus of 2 experienced observers from the
fluid-attenuated inversion recovery (discovery set) or dual-echo
(validation set) images using a semiautomated technique For the
measurement of these atrophy and lesion surrogates from MRI
scans our methods are well established regarding their operational
procedures validity and reliability34ndash39
Antigens Peptides were synthesized at the Biopolymers Facility
of the Department of Biological Chemistry and Molecular Phar-
macology of Harvard Medical School Recombinant proteins and
lipids were purchased from Sigma (St Louis MO) Abnova
(Taipei City Taiwan) Matreya LLC (Pleasant Gap PA) Avanti
Polar Lipids (Alabaster AL) Calbiochem (San Diego CA)
Chemicon (Temecula CA) GeneTex (San Antonio TX) Novus
Table 1 Demographic clinical and brain MRI data
Discovery seta Validation setb
No of patients with relapsing-remitting MS 21 14
Age y 406 6 80 471 6 81
Women n () 14 (67) 13 (93)
Disease duration y 68 6 50 127 6 81
EDSS score 14 6 12 17 6 19
Timed 25-ft walk s 47 6 06 83 6 102
BPFc 083 6 004 076 6 005
GMFc 049 6 004d 033 6 003e
WMF 033 6 002 NP
T2LVc mL 143 6 169 49 6 79
Abbreviations BPF 5 whole brain parenchymal fraction EDSS 5 Expanded Disability Sta-tus Scale GMF 5 cerebral gray matter fraction MS 5 multiple sclerosis NP 5 not per-formed T2LV 5 cerebral T2 hyperintense lesion volume WMF 5 global cerebral whitematter fractionValues represent mean 6 SD unless otherwise indicateda Fluid-attenuated inversion recovery 3T high resolutionbDual-echo 15T low resolutionc The 2 groups had different MRI acquisitionsource images (3T high resolution vs 15T lowresolution) and different software analysis pipelines leading to a difference in scalingbetween the MRI output metrics (see the methods section for more details)dWhole brain GMFeCortical GMF
2 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
Table 2 Serum immunoglobulin Gs associated with MRI measures of disease severity
Antigens
BPF GMF WMF T2LV
r p FDR r p FDR r p FDR r p FDR
1-P-2-5-o-V-sn-G-3-PC 2038 0087 0161 2034 0131 0197 2014 0540 1000 056 0008a 0056
15-ketocholestene 2030 0190 0370 2044 0047b 0066 024 0302 0854 030 0189 0247
9 HODE 2040 0073 0097 2051 0019b 0026b 016 0499 1000 045 0043a 0057
9 S-HODE 2042 0057 0067 2048 0027b 0026b 006 0797 1000 061 0003a 0033a
a-MSH 050 0021a 0040a 040 0070 0112 028 0214 0854 2004 0858 1000
ABPF 1-12 2023 0320 0610 2040 0070 0112 032 0155 0804 046 0035a 0057
ABPP 227 2030 0179 0305 2056 0009b 0025b 047 0033a 0156 049 0024a 0056
Ac phosphatase 035 0119 0194 047 0033a 0027a 2018 0443 1000 2020 0374 0516
ADPF 1-34 2020 0393 0823 2038 0089 0133 034 0126 0804 046 0035a 0057
ANP 013 0587 0969 037 0100 0197 2048 0027b 0142 2027 0242 0516
Asialoganglioside-GM1 2052 0016b 0040b 2060 0004b 0015b 007 0749 1000 032 0152 0209
Asialoganglioside-GM2 2040 0074 0097 2048 0027b 0026b 011 0648 1000 024 0292 0516
b-MSH 2047 0033b 0058 2038 0085 0133 2025 0277 0854 020 0376 0516
b-Amyloid 2021 0350 0610 2036 0107 0197 026 0247 0854 049 0025a 0056
b-Synuclein 001 0954 0969 2022 0343 0996 048 0029a 0142 042 0055 0094
bNGF 029 0204 0413 006 0783 0996 051 0018a 0122 2006 0807 1000
Brain D-erythrosphingosine 2047 0030b 0058 2056 0009b 0025b 010 0679 1000 037 0096 0146
Brain extract VII 2052 0016b 0040b 2059 0005b 0015b 006 0789 1000 053 0013a 0056
Brain L-a-phosphatidyl-ethanolamine 2044 0044b 0067 2046 0037b 0027b 2005 0837 1000 019 0407 0537
Brain polar lipid extract 013 0573 0969 2010 0657 0996 051 0019a 0122 018 0434 0537
Chorionic G 005 0826 0969 028 0223 0303 2046 0034b 0156 2010 0656 1000
CNPase aa 106ndash125 039 0080 0161 049 0026a 0026a 2014 0557 1000 2051 0017b 0056
CNPase aa 16ndash35 042 0056 0067 046 0037a 0027a 2002 0941 1000 2037 0103 0157
CNPase aa 195ndash214 031 0173 0305 050 0020a 0026a 2035 0119 0804 2035 0118 0157
CNPase aa 286ndash305 028 0227 0413 045 0039a 0027a 2033 0144 0804 2004 0873 1000
CNPase aa 376ndash395 019 0416 0823 043 0049a 0066 2049 0024b 0142 2038 0089 0146
Collagen II 025 0278 0610 046 0034a 0027a 2040 0072 0788 2032 0151 0209
Collagen IX 2067 0001b 0025b 2075 0001b 0002b 008 0741 1000 066 0001a 0033a
Collagen VIII 2027 0242 0521 2026 0251 0605 2005 0831 1000 044 0044a 0057
Collagen X 2036 0114 0194 2012 0595 0996 2052 0016b 0122 2006 0792 1000
Disialoganglioside-GD2 2040 0073 0097 2049 0023b 0026b 013 0589 1000 043 0055 0094
Disialogang-GD1B 2038 0088 0161 2050 0022b 0026b 018 0441 1000 031 0174 0247
Disialoganglioside GD1a 2051 0017b 0040b 2053 0013b 0026b 2005 0824 1000 042 0060 0146
Disialoganglioside GD3 2040 0069 0097 2054 0012b 0026b 020 0382 1000 051 0018a 0056
Galactocerebrosides 001 0952 0969 2027 0229 0303 059 0005a 0108 025 0265 0516
Ganglioside mixture 2053 0014b 0040b 2062 0003b 0015b 011 0644 1000 053 0014a 0056
Ganglioside-GM4 002 0929 0969 029 0203 0303 2056 0009b 0109 2018 0423 0537
Gelsolin 053 0013a 0040a 057 0008a 0025a 002 0934 1000 2048 0029b 0056
GNRH 2053 0014b 0040b 2043 0054 0072 2029 0196 0854 036 0113 0157
gpMBP 017 0449 0928 2004 0857 1000 048 0029a 0142 000 0988 1000
GT1a 2043 0053 0067 2048 0028b 0026b 003 0912 1000 032 0161 0247
Hemoglobin 015 0527 0969 2006 0784 0996 046 0037a 0203 028 0212 0516
HSP60 aa 195ndash214 023 0311 0610 004 0879 1000 044 0045a 0206 000 0989 1000
HSP60 aa 210ndash229 053 0015a 0040a 049 0024a 0026a 016 0493 1000 2013 0577 1000
ContinuedNeurology Neuroimmunology amp Neuroinflammation 3
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
Table 2 Continued
Antigens
BPF GMF WMF T2LV
r p FDR r p FDR r p FDR r p FDR
HSP60 aa 240ndash259 048 0028a 0058 052 0017a 0026a 000 0990 1000 2047 0030b 0056
HSP60 aa 421ndash440 010 0655 0969 039 0080 0133 2058 0006b 0108 2022 0334 0516
HSP60 aa 436ndash455 042 0061 0067 060 0004a 0015a 2031 0173 0804 2038 0094 0146
HSP70 aa 121ndash140 044 0046a 0067 036 0111 0197 024 0289 0854 2016 0496 0728
HSP70 aa 181ndash199 051 0017a 0040a 048 0027a 0026a 015 0525 1000 2020 0378 0516
HSP70 aa 210ndash229 006 0788 0969 027 0228 0303 2044 0045b 0206 2011 0640 1000
HSP70 aa 316ndash335 2028 0216 0413 2029 0209 0303 2004 0879 1000 044 0048a 0072
HSP70 aa 376ndash395 2042 0059 0067 2046 0035b 0027b 002 0942 1000 035 0121 0157
HSP70 aa 421ndash440 048 0030a 0058 065 0002a 0015a 2027 0229 0854 2034 0137 0209
HSP70 aa 61ndash80 2002 0941 0969 2029 0198 0303 056 0008a 0109 018 0441 0537
IL-12 2050 0021b 0040b 2045 0040b 0027b 2018 0429 1000 039 0083 0146
IL-3 2041 0063 0067 2051 0019b 0026b 013 0575 1000 052 0017a 0056
Insula 046 0038a 0067 033 0143 0303 034 0133 0804 2017 0473 0550
Intrinsic factor 036 0108 0194 044 0048a 0066 2009 0702 1000 000 0986 1000
Isoprostane F2a-I 2053 0014b 0040b 2055 0010b 0025b 2004 0854 1000 035 0115 0157
Lactosylceramide 2021 0368 0630 2041 0062 0112 039 0077 0788 060 0004a 0033a
LIF 2065 0002b 0025b 2059 0005b 0015b 2022 0336 0854 041 0064 0146
MMP2 042 0055 0067 061 0003a 0015a 2032 0153 0804 2039 0082 0146
MMP9 2055 0009b 0040b 2062 0003b 0015b 006 0804 1000 045 0038a 0057
MOG peptide aa 196ndash215 2005 0837 0969 2008 0736 0996 005 0836 1000 044 0045a 0057
Monosialogang GM2 2056 0008b 0040b 2048 0028b 0026b 2026 0260 0854 017 0460 0550
Myelin protein 2 peptide aa 31ndash50 035 0124 0194 048 0029a 0027a 2022 0337 0854 2033 0141 0209
Myelin protein 2 peptide aa 91ndash110 2009 0694 0969 014 0533 0996 2051 0019b 0122 2004 0878 1000
Neurofilament 160 kDa 032 0162 0305 039 0082 0133 2009 0688 1000 2049 0026b 0056
Neurofilament 200 kDa 043 0050 0067 045 0040a 0027a 003 0881 1000 2015 0516 0728
Neurofilament 68 kDa 000 0999 1000 2021 0359 0996 044 0044a 0206 045 0043a 0057
NOGO 016 0493 0928 2012 0617 0996 060 0004a 0108 021 0352 0516
Nonhydroxy fatty acid ceramide 2053 0014b 0040b 2059 0005b 0015b 005 0826 1000 044 0044a 0057
Occipital lobe 2045 0039b 0067 2043 0054 0072 2012 0599 1000 039 0083 0146
ORF26 protein 053 0013a 0040a 052 0015a 0026a 010 0653 1000 2030 0182 0247
Parietal lobe 049 0023a 0040a 058 0005a 0015a 2011 0641 1000 2025 0283 0516
PDGF-Ra 2007 0752 0969 017 0461 0996 2052 0016b 0122 2009 0687 1000
Pepstatin 2035 0116 0194 2046 0036b 0027b 016 0483 1000 025 0277 0516
Postcentral gyrus AD 044 0044a 0067 034 0132 0197 028 0214 0854 2012 0603 1000
Proteolipid protein peptide aa 125ndash141 2009 0685 0969 010 0678 0996 2041 0064 0788 2048 0029b 0056
Proteolipid protein peptide aa 150ndash163 044 0043a 0067 051 0019a 0026a 2006 0809 1000 2050 0020b 0056
Proteolipid protein peptide aa 161ndash180 022 0343 0610 031 0166 0303 2017 0475 1000 2044 0045b 0057
Proteolipid protein peptide aa 250ndash269 037 0103 0194 050 0020a 0026 2023 0321 0854 2037 0098 0146
RBP 025 0274 0610 029 0201 0303 2005 0844 1000 2057 0007b 0056
Ribonuclease 2002 0937 0969 2017 0472 0996 031 0175 0804 054 0011a 0056
S100b 033 0141 0258 011 0649 0996 052 0015a 0122 2022 0344 0516
Spectrin 048 0026a 0058 035 0122 0197 036 0105 0804 2001 0966 1000
Sulfatides 2047 0033b 0058 2049 0023b 0026b 2002 0922 1000 034 0133 0209
Tetrasialoganglioside-GQ1B 2043 0054 0067 2057 0007b 0025b 023 0319 0854 048 0027a 0056
Continued4 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
Biologicals (Littleton CO) Assay Designs (Ann Arbor MI) Pro-
Sci Inc (Poway CA) EMD Biosciences (San Diego CA) Cay-
man Chemical (Ann Arbor MI) HyTest (Turku Finland)
Meridian Life Science (Memphis TN) and Biodesign Interna-
tional (Saco ME) The antigens used in the construction of anti-
gen microarrays are listed in table e-1 at Neurologyorgnn
Antigen microarray production development and dataanalysis The antigens listed in table e-1 were spotted in replicatesof 6 on SuperEpoxy 2 slides (TeleChem Sunnyvale CA) using
an Arrayit NanoPrint 2 LM210 microarray printer (Arrayit Corpo-
ration Sunnyvale CA) and optimized spotting conditions as
described28303140 The microarrays were hybridized using an HS
4800Pro Hybridization Station (Tecan Maumlnnedorf Switzerland)
in which they were blocked with 1 bovine serum albumin for
1 hour and incubated for 2 hours at 37degC with the test serum at
a 1100 dilution in blocking buffer The arrays were then washed
and incubated for 45 minutes with a 1500 dilution of goat
anti-human immunoglobulin G (IgG) Cy3-conjugated and
goat anti-human IgM Cy5-conjugated detection antibodies (Jackson
ImmunoResearch Labs West Grove PA) The arrays were scanned
with a Tecan PowerScanner Repeated measurements indicate that
our antigen microarray technique is reproducible exhibiting
a coefficient of variation of 1336 1231
Background signal was subtracted and raw data were normal-
ized and analyzed using the GeneSpring software (Silicon Genetics
Redwood City CA) Antigen reactivity was defined by the mean
intensity of binding to the replicates of that antigen on the micro-
array and expressed as relative fluorescence units Scatter plots were
generated using linear regression models in the R statistical package
Pearson product-moment correlation coefficients were calculated
between the MRI measures (BPF GMF WMF T2LV) and the
weighted average of a group of statistically significant lipids The
weighted average of significant lipids was calculated using the for-
mula (SWiAiSWi) (SWi 5 1 i 5 1 2 3 ) where Wi is the
proportion of intensity of antigen Ai Ai is the observed intensity of
an antigen and SWi is the sum of the weights
RESULTS Serum IgG antibodies correlate with brain
MRI measures of disease severity To study the relation-ship between the peripheral immune response andMRI measures of disease severity we analyzed serum
antibody reactivity in those MS samples We analyzedIgG serum antibodies using a panel of antigensincluding CNS antigens heat shock proteins and lip-ids The association between the antibody reactivityagainst each antigen and 4 MRI measures of disease(T2LV BPF WMF and GMF) was investigatedusing Spearman correlation tests
We found significant associations between eachMRI measure of disease and different sets of IgG anti-body reactivity which are shown in table 2 Similarpatterns of antibody reactivity were linked to BPFand GMF consistent with the known dominant con-tribution of GM atrophy to whole brain atrophymeasures3741 Strikingly there was little overlapbetween the antibody reactivity associated withGMF and WMF suggesting that different immuno-pathologic processes contribute to tissue degenerationin these areas of the brain Similarly these profiles ofantibody reactivity were also different from thoseassociated with T2LV
Serum lipid-reactive IgGs are associated with increased
disease pathogenesis determined by MRI In evaluationof MRI measures and their link to disease BPFGMF and WMF decrease with disease progressionwhile T2LV increases with disease progressionAccordingly we analyzed the linkage between eachMRI measure and the significant antibody reactivitiesshown in table 2 We identified IgG antibody reac-tivities associated with increased tissue destruction asevidenced by decreased BPF values (figure 1) Strik-ingly we found that antibodies linked to diseasepathology as measured by BPF were enriched for reac-tivity against lipids Furthermore a selective increasein lipid-reactive antibodies linked to disease severitywas also observed when we analyzed all available MRImeasures (figure 2) Of note we did not detect an
Table 2 Continued
Antigens
BPF GMF WMF T2LV
r p FDR r p FDR r p FDR r p FDR
Thyrocalcitonin 2033 0143 0258 2008 0746 0996 2059 0005b 0108 004 0878 1000
TNPAL-galactocerebroside 007 0763 0969 2015 0506 0996 047 0030a 0142 005 0823 1000
Total cerebroside 2016 0498 0928 2041 0064 0112 050 0020a 0122 062 0003a 0033
Trisialogang GT1b 2034 0128 0194 2044 0043b 0066 015 0513 1000 030 0185 0247
Troponin I 2051 0020b 0040b 2053 0013b 0026b 2003 0900 1000 043 0050 0072
Ubiquitin 031 0166 0305 045 0039a 0027 2025 0282 0854 2029 0196 0247
VEGF 2050 0023b 0040b 2047 0031b 0027b 2013 0570 1000 040 0070 0146
Abbreviations BPF 5 whole brain parenchymal fraction FDR 5 false discovery ratendashadjusted significance value GMF 5 global cerebral gray matterfraction T2LV 5 cerebral T2 (fluid-attenuated inversion recovery) hyperintense lesion volume WMF 5 global cerebral white matter fractionDiscovery setap 005 (positive association)bp 005 (negative association)
Neurology Neuroimmunology amp Neuroinflammation 5
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
enrichment in lipid-reactive IgM in the group ofantibodies associated with increased diseasepathogenesis (not shown)
An index of serum IgG reactivity to lipids is associated
with MRI measures of disease severity To further inves-tigate the association between serum IgG reactivityto lipids and MRI measures of disease severity wecalculated an IgG anti-lipid index for each patientcorresponding to the information on each lipid-specific antibody listed in table 2 normalized by thestrength of its correlation with that MRI measureunder investigation The antibody reactivities tolipids used to calculate the lipid index are shown in
figure 3A With this approach we calculated one indexfor each of the MRI measures analyzed in this studyWe found a significant correlation between each IgGlipid antibody index and BPF GMF and T2LV(figure 3B) no significant correlation was found withWMF Moreover no significant correlations werefound between BPF WMF GMF and T2LVmeasures and anti-lipid antibody indexes based onIgM reactivity (not shown)
Finally we evaluated the performance of the lipidantibody indexes linked to BPF GMF and T2LV onan additional set of independent MS samples (valida-tion set table 1) We detected a similar trend to theone detected in the discovery set with regard to the
Figure 1 Antibody reactivity in serum is associated with decreased brain volume
Heatmap in which each column represents the mean immunoglobulin G antibody reactivity in a serum sample from a patient with multiple sclerosis sortedaccording to the whole brain parenchymal fraction (BPF) (indicated at the topmdasha lower BPF indicates more brain atrophy) and each row represents theantibody reactivity to an antigen according to the colorimetric scale shown The antibody reactivities included in this heatmap are listed in table 2
6 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
Figure 2 Association of serum IgG reactivity with MRI measures of disease severity
(A) Heatmap in which each column represents an MRI measure of either atrophy (BPF GMF WMF) or lesions (T2LV) andeach row represents the correlation to IgG serum antibody reactivity according to a colorimetric scale (B) Frequency oflipid-reactive antibodies linked to higher or lower MRI disease severity BPF 5 whole brain parenchymal fraction GMF 5
global cerebral gray matter fraction IgG 5 immunoglobulin G T2LV 5 cerebral T2 (fluid-attenuated inversion recovery)hyperintense lesion volume WMF 5 global cerebral white matter fraction
Neurology Neuroimmunology amp Neuroinflammation 7
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
correlation between BPF and GMF and their respec-tive lipid indexes and we validated the correlationbetween T2LV and its lipid index (figure 3C)
DISCUSSION Previous studies have investigated theassociation of MRI with immune activity in MS13ndash19
These studies have included cellular immune measures
Figure 3 Correlation of lipid indexes to MRI measures of disease severity in multiple sclerosis
(A) Heatmap inwhich each column represents anMRImeasure and each row represents the correlation to immunoglobulin G serumantibody reactivity to lipids accord-ing to a colorimetric scale (B) Scatter plots depicting the correlation between each lipid index andMRImeasures (BPF GMF and T2LV) in the discovery set (C) Scatterplots depicting the correlation between each lipid index andMRImeasures (BPF GMF and T2LV) in the validation set BPF5whole brain parenchymal fraction GMF5
cerebral gray matter fraction (see methods section for details) T2LV 5 cerebral T2 hyperintense lesion volume WMF 5 global cerebral white matter fraction
8 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
as well as oligoclonal bands or neurofilament-specificantibodies in CSF42ndash44 In the present study weanalyzed the association between serum antibodyprofiles detected with antigen microarrays and MRImeasures of disease including cerebral atrophy andT2 hyperintense lesions We found that specificantibody patterns are associated with different aspectsof MRI-defined disease pathology Our major findingwas that increased reactivity to lipids is associated withdifferent aspects of brain MRI measures of diseaseseverity Of note the specific set of lipids associatedwith atrophy differed from those associated withlesions Taken together these data suggest that anti-lipid antibodies in serum are related to MRI measuresof disease Our findings highlight the important role oflipid and lipid-specific immunity in the pathogenesis ofMS
Although a potential limitation of these studies isthe relatively small number of samples analyzed wehave validated our findings in an independent valida-tion cohort strengthening their significance In addi-tion further studies in large cohorts from patientsaffected by non-MS neurodegenerative diseases andhealthy controls may indicate whether the antibodyreactivities and lipid indexes identified in these stud-ies are exclusive to MS or are associated with addi-tional biological processes Indeed some of theseantibody reactivities contributing to the immune sig-natures described in this work have been found innewborns45
Different sets of antibody reactivities were associ-ated with GMF WMF BPF and T2LV The differ-ent antibody link between lesions and atrophy is inkeeping with the long-held view that brain atrophyin patients with MS is multifactorial and is onlyweakly related to lesions41 This has been underscoredin recent studies showing the complementary infor-mation obtained by combining lesion and atrophymeasures4647
Among the cerebral atrophy measures there was astriking overlap between those antibodies associatedwith GMF and BPF which separated them fromWMF results These findings are in keeping with pre-vious observations that whole brain atrophy is domi-nated by GM loss3741 In the early stages of MS thisGM atrophy selectively involves the deep GM nuclei34
Thus the similarities observed in the autoantibodieslinked to GMF and BPF might reflect the dominantcontribution and colinearity of GM atrophy to totalbrain atrophy The set of antibodies linked to GMFmay be the most relevant given that MRI studies haveshown that GM volume is more closely linked to phys-ical disability37 and cognitive impairment38 than areWM volume or lesion measures
Using antigen microarrays antibody reactivity tolipids has been detected in the CSF and serum of
patients with MS at different stages of the diseaseand such antibodies in the CSF have been linkedto disease progression and MRI involvement44
Although several mechanisms are thought to drivebrain inflammation and atrophy in MS46 the connec-tion between these mechanisms and the antibodiesdetected with our antigen microarrays is yet unknownOne possibility is that the serum autoantibodies iden-tified in our studies are not pathogenic and reflect theresult of immunization against self-antigens releasedfrom the CNS during the course of the disease Indeedneurofilament is released from damaged axons duringthe course of MS and both neurofilament lightchains48 and antibodies against it42 have been linkedto MRI measures of disease Moreover heat shockproteins are upregulated in different cell types duringinflammation and have been shown to have an impor-tant role as immunomodulators when released to theextracellular medium4950 Thus it is possible that heatshock proteinndashreactive antibodies reflect changes in theproduction and release of these immunomodulatorsduring the course of disease pathogenesis Similarlybioactive lipids and their products are released fromdamaged myelin and lipid-specific antibodies havebeen detected in patients with MS27293051 Lipids havean important role in the immune response both asbioactive molecules with immunomodulatory proper-ties and also as targets of the adaptive immuneresponse52 Moreover lipids have significant effectson the murine model of MS experimental autoim-mune encephalomyelitis2729305153 Indeed we recentlyfound that the glycolipid lactosylceramide activates abroad set of biological processes in astrocytes promot-ing neurodegeneration and inflammation53 Thus thelipid-reactive antibodies detected in this work mayreflect the release of myelin lipids in the context ofMS pathogenesis andor may directly contribute toimmune-mediated damage in CNS tissue Of notelipid-reactive antibodies are highly cross-reactive52 Inaddition the lipid-reactive antibodies detected in thiswork as associated with MRI measures of disease activ-ity were of the IgG class These are important points toconsider when evaluating a potential role of lipid-reactive antibodies inMS pathogenesis Further studiesare warranted to investigate whether bioactive lipidsoffer new therapeutic targets in MS
We have found that unique patterns of immunereactivity determined with antibody arrays are associ-ated with specific MRI measures of disease severityThese patterns agree with the interpretation that dif-ferent pathogenic mechanisms drive diverse diseaseprocesses reflected by these MRI measures These pat-terns also suggest a predominant role for lipids andlipid-specific immunity in MS pathology Furtherstudies are warranted to determine whether the earlyappearance of anti-lipid antibodies predicts the
Neurology Neuroimmunology amp Neuroinflammation 9
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
subsequent development of clinical and MRI-defineddisease worsening
AUTHOR CONTRIBUTIONSRohit Bakshi draftingrevising the manuscript study concept or design
analysis or interpretation of data acquisition of data study supervision
obtaining funding Ada Yeste study concept or design acquisition of
data Bonny Patel analysis or interpretation of data statistical analysis
Shahamat Tauhid analysis or interpretation of data Subhash Tummala
analysis or interpretation of data Roya Rahbari analysis or interpretation
of data statistical analysis Renxin Chu analysis or interpretation of data
Keren Regev analysis or interpretation of data acquisition of data
Pia Kivisaumlkk draftingrevising the manuscript study concept or design
contribution of vital reagentstoolspatients acquisition of data study
supervision Howard L Weiner study concept or design obtaining fund-
ing Francisco J Quintana draftingrevising the manuscript study con-
cept or design contribution of vital reagentstoolspatients study
supervision obtaining funding
ACKNOWLEDGMENTThe authors thank Dr Mohit Neema Dr Antonella Ceccarelli and
Dr Ashish Arora for valuable assistance at the early stages of this project
STUDY FUNDINGThis work was supported in part by grants to FJQ and HLW from
EMD Serono RG4111A1 and JF2161-A-5 from the National Multiple
Sclerosis Society PA0069 from the International Progressive MS Alli-
ance and by grants from the NIH (5R01NS055083-04) and NMSS
(RG3798A2) to RB
DISCLOSUREFJ Quintana serves on the editorial board for Systems Biomedicine
Inmunologia American Journal of Clinical and Experimental Immunology
is an associate editor for Immunology (UK) is an advisory board member
for Seminars in Immunopathology received research support from Harvard
Medical School BADERC NMSS A Yeste and B Patel report no
disclosures S Tauhid is managing editor for Journal of Neuroimaging
S Tummala R Rahbari R Chu and K Regev report no disclosures
P Kivisaumlkk received research support from EMD Serono R Bakshi is
editor-in-chief for Journal of Neuroimaging received consulting fees from
AbbVie Alkermes Biogen Novartis Questcor received research support
from Biogen EMD-Serono Novartis Sanofi-Genzyme Teva his spouse
holds stock in Biogen Inc H Weiner served on the advisory board
for The Guthy-Jackson Charitable Foundation Teva Pharmaceuticals
Industries Ltd Biogen Idec Novartis Sanofi-Aventis has consulted for
Therapix Bioven Novartis Serono Teva Sanofi received research support
from National Multiple Sclerosis Society Go to Neurologyorgnn for full
disclosure forms
Received July 27 2015 Accepted in final form December 8 2015
REFERENCES1 Weiner HL The challenge of multiple sclerosis how do
we cure a chronic heterogeneous disease Ann Neurol
200965239ndash248
2 Bakshi R Thompson AJ Rocca MA et al MRI in mul-
tiple sclerosis current status and future prospects Lancet
Neurol 20087615ndash625
3 Ceccarelli A Bakshi R Neema M MRI in multiple scle-
rosis a review of the current literature Curr Opin Neurol
201225402ndash409
4 Filippi M Rocca MA Barkhof F et al Association
between pathological and MRI findings in multiple scle-
rosis Lancet Neurol 201211349ndash360
5 Geurts JJ Calabrese M Fisher E Rudick RA Measure-
ment and clinical effect of grey matter pathology in mul-
tiple sclerosis Lancet Neurol 2012111082ndash1092
6 Klaver R De Vries HE Schenk GJ Geurts JJ Grey mat-
ter damage in multiple sclerosis a pathology perspective
Prion 2013766ndash75
7 Khoury S Bakshi R Cerebral pseudoatrophy or real atro-
phy after therapy in multiple sclerosis Ann Neurol 2010
68778ndash779
8 Hauser SL Chan JR Oksenberg JR Multiple sclerosis
prospects and promise Ann Neurol 201374317ndash327
9 McFarland HF Martin R Multiple sclerosis a complicated
picture of autoimmunity Nat Immunol 20078913ndash919
10 Nylander A Hafler DA Multiple sclerosis J Clin Invest
20121221180ndash1188
11 Sospedra M Martin R Immunology of multiple sclerosis
Ann Rev Immunol 200523683ndash747
12 Steinman L Immunology of relapse and remission in mul-
tiple sclerosis Ann Rev Immunol 201432257ndash281
13 Khoury SJ Guttmann CR Orav EJ Kikinis R Jolesz FA
Weiner HL Changes in activated T cells in the blood
correlate with disease activity in multiple sclerosis Arch
Neurol 2000571183ndash1189
14 Khoury SJ Orav EJ Guttmann CR Kikinis R Jolesz FA
Weiner HL Changes in serum levels of ICAM and TNF-
R correlate with disease activity in multiple sclerosis Neu-
rology 199953758ndash764
15 Laplaud DA Berthelot L Miqueu P et al Serial blood T
cell repertoire alterations in multiple sclerosis patients cor-
relation with clinical and MRI parameters J Neuroimmunol
2006177151ndash160
16 Makhlouf K Weiner HL Khoury SJ Increased percentage
of IL-121 monocytes in the blood correlates with the
presence of active MRI lesions in MS J Neuroimmunol
2001119145ndash149
17 Prat A Biernacki K Saroli T et al Kinin B1 receptor expres-
sion on multiple sclerosis mononuclear cells correlation with
magnetic resonance imaging T2-weighted lesion volume and
clinical disability Arch Neurol 200562795ndash800
18 Rinaldi L Gallo P Calabrese M et al Longitudinal anal-
ysis of immune cell phenotypes in early stage multiple
sclerosis distinctive patterns characterize MRI-active pa-
tients Brain 20061291993ndash2007
19 Tortorella P Lagana MM Saresella M et al Determinants
of disability in multiple sclerosis an immunological and
MRI study Biomed Res Int 20142014875768
20 Robinson WH DiGennaro C Hueber W et al Autoan-
tigen microarrays for multiplex characterization of autoan-
tibody responses Nat Med 20028295ndash301
21 Yeste A Quintana FJ Antigen microarrays for the study of
autoimmune diseases Clin Chem 2013591036ndash1044
22 Li QZ Xie C Wu T et al Identification of autoantibody
clusters that best predict lupus disease activity using glomer-
ular proteome arrays J Clin Invest 20051153428ndash3439
23 Hueber W Kidd BA Tomooka BH et al Antigen micro-
array profiling of autoantibodies in rheumatoid arthritis
Arthritis Rheum 2005522645ndash2655
24 Goldschmidt Y Sharon E Quintana FJ Cohen IR
Brandt A Adaptive methods for classification of biological
microarray data from multiple experiments Report no
MCS03-07 Rehovot Israel The Arthur and Rochelle Bel-
fer Institute of Mathematics and Computer Science 2003
25 Quintana FJ Getz G Hed G Domany E Cohen IR
Cluster analysis of human autoantibody reactivities in
health and in type 1 diabetes mellitus a bio-informatic
approach to immune complexity J Autoimmun 2003
2165ndash75
10 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
26 Lalive PH Menge T Barman I Cree BA Genain CP
Identification of new serum autoantibodies in neuromye-
litis optica using protein microarrays Neurology 200667
176ndash177
27 Kanter JL Narayana S Ho PP et al Lipid microarrays
identify key mediators of autoimmune brain inflamma-
tion Nat Med 200612138ndash143
28 Quintana FJ Farez MF Izquierdo G Lucas M Cohen IR
Weiner HL Antigen microarrays identify CNS-produced
autoantibodies in RRMS Neurology 201278532ndash539
29 Farez MF Quintana FJ Gandhi R Izquierdo G Lucas M
Weiner HL Toll-like receptor 2 and poly(ADP-ribose)
polymerase 1 promote central nervous system neuroin-
flammation in progressive EAE Nat Immunol 200910
958ndash964
30 Quintana FJ Farez MF Viglietta V et al Antigen micro-
arrays identify unique serum autoantibody signatures in
clinical and pathologic subtypes of multiple sclerosis Proc
Natl Acad Sci USA 200810518889ndash18894
31 Quintana FJ Patel B Yeste A et al Epitope spreading as
an early pathogenic event in pediatric multiple sclerosis
Neurology 2014832219ndash2226
32 Gauthier SA Glanz BI Mandel M Weiner HL A model
for the comprehensive investigation of a chronic autoim-
mune disease the multiple sclerosis CLIMB study Auto-
immun Rev 20065532ndash536
33 Polman CH Reingold SC Edan G et al Diagnostic criteria
for multiple sclerosis 2005 revisions to the ldquoMcDonald
Criteriardquo Ann Neurol 200558840ndash846
34 DellrsquoOglio E Ceccarelli A Glanz BI et al Quantification
of global cerebral atrophy in multiple sclerosis from 3T
MRI using SPM the role of misclassification errors
J Neuroimaging 201525191ndash199
35 Ceccarelli A Jackson JS Tauhid S et al The impact of
lesion in-painting and registration methods on voxel-based
morphometry in detecting regional cerebral gray matter
atrophy in multiple sclerosis AJNR Am J Neuroradiol
2012331579ndash1585
36 Cohen AB Neema M Arora A et al The relationships
among MRI-defined spinal cord involvement brain involve-
ment and disability in multiple sclerosis J Neuroimaging
201222122ndash128
37 Sanfilipo MP Benedict RH Sharma J Weinstock-
Guttman B Bakshi R The relationship between whole
brain volume and disability in multiple sclerosis a com-
parison of normalized gray vs white matter with misclas-
sification correction Neuroimage 2005261068ndash1077
38 Sanfilipo MP Benedict RH Weinstock-Guttman B
Bakshi R Gray and white matter brain atrophy and neuro-
psychological impairment in multiple sclerosis Neurology
200666685ndash692
39 Stankiewicz JM Glanz BI Healy BC et al Brain MRI
lesion load at 15T and 3T versus clinical status in multi-
ple sclerosis J Neuroimaging 201121e50ndashe56
40 Quintana FJ Hagedorn PH Elizur G Merbl Y
Domany E Cohen IR Functional immunomics micro-
array analysis of IgG autoantibody repertoires predicts the
future response of mice to induced diabetes Proc Natl
Acad Sci USA 2004101(suppl 2)14615ndash14621
41 Bermel RA Bakshi R The measurement and clinical
relevance of brain atrophy in multiple sclerosis Lancet
Neurol 20065158ndash170
42 Eikelenboom MJ Petzold A Lazeron RH et al Multiple
sclerosis neurofilament light chain antibodies are corre-
lated to cerebral atrophy Neurology 200360219ndash223
43 Magraner MJ Bosca I Simo-Castello M et al Brain atro-
phy and lesion load are related to CSF lipid-specific IgM
oligoclonal bands in clinically isolated syndromes Neuro-
radiol 2012545ndash12
44 Villar LM Sadaba MC Roldan E et al Intrathecal syn-
thesis of oligoclonal IgM against myelin lipids predicts an
aggressive disease course in MS J Clin Invest 2005115
187ndash194
45 Merbl Y Zucker-Toledano M Quintana FJ Cohen IR
Newborn humans manifest autoantibodies to defined self
molecules detected by antigen microarray informatics
J Clin Invest 2007117712ndash718
46 Sormani MP Arnold DL De Stefano N Treatment effect
on brain atrophy correlates with treatment effect on disa-
bility in multiple sclerosis Ann Neurol 20147543ndash49
47 Tauhid S Neema M Healy BC Weiner HL Bakshi R
MRI phenotypes based on cerebral lesions and atrophy in
patients with multiple sclerosis J Neurol Sci 2014346
250ndash254
48 Gnanapavan S Grant D Morant S et al Biomarker
report from the phase II lamotrigine trial in secondary
progressive MS neurofilament as a surrogate of disease
progression PLoS One 20138e70019
49 Quintana FJ Cohen IR Heat shock proteins as endoge-
nous adjuvants in sterile and septic inflammation
J Immunol 20051752777ndash2782
50 Quintana FJ Cohen IR The HSP60 immune system net-
work Trends Immunol 20113289ndash95
51 Ho PP Kanter JL Johnson AM et al Identification of
naturally occurring fatty acids of the myelin sheath that
resolve neuroinflammation Sci Transl Med 20124137ra73
52 Quintana FJ Yeste A Weiner HL Covacu R Lipids and
lipid-reactive antibodies as biomarkers for multiple sclero-
sis J Neuroimmunol 201224853ndash57
53 Mayo L Trauger SA Blain M et al Regulation of astro-
cyte activation by glycolipids drives chronic CNS inflam-
mation Nat Med 2014201147ndash1156
Neurology Neuroimmunology amp Neuroinflammation 11
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
1
Table e-1 Antigens used
Group Antigen Source
27 kDa Heat Shock Protein Stressgen
HSP 32 kDa Heat Shock Protein Stressgen
40 kDa Heat Shock Protein Stressgen
47 kDa Heat Shock Protein Stressgen
60 kDa Heat Shock Protein Stressgen
60 kDa Heat Shock Protein peptide aa 106ndash125 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 121ndash140 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 136ndash155 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 151ndash170 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 16ndash35 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 166ndash185 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
2
60 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 556ndash573 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
65 kDa Heat Shock Protein M tuberculosis Stressgen
70 kDa Heat Shock Protein Stressgen
70 kDa Heat Shock Protein peptide aa 106ndash125 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 121ndash140 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 136ndash155 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 151ndash170 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 16ndash35 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 166ndash185 Biopolymers Facility Harvard Medical School
3
70 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
4
70 kDa Heat Shock Protein peptide aa 556ndash575 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 571ndash590 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 586ndash605 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 601ndash620 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 616ndash635 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 631ndash640 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
71 kDa Heat Shock Protein M tuberculosis Biopolymers Facility Harvard Medical School
90 kDa Heat Shock Protein Stressgen
GroEL Stressgen
23-cyclic nucleotide 3-phosphodiesterase
peptide aa106-125 Biopolymers Facility Harvard Medical School
CNS
23-cyclic nucleotide 3-phosphodiesterase
peptide aa1-20 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa121-140 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa136-155 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa151-170 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa16-35 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa166-185 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase Biopolymers Facility Harvard Medical School
5
peptide aa181-200
23-cyclic nucleotide 3-phosphodiesterase
peptide aa196-215 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa211-230 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa226-245 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa241-260 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa256-275 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa271-290 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa286-305 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa301-320 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa31-50 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa316-335 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa331-350 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa346-365 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa361-380 Biopolymers Facility Harvard Medical School
6
23-cyclic nucleotide 3-phosphodiesterase
peptide aa376-395 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa391-410 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa406-421 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa46-65 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa61-80 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa76-95 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa91-110 Biopolymers Facility Harvard Medical School
Acetyl Cholinesterase Sigma Aldrich
ADAM-10 Sigma Aldrich
alpha-Cristallin Stressgen
beta-Cristallin Sigma Aldrich
bovine Myelin Basic Protein Sigma Aldrich
Brain Extract I Sigma Aldrich
Brain Extract II Sigma Aldrich
Brain Extract III Sigma Aldrich
Glial Filament Acidic Protein Research Diagnostic
guinea pig Myelin Basic Protein Sigma Aldrich
human Myelin Basic Protein Sigma Aldrich
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 106-125 Biopolymers Facility Harvard Medical School
7
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 121-140 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 136-155 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 151-170 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 166-185 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 181-200 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 91-110 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
106-125 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa 1- Biopolymers Facility Harvard Medical School
8
20
Myelinoligodendrocyte glycoprotein peptide aa
121-140 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
136-155 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
151-170 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
16-35 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
166-185 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
181-200 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
196-215 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
211-230 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
226-247 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
31-50 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
35-55 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
46-65 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
61-80 Biopolymers Facility Harvard Medical School
9
Myelinoligodendrocyte glycoprotein peptide aa
76-95 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
91-110 Biopolymers Facility Harvard Medical School
murine Myelin Basic Protein Sigma Aldrich
Myelin Associated Glycoprotein Sigma Aldrich
Myelin Basic Protein peptide aa 104-123 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 11-30 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 113-132 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 121-138 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 124-142 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 138-147 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 141-161 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 143-168 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 155-178 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 26-35 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 41-60 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 51-70 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 71-92 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 84-94 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 89-101 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 173-186 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 93-112 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 106-125 Biopolymers Facility Harvard Medical School
10
Myelin Protein 2 peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 121-132 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 91-110 Biopolymers Facility Harvard Medical School
Neurofilament 160kd Chemicon
Neurofilament 200kd Chemicon
Neurofilament 68kd Chemicon
Neuronal Enolase Calbiochem
Nicastrin GeneTex
NMDA receptor Novus Biologicals
NOGO Sigma Aldrich
Olygodendrocyte-Specific Protein peptide aa 106ndash
125 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 121ndash
140 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 136ndash
155 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 151ndash
170 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 16ndash
35 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 166ndash Biopolymers Facility Harvard Medical School
11
185
Olygodendrocyte-Specific Protein peptide aa 181ndash
199 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 195ndash
217 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 31ndash
50 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 46ndash
65 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 61ndash
80 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 76ndash
95 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 91ndash
110 Biopolymers Facility Harvard Medical School
Proteolipid Protein Abnova
Proteolipid Protein peptide aa 100-119 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 10-29 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 110-129 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 1-19 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 125-141 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-150 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-154 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 150-163 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 151-173 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 158-166 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 161-180 Biopolymers Facility Harvard Medical School
12
Proteolipid Protein peptide aa 178-191 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 180-199 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 190-209 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 20-39 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 205-220 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 215-232 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-239 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-249 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 248-259 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 250-269 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 265-277 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 35-50 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 40-59 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 50-69 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 65-84 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 80-99 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 91-110 Biopolymers Facility Harvard Medical School
Retinol Binding Protein Sigma Aldrich
S100beta protein Assay Designs
Super Oxide Dismutase Sigma Aldrich
Synuclein beta Sigma Aldrich
Synuclein gamma Sigma Aldrich
Amydgala ProSci Inc
Tissue Amydgala AD ProSci Inc
Brain lysate ProSci Inc
Brain Tissue Membrane ProSci Inc
Cerebellar pedunculus ProSci Inc
13
Cerebral meninges ProSci Inc
Corpus Callosum ProSci Inc
Corpus Callosum AD ProSci Inc
Diencephalon ProSci Inc
Fetal brain ProSci Inc
Frontal lobe ProSci Inc
Frontal lobe AD ProSci Inc
Hippocampus ProSci Inc
Hippocampus AD ProSci Inc
Insula ProSci Inc
Occipital lobe ProSci Inc
Occipital lobe AD ProSci Inc
Olfactory region ProSci Inc
Optic Nerve ProSci Inc
Parietal lobe ProSci Inc
Parietal lobe AD ProSci Inc
Pons ProSci Inc
Pons AD ProSci Inc
Postcentral gyrus ProSci Inc
Postcentral gyrus AD ProSci Inc
Precentral gyrus ProSci Inc
Precentral gyrus AD ProSci Inc
Spinal cord ProSci Inc
Temporal lobe ProSci Inc
Temporal lobe AD ProSci Inc
Thalamus ProSci Inc
Thalamus AD ProSci Inc
14
Amyloid beta Sigma Aldrich
AD
related Amyloid beta 10-20 Sigma Aldrich
Amyloid beta 1-12 Sigma Aldrich
Amyloid beta 12-28 Sigma Aldrich
Amyloid beta 1-23 Sigma Aldrich
Amyloid beta 1-38 Sigma Aldrich
Amyloid beta 17-40 Sigma Aldrich
Amyloid beta 25-35 Sigma Aldrich
Amyloid beta 34-42 Sigma Aldrich
Amyloid bri protein precursor 227 Sigma Aldrich
Amyloid DAN Protein Fragment 1-34 Sigma Aldrich
Amyloid Precursor Protein Sigma Aldrich
Amyloid protein no AB component Sigma Aldrich
Secreted amyloid precursor protein (SAP) beta Sigma Aldrich
Tau isoform variant 0N3R Sigma Aldrich
Tau isoform variant 1N3R Sigma Aldrich
Tau isoform variant 0N4R Sigma Aldrich
Tau isoform variant 2N3R Sigma Aldrich
Tau phospho Ser412 Sigma Aldrich
Tau phospho Ser441 Sigma Aldrich
Tau phospho Thr181 Sigma Aldrich
Tau Protein human EMD Biosciences
(plusmn)9-HODE Cayman Chemical
Lipids
1 Palmitoyl-2-(5oxo-Valeroyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
15a-hydroxycholestene Avanti Polar Lipids
15
15-ketocholestane Avanti Polar Lipids
15-ketocholestene Avanti Polar Lipids
1-Palmitoil-2-(9oxo-Nonanoyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Azelaoyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Glutaroyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
5 α-cholestane-3 β15 α-diol Avanti Polar Lipids
9(S)-HODE Cayman Chemical
Asialoganglioside-GM1 Sigma Aldrich
Asialoganglioside-GM2 Sigma Aldrich
Brain ceramides Avanti Polar Lipids
Brain D-erythrosphingosine Avanti Polar Lipids
Brain lysophosphatidylethanolamine Avanti Polar Lipids
Brain L-α-lysophosphatidylserine Avanti Polar Lipids
Brain L-α-phosphatidylcholine Avanti Polar Lipids
Brain L-α-phosphatidyl-ethanolamine Avanti Polar Lipids
Brain L-α-phosphatidylserine Avanti Polar Lipids
Brain polar lipid extract Avanti Polar Lipids
Brain sphingomyelin Avanti Polar Lipids
Brain sulfatide Avanti Polar Lipids
Brain total lipid extract Avanti Polar Lipids
Cardiolipin Sigma Aldrich
Ceramide Sigma Aldrich
Ceramide 1-phosphate Sigma Aldrich
Cholesterol Sigma Aldrich
16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-
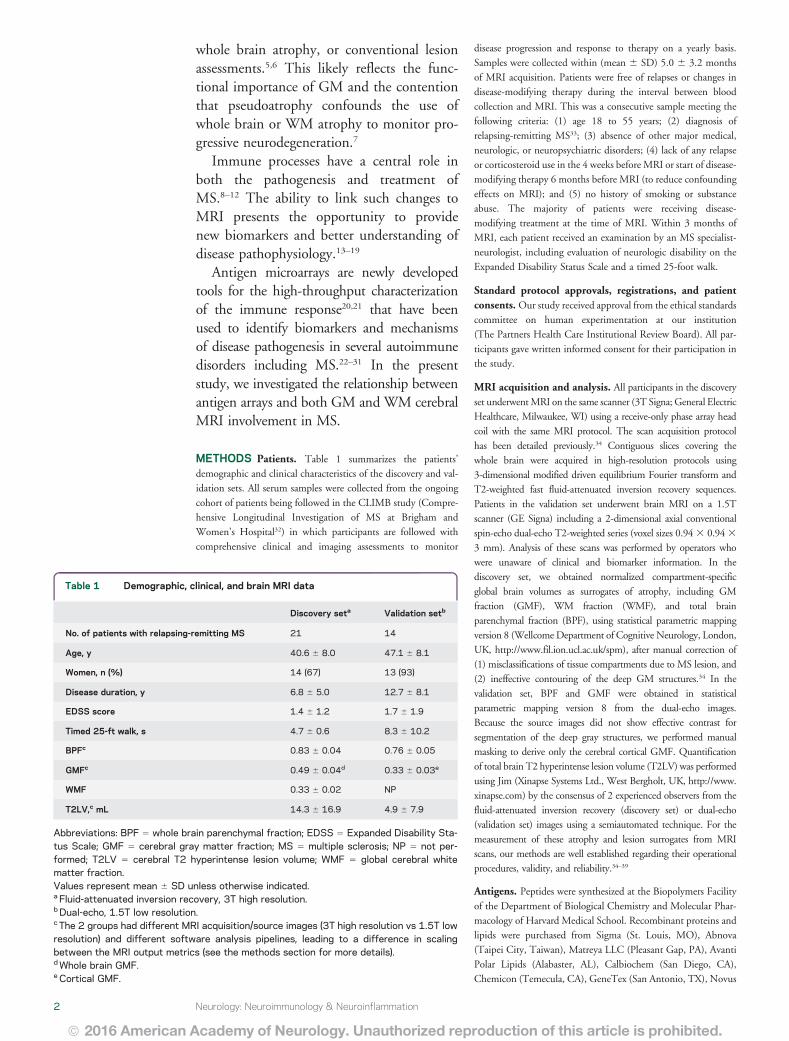
whole brain atrophy or conventional lesionassessments56 This likely reflects the func-tional importance of GM and the contentionthat pseudoatrophy confounds the use ofwhole brain or WM atrophy to monitor pro-gressive neurodegeneration7
Immune processes have a central role inboth the pathogenesis and treatment ofMS8ndash12 The ability to link such changes toMRI presents the opportunity to providenew biomarkers and better understanding ofdisease pathophysiology13ndash19
Antigen microarrays are newly developedtools for the high-throughput characterizationof the immune response2021 that have beenused to identify biomarkers and mechanismsof disease pathogenesis in several autoimmunedisorders including MS22ndash31 In the presentstudy we investigated the relationship betweenantigen arrays and both GM and WM cerebralMRI involvement in MS
METHODS Patients Table 1 summarizes the patientsrsquo
demographic and clinical characteristics of the discovery and val-
idation sets All serum samples were collected from the ongoing
cohort of patients being followed in the CLIMB study (Compre-
hensive Longitudinal Investigation of MS at Brigham and
Womenrsquos Hospital32) in which participants are followed with
comprehensive clinical and imaging assessments to monitor
disease progression and response to therapy on a yearly basis
Samples were collected within (mean 6 SD) 50 6 32 months
of MRI acquisition Patients were free of relapses or changes in
disease-modifying therapy during the interval between blood
collection and MRI This was a consecutive sample meeting the
following criteria (1) age 18 to 55 years (2) diagnosis of
relapsing-remitting MS33 (3) absence of other major medical
neurologic or neuropsychiatric disorders (4) lack of any relapse
or corticosteroid use in the 4 weeks before MRI or start of disease-
modifying therapy 6 months before MRI (to reduce confounding
effects on MRI) and (5) no history of smoking or substance
abuse The majority of patients were receiving disease-
modifying treatment at the time of MRI Within 3 months of
MRI each patient received an examination by an MS specialist-
neurologist including evaluation of neurologic disability on the
Expanded Disability Status Scale and a timed 25-foot walk
Standard protocol approvals registrations and patientconsents Our study received approval from the ethical standards
committee on human experimentation at our institution
(The Partners Health Care Institutional Review Board) All par-
ticipants gave written informed consent for their participation in
the study
MRI acquisition and analysis All participants in the discoveryset underwentMRI on the same scanner (3T Signa General Electric
Healthcare Milwaukee WI) using a receive-only phase array head
coil with the same MRI protocol The scan acquisition protocol
has been detailed previously34 Contiguous slices covering the
whole brain were acquired in high-resolution protocols using
3-dimensional modified driven equilibrium Fourier transform and
T2-weighted fast fluid-attenuated inversion recovery sequences
Patients in the validation set underwent brain MRI on a 15T
scanner (GE Signa) including a 2-dimensional axial conventional
spin-echo dual-echo T2-weighted series (voxel sizes 0943 0943
3 mm) Analysis of these scans was performed by operators who
were unaware of clinical and biomarker information In the
discovery set we obtained normalized compartment-specific
global brain volumes as surrogates of atrophy including GM
fraction (GMF) WM fraction (WMF) and total brain
parenchymal fraction (BPF) using statistical parametric mapping
version 8 (WellcomeDepartment of Cognitive Neurology London
UK httpwwwfilionuclacukspm) after manual correction of
(1) misclassifications of tissue compartments due to MS lesion and
(2) ineffective contouring of the deep GM structures34 In the
validation set BPF and GMF were obtained in statistical
parametric mapping version 8 from the dual-echo images
Because the source images did not show effective contrast for
segmentation of the deep gray structures we performed manual
masking to derive only the cerebral cortical GMF Quantification
of total brain T2 hyperintense lesion volume (T2LV) was performed
using Jim (Xinapse Systems Ltd West Bergholt UK httpwww
xinapsecom) by the consensus of 2 experienced observers from the
fluid-attenuated inversion recovery (discovery set) or dual-echo
(validation set) images using a semiautomated technique For the
measurement of these atrophy and lesion surrogates from MRI
scans our methods are well established regarding their operational
procedures validity and reliability34ndash39
Antigens Peptides were synthesized at the Biopolymers Facility
of the Department of Biological Chemistry and Molecular Phar-
macology of Harvard Medical School Recombinant proteins and
lipids were purchased from Sigma (St Louis MO) Abnova
(Taipei City Taiwan) Matreya LLC (Pleasant Gap PA) Avanti
Polar Lipids (Alabaster AL) Calbiochem (San Diego CA)
Chemicon (Temecula CA) GeneTex (San Antonio TX) Novus
Table 1 Demographic clinical and brain MRI data
Discovery seta Validation setb
No of patients with relapsing-remitting MS 21 14
Age y 406 6 80 471 6 81
Women n () 14 (67) 13 (93)
Disease duration y 68 6 50 127 6 81
EDSS score 14 6 12 17 6 19
Timed 25-ft walk s 47 6 06 83 6 102
BPFc 083 6 004 076 6 005
GMFc 049 6 004d 033 6 003e
WMF 033 6 002 NP
T2LVc mL 143 6 169 49 6 79
Abbreviations BPF 5 whole brain parenchymal fraction EDSS 5 Expanded Disability Sta-tus Scale GMF 5 cerebral gray matter fraction MS 5 multiple sclerosis NP 5 not per-formed T2LV 5 cerebral T2 hyperintense lesion volume WMF 5 global cerebral whitematter fractionValues represent mean 6 SD unless otherwise indicateda Fluid-attenuated inversion recovery 3T high resolutionbDual-echo 15T low resolutionc The 2 groups had different MRI acquisitionsource images (3T high resolution vs 15T lowresolution) and different software analysis pipelines leading to a difference in scalingbetween the MRI output metrics (see the methods section for more details)dWhole brain GMFeCortical GMF
2 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
Table 2 Serum immunoglobulin Gs associated with MRI measures of disease severity
Antigens
BPF GMF WMF T2LV
r p FDR r p FDR r p FDR r p FDR
1-P-2-5-o-V-sn-G-3-PC 2038 0087 0161 2034 0131 0197 2014 0540 1000 056 0008a 0056
15-ketocholestene 2030 0190 0370 2044 0047b 0066 024 0302 0854 030 0189 0247
9 HODE 2040 0073 0097 2051 0019b 0026b 016 0499 1000 045 0043a 0057
9 S-HODE 2042 0057 0067 2048 0027b 0026b 006 0797 1000 061 0003a 0033a
a-MSH 050 0021a 0040a 040 0070 0112 028 0214 0854 2004 0858 1000
ABPF 1-12 2023 0320 0610 2040 0070 0112 032 0155 0804 046 0035a 0057
ABPP 227 2030 0179 0305 2056 0009b 0025b 047 0033a 0156 049 0024a 0056
Ac phosphatase 035 0119 0194 047 0033a 0027a 2018 0443 1000 2020 0374 0516
ADPF 1-34 2020 0393 0823 2038 0089 0133 034 0126 0804 046 0035a 0057
ANP 013 0587 0969 037 0100 0197 2048 0027b 0142 2027 0242 0516
Asialoganglioside-GM1 2052 0016b 0040b 2060 0004b 0015b 007 0749 1000 032 0152 0209
Asialoganglioside-GM2 2040 0074 0097 2048 0027b 0026b 011 0648 1000 024 0292 0516
b-MSH 2047 0033b 0058 2038 0085 0133 2025 0277 0854 020 0376 0516
b-Amyloid 2021 0350 0610 2036 0107 0197 026 0247 0854 049 0025a 0056
b-Synuclein 001 0954 0969 2022 0343 0996 048 0029a 0142 042 0055 0094
bNGF 029 0204 0413 006 0783 0996 051 0018a 0122 2006 0807 1000
Brain D-erythrosphingosine 2047 0030b 0058 2056 0009b 0025b 010 0679 1000 037 0096 0146
Brain extract VII 2052 0016b 0040b 2059 0005b 0015b 006 0789 1000 053 0013a 0056
Brain L-a-phosphatidyl-ethanolamine 2044 0044b 0067 2046 0037b 0027b 2005 0837 1000 019 0407 0537
Brain polar lipid extract 013 0573 0969 2010 0657 0996 051 0019a 0122 018 0434 0537
Chorionic G 005 0826 0969 028 0223 0303 2046 0034b 0156 2010 0656 1000
CNPase aa 106ndash125 039 0080 0161 049 0026a 0026a 2014 0557 1000 2051 0017b 0056
CNPase aa 16ndash35 042 0056 0067 046 0037a 0027a 2002 0941 1000 2037 0103 0157
CNPase aa 195ndash214 031 0173 0305 050 0020a 0026a 2035 0119 0804 2035 0118 0157
CNPase aa 286ndash305 028 0227 0413 045 0039a 0027a 2033 0144 0804 2004 0873 1000
CNPase aa 376ndash395 019 0416 0823 043 0049a 0066 2049 0024b 0142 2038 0089 0146
Collagen II 025 0278 0610 046 0034a 0027a 2040 0072 0788 2032 0151 0209
Collagen IX 2067 0001b 0025b 2075 0001b 0002b 008 0741 1000 066 0001a 0033a
Collagen VIII 2027 0242 0521 2026 0251 0605 2005 0831 1000 044 0044a 0057
Collagen X 2036 0114 0194 2012 0595 0996 2052 0016b 0122 2006 0792 1000
Disialoganglioside-GD2 2040 0073 0097 2049 0023b 0026b 013 0589 1000 043 0055 0094
Disialogang-GD1B 2038 0088 0161 2050 0022b 0026b 018 0441 1000 031 0174 0247
Disialoganglioside GD1a 2051 0017b 0040b 2053 0013b 0026b 2005 0824 1000 042 0060 0146
Disialoganglioside GD3 2040 0069 0097 2054 0012b 0026b 020 0382 1000 051 0018a 0056
Galactocerebrosides 001 0952 0969 2027 0229 0303 059 0005a 0108 025 0265 0516
Ganglioside mixture 2053 0014b 0040b 2062 0003b 0015b 011 0644 1000 053 0014a 0056
Ganglioside-GM4 002 0929 0969 029 0203 0303 2056 0009b 0109 2018 0423 0537
Gelsolin 053 0013a 0040a 057 0008a 0025a 002 0934 1000 2048 0029b 0056
GNRH 2053 0014b 0040b 2043 0054 0072 2029 0196 0854 036 0113 0157
gpMBP 017 0449 0928 2004 0857 1000 048 0029a 0142 000 0988 1000
GT1a 2043 0053 0067 2048 0028b 0026b 003 0912 1000 032 0161 0247
Hemoglobin 015 0527 0969 2006 0784 0996 046 0037a 0203 028 0212 0516
HSP60 aa 195ndash214 023 0311 0610 004 0879 1000 044 0045a 0206 000 0989 1000
HSP60 aa 210ndash229 053 0015a 0040a 049 0024a 0026a 016 0493 1000 2013 0577 1000
ContinuedNeurology Neuroimmunology amp Neuroinflammation 3
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
Table 2 Continued
Antigens
BPF GMF WMF T2LV
r p FDR r p FDR r p FDR r p FDR
HSP60 aa 240ndash259 048 0028a 0058 052 0017a 0026a 000 0990 1000 2047 0030b 0056
HSP60 aa 421ndash440 010 0655 0969 039 0080 0133 2058 0006b 0108 2022 0334 0516
HSP60 aa 436ndash455 042 0061 0067 060 0004a 0015a 2031 0173 0804 2038 0094 0146
HSP70 aa 121ndash140 044 0046a 0067 036 0111 0197 024 0289 0854 2016 0496 0728
HSP70 aa 181ndash199 051 0017a 0040a 048 0027a 0026a 015 0525 1000 2020 0378 0516
HSP70 aa 210ndash229 006 0788 0969 027 0228 0303 2044 0045b 0206 2011 0640 1000
HSP70 aa 316ndash335 2028 0216 0413 2029 0209 0303 2004 0879 1000 044 0048a 0072
HSP70 aa 376ndash395 2042 0059 0067 2046 0035b 0027b 002 0942 1000 035 0121 0157
HSP70 aa 421ndash440 048 0030a 0058 065 0002a 0015a 2027 0229 0854 2034 0137 0209
HSP70 aa 61ndash80 2002 0941 0969 2029 0198 0303 056 0008a 0109 018 0441 0537
IL-12 2050 0021b 0040b 2045 0040b 0027b 2018 0429 1000 039 0083 0146
IL-3 2041 0063 0067 2051 0019b 0026b 013 0575 1000 052 0017a 0056
Insula 046 0038a 0067 033 0143 0303 034 0133 0804 2017 0473 0550
Intrinsic factor 036 0108 0194 044 0048a 0066 2009 0702 1000 000 0986 1000
Isoprostane F2a-I 2053 0014b 0040b 2055 0010b 0025b 2004 0854 1000 035 0115 0157
Lactosylceramide 2021 0368 0630 2041 0062 0112 039 0077 0788 060 0004a 0033a
LIF 2065 0002b 0025b 2059 0005b 0015b 2022 0336 0854 041 0064 0146
MMP2 042 0055 0067 061 0003a 0015a 2032 0153 0804 2039 0082 0146
MMP9 2055 0009b 0040b 2062 0003b 0015b 006 0804 1000 045 0038a 0057
MOG peptide aa 196ndash215 2005 0837 0969 2008 0736 0996 005 0836 1000 044 0045a 0057
Monosialogang GM2 2056 0008b 0040b 2048 0028b 0026b 2026 0260 0854 017 0460 0550
Myelin protein 2 peptide aa 31ndash50 035 0124 0194 048 0029a 0027a 2022 0337 0854 2033 0141 0209
Myelin protein 2 peptide aa 91ndash110 2009 0694 0969 014 0533 0996 2051 0019b 0122 2004 0878 1000
Neurofilament 160 kDa 032 0162 0305 039 0082 0133 2009 0688 1000 2049 0026b 0056
Neurofilament 200 kDa 043 0050 0067 045 0040a 0027a 003 0881 1000 2015 0516 0728
Neurofilament 68 kDa 000 0999 1000 2021 0359 0996 044 0044a 0206 045 0043a 0057
NOGO 016 0493 0928 2012 0617 0996 060 0004a 0108 021 0352 0516
Nonhydroxy fatty acid ceramide 2053 0014b 0040b 2059 0005b 0015b 005 0826 1000 044 0044a 0057
Occipital lobe 2045 0039b 0067 2043 0054 0072 2012 0599 1000 039 0083 0146
ORF26 protein 053 0013a 0040a 052 0015a 0026a 010 0653 1000 2030 0182 0247
Parietal lobe 049 0023a 0040a 058 0005a 0015a 2011 0641 1000 2025 0283 0516
PDGF-Ra 2007 0752 0969 017 0461 0996 2052 0016b 0122 2009 0687 1000
Pepstatin 2035 0116 0194 2046 0036b 0027b 016 0483 1000 025 0277 0516
Postcentral gyrus AD 044 0044a 0067 034 0132 0197 028 0214 0854 2012 0603 1000
Proteolipid protein peptide aa 125ndash141 2009 0685 0969 010 0678 0996 2041 0064 0788 2048 0029b 0056
Proteolipid protein peptide aa 150ndash163 044 0043a 0067 051 0019a 0026a 2006 0809 1000 2050 0020b 0056
Proteolipid protein peptide aa 161ndash180 022 0343 0610 031 0166 0303 2017 0475 1000 2044 0045b 0057
Proteolipid protein peptide aa 250ndash269 037 0103 0194 050 0020a 0026 2023 0321 0854 2037 0098 0146
RBP 025 0274 0610 029 0201 0303 2005 0844 1000 2057 0007b 0056
Ribonuclease 2002 0937 0969 2017 0472 0996 031 0175 0804 054 0011a 0056
S100b 033 0141 0258 011 0649 0996 052 0015a 0122 2022 0344 0516
Spectrin 048 0026a 0058 035 0122 0197 036 0105 0804 2001 0966 1000
Sulfatides 2047 0033b 0058 2049 0023b 0026b 2002 0922 1000 034 0133 0209
Tetrasialoganglioside-GQ1B 2043 0054 0067 2057 0007b 0025b 023 0319 0854 048 0027a 0056
Continued4 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
Biologicals (Littleton CO) Assay Designs (Ann Arbor MI) Pro-
Sci Inc (Poway CA) EMD Biosciences (San Diego CA) Cay-
man Chemical (Ann Arbor MI) HyTest (Turku Finland)
Meridian Life Science (Memphis TN) and Biodesign Interna-
tional (Saco ME) The antigens used in the construction of anti-
gen microarrays are listed in table e-1 at Neurologyorgnn
Antigen microarray production development and dataanalysis The antigens listed in table e-1 were spotted in replicatesof 6 on SuperEpoxy 2 slides (TeleChem Sunnyvale CA) using
an Arrayit NanoPrint 2 LM210 microarray printer (Arrayit Corpo-
ration Sunnyvale CA) and optimized spotting conditions as
described28303140 The microarrays were hybridized using an HS
4800Pro Hybridization Station (Tecan Maumlnnedorf Switzerland)
in which they were blocked with 1 bovine serum albumin for
1 hour and incubated for 2 hours at 37degC with the test serum at
a 1100 dilution in blocking buffer The arrays were then washed
and incubated for 45 minutes with a 1500 dilution of goat
anti-human immunoglobulin G (IgG) Cy3-conjugated and
goat anti-human IgM Cy5-conjugated detection antibodies (Jackson
ImmunoResearch Labs West Grove PA) The arrays were scanned
with a Tecan PowerScanner Repeated measurements indicate that
our antigen microarray technique is reproducible exhibiting
a coefficient of variation of 1336 1231
Background signal was subtracted and raw data were normal-
ized and analyzed using the GeneSpring software (Silicon Genetics
Redwood City CA) Antigen reactivity was defined by the mean
intensity of binding to the replicates of that antigen on the micro-
array and expressed as relative fluorescence units Scatter plots were
generated using linear regression models in the R statistical package
Pearson product-moment correlation coefficients were calculated
between the MRI measures (BPF GMF WMF T2LV) and the
weighted average of a group of statistically significant lipids The
weighted average of significant lipids was calculated using the for-
mula (SWiAiSWi) (SWi 5 1 i 5 1 2 3 ) where Wi is the
proportion of intensity of antigen Ai Ai is the observed intensity of
an antigen and SWi is the sum of the weights
RESULTS Serum IgG antibodies correlate with brain
MRI measures of disease severity To study the relation-ship between the peripheral immune response andMRI measures of disease severity we analyzed serum
antibody reactivity in those MS samples We analyzedIgG serum antibodies using a panel of antigensincluding CNS antigens heat shock proteins and lip-ids The association between the antibody reactivityagainst each antigen and 4 MRI measures of disease(T2LV BPF WMF and GMF) was investigatedusing Spearman correlation tests
We found significant associations between eachMRI measure of disease and different sets of IgG anti-body reactivity which are shown in table 2 Similarpatterns of antibody reactivity were linked to BPFand GMF consistent with the known dominant con-tribution of GM atrophy to whole brain atrophymeasures3741 Strikingly there was little overlapbetween the antibody reactivity associated withGMF and WMF suggesting that different immuno-pathologic processes contribute to tissue degenerationin these areas of the brain Similarly these profiles ofantibody reactivity were also different from thoseassociated with T2LV
Serum lipid-reactive IgGs are associated with increased
disease pathogenesis determined by MRI In evaluationof MRI measures and their link to disease BPFGMF and WMF decrease with disease progressionwhile T2LV increases with disease progressionAccordingly we analyzed the linkage between eachMRI measure and the significant antibody reactivitiesshown in table 2 We identified IgG antibody reac-tivities associated with increased tissue destruction asevidenced by decreased BPF values (figure 1) Strik-ingly we found that antibodies linked to diseasepathology as measured by BPF were enriched for reac-tivity against lipids Furthermore a selective increasein lipid-reactive antibodies linked to disease severitywas also observed when we analyzed all available MRImeasures (figure 2) Of note we did not detect an
Table 2 Continued
Antigens
BPF GMF WMF T2LV
r p FDR r p FDR r p FDR r p FDR
Thyrocalcitonin 2033 0143 0258 2008 0746 0996 2059 0005b 0108 004 0878 1000
TNPAL-galactocerebroside 007 0763 0969 2015 0506 0996 047 0030a 0142 005 0823 1000
Total cerebroside 2016 0498 0928 2041 0064 0112 050 0020a 0122 062 0003a 0033
Trisialogang GT1b 2034 0128 0194 2044 0043b 0066 015 0513 1000 030 0185 0247
Troponin I 2051 0020b 0040b 2053 0013b 0026b 2003 0900 1000 043 0050 0072
Ubiquitin 031 0166 0305 045 0039a 0027 2025 0282 0854 2029 0196 0247
VEGF 2050 0023b 0040b 2047 0031b 0027b 2013 0570 1000 040 0070 0146
Abbreviations BPF 5 whole brain parenchymal fraction FDR 5 false discovery ratendashadjusted significance value GMF 5 global cerebral gray matterfraction T2LV 5 cerebral T2 (fluid-attenuated inversion recovery) hyperintense lesion volume WMF 5 global cerebral white matter fractionDiscovery setap 005 (positive association)bp 005 (negative association)
Neurology Neuroimmunology amp Neuroinflammation 5
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
enrichment in lipid-reactive IgM in the group ofantibodies associated with increased diseasepathogenesis (not shown)
An index of serum IgG reactivity to lipids is associated
with MRI measures of disease severity To further inves-tigate the association between serum IgG reactivityto lipids and MRI measures of disease severity wecalculated an IgG anti-lipid index for each patientcorresponding to the information on each lipid-specific antibody listed in table 2 normalized by thestrength of its correlation with that MRI measureunder investigation The antibody reactivities tolipids used to calculate the lipid index are shown in
figure 3A With this approach we calculated one indexfor each of the MRI measures analyzed in this studyWe found a significant correlation between each IgGlipid antibody index and BPF GMF and T2LV(figure 3B) no significant correlation was found withWMF Moreover no significant correlations werefound between BPF WMF GMF and T2LVmeasures and anti-lipid antibody indexes based onIgM reactivity (not shown)
Finally we evaluated the performance of the lipidantibody indexes linked to BPF GMF and T2LV onan additional set of independent MS samples (valida-tion set table 1) We detected a similar trend to theone detected in the discovery set with regard to the
Figure 1 Antibody reactivity in serum is associated with decreased brain volume
Heatmap in which each column represents the mean immunoglobulin G antibody reactivity in a serum sample from a patient with multiple sclerosis sortedaccording to the whole brain parenchymal fraction (BPF) (indicated at the topmdasha lower BPF indicates more brain atrophy) and each row represents theantibody reactivity to an antigen according to the colorimetric scale shown The antibody reactivities included in this heatmap are listed in table 2
6 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
Figure 2 Association of serum IgG reactivity with MRI measures of disease severity
(A) Heatmap in which each column represents an MRI measure of either atrophy (BPF GMF WMF) or lesions (T2LV) andeach row represents the correlation to IgG serum antibody reactivity according to a colorimetric scale (B) Frequency oflipid-reactive antibodies linked to higher or lower MRI disease severity BPF 5 whole brain parenchymal fraction GMF 5
global cerebral gray matter fraction IgG 5 immunoglobulin G T2LV 5 cerebral T2 (fluid-attenuated inversion recovery)hyperintense lesion volume WMF 5 global cerebral white matter fraction
Neurology Neuroimmunology amp Neuroinflammation 7
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
correlation between BPF and GMF and their respec-tive lipid indexes and we validated the correlationbetween T2LV and its lipid index (figure 3C)
DISCUSSION Previous studies have investigated theassociation of MRI with immune activity in MS13ndash19
These studies have included cellular immune measures
Figure 3 Correlation of lipid indexes to MRI measures of disease severity in multiple sclerosis
(A) Heatmap inwhich each column represents anMRImeasure and each row represents the correlation to immunoglobulin G serumantibody reactivity to lipids accord-ing to a colorimetric scale (B) Scatter plots depicting the correlation between each lipid index andMRImeasures (BPF GMF and T2LV) in the discovery set (C) Scatterplots depicting the correlation between each lipid index andMRImeasures (BPF GMF and T2LV) in the validation set BPF5whole brain parenchymal fraction GMF5
cerebral gray matter fraction (see methods section for details) T2LV 5 cerebral T2 hyperintense lesion volume WMF 5 global cerebral white matter fraction
8 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
as well as oligoclonal bands or neurofilament-specificantibodies in CSF42ndash44 In the present study weanalyzed the association between serum antibodyprofiles detected with antigen microarrays and MRImeasures of disease including cerebral atrophy andT2 hyperintense lesions We found that specificantibody patterns are associated with different aspectsof MRI-defined disease pathology Our major findingwas that increased reactivity to lipids is associated withdifferent aspects of brain MRI measures of diseaseseverity Of note the specific set of lipids associatedwith atrophy differed from those associated withlesions Taken together these data suggest that anti-lipid antibodies in serum are related to MRI measuresof disease Our findings highlight the important role oflipid and lipid-specific immunity in the pathogenesis ofMS
Although a potential limitation of these studies isthe relatively small number of samples analyzed wehave validated our findings in an independent valida-tion cohort strengthening their significance In addi-tion further studies in large cohorts from patientsaffected by non-MS neurodegenerative diseases andhealthy controls may indicate whether the antibodyreactivities and lipid indexes identified in these stud-ies are exclusive to MS or are associated with addi-tional biological processes Indeed some of theseantibody reactivities contributing to the immune sig-natures described in this work have been found innewborns45
Different sets of antibody reactivities were associ-ated with GMF WMF BPF and T2LV The differ-ent antibody link between lesions and atrophy is inkeeping with the long-held view that brain atrophyin patients with MS is multifactorial and is onlyweakly related to lesions41 This has been underscoredin recent studies showing the complementary infor-mation obtained by combining lesion and atrophymeasures4647
Among the cerebral atrophy measures there was astriking overlap between those antibodies associatedwith GMF and BPF which separated them fromWMF results These findings are in keeping with pre-vious observations that whole brain atrophy is domi-nated by GM loss3741 In the early stages of MS thisGM atrophy selectively involves the deep GM nuclei34
Thus the similarities observed in the autoantibodieslinked to GMF and BPF might reflect the dominantcontribution and colinearity of GM atrophy to totalbrain atrophy The set of antibodies linked to GMFmay be the most relevant given that MRI studies haveshown that GM volume is more closely linked to phys-ical disability37 and cognitive impairment38 than areWM volume or lesion measures
Using antigen microarrays antibody reactivity tolipids has been detected in the CSF and serum of
patients with MS at different stages of the diseaseand such antibodies in the CSF have been linkedto disease progression and MRI involvement44
Although several mechanisms are thought to drivebrain inflammation and atrophy in MS46 the connec-tion between these mechanisms and the antibodiesdetected with our antigen microarrays is yet unknownOne possibility is that the serum autoantibodies iden-tified in our studies are not pathogenic and reflect theresult of immunization against self-antigens releasedfrom the CNS during the course of the disease Indeedneurofilament is released from damaged axons duringthe course of MS and both neurofilament lightchains48 and antibodies against it42 have been linkedto MRI measures of disease Moreover heat shockproteins are upregulated in different cell types duringinflammation and have been shown to have an impor-tant role as immunomodulators when released to theextracellular medium4950 Thus it is possible that heatshock proteinndashreactive antibodies reflect changes in theproduction and release of these immunomodulatorsduring the course of disease pathogenesis Similarlybioactive lipids and their products are released fromdamaged myelin and lipid-specific antibodies havebeen detected in patients with MS27293051 Lipids havean important role in the immune response both asbioactive molecules with immunomodulatory proper-ties and also as targets of the adaptive immuneresponse52 Moreover lipids have significant effectson the murine model of MS experimental autoim-mune encephalomyelitis2729305153 Indeed we recentlyfound that the glycolipid lactosylceramide activates abroad set of biological processes in astrocytes promot-ing neurodegeneration and inflammation53 Thus thelipid-reactive antibodies detected in this work mayreflect the release of myelin lipids in the context ofMS pathogenesis andor may directly contribute toimmune-mediated damage in CNS tissue Of notelipid-reactive antibodies are highly cross-reactive52 Inaddition the lipid-reactive antibodies detected in thiswork as associated with MRI measures of disease activ-ity were of the IgG class These are important points toconsider when evaluating a potential role of lipid-reactive antibodies inMS pathogenesis Further studiesare warranted to investigate whether bioactive lipidsoffer new therapeutic targets in MS
We have found that unique patterns of immunereactivity determined with antibody arrays are associ-ated with specific MRI measures of disease severityThese patterns agree with the interpretation that dif-ferent pathogenic mechanisms drive diverse diseaseprocesses reflected by these MRI measures These pat-terns also suggest a predominant role for lipids andlipid-specific immunity in MS pathology Furtherstudies are warranted to determine whether the earlyappearance of anti-lipid antibodies predicts the
Neurology Neuroimmunology amp Neuroinflammation 9
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
subsequent development of clinical and MRI-defineddisease worsening
AUTHOR CONTRIBUTIONSRohit Bakshi draftingrevising the manuscript study concept or design
analysis or interpretation of data acquisition of data study supervision
obtaining funding Ada Yeste study concept or design acquisition of
data Bonny Patel analysis or interpretation of data statistical analysis
Shahamat Tauhid analysis or interpretation of data Subhash Tummala
analysis or interpretation of data Roya Rahbari analysis or interpretation
of data statistical analysis Renxin Chu analysis or interpretation of data
Keren Regev analysis or interpretation of data acquisition of data
Pia Kivisaumlkk draftingrevising the manuscript study concept or design
contribution of vital reagentstoolspatients acquisition of data study
supervision Howard L Weiner study concept or design obtaining fund-
ing Francisco J Quintana draftingrevising the manuscript study con-
cept or design contribution of vital reagentstoolspatients study
supervision obtaining funding
ACKNOWLEDGMENTThe authors thank Dr Mohit Neema Dr Antonella Ceccarelli and
Dr Ashish Arora for valuable assistance at the early stages of this project
STUDY FUNDINGThis work was supported in part by grants to FJQ and HLW from
EMD Serono RG4111A1 and JF2161-A-5 from the National Multiple
Sclerosis Society PA0069 from the International Progressive MS Alli-
ance and by grants from the NIH (5R01NS055083-04) and NMSS
(RG3798A2) to RB
DISCLOSUREFJ Quintana serves on the editorial board for Systems Biomedicine
Inmunologia American Journal of Clinical and Experimental Immunology
is an associate editor for Immunology (UK) is an advisory board member
for Seminars in Immunopathology received research support from Harvard
Medical School BADERC NMSS A Yeste and B Patel report no
disclosures S Tauhid is managing editor for Journal of Neuroimaging
S Tummala R Rahbari R Chu and K Regev report no disclosures
P Kivisaumlkk received research support from EMD Serono R Bakshi is
editor-in-chief for Journal of Neuroimaging received consulting fees from
AbbVie Alkermes Biogen Novartis Questcor received research support
from Biogen EMD-Serono Novartis Sanofi-Genzyme Teva his spouse
holds stock in Biogen Inc H Weiner served on the advisory board
for The Guthy-Jackson Charitable Foundation Teva Pharmaceuticals
Industries Ltd Biogen Idec Novartis Sanofi-Aventis has consulted for
Therapix Bioven Novartis Serono Teva Sanofi received research support
from National Multiple Sclerosis Society Go to Neurologyorgnn for full
disclosure forms
Received July 27 2015 Accepted in final form December 8 2015
REFERENCES1 Weiner HL The challenge of multiple sclerosis how do
we cure a chronic heterogeneous disease Ann Neurol
200965239ndash248
2 Bakshi R Thompson AJ Rocca MA et al MRI in mul-
tiple sclerosis current status and future prospects Lancet
Neurol 20087615ndash625
3 Ceccarelli A Bakshi R Neema M MRI in multiple scle-
rosis a review of the current literature Curr Opin Neurol
201225402ndash409
4 Filippi M Rocca MA Barkhof F et al Association
between pathological and MRI findings in multiple scle-
rosis Lancet Neurol 201211349ndash360
5 Geurts JJ Calabrese M Fisher E Rudick RA Measure-
ment and clinical effect of grey matter pathology in mul-
tiple sclerosis Lancet Neurol 2012111082ndash1092
6 Klaver R De Vries HE Schenk GJ Geurts JJ Grey mat-
ter damage in multiple sclerosis a pathology perspective
Prion 2013766ndash75
7 Khoury S Bakshi R Cerebral pseudoatrophy or real atro-
phy after therapy in multiple sclerosis Ann Neurol 2010
68778ndash779
8 Hauser SL Chan JR Oksenberg JR Multiple sclerosis
prospects and promise Ann Neurol 201374317ndash327
9 McFarland HF Martin R Multiple sclerosis a complicated
picture of autoimmunity Nat Immunol 20078913ndash919
10 Nylander A Hafler DA Multiple sclerosis J Clin Invest
20121221180ndash1188
11 Sospedra M Martin R Immunology of multiple sclerosis
Ann Rev Immunol 200523683ndash747
12 Steinman L Immunology of relapse and remission in mul-
tiple sclerosis Ann Rev Immunol 201432257ndash281
13 Khoury SJ Guttmann CR Orav EJ Kikinis R Jolesz FA
Weiner HL Changes in activated T cells in the blood
correlate with disease activity in multiple sclerosis Arch
Neurol 2000571183ndash1189
14 Khoury SJ Orav EJ Guttmann CR Kikinis R Jolesz FA
Weiner HL Changes in serum levels of ICAM and TNF-
R correlate with disease activity in multiple sclerosis Neu-
rology 199953758ndash764
15 Laplaud DA Berthelot L Miqueu P et al Serial blood T
cell repertoire alterations in multiple sclerosis patients cor-
relation with clinical and MRI parameters J Neuroimmunol
2006177151ndash160
16 Makhlouf K Weiner HL Khoury SJ Increased percentage
of IL-121 monocytes in the blood correlates with the
presence of active MRI lesions in MS J Neuroimmunol
2001119145ndash149
17 Prat A Biernacki K Saroli T et al Kinin B1 receptor expres-
sion on multiple sclerosis mononuclear cells correlation with
magnetic resonance imaging T2-weighted lesion volume and
clinical disability Arch Neurol 200562795ndash800
18 Rinaldi L Gallo P Calabrese M et al Longitudinal anal-
ysis of immune cell phenotypes in early stage multiple
sclerosis distinctive patterns characterize MRI-active pa-
tients Brain 20061291993ndash2007
19 Tortorella P Lagana MM Saresella M et al Determinants
of disability in multiple sclerosis an immunological and
MRI study Biomed Res Int 20142014875768
20 Robinson WH DiGennaro C Hueber W et al Autoan-
tigen microarrays for multiplex characterization of autoan-
tibody responses Nat Med 20028295ndash301
21 Yeste A Quintana FJ Antigen microarrays for the study of
autoimmune diseases Clin Chem 2013591036ndash1044
22 Li QZ Xie C Wu T et al Identification of autoantibody
clusters that best predict lupus disease activity using glomer-
ular proteome arrays J Clin Invest 20051153428ndash3439
23 Hueber W Kidd BA Tomooka BH et al Antigen micro-
array profiling of autoantibodies in rheumatoid arthritis
Arthritis Rheum 2005522645ndash2655
24 Goldschmidt Y Sharon E Quintana FJ Cohen IR
Brandt A Adaptive methods for classification of biological
microarray data from multiple experiments Report no
MCS03-07 Rehovot Israel The Arthur and Rochelle Bel-
fer Institute of Mathematics and Computer Science 2003
25 Quintana FJ Getz G Hed G Domany E Cohen IR
Cluster analysis of human autoantibody reactivities in
health and in type 1 diabetes mellitus a bio-informatic
approach to immune complexity J Autoimmun 2003
2165ndash75
10 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
26 Lalive PH Menge T Barman I Cree BA Genain CP
Identification of new serum autoantibodies in neuromye-
litis optica using protein microarrays Neurology 200667
176ndash177
27 Kanter JL Narayana S Ho PP et al Lipid microarrays
identify key mediators of autoimmune brain inflamma-
tion Nat Med 200612138ndash143
28 Quintana FJ Farez MF Izquierdo G Lucas M Cohen IR
Weiner HL Antigen microarrays identify CNS-produced
autoantibodies in RRMS Neurology 201278532ndash539
29 Farez MF Quintana FJ Gandhi R Izquierdo G Lucas M
Weiner HL Toll-like receptor 2 and poly(ADP-ribose)
polymerase 1 promote central nervous system neuroin-
flammation in progressive EAE Nat Immunol 200910
958ndash964
30 Quintana FJ Farez MF Viglietta V et al Antigen micro-
arrays identify unique serum autoantibody signatures in
clinical and pathologic subtypes of multiple sclerosis Proc
Natl Acad Sci USA 200810518889ndash18894
31 Quintana FJ Patel B Yeste A et al Epitope spreading as
an early pathogenic event in pediatric multiple sclerosis
Neurology 2014832219ndash2226
32 Gauthier SA Glanz BI Mandel M Weiner HL A model
for the comprehensive investigation of a chronic autoim-
mune disease the multiple sclerosis CLIMB study Auto-
immun Rev 20065532ndash536
33 Polman CH Reingold SC Edan G et al Diagnostic criteria
for multiple sclerosis 2005 revisions to the ldquoMcDonald
Criteriardquo Ann Neurol 200558840ndash846
34 DellrsquoOglio E Ceccarelli A Glanz BI et al Quantification
of global cerebral atrophy in multiple sclerosis from 3T
MRI using SPM the role of misclassification errors
J Neuroimaging 201525191ndash199
35 Ceccarelli A Jackson JS Tauhid S et al The impact of
lesion in-painting and registration methods on voxel-based
morphometry in detecting regional cerebral gray matter
atrophy in multiple sclerosis AJNR Am J Neuroradiol
2012331579ndash1585
36 Cohen AB Neema M Arora A et al The relationships
among MRI-defined spinal cord involvement brain involve-
ment and disability in multiple sclerosis J Neuroimaging
201222122ndash128
37 Sanfilipo MP Benedict RH Sharma J Weinstock-
Guttman B Bakshi R The relationship between whole
brain volume and disability in multiple sclerosis a com-
parison of normalized gray vs white matter with misclas-
sification correction Neuroimage 2005261068ndash1077
38 Sanfilipo MP Benedict RH Weinstock-Guttman B
Bakshi R Gray and white matter brain atrophy and neuro-
psychological impairment in multiple sclerosis Neurology
200666685ndash692
39 Stankiewicz JM Glanz BI Healy BC et al Brain MRI
lesion load at 15T and 3T versus clinical status in multi-
ple sclerosis J Neuroimaging 201121e50ndashe56
40 Quintana FJ Hagedorn PH Elizur G Merbl Y
Domany E Cohen IR Functional immunomics micro-
array analysis of IgG autoantibody repertoires predicts the
future response of mice to induced diabetes Proc Natl
Acad Sci USA 2004101(suppl 2)14615ndash14621
41 Bermel RA Bakshi R The measurement and clinical
relevance of brain atrophy in multiple sclerosis Lancet
Neurol 20065158ndash170
42 Eikelenboom MJ Petzold A Lazeron RH et al Multiple
sclerosis neurofilament light chain antibodies are corre-
lated to cerebral atrophy Neurology 200360219ndash223
43 Magraner MJ Bosca I Simo-Castello M et al Brain atro-
phy and lesion load are related to CSF lipid-specific IgM
oligoclonal bands in clinically isolated syndromes Neuro-
radiol 2012545ndash12
44 Villar LM Sadaba MC Roldan E et al Intrathecal syn-
thesis of oligoclonal IgM against myelin lipids predicts an
aggressive disease course in MS J Clin Invest 2005115
187ndash194
45 Merbl Y Zucker-Toledano M Quintana FJ Cohen IR
Newborn humans manifest autoantibodies to defined self
molecules detected by antigen microarray informatics
J Clin Invest 2007117712ndash718
46 Sormani MP Arnold DL De Stefano N Treatment effect
on brain atrophy correlates with treatment effect on disa-
bility in multiple sclerosis Ann Neurol 20147543ndash49
47 Tauhid S Neema M Healy BC Weiner HL Bakshi R
MRI phenotypes based on cerebral lesions and atrophy in
patients with multiple sclerosis J Neurol Sci 2014346
250ndash254
48 Gnanapavan S Grant D Morant S et al Biomarker
report from the phase II lamotrigine trial in secondary
progressive MS neurofilament as a surrogate of disease
progression PLoS One 20138e70019
49 Quintana FJ Cohen IR Heat shock proteins as endoge-
nous adjuvants in sterile and septic inflammation
J Immunol 20051752777ndash2782
50 Quintana FJ Cohen IR The HSP60 immune system net-
work Trends Immunol 20113289ndash95
51 Ho PP Kanter JL Johnson AM et al Identification of
naturally occurring fatty acids of the myelin sheath that
resolve neuroinflammation Sci Transl Med 20124137ra73
52 Quintana FJ Yeste A Weiner HL Covacu R Lipids and
lipid-reactive antibodies as biomarkers for multiple sclero-
sis J Neuroimmunol 201224853ndash57
53 Mayo L Trauger SA Blain M et al Regulation of astro-
cyte activation by glycolipids drives chronic CNS inflam-
mation Nat Med 2014201147ndash1156
Neurology Neuroimmunology amp Neuroinflammation 11
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
1
Table e-1 Antigens used
Group Antigen Source
27 kDa Heat Shock Protein Stressgen
HSP 32 kDa Heat Shock Protein Stressgen
40 kDa Heat Shock Protein Stressgen
47 kDa Heat Shock Protein Stressgen
60 kDa Heat Shock Protein Stressgen
60 kDa Heat Shock Protein peptide aa 106ndash125 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 121ndash140 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 136ndash155 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 151ndash170 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 16ndash35 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 166ndash185 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
2
60 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 556ndash573 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
65 kDa Heat Shock Protein M tuberculosis Stressgen
70 kDa Heat Shock Protein Stressgen
70 kDa Heat Shock Protein peptide aa 106ndash125 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 121ndash140 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 136ndash155 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 151ndash170 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 16ndash35 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 166ndash185 Biopolymers Facility Harvard Medical School
3
70 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
4
70 kDa Heat Shock Protein peptide aa 556ndash575 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 571ndash590 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 586ndash605 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 601ndash620 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 616ndash635 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 631ndash640 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
71 kDa Heat Shock Protein M tuberculosis Biopolymers Facility Harvard Medical School
90 kDa Heat Shock Protein Stressgen
GroEL Stressgen
23-cyclic nucleotide 3-phosphodiesterase
peptide aa106-125 Biopolymers Facility Harvard Medical School
CNS
23-cyclic nucleotide 3-phosphodiesterase
peptide aa1-20 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa121-140 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa136-155 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa151-170 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa16-35 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa166-185 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase Biopolymers Facility Harvard Medical School
5
peptide aa181-200
23-cyclic nucleotide 3-phosphodiesterase
peptide aa196-215 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa211-230 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa226-245 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa241-260 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa256-275 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa271-290 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa286-305 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa301-320 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa31-50 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa316-335 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa331-350 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa346-365 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa361-380 Biopolymers Facility Harvard Medical School
6
23-cyclic nucleotide 3-phosphodiesterase
peptide aa376-395 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa391-410 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa406-421 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa46-65 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa61-80 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa76-95 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa91-110 Biopolymers Facility Harvard Medical School
Acetyl Cholinesterase Sigma Aldrich
ADAM-10 Sigma Aldrich
alpha-Cristallin Stressgen
beta-Cristallin Sigma Aldrich
bovine Myelin Basic Protein Sigma Aldrich
Brain Extract I Sigma Aldrich
Brain Extract II Sigma Aldrich
Brain Extract III Sigma Aldrich
Glial Filament Acidic Protein Research Diagnostic
guinea pig Myelin Basic Protein Sigma Aldrich
human Myelin Basic Protein Sigma Aldrich
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 106-125 Biopolymers Facility Harvard Medical School
7
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 121-140 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 136-155 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 151-170 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 166-185 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 181-200 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 91-110 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
106-125 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa 1- Biopolymers Facility Harvard Medical School
8
20
Myelinoligodendrocyte glycoprotein peptide aa
121-140 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
136-155 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
151-170 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
16-35 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
166-185 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
181-200 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
196-215 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
211-230 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
226-247 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
31-50 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
35-55 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
46-65 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
61-80 Biopolymers Facility Harvard Medical School
9
Myelinoligodendrocyte glycoprotein peptide aa
76-95 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
91-110 Biopolymers Facility Harvard Medical School
murine Myelin Basic Protein Sigma Aldrich
Myelin Associated Glycoprotein Sigma Aldrich
Myelin Basic Protein peptide aa 104-123 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 11-30 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 113-132 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 121-138 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 124-142 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 138-147 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 141-161 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 143-168 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 155-178 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 26-35 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 41-60 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 51-70 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 71-92 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 84-94 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 89-101 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 173-186 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 93-112 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 106-125 Biopolymers Facility Harvard Medical School
10
Myelin Protein 2 peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 121-132 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 91-110 Biopolymers Facility Harvard Medical School
Neurofilament 160kd Chemicon
Neurofilament 200kd Chemicon
Neurofilament 68kd Chemicon
Neuronal Enolase Calbiochem
Nicastrin GeneTex
NMDA receptor Novus Biologicals
NOGO Sigma Aldrich
Olygodendrocyte-Specific Protein peptide aa 106ndash
125 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 121ndash
140 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 136ndash
155 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 151ndash
170 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 16ndash
35 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 166ndash Biopolymers Facility Harvard Medical School
11
185
Olygodendrocyte-Specific Protein peptide aa 181ndash
199 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 195ndash
217 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 31ndash
50 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 46ndash
65 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 61ndash
80 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 76ndash
95 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 91ndash
110 Biopolymers Facility Harvard Medical School
Proteolipid Protein Abnova
Proteolipid Protein peptide aa 100-119 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 10-29 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 110-129 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 1-19 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 125-141 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-150 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-154 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 150-163 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 151-173 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 158-166 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 161-180 Biopolymers Facility Harvard Medical School
12
Proteolipid Protein peptide aa 178-191 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 180-199 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 190-209 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 20-39 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 205-220 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 215-232 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-239 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-249 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 248-259 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 250-269 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 265-277 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 35-50 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 40-59 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 50-69 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 65-84 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 80-99 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 91-110 Biopolymers Facility Harvard Medical School
Retinol Binding Protein Sigma Aldrich
S100beta protein Assay Designs
Super Oxide Dismutase Sigma Aldrich
Synuclein beta Sigma Aldrich
Synuclein gamma Sigma Aldrich
Amydgala ProSci Inc
Tissue Amydgala AD ProSci Inc
Brain lysate ProSci Inc
Brain Tissue Membrane ProSci Inc
Cerebellar pedunculus ProSci Inc
13
Cerebral meninges ProSci Inc
Corpus Callosum ProSci Inc
Corpus Callosum AD ProSci Inc
Diencephalon ProSci Inc
Fetal brain ProSci Inc
Frontal lobe ProSci Inc
Frontal lobe AD ProSci Inc
Hippocampus ProSci Inc
Hippocampus AD ProSci Inc
Insula ProSci Inc
Occipital lobe ProSci Inc
Occipital lobe AD ProSci Inc
Olfactory region ProSci Inc
Optic Nerve ProSci Inc
Parietal lobe ProSci Inc
Parietal lobe AD ProSci Inc
Pons ProSci Inc
Pons AD ProSci Inc
Postcentral gyrus ProSci Inc
Postcentral gyrus AD ProSci Inc
Precentral gyrus ProSci Inc
Precentral gyrus AD ProSci Inc
Spinal cord ProSci Inc
Temporal lobe ProSci Inc
Temporal lobe AD ProSci Inc
Thalamus ProSci Inc
Thalamus AD ProSci Inc
14
Amyloid beta Sigma Aldrich
AD
related Amyloid beta 10-20 Sigma Aldrich
Amyloid beta 1-12 Sigma Aldrich
Amyloid beta 12-28 Sigma Aldrich
Amyloid beta 1-23 Sigma Aldrich
Amyloid beta 1-38 Sigma Aldrich
Amyloid beta 17-40 Sigma Aldrich
Amyloid beta 25-35 Sigma Aldrich
Amyloid beta 34-42 Sigma Aldrich
Amyloid bri protein precursor 227 Sigma Aldrich
Amyloid DAN Protein Fragment 1-34 Sigma Aldrich
Amyloid Precursor Protein Sigma Aldrich
Amyloid protein no AB component Sigma Aldrich
Secreted amyloid precursor protein (SAP) beta Sigma Aldrich
Tau isoform variant 0N3R Sigma Aldrich
Tau isoform variant 1N3R Sigma Aldrich
Tau isoform variant 0N4R Sigma Aldrich
Tau isoform variant 2N3R Sigma Aldrich
Tau phospho Ser412 Sigma Aldrich
Tau phospho Ser441 Sigma Aldrich
Tau phospho Thr181 Sigma Aldrich
Tau Protein human EMD Biosciences
(plusmn)9-HODE Cayman Chemical
Lipids
1 Palmitoyl-2-(5oxo-Valeroyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
15a-hydroxycholestene Avanti Polar Lipids
15
15-ketocholestane Avanti Polar Lipids
15-ketocholestene Avanti Polar Lipids
1-Palmitoil-2-(9oxo-Nonanoyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Azelaoyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Glutaroyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
5 α-cholestane-3 β15 α-diol Avanti Polar Lipids
9(S)-HODE Cayman Chemical
Asialoganglioside-GM1 Sigma Aldrich
Asialoganglioside-GM2 Sigma Aldrich
Brain ceramides Avanti Polar Lipids
Brain D-erythrosphingosine Avanti Polar Lipids
Brain lysophosphatidylethanolamine Avanti Polar Lipids
Brain L-α-lysophosphatidylserine Avanti Polar Lipids
Brain L-α-phosphatidylcholine Avanti Polar Lipids
Brain L-α-phosphatidyl-ethanolamine Avanti Polar Lipids
Brain L-α-phosphatidylserine Avanti Polar Lipids
Brain polar lipid extract Avanti Polar Lipids
Brain sphingomyelin Avanti Polar Lipids
Brain sulfatide Avanti Polar Lipids
Brain total lipid extract Avanti Polar Lipids
Cardiolipin Sigma Aldrich
Ceramide Sigma Aldrich
Ceramide 1-phosphate Sigma Aldrich
Cholesterol Sigma Aldrich
16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-

Table 2 Serum immunoglobulin Gs associated with MRI measures of disease severity
Antigens
BPF GMF WMF T2LV
r p FDR r p FDR r p FDR r p FDR
1-P-2-5-o-V-sn-G-3-PC 2038 0087 0161 2034 0131 0197 2014 0540 1000 056 0008a 0056
15-ketocholestene 2030 0190 0370 2044 0047b 0066 024 0302 0854 030 0189 0247
9 HODE 2040 0073 0097 2051 0019b 0026b 016 0499 1000 045 0043a 0057
9 S-HODE 2042 0057 0067 2048 0027b 0026b 006 0797 1000 061 0003a 0033a
a-MSH 050 0021a 0040a 040 0070 0112 028 0214 0854 2004 0858 1000
ABPF 1-12 2023 0320 0610 2040 0070 0112 032 0155 0804 046 0035a 0057
ABPP 227 2030 0179 0305 2056 0009b 0025b 047 0033a 0156 049 0024a 0056
Ac phosphatase 035 0119 0194 047 0033a 0027a 2018 0443 1000 2020 0374 0516
ADPF 1-34 2020 0393 0823 2038 0089 0133 034 0126 0804 046 0035a 0057
ANP 013 0587 0969 037 0100 0197 2048 0027b 0142 2027 0242 0516
Asialoganglioside-GM1 2052 0016b 0040b 2060 0004b 0015b 007 0749 1000 032 0152 0209
Asialoganglioside-GM2 2040 0074 0097 2048 0027b 0026b 011 0648 1000 024 0292 0516
b-MSH 2047 0033b 0058 2038 0085 0133 2025 0277 0854 020 0376 0516
b-Amyloid 2021 0350 0610 2036 0107 0197 026 0247 0854 049 0025a 0056
b-Synuclein 001 0954 0969 2022 0343 0996 048 0029a 0142 042 0055 0094
bNGF 029 0204 0413 006 0783 0996 051 0018a 0122 2006 0807 1000
Brain D-erythrosphingosine 2047 0030b 0058 2056 0009b 0025b 010 0679 1000 037 0096 0146
Brain extract VII 2052 0016b 0040b 2059 0005b 0015b 006 0789 1000 053 0013a 0056
Brain L-a-phosphatidyl-ethanolamine 2044 0044b 0067 2046 0037b 0027b 2005 0837 1000 019 0407 0537
Brain polar lipid extract 013 0573 0969 2010 0657 0996 051 0019a 0122 018 0434 0537
Chorionic G 005 0826 0969 028 0223 0303 2046 0034b 0156 2010 0656 1000
CNPase aa 106ndash125 039 0080 0161 049 0026a 0026a 2014 0557 1000 2051 0017b 0056
CNPase aa 16ndash35 042 0056 0067 046 0037a 0027a 2002 0941 1000 2037 0103 0157
CNPase aa 195ndash214 031 0173 0305 050 0020a 0026a 2035 0119 0804 2035 0118 0157
CNPase aa 286ndash305 028 0227 0413 045 0039a 0027a 2033 0144 0804 2004 0873 1000
CNPase aa 376ndash395 019 0416 0823 043 0049a 0066 2049 0024b 0142 2038 0089 0146
Collagen II 025 0278 0610 046 0034a 0027a 2040 0072 0788 2032 0151 0209
Collagen IX 2067 0001b 0025b 2075 0001b 0002b 008 0741 1000 066 0001a 0033a
Collagen VIII 2027 0242 0521 2026 0251 0605 2005 0831 1000 044 0044a 0057
Collagen X 2036 0114 0194 2012 0595 0996 2052 0016b 0122 2006 0792 1000
Disialoganglioside-GD2 2040 0073 0097 2049 0023b 0026b 013 0589 1000 043 0055 0094
Disialogang-GD1B 2038 0088 0161 2050 0022b 0026b 018 0441 1000 031 0174 0247
Disialoganglioside GD1a 2051 0017b 0040b 2053 0013b 0026b 2005 0824 1000 042 0060 0146
Disialoganglioside GD3 2040 0069 0097 2054 0012b 0026b 020 0382 1000 051 0018a 0056
Galactocerebrosides 001 0952 0969 2027 0229 0303 059 0005a 0108 025 0265 0516
Ganglioside mixture 2053 0014b 0040b 2062 0003b 0015b 011 0644 1000 053 0014a 0056
Ganglioside-GM4 002 0929 0969 029 0203 0303 2056 0009b 0109 2018 0423 0537
Gelsolin 053 0013a 0040a 057 0008a 0025a 002 0934 1000 2048 0029b 0056
GNRH 2053 0014b 0040b 2043 0054 0072 2029 0196 0854 036 0113 0157
gpMBP 017 0449 0928 2004 0857 1000 048 0029a 0142 000 0988 1000
GT1a 2043 0053 0067 2048 0028b 0026b 003 0912 1000 032 0161 0247
Hemoglobin 015 0527 0969 2006 0784 0996 046 0037a 0203 028 0212 0516
HSP60 aa 195ndash214 023 0311 0610 004 0879 1000 044 0045a 0206 000 0989 1000
HSP60 aa 210ndash229 053 0015a 0040a 049 0024a 0026a 016 0493 1000 2013 0577 1000
ContinuedNeurology Neuroimmunology amp Neuroinflammation 3
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
Table 2 Continued
Antigens
BPF GMF WMF T2LV
r p FDR r p FDR r p FDR r p FDR
HSP60 aa 240ndash259 048 0028a 0058 052 0017a 0026a 000 0990 1000 2047 0030b 0056
HSP60 aa 421ndash440 010 0655 0969 039 0080 0133 2058 0006b 0108 2022 0334 0516
HSP60 aa 436ndash455 042 0061 0067 060 0004a 0015a 2031 0173 0804 2038 0094 0146
HSP70 aa 121ndash140 044 0046a 0067 036 0111 0197 024 0289 0854 2016 0496 0728
HSP70 aa 181ndash199 051 0017a 0040a 048 0027a 0026a 015 0525 1000 2020 0378 0516
HSP70 aa 210ndash229 006 0788 0969 027 0228 0303 2044 0045b 0206 2011 0640 1000
HSP70 aa 316ndash335 2028 0216 0413 2029 0209 0303 2004 0879 1000 044 0048a 0072
HSP70 aa 376ndash395 2042 0059 0067 2046 0035b 0027b 002 0942 1000 035 0121 0157
HSP70 aa 421ndash440 048 0030a 0058 065 0002a 0015a 2027 0229 0854 2034 0137 0209
HSP70 aa 61ndash80 2002 0941 0969 2029 0198 0303 056 0008a 0109 018 0441 0537
IL-12 2050 0021b 0040b 2045 0040b 0027b 2018 0429 1000 039 0083 0146
IL-3 2041 0063 0067 2051 0019b 0026b 013 0575 1000 052 0017a 0056
Insula 046 0038a 0067 033 0143 0303 034 0133 0804 2017 0473 0550
Intrinsic factor 036 0108 0194 044 0048a 0066 2009 0702 1000 000 0986 1000
Isoprostane F2a-I 2053 0014b 0040b 2055 0010b 0025b 2004 0854 1000 035 0115 0157
Lactosylceramide 2021 0368 0630 2041 0062 0112 039 0077 0788 060 0004a 0033a
LIF 2065 0002b 0025b 2059 0005b 0015b 2022 0336 0854 041 0064 0146
MMP2 042 0055 0067 061 0003a 0015a 2032 0153 0804 2039 0082 0146
MMP9 2055 0009b 0040b 2062 0003b 0015b 006 0804 1000 045 0038a 0057
MOG peptide aa 196ndash215 2005 0837 0969 2008 0736 0996 005 0836 1000 044 0045a 0057
Monosialogang GM2 2056 0008b 0040b 2048 0028b 0026b 2026 0260 0854 017 0460 0550
Myelin protein 2 peptide aa 31ndash50 035 0124 0194 048 0029a 0027a 2022 0337 0854 2033 0141 0209
Myelin protein 2 peptide aa 91ndash110 2009 0694 0969 014 0533 0996 2051 0019b 0122 2004 0878 1000
Neurofilament 160 kDa 032 0162 0305 039 0082 0133 2009 0688 1000 2049 0026b 0056
Neurofilament 200 kDa 043 0050 0067 045 0040a 0027a 003 0881 1000 2015 0516 0728
Neurofilament 68 kDa 000 0999 1000 2021 0359 0996 044 0044a 0206 045 0043a 0057
NOGO 016 0493 0928 2012 0617 0996 060 0004a 0108 021 0352 0516
Nonhydroxy fatty acid ceramide 2053 0014b 0040b 2059 0005b 0015b 005 0826 1000 044 0044a 0057
Occipital lobe 2045 0039b 0067 2043 0054 0072 2012 0599 1000 039 0083 0146
ORF26 protein 053 0013a 0040a 052 0015a 0026a 010 0653 1000 2030 0182 0247
Parietal lobe 049 0023a 0040a 058 0005a 0015a 2011 0641 1000 2025 0283 0516
PDGF-Ra 2007 0752 0969 017 0461 0996 2052 0016b 0122 2009 0687 1000
Pepstatin 2035 0116 0194 2046 0036b 0027b 016 0483 1000 025 0277 0516
Postcentral gyrus AD 044 0044a 0067 034 0132 0197 028 0214 0854 2012 0603 1000
Proteolipid protein peptide aa 125ndash141 2009 0685 0969 010 0678 0996 2041 0064 0788 2048 0029b 0056
Proteolipid protein peptide aa 150ndash163 044 0043a 0067 051 0019a 0026a 2006 0809 1000 2050 0020b 0056
Proteolipid protein peptide aa 161ndash180 022 0343 0610 031 0166 0303 2017 0475 1000 2044 0045b 0057
Proteolipid protein peptide aa 250ndash269 037 0103 0194 050 0020a 0026 2023 0321 0854 2037 0098 0146
RBP 025 0274 0610 029 0201 0303 2005 0844 1000 2057 0007b 0056
Ribonuclease 2002 0937 0969 2017 0472 0996 031 0175 0804 054 0011a 0056
S100b 033 0141 0258 011 0649 0996 052 0015a 0122 2022 0344 0516
Spectrin 048 0026a 0058 035 0122 0197 036 0105 0804 2001 0966 1000
Sulfatides 2047 0033b 0058 2049 0023b 0026b 2002 0922 1000 034 0133 0209
Tetrasialoganglioside-GQ1B 2043 0054 0067 2057 0007b 0025b 023 0319 0854 048 0027a 0056
Continued4 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
Biologicals (Littleton CO) Assay Designs (Ann Arbor MI) Pro-
Sci Inc (Poway CA) EMD Biosciences (San Diego CA) Cay-
man Chemical (Ann Arbor MI) HyTest (Turku Finland)
Meridian Life Science (Memphis TN) and Biodesign Interna-
tional (Saco ME) The antigens used in the construction of anti-
gen microarrays are listed in table e-1 at Neurologyorgnn
Antigen microarray production development and dataanalysis The antigens listed in table e-1 were spotted in replicatesof 6 on SuperEpoxy 2 slides (TeleChem Sunnyvale CA) using
an Arrayit NanoPrint 2 LM210 microarray printer (Arrayit Corpo-
ration Sunnyvale CA) and optimized spotting conditions as
described28303140 The microarrays were hybridized using an HS
4800Pro Hybridization Station (Tecan Maumlnnedorf Switzerland)
in which they were blocked with 1 bovine serum albumin for
1 hour and incubated for 2 hours at 37degC with the test serum at
a 1100 dilution in blocking buffer The arrays were then washed
and incubated for 45 minutes with a 1500 dilution of goat
anti-human immunoglobulin G (IgG) Cy3-conjugated and
goat anti-human IgM Cy5-conjugated detection antibodies (Jackson
ImmunoResearch Labs West Grove PA) The arrays were scanned
with a Tecan PowerScanner Repeated measurements indicate that
our antigen microarray technique is reproducible exhibiting
a coefficient of variation of 1336 1231
Background signal was subtracted and raw data were normal-
ized and analyzed using the GeneSpring software (Silicon Genetics
Redwood City CA) Antigen reactivity was defined by the mean
intensity of binding to the replicates of that antigen on the micro-
array and expressed as relative fluorescence units Scatter plots were
generated using linear regression models in the R statistical package
Pearson product-moment correlation coefficients were calculated
between the MRI measures (BPF GMF WMF T2LV) and the
weighted average of a group of statistically significant lipids The
weighted average of significant lipids was calculated using the for-
mula (SWiAiSWi) (SWi 5 1 i 5 1 2 3 ) where Wi is the
proportion of intensity of antigen Ai Ai is the observed intensity of
an antigen and SWi is the sum of the weights
RESULTS Serum IgG antibodies correlate with brain
MRI measures of disease severity To study the relation-ship between the peripheral immune response andMRI measures of disease severity we analyzed serum
antibody reactivity in those MS samples We analyzedIgG serum antibodies using a panel of antigensincluding CNS antigens heat shock proteins and lip-ids The association between the antibody reactivityagainst each antigen and 4 MRI measures of disease(T2LV BPF WMF and GMF) was investigatedusing Spearman correlation tests
We found significant associations between eachMRI measure of disease and different sets of IgG anti-body reactivity which are shown in table 2 Similarpatterns of antibody reactivity were linked to BPFand GMF consistent with the known dominant con-tribution of GM atrophy to whole brain atrophymeasures3741 Strikingly there was little overlapbetween the antibody reactivity associated withGMF and WMF suggesting that different immuno-pathologic processes contribute to tissue degenerationin these areas of the brain Similarly these profiles ofantibody reactivity were also different from thoseassociated with T2LV
Serum lipid-reactive IgGs are associated with increased
disease pathogenesis determined by MRI In evaluationof MRI measures and their link to disease BPFGMF and WMF decrease with disease progressionwhile T2LV increases with disease progressionAccordingly we analyzed the linkage between eachMRI measure and the significant antibody reactivitiesshown in table 2 We identified IgG antibody reac-tivities associated with increased tissue destruction asevidenced by decreased BPF values (figure 1) Strik-ingly we found that antibodies linked to diseasepathology as measured by BPF were enriched for reac-tivity against lipids Furthermore a selective increasein lipid-reactive antibodies linked to disease severitywas also observed when we analyzed all available MRImeasures (figure 2) Of note we did not detect an
Table 2 Continued
Antigens
BPF GMF WMF T2LV
r p FDR r p FDR r p FDR r p FDR
Thyrocalcitonin 2033 0143 0258 2008 0746 0996 2059 0005b 0108 004 0878 1000
TNPAL-galactocerebroside 007 0763 0969 2015 0506 0996 047 0030a 0142 005 0823 1000
Total cerebroside 2016 0498 0928 2041 0064 0112 050 0020a 0122 062 0003a 0033
Trisialogang GT1b 2034 0128 0194 2044 0043b 0066 015 0513 1000 030 0185 0247
Troponin I 2051 0020b 0040b 2053 0013b 0026b 2003 0900 1000 043 0050 0072
Ubiquitin 031 0166 0305 045 0039a 0027 2025 0282 0854 2029 0196 0247
VEGF 2050 0023b 0040b 2047 0031b 0027b 2013 0570 1000 040 0070 0146
Abbreviations BPF 5 whole brain parenchymal fraction FDR 5 false discovery ratendashadjusted significance value GMF 5 global cerebral gray matterfraction T2LV 5 cerebral T2 (fluid-attenuated inversion recovery) hyperintense lesion volume WMF 5 global cerebral white matter fractionDiscovery setap 005 (positive association)bp 005 (negative association)
Neurology Neuroimmunology amp Neuroinflammation 5
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
enrichment in lipid-reactive IgM in the group ofantibodies associated with increased diseasepathogenesis (not shown)
An index of serum IgG reactivity to lipids is associated
with MRI measures of disease severity To further inves-tigate the association between serum IgG reactivityto lipids and MRI measures of disease severity wecalculated an IgG anti-lipid index for each patientcorresponding to the information on each lipid-specific antibody listed in table 2 normalized by thestrength of its correlation with that MRI measureunder investigation The antibody reactivities tolipids used to calculate the lipid index are shown in
figure 3A With this approach we calculated one indexfor each of the MRI measures analyzed in this studyWe found a significant correlation between each IgGlipid antibody index and BPF GMF and T2LV(figure 3B) no significant correlation was found withWMF Moreover no significant correlations werefound between BPF WMF GMF and T2LVmeasures and anti-lipid antibody indexes based onIgM reactivity (not shown)
Finally we evaluated the performance of the lipidantibody indexes linked to BPF GMF and T2LV onan additional set of independent MS samples (valida-tion set table 1) We detected a similar trend to theone detected in the discovery set with regard to the
Figure 1 Antibody reactivity in serum is associated with decreased brain volume
Heatmap in which each column represents the mean immunoglobulin G antibody reactivity in a serum sample from a patient with multiple sclerosis sortedaccording to the whole brain parenchymal fraction (BPF) (indicated at the topmdasha lower BPF indicates more brain atrophy) and each row represents theantibody reactivity to an antigen according to the colorimetric scale shown The antibody reactivities included in this heatmap are listed in table 2
6 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
Figure 2 Association of serum IgG reactivity with MRI measures of disease severity
(A) Heatmap in which each column represents an MRI measure of either atrophy (BPF GMF WMF) or lesions (T2LV) andeach row represents the correlation to IgG serum antibody reactivity according to a colorimetric scale (B) Frequency oflipid-reactive antibodies linked to higher or lower MRI disease severity BPF 5 whole brain parenchymal fraction GMF 5
global cerebral gray matter fraction IgG 5 immunoglobulin G T2LV 5 cerebral T2 (fluid-attenuated inversion recovery)hyperintense lesion volume WMF 5 global cerebral white matter fraction
Neurology Neuroimmunology amp Neuroinflammation 7
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
correlation between BPF and GMF and their respec-tive lipid indexes and we validated the correlationbetween T2LV and its lipid index (figure 3C)
DISCUSSION Previous studies have investigated theassociation of MRI with immune activity in MS13ndash19
These studies have included cellular immune measures
Figure 3 Correlation of lipid indexes to MRI measures of disease severity in multiple sclerosis
(A) Heatmap inwhich each column represents anMRImeasure and each row represents the correlation to immunoglobulin G serumantibody reactivity to lipids accord-ing to a colorimetric scale (B) Scatter plots depicting the correlation between each lipid index andMRImeasures (BPF GMF and T2LV) in the discovery set (C) Scatterplots depicting the correlation between each lipid index andMRImeasures (BPF GMF and T2LV) in the validation set BPF5whole brain parenchymal fraction GMF5
cerebral gray matter fraction (see methods section for details) T2LV 5 cerebral T2 hyperintense lesion volume WMF 5 global cerebral white matter fraction
8 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
as well as oligoclonal bands or neurofilament-specificantibodies in CSF42ndash44 In the present study weanalyzed the association between serum antibodyprofiles detected with antigen microarrays and MRImeasures of disease including cerebral atrophy andT2 hyperintense lesions We found that specificantibody patterns are associated with different aspectsof MRI-defined disease pathology Our major findingwas that increased reactivity to lipids is associated withdifferent aspects of brain MRI measures of diseaseseverity Of note the specific set of lipids associatedwith atrophy differed from those associated withlesions Taken together these data suggest that anti-lipid antibodies in serum are related to MRI measuresof disease Our findings highlight the important role oflipid and lipid-specific immunity in the pathogenesis ofMS
Although a potential limitation of these studies isthe relatively small number of samples analyzed wehave validated our findings in an independent valida-tion cohort strengthening their significance In addi-tion further studies in large cohorts from patientsaffected by non-MS neurodegenerative diseases andhealthy controls may indicate whether the antibodyreactivities and lipid indexes identified in these stud-ies are exclusive to MS or are associated with addi-tional biological processes Indeed some of theseantibody reactivities contributing to the immune sig-natures described in this work have been found innewborns45
Different sets of antibody reactivities were associ-ated with GMF WMF BPF and T2LV The differ-ent antibody link between lesions and atrophy is inkeeping with the long-held view that brain atrophyin patients with MS is multifactorial and is onlyweakly related to lesions41 This has been underscoredin recent studies showing the complementary infor-mation obtained by combining lesion and atrophymeasures4647
Among the cerebral atrophy measures there was astriking overlap between those antibodies associatedwith GMF and BPF which separated them fromWMF results These findings are in keeping with pre-vious observations that whole brain atrophy is domi-nated by GM loss3741 In the early stages of MS thisGM atrophy selectively involves the deep GM nuclei34
Thus the similarities observed in the autoantibodieslinked to GMF and BPF might reflect the dominantcontribution and colinearity of GM atrophy to totalbrain atrophy The set of antibodies linked to GMFmay be the most relevant given that MRI studies haveshown that GM volume is more closely linked to phys-ical disability37 and cognitive impairment38 than areWM volume or lesion measures
Using antigen microarrays antibody reactivity tolipids has been detected in the CSF and serum of
patients with MS at different stages of the diseaseand such antibodies in the CSF have been linkedto disease progression and MRI involvement44
Although several mechanisms are thought to drivebrain inflammation and atrophy in MS46 the connec-tion between these mechanisms and the antibodiesdetected with our antigen microarrays is yet unknownOne possibility is that the serum autoantibodies iden-tified in our studies are not pathogenic and reflect theresult of immunization against self-antigens releasedfrom the CNS during the course of the disease Indeedneurofilament is released from damaged axons duringthe course of MS and both neurofilament lightchains48 and antibodies against it42 have been linkedto MRI measures of disease Moreover heat shockproteins are upregulated in different cell types duringinflammation and have been shown to have an impor-tant role as immunomodulators when released to theextracellular medium4950 Thus it is possible that heatshock proteinndashreactive antibodies reflect changes in theproduction and release of these immunomodulatorsduring the course of disease pathogenesis Similarlybioactive lipids and their products are released fromdamaged myelin and lipid-specific antibodies havebeen detected in patients with MS27293051 Lipids havean important role in the immune response both asbioactive molecules with immunomodulatory proper-ties and also as targets of the adaptive immuneresponse52 Moreover lipids have significant effectson the murine model of MS experimental autoim-mune encephalomyelitis2729305153 Indeed we recentlyfound that the glycolipid lactosylceramide activates abroad set of biological processes in astrocytes promot-ing neurodegeneration and inflammation53 Thus thelipid-reactive antibodies detected in this work mayreflect the release of myelin lipids in the context ofMS pathogenesis andor may directly contribute toimmune-mediated damage in CNS tissue Of notelipid-reactive antibodies are highly cross-reactive52 Inaddition the lipid-reactive antibodies detected in thiswork as associated with MRI measures of disease activ-ity were of the IgG class These are important points toconsider when evaluating a potential role of lipid-reactive antibodies inMS pathogenesis Further studiesare warranted to investigate whether bioactive lipidsoffer new therapeutic targets in MS
We have found that unique patterns of immunereactivity determined with antibody arrays are associ-ated with specific MRI measures of disease severityThese patterns agree with the interpretation that dif-ferent pathogenic mechanisms drive diverse diseaseprocesses reflected by these MRI measures These pat-terns also suggest a predominant role for lipids andlipid-specific immunity in MS pathology Furtherstudies are warranted to determine whether the earlyappearance of anti-lipid antibodies predicts the
Neurology Neuroimmunology amp Neuroinflammation 9
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
subsequent development of clinical and MRI-defineddisease worsening
AUTHOR CONTRIBUTIONSRohit Bakshi draftingrevising the manuscript study concept or design
analysis or interpretation of data acquisition of data study supervision
obtaining funding Ada Yeste study concept or design acquisition of
data Bonny Patel analysis or interpretation of data statistical analysis
Shahamat Tauhid analysis or interpretation of data Subhash Tummala
analysis or interpretation of data Roya Rahbari analysis or interpretation
of data statistical analysis Renxin Chu analysis or interpretation of data
Keren Regev analysis or interpretation of data acquisition of data
Pia Kivisaumlkk draftingrevising the manuscript study concept or design
contribution of vital reagentstoolspatients acquisition of data study
supervision Howard L Weiner study concept or design obtaining fund-
ing Francisco J Quintana draftingrevising the manuscript study con-
cept or design contribution of vital reagentstoolspatients study
supervision obtaining funding
ACKNOWLEDGMENTThe authors thank Dr Mohit Neema Dr Antonella Ceccarelli and
Dr Ashish Arora for valuable assistance at the early stages of this project
STUDY FUNDINGThis work was supported in part by grants to FJQ and HLW from
EMD Serono RG4111A1 and JF2161-A-5 from the National Multiple
Sclerosis Society PA0069 from the International Progressive MS Alli-
ance and by grants from the NIH (5R01NS055083-04) and NMSS
(RG3798A2) to RB
DISCLOSUREFJ Quintana serves on the editorial board for Systems Biomedicine
Inmunologia American Journal of Clinical and Experimental Immunology
is an associate editor for Immunology (UK) is an advisory board member
for Seminars in Immunopathology received research support from Harvard
Medical School BADERC NMSS A Yeste and B Patel report no
disclosures S Tauhid is managing editor for Journal of Neuroimaging
S Tummala R Rahbari R Chu and K Regev report no disclosures
P Kivisaumlkk received research support from EMD Serono R Bakshi is
editor-in-chief for Journal of Neuroimaging received consulting fees from
AbbVie Alkermes Biogen Novartis Questcor received research support
from Biogen EMD-Serono Novartis Sanofi-Genzyme Teva his spouse
holds stock in Biogen Inc H Weiner served on the advisory board
for The Guthy-Jackson Charitable Foundation Teva Pharmaceuticals
Industries Ltd Biogen Idec Novartis Sanofi-Aventis has consulted for
Therapix Bioven Novartis Serono Teva Sanofi received research support
from National Multiple Sclerosis Society Go to Neurologyorgnn for full
disclosure forms
Received July 27 2015 Accepted in final form December 8 2015
REFERENCES1 Weiner HL The challenge of multiple sclerosis how do
we cure a chronic heterogeneous disease Ann Neurol
200965239ndash248
2 Bakshi R Thompson AJ Rocca MA et al MRI in mul-
tiple sclerosis current status and future prospects Lancet
Neurol 20087615ndash625
3 Ceccarelli A Bakshi R Neema M MRI in multiple scle-
rosis a review of the current literature Curr Opin Neurol
201225402ndash409
4 Filippi M Rocca MA Barkhof F et al Association
between pathological and MRI findings in multiple scle-
rosis Lancet Neurol 201211349ndash360
5 Geurts JJ Calabrese M Fisher E Rudick RA Measure-
ment and clinical effect of grey matter pathology in mul-
tiple sclerosis Lancet Neurol 2012111082ndash1092
6 Klaver R De Vries HE Schenk GJ Geurts JJ Grey mat-
ter damage in multiple sclerosis a pathology perspective
Prion 2013766ndash75
7 Khoury S Bakshi R Cerebral pseudoatrophy or real atro-
phy after therapy in multiple sclerosis Ann Neurol 2010
68778ndash779
8 Hauser SL Chan JR Oksenberg JR Multiple sclerosis
prospects and promise Ann Neurol 201374317ndash327
9 McFarland HF Martin R Multiple sclerosis a complicated
picture of autoimmunity Nat Immunol 20078913ndash919
10 Nylander A Hafler DA Multiple sclerosis J Clin Invest
20121221180ndash1188
11 Sospedra M Martin R Immunology of multiple sclerosis
Ann Rev Immunol 200523683ndash747
12 Steinman L Immunology of relapse and remission in mul-
tiple sclerosis Ann Rev Immunol 201432257ndash281
13 Khoury SJ Guttmann CR Orav EJ Kikinis R Jolesz FA
Weiner HL Changes in activated T cells in the blood
correlate with disease activity in multiple sclerosis Arch
Neurol 2000571183ndash1189
14 Khoury SJ Orav EJ Guttmann CR Kikinis R Jolesz FA
Weiner HL Changes in serum levels of ICAM and TNF-
R correlate with disease activity in multiple sclerosis Neu-
rology 199953758ndash764
15 Laplaud DA Berthelot L Miqueu P et al Serial blood T
cell repertoire alterations in multiple sclerosis patients cor-
relation with clinical and MRI parameters J Neuroimmunol
2006177151ndash160
16 Makhlouf K Weiner HL Khoury SJ Increased percentage
of IL-121 monocytes in the blood correlates with the
presence of active MRI lesions in MS J Neuroimmunol
2001119145ndash149
17 Prat A Biernacki K Saroli T et al Kinin B1 receptor expres-
sion on multiple sclerosis mononuclear cells correlation with
magnetic resonance imaging T2-weighted lesion volume and
clinical disability Arch Neurol 200562795ndash800
18 Rinaldi L Gallo P Calabrese M et al Longitudinal anal-
ysis of immune cell phenotypes in early stage multiple
sclerosis distinctive patterns characterize MRI-active pa-
tients Brain 20061291993ndash2007
19 Tortorella P Lagana MM Saresella M et al Determinants
of disability in multiple sclerosis an immunological and
MRI study Biomed Res Int 20142014875768
20 Robinson WH DiGennaro C Hueber W et al Autoan-
tigen microarrays for multiplex characterization of autoan-
tibody responses Nat Med 20028295ndash301
21 Yeste A Quintana FJ Antigen microarrays for the study of
autoimmune diseases Clin Chem 2013591036ndash1044
22 Li QZ Xie C Wu T et al Identification of autoantibody
clusters that best predict lupus disease activity using glomer-
ular proteome arrays J Clin Invest 20051153428ndash3439
23 Hueber W Kidd BA Tomooka BH et al Antigen micro-
array profiling of autoantibodies in rheumatoid arthritis
Arthritis Rheum 2005522645ndash2655
24 Goldschmidt Y Sharon E Quintana FJ Cohen IR
Brandt A Adaptive methods for classification of biological
microarray data from multiple experiments Report no
MCS03-07 Rehovot Israel The Arthur and Rochelle Bel-
fer Institute of Mathematics and Computer Science 2003
25 Quintana FJ Getz G Hed G Domany E Cohen IR
Cluster analysis of human autoantibody reactivities in
health and in type 1 diabetes mellitus a bio-informatic
approach to immune complexity J Autoimmun 2003
2165ndash75
10 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
26 Lalive PH Menge T Barman I Cree BA Genain CP
Identification of new serum autoantibodies in neuromye-
litis optica using protein microarrays Neurology 200667
176ndash177
27 Kanter JL Narayana S Ho PP et al Lipid microarrays
identify key mediators of autoimmune brain inflamma-
tion Nat Med 200612138ndash143
28 Quintana FJ Farez MF Izquierdo G Lucas M Cohen IR
Weiner HL Antigen microarrays identify CNS-produced
autoantibodies in RRMS Neurology 201278532ndash539
29 Farez MF Quintana FJ Gandhi R Izquierdo G Lucas M
Weiner HL Toll-like receptor 2 and poly(ADP-ribose)
polymerase 1 promote central nervous system neuroin-
flammation in progressive EAE Nat Immunol 200910
958ndash964
30 Quintana FJ Farez MF Viglietta V et al Antigen micro-
arrays identify unique serum autoantibody signatures in
clinical and pathologic subtypes of multiple sclerosis Proc
Natl Acad Sci USA 200810518889ndash18894
31 Quintana FJ Patel B Yeste A et al Epitope spreading as
an early pathogenic event in pediatric multiple sclerosis
Neurology 2014832219ndash2226
32 Gauthier SA Glanz BI Mandel M Weiner HL A model
for the comprehensive investigation of a chronic autoim-
mune disease the multiple sclerosis CLIMB study Auto-
immun Rev 20065532ndash536
33 Polman CH Reingold SC Edan G et al Diagnostic criteria
for multiple sclerosis 2005 revisions to the ldquoMcDonald
Criteriardquo Ann Neurol 200558840ndash846
34 DellrsquoOglio E Ceccarelli A Glanz BI et al Quantification
of global cerebral atrophy in multiple sclerosis from 3T
MRI using SPM the role of misclassification errors
J Neuroimaging 201525191ndash199
35 Ceccarelli A Jackson JS Tauhid S et al The impact of
lesion in-painting and registration methods on voxel-based
morphometry in detecting regional cerebral gray matter
atrophy in multiple sclerosis AJNR Am J Neuroradiol
2012331579ndash1585
36 Cohen AB Neema M Arora A et al The relationships
among MRI-defined spinal cord involvement brain involve-
ment and disability in multiple sclerosis J Neuroimaging
201222122ndash128
37 Sanfilipo MP Benedict RH Sharma J Weinstock-
Guttman B Bakshi R The relationship between whole
brain volume and disability in multiple sclerosis a com-
parison of normalized gray vs white matter with misclas-
sification correction Neuroimage 2005261068ndash1077
38 Sanfilipo MP Benedict RH Weinstock-Guttman B
Bakshi R Gray and white matter brain atrophy and neuro-
psychological impairment in multiple sclerosis Neurology
200666685ndash692
39 Stankiewicz JM Glanz BI Healy BC et al Brain MRI
lesion load at 15T and 3T versus clinical status in multi-
ple sclerosis J Neuroimaging 201121e50ndashe56
40 Quintana FJ Hagedorn PH Elizur G Merbl Y
Domany E Cohen IR Functional immunomics micro-
array analysis of IgG autoantibody repertoires predicts the
future response of mice to induced diabetes Proc Natl
Acad Sci USA 2004101(suppl 2)14615ndash14621
41 Bermel RA Bakshi R The measurement and clinical
relevance of brain atrophy in multiple sclerosis Lancet
Neurol 20065158ndash170
42 Eikelenboom MJ Petzold A Lazeron RH et al Multiple
sclerosis neurofilament light chain antibodies are corre-
lated to cerebral atrophy Neurology 200360219ndash223
43 Magraner MJ Bosca I Simo-Castello M et al Brain atro-
phy and lesion load are related to CSF lipid-specific IgM
oligoclonal bands in clinically isolated syndromes Neuro-
radiol 2012545ndash12
44 Villar LM Sadaba MC Roldan E et al Intrathecal syn-
thesis of oligoclonal IgM against myelin lipids predicts an
aggressive disease course in MS J Clin Invest 2005115
187ndash194
45 Merbl Y Zucker-Toledano M Quintana FJ Cohen IR
Newborn humans manifest autoantibodies to defined self
molecules detected by antigen microarray informatics
J Clin Invest 2007117712ndash718
46 Sormani MP Arnold DL De Stefano N Treatment effect
on brain atrophy correlates with treatment effect on disa-
bility in multiple sclerosis Ann Neurol 20147543ndash49
47 Tauhid S Neema M Healy BC Weiner HL Bakshi R
MRI phenotypes based on cerebral lesions and atrophy in
patients with multiple sclerosis J Neurol Sci 2014346
250ndash254
48 Gnanapavan S Grant D Morant S et al Biomarker
report from the phase II lamotrigine trial in secondary
progressive MS neurofilament as a surrogate of disease
progression PLoS One 20138e70019
49 Quintana FJ Cohen IR Heat shock proteins as endoge-
nous adjuvants in sterile and septic inflammation
J Immunol 20051752777ndash2782
50 Quintana FJ Cohen IR The HSP60 immune system net-
work Trends Immunol 20113289ndash95
51 Ho PP Kanter JL Johnson AM et al Identification of
naturally occurring fatty acids of the myelin sheath that
resolve neuroinflammation Sci Transl Med 20124137ra73
52 Quintana FJ Yeste A Weiner HL Covacu R Lipids and
lipid-reactive antibodies as biomarkers for multiple sclero-
sis J Neuroimmunol 201224853ndash57
53 Mayo L Trauger SA Blain M et al Regulation of astro-
cyte activation by glycolipids drives chronic CNS inflam-
mation Nat Med 2014201147ndash1156
Neurology Neuroimmunology amp Neuroinflammation 11
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
1
Table e-1 Antigens used
Group Antigen Source
27 kDa Heat Shock Protein Stressgen
HSP 32 kDa Heat Shock Protein Stressgen
40 kDa Heat Shock Protein Stressgen
47 kDa Heat Shock Protein Stressgen
60 kDa Heat Shock Protein Stressgen
60 kDa Heat Shock Protein peptide aa 106ndash125 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 121ndash140 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 136ndash155 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 151ndash170 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 16ndash35 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 166ndash185 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
2
60 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 556ndash573 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
65 kDa Heat Shock Protein M tuberculosis Stressgen
70 kDa Heat Shock Protein Stressgen
70 kDa Heat Shock Protein peptide aa 106ndash125 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 121ndash140 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 136ndash155 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 151ndash170 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 16ndash35 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 166ndash185 Biopolymers Facility Harvard Medical School
3
70 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
4
70 kDa Heat Shock Protein peptide aa 556ndash575 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 571ndash590 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 586ndash605 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 601ndash620 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 616ndash635 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 631ndash640 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
71 kDa Heat Shock Protein M tuberculosis Biopolymers Facility Harvard Medical School
90 kDa Heat Shock Protein Stressgen
GroEL Stressgen
23-cyclic nucleotide 3-phosphodiesterase
peptide aa106-125 Biopolymers Facility Harvard Medical School
CNS
23-cyclic nucleotide 3-phosphodiesterase
peptide aa1-20 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa121-140 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa136-155 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa151-170 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa16-35 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa166-185 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase Biopolymers Facility Harvard Medical School
5
peptide aa181-200
23-cyclic nucleotide 3-phosphodiesterase
peptide aa196-215 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa211-230 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa226-245 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa241-260 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa256-275 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa271-290 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa286-305 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa301-320 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa31-50 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa316-335 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa331-350 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa346-365 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa361-380 Biopolymers Facility Harvard Medical School
6
23-cyclic nucleotide 3-phosphodiesterase
peptide aa376-395 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa391-410 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa406-421 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa46-65 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa61-80 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa76-95 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa91-110 Biopolymers Facility Harvard Medical School
Acetyl Cholinesterase Sigma Aldrich
ADAM-10 Sigma Aldrich
alpha-Cristallin Stressgen
beta-Cristallin Sigma Aldrich
bovine Myelin Basic Protein Sigma Aldrich
Brain Extract I Sigma Aldrich
Brain Extract II Sigma Aldrich
Brain Extract III Sigma Aldrich
Glial Filament Acidic Protein Research Diagnostic
guinea pig Myelin Basic Protein Sigma Aldrich
human Myelin Basic Protein Sigma Aldrich
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 106-125 Biopolymers Facility Harvard Medical School
7
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 121-140 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 136-155 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 151-170 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 166-185 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 181-200 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 91-110 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
106-125 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa 1- Biopolymers Facility Harvard Medical School
8
20
Myelinoligodendrocyte glycoprotein peptide aa
121-140 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
136-155 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
151-170 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
16-35 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
166-185 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
181-200 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
196-215 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
211-230 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
226-247 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
31-50 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
35-55 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
46-65 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
61-80 Biopolymers Facility Harvard Medical School
9
Myelinoligodendrocyte glycoprotein peptide aa
76-95 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
91-110 Biopolymers Facility Harvard Medical School
murine Myelin Basic Protein Sigma Aldrich
Myelin Associated Glycoprotein Sigma Aldrich
Myelin Basic Protein peptide aa 104-123 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 11-30 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 113-132 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 121-138 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 124-142 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 138-147 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 141-161 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 143-168 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 155-178 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 26-35 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 41-60 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 51-70 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 71-92 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 84-94 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 89-101 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 173-186 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 93-112 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 106-125 Biopolymers Facility Harvard Medical School
10
Myelin Protein 2 peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 121-132 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 91-110 Biopolymers Facility Harvard Medical School
Neurofilament 160kd Chemicon
Neurofilament 200kd Chemicon
Neurofilament 68kd Chemicon
Neuronal Enolase Calbiochem
Nicastrin GeneTex
NMDA receptor Novus Biologicals
NOGO Sigma Aldrich
Olygodendrocyte-Specific Protein peptide aa 106ndash
125 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 121ndash
140 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 136ndash
155 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 151ndash
170 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 16ndash
35 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 166ndash Biopolymers Facility Harvard Medical School
11
185
Olygodendrocyte-Specific Protein peptide aa 181ndash
199 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 195ndash
217 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 31ndash
50 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 46ndash
65 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 61ndash
80 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 76ndash
95 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 91ndash
110 Biopolymers Facility Harvard Medical School
Proteolipid Protein Abnova
Proteolipid Protein peptide aa 100-119 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 10-29 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 110-129 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 1-19 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 125-141 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-150 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-154 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 150-163 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 151-173 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 158-166 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 161-180 Biopolymers Facility Harvard Medical School
12
Proteolipid Protein peptide aa 178-191 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 180-199 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 190-209 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 20-39 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 205-220 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 215-232 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-239 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-249 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 248-259 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 250-269 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 265-277 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 35-50 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 40-59 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 50-69 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 65-84 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 80-99 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 91-110 Biopolymers Facility Harvard Medical School
Retinol Binding Protein Sigma Aldrich
S100beta protein Assay Designs
Super Oxide Dismutase Sigma Aldrich
Synuclein beta Sigma Aldrich
Synuclein gamma Sigma Aldrich
Amydgala ProSci Inc
Tissue Amydgala AD ProSci Inc
Brain lysate ProSci Inc
Brain Tissue Membrane ProSci Inc
Cerebellar pedunculus ProSci Inc
13
Cerebral meninges ProSci Inc
Corpus Callosum ProSci Inc
Corpus Callosum AD ProSci Inc
Diencephalon ProSci Inc
Fetal brain ProSci Inc
Frontal lobe ProSci Inc
Frontal lobe AD ProSci Inc
Hippocampus ProSci Inc
Hippocampus AD ProSci Inc
Insula ProSci Inc
Occipital lobe ProSci Inc
Occipital lobe AD ProSci Inc
Olfactory region ProSci Inc
Optic Nerve ProSci Inc
Parietal lobe ProSci Inc
Parietal lobe AD ProSci Inc
Pons ProSci Inc
Pons AD ProSci Inc
Postcentral gyrus ProSci Inc
Postcentral gyrus AD ProSci Inc
Precentral gyrus ProSci Inc
Precentral gyrus AD ProSci Inc
Spinal cord ProSci Inc
Temporal lobe ProSci Inc
Temporal lobe AD ProSci Inc
Thalamus ProSci Inc
Thalamus AD ProSci Inc
14
Amyloid beta Sigma Aldrich
AD
related Amyloid beta 10-20 Sigma Aldrich
Amyloid beta 1-12 Sigma Aldrich
Amyloid beta 12-28 Sigma Aldrich
Amyloid beta 1-23 Sigma Aldrich
Amyloid beta 1-38 Sigma Aldrich
Amyloid beta 17-40 Sigma Aldrich
Amyloid beta 25-35 Sigma Aldrich
Amyloid beta 34-42 Sigma Aldrich
Amyloid bri protein precursor 227 Sigma Aldrich
Amyloid DAN Protein Fragment 1-34 Sigma Aldrich
Amyloid Precursor Protein Sigma Aldrich
Amyloid protein no AB component Sigma Aldrich
Secreted amyloid precursor protein (SAP) beta Sigma Aldrich
Tau isoform variant 0N3R Sigma Aldrich
Tau isoform variant 1N3R Sigma Aldrich
Tau isoform variant 0N4R Sigma Aldrich
Tau isoform variant 2N3R Sigma Aldrich
Tau phospho Ser412 Sigma Aldrich
Tau phospho Ser441 Sigma Aldrich
Tau phospho Thr181 Sigma Aldrich
Tau Protein human EMD Biosciences
(plusmn)9-HODE Cayman Chemical
Lipids
1 Palmitoyl-2-(5oxo-Valeroyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
15a-hydroxycholestene Avanti Polar Lipids
15
15-ketocholestane Avanti Polar Lipids
15-ketocholestene Avanti Polar Lipids
1-Palmitoil-2-(9oxo-Nonanoyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Azelaoyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Glutaroyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
5 α-cholestane-3 β15 α-diol Avanti Polar Lipids
9(S)-HODE Cayman Chemical
Asialoganglioside-GM1 Sigma Aldrich
Asialoganglioside-GM2 Sigma Aldrich
Brain ceramides Avanti Polar Lipids
Brain D-erythrosphingosine Avanti Polar Lipids
Brain lysophosphatidylethanolamine Avanti Polar Lipids
Brain L-α-lysophosphatidylserine Avanti Polar Lipids
Brain L-α-phosphatidylcholine Avanti Polar Lipids
Brain L-α-phosphatidyl-ethanolamine Avanti Polar Lipids
Brain L-α-phosphatidylserine Avanti Polar Lipids
Brain polar lipid extract Avanti Polar Lipids
Brain sphingomyelin Avanti Polar Lipids
Brain sulfatide Avanti Polar Lipids
Brain total lipid extract Avanti Polar Lipids
Cardiolipin Sigma Aldrich
Ceramide Sigma Aldrich
Ceramide 1-phosphate Sigma Aldrich
Cholesterol Sigma Aldrich
16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-

Table 2 Continued
Antigens
BPF GMF WMF T2LV
r p FDR r p FDR r p FDR r p FDR
HSP60 aa 240ndash259 048 0028a 0058 052 0017a 0026a 000 0990 1000 2047 0030b 0056
HSP60 aa 421ndash440 010 0655 0969 039 0080 0133 2058 0006b 0108 2022 0334 0516
HSP60 aa 436ndash455 042 0061 0067 060 0004a 0015a 2031 0173 0804 2038 0094 0146
HSP70 aa 121ndash140 044 0046a 0067 036 0111 0197 024 0289 0854 2016 0496 0728
HSP70 aa 181ndash199 051 0017a 0040a 048 0027a 0026a 015 0525 1000 2020 0378 0516
HSP70 aa 210ndash229 006 0788 0969 027 0228 0303 2044 0045b 0206 2011 0640 1000
HSP70 aa 316ndash335 2028 0216 0413 2029 0209 0303 2004 0879 1000 044 0048a 0072
HSP70 aa 376ndash395 2042 0059 0067 2046 0035b 0027b 002 0942 1000 035 0121 0157
HSP70 aa 421ndash440 048 0030a 0058 065 0002a 0015a 2027 0229 0854 2034 0137 0209
HSP70 aa 61ndash80 2002 0941 0969 2029 0198 0303 056 0008a 0109 018 0441 0537
IL-12 2050 0021b 0040b 2045 0040b 0027b 2018 0429 1000 039 0083 0146
IL-3 2041 0063 0067 2051 0019b 0026b 013 0575 1000 052 0017a 0056
Insula 046 0038a 0067 033 0143 0303 034 0133 0804 2017 0473 0550
Intrinsic factor 036 0108 0194 044 0048a 0066 2009 0702 1000 000 0986 1000
Isoprostane F2a-I 2053 0014b 0040b 2055 0010b 0025b 2004 0854 1000 035 0115 0157
Lactosylceramide 2021 0368 0630 2041 0062 0112 039 0077 0788 060 0004a 0033a
LIF 2065 0002b 0025b 2059 0005b 0015b 2022 0336 0854 041 0064 0146
MMP2 042 0055 0067 061 0003a 0015a 2032 0153 0804 2039 0082 0146
MMP9 2055 0009b 0040b 2062 0003b 0015b 006 0804 1000 045 0038a 0057
MOG peptide aa 196ndash215 2005 0837 0969 2008 0736 0996 005 0836 1000 044 0045a 0057
Monosialogang GM2 2056 0008b 0040b 2048 0028b 0026b 2026 0260 0854 017 0460 0550
Myelin protein 2 peptide aa 31ndash50 035 0124 0194 048 0029a 0027a 2022 0337 0854 2033 0141 0209
Myelin protein 2 peptide aa 91ndash110 2009 0694 0969 014 0533 0996 2051 0019b 0122 2004 0878 1000
Neurofilament 160 kDa 032 0162 0305 039 0082 0133 2009 0688 1000 2049 0026b 0056
Neurofilament 200 kDa 043 0050 0067 045 0040a 0027a 003 0881 1000 2015 0516 0728
Neurofilament 68 kDa 000 0999 1000 2021 0359 0996 044 0044a 0206 045 0043a 0057
NOGO 016 0493 0928 2012 0617 0996 060 0004a 0108 021 0352 0516
Nonhydroxy fatty acid ceramide 2053 0014b 0040b 2059 0005b 0015b 005 0826 1000 044 0044a 0057
Occipital lobe 2045 0039b 0067 2043 0054 0072 2012 0599 1000 039 0083 0146
ORF26 protein 053 0013a 0040a 052 0015a 0026a 010 0653 1000 2030 0182 0247
Parietal lobe 049 0023a 0040a 058 0005a 0015a 2011 0641 1000 2025 0283 0516
PDGF-Ra 2007 0752 0969 017 0461 0996 2052 0016b 0122 2009 0687 1000
Pepstatin 2035 0116 0194 2046 0036b 0027b 016 0483 1000 025 0277 0516
Postcentral gyrus AD 044 0044a 0067 034 0132 0197 028 0214 0854 2012 0603 1000
Proteolipid protein peptide aa 125ndash141 2009 0685 0969 010 0678 0996 2041 0064 0788 2048 0029b 0056
Proteolipid protein peptide aa 150ndash163 044 0043a 0067 051 0019a 0026a 2006 0809 1000 2050 0020b 0056
Proteolipid protein peptide aa 161ndash180 022 0343 0610 031 0166 0303 2017 0475 1000 2044 0045b 0057
Proteolipid protein peptide aa 250ndash269 037 0103 0194 050 0020a 0026 2023 0321 0854 2037 0098 0146
RBP 025 0274 0610 029 0201 0303 2005 0844 1000 2057 0007b 0056
Ribonuclease 2002 0937 0969 2017 0472 0996 031 0175 0804 054 0011a 0056
S100b 033 0141 0258 011 0649 0996 052 0015a 0122 2022 0344 0516
Spectrin 048 0026a 0058 035 0122 0197 036 0105 0804 2001 0966 1000
Sulfatides 2047 0033b 0058 2049 0023b 0026b 2002 0922 1000 034 0133 0209
Tetrasialoganglioside-GQ1B 2043 0054 0067 2057 0007b 0025b 023 0319 0854 048 0027a 0056
Continued4 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
Biologicals (Littleton CO) Assay Designs (Ann Arbor MI) Pro-
Sci Inc (Poway CA) EMD Biosciences (San Diego CA) Cay-
man Chemical (Ann Arbor MI) HyTest (Turku Finland)
Meridian Life Science (Memphis TN) and Biodesign Interna-
tional (Saco ME) The antigens used in the construction of anti-
gen microarrays are listed in table e-1 at Neurologyorgnn
Antigen microarray production development and dataanalysis The antigens listed in table e-1 were spotted in replicatesof 6 on SuperEpoxy 2 slides (TeleChem Sunnyvale CA) using
an Arrayit NanoPrint 2 LM210 microarray printer (Arrayit Corpo-
ration Sunnyvale CA) and optimized spotting conditions as
described28303140 The microarrays were hybridized using an HS
4800Pro Hybridization Station (Tecan Maumlnnedorf Switzerland)
in which they were blocked with 1 bovine serum albumin for
1 hour and incubated for 2 hours at 37degC with the test serum at
a 1100 dilution in blocking buffer The arrays were then washed
and incubated for 45 minutes with a 1500 dilution of goat
anti-human immunoglobulin G (IgG) Cy3-conjugated and
goat anti-human IgM Cy5-conjugated detection antibodies (Jackson
ImmunoResearch Labs West Grove PA) The arrays were scanned
with a Tecan PowerScanner Repeated measurements indicate that
our antigen microarray technique is reproducible exhibiting
a coefficient of variation of 1336 1231
Background signal was subtracted and raw data were normal-
ized and analyzed using the GeneSpring software (Silicon Genetics
Redwood City CA) Antigen reactivity was defined by the mean
intensity of binding to the replicates of that antigen on the micro-
array and expressed as relative fluorescence units Scatter plots were
generated using linear regression models in the R statistical package
Pearson product-moment correlation coefficients were calculated
between the MRI measures (BPF GMF WMF T2LV) and the
weighted average of a group of statistically significant lipids The
weighted average of significant lipids was calculated using the for-
mula (SWiAiSWi) (SWi 5 1 i 5 1 2 3 ) where Wi is the
proportion of intensity of antigen Ai Ai is the observed intensity of
an antigen and SWi is the sum of the weights
RESULTS Serum IgG antibodies correlate with brain
MRI measures of disease severity To study the relation-ship between the peripheral immune response andMRI measures of disease severity we analyzed serum
antibody reactivity in those MS samples We analyzedIgG serum antibodies using a panel of antigensincluding CNS antigens heat shock proteins and lip-ids The association between the antibody reactivityagainst each antigen and 4 MRI measures of disease(T2LV BPF WMF and GMF) was investigatedusing Spearman correlation tests
We found significant associations between eachMRI measure of disease and different sets of IgG anti-body reactivity which are shown in table 2 Similarpatterns of antibody reactivity were linked to BPFand GMF consistent with the known dominant con-tribution of GM atrophy to whole brain atrophymeasures3741 Strikingly there was little overlapbetween the antibody reactivity associated withGMF and WMF suggesting that different immuno-pathologic processes contribute to tissue degenerationin these areas of the brain Similarly these profiles ofantibody reactivity were also different from thoseassociated with T2LV
Serum lipid-reactive IgGs are associated with increased
disease pathogenesis determined by MRI In evaluationof MRI measures and their link to disease BPFGMF and WMF decrease with disease progressionwhile T2LV increases with disease progressionAccordingly we analyzed the linkage between eachMRI measure and the significant antibody reactivitiesshown in table 2 We identified IgG antibody reac-tivities associated with increased tissue destruction asevidenced by decreased BPF values (figure 1) Strik-ingly we found that antibodies linked to diseasepathology as measured by BPF were enriched for reac-tivity against lipids Furthermore a selective increasein lipid-reactive antibodies linked to disease severitywas also observed when we analyzed all available MRImeasures (figure 2) Of note we did not detect an
Table 2 Continued
Antigens
BPF GMF WMF T2LV
r p FDR r p FDR r p FDR r p FDR
Thyrocalcitonin 2033 0143 0258 2008 0746 0996 2059 0005b 0108 004 0878 1000
TNPAL-galactocerebroside 007 0763 0969 2015 0506 0996 047 0030a 0142 005 0823 1000
Total cerebroside 2016 0498 0928 2041 0064 0112 050 0020a 0122 062 0003a 0033
Trisialogang GT1b 2034 0128 0194 2044 0043b 0066 015 0513 1000 030 0185 0247
Troponin I 2051 0020b 0040b 2053 0013b 0026b 2003 0900 1000 043 0050 0072
Ubiquitin 031 0166 0305 045 0039a 0027 2025 0282 0854 2029 0196 0247
VEGF 2050 0023b 0040b 2047 0031b 0027b 2013 0570 1000 040 0070 0146
Abbreviations BPF 5 whole brain parenchymal fraction FDR 5 false discovery ratendashadjusted significance value GMF 5 global cerebral gray matterfraction T2LV 5 cerebral T2 (fluid-attenuated inversion recovery) hyperintense lesion volume WMF 5 global cerebral white matter fractionDiscovery setap 005 (positive association)bp 005 (negative association)
Neurology Neuroimmunology amp Neuroinflammation 5
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
enrichment in lipid-reactive IgM in the group ofantibodies associated with increased diseasepathogenesis (not shown)
An index of serum IgG reactivity to lipids is associated
with MRI measures of disease severity To further inves-tigate the association between serum IgG reactivityto lipids and MRI measures of disease severity wecalculated an IgG anti-lipid index for each patientcorresponding to the information on each lipid-specific antibody listed in table 2 normalized by thestrength of its correlation with that MRI measureunder investigation The antibody reactivities tolipids used to calculate the lipid index are shown in
figure 3A With this approach we calculated one indexfor each of the MRI measures analyzed in this studyWe found a significant correlation between each IgGlipid antibody index and BPF GMF and T2LV(figure 3B) no significant correlation was found withWMF Moreover no significant correlations werefound between BPF WMF GMF and T2LVmeasures and anti-lipid antibody indexes based onIgM reactivity (not shown)
Finally we evaluated the performance of the lipidantibody indexes linked to BPF GMF and T2LV onan additional set of independent MS samples (valida-tion set table 1) We detected a similar trend to theone detected in the discovery set with regard to the
Figure 1 Antibody reactivity in serum is associated with decreased brain volume
Heatmap in which each column represents the mean immunoglobulin G antibody reactivity in a serum sample from a patient with multiple sclerosis sortedaccording to the whole brain parenchymal fraction (BPF) (indicated at the topmdasha lower BPF indicates more brain atrophy) and each row represents theantibody reactivity to an antigen according to the colorimetric scale shown The antibody reactivities included in this heatmap are listed in table 2
6 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
Figure 2 Association of serum IgG reactivity with MRI measures of disease severity
(A) Heatmap in which each column represents an MRI measure of either atrophy (BPF GMF WMF) or lesions (T2LV) andeach row represents the correlation to IgG serum antibody reactivity according to a colorimetric scale (B) Frequency oflipid-reactive antibodies linked to higher or lower MRI disease severity BPF 5 whole brain parenchymal fraction GMF 5
global cerebral gray matter fraction IgG 5 immunoglobulin G T2LV 5 cerebral T2 (fluid-attenuated inversion recovery)hyperintense lesion volume WMF 5 global cerebral white matter fraction
Neurology Neuroimmunology amp Neuroinflammation 7
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
correlation between BPF and GMF and their respec-tive lipid indexes and we validated the correlationbetween T2LV and its lipid index (figure 3C)
DISCUSSION Previous studies have investigated theassociation of MRI with immune activity in MS13ndash19
These studies have included cellular immune measures
Figure 3 Correlation of lipid indexes to MRI measures of disease severity in multiple sclerosis
(A) Heatmap inwhich each column represents anMRImeasure and each row represents the correlation to immunoglobulin G serumantibody reactivity to lipids accord-ing to a colorimetric scale (B) Scatter plots depicting the correlation between each lipid index andMRImeasures (BPF GMF and T2LV) in the discovery set (C) Scatterplots depicting the correlation between each lipid index andMRImeasures (BPF GMF and T2LV) in the validation set BPF5whole brain parenchymal fraction GMF5
cerebral gray matter fraction (see methods section for details) T2LV 5 cerebral T2 hyperintense lesion volume WMF 5 global cerebral white matter fraction
8 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
as well as oligoclonal bands or neurofilament-specificantibodies in CSF42ndash44 In the present study weanalyzed the association between serum antibodyprofiles detected with antigen microarrays and MRImeasures of disease including cerebral atrophy andT2 hyperintense lesions We found that specificantibody patterns are associated with different aspectsof MRI-defined disease pathology Our major findingwas that increased reactivity to lipids is associated withdifferent aspects of brain MRI measures of diseaseseverity Of note the specific set of lipids associatedwith atrophy differed from those associated withlesions Taken together these data suggest that anti-lipid antibodies in serum are related to MRI measuresof disease Our findings highlight the important role oflipid and lipid-specific immunity in the pathogenesis ofMS
Although a potential limitation of these studies isthe relatively small number of samples analyzed wehave validated our findings in an independent valida-tion cohort strengthening their significance In addi-tion further studies in large cohorts from patientsaffected by non-MS neurodegenerative diseases andhealthy controls may indicate whether the antibodyreactivities and lipid indexes identified in these stud-ies are exclusive to MS or are associated with addi-tional biological processes Indeed some of theseantibody reactivities contributing to the immune sig-natures described in this work have been found innewborns45
Different sets of antibody reactivities were associ-ated with GMF WMF BPF and T2LV The differ-ent antibody link between lesions and atrophy is inkeeping with the long-held view that brain atrophyin patients with MS is multifactorial and is onlyweakly related to lesions41 This has been underscoredin recent studies showing the complementary infor-mation obtained by combining lesion and atrophymeasures4647
Among the cerebral atrophy measures there was astriking overlap between those antibodies associatedwith GMF and BPF which separated them fromWMF results These findings are in keeping with pre-vious observations that whole brain atrophy is domi-nated by GM loss3741 In the early stages of MS thisGM atrophy selectively involves the deep GM nuclei34
Thus the similarities observed in the autoantibodieslinked to GMF and BPF might reflect the dominantcontribution and colinearity of GM atrophy to totalbrain atrophy The set of antibodies linked to GMFmay be the most relevant given that MRI studies haveshown that GM volume is more closely linked to phys-ical disability37 and cognitive impairment38 than areWM volume or lesion measures
Using antigen microarrays antibody reactivity tolipids has been detected in the CSF and serum of
patients with MS at different stages of the diseaseand such antibodies in the CSF have been linkedto disease progression and MRI involvement44
Although several mechanisms are thought to drivebrain inflammation and atrophy in MS46 the connec-tion between these mechanisms and the antibodiesdetected with our antigen microarrays is yet unknownOne possibility is that the serum autoantibodies iden-tified in our studies are not pathogenic and reflect theresult of immunization against self-antigens releasedfrom the CNS during the course of the disease Indeedneurofilament is released from damaged axons duringthe course of MS and both neurofilament lightchains48 and antibodies against it42 have been linkedto MRI measures of disease Moreover heat shockproteins are upregulated in different cell types duringinflammation and have been shown to have an impor-tant role as immunomodulators when released to theextracellular medium4950 Thus it is possible that heatshock proteinndashreactive antibodies reflect changes in theproduction and release of these immunomodulatorsduring the course of disease pathogenesis Similarlybioactive lipids and their products are released fromdamaged myelin and lipid-specific antibodies havebeen detected in patients with MS27293051 Lipids havean important role in the immune response both asbioactive molecules with immunomodulatory proper-ties and also as targets of the adaptive immuneresponse52 Moreover lipids have significant effectson the murine model of MS experimental autoim-mune encephalomyelitis2729305153 Indeed we recentlyfound that the glycolipid lactosylceramide activates abroad set of biological processes in astrocytes promot-ing neurodegeneration and inflammation53 Thus thelipid-reactive antibodies detected in this work mayreflect the release of myelin lipids in the context ofMS pathogenesis andor may directly contribute toimmune-mediated damage in CNS tissue Of notelipid-reactive antibodies are highly cross-reactive52 Inaddition the lipid-reactive antibodies detected in thiswork as associated with MRI measures of disease activ-ity were of the IgG class These are important points toconsider when evaluating a potential role of lipid-reactive antibodies inMS pathogenesis Further studiesare warranted to investigate whether bioactive lipidsoffer new therapeutic targets in MS
We have found that unique patterns of immunereactivity determined with antibody arrays are associ-ated with specific MRI measures of disease severityThese patterns agree with the interpretation that dif-ferent pathogenic mechanisms drive diverse diseaseprocesses reflected by these MRI measures These pat-terns also suggest a predominant role for lipids andlipid-specific immunity in MS pathology Furtherstudies are warranted to determine whether the earlyappearance of anti-lipid antibodies predicts the
Neurology Neuroimmunology amp Neuroinflammation 9
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
subsequent development of clinical and MRI-defineddisease worsening
AUTHOR CONTRIBUTIONSRohit Bakshi draftingrevising the manuscript study concept or design
analysis or interpretation of data acquisition of data study supervision
obtaining funding Ada Yeste study concept or design acquisition of
data Bonny Patel analysis or interpretation of data statistical analysis
Shahamat Tauhid analysis or interpretation of data Subhash Tummala
analysis or interpretation of data Roya Rahbari analysis or interpretation
of data statistical analysis Renxin Chu analysis or interpretation of data
Keren Regev analysis or interpretation of data acquisition of data
Pia Kivisaumlkk draftingrevising the manuscript study concept or design
contribution of vital reagentstoolspatients acquisition of data study
supervision Howard L Weiner study concept or design obtaining fund-
ing Francisco J Quintana draftingrevising the manuscript study con-
cept or design contribution of vital reagentstoolspatients study
supervision obtaining funding
ACKNOWLEDGMENTThe authors thank Dr Mohit Neema Dr Antonella Ceccarelli and
Dr Ashish Arora for valuable assistance at the early stages of this project
STUDY FUNDINGThis work was supported in part by grants to FJQ and HLW from
EMD Serono RG4111A1 and JF2161-A-5 from the National Multiple
Sclerosis Society PA0069 from the International Progressive MS Alli-
ance and by grants from the NIH (5R01NS055083-04) and NMSS
(RG3798A2) to RB
DISCLOSUREFJ Quintana serves on the editorial board for Systems Biomedicine
Inmunologia American Journal of Clinical and Experimental Immunology
is an associate editor for Immunology (UK) is an advisory board member
for Seminars in Immunopathology received research support from Harvard
Medical School BADERC NMSS A Yeste and B Patel report no
disclosures S Tauhid is managing editor for Journal of Neuroimaging
S Tummala R Rahbari R Chu and K Regev report no disclosures
P Kivisaumlkk received research support from EMD Serono R Bakshi is
editor-in-chief for Journal of Neuroimaging received consulting fees from
AbbVie Alkermes Biogen Novartis Questcor received research support
from Biogen EMD-Serono Novartis Sanofi-Genzyme Teva his spouse
holds stock in Biogen Inc H Weiner served on the advisory board
for The Guthy-Jackson Charitable Foundation Teva Pharmaceuticals
Industries Ltd Biogen Idec Novartis Sanofi-Aventis has consulted for
Therapix Bioven Novartis Serono Teva Sanofi received research support
from National Multiple Sclerosis Society Go to Neurologyorgnn for full
disclosure forms
Received July 27 2015 Accepted in final form December 8 2015
REFERENCES1 Weiner HL The challenge of multiple sclerosis how do
we cure a chronic heterogeneous disease Ann Neurol
200965239ndash248
2 Bakshi R Thompson AJ Rocca MA et al MRI in mul-
tiple sclerosis current status and future prospects Lancet
Neurol 20087615ndash625
3 Ceccarelli A Bakshi R Neema M MRI in multiple scle-
rosis a review of the current literature Curr Opin Neurol
201225402ndash409
4 Filippi M Rocca MA Barkhof F et al Association
between pathological and MRI findings in multiple scle-
rosis Lancet Neurol 201211349ndash360
5 Geurts JJ Calabrese M Fisher E Rudick RA Measure-
ment and clinical effect of grey matter pathology in mul-
tiple sclerosis Lancet Neurol 2012111082ndash1092
6 Klaver R De Vries HE Schenk GJ Geurts JJ Grey mat-
ter damage in multiple sclerosis a pathology perspective
Prion 2013766ndash75
7 Khoury S Bakshi R Cerebral pseudoatrophy or real atro-
phy after therapy in multiple sclerosis Ann Neurol 2010
68778ndash779
8 Hauser SL Chan JR Oksenberg JR Multiple sclerosis
prospects and promise Ann Neurol 201374317ndash327
9 McFarland HF Martin R Multiple sclerosis a complicated
picture of autoimmunity Nat Immunol 20078913ndash919
10 Nylander A Hafler DA Multiple sclerosis J Clin Invest
20121221180ndash1188
11 Sospedra M Martin R Immunology of multiple sclerosis
Ann Rev Immunol 200523683ndash747
12 Steinman L Immunology of relapse and remission in mul-
tiple sclerosis Ann Rev Immunol 201432257ndash281
13 Khoury SJ Guttmann CR Orav EJ Kikinis R Jolesz FA
Weiner HL Changes in activated T cells in the blood
correlate with disease activity in multiple sclerosis Arch
Neurol 2000571183ndash1189
14 Khoury SJ Orav EJ Guttmann CR Kikinis R Jolesz FA
Weiner HL Changes in serum levels of ICAM and TNF-
R correlate with disease activity in multiple sclerosis Neu-
rology 199953758ndash764
15 Laplaud DA Berthelot L Miqueu P et al Serial blood T
cell repertoire alterations in multiple sclerosis patients cor-
relation with clinical and MRI parameters J Neuroimmunol
2006177151ndash160
16 Makhlouf K Weiner HL Khoury SJ Increased percentage
of IL-121 monocytes in the blood correlates with the
presence of active MRI lesions in MS J Neuroimmunol
2001119145ndash149
17 Prat A Biernacki K Saroli T et al Kinin B1 receptor expres-
sion on multiple sclerosis mononuclear cells correlation with
magnetic resonance imaging T2-weighted lesion volume and
clinical disability Arch Neurol 200562795ndash800
18 Rinaldi L Gallo P Calabrese M et al Longitudinal anal-
ysis of immune cell phenotypes in early stage multiple
sclerosis distinctive patterns characterize MRI-active pa-
tients Brain 20061291993ndash2007
19 Tortorella P Lagana MM Saresella M et al Determinants
of disability in multiple sclerosis an immunological and
MRI study Biomed Res Int 20142014875768
20 Robinson WH DiGennaro C Hueber W et al Autoan-
tigen microarrays for multiplex characterization of autoan-
tibody responses Nat Med 20028295ndash301
21 Yeste A Quintana FJ Antigen microarrays for the study of
autoimmune diseases Clin Chem 2013591036ndash1044
22 Li QZ Xie C Wu T et al Identification of autoantibody
clusters that best predict lupus disease activity using glomer-
ular proteome arrays J Clin Invest 20051153428ndash3439
23 Hueber W Kidd BA Tomooka BH et al Antigen micro-
array profiling of autoantibodies in rheumatoid arthritis
Arthritis Rheum 2005522645ndash2655
24 Goldschmidt Y Sharon E Quintana FJ Cohen IR
Brandt A Adaptive methods for classification of biological
microarray data from multiple experiments Report no
MCS03-07 Rehovot Israel The Arthur and Rochelle Bel-
fer Institute of Mathematics and Computer Science 2003
25 Quintana FJ Getz G Hed G Domany E Cohen IR
Cluster analysis of human autoantibody reactivities in
health and in type 1 diabetes mellitus a bio-informatic
approach to immune complexity J Autoimmun 2003
2165ndash75
10 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
26 Lalive PH Menge T Barman I Cree BA Genain CP
Identification of new serum autoantibodies in neuromye-
litis optica using protein microarrays Neurology 200667
176ndash177
27 Kanter JL Narayana S Ho PP et al Lipid microarrays
identify key mediators of autoimmune brain inflamma-
tion Nat Med 200612138ndash143
28 Quintana FJ Farez MF Izquierdo G Lucas M Cohen IR
Weiner HL Antigen microarrays identify CNS-produced
autoantibodies in RRMS Neurology 201278532ndash539
29 Farez MF Quintana FJ Gandhi R Izquierdo G Lucas M
Weiner HL Toll-like receptor 2 and poly(ADP-ribose)
polymerase 1 promote central nervous system neuroin-
flammation in progressive EAE Nat Immunol 200910
958ndash964
30 Quintana FJ Farez MF Viglietta V et al Antigen micro-
arrays identify unique serum autoantibody signatures in
clinical and pathologic subtypes of multiple sclerosis Proc
Natl Acad Sci USA 200810518889ndash18894
31 Quintana FJ Patel B Yeste A et al Epitope spreading as
an early pathogenic event in pediatric multiple sclerosis
Neurology 2014832219ndash2226
32 Gauthier SA Glanz BI Mandel M Weiner HL A model
for the comprehensive investigation of a chronic autoim-
mune disease the multiple sclerosis CLIMB study Auto-
immun Rev 20065532ndash536
33 Polman CH Reingold SC Edan G et al Diagnostic criteria
for multiple sclerosis 2005 revisions to the ldquoMcDonald
Criteriardquo Ann Neurol 200558840ndash846
34 DellrsquoOglio E Ceccarelli A Glanz BI et al Quantification
of global cerebral atrophy in multiple sclerosis from 3T
MRI using SPM the role of misclassification errors
J Neuroimaging 201525191ndash199
35 Ceccarelli A Jackson JS Tauhid S et al The impact of
lesion in-painting and registration methods on voxel-based
morphometry in detecting regional cerebral gray matter
atrophy in multiple sclerosis AJNR Am J Neuroradiol
2012331579ndash1585
36 Cohen AB Neema M Arora A et al The relationships
among MRI-defined spinal cord involvement brain involve-
ment and disability in multiple sclerosis J Neuroimaging
201222122ndash128
37 Sanfilipo MP Benedict RH Sharma J Weinstock-
Guttman B Bakshi R The relationship between whole
brain volume and disability in multiple sclerosis a com-
parison of normalized gray vs white matter with misclas-
sification correction Neuroimage 2005261068ndash1077
38 Sanfilipo MP Benedict RH Weinstock-Guttman B
Bakshi R Gray and white matter brain atrophy and neuro-
psychological impairment in multiple sclerosis Neurology
200666685ndash692
39 Stankiewicz JM Glanz BI Healy BC et al Brain MRI
lesion load at 15T and 3T versus clinical status in multi-
ple sclerosis J Neuroimaging 201121e50ndashe56
40 Quintana FJ Hagedorn PH Elizur G Merbl Y
Domany E Cohen IR Functional immunomics micro-
array analysis of IgG autoantibody repertoires predicts the
future response of mice to induced diabetes Proc Natl
Acad Sci USA 2004101(suppl 2)14615ndash14621
41 Bermel RA Bakshi R The measurement and clinical
relevance of brain atrophy in multiple sclerosis Lancet
Neurol 20065158ndash170
42 Eikelenboom MJ Petzold A Lazeron RH et al Multiple
sclerosis neurofilament light chain antibodies are corre-
lated to cerebral atrophy Neurology 200360219ndash223
43 Magraner MJ Bosca I Simo-Castello M et al Brain atro-
phy and lesion load are related to CSF lipid-specific IgM
oligoclonal bands in clinically isolated syndromes Neuro-
radiol 2012545ndash12
44 Villar LM Sadaba MC Roldan E et al Intrathecal syn-
thesis of oligoclonal IgM against myelin lipids predicts an
aggressive disease course in MS J Clin Invest 2005115
187ndash194
45 Merbl Y Zucker-Toledano M Quintana FJ Cohen IR
Newborn humans manifest autoantibodies to defined self
molecules detected by antigen microarray informatics
J Clin Invest 2007117712ndash718
46 Sormani MP Arnold DL De Stefano N Treatment effect
on brain atrophy correlates with treatment effect on disa-
bility in multiple sclerosis Ann Neurol 20147543ndash49
47 Tauhid S Neema M Healy BC Weiner HL Bakshi R
MRI phenotypes based on cerebral lesions and atrophy in
patients with multiple sclerosis J Neurol Sci 2014346
250ndash254
48 Gnanapavan S Grant D Morant S et al Biomarker
report from the phase II lamotrigine trial in secondary
progressive MS neurofilament as a surrogate of disease
progression PLoS One 20138e70019
49 Quintana FJ Cohen IR Heat shock proteins as endoge-
nous adjuvants in sterile and septic inflammation
J Immunol 20051752777ndash2782
50 Quintana FJ Cohen IR The HSP60 immune system net-
work Trends Immunol 20113289ndash95
51 Ho PP Kanter JL Johnson AM et al Identification of
naturally occurring fatty acids of the myelin sheath that
resolve neuroinflammation Sci Transl Med 20124137ra73
52 Quintana FJ Yeste A Weiner HL Covacu R Lipids and
lipid-reactive antibodies as biomarkers for multiple sclero-
sis J Neuroimmunol 201224853ndash57
53 Mayo L Trauger SA Blain M et al Regulation of astro-
cyte activation by glycolipids drives chronic CNS inflam-
mation Nat Med 2014201147ndash1156
Neurology Neuroimmunology amp Neuroinflammation 11
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
1
Table e-1 Antigens used
Group Antigen Source
27 kDa Heat Shock Protein Stressgen
HSP 32 kDa Heat Shock Protein Stressgen
40 kDa Heat Shock Protein Stressgen
47 kDa Heat Shock Protein Stressgen
60 kDa Heat Shock Protein Stressgen
60 kDa Heat Shock Protein peptide aa 106ndash125 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 121ndash140 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 136ndash155 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 151ndash170 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 16ndash35 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 166ndash185 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
2
60 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 556ndash573 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
65 kDa Heat Shock Protein M tuberculosis Stressgen
70 kDa Heat Shock Protein Stressgen
70 kDa Heat Shock Protein peptide aa 106ndash125 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 121ndash140 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 136ndash155 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 151ndash170 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 16ndash35 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 166ndash185 Biopolymers Facility Harvard Medical School
3
70 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
4
70 kDa Heat Shock Protein peptide aa 556ndash575 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 571ndash590 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 586ndash605 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 601ndash620 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 616ndash635 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 631ndash640 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
71 kDa Heat Shock Protein M tuberculosis Biopolymers Facility Harvard Medical School
90 kDa Heat Shock Protein Stressgen
GroEL Stressgen
23-cyclic nucleotide 3-phosphodiesterase
peptide aa106-125 Biopolymers Facility Harvard Medical School
CNS
23-cyclic nucleotide 3-phosphodiesterase
peptide aa1-20 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa121-140 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa136-155 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa151-170 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa16-35 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa166-185 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase Biopolymers Facility Harvard Medical School
5
peptide aa181-200
23-cyclic nucleotide 3-phosphodiesterase
peptide aa196-215 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa211-230 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa226-245 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa241-260 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa256-275 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa271-290 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa286-305 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa301-320 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa31-50 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa316-335 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa331-350 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa346-365 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa361-380 Biopolymers Facility Harvard Medical School
6
23-cyclic nucleotide 3-phosphodiesterase
peptide aa376-395 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa391-410 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa406-421 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa46-65 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa61-80 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa76-95 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa91-110 Biopolymers Facility Harvard Medical School
Acetyl Cholinesterase Sigma Aldrich
ADAM-10 Sigma Aldrich
alpha-Cristallin Stressgen
beta-Cristallin Sigma Aldrich
bovine Myelin Basic Protein Sigma Aldrich
Brain Extract I Sigma Aldrich
Brain Extract II Sigma Aldrich
Brain Extract III Sigma Aldrich
Glial Filament Acidic Protein Research Diagnostic
guinea pig Myelin Basic Protein Sigma Aldrich
human Myelin Basic Protein Sigma Aldrich
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 106-125 Biopolymers Facility Harvard Medical School
7
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 121-140 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 136-155 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 151-170 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 166-185 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 181-200 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 91-110 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
106-125 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa 1- Biopolymers Facility Harvard Medical School
8
20
Myelinoligodendrocyte glycoprotein peptide aa
121-140 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
136-155 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
151-170 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
16-35 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
166-185 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
181-200 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
196-215 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
211-230 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
226-247 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
31-50 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
35-55 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
46-65 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
61-80 Biopolymers Facility Harvard Medical School
9
Myelinoligodendrocyte glycoprotein peptide aa
76-95 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
91-110 Biopolymers Facility Harvard Medical School
murine Myelin Basic Protein Sigma Aldrich
Myelin Associated Glycoprotein Sigma Aldrich
Myelin Basic Protein peptide aa 104-123 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 11-30 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 113-132 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 121-138 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 124-142 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 138-147 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 141-161 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 143-168 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 155-178 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 26-35 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 41-60 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 51-70 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 71-92 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 84-94 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 89-101 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 173-186 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 93-112 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 106-125 Biopolymers Facility Harvard Medical School
10
Myelin Protein 2 peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 121-132 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 91-110 Biopolymers Facility Harvard Medical School
Neurofilament 160kd Chemicon
Neurofilament 200kd Chemicon
Neurofilament 68kd Chemicon
Neuronal Enolase Calbiochem
Nicastrin GeneTex
NMDA receptor Novus Biologicals
NOGO Sigma Aldrich
Olygodendrocyte-Specific Protein peptide aa 106ndash
125 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 121ndash
140 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 136ndash
155 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 151ndash
170 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 16ndash
35 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 166ndash Biopolymers Facility Harvard Medical School
11
185
Olygodendrocyte-Specific Protein peptide aa 181ndash
199 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 195ndash
217 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 31ndash
50 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 46ndash
65 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 61ndash
80 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 76ndash
95 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 91ndash
110 Biopolymers Facility Harvard Medical School
Proteolipid Protein Abnova
Proteolipid Protein peptide aa 100-119 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 10-29 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 110-129 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 1-19 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 125-141 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-150 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-154 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 150-163 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 151-173 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 158-166 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 161-180 Biopolymers Facility Harvard Medical School
12
Proteolipid Protein peptide aa 178-191 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 180-199 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 190-209 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 20-39 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 205-220 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 215-232 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-239 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-249 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 248-259 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 250-269 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 265-277 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 35-50 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 40-59 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 50-69 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 65-84 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 80-99 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 91-110 Biopolymers Facility Harvard Medical School
Retinol Binding Protein Sigma Aldrich
S100beta protein Assay Designs
Super Oxide Dismutase Sigma Aldrich
Synuclein beta Sigma Aldrich
Synuclein gamma Sigma Aldrich
Amydgala ProSci Inc
Tissue Amydgala AD ProSci Inc
Brain lysate ProSci Inc
Brain Tissue Membrane ProSci Inc
Cerebellar pedunculus ProSci Inc
13
Cerebral meninges ProSci Inc
Corpus Callosum ProSci Inc
Corpus Callosum AD ProSci Inc
Diencephalon ProSci Inc
Fetal brain ProSci Inc
Frontal lobe ProSci Inc
Frontal lobe AD ProSci Inc
Hippocampus ProSci Inc
Hippocampus AD ProSci Inc
Insula ProSci Inc
Occipital lobe ProSci Inc
Occipital lobe AD ProSci Inc
Olfactory region ProSci Inc
Optic Nerve ProSci Inc
Parietal lobe ProSci Inc
Parietal lobe AD ProSci Inc
Pons ProSci Inc
Pons AD ProSci Inc
Postcentral gyrus ProSci Inc
Postcentral gyrus AD ProSci Inc
Precentral gyrus ProSci Inc
Precentral gyrus AD ProSci Inc
Spinal cord ProSci Inc
Temporal lobe ProSci Inc
Temporal lobe AD ProSci Inc
Thalamus ProSci Inc
Thalamus AD ProSci Inc
14
Amyloid beta Sigma Aldrich
AD
related Amyloid beta 10-20 Sigma Aldrich
Amyloid beta 1-12 Sigma Aldrich
Amyloid beta 12-28 Sigma Aldrich
Amyloid beta 1-23 Sigma Aldrich
Amyloid beta 1-38 Sigma Aldrich
Amyloid beta 17-40 Sigma Aldrich
Amyloid beta 25-35 Sigma Aldrich
Amyloid beta 34-42 Sigma Aldrich
Amyloid bri protein precursor 227 Sigma Aldrich
Amyloid DAN Protein Fragment 1-34 Sigma Aldrich
Amyloid Precursor Protein Sigma Aldrich
Amyloid protein no AB component Sigma Aldrich
Secreted amyloid precursor protein (SAP) beta Sigma Aldrich
Tau isoform variant 0N3R Sigma Aldrich
Tau isoform variant 1N3R Sigma Aldrich
Tau isoform variant 0N4R Sigma Aldrich
Tau isoform variant 2N3R Sigma Aldrich
Tau phospho Ser412 Sigma Aldrich
Tau phospho Ser441 Sigma Aldrich
Tau phospho Thr181 Sigma Aldrich
Tau Protein human EMD Biosciences
(plusmn)9-HODE Cayman Chemical
Lipids
1 Palmitoyl-2-(5oxo-Valeroyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
15a-hydroxycholestene Avanti Polar Lipids
15
15-ketocholestane Avanti Polar Lipids
15-ketocholestene Avanti Polar Lipids
1-Palmitoil-2-(9oxo-Nonanoyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Azelaoyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Glutaroyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
5 α-cholestane-3 β15 α-diol Avanti Polar Lipids
9(S)-HODE Cayman Chemical
Asialoganglioside-GM1 Sigma Aldrich
Asialoganglioside-GM2 Sigma Aldrich
Brain ceramides Avanti Polar Lipids
Brain D-erythrosphingosine Avanti Polar Lipids
Brain lysophosphatidylethanolamine Avanti Polar Lipids
Brain L-α-lysophosphatidylserine Avanti Polar Lipids
Brain L-α-phosphatidylcholine Avanti Polar Lipids
Brain L-α-phosphatidyl-ethanolamine Avanti Polar Lipids
Brain L-α-phosphatidylserine Avanti Polar Lipids
Brain polar lipid extract Avanti Polar Lipids
Brain sphingomyelin Avanti Polar Lipids
Brain sulfatide Avanti Polar Lipids
Brain total lipid extract Avanti Polar Lipids
Cardiolipin Sigma Aldrich
Ceramide Sigma Aldrich
Ceramide 1-phosphate Sigma Aldrich
Cholesterol Sigma Aldrich
16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-
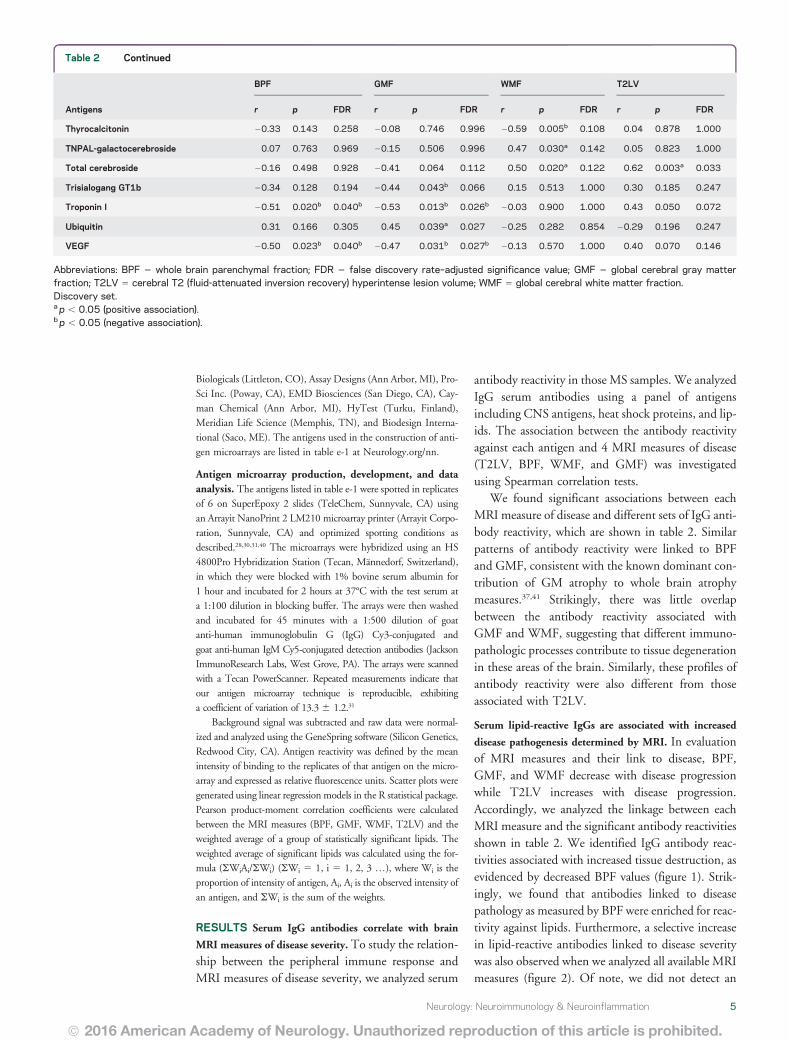
Biologicals (Littleton CO) Assay Designs (Ann Arbor MI) Pro-
Sci Inc (Poway CA) EMD Biosciences (San Diego CA) Cay-
man Chemical (Ann Arbor MI) HyTest (Turku Finland)
Meridian Life Science (Memphis TN) and Biodesign Interna-
tional (Saco ME) The antigens used in the construction of anti-
gen microarrays are listed in table e-1 at Neurologyorgnn
Antigen microarray production development and dataanalysis The antigens listed in table e-1 were spotted in replicatesof 6 on SuperEpoxy 2 slides (TeleChem Sunnyvale CA) using
an Arrayit NanoPrint 2 LM210 microarray printer (Arrayit Corpo-
ration Sunnyvale CA) and optimized spotting conditions as
described28303140 The microarrays were hybridized using an HS
4800Pro Hybridization Station (Tecan Maumlnnedorf Switzerland)
in which they were blocked with 1 bovine serum albumin for
1 hour and incubated for 2 hours at 37degC with the test serum at
a 1100 dilution in blocking buffer The arrays were then washed
and incubated for 45 minutes with a 1500 dilution of goat
anti-human immunoglobulin G (IgG) Cy3-conjugated and
goat anti-human IgM Cy5-conjugated detection antibodies (Jackson
ImmunoResearch Labs West Grove PA) The arrays were scanned
with a Tecan PowerScanner Repeated measurements indicate that
our antigen microarray technique is reproducible exhibiting
a coefficient of variation of 1336 1231
Background signal was subtracted and raw data were normal-
ized and analyzed using the GeneSpring software (Silicon Genetics
Redwood City CA) Antigen reactivity was defined by the mean
intensity of binding to the replicates of that antigen on the micro-
array and expressed as relative fluorescence units Scatter plots were
generated using linear regression models in the R statistical package
Pearson product-moment correlation coefficients were calculated
between the MRI measures (BPF GMF WMF T2LV) and the
weighted average of a group of statistically significant lipids The
weighted average of significant lipids was calculated using the for-
mula (SWiAiSWi) (SWi 5 1 i 5 1 2 3 ) where Wi is the
proportion of intensity of antigen Ai Ai is the observed intensity of
an antigen and SWi is the sum of the weights
RESULTS Serum IgG antibodies correlate with brain
MRI measures of disease severity To study the relation-ship between the peripheral immune response andMRI measures of disease severity we analyzed serum
antibody reactivity in those MS samples We analyzedIgG serum antibodies using a panel of antigensincluding CNS antigens heat shock proteins and lip-ids The association between the antibody reactivityagainst each antigen and 4 MRI measures of disease(T2LV BPF WMF and GMF) was investigatedusing Spearman correlation tests
We found significant associations between eachMRI measure of disease and different sets of IgG anti-body reactivity which are shown in table 2 Similarpatterns of antibody reactivity were linked to BPFand GMF consistent with the known dominant con-tribution of GM atrophy to whole brain atrophymeasures3741 Strikingly there was little overlapbetween the antibody reactivity associated withGMF and WMF suggesting that different immuno-pathologic processes contribute to tissue degenerationin these areas of the brain Similarly these profiles ofantibody reactivity were also different from thoseassociated with T2LV
Serum lipid-reactive IgGs are associated with increased
disease pathogenesis determined by MRI In evaluationof MRI measures and their link to disease BPFGMF and WMF decrease with disease progressionwhile T2LV increases with disease progressionAccordingly we analyzed the linkage between eachMRI measure and the significant antibody reactivitiesshown in table 2 We identified IgG antibody reac-tivities associated with increased tissue destruction asevidenced by decreased BPF values (figure 1) Strik-ingly we found that antibodies linked to diseasepathology as measured by BPF were enriched for reac-tivity against lipids Furthermore a selective increasein lipid-reactive antibodies linked to disease severitywas also observed when we analyzed all available MRImeasures (figure 2) Of note we did not detect an
Table 2 Continued
Antigens
BPF GMF WMF T2LV
r p FDR r p FDR r p FDR r p FDR
Thyrocalcitonin 2033 0143 0258 2008 0746 0996 2059 0005b 0108 004 0878 1000
TNPAL-galactocerebroside 007 0763 0969 2015 0506 0996 047 0030a 0142 005 0823 1000
Total cerebroside 2016 0498 0928 2041 0064 0112 050 0020a 0122 062 0003a 0033
Trisialogang GT1b 2034 0128 0194 2044 0043b 0066 015 0513 1000 030 0185 0247
Troponin I 2051 0020b 0040b 2053 0013b 0026b 2003 0900 1000 043 0050 0072
Ubiquitin 031 0166 0305 045 0039a 0027 2025 0282 0854 2029 0196 0247
VEGF 2050 0023b 0040b 2047 0031b 0027b 2013 0570 1000 040 0070 0146
Abbreviations BPF 5 whole brain parenchymal fraction FDR 5 false discovery ratendashadjusted significance value GMF 5 global cerebral gray matterfraction T2LV 5 cerebral T2 (fluid-attenuated inversion recovery) hyperintense lesion volume WMF 5 global cerebral white matter fractionDiscovery setap 005 (positive association)bp 005 (negative association)
Neurology Neuroimmunology amp Neuroinflammation 5
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
enrichment in lipid-reactive IgM in the group ofantibodies associated with increased diseasepathogenesis (not shown)
An index of serum IgG reactivity to lipids is associated
with MRI measures of disease severity To further inves-tigate the association between serum IgG reactivityto lipids and MRI measures of disease severity wecalculated an IgG anti-lipid index for each patientcorresponding to the information on each lipid-specific antibody listed in table 2 normalized by thestrength of its correlation with that MRI measureunder investigation The antibody reactivities tolipids used to calculate the lipid index are shown in
figure 3A With this approach we calculated one indexfor each of the MRI measures analyzed in this studyWe found a significant correlation between each IgGlipid antibody index and BPF GMF and T2LV(figure 3B) no significant correlation was found withWMF Moreover no significant correlations werefound between BPF WMF GMF and T2LVmeasures and anti-lipid antibody indexes based onIgM reactivity (not shown)
Finally we evaluated the performance of the lipidantibody indexes linked to BPF GMF and T2LV onan additional set of independent MS samples (valida-tion set table 1) We detected a similar trend to theone detected in the discovery set with regard to the
Figure 1 Antibody reactivity in serum is associated with decreased brain volume
Heatmap in which each column represents the mean immunoglobulin G antibody reactivity in a serum sample from a patient with multiple sclerosis sortedaccording to the whole brain parenchymal fraction (BPF) (indicated at the topmdasha lower BPF indicates more brain atrophy) and each row represents theantibody reactivity to an antigen according to the colorimetric scale shown The antibody reactivities included in this heatmap are listed in table 2
6 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
Figure 2 Association of serum IgG reactivity with MRI measures of disease severity
(A) Heatmap in which each column represents an MRI measure of either atrophy (BPF GMF WMF) or lesions (T2LV) andeach row represents the correlation to IgG serum antibody reactivity according to a colorimetric scale (B) Frequency oflipid-reactive antibodies linked to higher or lower MRI disease severity BPF 5 whole brain parenchymal fraction GMF 5
global cerebral gray matter fraction IgG 5 immunoglobulin G T2LV 5 cerebral T2 (fluid-attenuated inversion recovery)hyperintense lesion volume WMF 5 global cerebral white matter fraction
Neurology Neuroimmunology amp Neuroinflammation 7
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
correlation between BPF and GMF and their respec-tive lipid indexes and we validated the correlationbetween T2LV and its lipid index (figure 3C)
DISCUSSION Previous studies have investigated theassociation of MRI with immune activity in MS13ndash19
These studies have included cellular immune measures
Figure 3 Correlation of lipid indexes to MRI measures of disease severity in multiple sclerosis
(A) Heatmap inwhich each column represents anMRImeasure and each row represents the correlation to immunoglobulin G serumantibody reactivity to lipids accord-ing to a colorimetric scale (B) Scatter plots depicting the correlation between each lipid index andMRImeasures (BPF GMF and T2LV) in the discovery set (C) Scatterplots depicting the correlation between each lipid index andMRImeasures (BPF GMF and T2LV) in the validation set BPF5whole brain parenchymal fraction GMF5
cerebral gray matter fraction (see methods section for details) T2LV 5 cerebral T2 hyperintense lesion volume WMF 5 global cerebral white matter fraction
8 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
as well as oligoclonal bands or neurofilament-specificantibodies in CSF42ndash44 In the present study weanalyzed the association between serum antibodyprofiles detected with antigen microarrays and MRImeasures of disease including cerebral atrophy andT2 hyperintense lesions We found that specificantibody patterns are associated with different aspectsof MRI-defined disease pathology Our major findingwas that increased reactivity to lipids is associated withdifferent aspects of brain MRI measures of diseaseseverity Of note the specific set of lipids associatedwith atrophy differed from those associated withlesions Taken together these data suggest that anti-lipid antibodies in serum are related to MRI measuresof disease Our findings highlight the important role oflipid and lipid-specific immunity in the pathogenesis ofMS
Although a potential limitation of these studies isthe relatively small number of samples analyzed wehave validated our findings in an independent valida-tion cohort strengthening their significance In addi-tion further studies in large cohorts from patientsaffected by non-MS neurodegenerative diseases andhealthy controls may indicate whether the antibodyreactivities and lipid indexes identified in these stud-ies are exclusive to MS or are associated with addi-tional biological processes Indeed some of theseantibody reactivities contributing to the immune sig-natures described in this work have been found innewborns45
Different sets of antibody reactivities were associ-ated with GMF WMF BPF and T2LV The differ-ent antibody link between lesions and atrophy is inkeeping with the long-held view that brain atrophyin patients with MS is multifactorial and is onlyweakly related to lesions41 This has been underscoredin recent studies showing the complementary infor-mation obtained by combining lesion and atrophymeasures4647
Among the cerebral atrophy measures there was astriking overlap between those antibodies associatedwith GMF and BPF which separated them fromWMF results These findings are in keeping with pre-vious observations that whole brain atrophy is domi-nated by GM loss3741 In the early stages of MS thisGM atrophy selectively involves the deep GM nuclei34
Thus the similarities observed in the autoantibodieslinked to GMF and BPF might reflect the dominantcontribution and colinearity of GM atrophy to totalbrain atrophy The set of antibodies linked to GMFmay be the most relevant given that MRI studies haveshown that GM volume is more closely linked to phys-ical disability37 and cognitive impairment38 than areWM volume or lesion measures
Using antigen microarrays antibody reactivity tolipids has been detected in the CSF and serum of
patients with MS at different stages of the diseaseand such antibodies in the CSF have been linkedto disease progression and MRI involvement44
Although several mechanisms are thought to drivebrain inflammation and atrophy in MS46 the connec-tion between these mechanisms and the antibodiesdetected with our antigen microarrays is yet unknownOne possibility is that the serum autoantibodies iden-tified in our studies are not pathogenic and reflect theresult of immunization against self-antigens releasedfrom the CNS during the course of the disease Indeedneurofilament is released from damaged axons duringthe course of MS and both neurofilament lightchains48 and antibodies against it42 have been linkedto MRI measures of disease Moreover heat shockproteins are upregulated in different cell types duringinflammation and have been shown to have an impor-tant role as immunomodulators when released to theextracellular medium4950 Thus it is possible that heatshock proteinndashreactive antibodies reflect changes in theproduction and release of these immunomodulatorsduring the course of disease pathogenesis Similarlybioactive lipids and their products are released fromdamaged myelin and lipid-specific antibodies havebeen detected in patients with MS27293051 Lipids havean important role in the immune response both asbioactive molecules with immunomodulatory proper-ties and also as targets of the adaptive immuneresponse52 Moreover lipids have significant effectson the murine model of MS experimental autoim-mune encephalomyelitis2729305153 Indeed we recentlyfound that the glycolipid lactosylceramide activates abroad set of biological processes in astrocytes promot-ing neurodegeneration and inflammation53 Thus thelipid-reactive antibodies detected in this work mayreflect the release of myelin lipids in the context ofMS pathogenesis andor may directly contribute toimmune-mediated damage in CNS tissue Of notelipid-reactive antibodies are highly cross-reactive52 Inaddition the lipid-reactive antibodies detected in thiswork as associated with MRI measures of disease activ-ity were of the IgG class These are important points toconsider when evaluating a potential role of lipid-reactive antibodies inMS pathogenesis Further studiesare warranted to investigate whether bioactive lipidsoffer new therapeutic targets in MS
We have found that unique patterns of immunereactivity determined with antibody arrays are associ-ated with specific MRI measures of disease severityThese patterns agree with the interpretation that dif-ferent pathogenic mechanisms drive diverse diseaseprocesses reflected by these MRI measures These pat-terns also suggest a predominant role for lipids andlipid-specific immunity in MS pathology Furtherstudies are warranted to determine whether the earlyappearance of anti-lipid antibodies predicts the
Neurology Neuroimmunology amp Neuroinflammation 9
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
subsequent development of clinical and MRI-defineddisease worsening
AUTHOR CONTRIBUTIONSRohit Bakshi draftingrevising the manuscript study concept or design
analysis or interpretation of data acquisition of data study supervision
obtaining funding Ada Yeste study concept or design acquisition of
data Bonny Patel analysis or interpretation of data statistical analysis
Shahamat Tauhid analysis or interpretation of data Subhash Tummala
analysis or interpretation of data Roya Rahbari analysis or interpretation
of data statistical analysis Renxin Chu analysis or interpretation of data
Keren Regev analysis or interpretation of data acquisition of data
Pia Kivisaumlkk draftingrevising the manuscript study concept or design
contribution of vital reagentstoolspatients acquisition of data study
supervision Howard L Weiner study concept or design obtaining fund-
ing Francisco J Quintana draftingrevising the manuscript study con-
cept or design contribution of vital reagentstoolspatients study
supervision obtaining funding
ACKNOWLEDGMENTThe authors thank Dr Mohit Neema Dr Antonella Ceccarelli and
Dr Ashish Arora for valuable assistance at the early stages of this project
STUDY FUNDINGThis work was supported in part by grants to FJQ and HLW from
EMD Serono RG4111A1 and JF2161-A-5 from the National Multiple
Sclerosis Society PA0069 from the International Progressive MS Alli-
ance and by grants from the NIH (5R01NS055083-04) and NMSS
(RG3798A2) to RB
DISCLOSUREFJ Quintana serves on the editorial board for Systems Biomedicine
Inmunologia American Journal of Clinical and Experimental Immunology
is an associate editor for Immunology (UK) is an advisory board member
for Seminars in Immunopathology received research support from Harvard
Medical School BADERC NMSS A Yeste and B Patel report no
disclosures S Tauhid is managing editor for Journal of Neuroimaging
S Tummala R Rahbari R Chu and K Regev report no disclosures
P Kivisaumlkk received research support from EMD Serono R Bakshi is
editor-in-chief for Journal of Neuroimaging received consulting fees from
AbbVie Alkermes Biogen Novartis Questcor received research support
from Biogen EMD-Serono Novartis Sanofi-Genzyme Teva his spouse
holds stock in Biogen Inc H Weiner served on the advisory board
for The Guthy-Jackson Charitable Foundation Teva Pharmaceuticals
Industries Ltd Biogen Idec Novartis Sanofi-Aventis has consulted for
Therapix Bioven Novartis Serono Teva Sanofi received research support
from National Multiple Sclerosis Society Go to Neurologyorgnn for full
disclosure forms
Received July 27 2015 Accepted in final form December 8 2015
REFERENCES1 Weiner HL The challenge of multiple sclerosis how do
we cure a chronic heterogeneous disease Ann Neurol
200965239ndash248
2 Bakshi R Thompson AJ Rocca MA et al MRI in mul-
tiple sclerosis current status and future prospects Lancet
Neurol 20087615ndash625
3 Ceccarelli A Bakshi R Neema M MRI in multiple scle-
rosis a review of the current literature Curr Opin Neurol
201225402ndash409
4 Filippi M Rocca MA Barkhof F et al Association
between pathological and MRI findings in multiple scle-
rosis Lancet Neurol 201211349ndash360
5 Geurts JJ Calabrese M Fisher E Rudick RA Measure-
ment and clinical effect of grey matter pathology in mul-
tiple sclerosis Lancet Neurol 2012111082ndash1092
6 Klaver R De Vries HE Schenk GJ Geurts JJ Grey mat-
ter damage in multiple sclerosis a pathology perspective
Prion 2013766ndash75
7 Khoury S Bakshi R Cerebral pseudoatrophy or real atro-
phy after therapy in multiple sclerosis Ann Neurol 2010
68778ndash779
8 Hauser SL Chan JR Oksenberg JR Multiple sclerosis
prospects and promise Ann Neurol 201374317ndash327
9 McFarland HF Martin R Multiple sclerosis a complicated
picture of autoimmunity Nat Immunol 20078913ndash919
10 Nylander A Hafler DA Multiple sclerosis J Clin Invest
20121221180ndash1188
11 Sospedra M Martin R Immunology of multiple sclerosis
Ann Rev Immunol 200523683ndash747
12 Steinman L Immunology of relapse and remission in mul-
tiple sclerosis Ann Rev Immunol 201432257ndash281
13 Khoury SJ Guttmann CR Orav EJ Kikinis R Jolesz FA
Weiner HL Changes in activated T cells in the blood
correlate with disease activity in multiple sclerosis Arch
Neurol 2000571183ndash1189
14 Khoury SJ Orav EJ Guttmann CR Kikinis R Jolesz FA
Weiner HL Changes in serum levels of ICAM and TNF-
R correlate with disease activity in multiple sclerosis Neu-
rology 199953758ndash764
15 Laplaud DA Berthelot L Miqueu P et al Serial blood T
cell repertoire alterations in multiple sclerosis patients cor-
relation with clinical and MRI parameters J Neuroimmunol
2006177151ndash160
16 Makhlouf K Weiner HL Khoury SJ Increased percentage
of IL-121 monocytes in the blood correlates with the
presence of active MRI lesions in MS J Neuroimmunol
2001119145ndash149
17 Prat A Biernacki K Saroli T et al Kinin B1 receptor expres-
sion on multiple sclerosis mononuclear cells correlation with
magnetic resonance imaging T2-weighted lesion volume and
clinical disability Arch Neurol 200562795ndash800
18 Rinaldi L Gallo P Calabrese M et al Longitudinal anal-
ysis of immune cell phenotypes in early stage multiple
sclerosis distinctive patterns characterize MRI-active pa-
tients Brain 20061291993ndash2007
19 Tortorella P Lagana MM Saresella M et al Determinants
of disability in multiple sclerosis an immunological and
MRI study Biomed Res Int 20142014875768
20 Robinson WH DiGennaro C Hueber W et al Autoan-
tigen microarrays for multiplex characterization of autoan-
tibody responses Nat Med 20028295ndash301
21 Yeste A Quintana FJ Antigen microarrays for the study of
autoimmune diseases Clin Chem 2013591036ndash1044
22 Li QZ Xie C Wu T et al Identification of autoantibody
clusters that best predict lupus disease activity using glomer-
ular proteome arrays J Clin Invest 20051153428ndash3439
23 Hueber W Kidd BA Tomooka BH et al Antigen micro-
array profiling of autoantibodies in rheumatoid arthritis
Arthritis Rheum 2005522645ndash2655
24 Goldschmidt Y Sharon E Quintana FJ Cohen IR
Brandt A Adaptive methods for classification of biological
microarray data from multiple experiments Report no
MCS03-07 Rehovot Israel The Arthur and Rochelle Bel-
fer Institute of Mathematics and Computer Science 2003
25 Quintana FJ Getz G Hed G Domany E Cohen IR
Cluster analysis of human autoantibody reactivities in
health and in type 1 diabetes mellitus a bio-informatic
approach to immune complexity J Autoimmun 2003
2165ndash75
10 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
26 Lalive PH Menge T Barman I Cree BA Genain CP
Identification of new serum autoantibodies in neuromye-
litis optica using protein microarrays Neurology 200667
176ndash177
27 Kanter JL Narayana S Ho PP et al Lipid microarrays
identify key mediators of autoimmune brain inflamma-
tion Nat Med 200612138ndash143
28 Quintana FJ Farez MF Izquierdo G Lucas M Cohen IR
Weiner HL Antigen microarrays identify CNS-produced
autoantibodies in RRMS Neurology 201278532ndash539
29 Farez MF Quintana FJ Gandhi R Izquierdo G Lucas M
Weiner HL Toll-like receptor 2 and poly(ADP-ribose)
polymerase 1 promote central nervous system neuroin-
flammation in progressive EAE Nat Immunol 200910
958ndash964
30 Quintana FJ Farez MF Viglietta V et al Antigen micro-
arrays identify unique serum autoantibody signatures in
clinical and pathologic subtypes of multiple sclerosis Proc
Natl Acad Sci USA 200810518889ndash18894
31 Quintana FJ Patel B Yeste A et al Epitope spreading as
an early pathogenic event in pediatric multiple sclerosis
Neurology 2014832219ndash2226
32 Gauthier SA Glanz BI Mandel M Weiner HL A model
for the comprehensive investigation of a chronic autoim-
mune disease the multiple sclerosis CLIMB study Auto-
immun Rev 20065532ndash536
33 Polman CH Reingold SC Edan G et al Diagnostic criteria
for multiple sclerosis 2005 revisions to the ldquoMcDonald
Criteriardquo Ann Neurol 200558840ndash846
34 DellrsquoOglio E Ceccarelli A Glanz BI et al Quantification
of global cerebral atrophy in multiple sclerosis from 3T
MRI using SPM the role of misclassification errors
J Neuroimaging 201525191ndash199
35 Ceccarelli A Jackson JS Tauhid S et al The impact of
lesion in-painting and registration methods on voxel-based
morphometry in detecting regional cerebral gray matter
atrophy in multiple sclerosis AJNR Am J Neuroradiol
2012331579ndash1585
36 Cohen AB Neema M Arora A et al The relationships
among MRI-defined spinal cord involvement brain involve-
ment and disability in multiple sclerosis J Neuroimaging
201222122ndash128
37 Sanfilipo MP Benedict RH Sharma J Weinstock-
Guttman B Bakshi R The relationship between whole
brain volume and disability in multiple sclerosis a com-
parison of normalized gray vs white matter with misclas-
sification correction Neuroimage 2005261068ndash1077
38 Sanfilipo MP Benedict RH Weinstock-Guttman B
Bakshi R Gray and white matter brain atrophy and neuro-
psychological impairment in multiple sclerosis Neurology
200666685ndash692
39 Stankiewicz JM Glanz BI Healy BC et al Brain MRI
lesion load at 15T and 3T versus clinical status in multi-
ple sclerosis J Neuroimaging 201121e50ndashe56
40 Quintana FJ Hagedorn PH Elizur G Merbl Y
Domany E Cohen IR Functional immunomics micro-
array analysis of IgG autoantibody repertoires predicts the
future response of mice to induced diabetes Proc Natl
Acad Sci USA 2004101(suppl 2)14615ndash14621
41 Bermel RA Bakshi R The measurement and clinical
relevance of brain atrophy in multiple sclerosis Lancet
Neurol 20065158ndash170
42 Eikelenboom MJ Petzold A Lazeron RH et al Multiple
sclerosis neurofilament light chain antibodies are corre-
lated to cerebral atrophy Neurology 200360219ndash223
43 Magraner MJ Bosca I Simo-Castello M et al Brain atro-
phy and lesion load are related to CSF lipid-specific IgM
oligoclonal bands in clinically isolated syndromes Neuro-
radiol 2012545ndash12
44 Villar LM Sadaba MC Roldan E et al Intrathecal syn-
thesis of oligoclonal IgM against myelin lipids predicts an
aggressive disease course in MS J Clin Invest 2005115
187ndash194
45 Merbl Y Zucker-Toledano M Quintana FJ Cohen IR
Newborn humans manifest autoantibodies to defined self
molecules detected by antigen microarray informatics
J Clin Invest 2007117712ndash718
46 Sormani MP Arnold DL De Stefano N Treatment effect
on brain atrophy correlates with treatment effect on disa-
bility in multiple sclerosis Ann Neurol 20147543ndash49
47 Tauhid S Neema M Healy BC Weiner HL Bakshi R
MRI phenotypes based on cerebral lesions and atrophy in
patients with multiple sclerosis J Neurol Sci 2014346
250ndash254
48 Gnanapavan S Grant D Morant S et al Biomarker
report from the phase II lamotrigine trial in secondary
progressive MS neurofilament as a surrogate of disease
progression PLoS One 20138e70019
49 Quintana FJ Cohen IR Heat shock proteins as endoge-
nous adjuvants in sterile and septic inflammation
J Immunol 20051752777ndash2782
50 Quintana FJ Cohen IR The HSP60 immune system net-
work Trends Immunol 20113289ndash95
51 Ho PP Kanter JL Johnson AM et al Identification of
naturally occurring fatty acids of the myelin sheath that
resolve neuroinflammation Sci Transl Med 20124137ra73
52 Quintana FJ Yeste A Weiner HL Covacu R Lipids and
lipid-reactive antibodies as biomarkers for multiple sclero-
sis J Neuroimmunol 201224853ndash57
53 Mayo L Trauger SA Blain M et al Regulation of astro-
cyte activation by glycolipids drives chronic CNS inflam-
mation Nat Med 2014201147ndash1156
Neurology Neuroimmunology amp Neuroinflammation 11
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
1
Table e-1 Antigens used
Group Antigen Source
27 kDa Heat Shock Protein Stressgen
HSP 32 kDa Heat Shock Protein Stressgen
40 kDa Heat Shock Protein Stressgen
47 kDa Heat Shock Protein Stressgen
60 kDa Heat Shock Protein Stressgen
60 kDa Heat Shock Protein peptide aa 106ndash125 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 121ndash140 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 136ndash155 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 151ndash170 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 16ndash35 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 166ndash185 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
2
60 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 556ndash573 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
65 kDa Heat Shock Protein M tuberculosis Stressgen
70 kDa Heat Shock Protein Stressgen
70 kDa Heat Shock Protein peptide aa 106ndash125 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 121ndash140 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 136ndash155 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 151ndash170 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 16ndash35 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 166ndash185 Biopolymers Facility Harvard Medical School
3
70 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
4
70 kDa Heat Shock Protein peptide aa 556ndash575 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 571ndash590 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 586ndash605 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 601ndash620 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 616ndash635 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 631ndash640 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
71 kDa Heat Shock Protein M tuberculosis Biopolymers Facility Harvard Medical School
90 kDa Heat Shock Protein Stressgen
GroEL Stressgen
23-cyclic nucleotide 3-phosphodiesterase
peptide aa106-125 Biopolymers Facility Harvard Medical School
CNS
23-cyclic nucleotide 3-phosphodiesterase
peptide aa1-20 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa121-140 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa136-155 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa151-170 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa16-35 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa166-185 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase Biopolymers Facility Harvard Medical School
5
peptide aa181-200
23-cyclic nucleotide 3-phosphodiesterase
peptide aa196-215 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa211-230 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa226-245 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa241-260 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa256-275 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa271-290 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa286-305 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa301-320 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa31-50 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa316-335 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa331-350 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa346-365 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa361-380 Biopolymers Facility Harvard Medical School
6
23-cyclic nucleotide 3-phosphodiesterase
peptide aa376-395 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa391-410 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa406-421 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa46-65 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa61-80 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa76-95 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa91-110 Biopolymers Facility Harvard Medical School
Acetyl Cholinesterase Sigma Aldrich
ADAM-10 Sigma Aldrich
alpha-Cristallin Stressgen
beta-Cristallin Sigma Aldrich
bovine Myelin Basic Protein Sigma Aldrich
Brain Extract I Sigma Aldrich
Brain Extract II Sigma Aldrich
Brain Extract III Sigma Aldrich
Glial Filament Acidic Protein Research Diagnostic
guinea pig Myelin Basic Protein Sigma Aldrich
human Myelin Basic Protein Sigma Aldrich
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 106-125 Biopolymers Facility Harvard Medical School
7
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 121-140 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 136-155 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 151-170 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 166-185 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 181-200 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 91-110 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
106-125 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa 1- Biopolymers Facility Harvard Medical School
8
20
Myelinoligodendrocyte glycoprotein peptide aa
121-140 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
136-155 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
151-170 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
16-35 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
166-185 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
181-200 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
196-215 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
211-230 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
226-247 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
31-50 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
35-55 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
46-65 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
61-80 Biopolymers Facility Harvard Medical School
9
Myelinoligodendrocyte glycoprotein peptide aa
76-95 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
91-110 Biopolymers Facility Harvard Medical School
murine Myelin Basic Protein Sigma Aldrich
Myelin Associated Glycoprotein Sigma Aldrich
Myelin Basic Protein peptide aa 104-123 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 11-30 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 113-132 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 121-138 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 124-142 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 138-147 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 141-161 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 143-168 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 155-178 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 26-35 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 41-60 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 51-70 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 71-92 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 84-94 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 89-101 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 173-186 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 93-112 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 106-125 Biopolymers Facility Harvard Medical School
10
Myelin Protein 2 peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 121-132 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 91-110 Biopolymers Facility Harvard Medical School
Neurofilament 160kd Chemicon
Neurofilament 200kd Chemicon
Neurofilament 68kd Chemicon
Neuronal Enolase Calbiochem
Nicastrin GeneTex
NMDA receptor Novus Biologicals
NOGO Sigma Aldrich
Olygodendrocyte-Specific Protein peptide aa 106ndash
125 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 121ndash
140 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 136ndash
155 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 151ndash
170 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 16ndash
35 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 166ndash Biopolymers Facility Harvard Medical School
11
185
Olygodendrocyte-Specific Protein peptide aa 181ndash
199 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 195ndash
217 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 31ndash
50 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 46ndash
65 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 61ndash
80 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 76ndash
95 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 91ndash
110 Biopolymers Facility Harvard Medical School
Proteolipid Protein Abnova
Proteolipid Protein peptide aa 100-119 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 10-29 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 110-129 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 1-19 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 125-141 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-150 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-154 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 150-163 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 151-173 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 158-166 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 161-180 Biopolymers Facility Harvard Medical School
12
Proteolipid Protein peptide aa 178-191 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 180-199 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 190-209 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 20-39 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 205-220 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 215-232 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-239 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-249 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 248-259 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 250-269 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 265-277 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 35-50 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 40-59 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 50-69 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 65-84 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 80-99 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 91-110 Biopolymers Facility Harvard Medical School
Retinol Binding Protein Sigma Aldrich
S100beta protein Assay Designs
Super Oxide Dismutase Sigma Aldrich
Synuclein beta Sigma Aldrich
Synuclein gamma Sigma Aldrich
Amydgala ProSci Inc
Tissue Amydgala AD ProSci Inc
Brain lysate ProSci Inc
Brain Tissue Membrane ProSci Inc
Cerebellar pedunculus ProSci Inc
13
Cerebral meninges ProSci Inc
Corpus Callosum ProSci Inc
Corpus Callosum AD ProSci Inc
Diencephalon ProSci Inc
Fetal brain ProSci Inc
Frontal lobe ProSci Inc
Frontal lobe AD ProSci Inc
Hippocampus ProSci Inc
Hippocampus AD ProSci Inc
Insula ProSci Inc
Occipital lobe ProSci Inc
Occipital lobe AD ProSci Inc
Olfactory region ProSci Inc
Optic Nerve ProSci Inc
Parietal lobe ProSci Inc
Parietal lobe AD ProSci Inc
Pons ProSci Inc
Pons AD ProSci Inc
Postcentral gyrus ProSci Inc
Postcentral gyrus AD ProSci Inc
Precentral gyrus ProSci Inc
Precentral gyrus AD ProSci Inc
Spinal cord ProSci Inc
Temporal lobe ProSci Inc
Temporal lobe AD ProSci Inc
Thalamus ProSci Inc
Thalamus AD ProSci Inc
14
Amyloid beta Sigma Aldrich
AD
related Amyloid beta 10-20 Sigma Aldrich
Amyloid beta 1-12 Sigma Aldrich
Amyloid beta 12-28 Sigma Aldrich
Amyloid beta 1-23 Sigma Aldrich
Amyloid beta 1-38 Sigma Aldrich
Amyloid beta 17-40 Sigma Aldrich
Amyloid beta 25-35 Sigma Aldrich
Amyloid beta 34-42 Sigma Aldrich
Amyloid bri protein precursor 227 Sigma Aldrich
Amyloid DAN Protein Fragment 1-34 Sigma Aldrich
Amyloid Precursor Protein Sigma Aldrich
Amyloid protein no AB component Sigma Aldrich
Secreted amyloid precursor protein (SAP) beta Sigma Aldrich
Tau isoform variant 0N3R Sigma Aldrich
Tau isoform variant 1N3R Sigma Aldrich
Tau isoform variant 0N4R Sigma Aldrich
Tau isoform variant 2N3R Sigma Aldrich
Tau phospho Ser412 Sigma Aldrich
Tau phospho Ser441 Sigma Aldrich
Tau phospho Thr181 Sigma Aldrich
Tau Protein human EMD Biosciences
(plusmn)9-HODE Cayman Chemical
Lipids
1 Palmitoyl-2-(5oxo-Valeroyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
15a-hydroxycholestene Avanti Polar Lipids
15
15-ketocholestane Avanti Polar Lipids
15-ketocholestene Avanti Polar Lipids
1-Palmitoil-2-(9oxo-Nonanoyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Azelaoyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Glutaroyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
5 α-cholestane-3 β15 α-diol Avanti Polar Lipids
9(S)-HODE Cayman Chemical
Asialoganglioside-GM1 Sigma Aldrich
Asialoganglioside-GM2 Sigma Aldrich
Brain ceramides Avanti Polar Lipids
Brain D-erythrosphingosine Avanti Polar Lipids
Brain lysophosphatidylethanolamine Avanti Polar Lipids
Brain L-α-lysophosphatidylserine Avanti Polar Lipids
Brain L-α-phosphatidylcholine Avanti Polar Lipids
Brain L-α-phosphatidyl-ethanolamine Avanti Polar Lipids
Brain L-α-phosphatidylserine Avanti Polar Lipids
Brain polar lipid extract Avanti Polar Lipids
Brain sphingomyelin Avanti Polar Lipids
Brain sulfatide Avanti Polar Lipids
Brain total lipid extract Avanti Polar Lipids
Cardiolipin Sigma Aldrich
Ceramide Sigma Aldrich
Ceramide 1-phosphate Sigma Aldrich
Cholesterol Sigma Aldrich
16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-

enrichment in lipid-reactive IgM in the group ofantibodies associated with increased diseasepathogenesis (not shown)
An index of serum IgG reactivity to lipids is associated
with MRI measures of disease severity To further inves-tigate the association between serum IgG reactivityto lipids and MRI measures of disease severity wecalculated an IgG anti-lipid index for each patientcorresponding to the information on each lipid-specific antibody listed in table 2 normalized by thestrength of its correlation with that MRI measureunder investigation The antibody reactivities tolipids used to calculate the lipid index are shown in
figure 3A With this approach we calculated one indexfor each of the MRI measures analyzed in this studyWe found a significant correlation between each IgGlipid antibody index and BPF GMF and T2LV(figure 3B) no significant correlation was found withWMF Moreover no significant correlations werefound between BPF WMF GMF and T2LVmeasures and anti-lipid antibody indexes based onIgM reactivity (not shown)
Finally we evaluated the performance of the lipidantibody indexes linked to BPF GMF and T2LV onan additional set of independent MS samples (valida-tion set table 1) We detected a similar trend to theone detected in the discovery set with regard to the
Figure 1 Antibody reactivity in serum is associated with decreased brain volume
Heatmap in which each column represents the mean immunoglobulin G antibody reactivity in a serum sample from a patient with multiple sclerosis sortedaccording to the whole brain parenchymal fraction (BPF) (indicated at the topmdasha lower BPF indicates more brain atrophy) and each row represents theantibody reactivity to an antigen according to the colorimetric scale shown The antibody reactivities included in this heatmap are listed in table 2
6 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
Figure 2 Association of serum IgG reactivity with MRI measures of disease severity
(A) Heatmap in which each column represents an MRI measure of either atrophy (BPF GMF WMF) or lesions (T2LV) andeach row represents the correlation to IgG serum antibody reactivity according to a colorimetric scale (B) Frequency oflipid-reactive antibodies linked to higher or lower MRI disease severity BPF 5 whole brain parenchymal fraction GMF 5
global cerebral gray matter fraction IgG 5 immunoglobulin G T2LV 5 cerebral T2 (fluid-attenuated inversion recovery)hyperintense lesion volume WMF 5 global cerebral white matter fraction
Neurology Neuroimmunology amp Neuroinflammation 7
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
correlation between BPF and GMF and their respec-tive lipid indexes and we validated the correlationbetween T2LV and its lipid index (figure 3C)
DISCUSSION Previous studies have investigated theassociation of MRI with immune activity in MS13ndash19
These studies have included cellular immune measures
Figure 3 Correlation of lipid indexes to MRI measures of disease severity in multiple sclerosis
(A) Heatmap inwhich each column represents anMRImeasure and each row represents the correlation to immunoglobulin G serumantibody reactivity to lipids accord-ing to a colorimetric scale (B) Scatter plots depicting the correlation between each lipid index andMRImeasures (BPF GMF and T2LV) in the discovery set (C) Scatterplots depicting the correlation between each lipid index andMRImeasures (BPF GMF and T2LV) in the validation set BPF5whole brain parenchymal fraction GMF5
cerebral gray matter fraction (see methods section for details) T2LV 5 cerebral T2 hyperintense lesion volume WMF 5 global cerebral white matter fraction
8 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
as well as oligoclonal bands or neurofilament-specificantibodies in CSF42ndash44 In the present study weanalyzed the association between serum antibodyprofiles detected with antigen microarrays and MRImeasures of disease including cerebral atrophy andT2 hyperintense lesions We found that specificantibody patterns are associated with different aspectsof MRI-defined disease pathology Our major findingwas that increased reactivity to lipids is associated withdifferent aspects of brain MRI measures of diseaseseverity Of note the specific set of lipids associatedwith atrophy differed from those associated withlesions Taken together these data suggest that anti-lipid antibodies in serum are related to MRI measuresof disease Our findings highlight the important role oflipid and lipid-specific immunity in the pathogenesis ofMS
Although a potential limitation of these studies isthe relatively small number of samples analyzed wehave validated our findings in an independent valida-tion cohort strengthening their significance In addi-tion further studies in large cohorts from patientsaffected by non-MS neurodegenerative diseases andhealthy controls may indicate whether the antibodyreactivities and lipid indexes identified in these stud-ies are exclusive to MS or are associated with addi-tional biological processes Indeed some of theseantibody reactivities contributing to the immune sig-natures described in this work have been found innewborns45
Different sets of antibody reactivities were associ-ated with GMF WMF BPF and T2LV The differ-ent antibody link between lesions and atrophy is inkeeping with the long-held view that brain atrophyin patients with MS is multifactorial and is onlyweakly related to lesions41 This has been underscoredin recent studies showing the complementary infor-mation obtained by combining lesion and atrophymeasures4647
Among the cerebral atrophy measures there was astriking overlap between those antibodies associatedwith GMF and BPF which separated them fromWMF results These findings are in keeping with pre-vious observations that whole brain atrophy is domi-nated by GM loss3741 In the early stages of MS thisGM atrophy selectively involves the deep GM nuclei34
Thus the similarities observed in the autoantibodieslinked to GMF and BPF might reflect the dominantcontribution and colinearity of GM atrophy to totalbrain atrophy The set of antibodies linked to GMFmay be the most relevant given that MRI studies haveshown that GM volume is more closely linked to phys-ical disability37 and cognitive impairment38 than areWM volume or lesion measures
Using antigen microarrays antibody reactivity tolipids has been detected in the CSF and serum of
patients with MS at different stages of the diseaseand such antibodies in the CSF have been linkedto disease progression and MRI involvement44
Although several mechanisms are thought to drivebrain inflammation and atrophy in MS46 the connec-tion between these mechanisms and the antibodiesdetected with our antigen microarrays is yet unknownOne possibility is that the serum autoantibodies iden-tified in our studies are not pathogenic and reflect theresult of immunization against self-antigens releasedfrom the CNS during the course of the disease Indeedneurofilament is released from damaged axons duringthe course of MS and both neurofilament lightchains48 and antibodies against it42 have been linkedto MRI measures of disease Moreover heat shockproteins are upregulated in different cell types duringinflammation and have been shown to have an impor-tant role as immunomodulators when released to theextracellular medium4950 Thus it is possible that heatshock proteinndashreactive antibodies reflect changes in theproduction and release of these immunomodulatorsduring the course of disease pathogenesis Similarlybioactive lipids and their products are released fromdamaged myelin and lipid-specific antibodies havebeen detected in patients with MS27293051 Lipids havean important role in the immune response both asbioactive molecules with immunomodulatory proper-ties and also as targets of the adaptive immuneresponse52 Moreover lipids have significant effectson the murine model of MS experimental autoim-mune encephalomyelitis2729305153 Indeed we recentlyfound that the glycolipid lactosylceramide activates abroad set of biological processes in astrocytes promot-ing neurodegeneration and inflammation53 Thus thelipid-reactive antibodies detected in this work mayreflect the release of myelin lipids in the context ofMS pathogenesis andor may directly contribute toimmune-mediated damage in CNS tissue Of notelipid-reactive antibodies are highly cross-reactive52 Inaddition the lipid-reactive antibodies detected in thiswork as associated with MRI measures of disease activ-ity were of the IgG class These are important points toconsider when evaluating a potential role of lipid-reactive antibodies inMS pathogenesis Further studiesare warranted to investigate whether bioactive lipidsoffer new therapeutic targets in MS
We have found that unique patterns of immunereactivity determined with antibody arrays are associ-ated with specific MRI measures of disease severityThese patterns agree with the interpretation that dif-ferent pathogenic mechanisms drive diverse diseaseprocesses reflected by these MRI measures These pat-terns also suggest a predominant role for lipids andlipid-specific immunity in MS pathology Furtherstudies are warranted to determine whether the earlyappearance of anti-lipid antibodies predicts the
Neurology Neuroimmunology amp Neuroinflammation 9
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
subsequent development of clinical and MRI-defineddisease worsening
AUTHOR CONTRIBUTIONSRohit Bakshi draftingrevising the manuscript study concept or design
analysis or interpretation of data acquisition of data study supervision
obtaining funding Ada Yeste study concept or design acquisition of
data Bonny Patel analysis or interpretation of data statistical analysis
Shahamat Tauhid analysis or interpretation of data Subhash Tummala
analysis or interpretation of data Roya Rahbari analysis or interpretation
of data statistical analysis Renxin Chu analysis or interpretation of data
Keren Regev analysis or interpretation of data acquisition of data
Pia Kivisaumlkk draftingrevising the manuscript study concept or design
contribution of vital reagentstoolspatients acquisition of data study
supervision Howard L Weiner study concept or design obtaining fund-
ing Francisco J Quintana draftingrevising the manuscript study con-
cept or design contribution of vital reagentstoolspatients study
supervision obtaining funding
ACKNOWLEDGMENTThe authors thank Dr Mohit Neema Dr Antonella Ceccarelli and
Dr Ashish Arora for valuable assistance at the early stages of this project
STUDY FUNDINGThis work was supported in part by grants to FJQ and HLW from
EMD Serono RG4111A1 and JF2161-A-5 from the National Multiple
Sclerosis Society PA0069 from the International Progressive MS Alli-
ance and by grants from the NIH (5R01NS055083-04) and NMSS
(RG3798A2) to RB
DISCLOSUREFJ Quintana serves on the editorial board for Systems Biomedicine
Inmunologia American Journal of Clinical and Experimental Immunology
is an associate editor for Immunology (UK) is an advisory board member
for Seminars in Immunopathology received research support from Harvard
Medical School BADERC NMSS A Yeste and B Patel report no
disclosures S Tauhid is managing editor for Journal of Neuroimaging
S Tummala R Rahbari R Chu and K Regev report no disclosures
P Kivisaumlkk received research support from EMD Serono R Bakshi is
editor-in-chief for Journal of Neuroimaging received consulting fees from
AbbVie Alkermes Biogen Novartis Questcor received research support
from Biogen EMD-Serono Novartis Sanofi-Genzyme Teva his spouse
holds stock in Biogen Inc H Weiner served on the advisory board
for The Guthy-Jackson Charitable Foundation Teva Pharmaceuticals
Industries Ltd Biogen Idec Novartis Sanofi-Aventis has consulted for
Therapix Bioven Novartis Serono Teva Sanofi received research support
from National Multiple Sclerosis Society Go to Neurologyorgnn for full
disclosure forms
Received July 27 2015 Accepted in final form December 8 2015
REFERENCES1 Weiner HL The challenge of multiple sclerosis how do
we cure a chronic heterogeneous disease Ann Neurol
200965239ndash248
2 Bakshi R Thompson AJ Rocca MA et al MRI in mul-
tiple sclerosis current status and future prospects Lancet
Neurol 20087615ndash625
3 Ceccarelli A Bakshi R Neema M MRI in multiple scle-
rosis a review of the current literature Curr Opin Neurol
201225402ndash409
4 Filippi M Rocca MA Barkhof F et al Association
between pathological and MRI findings in multiple scle-
rosis Lancet Neurol 201211349ndash360
5 Geurts JJ Calabrese M Fisher E Rudick RA Measure-
ment and clinical effect of grey matter pathology in mul-
tiple sclerosis Lancet Neurol 2012111082ndash1092
6 Klaver R De Vries HE Schenk GJ Geurts JJ Grey mat-
ter damage in multiple sclerosis a pathology perspective
Prion 2013766ndash75
7 Khoury S Bakshi R Cerebral pseudoatrophy or real atro-
phy after therapy in multiple sclerosis Ann Neurol 2010
68778ndash779
8 Hauser SL Chan JR Oksenberg JR Multiple sclerosis
prospects and promise Ann Neurol 201374317ndash327
9 McFarland HF Martin R Multiple sclerosis a complicated
picture of autoimmunity Nat Immunol 20078913ndash919
10 Nylander A Hafler DA Multiple sclerosis J Clin Invest
20121221180ndash1188
11 Sospedra M Martin R Immunology of multiple sclerosis
Ann Rev Immunol 200523683ndash747
12 Steinman L Immunology of relapse and remission in mul-
tiple sclerosis Ann Rev Immunol 201432257ndash281
13 Khoury SJ Guttmann CR Orav EJ Kikinis R Jolesz FA
Weiner HL Changes in activated T cells in the blood
correlate with disease activity in multiple sclerosis Arch
Neurol 2000571183ndash1189
14 Khoury SJ Orav EJ Guttmann CR Kikinis R Jolesz FA
Weiner HL Changes in serum levels of ICAM and TNF-
R correlate with disease activity in multiple sclerosis Neu-
rology 199953758ndash764
15 Laplaud DA Berthelot L Miqueu P et al Serial blood T
cell repertoire alterations in multiple sclerosis patients cor-
relation with clinical and MRI parameters J Neuroimmunol
2006177151ndash160
16 Makhlouf K Weiner HL Khoury SJ Increased percentage
of IL-121 monocytes in the blood correlates with the
presence of active MRI lesions in MS J Neuroimmunol
2001119145ndash149
17 Prat A Biernacki K Saroli T et al Kinin B1 receptor expres-
sion on multiple sclerosis mononuclear cells correlation with
magnetic resonance imaging T2-weighted lesion volume and
clinical disability Arch Neurol 200562795ndash800
18 Rinaldi L Gallo P Calabrese M et al Longitudinal anal-
ysis of immune cell phenotypes in early stage multiple
sclerosis distinctive patterns characterize MRI-active pa-
tients Brain 20061291993ndash2007
19 Tortorella P Lagana MM Saresella M et al Determinants
of disability in multiple sclerosis an immunological and
MRI study Biomed Res Int 20142014875768
20 Robinson WH DiGennaro C Hueber W et al Autoan-
tigen microarrays for multiplex characterization of autoan-
tibody responses Nat Med 20028295ndash301
21 Yeste A Quintana FJ Antigen microarrays for the study of
autoimmune diseases Clin Chem 2013591036ndash1044
22 Li QZ Xie C Wu T et al Identification of autoantibody
clusters that best predict lupus disease activity using glomer-
ular proteome arrays J Clin Invest 20051153428ndash3439
23 Hueber W Kidd BA Tomooka BH et al Antigen micro-
array profiling of autoantibodies in rheumatoid arthritis
Arthritis Rheum 2005522645ndash2655
24 Goldschmidt Y Sharon E Quintana FJ Cohen IR
Brandt A Adaptive methods for classification of biological
microarray data from multiple experiments Report no
MCS03-07 Rehovot Israel The Arthur and Rochelle Bel-
fer Institute of Mathematics and Computer Science 2003
25 Quintana FJ Getz G Hed G Domany E Cohen IR
Cluster analysis of human autoantibody reactivities in
health and in type 1 diabetes mellitus a bio-informatic
approach to immune complexity J Autoimmun 2003
2165ndash75
10 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
26 Lalive PH Menge T Barman I Cree BA Genain CP
Identification of new serum autoantibodies in neuromye-
litis optica using protein microarrays Neurology 200667
176ndash177
27 Kanter JL Narayana S Ho PP et al Lipid microarrays
identify key mediators of autoimmune brain inflamma-
tion Nat Med 200612138ndash143
28 Quintana FJ Farez MF Izquierdo G Lucas M Cohen IR
Weiner HL Antigen microarrays identify CNS-produced
autoantibodies in RRMS Neurology 201278532ndash539
29 Farez MF Quintana FJ Gandhi R Izquierdo G Lucas M
Weiner HL Toll-like receptor 2 and poly(ADP-ribose)
polymerase 1 promote central nervous system neuroin-
flammation in progressive EAE Nat Immunol 200910
958ndash964
30 Quintana FJ Farez MF Viglietta V et al Antigen micro-
arrays identify unique serum autoantibody signatures in
clinical and pathologic subtypes of multiple sclerosis Proc
Natl Acad Sci USA 200810518889ndash18894
31 Quintana FJ Patel B Yeste A et al Epitope spreading as
an early pathogenic event in pediatric multiple sclerosis
Neurology 2014832219ndash2226
32 Gauthier SA Glanz BI Mandel M Weiner HL A model
for the comprehensive investigation of a chronic autoim-
mune disease the multiple sclerosis CLIMB study Auto-
immun Rev 20065532ndash536
33 Polman CH Reingold SC Edan G et al Diagnostic criteria
for multiple sclerosis 2005 revisions to the ldquoMcDonald
Criteriardquo Ann Neurol 200558840ndash846
34 DellrsquoOglio E Ceccarelli A Glanz BI et al Quantification
of global cerebral atrophy in multiple sclerosis from 3T
MRI using SPM the role of misclassification errors
J Neuroimaging 201525191ndash199
35 Ceccarelli A Jackson JS Tauhid S et al The impact of
lesion in-painting and registration methods on voxel-based
morphometry in detecting regional cerebral gray matter
atrophy in multiple sclerosis AJNR Am J Neuroradiol
2012331579ndash1585
36 Cohen AB Neema M Arora A et al The relationships
among MRI-defined spinal cord involvement brain involve-
ment and disability in multiple sclerosis J Neuroimaging
201222122ndash128
37 Sanfilipo MP Benedict RH Sharma J Weinstock-
Guttman B Bakshi R The relationship between whole
brain volume and disability in multiple sclerosis a com-
parison of normalized gray vs white matter with misclas-
sification correction Neuroimage 2005261068ndash1077
38 Sanfilipo MP Benedict RH Weinstock-Guttman B
Bakshi R Gray and white matter brain atrophy and neuro-
psychological impairment in multiple sclerosis Neurology
200666685ndash692
39 Stankiewicz JM Glanz BI Healy BC et al Brain MRI
lesion load at 15T and 3T versus clinical status in multi-
ple sclerosis J Neuroimaging 201121e50ndashe56
40 Quintana FJ Hagedorn PH Elizur G Merbl Y
Domany E Cohen IR Functional immunomics micro-
array analysis of IgG autoantibody repertoires predicts the
future response of mice to induced diabetes Proc Natl
Acad Sci USA 2004101(suppl 2)14615ndash14621
41 Bermel RA Bakshi R The measurement and clinical
relevance of brain atrophy in multiple sclerosis Lancet
Neurol 20065158ndash170
42 Eikelenboom MJ Petzold A Lazeron RH et al Multiple
sclerosis neurofilament light chain antibodies are corre-
lated to cerebral atrophy Neurology 200360219ndash223
43 Magraner MJ Bosca I Simo-Castello M et al Brain atro-
phy and lesion load are related to CSF lipid-specific IgM
oligoclonal bands in clinically isolated syndromes Neuro-
radiol 2012545ndash12
44 Villar LM Sadaba MC Roldan E et al Intrathecal syn-
thesis of oligoclonal IgM against myelin lipids predicts an
aggressive disease course in MS J Clin Invest 2005115
187ndash194
45 Merbl Y Zucker-Toledano M Quintana FJ Cohen IR
Newborn humans manifest autoantibodies to defined self
molecules detected by antigen microarray informatics
J Clin Invest 2007117712ndash718
46 Sormani MP Arnold DL De Stefano N Treatment effect
on brain atrophy correlates with treatment effect on disa-
bility in multiple sclerosis Ann Neurol 20147543ndash49
47 Tauhid S Neema M Healy BC Weiner HL Bakshi R
MRI phenotypes based on cerebral lesions and atrophy in
patients with multiple sclerosis J Neurol Sci 2014346
250ndash254
48 Gnanapavan S Grant D Morant S et al Biomarker
report from the phase II lamotrigine trial in secondary
progressive MS neurofilament as a surrogate of disease
progression PLoS One 20138e70019
49 Quintana FJ Cohen IR Heat shock proteins as endoge-
nous adjuvants in sterile and septic inflammation
J Immunol 20051752777ndash2782
50 Quintana FJ Cohen IR The HSP60 immune system net-
work Trends Immunol 20113289ndash95
51 Ho PP Kanter JL Johnson AM et al Identification of
naturally occurring fatty acids of the myelin sheath that
resolve neuroinflammation Sci Transl Med 20124137ra73
52 Quintana FJ Yeste A Weiner HL Covacu R Lipids and
lipid-reactive antibodies as biomarkers for multiple sclero-
sis J Neuroimmunol 201224853ndash57
53 Mayo L Trauger SA Blain M et al Regulation of astro-
cyte activation by glycolipids drives chronic CNS inflam-
mation Nat Med 2014201147ndash1156
Neurology Neuroimmunology amp Neuroinflammation 11
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
1
Table e-1 Antigens used
Group Antigen Source
27 kDa Heat Shock Protein Stressgen
HSP 32 kDa Heat Shock Protein Stressgen
40 kDa Heat Shock Protein Stressgen
47 kDa Heat Shock Protein Stressgen
60 kDa Heat Shock Protein Stressgen
60 kDa Heat Shock Protein peptide aa 106ndash125 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 121ndash140 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 136ndash155 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 151ndash170 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 16ndash35 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 166ndash185 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
2
60 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 556ndash573 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
65 kDa Heat Shock Protein M tuberculosis Stressgen
70 kDa Heat Shock Protein Stressgen
70 kDa Heat Shock Protein peptide aa 106ndash125 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 121ndash140 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 136ndash155 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 151ndash170 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 16ndash35 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 166ndash185 Biopolymers Facility Harvard Medical School
3
70 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
4
70 kDa Heat Shock Protein peptide aa 556ndash575 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 571ndash590 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 586ndash605 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 601ndash620 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 616ndash635 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 631ndash640 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
71 kDa Heat Shock Protein M tuberculosis Biopolymers Facility Harvard Medical School
90 kDa Heat Shock Protein Stressgen
GroEL Stressgen
23-cyclic nucleotide 3-phosphodiesterase
peptide aa106-125 Biopolymers Facility Harvard Medical School
CNS
23-cyclic nucleotide 3-phosphodiesterase
peptide aa1-20 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa121-140 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa136-155 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa151-170 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa16-35 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa166-185 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase Biopolymers Facility Harvard Medical School
5
peptide aa181-200
23-cyclic nucleotide 3-phosphodiesterase
peptide aa196-215 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa211-230 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa226-245 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa241-260 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa256-275 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa271-290 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa286-305 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa301-320 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa31-50 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa316-335 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa331-350 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa346-365 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa361-380 Biopolymers Facility Harvard Medical School
6
23-cyclic nucleotide 3-phosphodiesterase
peptide aa376-395 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa391-410 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa406-421 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa46-65 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa61-80 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa76-95 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa91-110 Biopolymers Facility Harvard Medical School
Acetyl Cholinesterase Sigma Aldrich
ADAM-10 Sigma Aldrich
alpha-Cristallin Stressgen
beta-Cristallin Sigma Aldrich
bovine Myelin Basic Protein Sigma Aldrich
Brain Extract I Sigma Aldrich
Brain Extract II Sigma Aldrich
Brain Extract III Sigma Aldrich
Glial Filament Acidic Protein Research Diagnostic
guinea pig Myelin Basic Protein Sigma Aldrich
human Myelin Basic Protein Sigma Aldrich
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 106-125 Biopolymers Facility Harvard Medical School
7
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 121-140 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 136-155 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 151-170 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 166-185 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 181-200 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 91-110 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
106-125 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa 1- Biopolymers Facility Harvard Medical School
8
20
Myelinoligodendrocyte glycoprotein peptide aa
121-140 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
136-155 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
151-170 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
16-35 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
166-185 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
181-200 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
196-215 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
211-230 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
226-247 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
31-50 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
35-55 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
46-65 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
61-80 Biopolymers Facility Harvard Medical School
9
Myelinoligodendrocyte glycoprotein peptide aa
76-95 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
91-110 Biopolymers Facility Harvard Medical School
murine Myelin Basic Protein Sigma Aldrich
Myelin Associated Glycoprotein Sigma Aldrich
Myelin Basic Protein peptide aa 104-123 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 11-30 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 113-132 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 121-138 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 124-142 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 138-147 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 141-161 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 143-168 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 155-178 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 26-35 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 41-60 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 51-70 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 71-92 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 84-94 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 89-101 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 173-186 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 93-112 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 106-125 Biopolymers Facility Harvard Medical School
10
Myelin Protein 2 peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 121-132 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 91-110 Biopolymers Facility Harvard Medical School
Neurofilament 160kd Chemicon
Neurofilament 200kd Chemicon
Neurofilament 68kd Chemicon
Neuronal Enolase Calbiochem
Nicastrin GeneTex
NMDA receptor Novus Biologicals
NOGO Sigma Aldrich
Olygodendrocyte-Specific Protein peptide aa 106ndash
125 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 121ndash
140 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 136ndash
155 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 151ndash
170 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 16ndash
35 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 166ndash Biopolymers Facility Harvard Medical School
11
185
Olygodendrocyte-Specific Protein peptide aa 181ndash
199 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 195ndash
217 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 31ndash
50 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 46ndash
65 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 61ndash
80 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 76ndash
95 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 91ndash
110 Biopolymers Facility Harvard Medical School
Proteolipid Protein Abnova
Proteolipid Protein peptide aa 100-119 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 10-29 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 110-129 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 1-19 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 125-141 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-150 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-154 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 150-163 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 151-173 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 158-166 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 161-180 Biopolymers Facility Harvard Medical School
12
Proteolipid Protein peptide aa 178-191 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 180-199 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 190-209 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 20-39 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 205-220 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 215-232 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-239 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-249 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 248-259 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 250-269 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 265-277 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 35-50 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 40-59 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 50-69 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 65-84 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 80-99 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 91-110 Biopolymers Facility Harvard Medical School
Retinol Binding Protein Sigma Aldrich
S100beta protein Assay Designs
Super Oxide Dismutase Sigma Aldrich
Synuclein beta Sigma Aldrich
Synuclein gamma Sigma Aldrich
Amydgala ProSci Inc
Tissue Amydgala AD ProSci Inc
Brain lysate ProSci Inc
Brain Tissue Membrane ProSci Inc
Cerebellar pedunculus ProSci Inc
13
Cerebral meninges ProSci Inc
Corpus Callosum ProSci Inc
Corpus Callosum AD ProSci Inc
Diencephalon ProSci Inc
Fetal brain ProSci Inc
Frontal lobe ProSci Inc
Frontal lobe AD ProSci Inc
Hippocampus ProSci Inc
Hippocampus AD ProSci Inc
Insula ProSci Inc
Occipital lobe ProSci Inc
Occipital lobe AD ProSci Inc
Olfactory region ProSci Inc
Optic Nerve ProSci Inc
Parietal lobe ProSci Inc
Parietal lobe AD ProSci Inc
Pons ProSci Inc
Pons AD ProSci Inc
Postcentral gyrus ProSci Inc
Postcentral gyrus AD ProSci Inc
Precentral gyrus ProSci Inc
Precentral gyrus AD ProSci Inc
Spinal cord ProSci Inc
Temporal lobe ProSci Inc
Temporal lobe AD ProSci Inc
Thalamus ProSci Inc
Thalamus AD ProSci Inc
14
Amyloid beta Sigma Aldrich
AD
related Amyloid beta 10-20 Sigma Aldrich
Amyloid beta 1-12 Sigma Aldrich
Amyloid beta 12-28 Sigma Aldrich
Amyloid beta 1-23 Sigma Aldrich
Amyloid beta 1-38 Sigma Aldrich
Amyloid beta 17-40 Sigma Aldrich
Amyloid beta 25-35 Sigma Aldrich
Amyloid beta 34-42 Sigma Aldrich
Amyloid bri protein precursor 227 Sigma Aldrich
Amyloid DAN Protein Fragment 1-34 Sigma Aldrich
Amyloid Precursor Protein Sigma Aldrich
Amyloid protein no AB component Sigma Aldrich
Secreted amyloid precursor protein (SAP) beta Sigma Aldrich
Tau isoform variant 0N3R Sigma Aldrich
Tau isoform variant 1N3R Sigma Aldrich
Tau isoform variant 0N4R Sigma Aldrich
Tau isoform variant 2N3R Sigma Aldrich
Tau phospho Ser412 Sigma Aldrich
Tau phospho Ser441 Sigma Aldrich
Tau phospho Thr181 Sigma Aldrich
Tau Protein human EMD Biosciences
(plusmn)9-HODE Cayman Chemical
Lipids
1 Palmitoyl-2-(5oxo-Valeroyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
15a-hydroxycholestene Avanti Polar Lipids
15
15-ketocholestane Avanti Polar Lipids
15-ketocholestene Avanti Polar Lipids
1-Palmitoil-2-(9oxo-Nonanoyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Azelaoyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Glutaroyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
5 α-cholestane-3 β15 α-diol Avanti Polar Lipids
9(S)-HODE Cayman Chemical
Asialoganglioside-GM1 Sigma Aldrich
Asialoganglioside-GM2 Sigma Aldrich
Brain ceramides Avanti Polar Lipids
Brain D-erythrosphingosine Avanti Polar Lipids
Brain lysophosphatidylethanolamine Avanti Polar Lipids
Brain L-α-lysophosphatidylserine Avanti Polar Lipids
Brain L-α-phosphatidylcholine Avanti Polar Lipids
Brain L-α-phosphatidyl-ethanolamine Avanti Polar Lipids
Brain L-α-phosphatidylserine Avanti Polar Lipids
Brain polar lipid extract Avanti Polar Lipids
Brain sphingomyelin Avanti Polar Lipids
Brain sulfatide Avanti Polar Lipids
Brain total lipid extract Avanti Polar Lipids
Cardiolipin Sigma Aldrich
Ceramide Sigma Aldrich
Ceramide 1-phosphate Sigma Aldrich
Cholesterol Sigma Aldrich
16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-

Figure 2 Association of serum IgG reactivity with MRI measures of disease severity
(A) Heatmap in which each column represents an MRI measure of either atrophy (BPF GMF WMF) or lesions (T2LV) andeach row represents the correlation to IgG serum antibody reactivity according to a colorimetric scale (B) Frequency oflipid-reactive antibodies linked to higher or lower MRI disease severity BPF 5 whole brain parenchymal fraction GMF 5
global cerebral gray matter fraction IgG 5 immunoglobulin G T2LV 5 cerebral T2 (fluid-attenuated inversion recovery)hyperintense lesion volume WMF 5 global cerebral white matter fraction
Neurology Neuroimmunology amp Neuroinflammation 7
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
correlation between BPF and GMF and their respec-tive lipid indexes and we validated the correlationbetween T2LV and its lipid index (figure 3C)
DISCUSSION Previous studies have investigated theassociation of MRI with immune activity in MS13ndash19
These studies have included cellular immune measures
Figure 3 Correlation of lipid indexes to MRI measures of disease severity in multiple sclerosis
(A) Heatmap inwhich each column represents anMRImeasure and each row represents the correlation to immunoglobulin G serumantibody reactivity to lipids accord-ing to a colorimetric scale (B) Scatter plots depicting the correlation between each lipid index andMRImeasures (BPF GMF and T2LV) in the discovery set (C) Scatterplots depicting the correlation between each lipid index andMRImeasures (BPF GMF and T2LV) in the validation set BPF5whole brain parenchymal fraction GMF5
cerebral gray matter fraction (see methods section for details) T2LV 5 cerebral T2 hyperintense lesion volume WMF 5 global cerebral white matter fraction
8 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
as well as oligoclonal bands or neurofilament-specificantibodies in CSF42ndash44 In the present study weanalyzed the association between serum antibodyprofiles detected with antigen microarrays and MRImeasures of disease including cerebral atrophy andT2 hyperintense lesions We found that specificantibody patterns are associated with different aspectsof MRI-defined disease pathology Our major findingwas that increased reactivity to lipids is associated withdifferent aspects of brain MRI measures of diseaseseverity Of note the specific set of lipids associatedwith atrophy differed from those associated withlesions Taken together these data suggest that anti-lipid antibodies in serum are related to MRI measuresof disease Our findings highlight the important role oflipid and lipid-specific immunity in the pathogenesis ofMS
Although a potential limitation of these studies isthe relatively small number of samples analyzed wehave validated our findings in an independent valida-tion cohort strengthening their significance In addi-tion further studies in large cohorts from patientsaffected by non-MS neurodegenerative diseases andhealthy controls may indicate whether the antibodyreactivities and lipid indexes identified in these stud-ies are exclusive to MS or are associated with addi-tional biological processes Indeed some of theseantibody reactivities contributing to the immune sig-natures described in this work have been found innewborns45
Different sets of antibody reactivities were associ-ated with GMF WMF BPF and T2LV The differ-ent antibody link between lesions and atrophy is inkeeping with the long-held view that brain atrophyin patients with MS is multifactorial and is onlyweakly related to lesions41 This has been underscoredin recent studies showing the complementary infor-mation obtained by combining lesion and atrophymeasures4647
Among the cerebral atrophy measures there was astriking overlap between those antibodies associatedwith GMF and BPF which separated them fromWMF results These findings are in keeping with pre-vious observations that whole brain atrophy is domi-nated by GM loss3741 In the early stages of MS thisGM atrophy selectively involves the deep GM nuclei34
Thus the similarities observed in the autoantibodieslinked to GMF and BPF might reflect the dominantcontribution and colinearity of GM atrophy to totalbrain atrophy The set of antibodies linked to GMFmay be the most relevant given that MRI studies haveshown that GM volume is more closely linked to phys-ical disability37 and cognitive impairment38 than areWM volume or lesion measures
Using antigen microarrays antibody reactivity tolipids has been detected in the CSF and serum of
patients with MS at different stages of the diseaseand such antibodies in the CSF have been linkedto disease progression and MRI involvement44
Although several mechanisms are thought to drivebrain inflammation and atrophy in MS46 the connec-tion between these mechanisms and the antibodiesdetected with our antigen microarrays is yet unknownOne possibility is that the serum autoantibodies iden-tified in our studies are not pathogenic and reflect theresult of immunization against self-antigens releasedfrom the CNS during the course of the disease Indeedneurofilament is released from damaged axons duringthe course of MS and both neurofilament lightchains48 and antibodies against it42 have been linkedto MRI measures of disease Moreover heat shockproteins are upregulated in different cell types duringinflammation and have been shown to have an impor-tant role as immunomodulators when released to theextracellular medium4950 Thus it is possible that heatshock proteinndashreactive antibodies reflect changes in theproduction and release of these immunomodulatorsduring the course of disease pathogenesis Similarlybioactive lipids and their products are released fromdamaged myelin and lipid-specific antibodies havebeen detected in patients with MS27293051 Lipids havean important role in the immune response both asbioactive molecules with immunomodulatory proper-ties and also as targets of the adaptive immuneresponse52 Moreover lipids have significant effectson the murine model of MS experimental autoim-mune encephalomyelitis2729305153 Indeed we recentlyfound that the glycolipid lactosylceramide activates abroad set of biological processes in astrocytes promot-ing neurodegeneration and inflammation53 Thus thelipid-reactive antibodies detected in this work mayreflect the release of myelin lipids in the context ofMS pathogenesis andor may directly contribute toimmune-mediated damage in CNS tissue Of notelipid-reactive antibodies are highly cross-reactive52 Inaddition the lipid-reactive antibodies detected in thiswork as associated with MRI measures of disease activ-ity were of the IgG class These are important points toconsider when evaluating a potential role of lipid-reactive antibodies inMS pathogenesis Further studiesare warranted to investigate whether bioactive lipidsoffer new therapeutic targets in MS
We have found that unique patterns of immunereactivity determined with antibody arrays are associ-ated with specific MRI measures of disease severityThese patterns agree with the interpretation that dif-ferent pathogenic mechanisms drive diverse diseaseprocesses reflected by these MRI measures These pat-terns also suggest a predominant role for lipids andlipid-specific immunity in MS pathology Furtherstudies are warranted to determine whether the earlyappearance of anti-lipid antibodies predicts the
Neurology Neuroimmunology amp Neuroinflammation 9
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
subsequent development of clinical and MRI-defineddisease worsening
AUTHOR CONTRIBUTIONSRohit Bakshi draftingrevising the manuscript study concept or design
analysis or interpretation of data acquisition of data study supervision
obtaining funding Ada Yeste study concept or design acquisition of
data Bonny Patel analysis or interpretation of data statistical analysis
Shahamat Tauhid analysis or interpretation of data Subhash Tummala
analysis or interpretation of data Roya Rahbari analysis or interpretation
of data statistical analysis Renxin Chu analysis or interpretation of data
Keren Regev analysis or interpretation of data acquisition of data
Pia Kivisaumlkk draftingrevising the manuscript study concept or design
contribution of vital reagentstoolspatients acquisition of data study
supervision Howard L Weiner study concept or design obtaining fund-
ing Francisco J Quintana draftingrevising the manuscript study con-
cept or design contribution of vital reagentstoolspatients study
supervision obtaining funding
ACKNOWLEDGMENTThe authors thank Dr Mohit Neema Dr Antonella Ceccarelli and
Dr Ashish Arora for valuable assistance at the early stages of this project
STUDY FUNDINGThis work was supported in part by grants to FJQ and HLW from
EMD Serono RG4111A1 and JF2161-A-5 from the National Multiple
Sclerosis Society PA0069 from the International Progressive MS Alli-
ance and by grants from the NIH (5R01NS055083-04) and NMSS
(RG3798A2) to RB
DISCLOSUREFJ Quintana serves on the editorial board for Systems Biomedicine
Inmunologia American Journal of Clinical and Experimental Immunology
is an associate editor for Immunology (UK) is an advisory board member
for Seminars in Immunopathology received research support from Harvard
Medical School BADERC NMSS A Yeste and B Patel report no
disclosures S Tauhid is managing editor for Journal of Neuroimaging
S Tummala R Rahbari R Chu and K Regev report no disclosures
P Kivisaumlkk received research support from EMD Serono R Bakshi is
editor-in-chief for Journal of Neuroimaging received consulting fees from
AbbVie Alkermes Biogen Novartis Questcor received research support
from Biogen EMD-Serono Novartis Sanofi-Genzyme Teva his spouse
holds stock in Biogen Inc H Weiner served on the advisory board
for The Guthy-Jackson Charitable Foundation Teva Pharmaceuticals
Industries Ltd Biogen Idec Novartis Sanofi-Aventis has consulted for
Therapix Bioven Novartis Serono Teva Sanofi received research support
from National Multiple Sclerosis Society Go to Neurologyorgnn for full
disclosure forms
Received July 27 2015 Accepted in final form December 8 2015
REFERENCES1 Weiner HL The challenge of multiple sclerosis how do
we cure a chronic heterogeneous disease Ann Neurol
200965239ndash248
2 Bakshi R Thompson AJ Rocca MA et al MRI in mul-
tiple sclerosis current status and future prospects Lancet
Neurol 20087615ndash625
3 Ceccarelli A Bakshi R Neema M MRI in multiple scle-
rosis a review of the current literature Curr Opin Neurol
201225402ndash409
4 Filippi M Rocca MA Barkhof F et al Association
between pathological and MRI findings in multiple scle-
rosis Lancet Neurol 201211349ndash360
5 Geurts JJ Calabrese M Fisher E Rudick RA Measure-
ment and clinical effect of grey matter pathology in mul-
tiple sclerosis Lancet Neurol 2012111082ndash1092
6 Klaver R De Vries HE Schenk GJ Geurts JJ Grey mat-
ter damage in multiple sclerosis a pathology perspective
Prion 2013766ndash75
7 Khoury S Bakshi R Cerebral pseudoatrophy or real atro-
phy after therapy in multiple sclerosis Ann Neurol 2010
68778ndash779
8 Hauser SL Chan JR Oksenberg JR Multiple sclerosis
prospects and promise Ann Neurol 201374317ndash327
9 McFarland HF Martin R Multiple sclerosis a complicated
picture of autoimmunity Nat Immunol 20078913ndash919
10 Nylander A Hafler DA Multiple sclerosis J Clin Invest
20121221180ndash1188
11 Sospedra M Martin R Immunology of multiple sclerosis
Ann Rev Immunol 200523683ndash747
12 Steinman L Immunology of relapse and remission in mul-
tiple sclerosis Ann Rev Immunol 201432257ndash281
13 Khoury SJ Guttmann CR Orav EJ Kikinis R Jolesz FA
Weiner HL Changes in activated T cells in the blood
correlate with disease activity in multiple sclerosis Arch
Neurol 2000571183ndash1189
14 Khoury SJ Orav EJ Guttmann CR Kikinis R Jolesz FA
Weiner HL Changes in serum levels of ICAM and TNF-
R correlate with disease activity in multiple sclerosis Neu-
rology 199953758ndash764
15 Laplaud DA Berthelot L Miqueu P et al Serial blood T
cell repertoire alterations in multiple sclerosis patients cor-
relation with clinical and MRI parameters J Neuroimmunol
2006177151ndash160
16 Makhlouf K Weiner HL Khoury SJ Increased percentage
of IL-121 monocytes in the blood correlates with the
presence of active MRI lesions in MS J Neuroimmunol
2001119145ndash149
17 Prat A Biernacki K Saroli T et al Kinin B1 receptor expres-
sion on multiple sclerosis mononuclear cells correlation with
magnetic resonance imaging T2-weighted lesion volume and
clinical disability Arch Neurol 200562795ndash800
18 Rinaldi L Gallo P Calabrese M et al Longitudinal anal-
ysis of immune cell phenotypes in early stage multiple
sclerosis distinctive patterns characterize MRI-active pa-
tients Brain 20061291993ndash2007
19 Tortorella P Lagana MM Saresella M et al Determinants
of disability in multiple sclerosis an immunological and
MRI study Biomed Res Int 20142014875768
20 Robinson WH DiGennaro C Hueber W et al Autoan-
tigen microarrays for multiplex characterization of autoan-
tibody responses Nat Med 20028295ndash301
21 Yeste A Quintana FJ Antigen microarrays for the study of
autoimmune diseases Clin Chem 2013591036ndash1044
22 Li QZ Xie C Wu T et al Identification of autoantibody
clusters that best predict lupus disease activity using glomer-
ular proteome arrays J Clin Invest 20051153428ndash3439
23 Hueber W Kidd BA Tomooka BH et al Antigen micro-
array profiling of autoantibodies in rheumatoid arthritis
Arthritis Rheum 2005522645ndash2655
24 Goldschmidt Y Sharon E Quintana FJ Cohen IR
Brandt A Adaptive methods for classification of biological
microarray data from multiple experiments Report no
MCS03-07 Rehovot Israel The Arthur and Rochelle Bel-
fer Institute of Mathematics and Computer Science 2003
25 Quintana FJ Getz G Hed G Domany E Cohen IR
Cluster analysis of human autoantibody reactivities in
health and in type 1 diabetes mellitus a bio-informatic
approach to immune complexity J Autoimmun 2003
2165ndash75
10 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
26 Lalive PH Menge T Barman I Cree BA Genain CP
Identification of new serum autoantibodies in neuromye-
litis optica using protein microarrays Neurology 200667
176ndash177
27 Kanter JL Narayana S Ho PP et al Lipid microarrays
identify key mediators of autoimmune brain inflamma-
tion Nat Med 200612138ndash143
28 Quintana FJ Farez MF Izquierdo G Lucas M Cohen IR
Weiner HL Antigen microarrays identify CNS-produced
autoantibodies in RRMS Neurology 201278532ndash539
29 Farez MF Quintana FJ Gandhi R Izquierdo G Lucas M
Weiner HL Toll-like receptor 2 and poly(ADP-ribose)
polymerase 1 promote central nervous system neuroin-
flammation in progressive EAE Nat Immunol 200910
958ndash964
30 Quintana FJ Farez MF Viglietta V et al Antigen micro-
arrays identify unique serum autoantibody signatures in
clinical and pathologic subtypes of multiple sclerosis Proc
Natl Acad Sci USA 200810518889ndash18894
31 Quintana FJ Patel B Yeste A et al Epitope spreading as
an early pathogenic event in pediatric multiple sclerosis
Neurology 2014832219ndash2226
32 Gauthier SA Glanz BI Mandel M Weiner HL A model
for the comprehensive investigation of a chronic autoim-
mune disease the multiple sclerosis CLIMB study Auto-
immun Rev 20065532ndash536
33 Polman CH Reingold SC Edan G et al Diagnostic criteria
for multiple sclerosis 2005 revisions to the ldquoMcDonald
Criteriardquo Ann Neurol 200558840ndash846
34 DellrsquoOglio E Ceccarelli A Glanz BI et al Quantification
of global cerebral atrophy in multiple sclerosis from 3T
MRI using SPM the role of misclassification errors
J Neuroimaging 201525191ndash199
35 Ceccarelli A Jackson JS Tauhid S et al The impact of
lesion in-painting and registration methods on voxel-based
morphometry in detecting regional cerebral gray matter
atrophy in multiple sclerosis AJNR Am J Neuroradiol
2012331579ndash1585
36 Cohen AB Neema M Arora A et al The relationships
among MRI-defined spinal cord involvement brain involve-
ment and disability in multiple sclerosis J Neuroimaging
201222122ndash128
37 Sanfilipo MP Benedict RH Sharma J Weinstock-
Guttman B Bakshi R The relationship between whole
brain volume and disability in multiple sclerosis a com-
parison of normalized gray vs white matter with misclas-
sification correction Neuroimage 2005261068ndash1077
38 Sanfilipo MP Benedict RH Weinstock-Guttman B
Bakshi R Gray and white matter brain atrophy and neuro-
psychological impairment in multiple sclerosis Neurology
200666685ndash692
39 Stankiewicz JM Glanz BI Healy BC et al Brain MRI
lesion load at 15T and 3T versus clinical status in multi-
ple sclerosis J Neuroimaging 201121e50ndashe56
40 Quintana FJ Hagedorn PH Elizur G Merbl Y
Domany E Cohen IR Functional immunomics micro-
array analysis of IgG autoantibody repertoires predicts the
future response of mice to induced diabetes Proc Natl
Acad Sci USA 2004101(suppl 2)14615ndash14621
41 Bermel RA Bakshi R The measurement and clinical
relevance of brain atrophy in multiple sclerosis Lancet
Neurol 20065158ndash170
42 Eikelenboom MJ Petzold A Lazeron RH et al Multiple
sclerosis neurofilament light chain antibodies are corre-
lated to cerebral atrophy Neurology 200360219ndash223
43 Magraner MJ Bosca I Simo-Castello M et al Brain atro-
phy and lesion load are related to CSF lipid-specific IgM
oligoclonal bands in clinically isolated syndromes Neuro-
radiol 2012545ndash12
44 Villar LM Sadaba MC Roldan E et al Intrathecal syn-
thesis of oligoclonal IgM against myelin lipids predicts an
aggressive disease course in MS J Clin Invest 2005115
187ndash194
45 Merbl Y Zucker-Toledano M Quintana FJ Cohen IR
Newborn humans manifest autoantibodies to defined self
molecules detected by antigen microarray informatics
J Clin Invest 2007117712ndash718
46 Sormani MP Arnold DL De Stefano N Treatment effect
on brain atrophy correlates with treatment effect on disa-
bility in multiple sclerosis Ann Neurol 20147543ndash49
47 Tauhid S Neema M Healy BC Weiner HL Bakshi R
MRI phenotypes based on cerebral lesions and atrophy in
patients with multiple sclerosis J Neurol Sci 2014346
250ndash254
48 Gnanapavan S Grant D Morant S et al Biomarker
report from the phase II lamotrigine trial in secondary
progressive MS neurofilament as a surrogate of disease
progression PLoS One 20138e70019
49 Quintana FJ Cohen IR Heat shock proteins as endoge-
nous adjuvants in sterile and septic inflammation
J Immunol 20051752777ndash2782
50 Quintana FJ Cohen IR The HSP60 immune system net-
work Trends Immunol 20113289ndash95
51 Ho PP Kanter JL Johnson AM et al Identification of
naturally occurring fatty acids of the myelin sheath that
resolve neuroinflammation Sci Transl Med 20124137ra73
52 Quintana FJ Yeste A Weiner HL Covacu R Lipids and
lipid-reactive antibodies as biomarkers for multiple sclero-
sis J Neuroimmunol 201224853ndash57
53 Mayo L Trauger SA Blain M et al Regulation of astro-
cyte activation by glycolipids drives chronic CNS inflam-
mation Nat Med 2014201147ndash1156
Neurology Neuroimmunology amp Neuroinflammation 11
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
1
Table e-1 Antigens used
Group Antigen Source
27 kDa Heat Shock Protein Stressgen
HSP 32 kDa Heat Shock Protein Stressgen
40 kDa Heat Shock Protein Stressgen
47 kDa Heat Shock Protein Stressgen
60 kDa Heat Shock Protein Stressgen
60 kDa Heat Shock Protein peptide aa 106ndash125 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 121ndash140 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 136ndash155 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 151ndash170 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 16ndash35 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 166ndash185 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
2
60 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 556ndash573 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
65 kDa Heat Shock Protein M tuberculosis Stressgen
70 kDa Heat Shock Protein Stressgen
70 kDa Heat Shock Protein peptide aa 106ndash125 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 121ndash140 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 136ndash155 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 151ndash170 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 16ndash35 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 166ndash185 Biopolymers Facility Harvard Medical School
3
70 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
4
70 kDa Heat Shock Protein peptide aa 556ndash575 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 571ndash590 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 586ndash605 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 601ndash620 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 616ndash635 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 631ndash640 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
71 kDa Heat Shock Protein M tuberculosis Biopolymers Facility Harvard Medical School
90 kDa Heat Shock Protein Stressgen
GroEL Stressgen
23-cyclic nucleotide 3-phosphodiesterase
peptide aa106-125 Biopolymers Facility Harvard Medical School
CNS
23-cyclic nucleotide 3-phosphodiesterase
peptide aa1-20 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa121-140 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa136-155 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa151-170 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa16-35 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa166-185 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase Biopolymers Facility Harvard Medical School
5
peptide aa181-200
23-cyclic nucleotide 3-phosphodiesterase
peptide aa196-215 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa211-230 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa226-245 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa241-260 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa256-275 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa271-290 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa286-305 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa301-320 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa31-50 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa316-335 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa331-350 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa346-365 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa361-380 Biopolymers Facility Harvard Medical School
6
23-cyclic nucleotide 3-phosphodiesterase
peptide aa376-395 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa391-410 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa406-421 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa46-65 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa61-80 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa76-95 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa91-110 Biopolymers Facility Harvard Medical School
Acetyl Cholinesterase Sigma Aldrich
ADAM-10 Sigma Aldrich
alpha-Cristallin Stressgen
beta-Cristallin Sigma Aldrich
bovine Myelin Basic Protein Sigma Aldrich
Brain Extract I Sigma Aldrich
Brain Extract II Sigma Aldrich
Brain Extract III Sigma Aldrich
Glial Filament Acidic Protein Research Diagnostic
guinea pig Myelin Basic Protein Sigma Aldrich
human Myelin Basic Protein Sigma Aldrich
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 106-125 Biopolymers Facility Harvard Medical School
7
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 121-140 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 136-155 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 151-170 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 166-185 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 181-200 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 91-110 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
106-125 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa 1- Biopolymers Facility Harvard Medical School
8
20
Myelinoligodendrocyte glycoprotein peptide aa
121-140 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
136-155 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
151-170 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
16-35 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
166-185 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
181-200 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
196-215 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
211-230 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
226-247 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
31-50 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
35-55 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
46-65 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
61-80 Biopolymers Facility Harvard Medical School
9
Myelinoligodendrocyte glycoprotein peptide aa
76-95 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
91-110 Biopolymers Facility Harvard Medical School
murine Myelin Basic Protein Sigma Aldrich
Myelin Associated Glycoprotein Sigma Aldrich
Myelin Basic Protein peptide aa 104-123 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 11-30 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 113-132 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 121-138 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 124-142 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 138-147 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 141-161 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 143-168 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 155-178 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 26-35 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 41-60 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 51-70 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 71-92 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 84-94 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 89-101 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 173-186 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 93-112 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 106-125 Biopolymers Facility Harvard Medical School
10
Myelin Protein 2 peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 121-132 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 91-110 Biopolymers Facility Harvard Medical School
Neurofilament 160kd Chemicon
Neurofilament 200kd Chemicon
Neurofilament 68kd Chemicon
Neuronal Enolase Calbiochem
Nicastrin GeneTex
NMDA receptor Novus Biologicals
NOGO Sigma Aldrich
Olygodendrocyte-Specific Protein peptide aa 106ndash
125 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 121ndash
140 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 136ndash
155 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 151ndash
170 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 16ndash
35 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 166ndash Biopolymers Facility Harvard Medical School
11
185
Olygodendrocyte-Specific Protein peptide aa 181ndash
199 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 195ndash
217 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 31ndash
50 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 46ndash
65 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 61ndash
80 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 76ndash
95 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 91ndash
110 Biopolymers Facility Harvard Medical School
Proteolipid Protein Abnova
Proteolipid Protein peptide aa 100-119 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 10-29 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 110-129 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 1-19 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 125-141 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-150 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-154 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 150-163 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 151-173 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 158-166 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 161-180 Biopolymers Facility Harvard Medical School
12
Proteolipid Protein peptide aa 178-191 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 180-199 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 190-209 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 20-39 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 205-220 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 215-232 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-239 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-249 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 248-259 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 250-269 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 265-277 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 35-50 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 40-59 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 50-69 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 65-84 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 80-99 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 91-110 Biopolymers Facility Harvard Medical School
Retinol Binding Protein Sigma Aldrich
S100beta protein Assay Designs
Super Oxide Dismutase Sigma Aldrich
Synuclein beta Sigma Aldrich
Synuclein gamma Sigma Aldrich
Amydgala ProSci Inc
Tissue Amydgala AD ProSci Inc
Brain lysate ProSci Inc
Brain Tissue Membrane ProSci Inc
Cerebellar pedunculus ProSci Inc
13
Cerebral meninges ProSci Inc
Corpus Callosum ProSci Inc
Corpus Callosum AD ProSci Inc
Diencephalon ProSci Inc
Fetal brain ProSci Inc
Frontal lobe ProSci Inc
Frontal lobe AD ProSci Inc
Hippocampus ProSci Inc
Hippocampus AD ProSci Inc
Insula ProSci Inc
Occipital lobe ProSci Inc
Occipital lobe AD ProSci Inc
Olfactory region ProSci Inc
Optic Nerve ProSci Inc
Parietal lobe ProSci Inc
Parietal lobe AD ProSci Inc
Pons ProSci Inc
Pons AD ProSci Inc
Postcentral gyrus ProSci Inc
Postcentral gyrus AD ProSci Inc
Precentral gyrus ProSci Inc
Precentral gyrus AD ProSci Inc
Spinal cord ProSci Inc
Temporal lobe ProSci Inc
Temporal lobe AD ProSci Inc
Thalamus ProSci Inc
Thalamus AD ProSci Inc
14
Amyloid beta Sigma Aldrich
AD
related Amyloid beta 10-20 Sigma Aldrich
Amyloid beta 1-12 Sigma Aldrich
Amyloid beta 12-28 Sigma Aldrich
Amyloid beta 1-23 Sigma Aldrich
Amyloid beta 1-38 Sigma Aldrich
Amyloid beta 17-40 Sigma Aldrich
Amyloid beta 25-35 Sigma Aldrich
Amyloid beta 34-42 Sigma Aldrich
Amyloid bri protein precursor 227 Sigma Aldrich
Amyloid DAN Protein Fragment 1-34 Sigma Aldrich
Amyloid Precursor Protein Sigma Aldrich
Amyloid protein no AB component Sigma Aldrich
Secreted amyloid precursor protein (SAP) beta Sigma Aldrich
Tau isoform variant 0N3R Sigma Aldrich
Tau isoform variant 1N3R Sigma Aldrich
Tau isoform variant 0N4R Sigma Aldrich
Tau isoform variant 2N3R Sigma Aldrich
Tau phospho Ser412 Sigma Aldrich
Tau phospho Ser441 Sigma Aldrich
Tau phospho Thr181 Sigma Aldrich
Tau Protein human EMD Biosciences
(plusmn)9-HODE Cayman Chemical
Lipids
1 Palmitoyl-2-(5oxo-Valeroyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
15a-hydroxycholestene Avanti Polar Lipids
15
15-ketocholestane Avanti Polar Lipids
15-ketocholestene Avanti Polar Lipids
1-Palmitoil-2-(9oxo-Nonanoyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Azelaoyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Glutaroyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
5 α-cholestane-3 β15 α-diol Avanti Polar Lipids
9(S)-HODE Cayman Chemical
Asialoganglioside-GM1 Sigma Aldrich
Asialoganglioside-GM2 Sigma Aldrich
Brain ceramides Avanti Polar Lipids
Brain D-erythrosphingosine Avanti Polar Lipids
Brain lysophosphatidylethanolamine Avanti Polar Lipids
Brain L-α-lysophosphatidylserine Avanti Polar Lipids
Brain L-α-phosphatidylcholine Avanti Polar Lipids
Brain L-α-phosphatidyl-ethanolamine Avanti Polar Lipids
Brain L-α-phosphatidylserine Avanti Polar Lipids
Brain polar lipid extract Avanti Polar Lipids
Brain sphingomyelin Avanti Polar Lipids
Brain sulfatide Avanti Polar Lipids
Brain total lipid extract Avanti Polar Lipids
Cardiolipin Sigma Aldrich
Ceramide Sigma Aldrich
Ceramide 1-phosphate Sigma Aldrich
Cholesterol Sigma Aldrich
16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-

correlation between BPF and GMF and their respec-tive lipid indexes and we validated the correlationbetween T2LV and its lipid index (figure 3C)
DISCUSSION Previous studies have investigated theassociation of MRI with immune activity in MS13ndash19
These studies have included cellular immune measures
Figure 3 Correlation of lipid indexes to MRI measures of disease severity in multiple sclerosis
(A) Heatmap inwhich each column represents anMRImeasure and each row represents the correlation to immunoglobulin G serumantibody reactivity to lipids accord-ing to a colorimetric scale (B) Scatter plots depicting the correlation between each lipid index andMRImeasures (BPF GMF and T2LV) in the discovery set (C) Scatterplots depicting the correlation between each lipid index andMRImeasures (BPF GMF and T2LV) in the validation set BPF5whole brain parenchymal fraction GMF5
cerebral gray matter fraction (see methods section for details) T2LV 5 cerebral T2 hyperintense lesion volume WMF 5 global cerebral white matter fraction
8 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
as well as oligoclonal bands or neurofilament-specificantibodies in CSF42ndash44 In the present study weanalyzed the association between serum antibodyprofiles detected with antigen microarrays and MRImeasures of disease including cerebral atrophy andT2 hyperintense lesions We found that specificantibody patterns are associated with different aspectsof MRI-defined disease pathology Our major findingwas that increased reactivity to lipids is associated withdifferent aspects of brain MRI measures of diseaseseverity Of note the specific set of lipids associatedwith atrophy differed from those associated withlesions Taken together these data suggest that anti-lipid antibodies in serum are related to MRI measuresof disease Our findings highlight the important role oflipid and lipid-specific immunity in the pathogenesis ofMS
Although a potential limitation of these studies isthe relatively small number of samples analyzed wehave validated our findings in an independent valida-tion cohort strengthening their significance In addi-tion further studies in large cohorts from patientsaffected by non-MS neurodegenerative diseases andhealthy controls may indicate whether the antibodyreactivities and lipid indexes identified in these stud-ies are exclusive to MS or are associated with addi-tional biological processes Indeed some of theseantibody reactivities contributing to the immune sig-natures described in this work have been found innewborns45
Different sets of antibody reactivities were associ-ated with GMF WMF BPF and T2LV The differ-ent antibody link between lesions and atrophy is inkeeping with the long-held view that brain atrophyin patients with MS is multifactorial and is onlyweakly related to lesions41 This has been underscoredin recent studies showing the complementary infor-mation obtained by combining lesion and atrophymeasures4647
Among the cerebral atrophy measures there was astriking overlap between those antibodies associatedwith GMF and BPF which separated them fromWMF results These findings are in keeping with pre-vious observations that whole brain atrophy is domi-nated by GM loss3741 In the early stages of MS thisGM atrophy selectively involves the deep GM nuclei34
Thus the similarities observed in the autoantibodieslinked to GMF and BPF might reflect the dominantcontribution and colinearity of GM atrophy to totalbrain atrophy The set of antibodies linked to GMFmay be the most relevant given that MRI studies haveshown that GM volume is more closely linked to phys-ical disability37 and cognitive impairment38 than areWM volume or lesion measures
Using antigen microarrays antibody reactivity tolipids has been detected in the CSF and serum of
patients with MS at different stages of the diseaseand such antibodies in the CSF have been linkedto disease progression and MRI involvement44
Although several mechanisms are thought to drivebrain inflammation and atrophy in MS46 the connec-tion between these mechanisms and the antibodiesdetected with our antigen microarrays is yet unknownOne possibility is that the serum autoantibodies iden-tified in our studies are not pathogenic and reflect theresult of immunization against self-antigens releasedfrom the CNS during the course of the disease Indeedneurofilament is released from damaged axons duringthe course of MS and both neurofilament lightchains48 and antibodies against it42 have been linkedto MRI measures of disease Moreover heat shockproteins are upregulated in different cell types duringinflammation and have been shown to have an impor-tant role as immunomodulators when released to theextracellular medium4950 Thus it is possible that heatshock proteinndashreactive antibodies reflect changes in theproduction and release of these immunomodulatorsduring the course of disease pathogenesis Similarlybioactive lipids and their products are released fromdamaged myelin and lipid-specific antibodies havebeen detected in patients with MS27293051 Lipids havean important role in the immune response both asbioactive molecules with immunomodulatory proper-ties and also as targets of the adaptive immuneresponse52 Moreover lipids have significant effectson the murine model of MS experimental autoim-mune encephalomyelitis2729305153 Indeed we recentlyfound that the glycolipid lactosylceramide activates abroad set of biological processes in astrocytes promot-ing neurodegeneration and inflammation53 Thus thelipid-reactive antibodies detected in this work mayreflect the release of myelin lipids in the context ofMS pathogenesis andor may directly contribute toimmune-mediated damage in CNS tissue Of notelipid-reactive antibodies are highly cross-reactive52 Inaddition the lipid-reactive antibodies detected in thiswork as associated with MRI measures of disease activ-ity were of the IgG class These are important points toconsider when evaluating a potential role of lipid-reactive antibodies inMS pathogenesis Further studiesare warranted to investigate whether bioactive lipidsoffer new therapeutic targets in MS
We have found that unique patterns of immunereactivity determined with antibody arrays are associ-ated with specific MRI measures of disease severityThese patterns agree with the interpretation that dif-ferent pathogenic mechanisms drive diverse diseaseprocesses reflected by these MRI measures These pat-terns also suggest a predominant role for lipids andlipid-specific immunity in MS pathology Furtherstudies are warranted to determine whether the earlyappearance of anti-lipid antibodies predicts the
Neurology Neuroimmunology amp Neuroinflammation 9
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
subsequent development of clinical and MRI-defineddisease worsening
AUTHOR CONTRIBUTIONSRohit Bakshi draftingrevising the manuscript study concept or design
analysis or interpretation of data acquisition of data study supervision
obtaining funding Ada Yeste study concept or design acquisition of
data Bonny Patel analysis or interpretation of data statistical analysis
Shahamat Tauhid analysis or interpretation of data Subhash Tummala
analysis or interpretation of data Roya Rahbari analysis or interpretation
of data statistical analysis Renxin Chu analysis or interpretation of data
Keren Regev analysis or interpretation of data acquisition of data
Pia Kivisaumlkk draftingrevising the manuscript study concept or design
contribution of vital reagentstoolspatients acquisition of data study
supervision Howard L Weiner study concept or design obtaining fund-
ing Francisco J Quintana draftingrevising the manuscript study con-
cept or design contribution of vital reagentstoolspatients study
supervision obtaining funding
ACKNOWLEDGMENTThe authors thank Dr Mohit Neema Dr Antonella Ceccarelli and
Dr Ashish Arora for valuable assistance at the early stages of this project
STUDY FUNDINGThis work was supported in part by grants to FJQ and HLW from
EMD Serono RG4111A1 and JF2161-A-5 from the National Multiple
Sclerosis Society PA0069 from the International Progressive MS Alli-
ance and by grants from the NIH (5R01NS055083-04) and NMSS
(RG3798A2) to RB
DISCLOSUREFJ Quintana serves on the editorial board for Systems Biomedicine
Inmunologia American Journal of Clinical and Experimental Immunology
is an associate editor for Immunology (UK) is an advisory board member
for Seminars in Immunopathology received research support from Harvard
Medical School BADERC NMSS A Yeste and B Patel report no
disclosures S Tauhid is managing editor for Journal of Neuroimaging
S Tummala R Rahbari R Chu and K Regev report no disclosures
P Kivisaumlkk received research support from EMD Serono R Bakshi is
editor-in-chief for Journal of Neuroimaging received consulting fees from
AbbVie Alkermes Biogen Novartis Questcor received research support
from Biogen EMD-Serono Novartis Sanofi-Genzyme Teva his spouse
holds stock in Biogen Inc H Weiner served on the advisory board
for The Guthy-Jackson Charitable Foundation Teva Pharmaceuticals
Industries Ltd Biogen Idec Novartis Sanofi-Aventis has consulted for
Therapix Bioven Novartis Serono Teva Sanofi received research support
from National Multiple Sclerosis Society Go to Neurologyorgnn for full
disclosure forms
Received July 27 2015 Accepted in final form December 8 2015
REFERENCES1 Weiner HL The challenge of multiple sclerosis how do
we cure a chronic heterogeneous disease Ann Neurol
200965239ndash248
2 Bakshi R Thompson AJ Rocca MA et al MRI in mul-
tiple sclerosis current status and future prospects Lancet
Neurol 20087615ndash625
3 Ceccarelli A Bakshi R Neema M MRI in multiple scle-
rosis a review of the current literature Curr Opin Neurol
201225402ndash409
4 Filippi M Rocca MA Barkhof F et al Association
between pathological and MRI findings in multiple scle-
rosis Lancet Neurol 201211349ndash360
5 Geurts JJ Calabrese M Fisher E Rudick RA Measure-
ment and clinical effect of grey matter pathology in mul-
tiple sclerosis Lancet Neurol 2012111082ndash1092
6 Klaver R De Vries HE Schenk GJ Geurts JJ Grey mat-
ter damage in multiple sclerosis a pathology perspective
Prion 2013766ndash75
7 Khoury S Bakshi R Cerebral pseudoatrophy or real atro-
phy after therapy in multiple sclerosis Ann Neurol 2010
68778ndash779
8 Hauser SL Chan JR Oksenberg JR Multiple sclerosis
prospects and promise Ann Neurol 201374317ndash327
9 McFarland HF Martin R Multiple sclerosis a complicated
picture of autoimmunity Nat Immunol 20078913ndash919
10 Nylander A Hafler DA Multiple sclerosis J Clin Invest
20121221180ndash1188
11 Sospedra M Martin R Immunology of multiple sclerosis
Ann Rev Immunol 200523683ndash747
12 Steinman L Immunology of relapse and remission in mul-
tiple sclerosis Ann Rev Immunol 201432257ndash281
13 Khoury SJ Guttmann CR Orav EJ Kikinis R Jolesz FA
Weiner HL Changes in activated T cells in the blood
correlate with disease activity in multiple sclerosis Arch
Neurol 2000571183ndash1189
14 Khoury SJ Orav EJ Guttmann CR Kikinis R Jolesz FA
Weiner HL Changes in serum levels of ICAM and TNF-
R correlate with disease activity in multiple sclerosis Neu-
rology 199953758ndash764
15 Laplaud DA Berthelot L Miqueu P et al Serial blood T
cell repertoire alterations in multiple sclerosis patients cor-
relation with clinical and MRI parameters J Neuroimmunol
2006177151ndash160
16 Makhlouf K Weiner HL Khoury SJ Increased percentage
of IL-121 monocytes in the blood correlates with the
presence of active MRI lesions in MS J Neuroimmunol
2001119145ndash149
17 Prat A Biernacki K Saroli T et al Kinin B1 receptor expres-
sion on multiple sclerosis mononuclear cells correlation with
magnetic resonance imaging T2-weighted lesion volume and
clinical disability Arch Neurol 200562795ndash800
18 Rinaldi L Gallo P Calabrese M et al Longitudinal anal-
ysis of immune cell phenotypes in early stage multiple
sclerosis distinctive patterns characterize MRI-active pa-
tients Brain 20061291993ndash2007
19 Tortorella P Lagana MM Saresella M et al Determinants
of disability in multiple sclerosis an immunological and
MRI study Biomed Res Int 20142014875768
20 Robinson WH DiGennaro C Hueber W et al Autoan-
tigen microarrays for multiplex characterization of autoan-
tibody responses Nat Med 20028295ndash301
21 Yeste A Quintana FJ Antigen microarrays for the study of
autoimmune diseases Clin Chem 2013591036ndash1044
22 Li QZ Xie C Wu T et al Identification of autoantibody
clusters that best predict lupus disease activity using glomer-
ular proteome arrays J Clin Invest 20051153428ndash3439
23 Hueber W Kidd BA Tomooka BH et al Antigen micro-
array profiling of autoantibodies in rheumatoid arthritis
Arthritis Rheum 2005522645ndash2655
24 Goldschmidt Y Sharon E Quintana FJ Cohen IR
Brandt A Adaptive methods for classification of biological
microarray data from multiple experiments Report no
MCS03-07 Rehovot Israel The Arthur and Rochelle Bel-
fer Institute of Mathematics and Computer Science 2003
25 Quintana FJ Getz G Hed G Domany E Cohen IR
Cluster analysis of human autoantibody reactivities in
health and in type 1 diabetes mellitus a bio-informatic
approach to immune complexity J Autoimmun 2003
2165ndash75
10 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
26 Lalive PH Menge T Barman I Cree BA Genain CP
Identification of new serum autoantibodies in neuromye-
litis optica using protein microarrays Neurology 200667
176ndash177
27 Kanter JL Narayana S Ho PP et al Lipid microarrays
identify key mediators of autoimmune brain inflamma-
tion Nat Med 200612138ndash143
28 Quintana FJ Farez MF Izquierdo G Lucas M Cohen IR
Weiner HL Antigen microarrays identify CNS-produced
autoantibodies in RRMS Neurology 201278532ndash539
29 Farez MF Quintana FJ Gandhi R Izquierdo G Lucas M
Weiner HL Toll-like receptor 2 and poly(ADP-ribose)
polymerase 1 promote central nervous system neuroin-
flammation in progressive EAE Nat Immunol 200910
958ndash964
30 Quintana FJ Farez MF Viglietta V et al Antigen micro-
arrays identify unique serum autoantibody signatures in
clinical and pathologic subtypes of multiple sclerosis Proc
Natl Acad Sci USA 200810518889ndash18894
31 Quintana FJ Patel B Yeste A et al Epitope spreading as
an early pathogenic event in pediatric multiple sclerosis
Neurology 2014832219ndash2226
32 Gauthier SA Glanz BI Mandel M Weiner HL A model
for the comprehensive investigation of a chronic autoim-
mune disease the multiple sclerosis CLIMB study Auto-
immun Rev 20065532ndash536
33 Polman CH Reingold SC Edan G et al Diagnostic criteria
for multiple sclerosis 2005 revisions to the ldquoMcDonald
Criteriardquo Ann Neurol 200558840ndash846
34 DellrsquoOglio E Ceccarelli A Glanz BI et al Quantification
of global cerebral atrophy in multiple sclerosis from 3T
MRI using SPM the role of misclassification errors
J Neuroimaging 201525191ndash199
35 Ceccarelli A Jackson JS Tauhid S et al The impact of
lesion in-painting and registration methods on voxel-based
morphometry in detecting regional cerebral gray matter
atrophy in multiple sclerosis AJNR Am J Neuroradiol
2012331579ndash1585
36 Cohen AB Neema M Arora A et al The relationships
among MRI-defined spinal cord involvement brain involve-
ment and disability in multiple sclerosis J Neuroimaging
201222122ndash128
37 Sanfilipo MP Benedict RH Sharma J Weinstock-
Guttman B Bakshi R The relationship between whole
brain volume and disability in multiple sclerosis a com-
parison of normalized gray vs white matter with misclas-
sification correction Neuroimage 2005261068ndash1077
38 Sanfilipo MP Benedict RH Weinstock-Guttman B
Bakshi R Gray and white matter brain atrophy and neuro-
psychological impairment in multiple sclerosis Neurology
200666685ndash692
39 Stankiewicz JM Glanz BI Healy BC et al Brain MRI
lesion load at 15T and 3T versus clinical status in multi-
ple sclerosis J Neuroimaging 201121e50ndashe56
40 Quintana FJ Hagedorn PH Elizur G Merbl Y
Domany E Cohen IR Functional immunomics micro-
array analysis of IgG autoantibody repertoires predicts the
future response of mice to induced diabetes Proc Natl
Acad Sci USA 2004101(suppl 2)14615ndash14621
41 Bermel RA Bakshi R The measurement and clinical
relevance of brain atrophy in multiple sclerosis Lancet
Neurol 20065158ndash170
42 Eikelenboom MJ Petzold A Lazeron RH et al Multiple
sclerosis neurofilament light chain antibodies are corre-
lated to cerebral atrophy Neurology 200360219ndash223
43 Magraner MJ Bosca I Simo-Castello M et al Brain atro-
phy and lesion load are related to CSF lipid-specific IgM
oligoclonal bands in clinically isolated syndromes Neuro-
radiol 2012545ndash12
44 Villar LM Sadaba MC Roldan E et al Intrathecal syn-
thesis of oligoclonal IgM against myelin lipids predicts an
aggressive disease course in MS J Clin Invest 2005115
187ndash194
45 Merbl Y Zucker-Toledano M Quintana FJ Cohen IR
Newborn humans manifest autoantibodies to defined self
molecules detected by antigen microarray informatics
J Clin Invest 2007117712ndash718
46 Sormani MP Arnold DL De Stefano N Treatment effect
on brain atrophy correlates with treatment effect on disa-
bility in multiple sclerosis Ann Neurol 20147543ndash49
47 Tauhid S Neema M Healy BC Weiner HL Bakshi R
MRI phenotypes based on cerebral lesions and atrophy in
patients with multiple sclerosis J Neurol Sci 2014346
250ndash254
48 Gnanapavan S Grant D Morant S et al Biomarker
report from the phase II lamotrigine trial in secondary
progressive MS neurofilament as a surrogate of disease
progression PLoS One 20138e70019
49 Quintana FJ Cohen IR Heat shock proteins as endoge-
nous adjuvants in sterile and septic inflammation
J Immunol 20051752777ndash2782
50 Quintana FJ Cohen IR The HSP60 immune system net-
work Trends Immunol 20113289ndash95
51 Ho PP Kanter JL Johnson AM et al Identification of
naturally occurring fatty acids of the myelin sheath that
resolve neuroinflammation Sci Transl Med 20124137ra73
52 Quintana FJ Yeste A Weiner HL Covacu R Lipids and
lipid-reactive antibodies as biomarkers for multiple sclero-
sis J Neuroimmunol 201224853ndash57
53 Mayo L Trauger SA Blain M et al Regulation of astro-
cyte activation by glycolipids drives chronic CNS inflam-
mation Nat Med 2014201147ndash1156
Neurology Neuroimmunology amp Neuroinflammation 11
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
1
Table e-1 Antigens used
Group Antigen Source
27 kDa Heat Shock Protein Stressgen
HSP 32 kDa Heat Shock Protein Stressgen
40 kDa Heat Shock Protein Stressgen
47 kDa Heat Shock Protein Stressgen
60 kDa Heat Shock Protein Stressgen
60 kDa Heat Shock Protein peptide aa 106ndash125 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 121ndash140 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 136ndash155 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 151ndash170 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 16ndash35 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 166ndash185 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
2
60 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 556ndash573 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
65 kDa Heat Shock Protein M tuberculosis Stressgen
70 kDa Heat Shock Protein Stressgen
70 kDa Heat Shock Protein peptide aa 106ndash125 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 121ndash140 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 136ndash155 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 151ndash170 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 16ndash35 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 166ndash185 Biopolymers Facility Harvard Medical School
3
70 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
4
70 kDa Heat Shock Protein peptide aa 556ndash575 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 571ndash590 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 586ndash605 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 601ndash620 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 616ndash635 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 631ndash640 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
71 kDa Heat Shock Protein M tuberculosis Biopolymers Facility Harvard Medical School
90 kDa Heat Shock Protein Stressgen
GroEL Stressgen
23-cyclic nucleotide 3-phosphodiesterase
peptide aa106-125 Biopolymers Facility Harvard Medical School
CNS
23-cyclic nucleotide 3-phosphodiesterase
peptide aa1-20 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa121-140 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa136-155 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa151-170 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa16-35 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa166-185 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase Biopolymers Facility Harvard Medical School
5
peptide aa181-200
23-cyclic nucleotide 3-phosphodiesterase
peptide aa196-215 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa211-230 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa226-245 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa241-260 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa256-275 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa271-290 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa286-305 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa301-320 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa31-50 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa316-335 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa331-350 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa346-365 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa361-380 Biopolymers Facility Harvard Medical School
6
23-cyclic nucleotide 3-phosphodiesterase
peptide aa376-395 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa391-410 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa406-421 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa46-65 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa61-80 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa76-95 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa91-110 Biopolymers Facility Harvard Medical School
Acetyl Cholinesterase Sigma Aldrich
ADAM-10 Sigma Aldrich
alpha-Cristallin Stressgen
beta-Cristallin Sigma Aldrich
bovine Myelin Basic Protein Sigma Aldrich
Brain Extract I Sigma Aldrich
Brain Extract II Sigma Aldrich
Brain Extract III Sigma Aldrich
Glial Filament Acidic Protein Research Diagnostic
guinea pig Myelin Basic Protein Sigma Aldrich
human Myelin Basic Protein Sigma Aldrich
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 106-125 Biopolymers Facility Harvard Medical School
7
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 121-140 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 136-155 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 151-170 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 166-185 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 181-200 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 91-110 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
106-125 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa 1- Biopolymers Facility Harvard Medical School
8
20
Myelinoligodendrocyte glycoprotein peptide aa
121-140 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
136-155 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
151-170 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
16-35 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
166-185 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
181-200 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
196-215 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
211-230 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
226-247 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
31-50 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
35-55 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
46-65 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
61-80 Biopolymers Facility Harvard Medical School
9
Myelinoligodendrocyte glycoprotein peptide aa
76-95 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
91-110 Biopolymers Facility Harvard Medical School
murine Myelin Basic Protein Sigma Aldrich
Myelin Associated Glycoprotein Sigma Aldrich
Myelin Basic Protein peptide aa 104-123 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 11-30 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 113-132 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 121-138 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 124-142 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 138-147 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 141-161 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 143-168 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 155-178 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 26-35 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 41-60 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 51-70 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 71-92 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 84-94 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 89-101 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 173-186 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 93-112 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 106-125 Biopolymers Facility Harvard Medical School
10
Myelin Protein 2 peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 121-132 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 91-110 Biopolymers Facility Harvard Medical School
Neurofilament 160kd Chemicon
Neurofilament 200kd Chemicon
Neurofilament 68kd Chemicon
Neuronal Enolase Calbiochem
Nicastrin GeneTex
NMDA receptor Novus Biologicals
NOGO Sigma Aldrich
Olygodendrocyte-Specific Protein peptide aa 106ndash
125 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 121ndash
140 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 136ndash
155 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 151ndash
170 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 16ndash
35 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 166ndash Biopolymers Facility Harvard Medical School
11
185
Olygodendrocyte-Specific Protein peptide aa 181ndash
199 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 195ndash
217 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 31ndash
50 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 46ndash
65 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 61ndash
80 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 76ndash
95 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 91ndash
110 Biopolymers Facility Harvard Medical School
Proteolipid Protein Abnova
Proteolipid Protein peptide aa 100-119 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 10-29 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 110-129 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 1-19 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 125-141 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-150 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-154 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 150-163 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 151-173 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 158-166 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 161-180 Biopolymers Facility Harvard Medical School
12
Proteolipid Protein peptide aa 178-191 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 180-199 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 190-209 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 20-39 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 205-220 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 215-232 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-239 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-249 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 248-259 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 250-269 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 265-277 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 35-50 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 40-59 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 50-69 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 65-84 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 80-99 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 91-110 Biopolymers Facility Harvard Medical School
Retinol Binding Protein Sigma Aldrich
S100beta protein Assay Designs
Super Oxide Dismutase Sigma Aldrich
Synuclein beta Sigma Aldrich
Synuclein gamma Sigma Aldrich
Amydgala ProSci Inc
Tissue Amydgala AD ProSci Inc
Brain lysate ProSci Inc
Brain Tissue Membrane ProSci Inc
Cerebellar pedunculus ProSci Inc
13
Cerebral meninges ProSci Inc
Corpus Callosum ProSci Inc
Corpus Callosum AD ProSci Inc
Diencephalon ProSci Inc
Fetal brain ProSci Inc
Frontal lobe ProSci Inc
Frontal lobe AD ProSci Inc
Hippocampus ProSci Inc
Hippocampus AD ProSci Inc
Insula ProSci Inc
Occipital lobe ProSci Inc
Occipital lobe AD ProSci Inc
Olfactory region ProSci Inc
Optic Nerve ProSci Inc
Parietal lobe ProSci Inc
Parietal lobe AD ProSci Inc
Pons ProSci Inc
Pons AD ProSci Inc
Postcentral gyrus ProSci Inc
Postcentral gyrus AD ProSci Inc
Precentral gyrus ProSci Inc
Precentral gyrus AD ProSci Inc
Spinal cord ProSci Inc
Temporal lobe ProSci Inc
Temporal lobe AD ProSci Inc
Thalamus ProSci Inc
Thalamus AD ProSci Inc
14
Amyloid beta Sigma Aldrich
AD
related Amyloid beta 10-20 Sigma Aldrich
Amyloid beta 1-12 Sigma Aldrich
Amyloid beta 12-28 Sigma Aldrich
Amyloid beta 1-23 Sigma Aldrich
Amyloid beta 1-38 Sigma Aldrich
Amyloid beta 17-40 Sigma Aldrich
Amyloid beta 25-35 Sigma Aldrich
Amyloid beta 34-42 Sigma Aldrich
Amyloid bri protein precursor 227 Sigma Aldrich
Amyloid DAN Protein Fragment 1-34 Sigma Aldrich
Amyloid Precursor Protein Sigma Aldrich
Amyloid protein no AB component Sigma Aldrich
Secreted amyloid precursor protein (SAP) beta Sigma Aldrich
Tau isoform variant 0N3R Sigma Aldrich
Tau isoform variant 1N3R Sigma Aldrich
Tau isoform variant 0N4R Sigma Aldrich
Tau isoform variant 2N3R Sigma Aldrich
Tau phospho Ser412 Sigma Aldrich
Tau phospho Ser441 Sigma Aldrich
Tau phospho Thr181 Sigma Aldrich
Tau Protein human EMD Biosciences
(plusmn)9-HODE Cayman Chemical
Lipids
1 Palmitoyl-2-(5oxo-Valeroyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
15a-hydroxycholestene Avanti Polar Lipids
15
15-ketocholestane Avanti Polar Lipids
15-ketocholestene Avanti Polar Lipids
1-Palmitoil-2-(9oxo-Nonanoyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Azelaoyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Glutaroyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
5 α-cholestane-3 β15 α-diol Avanti Polar Lipids
9(S)-HODE Cayman Chemical
Asialoganglioside-GM1 Sigma Aldrich
Asialoganglioside-GM2 Sigma Aldrich
Brain ceramides Avanti Polar Lipids
Brain D-erythrosphingosine Avanti Polar Lipids
Brain lysophosphatidylethanolamine Avanti Polar Lipids
Brain L-α-lysophosphatidylserine Avanti Polar Lipids
Brain L-α-phosphatidylcholine Avanti Polar Lipids
Brain L-α-phosphatidyl-ethanolamine Avanti Polar Lipids
Brain L-α-phosphatidylserine Avanti Polar Lipids
Brain polar lipid extract Avanti Polar Lipids
Brain sphingomyelin Avanti Polar Lipids
Brain sulfatide Avanti Polar Lipids
Brain total lipid extract Avanti Polar Lipids
Cardiolipin Sigma Aldrich
Ceramide Sigma Aldrich
Ceramide 1-phosphate Sigma Aldrich
Cholesterol Sigma Aldrich
16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-

as well as oligoclonal bands or neurofilament-specificantibodies in CSF42ndash44 In the present study weanalyzed the association between serum antibodyprofiles detected with antigen microarrays and MRImeasures of disease including cerebral atrophy andT2 hyperintense lesions We found that specificantibody patterns are associated with different aspectsof MRI-defined disease pathology Our major findingwas that increased reactivity to lipids is associated withdifferent aspects of brain MRI measures of diseaseseverity Of note the specific set of lipids associatedwith atrophy differed from those associated withlesions Taken together these data suggest that anti-lipid antibodies in serum are related to MRI measuresof disease Our findings highlight the important role oflipid and lipid-specific immunity in the pathogenesis ofMS
Although a potential limitation of these studies isthe relatively small number of samples analyzed wehave validated our findings in an independent valida-tion cohort strengthening their significance In addi-tion further studies in large cohorts from patientsaffected by non-MS neurodegenerative diseases andhealthy controls may indicate whether the antibodyreactivities and lipid indexes identified in these stud-ies are exclusive to MS or are associated with addi-tional biological processes Indeed some of theseantibody reactivities contributing to the immune sig-natures described in this work have been found innewborns45
Different sets of antibody reactivities were associ-ated with GMF WMF BPF and T2LV The differ-ent antibody link between lesions and atrophy is inkeeping with the long-held view that brain atrophyin patients with MS is multifactorial and is onlyweakly related to lesions41 This has been underscoredin recent studies showing the complementary infor-mation obtained by combining lesion and atrophymeasures4647
Among the cerebral atrophy measures there was astriking overlap between those antibodies associatedwith GMF and BPF which separated them fromWMF results These findings are in keeping with pre-vious observations that whole brain atrophy is domi-nated by GM loss3741 In the early stages of MS thisGM atrophy selectively involves the deep GM nuclei34
Thus the similarities observed in the autoantibodieslinked to GMF and BPF might reflect the dominantcontribution and colinearity of GM atrophy to totalbrain atrophy The set of antibodies linked to GMFmay be the most relevant given that MRI studies haveshown that GM volume is more closely linked to phys-ical disability37 and cognitive impairment38 than areWM volume or lesion measures
Using antigen microarrays antibody reactivity tolipids has been detected in the CSF and serum of
patients with MS at different stages of the diseaseand such antibodies in the CSF have been linkedto disease progression and MRI involvement44
Although several mechanisms are thought to drivebrain inflammation and atrophy in MS46 the connec-tion between these mechanisms and the antibodiesdetected with our antigen microarrays is yet unknownOne possibility is that the serum autoantibodies iden-tified in our studies are not pathogenic and reflect theresult of immunization against self-antigens releasedfrom the CNS during the course of the disease Indeedneurofilament is released from damaged axons duringthe course of MS and both neurofilament lightchains48 and antibodies against it42 have been linkedto MRI measures of disease Moreover heat shockproteins are upregulated in different cell types duringinflammation and have been shown to have an impor-tant role as immunomodulators when released to theextracellular medium4950 Thus it is possible that heatshock proteinndashreactive antibodies reflect changes in theproduction and release of these immunomodulatorsduring the course of disease pathogenesis Similarlybioactive lipids and their products are released fromdamaged myelin and lipid-specific antibodies havebeen detected in patients with MS27293051 Lipids havean important role in the immune response both asbioactive molecules with immunomodulatory proper-ties and also as targets of the adaptive immuneresponse52 Moreover lipids have significant effectson the murine model of MS experimental autoim-mune encephalomyelitis2729305153 Indeed we recentlyfound that the glycolipid lactosylceramide activates abroad set of biological processes in astrocytes promot-ing neurodegeneration and inflammation53 Thus thelipid-reactive antibodies detected in this work mayreflect the release of myelin lipids in the context ofMS pathogenesis andor may directly contribute toimmune-mediated damage in CNS tissue Of notelipid-reactive antibodies are highly cross-reactive52 Inaddition the lipid-reactive antibodies detected in thiswork as associated with MRI measures of disease activ-ity were of the IgG class These are important points toconsider when evaluating a potential role of lipid-reactive antibodies inMS pathogenesis Further studiesare warranted to investigate whether bioactive lipidsoffer new therapeutic targets in MS
We have found that unique patterns of immunereactivity determined with antibody arrays are associ-ated with specific MRI measures of disease severityThese patterns agree with the interpretation that dif-ferent pathogenic mechanisms drive diverse diseaseprocesses reflected by these MRI measures These pat-terns also suggest a predominant role for lipids andlipid-specific immunity in MS pathology Furtherstudies are warranted to determine whether the earlyappearance of anti-lipid antibodies predicts the
Neurology Neuroimmunology amp Neuroinflammation 9
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
subsequent development of clinical and MRI-defineddisease worsening
AUTHOR CONTRIBUTIONSRohit Bakshi draftingrevising the manuscript study concept or design
analysis or interpretation of data acquisition of data study supervision
obtaining funding Ada Yeste study concept or design acquisition of
data Bonny Patel analysis or interpretation of data statistical analysis
Shahamat Tauhid analysis or interpretation of data Subhash Tummala
analysis or interpretation of data Roya Rahbari analysis or interpretation
of data statistical analysis Renxin Chu analysis or interpretation of data
Keren Regev analysis or interpretation of data acquisition of data
Pia Kivisaumlkk draftingrevising the manuscript study concept or design
contribution of vital reagentstoolspatients acquisition of data study
supervision Howard L Weiner study concept or design obtaining fund-
ing Francisco J Quintana draftingrevising the manuscript study con-
cept or design contribution of vital reagentstoolspatients study
supervision obtaining funding
ACKNOWLEDGMENTThe authors thank Dr Mohit Neema Dr Antonella Ceccarelli and
Dr Ashish Arora for valuable assistance at the early stages of this project
STUDY FUNDINGThis work was supported in part by grants to FJQ and HLW from
EMD Serono RG4111A1 and JF2161-A-5 from the National Multiple
Sclerosis Society PA0069 from the International Progressive MS Alli-
ance and by grants from the NIH (5R01NS055083-04) and NMSS
(RG3798A2) to RB
DISCLOSUREFJ Quintana serves on the editorial board for Systems Biomedicine
Inmunologia American Journal of Clinical and Experimental Immunology
is an associate editor for Immunology (UK) is an advisory board member
for Seminars in Immunopathology received research support from Harvard
Medical School BADERC NMSS A Yeste and B Patel report no
disclosures S Tauhid is managing editor for Journal of Neuroimaging
S Tummala R Rahbari R Chu and K Regev report no disclosures
P Kivisaumlkk received research support from EMD Serono R Bakshi is
editor-in-chief for Journal of Neuroimaging received consulting fees from
AbbVie Alkermes Biogen Novartis Questcor received research support
from Biogen EMD-Serono Novartis Sanofi-Genzyme Teva his spouse
holds stock in Biogen Inc H Weiner served on the advisory board
for The Guthy-Jackson Charitable Foundation Teva Pharmaceuticals
Industries Ltd Biogen Idec Novartis Sanofi-Aventis has consulted for
Therapix Bioven Novartis Serono Teva Sanofi received research support
from National Multiple Sclerosis Society Go to Neurologyorgnn for full
disclosure forms
Received July 27 2015 Accepted in final form December 8 2015
REFERENCES1 Weiner HL The challenge of multiple sclerosis how do
we cure a chronic heterogeneous disease Ann Neurol
200965239ndash248
2 Bakshi R Thompson AJ Rocca MA et al MRI in mul-
tiple sclerosis current status and future prospects Lancet
Neurol 20087615ndash625
3 Ceccarelli A Bakshi R Neema M MRI in multiple scle-
rosis a review of the current literature Curr Opin Neurol
201225402ndash409
4 Filippi M Rocca MA Barkhof F et al Association
between pathological and MRI findings in multiple scle-
rosis Lancet Neurol 201211349ndash360
5 Geurts JJ Calabrese M Fisher E Rudick RA Measure-
ment and clinical effect of grey matter pathology in mul-
tiple sclerosis Lancet Neurol 2012111082ndash1092
6 Klaver R De Vries HE Schenk GJ Geurts JJ Grey mat-
ter damage in multiple sclerosis a pathology perspective
Prion 2013766ndash75
7 Khoury S Bakshi R Cerebral pseudoatrophy or real atro-
phy after therapy in multiple sclerosis Ann Neurol 2010
68778ndash779
8 Hauser SL Chan JR Oksenberg JR Multiple sclerosis
prospects and promise Ann Neurol 201374317ndash327
9 McFarland HF Martin R Multiple sclerosis a complicated
picture of autoimmunity Nat Immunol 20078913ndash919
10 Nylander A Hafler DA Multiple sclerosis J Clin Invest
20121221180ndash1188
11 Sospedra M Martin R Immunology of multiple sclerosis
Ann Rev Immunol 200523683ndash747
12 Steinman L Immunology of relapse and remission in mul-
tiple sclerosis Ann Rev Immunol 201432257ndash281
13 Khoury SJ Guttmann CR Orav EJ Kikinis R Jolesz FA
Weiner HL Changes in activated T cells in the blood
correlate with disease activity in multiple sclerosis Arch
Neurol 2000571183ndash1189
14 Khoury SJ Orav EJ Guttmann CR Kikinis R Jolesz FA
Weiner HL Changes in serum levels of ICAM and TNF-
R correlate with disease activity in multiple sclerosis Neu-
rology 199953758ndash764
15 Laplaud DA Berthelot L Miqueu P et al Serial blood T
cell repertoire alterations in multiple sclerosis patients cor-
relation with clinical and MRI parameters J Neuroimmunol
2006177151ndash160
16 Makhlouf K Weiner HL Khoury SJ Increased percentage
of IL-121 monocytes in the blood correlates with the
presence of active MRI lesions in MS J Neuroimmunol
2001119145ndash149
17 Prat A Biernacki K Saroli T et al Kinin B1 receptor expres-
sion on multiple sclerosis mononuclear cells correlation with
magnetic resonance imaging T2-weighted lesion volume and
clinical disability Arch Neurol 200562795ndash800
18 Rinaldi L Gallo P Calabrese M et al Longitudinal anal-
ysis of immune cell phenotypes in early stage multiple
sclerosis distinctive patterns characterize MRI-active pa-
tients Brain 20061291993ndash2007
19 Tortorella P Lagana MM Saresella M et al Determinants
of disability in multiple sclerosis an immunological and
MRI study Biomed Res Int 20142014875768
20 Robinson WH DiGennaro C Hueber W et al Autoan-
tigen microarrays for multiplex characterization of autoan-
tibody responses Nat Med 20028295ndash301
21 Yeste A Quintana FJ Antigen microarrays for the study of
autoimmune diseases Clin Chem 2013591036ndash1044
22 Li QZ Xie C Wu T et al Identification of autoantibody
clusters that best predict lupus disease activity using glomer-
ular proteome arrays J Clin Invest 20051153428ndash3439
23 Hueber W Kidd BA Tomooka BH et al Antigen micro-
array profiling of autoantibodies in rheumatoid arthritis
Arthritis Rheum 2005522645ndash2655
24 Goldschmidt Y Sharon E Quintana FJ Cohen IR
Brandt A Adaptive methods for classification of biological
microarray data from multiple experiments Report no
MCS03-07 Rehovot Israel The Arthur and Rochelle Bel-
fer Institute of Mathematics and Computer Science 2003
25 Quintana FJ Getz G Hed G Domany E Cohen IR
Cluster analysis of human autoantibody reactivities in
health and in type 1 diabetes mellitus a bio-informatic
approach to immune complexity J Autoimmun 2003
2165ndash75
10 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
26 Lalive PH Menge T Barman I Cree BA Genain CP
Identification of new serum autoantibodies in neuromye-
litis optica using protein microarrays Neurology 200667
176ndash177
27 Kanter JL Narayana S Ho PP et al Lipid microarrays
identify key mediators of autoimmune brain inflamma-
tion Nat Med 200612138ndash143
28 Quintana FJ Farez MF Izquierdo G Lucas M Cohen IR
Weiner HL Antigen microarrays identify CNS-produced
autoantibodies in RRMS Neurology 201278532ndash539
29 Farez MF Quintana FJ Gandhi R Izquierdo G Lucas M
Weiner HL Toll-like receptor 2 and poly(ADP-ribose)
polymerase 1 promote central nervous system neuroin-
flammation in progressive EAE Nat Immunol 200910
958ndash964
30 Quintana FJ Farez MF Viglietta V et al Antigen micro-
arrays identify unique serum autoantibody signatures in
clinical and pathologic subtypes of multiple sclerosis Proc
Natl Acad Sci USA 200810518889ndash18894
31 Quintana FJ Patel B Yeste A et al Epitope spreading as
an early pathogenic event in pediatric multiple sclerosis
Neurology 2014832219ndash2226
32 Gauthier SA Glanz BI Mandel M Weiner HL A model
for the comprehensive investigation of a chronic autoim-
mune disease the multiple sclerosis CLIMB study Auto-
immun Rev 20065532ndash536
33 Polman CH Reingold SC Edan G et al Diagnostic criteria
for multiple sclerosis 2005 revisions to the ldquoMcDonald
Criteriardquo Ann Neurol 200558840ndash846
34 DellrsquoOglio E Ceccarelli A Glanz BI et al Quantification
of global cerebral atrophy in multiple sclerosis from 3T
MRI using SPM the role of misclassification errors
J Neuroimaging 201525191ndash199
35 Ceccarelli A Jackson JS Tauhid S et al The impact of
lesion in-painting and registration methods on voxel-based
morphometry in detecting regional cerebral gray matter
atrophy in multiple sclerosis AJNR Am J Neuroradiol
2012331579ndash1585
36 Cohen AB Neema M Arora A et al The relationships
among MRI-defined spinal cord involvement brain involve-
ment and disability in multiple sclerosis J Neuroimaging
201222122ndash128
37 Sanfilipo MP Benedict RH Sharma J Weinstock-
Guttman B Bakshi R The relationship between whole
brain volume and disability in multiple sclerosis a com-
parison of normalized gray vs white matter with misclas-
sification correction Neuroimage 2005261068ndash1077
38 Sanfilipo MP Benedict RH Weinstock-Guttman B
Bakshi R Gray and white matter brain atrophy and neuro-
psychological impairment in multiple sclerosis Neurology
200666685ndash692
39 Stankiewicz JM Glanz BI Healy BC et al Brain MRI
lesion load at 15T and 3T versus clinical status in multi-
ple sclerosis J Neuroimaging 201121e50ndashe56
40 Quintana FJ Hagedorn PH Elizur G Merbl Y
Domany E Cohen IR Functional immunomics micro-
array analysis of IgG autoantibody repertoires predicts the
future response of mice to induced diabetes Proc Natl
Acad Sci USA 2004101(suppl 2)14615ndash14621
41 Bermel RA Bakshi R The measurement and clinical
relevance of brain atrophy in multiple sclerosis Lancet
Neurol 20065158ndash170
42 Eikelenboom MJ Petzold A Lazeron RH et al Multiple
sclerosis neurofilament light chain antibodies are corre-
lated to cerebral atrophy Neurology 200360219ndash223
43 Magraner MJ Bosca I Simo-Castello M et al Brain atro-
phy and lesion load are related to CSF lipid-specific IgM
oligoclonal bands in clinically isolated syndromes Neuro-
radiol 2012545ndash12
44 Villar LM Sadaba MC Roldan E et al Intrathecal syn-
thesis of oligoclonal IgM against myelin lipids predicts an
aggressive disease course in MS J Clin Invest 2005115
187ndash194
45 Merbl Y Zucker-Toledano M Quintana FJ Cohen IR
Newborn humans manifest autoantibodies to defined self
molecules detected by antigen microarray informatics
J Clin Invest 2007117712ndash718
46 Sormani MP Arnold DL De Stefano N Treatment effect
on brain atrophy correlates with treatment effect on disa-
bility in multiple sclerosis Ann Neurol 20147543ndash49
47 Tauhid S Neema M Healy BC Weiner HL Bakshi R
MRI phenotypes based on cerebral lesions and atrophy in
patients with multiple sclerosis J Neurol Sci 2014346
250ndash254
48 Gnanapavan S Grant D Morant S et al Biomarker
report from the phase II lamotrigine trial in secondary
progressive MS neurofilament as a surrogate of disease
progression PLoS One 20138e70019
49 Quintana FJ Cohen IR Heat shock proteins as endoge-
nous adjuvants in sterile and septic inflammation
J Immunol 20051752777ndash2782
50 Quintana FJ Cohen IR The HSP60 immune system net-
work Trends Immunol 20113289ndash95
51 Ho PP Kanter JL Johnson AM et al Identification of
naturally occurring fatty acids of the myelin sheath that
resolve neuroinflammation Sci Transl Med 20124137ra73
52 Quintana FJ Yeste A Weiner HL Covacu R Lipids and
lipid-reactive antibodies as biomarkers for multiple sclero-
sis J Neuroimmunol 201224853ndash57
53 Mayo L Trauger SA Blain M et al Regulation of astro-
cyte activation by glycolipids drives chronic CNS inflam-
mation Nat Med 2014201147ndash1156
Neurology Neuroimmunology amp Neuroinflammation 11
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
1
Table e-1 Antigens used
Group Antigen Source
27 kDa Heat Shock Protein Stressgen
HSP 32 kDa Heat Shock Protein Stressgen
40 kDa Heat Shock Protein Stressgen
47 kDa Heat Shock Protein Stressgen
60 kDa Heat Shock Protein Stressgen
60 kDa Heat Shock Protein peptide aa 106ndash125 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 121ndash140 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 136ndash155 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 151ndash170 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 16ndash35 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 166ndash185 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
2
60 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 556ndash573 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
65 kDa Heat Shock Protein M tuberculosis Stressgen
70 kDa Heat Shock Protein Stressgen
70 kDa Heat Shock Protein peptide aa 106ndash125 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 121ndash140 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 136ndash155 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 151ndash170 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 16ndash35 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 166ndash185 Biopolymers Facility Harvard Medical School
3
70 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
4
70 kDa Heat Shock Protein peptide aa 556ndash575 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 571ndash590 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 586ndash605 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 601ndash620 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 616ndash635 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 631ndash640 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
71 kDa Heat Shock Protein M tuberculosis Biopolymers Facility Harvard Medical School
90 kDa Heat Shock Protein Stressgen
GroEL Stressgen
23-cyclic nucleotide 3-phosphodiesterase
peptide aa106-125 Biopolymers Facility Harvard Medical School
CNS
23-cyclic nucleotide 3-phosphodiesterase
peptide aa1-20 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa121-140 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa136-155 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa151-170 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa16-35 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa166-185 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase Biopolymers Facility Harvard Medical School
5
peptide aa181-200
23-cyclic nucleotide 3-phosphodiesterase
peptide aa196-215 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa211-230 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa226-245 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa241-260 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa256-275 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa271-290 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa286-305 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa301-320 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa31-50 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa316-335 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa331-350 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa346-365 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa361-380 Biopolymers Facility Harvard Medical School
6
23-cyclic nucleotide 3-phosphodiesterase
peptide aa376-395 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa391-410 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa406-421 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa46-65 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa61-80 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa76-95 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa91-110 Biopolymers Facility Harvard Medical School
Acetyl Cholinesterase Sigma Aldrich
ADAM-10 Sigma Aldrich
alpha-Cristallin Stressgen
beta-Cristallin Sigma Aldrich
bovine Myelin Basic Protein Sigma Aldrich
Brain Extract I Sigma Aldrich
Brain Extract II Sigma Aldrich
Brain Extract III Sigma Aldrich
Glial Filament Acidic Protein Research Diagnostic
guinea pig Myelin Basic Protein Sigma Aldrich
human Myelin Basic Protein Sigma Aldrich
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 106-125 Biopolymers Facility Harvard Medical School
7
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 121-140 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 136-155 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 151-170 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 166-185 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 181-200 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 91-110 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
106-125 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa 1- Biopolymers Facility Harvard Medical School
8
20
Myelinoligodendrocyte glycoprotein peptide aa
121-140 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
136-155 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
151-170 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
16-35 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
166-185 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
181-200 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
196-215 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
211-230 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
226-247 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
31-50 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
35-55 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
46-65 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
61-80 Biopolymers Facility Harvard Medical School
9
Myelinoligodendrocyte glycoprotein peptide aa
76-95 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
91-110 Biopolymers Facility Harvard Medical School
murine Myelin Basic Protein Sigma Aldrich
Myelin Associated Glycoprotein Sigma Aldrich
Myelin Basic Protein peptide aa 104-123 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 11-30 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 113-132 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 121-138 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 124-142 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 138-147 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 141-161 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 143-168 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 155-178 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 26-35 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 41-60 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 51-70 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 71-92 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 84-94 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 89-101 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 173-186 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 93-112 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 106-125 Biopolymers Facility Harvard Medical School
10
Myelin Protein 2 peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 121-132 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 91-110 Biopolymers Facility Harvard Medical School
Neurofilament 160kd Chemicon
Neurofilament 200kd Chemicon
Neurofilament 68kd Chemicon
Neuronal Enolase Calbiochem
Nicastrin GeneTex
NMDA receptor Novus Biologicals
NOGO Sigma Aldrich
Olygodendrocyte-Specific Protein peptide aa 106ndash
125 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 121ndash
140 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 136ndash
155 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 151ndash
170 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 16ndash
35 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 166ndash Biopolymers Facility Harvard Medical School
11
185
Olygodendrocyte-Specific Protein peptide aa 181ndash
199 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 195ndash
217 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 31ndash
50 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 46ndash
65 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 61ndash
80 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 76ndash
95 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 91ndash
110 Biopolymers Facility Harvard Medical School
Proteolipid Protein Abnova
Proteolipid Protein peptide aa 100-119 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 10-29 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 110-129 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 1-19 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 125-141 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-150 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-154 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 150-163 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 151-173 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 158-166 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 161-180 Biopolymers Facility Harvard Medical School
12
Proteolipid Protein peptide aa 178-191 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 180-199 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 190-209 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 20-39 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 205-220 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 215-232 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-239 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-249 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 248-259 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 250-269 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 265-277 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 35-50 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 40-59 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 50-69 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 65-84 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 80-99 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 91-110 Biopolymers Facility Harvard Medical School
Retinol Binding Protein Sigma Aldrich
S100beta protein Assay Designs
Super Oxide Dismutase Sigma Aldrich
Synuclein beta Sigma Aldrich
Synuclein gamma Sigma Aldrich
Amydgala ProSci Inc
Tissue Amydgala AD ProSci Inc
Brain lysate ProSci Inc
Brain Tissue Membrane ProSci Inc
Cerebellar pedunculus ProSci Inc
13
Cerebral meninges ProSci Inc
Corpus Callosum ProSci Inc
Corpus Callosum AD ProSci Inc
Diencephalon ProSci Inc
Fetal brain ProSci Inc
Frontal lobe ProSci Inc
Frontal lobe AD ProSci Inc
Hippocampus ProSci Inc
Hippocampus AD ProSci Inc
Insula ProSci Inc
Occipital lobe ProSci Inc
Occipital lobe AD ProSci Inc
Olfactory region ProSci Inc
Optic Nerve ProSci Inc
Parietal lobe ProSci Inc
Parietal lobe AD ProSci Inc
Pons ProSci Inc
Pons AD ProSci Inc
Postcentral gyrus ProSci Inc
Postcentral gyrus AD ProSci Inc
Precentral gyrus ProSci Inc
Precentral gyrus AD ProSci Inc
Spinal cord ProSci Inc
Temporal lobe ProSci Inc
Temporal lobe AD ProSci Inc
Thalamus ProSci Inc
Thalamus AD ProSci Inc
14
Amyloid beta Sigma Aldrich
AD
related Amyloid beta 10-20 Sigma Aldrich
Amyloid beta 1-12 Sigma Aldrich
Amyloid beta 12-28 Sigma Aldrich
Amyloid beta 1-23 Sigma Aldrich
Amyloid beta 1-38 Sigma Aldrich
Amyloid beta 17-40 Sigma Aldrich
Amyloid beta 25-35 Sigma Aldrich
Amyloid beta 34-42 Sigma Aldrich
Amyloid bri protein precursor 227 Sigma Aldrich
Amyloid DAN Protein Fragment 1-34 Sigma Aldrich
Amyloid Precursor Protein Sigma Aldrich
Amyloid protein no AB component Sigma Aldrich
Secreted amyloid precursor protein (SAP) beta Sigma Aldrich
Tau isoform variant 0N3R Sigma Aldrich
Tau isoform variant 1N3R Sigma Aldrich
Tau isoform variant 0N4R Sigma Aldrich
Tau isoform variant 2N3R Sigma Aldrich
Tau phospho Ser412 Sigma Aldrich
Tau phospho Ser441 Sigma Aldrich
Tau phospho Thr181 Sigma Aldrich
Tau Protein human EMD Biosciences
(plusmn)9-HODE Cayman Chemical
Lipids
1 Palmitoyl-2-(5oxo-Valeroyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
15a-hydroxycholestene Avanti Polar Lipids
15
15-ketocholestane Avanti Polar Lipids
15-ketocholestene Avanti Polar Lipids
1-Palmitoil-2-(9oxo-Nonanoyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Azelaoyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Glutaroyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
5 α-cholestane-3 β15 α-diol Avanti Polar Lipids
9(S)-HODE Cayman Chemical
Asialoganglioside-GM1 Sigma Aldrich
Asialoganglioside-GM2 Sigma Aldrich
Brain ceramides Avanti Polar Lipids
Brain D-erythrosphingosine Avanti Polar Lipids
Brain lysophosphatidylethanolamine Avanti Polar Lipids
Brain L-α-lysophosphatidylserine Avanti Polar Lipids
Brain L-α-phosphatidylcholine Avanti Polar Lipids
Brain L-α-phosphatidyl-ethanolamine Avanti Polar Lipids
Brain L-α-phosphatidylserine Avanti Polar Lipids
Brain polar lipid extract Avanti Polar Lipids
Brain sphingomyelin Avanti Polar Lipids
Brain sulfatide Avanti Polar Lipids
Brain total lipid extract Avanti Polar Lipids
Cardiolipin Sigma Aldrich
Ceramide Sigma Aldrich
Ceramide 1-phosphate Sigma Aldrich
Cholesterol Sigma Aldrich
16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-

subsequent development of clinical and MRI-defineddisease worsening
AUTHOR CONTRIBUTIONSRohit Bakshi draftingrevising the manuscript study concept or design
analysis or interpretation of data acquisition of data study supervision
obtaining funding Ada Yeste study concept or design acquisition of
data Bonny Patel analysis or interpretation of data statistical analysis
Shahamat Tauhid analysis or interpretation of data Subhash Tummala
analysis or interpretation of data Roya Rahbari analysis or interpretation
of data statistical analysis Renxin Chu analysis or interpretation of data
Keren Regev analysis or interpretation of data acquisition of data
Pia Kivisaumlkk draftingrevising the manuscript study concept or design
contribution of vital reagentstoolspatients acquisition of data study
supervision Howard L Weiner study concept or design obtaining fund-
ing Francisco J Quintana draftingrevising the manuscript study con-
cept or design contribution of vital reagentstoolspatients study
supervision obtaining funding
ACKNOWLEDGMENTThe authors thank Dr Mohit Neema Dr Antonella Ceccarelli and
Dr Ashish Arora for valuable assistance at the early stages of this project
STUDY FUNDINGThis work was supported in part by grants to FJQ and HLW from
EMD Serono RG4111A1 and JF2161-A-5 from the National Multiple
Sclerosis Society PA0069 from the International Progressive MS Alli-
ance and by grants from the NIH (5R01NS055083-04) and NMSS
(RG3798A2) to RB
DISCLOSUREFJ Quintana serves on the editorial board for Systems Biomedicine
Inmunologia American Journal of Clinical and Experimental Immunology
is an associate editor for Immunology (UK) is an advisory board member
for Seminars in Immunopathology received research support from Harvard
Medical School BADERC NMSS A Yeste and B Patel report no
disclosures S Tauhid is managing editor for Journal of Neuroimaging
S Tummala R Rahbari R Chu and K Regev report no disclosures
P Kivisaumlkk received research support from EMD Serono R Bakshi is
editor-in-chief for Journal of Neuroimaging received consulting fees from
AbbVie Alkermes Biogen Novartis Questcor received research support
from Biogen EMD-Serono Novartis Sanofi-Genzyme Teva his spouse
holds stock in Biogen Inc H Weiner served on the advisory board
for The Guthy-Jackson Charitable Foundation Teva Pharmaceuticals
Industries Ltd Biogen Idec Novartis Sanofi-Aventis has consulted for
Therapix Bioven Novartis Serono Teva Sanofi received research support
from National Multiple Sclerosis Society Go to Neurologyorgnn for full
disclosure forms
Received July 27 2015 Accepted in final form December 8 2015
REFERENCES1 Weiner HL The challenge of multiple sclerosis how do
we cure a chronic heterogeneous disease Ann Neurol
200965239ndash248
2 Bakshi R Thompson AJ Rocca MA et al MRI in mul-
tiple sclerosis current status and future prospects Lancet
Neurol 20087615ndash625
3 Ceccarelli A Bakshi R Neema M MRI in multiple scle-
rosis a review of the current literature Curr Opin Neurol
201225402ndash409
4 Filippi M Rocca MA Barkhof F et al Association
between pathological and MRI findings in multiple scle-
rosis Lancet Neurol 201211349ndash360
5 Geurts JJ Calabrese M Fisher E Rudick RA Measure-
ment and clinical effect of grey matter pathology in mul-
tiple sclerosis Lancet Neurol 2012111082ndash1092
6 Klaver R De Vries HE Schenk GJ Geurts JJ Grey mat-
ter damage in multiple sclerosis a pathology perspective
Prion 2013766ndash75
7 Khoury S Bakshi R Cerebral pseudoatrophy or real atro-
phy after therapy in multiple sclerosis Ann Neurol 2010
68778ndash779
8 Hauser SL Chan JR Oksenberg JR Multiple sclerosis
prospects and promise Ann Neurol 201374317ndash327
9 McFarland HF Martin R Multiple sclerosis a complicated
picture of autoimmunity Nat Immunol 20078913ndash919
10 Nylander A Hafler DA Multiple sclerosis J Clin Invest
20121221180ndash1188
11 Sospedra M Martin R Immunology of multiple sclerosis
Ann Rev Immunol 200523683ndash747
12 Steinman L Immunology of relapse and remission in mul-
tiple sclerosis Ann Rev Immunol 201432257ndash281
13 Khoury SJ Guttmann CR Orav EJ Kikinis R Jolesz FA
Weiner HL Changes in activated T cells in the blood
correlate with disease activity in multiple sclerosis Arch
Neurol 2000571183ndash1189
14 Khoury SJ Orav EJ Guttmann CR Kikinis R Jolesz FA
Weiner HL Changes in serum levels of ICAM and TNF-
R correlate with disease activity in multiple sclerosis Neu-
rology 199953758ndash764
15 Laplaud DA Berthelot L Miqueu P et al Serial blood T
cell repertoire alterations in multiple sclerosis patients cor-
relation with clinical and MRI parameters J Neuroimmunol
2006177151ndash160
16 Makhlouf K Weiner HL Khoury SJ Increased percentage
of IL-121 monocytes in the blood correlates with the
presence of active MRI lesions in MS J Neuroimmunol
2001119145ndash149
17 Prat A Biernacki K Saroli T et al Kinin B1 receptor expres-
sion on multiple sclerosis mononuclear cells correlation with
magnetic resonance imaging T2-weighted lesion volume and
clinical disability Arch Neurol 200562795ndash800
18 Rinaldi L Gallo P Calabrese M et al Longitudinal anal-
ysis of immune cell phenotypes in early stage multiple
sclerosis distinctive patterns characterize MRI-active pa-
tients Brain 20061291993ndash2007
19 Tortorella P Lagana MM Saresella M et al Determinants
of disability in multiple sclerosis an immunological and
MRI study Biomed Res Int 20142014875768
20 Robinson WH DiGennaro C Hueber W et al Autoan-
tigen microarrays for multiplex characterization of autoan-
tibody responses Nat Med 20028295ndash301
21 Yeste A Quintana FJ Antigen microarrays for the study of
autoimmune diseases Clin Chem 2013591036ndash1044
22 Li QZ Xie C Wu T et al Identification of autoantibody
clusters that best predict lupus disease activity using glomer-
ular proteome arrays J Clin Invest 20051153428ndash3439
23 Hueber W Kidd BA Tomooka BH et al Antigen micro-
array profiling of autoantibodies in rheumatoid arthritis
Arthritis Rheum 2005522645ndash2655
24 Goldschmidt Y Sharon E Quintana FJ Cohen IR
Brandt A Adaptive methods for classification of biological
microarray data from multiple experiments Report no
MCS03-07 Rehovot Israel The Arthur and Rochelle Bel-
fer Institute of Mathematics and Computer Science 2003
25 Quintana FJ Getz G Hed G Domany E Cohen IR
Cluster analysis of human autoantibody reactivities in
health and in type 1 diabetes mellitus a bio-informatic
approach to immune complexity J Autoimmun 2003
2165ndash75
10 Neurology Neuroimmunology amp Neuroinflammation
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
26 Lalive PH Menge T Barman I Cree BA Genain CP
Identification of new serum autoantibodies in neuromye-
litis optica using protein microarrays Neurology 200667
176ndash177
27 Kanter JL Narayana S Ho PP et al Lipid microarrays
identify key mediators of autoimmune brain inflamma-
tion Nat Med 200612138ndash143
28 Quintana FJ Farez MF Izquierdo G Lucas M Cohen IR
Weiner HL Antigen microarrays identify CNS-produced
autoantibodies in RRMS Neurology 201278532ndash539
29 Farez MF Quintana FJ Gandhi R Izquierdo G Lucas M
Weiner HL Toll-like receptor 2 and poly(ADP-ribose)
polymerase 1 promote central nervous system neuroin-
flammation in progressive EAE Nat Immunol 200910
958ndash964
30 Quintana FJ Farez MF Viglietta V et al Antigen micro-
arrays identify unique serum autoantibody signatures in
clinical and pathologic subtypes of multiple sclerosis Proc
Natl Acad Sci USA 200810518889ndash18894
31 Quintana FJ Patel B Yeste A et al Epitope spreading as
an early pathogenic event in pediatric multiple sclerosis
Neurology 2014832219ndash2226
32 Gauthier SA Glanz BI Mandel M Weiner HL A model
for the comprehensive investigation of a chronic autoim-
mune disease the multiple sclerosis CLIMB study Auto-
immun Rev 20065532ndash536
33 Polman CH Reingold SC Edan G et al Diagnostic criteria
for multiple sclerosis 2005 revisions to the ldquoMcDonald
Criteriardquo Ann Neurol 200558840ndash846
34 DellrsquoOglio E Ceccarelli A Glanz BI et al Quantification
of global cerebral atrophy in multiple sclerosis from 3T
MRI using SPM the role of misclassification errors
J Neuroimaging 201525191ndash199
35 Ceccarelli A Jackson JS Tauhid S et al The impact of
lesion in-painting and registration methods on voxel-based
morphometry in detecting regional cerebral gray matter
atrophy in multiple sclerosis AJNR Am J Neuroradiol
2012331579ndash1585
36 Cohen AB Neema M Arora A et al The relationships
among MRI-defined spinal cord involvement brain involve-
ment and disability in multiple sclerosis J Neuroimaging
201222122ndash128
37 Sanfilipo MP Benedict RH Sharma J Weinstock-
Guttman B Bakshi R The relationship between whole
brain volume and disability in multiple sclerosis a com-
parison of normalized gray vs white matter with misclas-
sification correction Neuroimage 2005261068ndash1077
38 Sanfilipo MP Benedict RH Weinstock-Guttman B
Bakshi R Gray and white matter brain atrophy and neuro-
psychological impairment in multiple sclerosis Neurology
200666685ndash692
39 Stankiewicz JM Glanz BI Healy BC et al Brain MRI
lesion load at 15T and 3T versus clinical status in multi-
ple sclerosis J Neuroimaging 201121e50ndashe56
40 Quintana FJ Hagedorn PH Elizur G Merbl Y
Domany E Cohen IR Functional immunomics micro-
array analysis of IgG autoantibody repertoires predicts the
future response of mice to induced diabetes Proc Natl
Acad Sci USA 2004101(suppl 2)14615ndash14621
41 Bermel RA Bakshi R The measurement and clinical
relevance of brain atrophy in multiple sclerosis Lancet
Neurol 20065158ndash170
42 Eikelenboom MJ Petzold A Lazeron RH et al Multiple
sclerosis neurofilament light chain antibodies are corre-
lated to cerebral atrophy Neurology 200360219ndash223
43 Magraner MJ Bosca I Simo-Castello M et al Brain atro-
phy and lesion load are related to CSF lipid-specific IgM
oligoclonal bands in clinically isolated syndromes Neuro-
radiol 2012545ndash12
44 Villar LM Sadaba MC Roldan E et al Intrathecal syn-
thesis of oligoclonal IgM against myelin lipids predicts an
aggressive disease course in MS J Clin Invest 2005115
187ndash194
45 Merbl Y Zucker-Toledano M Quintana FJ Cohen IR
Newborn humans manifest autoantibodies to defined self
molecules detected by antigen microarray informatics
J Clin Invest 2007117712ndash718
46 Sormani MP Arnold DL De Stefano N Treatment effect
on brain atrophy correlates with treatment effect on disa-
bility in multiple sclerosis Ann Neurol 20147543ndash49
47 Tauhid S Neema M Healy BC Weiner HL Bakshi R
MRI phenotypes based on cerebral lesions and atrophy in
patients with multiple sclerosis J Neurol Sci 2014346
250ndash254
48 Gnanapavan S Grant D Morant S et al Biomarker
report from the phase II lamotrigine trial in secondary
progressive MS neurofilament as a surrogate of disease
progression PLoS One 20138e70019
49 Quintana FJ Cohen IR Heat shock proteins as endoge-
nous adjuvants in sterile and septic inflammation
J Immunol 20051752777ndash2782
50 Quintana FJ Cohen IR The HSP60 immune system net-
work Trends Immunol 20113289ndash95
51 Ho PP Kanter JL Johnson AM et al Identification of
naturally occurring fatty acids of the myelin sheath that
resolve neuroinflammation Sci Transl Med 20124137ra73
52 Quintana FJ Yeste A Weiner HL Covacu R Lipids and
lipid-reactive antibodies as biomarkers for multiple sclero-
sis J Neuroimmunol 201224853ndash57
53 Mayo L Trauger SA Blain M et al Regulation of astro-
cyte activation by glycolipids drives chronic CNS inflam-
mation Nat Med 2014201147ndash1156
Neurology Neuroimmunology amp Neuroinflammation 11
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
1
Table e-1 Antigens used
Group Antigen Source
27 kDa Heat Shock Protein Stressgen
HSP 32 kDa Heat Shock Protein Stressgen
40 kDa Heat Shock Protein Stressgen
47 kDa Heat Shock Protein Stressgen
60 kDa Heat Shock Protein Stressgen
60 kDa Heat Shock Protein peptide aa 106ndash125 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 121ndash140 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 136ndash155 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 151ndash170 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 16ndash35 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 166ndash185 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
2
60 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 556ndash573 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
65 kDa Heat Shock Protein M tuberculosis Stressgen
70 kDa Heat Shock Protein Stressgen
70 kDa Heat Shock Protein peptide aa 106ndash125 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 121ndash140 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 136ndash155 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 151ndash170 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 16ndash35 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 166ndash185 Biopolymers Facility Harvard Medical School
3
70 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
4
70 kDa Heat Shock Protein peptide aa 556ndash575 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 571ndash590 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 586ndash605 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 601ndash620 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 616ndash635 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 631ndash640 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
71 kDa Heat Shock Protein M tuberculosis Biopolymers Facility Harvard Medical School
90 kDa Heat Shock Protein Stressgen
GroEL Stressgen
23-cyclic nucleotide 3-phosphodiesterase
peptide aa106-125 Biopolymers Facility Harvard Medical School
CNS
23-cyclic nucleotide 3-phosphodiesterase
peptide aa1-20 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa121-140 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa136-155 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa151-170 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa16-35 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa166-185 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase Biopolymers Facility Harvard Medical School
5
peptide aa181-200
23-cyclic nucleotide 3-phosphodiesterase
peptide aa196-215 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa211-230 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa226-245 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa241-260 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa256-275 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa271-290 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa286-305 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa301-320 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa31-50 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa316-335 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa331-350 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa346-365 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa361-380 Biopolymers Facility Harvard Medical School
6
23-cyclic nucleotide 3-phosphodiesterase
peptide aa376-395 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa391-410 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa406-421 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa46-65 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa61-80 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa76-95 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa91-110 Biopolymers Facility Harvard Medical School
Acetyl Cholinesterase Sigma Aldrich
ADAM-10 Sigma Aldrich
alpha-Cristallin Stressgen
beta-Cristallin Sigma Aldrich
bovine Myelin Basic Protein Sigma Aldrich
Brain Extract I Sigma Aldrich
Brain Extract II Sigma Aldrich
Brain Extract III Sigma Aldrich
Glial Filament Acidic Protein Research Diagnostic
guinea pig Myelin Basic Protein Sigma Aldrich
human Myelin Basic Protein Sigma Aldrich
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 106-125 Biopolymers Facility Harvard Medical School
7
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 121-140 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 136-155 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 151-170 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 166-185 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 181-200 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 91-110 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
106-125 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa 1- Biopolymers Facility Harvard Medical School
8
20
Myelinoligodendrocyte glycoprotein peptide aa
121-140 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
136-155 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
151-170 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
16-35 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
166-185 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
181-200 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
196-215 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
211-230 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
226-247 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
31-50 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
35-55 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
46-65 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
61-80 Biopolymers Facility Harvard Medical School
9
Myelinoligodendrocyte glycoprotein peptide aa
76-95 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
91-110 Biopolymers Facility Harvard Medical School
murine Myelin Basic Protein Sigma Aldrich
Myelin Associated Glycoprotein Sigma Aldrich
Myelin Basic Protein peptide aa 104-123 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 11-30 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 113-132 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 121-138 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 124-142 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 138-147 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 141-161 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 143-168 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 155-178 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 26-35 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 41-60 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 51-70 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 71-92 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 84-94 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 89-101 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 173-186 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 93-112 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 106-125 Biopolymers Facility Harvard Medical School
10
Myelin Protein 2 peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 121-132 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 91-110 Biopolymers Facility Harvard Medical School
Neurofilament 160kd Chemicon
Neurofilament 200kd Chemicon
Neurofilament 68kd Chemicon
Neuronal Enolase Calbiochem
Nicastrin GeneTex
NMDA receptor Novus Biologicals
NOGO Sigma Aldrich
Olygodendrocyte-Specific Protein peptide aa 106ndash
125 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 121ndash
140 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 136ndash
155 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 151ndash
170 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 16ndash
35 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 166ndash Biopolymers Facility Harvard Medical School
11
185
Olygodendrocyte-Specific Protein peptide aa 181ndash
199 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 195ndash
217 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 31ndash
50 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 46ndash
65 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 61ndash
80 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 76ndash
95 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 91ndash
110 Biopolymers Facility Harvard Medical School
Proteolipid Protein Abnova
Proteolipid Protein peptide aa 100-119 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 10-29 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 110-129 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 1-19 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 125-141 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-150 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-154 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 150-163 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 151-173 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 158-166 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 161-180 Biopolymers Facility Harvard Medical School
12
Proteolipid Protein peptide aa 178-191 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 180-199 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 190-209 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 20-39 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 205-220 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 215-232 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-239 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-249 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 248-259 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 250-269 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 265-277 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 35-50 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 40-59 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 50-69 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 65-84 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 80-99 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 91-110 Biopolymers Facility Harvard Medical School
Retinol Binding Protein Sigma Aldrich
S100beta protein Assay Designs
Super Oxide Dismutase Sigma Aldrich
Synuclein beta Sigma Aldrich
Synuclein gamma Sigma Aldrich
Amydgala ProSci Inc
Tissue Amydgala AD ProSci Inc
Brain lysate ProSci Inc
Brain Tissue Membrane ProSci Inc
Cerebellar pedunculus ProSci Inc
13
Cerebral meninges ProSci Inc
Corpus Callosum ProSci Inc
Corpus Callosum AD ProSci Inc
Diencephalon ProSci Inc
Fetal brain ProSci Inc
Frontal lobe ProSci Inc
Frontal lobe AD ProSci Inc
Hippocampus ProSci Inc
Hippocampus AD ProSci Inc
Insula ProSci Inc
Occipital lobe ProSci Inc
Occipital lobe AD ProSci Inc
Olfactory region ProSci Inc
Optic Nerve ProSci Inc
Parietal lobe ProSci Inc
Parietal lobe AD ProSci Inc
Pons ProSci Inc
Pons AD ProSci Inc
Postcentral gyrus ProSci Inc
Postcentral gyrus AD ProSci Inc
Precentral gyrus ProSci Inc
Precentral gyrus AD ProSci Inc
Spinal cord ProSci Inc
Temporal lobe ProSci Inc
Temporal lobe AD ProSci Inc
Thalamus ProSci Inc
Thalamus AD ProSci Inc
14
Amyloid beta Sigma Aldrich
AD
related Amyloid beta 10-20 Sigma Aldrich
Amyloid beta 1-12 Sigma Aldrich
Amyloid beta 12-28 Sigma Aldrich
Amyloid beta 1-23 Sigma Aldrich
Amyloid beta 1-38 Sigma Aldrich
Amyloid beta 17-40 Sigma Aldrich
Amyloid beta 25-35 Sigma Aldrich
Amyloid beta 34-42 Sigma Aldrich
Amyloid bri protein precursor 227 Sigma Aldrich
Amyloid DAN Protein Fragment 1-34 Sigma Aldrich
Amyloid Precursor Protein Sigma Aldrich
Amyloid protein no AB component Sigma Aldrich
Secreted amyloid precursor protein (SAP) beta Sigma Aldrich
Tau isoform variant 0N3R Sigma Aldrich
Tau isoform variant 1N3R Sigma Aldrich
Tau isoform variant 0N4R Sigma Aldrich
Tau isoform variant 2N3R Sigma Aldrich
Tau phospho Ser412 Sigma Aldrich
Tau phospho Ser441 Sigma Aldrich
Tau phospho Thr181 Sigma Aldrich
Tau Protein human EMD Biosciences
(plusmn)9-HODE Cayman Chemical
Lipids
1 Palmitoyl-2-(5oxo-Valeroyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
15a-hydroxycholestene Avanti Polar Lipids
15
15-ketocholestane Avanti Polar Lipids
15-ketocholestene Avanti Polar Lipids
1-Palmitoil-2-(9oxo-Nonanoyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Azelaoyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Glutaroyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
5 α-cholestane-3 β15 α-diol Avanti Polar Lipids
9(S)-HODE Cayman Chemical
Asialoganglioside-GM1 Sigma Aldrich
Asialoganglioside-GM2 Sigma Aldrich
Brain ceramides Avanti Polar Lipids
Brain D-erythrosphingosine Avanti Polar Lipids
Brain lysophosphatidylethanolamine Avanti Polar Lipids
Brain L-α-lysophosphatidylserine Avanti Polar Lipids
Brain L-α-phosphatidylcholine Avanti Polar Lipids
Brain L-α-phosphatidyl-ethanolamine Avanti Polar Lipids
Brain L-α-phosphatidylserine Avanti Polar Lipids
Brain polar lipid extract Avanti Polar Lipids
Brain sphingomyelin Avanti Polar Lipids
Brain sulfatide Avanti Polar Lipids
Brain total lipid extract Avanti Polar Lipids
Cardiolipin Sigma Aldrich
Ceramide Sigma Aldrich
Ceramide 1-phosphate Sigma Aldrich
Cholesterol Sigma Aldrich
16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-

26 Lalive PH Menge T Barman I Cree BA Genain CP
Identification of new serum autoantibodies in neuromye-
litis optica using protein microarrays Neurology 200667
176ndash177
27 Kanter JL Narayana S Ho PP et al Lipid microarrays
identify key mediators of autoimmune brain inflamma-
tion Nat Med 200612138ndash143
28 Quintana FJ Farez MF Izquierdo G Lucas M Cohen IR
Weiner HL Antigen microarrays identify CNS-produced
autoantibodies in RRMS Neurology 201278532ndash539
29 Farez MF Quintana FJ Gandhi R Izquierdo G Lucas M
Weiner HL Toll-like receptor 2 and poly(ADP-ribose)
polymerase 1 promote central nervous system neuroin-
flammation in progressive EAE Nat Immunol 200910
958ndash964
30 Quintana FJ Farez MF Viglietta V et al Antigen micro-
arrays identify unique serum autoantibody signatures in
clinical and pathologic subtypes of multiple sclerosis Proc
Natl Acad Sci USA 200810518889ndash18894
31 Quintana FJ Patel B Yeste A et al Epitope spreading as
an early pathogenic event in pediatric multiple sclerosis
Neurology 2014832219ndash2226
32 Gauthier SA Glanz BI Mandel M Weiner HL A model
for the comprehensive investigation of a chronic autoim-
mune disease the multiple sclerosis CLIMB study Auto-
immun Rev 20065532ndash536
33 Polman CH Reingold SC Edan G et al Diagnostic criteria
for multiple sclerosis 2005 revisions to the ldquoMcDonald
Criteriardquo Ann Neurol 200558840ndash846
34 DellrsquoOglio E Ceccarelli A Glanz BI et al Quantification
of global cerebral atrophy in multiple sclerosis from 3T
MRI using SPM the role of misclassification errors
J Neuroimaging 201525191ndash199
35 Ceccarelli A Jackson JS Tauhid S et al The impact of
lesion in-painting and registration methods on voxel-based
morphometry in detecting regional cerebral gray matter
atrophy in multiple sclerosis AJNR Am J Neuroradiol
2012331579ndash1585
36 Cohen AB Neema M Arora A et al The relationships
among MRI-defined spinal cord involvement brain involve-
ment and disability in multiple sclerosis J Neuroimaging
201222122ndash128
37 Sanfilipo MP Benedict RH Sharma J Weinstock-
Guttman B Bakshi R The relationship between whole
brain volume and disability in multiple sclerosis a com-
parison of normalized gray vs white matter with misclas-
sification correction Neuroimage 2005261068ndash1077
38 Sanfilipo MP Benedict RH Weinstock-Guttman B
Bakshi R Gray and white matter brain atrophy and neuro-
psychological impairment in multiple sclerosis Neurology
200666685ndash692
39 Stankiewicz JM Glanz BI Healy BC et al Brain MRI
lesion load at 15T and 3T versus clinical status in multi-
ple sclerosis J Neuroimaging 201121e50ndashe56
40 Quintana FJ Hagedorn PH Elizur G Merbl Y
Domany E Cohen IR Functional immunomics micro-
array analysis of IgG autoantibody repertoires predicts the
future response of mice to induced diabetes Proc Natl
Acad Sci USA 2004101(suppl 2)14615ndash14621
41 Bermel RA Bakshi R The measurement and clinical
relevance of brain atrophy in multiple sclerosis Lancet
Neurol 20065158ndash170
42 Eikelenboom MJ Petzold A Lazeron RH et al Multiple
sclerosis neurofilament light chain antibodies are corre-
lated to cerebral atrophy Neurology 200360219ndash223
43 Magraner MJ Bosca I Simo-Castello M et al Brain atro-
phy and lesion load are related to CSF lipid-specific IgM
oligoclonal bands in clinically isolated syndromes Neuro-
radiol 2012545ndash12
44 Villar LM Sadaba MC Roldan E et al Intrathecal syn-
thesis of oligoclonal IgM against myelin lipids predicts an
aggressive disease course in MS J Clin Invest 2005115
187ndash194
45 Merbl Y Zucker-Toledano M Quintana FJ Cohen IR
Newborn humans manifest autoantibodies to defined self
molecules detected by antigen microarray informatics
J Clin Invest 2007117712ndash718
46 Sormani MP Arnold DL De Stefano N Treatment effect
on brain atrophy correlates with treatment effect on disa-
bility in multiple sclerosis Ann Neurol 20147543ndash49
47 Tauhid S Neema M Healy BC Weiner HL Bakshi R
MRI phenotypes based on cerebral lesions and atrophy in
patients with multiple sclerosis J Neurol Sci 2014346
250ndash254
48 Gnanapavan S Grant D Morant S et al Biomarker
report from the phase II lamotrigine trial in secondary
progressive MS neurofilament as a surrogate of disease
progression PLoS One 20138e70019
49 Quintana FJ Cohen IR Heat shock proteins as endoge-
nous adjuvants in sterile and septic inflammation
J Immunol 20051752777ndash2782
50 Quintana FJ Cohen IR The HSP60 immune system net-
work Trends Immunol 20113289ndash95
51 Ho PP Kanter JL Johnson AM et al Identification of
naturally occurring fatty acids of the myelin sheath that
resolve neuroinflammation Sci Transl Med 20124137ra73
52 Quintana FJ Yeste A Weiner HL Covacu R Lipids and
lipid-reactive antibodies as biomarkers for multiple sclero-
sis J Neuroimmunol 201224853ndash57
53 Mayo L Trauger SA Blain M et al Regulation of astro-
cyte activation by glycolipids drives chronic CNS inflam-
mation Nat Med 2014201147ndash1156
Neurology Neuroimmunology amp Neuroinflammation 11
ordf 2016 American Academy of Neurology Unauthorized reproduction of this article is prohibited
1
Table e-1 Antigens used
Group Antigen Source
27 kDa Heat Shock Protein Stressgen
HSP 32 kDa Heat Shock Protein Stressgen
40 kDa Heat Shock Protein Stressgen
47 kDa Heat Shock Protein Stressgen
60 kDa Heat Shock Protein Stressgen
60 kDa Heat Shock Protein peptide aa 106ndash125 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 121ndash140 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 136ndash155 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 151ndash170 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 16ndash35 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 166ndash185 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
2
60 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 556ndash573 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
65 kDa Heat Shock Protein M tuberculosis Stressgen
70 kDa Heat Shock Protein Stressgen
70 kDa Heat Shock Protein peptide aa 106ndash125 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 121ndash140 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 136ndash155 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 151ndash170 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 16ndash35 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 166ndash185 Biopolymers Facility Harvard Medical School
3
70 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
4
70 kDa Heat Shock Protein peptide aa 556ndash575 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 571ndash590 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 586ndash605 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 601ndash620 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 616ndash635 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 631ndash640 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
71 kDa Heat Shock Protein M tuberculosis Biopolymers Facility Harvard Medical School
90 kDa Heat Shock Protein Stressgen
GroEL Stressgen
23-cyclic nucleotide 3-phosphodiesterase
peptide aa106-125 Biopolymers Facility Harvard Medical School
CNS
23-cyclic nucleotide 3-phosphodiesterase
peptide aa1-20 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa121-140 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa136-155 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa151-170 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa16-35 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa166-185 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase Biopolymers Facility Harvard Medical School
5
peptide aa181-200
23-cyclic nucleotide 3-phosphodiesterase
peptide aa196-215 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa211-230 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa226-245 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa241-260 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa256-275 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa271-290 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa286-305 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa301-320 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa31-50 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa316-335 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa331-350 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa346-365 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa361-380 Biopolymers Facility Harvard Medical School
6
23-cyclic nucleotide 3-phosphodiesterase
peptide aa376-395 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa391-410 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa406-421 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa46-65 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa61-80 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa76-95 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa91-110 Biopolymers Facility Harvard Medical School
Acetyl Cholinesterase Sigma Aldrich
ADAM-10 Sigma Aldrich
alpha-Cristallin Stressgen
beta-Cristallin Sigma Aldrich
bovine Myelin Basic Protein Sigma Aldrich
Brain Extract I Sigma Aldrich
Brain Extract II Sigma Aldrich
Brain Extract III Sigma Aldrich
Glial Filament Acidic Protein Research Diagnostic
guinea pig Myelin Basic Protein Sigma Aldrich
human Myelin Basic Protein Sigma Aldrich
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 106-125 Biopolymers Facility Harvard Medical School
7
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 121-140 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 136-155 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 151-170 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 166-185 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 181-200 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 91-110 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
106-125 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa 1- Biopolymers Facility Harvard Medical School
8
20
Myelinoligodendrocyte glycoprotein peptide aa
121-140 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
136-155 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
151-170 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
16-35 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
166-185 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
181-200 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
196-215 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
211-230 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
226-247 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
31-50 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
35-55 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
46-65 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
61-80 Biopolymers Facility Harvard Medical School
9
Myelinoligodendrocyte glycoprotein peptide aa
76-95 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
91-110 Biopolymers Facility Harvard Medical School
murine Myelin Basic Protein Sigma Aldrich
Myelin Associated Glycoprotein Sigma Aldrich
Myelin Basic Protein peptide aa 104-123 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 11-30 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 113-132 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 121-138 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 124-142 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 138-147 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 141-161 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 143-168 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 155-178 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 26-35 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 41-60 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 51-70 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 71-92 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 84-94 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 89-101 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 173-186 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 93-112 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 106-125 Biopolymers Facility Harvard Medical School
10
Myelin Protein 2 peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 121-132 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 91-110 Biopolymers Facility Harvard Medical School
Neurofilament 160kd Chemicon
Neurofilament 200kd Chemicon
Neurofilament 68kd Chemicon
Neuronal Enolase Calbiochem
Nicastrin GeneTex
NMDA receptor Novus Biologicals
NOGO Sigma Aldrich
Olygodendrocyte-Specific Protein peptide aa 106ndash
125 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 121ndash
140 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 136ndash
155 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 151ndash
170 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 16ndash
35 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 166ndash Biopolymers Facility Harvard Medical School
11
185
Olygodendrocyte-Specific Protein peptide aa 181ndash
199 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 195ndash
217 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 31ndash
50 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 46ndash
65 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 61ndash
80 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 76ndash
95 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 91ndash
110 Biopolymers Facility Harvard Medical School
Proteolipid Protein Abnova
Proteolipid Protein peptide aa 100-119 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 10-29 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 110-129 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 1-19 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 125-141 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-150 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-154 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 150-163 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 151-173 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 158-166 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 161-180 Biopolymers Facility Harvard Medical School
12
Proteolipid Protein peptide aa 178-191 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 180-199 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 190-209 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 20-39 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 205-220 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 215-232 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-239 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-249 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 248-259 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 250-269 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 265-277 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 35-50 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 40-59 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 50-69 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 65-84 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 80-99 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 91-110 Biopolymers Facility Harvard Medical School
Retinol Binding Protein Sigma Aldrich
S100beta protein Assay Designs
Super Oxide Dismutase Sigma Aldrich
Synuclein beta Sigma Aldrich
Synuclein gamma Sigma Aldrich
Amydgala ProSci Inc
Tissue Amydgala AD ProSci Inc
Brain lysate ProSci Inc
Brain Tissue Membrane ProSci Inc
Cerebellar pedunculus ProSci Inc
13
Cerebral meninges ProSci Inc
Corpus Callosum ProSci Inc
Corpus Callosum AD ProSci Inc
Diencephalon ProSci Inc
Fetal brain ProSci Inc
Frontal lobe ProSci Inc
Frontal lobe AD ProSci Inc
Hippocampus ProSci Inc
Hippocampus AD ProSci Inc
Insula ProSci Inc
Occipital lobe ProSci Inc
Occipital lobe AD ProSci Inc
Olfactory region ProSci Inc
Optic Nerve ProSci Inc
Parietal lobe ProSci Inc
Parietal lobe AD ProSci Inc
Pons ProSci Inc
Pons AD ProSci Inc
Postcentral gyrus ProSci Inc
Postcentral gyrus AD ProSci Inc
Precentral gyrus ProSci Inc
Precentral gyrus AD ProSci Inc
Spinal cord ProSci Inc
Temporal lobe ProSci Inc
Temporal lobe AD ProSci Inc
Thalamus ProSci Inc
Thalamus AD ProSci Inc
14
Amyloid beta Sigma Aldrich
AD
related Amyloid beta 10-20 Sigma Aldrich
Amyloid beta 1-12 Sigma Aldrich
Amyloid beta 12-28 Sigma Aldrich
Amyloid beta 1-23 Sigma Aldrich
Amyloid beta 1-38 Sigma Aldrich
Amyloid beta 17-40 Sigma Aldrich
Amyloid beta 25-35 Sigma Aldrich
Amyloid beta 34-42 Sigma Aldrich
Amyloid bri protein precursor 227 Sigma Aldrich
Amyloid DAN Protein Fragment 1-34 Sigma Aldrich
Amyloid Precursor Protein Sigma Aldrich
Amyloid protein no AB component Sigma Aldrich
Secreted amyloid precursor protein (SAP) beta Sigma Aldrich
Tau isoform variant 0N3R Sigma Aldrich
Tau isoform variant 1N3R Sigma Aldrich
Tau isoform variant 0N4R Sigma Aldrich
Tau isoform variant 2N3R Sigma Aldrich
Tau phospho Ser412 Sigma Aldrich
Tau phospho Ser441 Sigma Aldrich
Tau phospho Thr181 Sigma Aldrich
Tau Protein human EMD Biosciences
(plusmn)9-HODE Cayman Chemical
Lipids
1 Palmitoyl-2-(5oxo-Valeroyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
15a-hydroxycholestene Avanti Polar Lipids
15
15-ketocholestane Avanti Polar Lipids
15-ketocholestene Avanti Polar Lipids
1-Palmitoil-2-(9oxo-Nonanoyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Azelaoyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Glutaroyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
5 α-cholestane-3 β15 α-diol Avanti Polar Lipids
9(S)-HODE Cayman Chemical
Asialoganglioside-GM1 Sigma Aldrich
Asialoganglioside-GM2 Sigma Aldrich
Brain ceramides Avanti Polar Lipids
Brain D-erythrosphingosine Avanti Polar Lipids
Brain lysophosphatidylethanolamine Avanti Polar Lipids
Brain L-α-lysophosphatidylserine Avanti Polar Lipids
Brain L-α-phosphatidylcholine Avanti Polar Lipids
Brain L-α-phosphatidyl-ethanolamine Avanti Polar Lipids
Brain L-α-phosphatidylserine Avanti Polar Lipids
Brain polar lipid extract Avanti Polar Lipids
Brain sphingomyelin Avanti Polar Lipids
Brain sulfatide Avanti Polar Lipids
Brain total lipid extract Avanti Polar Lipids
Cardiolipin Sigma Aldrich
Ceramide Sigma Aldrich
Ceramide 1-phosphate Sigma Aldrich
Cholesterol Sigma Aldrich
16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-
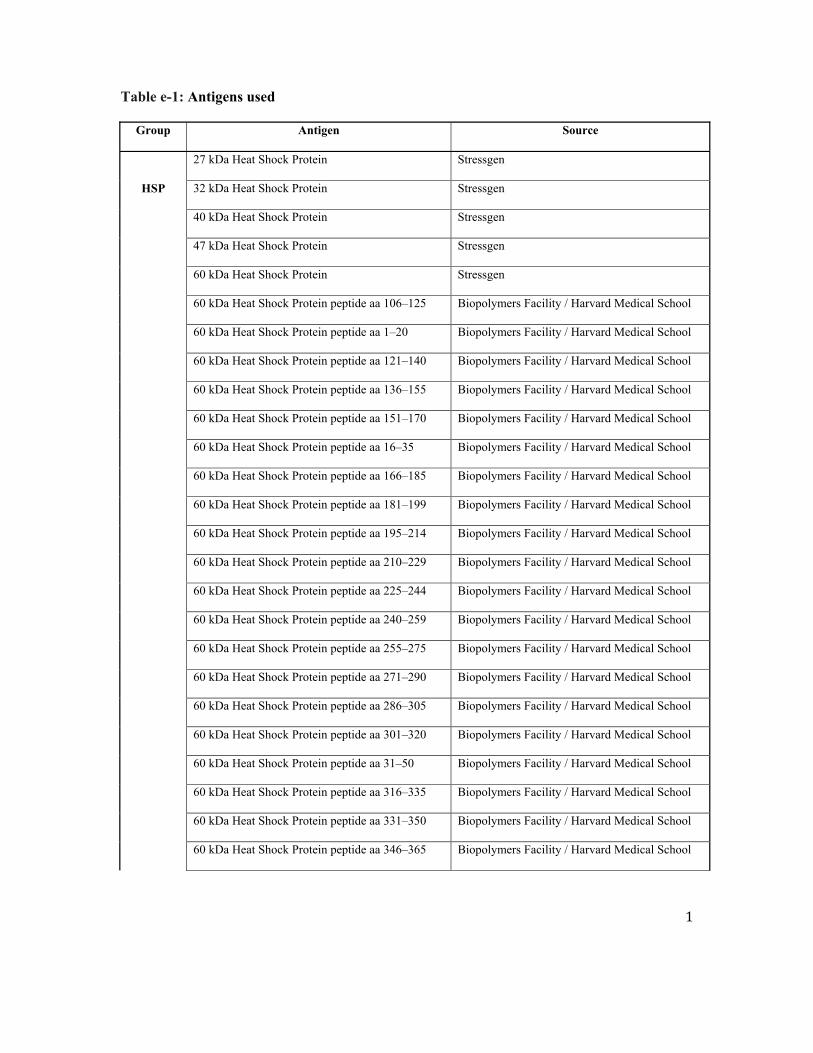
1
Table e-1 Antigens used
Group Antigen Source
27 kDa Heat Shock Protein Stressgen
HSP 32 kDa Heat Shock Protein Stressgen
40 kDa Heat Shock Protein Stressgen
47 kDa Heat Shock Protein Stressgen
60 kDa Heat Shock Protein Stressgen
60 kDa Heat Shock Protein peptide aa 106ndash125 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 121ndash140 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 136ndash155 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 151ndash170 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 16ndash35 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 166ndash185 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
2
60 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 556ndash573 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
65 kDa Heat Shock Protein M tuberculosis Stressgen
70 kDa Heat Shock Protein Stressgen
70 kDa Heat Shock Protein peptide aa 106ndash125 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 121ndash140 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 136ndash155 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 151ndash170 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 16ndash35 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 166ndash185 Biopolymers Facility Harvard Medical School
3
70 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
4
70 kDa Heat Shock Protein peptide aa 556ndash575 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 571ndash590 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 586ndash605 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 601ndash620 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 616ndash635 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 631ndash640 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
71 kDa Heat Shock Protein M tuberculosis Biopolymers Facility Harvard Medical School
90 kDa Heat Shock Protein Stressgen
GroEL Stressgen
23-cyclic nucleotide 3-phosphodiesterase
peptide aa106-125 Biopolymers Facility Harvard Medical School
CNS
23-cyclic nucleotide 3-phosphodiesterase
peptide aa1-20 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa121-140 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa136-155 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa151-170 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa16-35 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa166-185 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase Biopolymers Facility Harvard Medical School
5
peptide aa181-200
23-cyclic nucleotide 3-phosphodiesterase
peptide aa196-215 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa211-230 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa226-245 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa241-260 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa256-275 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa271-290 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa286-305 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa301-320 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa31-50 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa316-335 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa331-350 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa346-365 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa361-380 Biopolymers Facility Harvard Medical School
6
23-cyclic nucleotide 3-phosphodiesterase
peptide aa376-395 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa391-410 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa406-421 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa46-65 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa61-80 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa76-95 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa91-110 Biopolymers Facility Harvard Medical School
Acetyl Cholinesterase Sigma Aldrich
ADAM-10 Sigma Aldrich
alpha-Cristallin Stressgen
beta-Cristallin Sigma Aldrich
bovine Myelin Basic Protein Sigma Aldrich
Brain Extract I Sigma Aldrich
Brain Extract II Sigma Aldrich
Brain Extract III Sigma Aldrich
Glial Filament Acidic Protein Research Diagnostic
guinea pig Myelin Basic Protein Sigma Aldrich
human Myelin Basic Protein Sigma Aldrich
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 106-125 Biopolymers Facility Harvard Medical School
7
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 121-140 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 136-155 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 151-170 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 166-185 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 181-200 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 91-110 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
106-125 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa 1- Biopolymers Facility Harvard Medical School
8
20
Myelinoligodendrocyte glycoprotein peptide aa
121-140 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
136-155 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
151-170 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
16-35 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
166-185 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
181-200 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
196-215 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
211-230 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
226-247 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
31-50 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
35-55 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
46-65 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
61-80 Biopolymers Facility Harvard Medical School
9
Myelinoligodendrocyte glycoprotein peptide aa
76-95 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
91-110 Biopolymers Facility Harvard Medical School
murine Myelin Basic Protein Sigma Aldrich
Myelin Associated Glycoprotein Sigma Aldrich
Myelin Basic Protein peptide aa 104-123 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 11-30 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 113-132 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 121-138 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 124-142 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 138-147 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 141-161 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 143-168 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 155-178 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 26-35 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 41-60 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 51-70 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 71-92 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 84-94 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 89-101 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 173-186 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 93-112 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 106-125 Biopolymers Facility Harvard Medical School
10
Myelin Protein 2 peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 121-132 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 91-110 Biopolymers Facility Harvard Medical School
Neurofilament 160kd Chemicon
Neurofilament 200kd Chemicon
Neurofilament 68kd Chemicon
Neuronal Enolase Calbiochem
Nicastrin GeneTex
NMDA receptor Novus Biologicals
NOGO Sigma Aldrich
Olygodendrocyte-Specific Protein peptide aa 106ndash
125 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 121ndash
140 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 136ndash
155 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 151ndash
170 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 16ndash
35 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 166ndash Biopolymers Facility Harvard Medical School
11
185
Olygodendrocyte-Specific Protein peptide aa 181ndash
199 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 195ndash
217 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 31ndash
50 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 46ndash
65 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 61ndash
80 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 76ndash
95 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 91ndash
110 Biopolymers Facility Harvard Medical School
Proteolipid Protein Abnova
Proteolipid Protein peptide aa 100-119 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 10-29 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 110-129 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 1-19 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 125-141 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-150 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-154 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 150-163 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 151-173 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 158-166 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 161-180 Biopolymers Facility Harvard Medical School
12
Proteolipid Protein peptide aa 178-191 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 180-199 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 190-209 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 20-39 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 205-220 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 215-232 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-239 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-249 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 248-259 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 250-269 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 265-277 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 35-50 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 40-59 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 50-69 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 65-84 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 80-99 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 91-110 Biopolymers Facility Harvard Medical School
Retinol Binding Protein Sigma Aldrich
S100beta protein Assay Designs
Super Oxide Dismutase Sigma Aldrich
Synuclein beta Sigma Aldrich
Synuclein gamma Sigma Aldrich
Amydgala ProSci Inc
Tissue Amydgala AD ProSci Inc
Brain lysate ProSci Inc
Brain Tissue Membrane ProSci Inc
Cerebellar pedunculus ProSci Inc
13
Cerebral meninges ProSci Inc
Corpus Callosum ProSci Inc
Corpus Callosum AD ProSci Inc
Diencephalon ProSci Inc
Fetal brain ProSci Inc
Frontal lobe ProSci Inc
Frontal lobe AD ProSci Inc
Hippocampus ProSci Inc
Hippocampus AD ProSci Inc
Insula ProSci Inc
Occipital lobe ProSci Inc
Occipital lobe AD ProSci Inc
Olfactory region ProSci Inc
Optic Nerve ProSci Inc
Parietal lobe ProSci Inc
Parietal lobe AD ProSci Inc
Pons ProSci Inc
Pons AD ProSci Inc
Postcentral gyrus ProSci Inc
Postcentral gyrus AD ProSci Inc
Precentral gyrus ProSci Inc
Precentral gyrus AD ProSci Inc
Spinal cord ProSci Inc
Temporal lobe ProSci Inc
Temporal lobe AD ProSci Inc
Thalamus ProSci Inc
Thalamus AD ProSci Inc
14
Amyloid beta Sigma Aldrich
AD
related Amyloid beta 10-20 Sigma Aldrich
Amyloid beta 1-12 Sigma Aldrich
Amyloid beta 12-28 Sigma Aldrich
Amyloid beta 1-23 Sigma Aldrich
Amyloid beta 1-38 Sigma Aldrich
Amyloid beta 17-40 Sigma Aldrich
Amyloid beta 25-35 Sigma Aldrich
Amyloid beta 34-42 Sigma Aldrich
Amyloid bri protein precursor 227 Sigma Aldrich
Amyloid DAN Protein Fragment 1-34 Sigma Aldrich
Amyloid Precursor Protein Sigma Aldrich
Amyloid protein no AB component Sigma Aldrich
Secreted amyloid precursor protein (SAP) beta Sigma Aldrich
Tau isoform variant 0N3R Sigma Aldrich
Tau isoform variant 1N3R Sigma Aldrich
Tau isoform variant 0N4R Sigma Aldrich
Tau isoform variant 2N3R Sigma Aldrich
Tau phospho Ser412 Sigma Aldrich
Tau phospho Ser441 Sigma Aldrich
Tau phospho Thr181 Sigma Aldrich
Tau Protein human EMD Biosciences
(plusmn)9-HODE Cayman Chemical
Lipids
1 Palmitoyl-2-(5oxo-Valeroyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
15a-hydroxycholestene Avanti Polar Lipids
15
15-ketocholestane Avanti Polar Lipids
15-ketocholestene Avanti Polar Lipids
1-Palmitoil-2-(9oxo-Nonanoyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Azelaoyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Glutaroyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
5 α-cholestane-3 β15 α-diol Avanti Polar Lipids
9(S)-HODE Cayman Chemical
Asialoganglioside-GM1 Sigma Aldrich
Asialoganglioside-GM2 Sigma Aldrich
Brain ceramides Avanti Polar Lipids
Brain D-erythrosphingosine Avanti Polar Lipids
Brain lysophosphatidylethanolamine Avanti Polar Lipids
Brain L-α-lysophosphatidylserine Avanti Polar Lipids
Brain L-α-phosphatidylcholine Avanti Polar Lipids
Brain L-α-phosphatidyl-ethanolamine Avanti Polar Lipids
Brain L-α-phosphatidylserine Avanti Polar Lipids
Brain polar lipid extract Avanti Polar Lipids
Brain sphingomyelin Avanti Polar Lipids
Brain sulfatide Avanti Polar Lipids
Brain total lipid extract Avanti Polar Lipids
Cardiolipin Sigma Aldrich
Ceramide Sigma Aldrich
Ceramide 1-phosphate Sigma Aldrich
Cholesterol Sigma Aldrich
16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-

2
60 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 556ndash573 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
60 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
65 kDa Heat Shock Protein M tuberculosis Stressgen
70 kDa Heat Shock Protein Stressgen
70 kDa Heat Shock Protein peptide aa 106ndash125 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 121ndash140 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 136ndash155 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 151ndash170 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 16ndash35 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 166ndash185 Biopolymers Facility Harvard Medical School
3
70 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
4
70 kDa Heat Shock Protein peptide aa 556ndash575 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 571ndash590 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 586ndash605 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 601ndash620 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 616ndash635 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 631ndash640 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
71 kDa Heat Shock Protein M tuberculosis Biopolymers Facility Harvard Medical School
90 kDa Heat Shock Protein Stressgen
GroEL Stressgen
23-cyclic nucleotide 3-phosphodiesterase
peptide aa106-125 Biopolymers Facility Harvard Medical School
CNS
23-cyclic nucleotide 3-phosphodiesterase
peptide aa1-20 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa121-140 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa136-155 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa151-170 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa16-35 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa166-185 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase Biopolymers Facility Harvard Medical School
5
peptide aa181-200
23-cyclic nucleotide 3-phosphodiesterase
peptide aa196-215 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa211-230 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa226-245 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa241-260 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa256-275 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa271-290 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa286-305 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa301-320 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa31-50 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa316-335 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa331-350 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa346-365 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa361-380 Biopolymers Facility Harvard Medical School
6
23-cyclic nucleotide 3-phosphodiesterase
peptide aa376-395 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa391-410 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa406-421 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa46-65 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa61-80 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa76-95 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa91-110 Biopolymers Facility Harvard Medical School
Acetyl Cholinesterase Sigma Aldrich
ADAM-10 Sigma Aldrich
alpha-Cristallin Stressgen
beta-Cristallin Sigma Aldrich
bovine Myelin Basic Protein Sigma Aldrich
Brain Extract I Sigma Aldrich
Brain Extract II Sigma Aldrich
Brain Extract III Sigma Aldrich
Glial Filament Acidic Protein Research Diagnostic
guinea pig Myelin Basic Protein Sigma Aldrich
human Myelin Basic Protein Sigma Aldrich
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 106-125 Biopolymers Facility Harvard Medical School
7
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 121-140 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 136-155 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 151-170 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 166-185 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 181-200 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 91-110 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
106-125 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa 1- Biopolymers Facility Harvard Medical School
8
20
Myelinoligodendrocyte glycoprotein peptide aa
121-140 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
136-155 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
151-170 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
16-35 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
166-185 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
181-200 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
196-215 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
211-230 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
226-247 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
31-50 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
35-55 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
46-65 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
61-80 Biopolymers Facility Harvard Medical School
9
Myelinoligodendrocyte glycoprotein peptide aa
76-95 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
91-110 Biopolymers Facility Harvard Medical School
murine Myelin Basic Protein Sigma Aldrich
Myelin Associated Glycoprotein Sigma Aldrich
Myelin Basic Protein peptide aa 104-123 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 11-30 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 113-132 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 121-138 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 124-142 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 138-147 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 141-161 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 143-168 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 155-178 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 26-35 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 41-60 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 51-70 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 71-92 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 84-94 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 89-101 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 173-186 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 93-112 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 106-125 Biopolymers Facility Harvard Medical School
10
Myelin Protein 2 peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 121-132 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 91-110 Biopolymers Facility Harvard Medical School
Neurofilament 160kd Chemicon
Neurofilament 200kd Chemicon
Neurofilament 68kd Chemicon
Neuronal Enolase Calbiochem
Nicastrin GeneTex
NMDA receptor Novus Biologicals
NOGO Sigma Aldrich
Olygodendrocyte-Specific Protein peptide aa 106ndash
125 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 121ndash
140 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 136ndash
155 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 151ndash
170 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 16ndash
35 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 166ndash Biopolymers Facility Harvard Medical School
11
185
Olygodendrocyte-Specific Protein peptide aa 181ndash
199 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 195ndash
217 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 31ndash
50 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 46ndash
65 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 61ndash
80 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 76ndash
95 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 91ndash
110 Biopolymers Facility Harvard Medical School
Proteolipid Protein Abnova
Proteolipid Protein peptide aa 100-119 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 10-29 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 110-129 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 1-19 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 125-141 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-150 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-154 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 150-163 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 151-173 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 158-166 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 161-180 Biopolymers Facility Harvard Medical School
12
Proteolipid Protein peptide aa 178-191 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 180-199 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 190-209 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 20-39 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 205-220 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 215-232 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-239 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-249 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 248-259 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 250-269 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 265-277 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 35-50 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 40-59 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 50-69 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 65-84 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 80-99 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 91-110 Biopolymers Facility Harvard Medical School
Retinol Binding Protein Sigma Aldrich
S100beta protein Assay Designs
Super Oxide Dismutase Sigma Aldrich
Synuclein beta Sigma Aldrich
Synuclein gamma Sigma Aldrich
Amydgala ProSci Inc
Tissue Amydgala AD ProSci Inc
Brain lysate ProSci Inc
Brain Tissue Membrane ProSci Inc
Cerebellar pedunculus ProSci Inc
13
Cerebral meninges ProSci Inc
Corpus Callosum ProSci Inc
Corpus Callosum AD ProSci Inc
Diencephalon ProSci Inc
Fetal brain ProSci Inc
Frontal lobe ProSci Inc
Frontal lobe AD ProSci Inc
Hippocampus ProSci Inc
Hippocampus AD ProSci Inc
Insula ProSci Inc
Occipital lobe ProSci Inc
Occipital lobe AD ProSci Inc
Olfactory region ProSci Inc
Optic Nerve ProSci Inc
Parietal lobe ProSci Inc
Parietal lobe AD ProSci Inc
Pons ProSci Inc
Pons AD ProSci Inc
Postcentral gyrus ProSci Inc
Postcentral gyrus AD ProSci Inc
Precentral gyrus ProSci Inc
Precentral gyrus AD ProSci Inc
Spinal cord ProSci Inc
Temporal lobe ProSci Inc
Temporal lobe AD ProSci Inc
Thalamus ProSci Inc
Thalamus AD ProSci Inc
14
Amyloid beta Sigma Aldrich
AD
related Amyloid beta 10-20 Sigma Aldrich
Amyloid beta 1-12 Sigma Aldrich
Amyloid beta 12-28 Sigma Aldrich
Amyloid beta 1-23 Sigma Aldrich
Amyloid beta 1-38 Sigma Aldrich
Amyloid beta 17-40 Sigma Aldrich
Amyloid beta 25-35 Sigma Aldrich
Amyloid beta 34-42 Sigma Aldrich
Amyloid bri protein precursor 227 Sigma Aldrich
Amyloid DAN Protein Fragment 1-34 Sigma Aldrich
Amyloid Precursor Protein Sigma Aldrich
Amyloid protein no AB component Sigma Aldrich
Secreted amyloid precursor protein (SAP) beta Sigma Aldrich
Tau isoform variant 0N3R Sigma Aldrich
Tau isoform variant 1N3R Sigma Aldrich
Tau isoform variant 0N4R Sigma Aldrich
Tau isoform variant 2N3R Sigma Aldrich
Tau phospho Ser412 Sigma Aldrich
Tau phospho Ser441 Sigma Aldrich
Tau phospho Thr181 Sigma Aldrich
Tau Protein human EMD Biosciences
(plusmn)9-HODE Cayman Chemical
Lipids
1 Palmitoyl-2-(5oxo-Valeroyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
15a-hydroxycholestene Avanti Polar Lipids
15
15-ketocholestane Avanti Polar Lipids
15-ketocholestene Avanti Polar Lipids
1-Palmitoil-2-(9oxo-Nonanoyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Azelaoyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Glutaroyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
5 α-cholestane-3 β15 α-diol Avanti Polar Lipids
9(S)-HODE Cayman Chemical
Asialoganglioside-GM1 Sigma Aldrich
Asialoganglioside-GM2 Sigma Aldrich
Brain ceramides Avanti Polar Lipids
Brain D-erythrosphingosine Avanti Polar Lipids
Brain lysophosphatidylethanolamine Avanti Polar Lipids
Brain L-α-lysophosphatidylserine Avanti Polar Lipids
Brain L-α-phosphatidylcholine Avanti Polar Lipids
Brain L-α-phosphatidyl-ethanolamine Avanti Polar Lipids
Brain L-α-phosphatidylserine Avanti Polar Lipids
Brain polar lipid extract Avanti Polar Lipids
Brain sphingomyelin Avanti Polar Lipids
Brain sulfatide Avanti Polar Lipids
Brain total lipid extract Avanti Polar Lipids
Cardiolipin Sigma Aldrich
Ceramide Sigma Aldrich
Ceramide 1-phosphate Sigma Aldrich
Cholesterol Sigma Aldrich
16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-

3
70 kDa Heat Shock Protein peptide aa 181ndash199 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 195ndash214 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 210ndash229 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 225ndash244 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 240ndash259 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 255ndash275 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 271ndash290 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 286ndash305 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 301ndash320 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 31ndash50 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 316ndash335 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 331ndash350 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 346ndash365 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 361ndash380 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 376ndash395 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 391ndash410 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 406ndash425 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 421ndash440 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 436ndash455 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 451ndash470 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 466ndash485 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 46ndash65 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 481ndash500 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 496ndash515 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 511ndash530 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 526ndash545 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 541ndash560 Biopolymers Facility Harvard Medical School
4
70 kDa Heat Shock Protein peptide aa 556ndash575 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 571ndash590 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 586ndash605 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 601ndash620 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 616ndash635 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 631ndash640 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
71 kDa Heat Shock Protein M tuberculosis Biopolymers Facility Harvard Medical School
90 kDa Heat Shock Protein Stressgen
GroEL Stressgen
23-cyclic nucleotide 3-phosphodiesterase
peptide aa106-125 Biopolymers Facility Harvard Medical School
CNS
23-cyclic nucleotide 3-phosphodiesterase
peptide aa1-20 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa121-140 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa136-155 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa151-170 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa16-35 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa166-185 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase Biopolymers Facility Harvard Medical School
5
peptide aa181-200
23-cyclic nucleotide 3-phosphodiesterase
peptide aa196-215 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa211-230 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa226-245 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa241-260 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa256-275 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa271-290 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa286-305 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa301-320 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa31-50 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa316-335 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa331-350 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa346-365 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa361-380 Biopolymers Facility Harvard Medical School
6
23-cyclic nucleotide 3-phosphodiesterase
peptide aa376-395 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa391-410 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa406-421 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa46-65 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa61-80 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa76-95 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa91-110 Biopolymers Facility Harvard Medical School
Acetyl Cholinesterase Sigma Aldrich
ADAM-10 Sigma Aldrich
alpha-Cristallin Stressgen
beta-Cristallin Sigma Aldrich
bovine Myelin Basic Protein Sigma Aldrich
Brain Extract I Sigma Aldrich
Brain Extract II Sigma Aldrich
Brain Extract III Sigma Aldrich
Glial Filament Acidic Protein Research Diagnostic
guinea pig Myelin Basic Protein Sigma Aldrich
human Myelin Basic Protein Sigma Aldrich
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 106-125 Biopolymers Facility Harvard Medical School
7
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 121-140 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 136-155 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 151-170 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 166-185 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 181-200 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 91-110 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
106-125 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa 1- Biopolymers Facility Harvard Medical School
8
20
Myelinoligodendrocyte glycoprotein peptide aa
121-140 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
136-155 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
151-170 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
16-35 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
166-185 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
181-200 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
196-215 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
211-230 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
226-247 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
31-50 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
35-55 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
46-65 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
61-80 Biopolymers Facility Harvard Medical School
9
Myelinoligodendrocyte glycoprotein peptide aa
76-95 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
91-110 Biopolymers Facility Harvard Medical School
murine Myelin Basic Protein Sigma Aldrich
Myelin Associated Glycoprotein Sigma Aldrich
Myelin Basic Protein peptide aa 104-123 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 11-30 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 113-132 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 121-138 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 124-142 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 138-147 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 141-161 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 143-168 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 155-178 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 26-35 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 41-60 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 51-70 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 71-92 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 84-94 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 89-101 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 173-186 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 93-112 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 106-125 Biopolymers Facility Harvard Medical School
10
Myelin Protein 2 peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 121-132 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 91-110 Biopolymers Facility Harvard Medical School
Neurofilament 160kd Chemicon
Neurofilament 200kd Chemicon
Neurofilament 68kd Chemicon
Neuronal Enolase Calbiochem
Nicastrin GeneTex
NMDA receptor Novus Biologicals
NOGO Sigma Aldrich
Olygodendrocyte-Specific Protein peptide aa 106ndash
125 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 121ndash
140 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 136ndash
155 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 151ndash
170 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 16ndash
35 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 166ndash Biopolymers Facility Harvard Medical School
11
185
Olygodendrocyte-Specific Protein peptide aa 181ndash
199 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 195ndash
217 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 31ndash
50 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 46ndash
65 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 61ndash
80 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 76ndash
95 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 91ndash
110 Biopolymers Facility Harvard Medical School
Proteolipid Protein Abnova
Proteolipid Protein peptide aa 100-119 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 10-29 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 110-129 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 1-19 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 125-141 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-150 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-154 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 150-163 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 151-173 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 158-166 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 161-180 Biopolymers Facility Harvard Medical School
12
Proteolipid Protein peptide aa 178-191 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 180-199 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 190-209 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 20-39 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 205-220 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 215-232 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-239 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-249 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 248-259 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 250-269 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 265-277 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 35-50 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 40-59 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 50-69 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 65-84 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 80-99 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 91-110 Biopolymers Facility Harvard Medical School
Retinol Binding Protein Sigma Aldrich
S100beta protein Assay Designs
Super Oxide Dismutase Sigma Aldrich
Synuclein beta Sigma Aldrich
Synuclein gamma Sigma Aldrich
Amydgala ProSci Inc
Tissue Amydgala AD ProSci Inc
Brain lysate ProSci Inc
Brain Tissue Membrane ProSci Inc
Cerebellar pedunculus ProSci Inc
13
Cerebral meninges ProSci Inc
Corpus Callosum ProSci Inc
Corpus Callosum AD ProSci Inc
Diencephalon ProSci Inc
Fetal brain ProSci Inc
Frontal lobe ProSci Inc
Frontal lobe AD ProSci Inc
Hippocampus ProSci Inc
Hippocampus AD ProSci Inc
Insula ProSci Inc
Occipital lobe ProSci Inc
Occipital lobe AD ProSci Inc
Olfactory region ProSci Inc
Optic Nerve ProSci Inc
Parietal lobe ProSci Inc
Parietal lobe AD ProSci Inc
Pons ProSci Inc
Pons AD ProSci Inc
Postcentral gyrus ProSci Inc
Postcentral gyrus AD ProSci Inc
Precentral gyrus ProSci Inc
Precentral gyrus AD ProSci Inc
Spinal cord ProSci Inc
Temporal lobe ProSci Inc
Temporal lobe AD ProSci Inc
Thalamus ProSci Inc
Thalamus AD ProSci Inc
14
Amyloid beta Sigma Aldrich
AD
related Amyloid beta 10-20 Sigma Aldrich
Amyloid beta 1-12 Sigma Aldrich
Amyloid beta 12-28 Sigma Aldrich
Amyloid beta 1-23 Sigma Aldrich
Amyloid beta 1-38 Sigma Aldrich
Amyloid beta 17-40 Sigma Aldrich
Amyloid beta 25-35 Sigma Aldrich
Amyloid beta 34-42 Sigma Aldrich
Amyloid bri protein precursor 227 Sigma Aldrich
Amyloid DAN Protein Fragment 1-34 Sigma Aldrich
Amyloid Precursor Protein Sigma Aldrich
Amyloid protein no AB component Sigma Aldrich
Secreted amyloid precursor protein (SAP) beta Sigma Aldrich
Tau isoform variant 0N3R Sigma Aldrich
Tau isoform variant 1N3R Sigma Aldrich
Tau isoform variant 0N4R Sigma Aldrich
Tau isoform variant 2N3R Sigma Aldrich
Tau phospho Ser412 Sigma Aldrich
Tau phospho Ser441 Sigma Aldrich
Tau phospho Thr181 Sigma Aldrich
Tau Protein human EMD Biosciences
(plusmn)9-HODE Cayman Chemical
Lipids
1 Palmitoyl-2-(5oxo-Valeroyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
15a-hydroxycholestene Avanti Polar Lipids
15
15-ketocholestane Avanti Polar Lipids
15-ketocholestene Avanti Polar Lipids
1-Palmitoil-2-(9oxo-Nonanoyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Azelaoyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Glutaroyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
5 α-cholestane-3 β15 α-diol Avanti Polar Lipids
9(S)-HODE Cayman Chemical
Asialoganglioside-GM1 Sigma Aldrich
Asialoganglioside-GM2 Sigma Aldrich
Brain ceramides Avanti Polar Lipids
Brain D-erythrosphingosine Avanti Polar Lipids
Brain lysophosphatidylethanolamine Avanti Polar Lipids
Brain L-α-lysophosphatidylserine Avanti Polar Lipids
Brain L-α-phosphatidylcholine Avanti Polar Lipids
Brain L-α-phosphatidyl-ethanolamine Avanti Polar Lipids
Brain L-α-phosphatidylserine Avanti Polar Lipids
Brain polar lipid extract Avanti Polar Lipids
Brain sphingomyelin Avanti Polar Lipids
Brain sulfatide Avanti Polar Lipids
Brain total lipid extract Avanti Polar Lipids
Cardiolipin Sigma Aldrich
Ceramide Sigma Aldrich
Ceramide 1-phosphate Sigma Aldrich
Cholesterol Sigma Aldrich
16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-

4
70 kDa Heat Shock Protein peptide aa 556ndash575 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 571ndash590 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 586ndash605 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 601ndash620 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 616ndash635 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 61ndash80 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 631ndash640 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 76ndash95 Biopolymers Facility Harvard Medical School
70 kDa Heat Shock Protein peptide aa 91ndash110 Biopolymers Facility Harvard Medical School
71 kDa Heat Shock Protein M tuberculosis Biopolymers Facility Harvard Medical School
90 kDa Heat Shock Protein Stressgen
GroEL Stressgen
23-cyclic nucleotide 3-phosphodiesterase
peptide aa106-125 Biopolymers Facility Harvard Medical School
CNS
23-cyclic nucleotide 3-phosphodiesterase
peptide aa1-20 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa121-140 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa136-155 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa151-170 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa16-35 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa166-185 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase Biopolymers Facility Harvard Medical School
5
peptide aa181-200
23-cyclic nucleotide 3-phosphodiesterase
peptide aa196-215 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa211-230 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa226-245 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa241-260 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa256-275 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa271-290 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa286-305 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa301-320 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa31-50 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa316-335 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa331-350 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa346-365 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa361-380 Biopolymers Facility Harvard Medical School
6
23-cyclic nucleotide 3-phosphodiesterase
peptide aa376-395 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa391-410 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa406-421 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa46-65 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa61-80 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa76-95 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa91-110 Biopolymers Facility Harvard Medical School
Acetyl Cholinesterase Sigma Aldrich
ADAM-10 Sigma Aldrich
alpha-Cristallin Stressgen
beta-Cristallin Sigma Aldrich
bovine Myelin Basic Protein Sigma Aldrich
Brain Extract I Sigma Aldrich
Brain Extract II Sigma Aldrich
Brain Extract III Sigma Aldrich
Glial Filament Acidic Protein Research Diagnostic
guinea pig Myelin Basic Protein Sigma Aldrich
human Myelin Basic Protein Sigma Aldrich
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 106-125 Biopolymers Facility Harvard Medical School
7
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 121-140 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 136-155 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 151-170 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 166-185 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 181-200 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 91-110 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
106-125 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa 1- Biopolymers Facility Harvard Medical School
8
20
Myelinoligodendrocyte glycoprotein peptide aa
121-140 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
136-155 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
151-170 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
16-35 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
166-185 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
181-200 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
196-215 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
211-230 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
226-247 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
31-50 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
35-55 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
46-65 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
61-80 Biopolymers Facility Harvard Medical School
9
Myelinoligodendrocyte glycoprotein peptide aa
76-95 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
91-110 Biopolymers Facility Harvard Medical School
murine Myelin Basic Protein Sigma Aldrich
Myelin Associated Glycoprotein Sigma Aldrich
Myelin Basic Protein peptide aa 104-123 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 11-30 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 113-132 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 121-138 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 124-142 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 138-147 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 141-161 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 143-168 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 155-178 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 26-35 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 41-60 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 51-70 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 71-92 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 84-94 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 89-101 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 173-186 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 93-112 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 106-125 Biopolymers Facility Harvard Medical School
10
Myelin Protein 2 peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 121-132 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 91-110 Biopolymers Facility Harvard Medical School
Neurofilament 160kd Chemicon
Neurofilament 200kd Chemicon
Neurofilament 68kd Chemicon
Neuronal Enolase Calbiochem
Nicastrin GeneTex
NMDA receptor Novus Biologicals
NOGO Sigma Aldrich
Olygodendrocyte-Specific Protein peptide aa 106ndash
125 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 121ndash
140 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 136ndash
155 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 151ndash
170 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 16ndash
35 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 166ndash Biopolymers Facility Harvard Medical School
11
185
Olygodendrocyte-Specific Protein peptide aa 181ndash
199 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 195ndash
217 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 31ndash
50 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 46ndash
65 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 61ndash
80 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 76ndash
95 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 91ndash
110 Biopolymers Facility Harvard Medical School
Proteolipid Protein Abnova
Proteolipid Protein peptide aa 100-119 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 10-29 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 110-129 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 1-19 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 125-141 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-150 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-154 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 150-163 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 151-173 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 158-166 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 161-180 Biopolymers Facility Harvard Medical School
12
Proteolipid Protein peptide aa 178-191 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 180-199 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 190-209 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 20-39 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 205-220 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 215-232 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-239 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-249 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 248-259 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 250-269 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 265-277 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 35-50 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 40-59 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 50-69 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 65-84 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 80-99 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 91-110 Biopolymers Facility Harvard Medical School
Retinol Binding Protein Sigma Aldrich
S100beta protein Assay Designs
Super Oxide Dismutase Sigma Aldrich
Synuclein beta Sigma Aldrich
Synuclein gamma Sigma Aldrich
Amydgala ProSci Inc
Tissue Amydgala AD ProSci Inc
Brain lysate ProSci Inc
Brain Tissue Membrane ProSci Inc
Cerebellar pedunculus ProSci Inc
13
Cerebral meninges ProSci Inc
Corpus Callosum ProSci Inc
Corpus Callosum AD ProSci Inc
Diencephalon ProSci Inc
Fetal brain ProSci Inc
Frontal lobe ProSci Inc
Frontal lobe AD ProSci Inc
Hippocampus ProSci Inc
Hippocampus AD ProSci Inc
Insula ProSci Inc
Occipital lobe ProSci Inc
Occipital lobe AD ProSci Inc
Olfactory region ProSci Inc
Optic Nerve ProSci Inc
Parietal lobe ProSci Inc
Parietal lobe AD ProSci Inc
Pons ProSci Inc
Pons AD ProSci Inc
Postcentral gyrus ProSci Inc
Postcentral gyrus AD ProSci Inc
Precentral gyrus ProSci Inc
Precentral gyrus AD ProSci Inc
Spinal cord ProSci Inc
Temporal lobe ProSci Inc
Temporal lobe AD ProSci Inc
Thalamus ProSci Inc
Thalamus AD ProSci Inc
14
Amyloid beta Sigma Aldrich
AD
related Amyloid beta 10-20 Sigma Aldrich
Amyloid beta 1-12 Sigma Aldrich
Amyloid beta 12-28 Sigma Aldrich
Amyloid beta 1-23 Sigma Aldrich
Amyloid beta 1-38 Sigma Aldrich
Amyloid beta 17-40 Sigma Aldrich
Amyloid beta 25-35 Sigma Aldrich
Amyloid beta 34-42 Sigma Aldrich
Amyloid bri protein precursor 227 Sigma Aldrich
Amyloid DAN Protein Fragment 1-34 Sigma Aldrich
Amyloid Precursor Protein Sigma Aldrich
Amyloid protein no AB component Sigma Aldrich
Secreted amyloid precursor protein (SAP) beta Sigma Aldrich
Tau isoform variant 0N3R Sigma Aldrich
Tau isoform variant 1N3R Sigma Aldrich
Tau isoform variant 0N4R Sigma Aldrich
Tau isoform variant 2N3R Sigma Aldrich
Tau phospho Ser412 Sigma Aldrich
Tau phospho Ser441 Sigma Aldrich
Tau phospho Thr181 Sigma Aldrich
Tau Protein human EMD Biosciences
(plusmn)9-HODE Cayman Chemical
Lipids
1 Palmitoyl-2-(5oxo-Valeroyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
15a-hydroxycholestene Avanti Polar Lipids
15
15-ketocholestane Avanti Polar Lipids
15-ketocholestene Avanti Polar Lipids
1-Palmitoil-2-(9oxo-Nonanoyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Azelaoyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Glutaroyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
5 α-cholestane-3 β15 α-diol Avanti Polar Lipids
9(S)-HODE Cayman Chemical
Asialoganglioside-GM1 Sigma Aldrich
Asialoganglioside-GM2 Sigma Aldrich
Brain ceramides Avanti Polar Lipids
Brain D-erythrosphingosine Avanti Polar Lipids
Brain lysophosphatidylethanolamine Avanti Polar Lipids
Brain L-α-lysophosphatidylserine Avanti Polar Lipids
Brain L-α-phosphatidylcholine Avanti Polar Lipids
Brain L-α-phosphatidyl-ethanolamine Avanti Polar Lipids
Brain L-α-phosphatidylserine Avanti Polar Lipids
Brain polar lipid extract Avanti Polar Lipids
Brain sphingomyelin Avanti Polar Lipids
Brain sulfatide Avanti Polar Lipids
Brain total lipid extract Avanti Polar Lipids
Cardiolipin Sigma Aldrich
Ceramide Sigma Aldrich
Ceramide 1-phosphate Sigma Aldrich
Cholesterol Sigma Aldrich
16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-

5
peptide aa181-200
23-cyclic nucleotide 3-phosphodiesterase
peptide aa196-215 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa211-230 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa226-245 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa241-260 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa256-275 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa271-290 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa286-305 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa301-320 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa31-50 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa316-335 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa331-350 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa346-365 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa361-380 Biopolymers Facility Harvard Medical School
6
23-cyclic nucleotide 3-phosphodiesterase
peptide aa376-395 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa391-410 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa406-421 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa46-65 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa61-80 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa76-95 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa91-110 Biopolymers Facility Harvard Medical School
Acetyl Cholinesterase Sigma Aldrich
ADAM-10 Sigma Aldrich
alpha-Cristallin Stressgen
beta-Cristallin Sigma Aldrich
bovine Myelin Basic Protein Sigma Aldrich
Brain Extract I Sigma Aldrich
Brain Extract II Sigma Aldrich
Brain Extract III Sigma Aldrich
Glial Filament Acidic Protein Research Diagnostic
guinea pig Myelin Basic Protein Sigma Aldrich
human Myelin Basic Protein Sigma Aldrich
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 106-125 Biopolymers Facility Harvard Medical School
7
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 121-140 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 136-155 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 151-170 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 166-185 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 181-200 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 91-110 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
106-125 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa 1- Biopolymers Facility Harvard Medical School
8
20
Myelinoligodendrocyte glycoprotein peptide aa
121-140 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
136-155 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
151-170 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
16-35 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
166-185 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
181-200 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
196-215 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
211-230 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
226-247 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
31-50 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
35-55 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
46-65 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
61-80 Biopolymers Facility Harvard Medical School
9
Myelinoligodendrocyte glycoprotein peptide aa
76-95 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
91-110 Biopolymers Facility Harvard Medical School
murine Myelin Basic Protein Sigma Aldrich
Myelin Associated Glycoprotein Sigma Aldrich
Myelin Basic Protein peptide aa 104-123 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 11-30 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 113-132 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 121-138 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 124-142 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 138-147 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 141-161 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 143-168 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 155-178 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 26-35 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 41-60 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 51-70 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 71-92 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 84-94 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 89-101 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 173-186 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 93-112 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 106-125 Biopolymers Facility Harvard Medical School
10
Myelin Protein 2 peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 121-132 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 91-110 Biopolymers Facility Harvard Medical School
Neurofilament 160kd Chemicon
Neurofilament 200kd Chemicon
Neurofilament 68kd Chemicon
Neuronal Enolase Calbiochem
Nicastrin GeneTex
NMDA receptor Novus Biologicals
NOGO Sigma Aldrich
Olygodendrocyte-Specific Protein peptide aa 106ndash
125 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 121ndash
140 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 136ndash
155 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 151ndash
170 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 16ndash
35 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 166ndash Biopolymers Facility Harvard Medical School
11
185
Olygodendrocyte-Specific Protein peptide aa 181ndash
199 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 195ndash
217 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 31ndash
50 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 46ndash
65 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 61ndash
80 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 76ndash
95 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 91ndash
110 Biopolymers Facility Harvard Medical School
Proteolipid Protein Abnova
Proteolipid Protein peptide aa 100-119 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 10-29 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 110-129 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 1-19 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 125-141 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-150 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-154 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 150-163 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 151-173 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 158-166 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 161-180 Biopolymers Facility Harvard Medical School
12
Proteolipid Protein peptide aa 178-191 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 180-199 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 190-209 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 20-39 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 205-220 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 215-232 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-239 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-249 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 248-259 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 250-269 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 265-277 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 35-50 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 40-59 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 50-69 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 65-84 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 80-99 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 91-110 Biopolymers Facility Harvard Medical School
Retinol Binding Protein Sigma Aldrich
S100beta protein Assay Designs
Super Oxide Dismutase Sigma Aldrich
Synuclein beta Sigma Aldrich
Synuclein gamma Sigma Aldrich
Amydgala ProSci Inc
Tissue Amydgala AD ProSci Inc
Brain lysate ProSci Inc
Brain Tissue Membrane ProSci Inc
Cerebellar pedunculus ProSci Inc
13
Cerebral meninges ProSci Inc
Corpus Callosum ProSci Inc
Corpus Callosum AD ProSci Inc
Diencephalon ProSci Inc
Fetal brain ProSci Inc
Frontal lobe ProSci Inc
Frontal lobe AD ProSci Inc
Hippocampus ProSci Inc
Hippocampus AD ProSci Inc
Insula ProSci Inc
Occipital lobe ProSci Inc
Occipital lobe AD ProSci Inc
Olfactory region ProSci Inc
Optic Nerve ProSci Inc
Parietal lobe ProSci Inc
Parietal lobe AD ProSci Inc
Pons ProSci Inc
Pons AD ProSci Inc
Postcentral gyrus ProSci Inc
Postcentral gyrus AD ProSci Inc
Precentral gyrus ProSci Inc
Precentral gyrus AD ProSci Inc
Spinal cord ProSci Inc
Temporal lobe ProSci Inc
Temporal lobe AD ProSci Inc
Thalamus ProSci Inc
Thalamus AD ProSci Inc
14
Amyloid beta Sigma Aldrich
AD
related Amyloid beta 10-20 Sigma Aldrich
Amyloid beta 1-12 Sigma Aldrich
Amyloid beta 12-28 Sigma Aldrich
Amyloid beta 1-23 Sigma Aldrich
Amyloid beta 1-38 Sigma Aldrich
Amyloid beta 17-40 Sigma Aldrich
Amyloid beta 25-35 Sigma Aldrich
Amyloid beta 34-42 Sigma Aldrich
Amyloid bri protein precursor 227 Sigma Aldrich
Amyloid DAN Protein Fragment 1-34 Sigma Aldrich
Amyloid Precursor Protein Sigma Aldrich
Amyloid protein no AB component Sigma Aldrich
Secreted amyloid precursor protein (SAP) beta Sigma Aldrich
Tau isoform variant 0N3R Sigma Aldrich
Tau isoform variant 1N3R Sigma Aldrich
Tau isoform variant 0N4R Sigma Aldrich
Tau isoform variant 2N3R Sigma Aldrich
Tau phospho Ser412 Sigma Aldrich
Tau phospho Ser441 Sigma Aldrich
Tau phospho Thr181 Sigma Aldrich
Tau Protein human EMD Biosciences
(plusmn)9-HODE Cayman Chemical
Lipids
1 Palmitoyl-2-(5oxo-Valeroyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
15a-hydroxycholestene Avanti Polar Lipids
15
15-ketocholestane Avanti Polar Lipids
15-ketocholestene Avanti Polar Lipids
1-Palmitoil-2-(9oxo-Nonanoyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Azelaoyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Glutaroyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
5 α-cholestane-3 β15 α-diol Avanti Polar Lipids
9(S)-HODE Cayman Chemical
Asialoganglioside-GM1 Sigma Aldrich
Asialoganglioside-GM2 Sigma Aldrich
Brain ceramides Avanti Polar Lipids
Brain D-erythrosphingosine Avanti Polar Lipids
Brain lysophosphatidylethanolamine Avanti Polar Lipids
Brain L-α-lysophosphatidylserine Avanti Polar Lipids
Brain L-α-phosphatidylcholine Avanti Polar Lipids
Brain L-α-phosphatidyl-ethanolamine Avanti Polar Lipids
Brain L-α-phosphatidylserine Avanti Polar Lipids
Brain polar lipid extract Avanti Polar Lipids
Brain sphingomyelin Avanti Polar Lipids
Brain sulfatide Avanti Polar Lipids
Brain total lipid extract Avanti Polar Lipids
Cardiolipin Sigma Aldrich
Ceramide Sigma Aldrich
Ceramide 1-phosphate Sigma Aldrich
Cholesterol Sigma Aldrich
16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-

6
23-cyclic nucleotide 3-phosphodiesterase
peptide aa376-395 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa391-410 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa406-421 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa46-65 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa61-80 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa76-95 Biopolymers Facility Harvard Medical School
23-cyclic nucleotide 3-phosphodiesterase
peptide aa91-110 Biopolymers Facility Harvard Medical School
Acetyl Cholinesterase Sigma Aldrich
ADAM-10 Sigma Aldrich
alpha-Cristallin Stressgen
beta-Cristallin Sigma Aldrich
bovine Myelin Basic Protein Sigma Aldrich
Brain Extract I Sigma Aldrich
Brain Extract II Sigma Aldrich
Brain Extract III Sigma Aldrich
Glial Filament Acidic Protein Research Diagnostic
guinea pig Myelin Basic Protein Sigma Aldrich
human Myelin Basic Protein Sigma Aldrich
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 106-125 Biopolymers Facility Harvard Medical School
7
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 121-140 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 136-155 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 151-170 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 166-185 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 181-200 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 91-110 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
106-125 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa 1- Biopolymers Facility Harvard Medical School
8
20
Myelinoligodendrocyte glycoprotein peptide aa
121-140 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
136-155 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
151-170 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
16-35 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
166-185 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
181-200 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
196-215 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
211-230 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
226-247 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
31-50 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
35-55 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
46-65 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
61-80 Biopolymers Facility Harvard Medical School
9
Myelinoligodendrocyte glycoprotein peptide aa
76-95 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
91-110 Biopolymers Facility Harvard Medical School
murine Myelin Basic Protein Sigma Aldrich
Myelin Associated Glycoprotein Sigma Aldrich
Myelin Basic Protein peptide aa 104-123 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 11-30 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 113-132 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 121-138 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 124-142 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 138-147 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 141-161 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 143-168 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 155-178 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 26-35 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 41-60 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 51-70 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 71-92 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 84-94 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 89-101 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 173-186 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 93-112 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 106-125 Biopolymers Facility Harvard Medical School
10
Myelin Protein 2 peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 121-132 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 91-110 Biopolymers Facility Harvard Medical School
Neurofilament 160kd Chemicon
Neurofilament 200kd Chemicon
Neurofilament 68kd Chemicon
Neuronal Enolase Calbiochem
Nicastrin GeneTex
NMDA receptor Novus Biologicals
NOGO Sigma Aldrich
Olygodendrocyte-Specific Protein peptide aa 106ndash
125 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 121ndash
140 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 136ndash
155 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 151ndash
170 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 16ndash
35 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 166ndash Biopolymers Facility Harvard Medical School
11
185
Olygodendrocyte-Specific Protein peptide aa 181ndash
199 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 195ndash
217 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 31ndash
50 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 46ndash
65 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 61ndash
80 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 76ndash
95 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 91ndash
110 Biopolymers Facility Harvard Medical School
Proteolipid Protein Abnova
Proteolipid Protein peptide aa 100-119 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 10-29 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 110-129 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 1-19 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 125-141 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-150 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-154 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 150-163 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 151-173 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 158-166 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 161-180 Biopolymers Facility Harvard Medical School
12
Proteolipid Protein peptide aa 178-191 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 180-199 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 190-209 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 20-39 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 205-220 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 215-232 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-239 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-249 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 248-259 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 250-269 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 265-277 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 35-50 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 40-59 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 50-69 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 65-84 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 80-99 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 91-110 Biopolymers Facility Harvard Medical School
Retinol Binding Protein Sigma Aldrich
S100beta protein Assay Designs
Super Oxide Dismutase Sigma Aldrich
Synuclein beta Sigma Aldrich
Synuclein gamma Sigma Aldrich
Amydgala ProSci Inc
Tissue Amydgala AD ProSci Inc
Brain lysate ProSci Inc
Brain Tissue Membrane ProSci Inc
Cerebellar pedunculus ProSci Inc
13
Cerebral meninges ProSci Inc
Corpus Callosum ProSci Inc
Corpus Callosum AD ProSci Inc
Diencephalon ProSci Inc
Fetal brain ProSci Inc
Frontal lobe ProSci Inc
Frontal lobe AD ProSci Inc
Hippocampus ProSci Inc
Hippocampus AD ProSci Inc
Insula ProSci Inc
Occipital lobe ProSci Inc
Occipital lobe AD ProSci Inc
Olfactory region ProSci Inc
Optic Nerve ProSci Inc
Parietal lobe ProSci Inc
Parietal lobe AD ProSci Inc
Pons ProSci Inc
Pons AD ProSci Inc
Postcentral gyrus ProSci Inc
Postcentral gyrus AD ProSci Inc
Precentral gyrus ProSci Inc
Precentral gyrus AD ProSci Inc
Spinal cord ProSci Inc
Temporal lobe ProSci Inc
Temporal lobe AD ProSci Inc
Thalamus ProSci Inc
Thalamus AD ProSci Inc
14
Amyloid beta Sigma Aldrich
AD
related Amyloid beta 10-20 Sigma Aldrich
Amyloid beta 1-12 Sigma Aldrich
Amyloid beta 12-28 Sigma Aldrich
Amyloid beta 1-23 Sigma Aldrich
Amyloid beta 1-38 Sigma Aldrich
Amyloid beta 17-40 Sigma Aldrich
Amyloid beta 25-35 Sigma Aldrich
Amyloid beta 34-42 Sigma Aldrich
Amyloid bri protein precursor 227 Sigma Aldrich
Amyloid DAN Protein Fragment 1-34 Sigma Aldrich
Amyloid Precursor Protein Sigma Aldrich
Amyloid protein no AB component Sigma Aldrich
Secreted amyloid precursor protein (SAP) beta Sigma Aldrich
Tau isoform variant 0N3R Sigma Aldrich
Tau isoform variant 1N3R Sigma Aldrich
Tau isoform variant 0N4R Sigma Aldrich
Tau isoform variant 2N3R Sigma Aldrich
Tau phospho Ser412 Sigma Aldrich
Tau phospho Ser441 Sigma Aldrich
Tau phospho Thr181 Sigma Aldrich
Tau Protein human EMD Biosciences
(plusmn)9-HODE Cayman Chemical
Lipids
1 Palmitoyl-2-(5oxo-Valeroyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
15a-hydroxycholestene Avanti Polar Lipids
15
15-ketocholestane Avanti Polar Lipids
15-ketocholestene Avanti Polar Lipids
1-Palmitoil-2-(9oxo-Nonanoyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Azelaoyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Glutaroyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
5 α-cholestane-3 β15 α-diol Avanti Polar Lipids
9(S)-HODE Cayman Chemical
Asialoganglioside-GM1 Sigma Aldrich
Asialoganglioside-GM2 Sigma Aldrich
Brain ceramides Avanti Polar Lipids
Brain D-erythrosphingosine Avanti Polar Lipids
Brain lysophosphatidylethanolamine Avanti Polar Lipids
Brain L-α-lysophosphatidylserine Avanti Polar Lipids
Brain L-α-phosphatidylcholine Avanti Polar Lipids
Brain L-α-phosphatidyl-ethanolamine Avanti Polar Lipids
Brain L-α-phosphatidylserine Avanti Polar Lipids
Brain polar lipid extract Avanti Polar Lipids
Brain sphingomyelin Avanti Polar Lipids
Brain sulfatide Avanti Polar Lipids
Brain total lipid extract Avanti Polar Lipids
Cardiolipin Sigma Aldrich
Ceramide Sigma Aldrich
Ceramide 1-phosphate Sigma Aldrich
Cholesterol Sigma Aldrich
16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-

7
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 121-140 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 136-155 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 151-170 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 166-185 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 181-200 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin-Associated Oligodendrocytic Basic Protein
peptide aa 91-110 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
106-125 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa 1- Biopolymers Facility Harvard Medical School
8
20
Myelinoligodendrocyte glycoprotein peptide aa
121-140 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
136-155 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
151-170 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
16-35 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
166-185 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
181-200 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
196-215 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
211-230 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
226-247 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
31-50 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
35-55 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
46-65 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
61-80 Biopolymers Facility Harvard Medical School
9
Myelinoligodendrocyte glycoprotein peptide aa
76-95 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
91-110 Biopolymers Facility Harvard Medical School
murine Myelin Basic Protein Sigma Aldrich
Myelin Associated Glycoprotein Sigma Aldrich
Myelin Basic Protein peptide aa 104-123 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 11-30 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 113-132 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 121-138 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 124-142 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 138-147 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 141-161 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 143-168 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 155-178 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 26-35 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 41-60 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 51-70 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 71-92 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 84-94 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 89-101 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 173-186 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 93-112 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 106-125 Biopolymers Facility Harvard Medical School
10
Myelin Protein 2 peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 121-132 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 91-110 Biopolymers Facility Harvard Medical School
Neurofilament 160kd Chemicon
Neurofilament 200kd Chemicon
Neurofilament 68kd Chemicon
Neuronal Enolase Calbiochem
Nicastrin GeneTex
NMDA receptor Novus Biologicals
NOGO Sigma Aldrich
Olygodendrocyte-Specific Protein peptide aa 106ndash
125 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 121ndash
140 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 136ndash
155 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 151ndash
170 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 16ndash
35 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 166ndash Biopolymers Facility Harvard Medical School
11
185
Olygodendrocyte-Specific Protein peptide aa 181ndash
199 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 195ndash
217 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 31ndash
50 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 46ndash
65 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 61ndash
80 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 76ndash
95 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 91ndash
110 Biopolymers Facility Harvard Medical School
Proteolipid Protein Abnova
Proteolipid Protein peptide aa 100-119 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 10-29 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 110-129 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 1-19 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 125-141 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-150 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-154 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 150-163 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 151-173 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 158-166 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 161-180 Biopolymers Facility Harvard Medical School
12
Proteolipid Protein peptide aa 178-191 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 180-199 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 190-209 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 20-39 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 205-220 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 215-232 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-239 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-249 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 248-259 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 250-269 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 265-277 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 35-50 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 40-59 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 50-69 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 65-84 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 80-99 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 91-110 Biopolymers Facility Harvard Medical School
Retinol Binding Protein Sigma Aldrich
S100beta protein Assay Designs
Super Oxide Dismutase Sigma Aldrich
Synuclein beta Sigma Aldrich
Synuclein gamma Sigma Aldrich
Amydgala ProSci Inc
Tissue Amydgala AD ProSci Inc
Brain lysate ProSci Inc
Brain Tissue Membrane ProSci Inc
Cerebellar pedunculus ProSci Inc
13
Cerebral meninges ProSci Inc
Corpus Callosum ProSci Inc
Corpus Callosum AD ProSci Inc
Diencephalon ProSci Inc
Fetal brain ProSci Inc
Frontal lobe ProSci Inc
Frontal lobe AD ProSci Inc
Hippocampus ProSci Inc
Hippocampus AD ProSci Inc
Insula ProSci Inc
Occipital lobe ProSci Inc
Occipital lobe AD ProSci Inc
Olfactory region ProSci Inc
Optic Nerve ProSci Inc
Parietal lobe ProSci Inc
Parietal lobe AD ProSci Inc
Pons ProSci Inc
Pons AD ProSci Inc
Postcentral gyrus ProSci Inc
Postcentral gyrus AD ProSci Inc
Precentral gyrus ProSci Inc
Precentral gyrus AD ProSci Inc
Spinal cord ProSci Inc
Temporal lobe ProSci Inc
Temporal lobe AD ProSci Inc
Thalamus ProSci Inc
Thalamus AD ProSci Inc
14
Amyloid beta Sigma Aldrich
AD
related Amyloid beta 10-20 Sigma Aldrich
Amyloid beta 1-12 Sigma Aldrich
Amyloid beta 12-28 Sigma Aldrich
Amyloid beta 1-23 Sigma Aldrich
Amyloid beta 1-38 Sigma Aldrich
Amyloid beta 17-40 Sigma Aldrich
Amyloid beta 25-35 Sigma Aldrich
Amyloid beta 34-42 Sigma Aldrich
Amyloid bri protein precursor 227 Sigma Aldrich
Amyloid DAN Protein Fragment 1-34 Sigma Aldrich
Amyloid Precursor Protein Sigma Aldrich
Amyloid protein no AB component Sigma Aldrich
Secreted amyloid precursor protein (SAP) beta Sigma Aldrich
Tau isoform variant 0N3R Sigma Aldrich
Tau isoform variant 1N3R Sigma Aldrich
Tau isoform variant 0N4R Sigma Aldrich
Tau isoform variant 2N3R Sigma Aldrich
Tau phospho Ser412 Sigma Aldrich
Tau phospho Ser441 Sigma Aldrich
Tau phospho Thr181 Sigma Aldrich
Tau Protein human EMD Biosciences
(plusmn)9-HODE Cayman Chemical
Lipids
1 Palmitoyl-2-(5oxo-Valeroyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
15a-hydroxycholestene Avanti Polar Lipids
15
15-ketocholestane Avanti Polar Lipids
15-ketocholestene Avanti Polar Lipids
1-Palmitoil-2-(9oxo-Nonanoyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Azelaoyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Glutaroyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
5 α-cholestane-3 β15 α-diol Avanti Polar Lipids
9(S)-HODE Cayman Chemical
Asialoganglioside-GM1 Sigma Aldrich
Asialoganglioside-GM2 Sigma Aldrich
Brain ceramides Avanti Polar Lipids
Brain D-erythrosphingosine Avanti Polar Lipids
Brain lysophosphatidylethanolamine Avanti Polar Lipids
Brain L-α-lysophosphatidylserine Avanti Polar Lipids
Brain L-α-phosphatidylcholine Avanti Polar Lipids
Brain L-α-phosphatidyl-ethanolamine Avanti Polar Lipids
Brain L-α-phosphatidylserine Avanti Polar Lipids
Brain polar lipid extract Avanti Polar Lipids
Brain sphingomyelin Avanti Polar Lipids
Brain sulfatide Avanti Polar Lipids
Brain total lipid extract Avanti Polar Lipids
Cardiolipin Sigma Aldrich
Ceramide Sigma Aldrich
Ceramide 1-phosphate Sigma Aldrich
Cholesterol Sigma Aldrich
16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-

8
20
Myelinoligodendrocyte glycoprotein peptide aa
121-140 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
136-155 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
151-170 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
16-35 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
166-185 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
181-200 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
196-215 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
211-230 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
226-247 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
31-50 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
35-55 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
46-65 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
61-80 Biopolymers Facility Harvard Medical School
9
Myelinoligodendrocyte glycoprotein peptide aa
76-95 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
91-110 Biopolymers Facility Harvard Medical School
murine Myelin Basic Protein Sigma Aldrich
Myelin Associated Glycoprotein Sigma Aldrich
Myelin Basic Protein peptide aa 104-123 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 11-30 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 113-132 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 121-138 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 124-142 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 138-147 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 141-161 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 143-168 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 155-178 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 26-35 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 41-60 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 51-70 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 71-92 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 84-94 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 89-101 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 173-186 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 93-112 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 106-125 Biopolymers Facility Harvard Medical School
10
Myelin Protein 2 peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 121-132 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 91-110 Biopolymers Facility Harvard Medical School
Neurofilament 160kd Chemicon
Neurofilament 200kd Chemicon
Neurofilament 68kd Chemicon
Neuronal Enolase Calbiochem
Nicastrin GeneTex
NMDA receptor Novus Biologicals
NOGO Sigma Aldrich
Olygodendrocyte-Specific Protein peptide aa 106ndash
125 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 121ndash
140 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 136ndash
155 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 151ndash
170 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 16ndash
35 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 166ndash Biopolymers Facility Harvard Medical School
11
185
Olygodendrocyte-Specific Protein peptide aa 181ndash
199 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 195ndash
217 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 31ndash
50 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 46ndash
65 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 61ndash
80 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 76ndash
95 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 91ndash
110 Biopolymers Facility Harvard Medical School
Proteolipid Protein Abnova
Proteolipid Protein peptide aa 100-119 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 10-29 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 110-129 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 1-19 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 125-141 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-150 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-154 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 150-163 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 151-173 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 158-166 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 161-180 Biopolymers Facility Harvard Medical School
12
Proteolipid Protein peptide aa 178-191 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 180-199 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 190-209 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 20-39 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 205-220 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 215-232 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-239 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-249 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 248-259 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 250-269 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 265-277 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 35-50 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 40-59 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 50-69 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 65-84 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 80-99 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 91-110 Biopolymers Facility Harvard Medical School
Retinol Binding Protein Sigma Aldrich
S100beta protein Assay Designs
Super Oxide Dismutase Sigma Aldrich
Synuclein beta Sigma Aldrich
Synuclein gamma Sigma Aldrich
Amydgala ProSci Inc
Tissue Amydgala AD ProSci Inc
Brain lysate ProSci Inc
Brain Tissue Membrane ProSci Inc
Cerebellar pedunculus ProSci Inc
13
Cerebral meninges ProSci Inc
Corpus Callosum ProSci Inc
Corpus Callosum AD ProSci Inc
Diencephalon ProSci Inc
Fetal brain ProSci Inc
Frontal lobe ProSci Inc
Frontal lobe AD ProSci Inc
Hippocampus ProSci Inc
Hippocampus AD ProSci Inc
Insula ProSci Inc
Occipital lobe ProSci Inc
Occipital lobe AD ProSci Inc
Olfactory region ProSci Inc
Optic Nerve ProSci Inc
Parietal lobe ProSci Inc
Parietal lobe AD ProSci Inc
Pons ProSci Inc
Pons AD ProSci Inc
Postcentral gyrus ProSci Inc
Postcentral gyrus AD ProSci Inc
Precentral gyrus ProSci Inc
Precentral gyrus AD ProSci Inc
Spinal cord ProSci Inc
Temporal lobe ProSci Inc
Temporal lobe AD ProSci Inc
Thalamus ProSci Inc
Thalamus AD ProSci Inc
14
Amyloid beta Sigma Aldrich
AD
related Amyloid beta 10-20 Sigma Aldrich
Amyloid beta 1-12 Sigma Aldrich
Amyloid beta 12-28 Sigma Aldrich
Amyloid beta 1-23 Sigma Aldrich
Amyloid beta 1-38 Sigma Aldrich
Amyloid beta 17-40 Sigma Aldrich
Amyloid beta 25-35 Sigma Aldrich
Amyloid beta 34-42 Sigma Aldrich
Amyloid bri protein precursor 227 Sigma Aldrich
Amyloid DAN Protein Fragment 1-34 Sigma Aldrich
Amyloid Precursor Protein Sigma Aldrich
Amyloid protein no AB component Sigma Aldrich
Secreted amyloid precursor protein (SAP) beta Sigma Aldrich
Tau isoform variant 0N3R Sigma Aldrich
Tau isoform variant 1N3R Sigma Aldrich
Tau isoform variant 0N4R Sigma Aldrich
Tau isoform variant 2N3R Sigma Aldrich
Tau phospho Ser412 Sigma Aldrich
Tau phospho Ser441 Sigma Aldrich
Tau phospho Thr181 Sigma Aldrich
Tau Protein human EMD Biosciences
(plusmn)9-HODE Cayman Chemical
Lipids
1 Palmitoyl-2-(5oxo-Valeroyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
15a-hydroxycholestene Avanti Polar Lipids
15
15-ketocholestane Avanti Polar Lipids
15-ketocholestene Avanti Polar Lipids
1-Palmitoil-2-(9oxo-Nonanoyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Azelaoyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Glutaroyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
5 α-cholestane-3 β15 α-diol Avanti Polar Lipids
9(S)-HODE Cayman Chemical
Asialoganglioside-GM1 Sigma Aldrich
Asialoganglioside-GM2 Sigma Aldrich
Brain ceramides Avanti Polar Lipids
Brain D-erythrosphingosine Avanti Polar Lipids
Brain lysophosphatidylethanolamine Avanti Polar Lipids
Brain L-α-lysophosphatidylserine Avanti Polar Lipids
Brain L-α-phosphatidylcholine Avanti Polar Lipids
Brain L-α-phosphatidyl-ethanolamine Avanti Polar Lipids
Brain L-α-phosphatidylserine Avanti Polar Lipids
Brain polar lipid extract Avanti Polar Lipids
Brain sphingomyelin Avanti Polar Lipids
Brain sulfatide Avanti Polar Lipids
Brain total lipid extract Avanti Polar Lipids
Cardiolipin Sigma Aldrich
Ceramide Sigma Aldrich
Ceramide 1-phosphate Sigma Aldrich
Cholesterol Sigma Aldrich
16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-

9
Myelinoligodendrocyte glycoprotein peptide aa
76-95 Biopolymers Facility Harvard Medical School
Myelinoligodendrocyte glycoprotein peptide aa
91-110 Biopolymers Facility Harvard Medical School
murine Myelin Basic Protein Sigma Aldrich
Myelin Associated Glycoprotein Sigma Aldrich
Myelin Basic Protein peptide aa 104-123 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 11-30 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 113-132 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 121-138 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 124-142 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 138-147 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 141-161 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 143-168 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 155-178 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 26-35 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 41-60 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 51-70 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 71-92 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 84-94 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 89-101 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 173-186 Biopolymers Facility Harvard Medical School
Myelin Basic Protein peptide aa 93-112 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 106-125 Biopolymers Facility Harvard Medical School
10
Myelin Protein 2 peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 121-132 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 91-110 Biopolymers Facility Harvard Medical School
Neurofilament 160kd Chemicon
Neurofilament 200kd Chemicon
Neurofilament 68kd Chemicon
Neuronal Enolase Calbiochem
Nicastrin GeneTex
NMDA receptor Novus Biologicals
NOGO Sigma Aldrich
Olygodendrocyte-Specific Protein peptide aa 106ndash
125 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 121ndash
140 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 136ndash
155 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 151ndash
170 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 16ndash
35 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 166ndash Biopolymers Facility Harvard Medical School
11
185
Olygodendrocyte-Specific Protein peptide aa 181ndash
199 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 195ndash
217 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 31ndash
50 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 46ndash
65 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 61ndash
80 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 76ndash
95 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 91ndash
110 Biopolymers Facility Harvard Medical School
Proteolipid Protein Abnova
Proteolipid Protein peptide aa 100-119 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 10-29 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 110-129 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 1-19 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 125-141 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-150 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-154 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 150-163 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 151-173 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 158-166 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 161-180 Biopolymers Facility Harvard Medical School
12
Proteolipid Protein peptide aa 178-191 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 180-199 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 190-209 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 20-39 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 205-220 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 215-232 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-239 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-249 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 248-259 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 250-269 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 265-277 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 35-50 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 40-59 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 50-69 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 65-84 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 80-99 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 91-110 Biopolymers Facility Harvard Medical School
Retinol Binding Protein Sigma Aldrich
S100beta protein Assay Designs
Super Oxide Dismutase Sigma Aldrich
Synuclein beta Sigma Aldrich
Synuclein gamma Sigma Aldrich
Amydgala ProSci Inc
Tissue Amydgala AD ProSci Inc
Brain lysate ProSci Inc
Brain Tissue Membrane ProSci Inc
Cerebellar pedunculus ProSci Inc
13
Cerebral meninges ProSci Inc
Corpus Callosum ProSci Inc
Corpus Callosum AD ProSci Inc
Diencephalon ProSci Inc
Fetal brain ProSci Inc
Frontal lobe ProSci Inc
Frontal lobe AD ProSci Inc
Hippocampus ProSci Inc
Hippocampus AD ProSci Inc
Insula ProSci Inc
Occipital lobe ProSci Inc
Occipital lobe AD ProSci Inc
Olfactory region ProSci Inc
Optic Nerve ProSci Inc
Parietal lobe ProSci Inc
Parietal lobe AD ProSci Inc
Pons ProSci Inc
Pons AD ProSci Inc
Postcentral gyrus ProSci Inc
Postcentral gyrus AD ProSci Inc
Precentral gyrus ProSci Inc
Precentral gyrus AD ProSci Inc
Spinal cord ProSci Inc
Temporal lobe ProSci Inc
Temporal lobe AD ProSci Inc
Thalamus ProSci Inc
Thalamus AD ProSci Inc
14
Amyloid beta Sigma Aldrich
AD
related Amyloid beta 10-20 Sigma Aldrich
Amyloid beta 1-12 Sigma Aldrich
Amyloid beta 12-28 Sigma Aldrich
Amyloid beta 1-23 Sigma Aldrich
Amyloid beta 1-38 Sigma Aldrich
Amyloid beta 17-40 Sigma Aldrich
Amyloid beta 25-35 Sigma Aldrich
Amyloid beta 34-42 Sigma Aldrich
Amyloid bri protein precursor 227 Sigma Aldrich
Amyloid DAN Protein Fragment 1-34 Sigma Aldrich
Amyloid Precursor Protein Sigma Aldrich
Amyloid protein no AB component Sigma Aldrich
Secreted amyloid precursor protein (SAP) beta Sigma Aldrich
Tau isoform variant 0N3R Sigma Aldrich
Tau isoform variant 1N3R Sigma Aldrich
Tau isoform variant 0N4R Sigma Aldrich
Tau isoform variant 2N3R Sigma Aldrich
Tau phospho Ser412 Sigma Aldrich
Tau phospho Ser441 Sigma Aldrich
Tau phospho Thr181 Sigma Aldrich
Tau Protein human EMD Biosciences
(plusmn)9-HODE Cayman Chemical
Lipids
1 Palmitoyl-2-(5oxo-Valeroyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
15a-hydroxycholestene Avanti Polar Lipids
15
15-ketocholestane Avanti Polar Lipids
15-ketocholestene Avanti Polar Lipids
1-Palmitoil-2-(9oxo-Nonanoyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Azelaoyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Glutaroyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
5 α-cholestane-3 β15 α-diol Avanti Polar Lipids
9(S)-HODE Cayman Chemical
Asialoganglioside-GM1 Sigma Aldrich
Asialoganglioside-GM2 Sigma Aldrich
Brain ceramides Avanti Polar Lipids
Brain D-erythrosphingosine Avanti Polar Lipids
Brain lysophosphatidylethanolamine Avanti Polar Lipids
Brain L-α-lysophosphatidylserine Avanti Polar Lipids
Brain L-α-phosphatidylcholine Avanti Polar Lipids
Brain L-α-phosphatidyl-ethanolamine Avanti Polar Lipids
Brain L-α-phosphatidylserine Avanti Polar Lipids
Brain polar lipid extract Avanti Polar Lipids
Brain sphingomyelin Avanti Polar Lipids
Brain sulfatide Avanti Polar Lipids
Brain total lipid extract Avanti Polar Lipids
Cardiolipin Sigma Aldrich
Ceramide Sigma Aldrich
Ceramide 1-phosphate Sigma Aldrich
Cholesterol Sigma Aldrich
16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-

10
Myelin Protein 2 peptide aa 1-20 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 121-132 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 16-35 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 31-50 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 46-65 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 61-80 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 76-95 Biopolymers Facility Harvard Medical School
Myelin Protein 2 peptide aa 91-110 Biopolymers Facility Harvard Medical School
Neurofilament 160kd Chemicon
Neurofilament 200kd Chemicon
Neurofilament 68kd Chemicon
Neuronal Enolase Calbiochem
Nicastrin GeneTex
NMDA receptor Novus Biologicals
NOGO Sigma Aldrich
Olygodendrocyte-Specific Protein peptide aa 106ndash
125 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 1ndash20 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 121ndash
140 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 136ndash
155 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 151ndash
170 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 16ndash
35 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 166ndash Biopolymers Facility Harvard Medical School
11
185
Olygodendrocyte-Specific Protein peptide aa 181ndash
199 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 195ndash
217 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 31ndash
50 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 46ndash
65 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 61ndash
80 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 76ndash
95 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 91ndash
110 Biopolymers Facility Harvard Medical School
Proteolipid Protein Abnova
Proteolipid Protein peptide aa 100-119 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 10-29 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 110-129 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 1-19 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 125-141 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-150 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-154 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 150-163 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 151-173 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 158-166 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 161-180 Biopolymers Facility Harvard Medical School
12
Proteolipid Protein peptide aa 178-191 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 180-199 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 190-209 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 20-39 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 205-220 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 215-232 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-239 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-249 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 248-259 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 250-269 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 265-277 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 35-50 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 40-59 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 50-69 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 65-84 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 80-99 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 91-110 Biopolymers Facility Harvard Medical School
Retinol Binding Protein Sigma Aldrich
S100beta protein Assay Designs
Super Oxide Dismutase Sigma Aldrich
Synuclein beta Sigma Aldrich
Synuclein gamma Sigma Aldrich
Amydgala ProSci Inc
Tissue Amydgala AD ProSci Inc
Brain lysate ProSci Inc
Brain Tissue Membrane ProSci Inc
Cerebellar pedunculus ProSci Inc
13
Cerebral meninges ProSci Inc
Corpus Callosum ProSci Inc
Corpus Callosum AD ProSci Inc
Diencephalon ProSci Inc
Fetal brain ProSci Inc
Frontal lobe ProSci Inc
Frontal lobe AD ProSci Inc
Hippocampus ProSci Inc
Hippocampus AD ProSci Inc
Insula ProSci Inc
Occipital lobe ProSci Inc
Occipital lobe AD ProSci Inc
Olfactory region ProSci Inc
Optic Nerve ProSci Inc
Parietal lobe ProSci Inc
Parietal lobe AD ProSci Inc
Pons ProSci Inc
Pons AD ProSci Inc
Postcentral gyrus ProSci Inc
Postcentral gyrus AD ProSci Inc
Precentral gyrus ProSci Inc
Precentral gyrus AD ProSci Inc
Spinal cord ProSci Inc
Temporal lobe ProSci Inc
Temporal lobe AD ProSci Inc
Thalamus ProSci Inc
Thalamus AD ProSci Inc
14
Amyloid beta Sigma Aldrich
AD
related Amyloid beta 10-20 Sigma Aldrich
Amyloid beta 1-12 Sigma Aldrich
Amyloid beta 12-28 Sigma Aldrich
Amyloid beta 1-23 Sigma Aldrich
Amyloid beta 1-38 Sigma Aldrich
Amyloid beta 17-40 Sigma Aldrich
Amyloid beta 25-35 Sigma Aldrich
Amyloid beta 34-42 Sigma Aldrich
Amyloid bri protein precursor 227 Sigma Aldrich
Amyloid DAN Protein Fragment 1-34 Sigma Aldrich
Amyloid Precursor Protein Sigma Aldrich
Amyloid protein no AB component Sigma Aldrich
Secreted amyloid precursor protein (SAP) beta Sigma Aldrich
Tau isoform variant 0N3R Sigma Aldrich
Tau isoform variant 1N3R Sigma Aldrich
Tau isoform variant 0N4R Sigma Aldrich
Tau isoform variant 2N3R Sigma Aldrich
Tau phospho Ser412 Sigma Aldrich
Tau phospho Ser441 Sigma Aldrich
Tau phospho Thr181 Sigma Aldrich
Tau Protein human EMD Biosciences
(plusmn)9-HODE Cayman Chemical
Lipids
1 Palmitoyl-2-(5oxo-Valeroyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
15a-hydroxycholestene Avanti Polar Lipids
15
15-ketocholestane Avanti Polar Lipids
15-ketocholestene Avanti Polar Lipids
1-Palmitoil-2-(9oxo-Nonanoyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Azelaoyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Glutaroyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
5 α-cholestane-3 β15 α-diol Avanti Polar Lipids
9(S)-HODE Cayman Chemical
Asialoganglioside-GM1 Sigma Aldrich
Asialoganglioside-GM2 Sigma Aldrich
Brain ceramides Avanti Polar Lipids
Brain D-erythrosphingosine Avanti Polar Lipids
Brain lysophosphatidylethanolamine Avanti Polar Lipids
Brain L-α-lysophosphatidylserine Avanti Polar Lipids
Brain L-α-phosphatidylcholine Avanti Polar Lipids
Brain L-α-phosphatidyl-ethanolamine Avanti Polar Lipids
Brain L-α-phosphatidylserine Avanti Polar Lipids
Brain polar lipid extract Avanti Polar Lipids
Brain sphingomyelin Avanti Polar Lipids
Brain sulfatide Avanti Polar Lipids
Brain total lipid extract Avanti Polar Lipids
Cardiolipin Sigma Aldrich
Ceramide Sigma Aldrich
Ceramide 1-phosphate Sigma Aldrich
Cholesterol Sigma Aldrich
16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-
11
185
Olygodendrocyte-Specific Protein peptide aa 181ndash
199 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 195ndash
217 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 31ndash
50 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 46ndash
65 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 61ndash
80 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 76ndash
95 Biopolymers Facility Harvard Medical School
Olygodendrocyte-Specific Protein peptide aa 91ndash
110 Biopolymers Facility Harvard Medical School
Proteolipid Protein Abnova
Proteolipid Protein peptide aa 100-119 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 10-29 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 110-129 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 1-19 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 125-141 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-150 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 137-154 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 150-163 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 151-173 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 158-166 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 161-180 Biopolymers Facility Harvard Medical School
12
Proteolipid Protein peptide aa 178-191 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 180-199 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 190-209 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 20-39 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 205-220 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 215-232 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-239 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-249 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 248-259 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 250-269 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 265-277 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 35-50 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 40-59 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 50-69 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 65-84 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 80-99 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 91-110 Biopolymers Facility Harvard Medical School
Retinol Binding Protein Sigma Aldrich
S100beta protein Assay Designs
Super Oxide Dismutase Sigma Aldrich
Synuclein beta Sigma Aldrich
Synuclein gamma Sigma Aldrich
Amydgala ProSci Inc
Tissue Amydgala AD ProSci Inc
Brain lysate ProSci Inc
Brain Tissue Membrane ProSci Inc
Cerebellar pedunculus ProSci Inc
13
Cerebral meninges ProSci Inc
Corpus Callosum ProSci Inc
Corpus Callosum AD ProSci Inc
Diencephalon ProSci Inc
Fetal brain ProSci Inc
Frontal lobe ProSci Inc
Frontal lobe AD ProSci Inc
Hippocampus ProSci Inc
Hippocampus AD ProSci Inc
Insula ProSci Inc
Occipital lobe ProSci Inc
Occipital lobe AD ProSci Inc
Olfactory region ProSci Inc
Optic Nerve ProSci Inc
Parietal lobe ProSci Inc
Parietal lobe AD ProSci Inc
Pons ProSci Inc
Pons AD ProSci Inc
Postcentral gyrus ProSci Inc
Postcentral gyrus AD ProSci Inc
Precentral gyrus ProSci Inc
Precentral gyrus AD ProSci Inc
Spinal cord ProSci Inc
Temporal lobe ProSci Inc
Temporal lobe AD ProSci Inc
Thalamus ProSci Inc
Thalamus AD ProSci Inc
14
Amyloid beta Sigma Aldrich
AD
related Amyloid beta 10-20 Sigma Aldrich
Amyloid beta 1-12 Sigma Aldrich
Amyloid beta 12-28 Sigma Aldrich
Amyloid beta 1-23 Sigma Aldrich
Amyloid beta 1-38 Sigma Aldrich
Amyloid beta 17-40 Sigma Aldrich
Amyloid beta 25-35 Sigma Aldrich
Amyloid beta 34-42 Sigma Aldrich
Amyloid bri protein precursor 227 Sigma Aldrich
Amyloid DAN Protein Fragment 1-34 Sigma Aldrich
Amyloid Precursor Protein Sigma Aldrich
Amyloid protein no AB component Sigma Aldrich
Secreted amyloid precursor protein (SAP) beta Sigma Aldrich
Tau isoform variant 0N3R Sigma Aldrich
Tau isoform variant 1N3R Sigma Aldrich
Tau isoform variant 0N4R Sigma Aldrich
Tau isoform variant 2N3R Sigma Aldrich
Tau phospho Ser412 Sigma Aldrich
Tau phospho Ser441 Sigma Aldrich
Tau phospho Thr181 Sigma Aldrich
Tau Protein human EMD Biosciences
(plusmn)9-HODE Cayman Chemical
Lipids
1 Palmitoyl-2-(5oxo-Valeroyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
15a-hydroxycholestene Avanti Polar Lipids
15
15-ketocholestane Avanti Polar Lipids
15-ketocholestene Avanti Polar Lipids
1-Palmitoil-2-(9oxo-Nonanoyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Azelaoyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Glutaroyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
5 α-cholestane-3 β15 α-diol Avanti Polar Lipids
9(S)-HODE Cayman Chemical
Asialoganglioside-GM1 Sigma Aldrich
Asialoganglioside-GM2 Sigma Aldrich
Brain ceramides Avanti Polar Lipids
Brain D-erythrosphingosine Avanti Polar Lipids
Brain lysophosphatidylethanolamine Avanti Polar Lipids
Brain L-α-lysophosphatidylserine Avanti Polar Lipids
Brain L-α-phosphatidylcholine Avanti Polar Lipids
Brain L-α-phosphatidyl-ethanolamine Avanti Polar Lipids
Brain L-α-phosphatidylserine Avanti Polar Lipids
Brain polar lipid extract Avanti Polar Lipids
Brain sphingomyelin Avanti Polar Lipids
Brain sulfatide Avanti Polar Lipids
Brain total lipid extract Avanti Polar Lipids
Cardiolipin Sigma Aldrich
Ceramide Sigma Aldrich
Ceramide 1-phosphate Sigma Aldrich
Cholesterol Sigma Aldrich
16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-

12
Proteolipid Protein peptide aa 178-191 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 180-199 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 190-209 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 20-39 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 205-220 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 215-232 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-239 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 220-249 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 248-259 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 250-269 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 265-277 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 35-50 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 40-59 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 50-69 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 65-84 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 80-99 Biopolymers Facility Harvard Medical School
Proteolipid Protein peptide aa 91-110 Biopolymers Facility Harvard Medical School
Retinol Binding Protein Sigma Aldrich
S100beta protein Assay Designs
Super Oxide Dismutase Sigma Aldrich
Synuclein beta Sigma Aldrich
Synuclein gamma Sigma Aldrich
Amydgala ProSci Inc
Tissue Amydgala AD ProSci Inc
Brain lysate ProSci Inc
Brain Tissue Membrane ProSci Inc
Cerebellar pedunculus ProSci Inc
13
Cerebral meninges ProSci Inc
Corpus Callosum ProSci Inc
Corpus Callosum AD ProSci Inc
Diencephalon ProSci Inc
Fetal brain ProSci Inc
Frontal lobe ProSci Inc
Frontal lobe AD ProSci Inc
Hippocampus ProSci Inc
Hippocampus AD ProSci Inc
Insula ProSci Inc
Occipital lobe ProSci Inc
Occipital lobe AD ProSci Inc
Olfactory region ProSci Inc
Optic Nerve ProSci Inc
Parietal lobe ProSci Inc
Parietal lobe AD ProSci Inc
Pons ProSci Inc
Pons AD ProSci Inc
Postcentral gyrus ProSci Inc
Postcentral gyrus AD ProSci Inc
Precentral gyrus ProSci Inc
Precentral gyrus AD ProSci Inc
Spinal cord ProSci Inc
Temporal lobe ProSci Inc
Temporal lobe AD ProSci Inc
Thalamus ProSci Inc
Thalamus AD ProSci Inc
14
Amyloid beta Sigma Aldrich
AD
related Amyloid beta 10-20 Sigma Aldrich
Amyloid beta 1-12 Sigma Aldrich
Amyloid beta 12-28 Sigma Aldrich
Amyloid beta 1-23 Sigma Aldrich
Amyloid beta 1-38 Sigma Aldrich
Amyloid beta 17-40 Sigma Aldrich
Amyloid beta 25-35 Sigma Aldrich
Amyloid beta 34-42 Sigma Aldrich
Amyloid bri protein precursor 227 Sigma Aldrich
Amyloid DAN Protein Fragment 1-34 Sigma Aldrich
Amyloid Precursor Protein Sigma Aldrich
Amyloid protein no AB component Sigma Aldrich
Secreted amyloid precursor protein (SAP) beta Sigma Aldrich
Tau isoform variant 0N3R Sigma Aldrich
Tau isoform variant 1N3R Sigma Aldrich
Tau isoform variant 0N4R Sigma Aldrich
Tau isoform variant 2N3R Sigma Aldrich
Tau phospho Ser412 Sigma Aldrich
Tau phospho Ser441 Sigma Aldrich
Tau phospho Thr181 Sigma Aldrich
Tau Protein human EMD Biosciences
(plusmn)9-HODE Cayman Chemical
Lipids
1 Palmitoyl-2-(5oxo-Valeroyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
15a-hydroxycholestene Avanti Polar Lipids
15
15-ketocholestane Avanti Polar Lipids
15-ketocholestene Avanti Polar Lipids
1-Palmitoil-2-(9oxo-Nonanoyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Azelaoyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Glutaroyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
5 α-cholestane-3 β15 α-diol Avanti Polar Lipids
9(S)-HODE Cayman Chemical
Asialoganglioside-GM1 Sigma Aldrich
Asialoganglioside-GM2 Sigma Aldrich
Brain ceramides Avanti Polar Lipids
Brain D-erythrosphingosine Avanti Polar Lipids
Brain lysophosphatidylethanolamine Avanti Polar Lipids
Brain L-α-lysophosphatidylserine Avanti Polar Lipids
Brain L-α-phosphatidylcholine Avanti Polar Lipids
Brain L-α-phosphatidyl-ethanolamine Avanti Polar Lipids
Brain L-α-phosphatidylserine Avanti Polar Lipids
Brain polar lipid extract Avanti Polar Lipids
Brain sphingomyelin Avanti Polar Lipids
Brain sulfatide Avanti Polar Lipids
Brain total lipid extract Avanti Polar Lipids
Cardiolipin Sigma Aldrich
Ceramide Sigma Aldrich
Ceramide 1-phosphate Sigma Aldrich
Cholesterol Sigma Aldrich
16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-

13
Cerebral meninges ProSci Inc
Corpus Callosum ProSci Inc
Corpus Callosum AD ProSci Inc
Diencephalon ProSci Inc
Fetal brain ProSci Inc
Frontal lobe ProSci Inc
Frontal lobe AD ProSci Inc
Hippocampus ProSci Inc
Hippocampus AD ProSci Inc
Insula ProSci Inc
Occipital lobe ProSci Inc
Occipital lobe AD ProSci Inc
Olfactory region ProSci Inc
Optic Nerve ProSci Inc
Parietal lobe ProSci Inc
Parietal lobe AD ProSci Inc
Pons ProSci Inc
Pons AD ProSci Inc
Postcentral gyrus ProSci Inc
Postcentral gyrus AD ProSci Inc
Precentral gyrus ProSci Inc
Precentral gyrus AD ProSci Inc
Spinal cord ProSci Inc
Temporal lobe ProSci Inc
Temporal lobe AD ProSci Inc
Thalamus ProSci Inc
Thalamus AD ProSci Inc
14
Amyloid beta Sigma Aldrich
AD
related Amyloid beta 10-20 Sigma Aldrich
Amyloid beta 1-12 Sigma Aldrich
Amyloid beta 12-28 Sigma Aldrich
Amyloid beta 1-23 Sigma Aldrich
Amyloid beta 1-38 Sigma Aldrich
Amyloid beta 17-40 Sigma Aldrich
Amyloid beta 25-35 Sigma Aldrich
Amyloid beta 34-42 Sigma Aldrich
Amyloid bri protein precursor 227 Sigma Aldrich
Amyloid DAN Protein Fragment 1-34 Sigma Aldrich
Amyloid Precursor Protein Sigma Aldrich
Amyloid protein no AB component Sigma Aldrich
Secreted amyloid precursor protein (SAP) beta Sigma Aldrich
Tau isoform variant 0N3R Sigma Aldrich
Tau isoform variant 1N3R Sigma Aldrich
Tau isoform variant 0N4R Sigma Aldrich
Tau isoform variant 2N3R Sigma Aldrich
Tau phospho Ser412 Sigma Aldrich
Tau phospho Ser441 Sigma Aldrich
Tau phospho Thr181 Sigma Aldrich
Tau Protein human EMD Biosciences
(plusmn)9-HODE Cayman Chemical
Lipids
1 Palmitoyl-2-(5oxo-Valeroyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
15a-hydroxycholestene Avanti Polar Lipids
15
15-ketocholestane Avanti Polar Lipids
15-ketocholestene Avanti Polar Lipids
1-Palmitoil-2-(9oxo-Nonanoyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Azelaoyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Glutaroyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
5 α-cholestane-3 β15 α-diol Avanti Polar Lipids
9(S)-HODE Cayman Chemical
Asialoganglioside-GM1 Sigma Aldrich
Asialoganglioside-GM2 Sigma Aldrich
Brain ceramides Avanti Polar Lipids
Brain D-erythrosphingosine Avanti Polar Lipids
Brain lysophosphatidylethanolamine Avanti Polar Lipids
Brain L-α-lysophosphatidylserine Avanti Polar Lipids
Brain L-α-phosphatidylcholine Avanti Polar Lipids
Brain L-α-phosphatidyl-ethanolamine Avanti Polar Lipids
Brain L-α-phosphatidylserine Avanti Polar Lipids
Brain polar lipid extract Avanti Polar Lipids
Brain sphingomyelin Avanti Polar Lipids
Brain sulfatide Avanti Polar Lipids
Brain total lipid extract Avanti Polar Lipids
Cardiolipin Sigma Aldrich
Ceramide Sigma Aldrich
Ceramide 1-phosphate Sigma Aldrich
Cholesterol Sigma Aldrich
16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-

14
Amyloid beta Sigma Aldrich
AD
related Amyloid beta 10-20 Sigma Aldrich
Amyloid beta 1-12 Sigma Aldrich
Amyloid beta 12-28 Sigma Aldrich
Amyloid beta 1-23 Sigma Aldrich
Amyloid beta 1-38 Sigma Aldrich
Amyloid beta 17-40 Sigma Aldrich
Amyloid beta 25-35 Sigma Aldrich
Amyloid beta 34-42 Sigma Aldrich
Amyloid bri protein precursor 227 Sigma Aldrich
Amyloid DAN Protein Fragment 1-34 Sigma Aldrich
Amyloid Precursor Protein Sigma Aldrich
Amyloid protein no AB component Sigma Aldrich
Secreted amyloid precursor protein (SAP) beta Sigma Aldrich
Tau isoform variant 0N3R Sigma Aldrich
Tau isoform variant 1N3R Sigma Aldrich
Tau isoform variant 0N4R Sigma Aldrich
Tau isoform variant 2N3R Sigma Aldrich
Tau phospho Ser412 Sigma Aldrich
Tau phospho Ser441 Sigma Aldrich
Tau phospho Thr181 Sigma Aldrich
Tau Protein human EMD Biosciences
(plusmn)9-HODE Cayman Chemical
Lipids
1 Palmitoyl-2-(5oxo-Valeroyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
15a-hydroxycholestene Avanti Polar Lipids
15
15-ketocholestane Avanti Polar Lipids
15-ketocholestene Avanti Polar Lipids
1-Palmitoil-2-(9oxo-Nonanoyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Azelaoyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Glutaroyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
5 α-cholestane-3 β15 α-diol Avanti Polar Lipids
9(S)-HODE Cayman Chemical
Asialoganglioside-GM1 Sigma Aldrich
Asialoganglioside-GM2 Sigma Aldrich
Brain ceramides Avanti Polar Lipids
Brain D-erythrosphingosine Avanti Polar Lipids
Brain lysophosphatidylethanolamine Avanti Polar Lipids
Brain L-α-lysophosphatidylserine Avanti Polar Lipids
Brain L-α-phosphatidylcholine Avanti Polar Lipids
Brain L-α-phosphatidyl-ethanolamine Avanti Polar Lipids
Brain L-α-phosphatidylserine Avanti Polar Lipids
Brain polar lipid extract Avanti Polar Lipids
Brain sphingomyelin Avanti Polar Lipids
Brain sulfatide Avanti Polar Lipids
Brain total lipid extract Avanti Polar Lipids
Cardiolipin Sigma Aldrich
Ceramide Sigma Aldrich
Ceramide 1-phosphate Sigma Aldrich
Cholesterol Sigma Aldrich
16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-
15
15-ketocholestane Avanti Polar Lipids
15-ketocholestene Avanti Polar Lipids
1-Palmitoil-2-(9oxo-Nonanoyl)-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Azelaoyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
1-Palmitoil-2-Glutaroyl-sn-Glycero-3-
Phosphocholine Avanti Polar Lipids
5 α-cholestane-3 β15 α-diol Avanti Polar Lipids
9(S)-HODE Cayman Chemical
Asialoganglioside-GM1 Sigma Aldrich
Asialoganglioside-GM2 Sigma Aldrich
Brain ceramides Avanti Polar Lipids
Brain D-erythrosphingosine Avanti Polar Lipids
Brain lysophosphatidylethanolamine Avanti Polar Lipids
Brain L-α-lysophosphatidylserine Avanti Polar Lipids
Brain L-α-phosphatidylcholine Avanti Polar Lipids
Brain L-α-phosphatidyl-ethanolamine Avanti Polar Lipids
Brain L-α-phosphatidylserine Avanti Polar Lipids
Brain polar lipid extract Avanti Polar Lipids
Brain sphingomyelin Avanti Polar Lipids
Brain sulfatide Avanti Polar Lipids
Brain total lipid extract Avanti Polar Lipids
Cardiolipin Sigma Aldrich
Ceramide Sigma Aldrich
Ceramide 1-phosphate Sigma Aldrich
Cholesterol Sigma Aldrich
16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-

16
Disialogaglioside-GD1B Sigma Aldrich
Disialogaglioside-GD2 Sigma Aldrich
Disialoganglioside GD1a Sigma Aldrich
Disialoganglioside GD3 HyTest
Fucosyl-GM1 Calbiochem
Galactocerebrosides Sigma Aldrich
Ganglioside Mixture Sigma Aldrich
Ganglioside-GM4 Calbiochem
Gangliotetraosylceramide asialo-GM1 Avanti Polar Lipids
HDL Sigma Aldrich
Hexacosanoic acid (26) Sigma Aldrich
Hydroxy fatty acid ceramide Sigma Aldrich
Isoprostane F2 I Cayman Chemical
Lactocerebrosides Sigma Aldrich
Lactosylceramide Calbiochem
LDL Sigma Aldrich
Lipid A diphosphoryl from Salmonella enterica Sigma Aldrich
Lipopolysaccharides from Escherichia coli Sigma Aldrich
Lipopolysaccharides from Pseudomona aeruginosa Sigma Aldrich
Lipopolysaccharides from Salmonella enterica Sigma Aldrich
Lyso-GM1 Calbiochem
Monosialoganglioside GM1 Sigma Aldrich
Monosialoganglioside GM2 Sigma Aldrich
Monosialoganglioside GM3 Meridian
N-Hexanoyl-D-sphingosin Sigma Aldrich
Non-hydroxy fatty acid ceramide Sigma Aldrich
Phosphatidylinositol-4 phosphate Sigma Aldrich
17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-

17
Squalene Sigma Aldrich
Sulfatides Sigma Aldrich
Tetracosanoic acid (24) Sigma Aldrich
Tetrasialoganglioside-GQ1B Calbiochem
TNPAL Galactocerebroside Sigma Aldrich
Total brain gangliosides Avanti Polar Lipids
Total cerebroside Avanti Polar Lipids
Trisialoganglioside GT1a HyTest
Trisialoganglioside-GT1B Sigma Aldrich
DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-

DOI 101212NXI000000000000020020163 Neurol Neuroimmunol Neuroinflamm
Rohit Bakshi Ada Yeste Bonny Patel et al Serum lipid antibodies are associated with cerebral tissue damage in multiple sclerosis
This information is current as of January 27 2016
ServicesUpdated Information amp
httpnnneurologyorgcontent32e200fullhtmlincluding high resolution figures can be found at
Supplementary Material httpnnneurologyorgcontentsuppl2016012732e200DC1html
Supplementary material can be found at
References httpnnneurologyorgcontent32e200fullhtmlref-list-1
This article cites 52 articles 7 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionmultiple_sclerosisMultiple sclerosis
httpnnneurologyorgcgicollectionautoimmune_diseasesAutoimmune diseasesfollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
2016 American Academy of Neurology All rights reserved Online ISSN 2332-7812Published since April 2014 it is an open-access online-only continuous publication journal Copyright copy
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
- BakshiLipidsMRIN216
- Table_e-1
-
Related Documents
![Serum Antibodies to Benzo(a)pyrene Diol Epoxide-DNA ......[CANCER RESEARCH 58. 4122-4126. September 15. 1998] Serum Antibodies to Benzo(a)pyrene Diol Epoxide-DNA Adducts in the General](https://static.cupdf.com/doc/110x72/60785c3b392baf57e50e7143/serum-antibodies-to-benzoapyrene-diol-epoxide-dna-cancer-research-58.jpg)










