1227 1 Hematology Division, Department of Oncology, Transplants, and New Advances in Medicine, and 3 Laboratory of Bioactive Polymeric Materials for Biomedical and Environmental Applications (BIOlab) UdR INSTM, Department of Chemistry and Industrial Chemistry, University of Pisa, Italy. 2 CUCCS-RRMR, Regione Toscana, AOUP and University of Pisa, Italy. 4 First Department of Pathology, Kansai Medical University, Moriguchi, Osaka, Japan. * Authors contribute equally to this article. Selective Culture of Mesodermal Progenitor Cells Luisa Trombi, 1,2, * Simone Pacini, 1, * Marina Montali, 1 Rita Fazzi, 1 Federica Chiellini, 3 Susumu Ikehara, 4 and Mario Petrini 1,2 We have recently identified mesodermal progenitor cells (MPCs) isolated from adult human bone marrow. These cells show unusual phenotypes, having putative embryonic markers and aldehyde dehydrogenase (ALDH) ac- tivity. Interestingly, these resting cells, which have been selected by culturing them in the presence of adult human serum, can easily be induced to differentiate into mature mesenchymal stromal cells (MSCs) after substituting the adult human serum for fetal bovine serum (FBS) or human cord serum. MPC-derived MSCs are, in turn, able to differentiate toward osteoblasts, chondrocytes, and adipocytes. Furthermore, MPCs are able to differentiate into endothelial cells. MPCs have been proven to be strongly adherent to plastic culture bottles and to be trypsin-resistant. In the present article, we show a simple and inexpensive method to isolate highly selected mesodermal progenitors from bone marrow or cord blood. The optimization of standard culture con- ditions (using commercial human AB sera and appropriate concentrations for cell seeding in plastics) allows a pure population of MPCs to be obtained even after a short culture period. We believe that this simple, repeatable, and standardized method will facilitate studies on MPCs. Introduction M esodermal progenitor cells (MPCs) have recently been isolated when culturing human bone marrow- derived cells in the presence of autologous serum (hAS) [1]. These adherent cells have been identified among typi- cal mesenchymal stromal cells (MSCs) when cultured in a medium supplemented with hAS. MPCs differ from MSCs in terms of their morphology (large, round cells) and of their qui- escent status (Ki67-negative). MPCs are able to differentiate into mesenchymal cells when cultured in FBS or human cord serum (hCS), and have been proven to be capable of generat- ing CFU-F and of differentiating into mesodermal lineages (adipocytes, chondrocytes, and osteoblasts), as described for MSCs [2–4]. MPCs do not directly differentiate into the meso- dermal lineage (osteogenic, chondrogenic, adipogenic) even in the presence of appropriate stimuli. Moreover, these cells may differentiate into endothelial cells when cultured in an appropriate VEGF-containing medium similar to those used in the differentiation of embryonic cells [5]. MPCs are positive for CD105 (but negative for CD90), express high levels of aldehyde dehydrogenase (ALDH) ac- tivity, and are positive for several markers, such as SSEA4, Oct-4, and NANOG, which are also expressed in embryonic cells. Nevertheless, because of their strong plastic adherence and trypsin resistance these cells are isolated from mes- enchymal cells, and this property has been used to enrich MPCs from MSCs. In this article, by screening several commercial sera, we have been able to select an optimal serum for MPC cul- ture, thus reducing the culture variability related to the use of autologous serum. Using these modified culture condi- tions, it has been possible to reach a selective culture of MPCs. Furthermore, by this simple and repeatable method, we are able to identify and isolate the MPCs from cord blood. The increased ability to recover MPCs is essential to further characterize these cells and evaluate their in vivo potentiality. STEM CELLS AND DEVELOPMENT Volume 18, Number 8, 2009 © Mary Ann Liebert, Inc. DOI: 10.1089/scd.2009.0054 ORIGINAL RESEARCH REPORT

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1227
1 Hematology Division, Department of Oncology, Transplants, and New Advances in Medicine, and 3Laboratory of Bioactive
Polymeric Materials for Biomedical and Environmental Applications (BIOlab) UdR INSTM, Department of Chemistry and Industrial Chemistry, University of Pisa, Italy.
2 CUCCS-RRMR, Regione Toscana, AOUP and University of Pisa, Italy.
4 First Department of Pathology, Kansai Medical University, Moriguchi, Osaka, Japan.
* Authors contribute equally to this article.
Selective Culture of Mesodermal Progenitor Cells
Luisa Trombi,1,2,* Simone Pacini,1,* Marina Montali,1 Rita Fazzi,1 Federica Chiellini,3 Susumu Ikehara,4 and Mario Petrini1,2
We have recently identifi ed mesodermal progenitor cells (MPCs) isolated from adult human bone marrow. These cells show unusual phenotypes, having putative embryonic markers and aldehyde dehydrogenase (ALDH) ac-tivity. Interestingly, these resting cells, which have been selected by culturing them in the presence of adult human serum, can easily be induced to differentiate into mature mesenchymal stromal cells (MSCs) after substituting the adult human serum for fetal bovine serum (FBS) or human cord serum. MPC-derived MSCs are, in turn, able to differentiate toward osteoblasts, chondrocytes, and adipocytes. Furthermore, MPCs are able to differentiate into endothelial cells. MPCs have been proven to be strongly adherent to plastic culture bottles and to be trypsin-resistant. In the present article, we show a simple and inexpensive method to isolate highly selected mesodermal progenitors from bone marrow or cord blood. The optimization of standard culture con-ditions (using commercial human AB sera and appropriate concentrations for cell seeding in plastics) allows a pure population of MPCs to be obtained even after a short culture period. We believe that this simple, repeatable, and standardized method will facilitate studies on MPCs.
Introduction
Mesodermal progenitor cells (MPCs) have recently been isolated when culturing human bone marrow-
derived cells in the presence of autologous serum (hAS) [1].These adherent cells have been identifi ed among typi-
cal mesenchymal stromal cells (MSCs) when cultured in a medium supplemented with hAS. MPCs differ from MSCs in terms of their morphology (large, round cells) and of their qui-escent status (Ki67-negative). MPCs are able to differentiate into mesenchymal cells when cultured in FBS or human cord serum (hCS), and have been proven to be capable of generat-ing CFU-F and of differentiating into mesodermal lineages (adipocytes, chondrocytes, and osteoblasts), as described for MSCs [2–4]. MPCs do not directly differentiate into the meso-dermal lineage (osteogenic, chondrogenic, adipogenic) even in the presence of appropriate stimuli. Moreover, these cells may differentiate into endothelial cells when cultured in an appropriate VEGF-containing medium similar to those used in the differentiation of embryonic cells [5].
MPCs are positive for CD105 (but negative for CD90), express high levels of aldehyde dehydrogenase (ALDH) ac-tivity, and are positive for several markers, such as SSEA4, Oct-4, and NANOG, which are also expressed in embryonic cells. Nevertheless, because of their strong plastic adherence and trypsin resistance these cells are isolated from mes-enchymal cells, and this property has been used to enrich MPCs from MSCs.
In this article, by screening several commercial sera, we have been able to select an optimal serum for MPC cul-ture, thus reducing the culture variability related to the use of autologous serum. Using these modifi ed culture condi-tions, it has been possible to reach a selective culture of MPCs.
Furthermore, by this simple and repeatable method, we are able to identify and isolate the MPCs from cord blood.
The increased ability to recover MPCs is essential to further characterize these cells and evaluate their in vivo potentiality.
STEM CELLS AND DEVELOPMENTVolume 18, Number 8, 2009© Mary Ann Liebert, Inc.DOI: 10.1089/scd.2009.0054
ORIGINAL RESEARCH REPORT
13-SCD-2009_0054.indd 1227 10/8/2009 3:20:02 PM

TROMBI ET AL.1228
Select®. Cells were washed with D-PBS 59, supplemented with 0.1% NaN3 (Sigma) and 0.05% bovine serum albumin (BSA; Sigma), and incubated with direct fl uorochrome-con-jugated monoclonal antibodies for 30 min at 4°C, accord-ing to the manufacturer’s instructions. After incubation, the cells were washed in D-PBS/NaN3/BSA and 30,000 events were acquired by FACSCantoTM II cytometer (Becton Dickinson, San Jose, CA, USA) and analyzed by FACSDivaTM analysis software. Cells were incubated with CD105 PE-conjugated and CD90 PE/Cy5-conjugated antibodies (all from Becton Dickinson) to analyze the percentages of MPCs and MSCs under different culture conditions. ALDH activity was assayed on MPCs by ALDEFLUOR® kit (Stem Cell, Vancouver, Canada) according to the manufacturer’s instructions, and the cells were gated on the basis of CD105 and C90 expression.
Tri-color immunofl uorescence. PhABS cultures of BMMNCs were performed on Permanox® double-chamber slides (Nunc, Rochester, NY, USA) as described earlier. After 8–10 days, the slides were fi xed for 15 min in periodate–lysine–paraformaldehyde (PLP) and made permeable by Triton X-100 0.05% for 30 min. Immunofl uorescence was performed using anti-Oct-3/4 (Santa Cruz, CA, USA) or anti-NANOG (Becton Dickinson) as primary antibodies and revealed by a Goat anti-mouse SFX kit (Invitrogen), according to the man-ufacturer’s instructions, using Alexa Fluor® 488 anti-mouse IgG. Subsequently, the slides were stained by Phalloidin Alexa Fluor® 555-conjugated (Invitrogen) for 30 min to re-veal actin organization, and mounted in Prolong® Gold antifade reagent with 4′,6-diamidino-2-phenylindole (DAPI) (Invitrogen) to allow nucleus localization. Pictures were taken and combined by standard fl uorescence DMR Leica microscope (Leica, Wetzlar, Germany) equipped with Leica CW4000 image software (Leica).
Incorporation of 5-bromo-2′-deoxyuridine (BrdU) was evaluated by a Roche labeling and detection kit I (Roche, Penzberg, Germany) to confi rm that the MPCs were quiescent.
RT-PCR for Oct-3/4 and NANOG. Total RNA was extracted from detached MPCs or MSC using an RNeasy Mini Kit (Quiagen, Hilden, GER) and reverse transcripted using an iScriptTM cDNA synthesis kit (Biorad, Hercules, CA, USA). PCRs were performed by a Taq DNA Polymerase kit from Invitrogen using primer pairs from a Human Pluripotent Stem Cell Assessment kit (R&D Systems, Minneapolis, MN, USA), according to the manufacturer’s instructions.
Differentiation potential of MPCs cultured under selective conditions
Mesenchymal differentiation. MPCs from PhABS cultures were plated, in quadruplicate, in 24-well TC plates at 5,000 cells/well in DMEM-10% FBS to induce mesenchymal pre-differentiation. They were thus maintained for 8–10 days, after which media from three wells were replaced with ter-minal differentiation media: NH OsteoDiff, NH AdipoDiff media from Miltenyi Biotech or detached to perform pel-let culture in NH ChondroDiff media (Miltenyi; Bergisch Gadbach, Germany). The MPCs were then maintained in the new media for at least 3 weeks. Differentiation was revealed by Von Kossa, Oil Red staining of methanol fi xed cultures, or Alcian Blue staining of paraffi n-embedded pellet cultures.
Materials and Methods
Evaluation of sera
After written consent, bone marrow aspirates were obtained under local anesthesia from fi ve patients, who were undergoing cardiac surgery. Bone marrow mononu-clear cells (BMMNCs) were collected by density gradient centrifugation on Histopaque® 1077 (Sigma, St. Louis, MO, USA), washed twice in D-PBS (Invitrogen, Eugene, OR, USA), and plated in tissue culture-treated six-well plates (Becton Dickinson, Franklin Lakes, NJ, USA) containing low-glucose Dulbecco’s modifi ed Eagle medium (DMEM) supplemented with 2 mM l-glutamine, 100 μg/mL gentamicin, and 5% human Autologous Serum (hAS) or 10% Fetal Bovine Serum (FBS, Invitrogen) or 5%–10% of commercial pooled human AB-type sera (PhABS) at a cell density of 2 × 105/cm2. The medium was changed after 48 h to remove nonadherent cells, and then twice per week. After 10–15 days, plates were morphologically scored for MPCs under an inverted microscope. We evaluated four different batches of commer-cial sera from three different manufacturers; two batches of human sera (male only) from Lonza (Walkersville, MD, USA), one batch of human serum plus “Off the Clot” (GMP grade) from LiStarFish (Cernusco S/N, Milan, Italy), and one batch from Euroclone (Siziano, Pavia, Italy).
Evaluation of culture conditions for MPC isolation
Effect of cellular seeding density. BMMNCs from fi ve differ-ent donors were cultured in DMEM supplemented with 10% of pooled human AB sera (PhABS) from Lonza, at three dif-ferent cell densities: 2, 4 and 8 × 105/cm2 in six-well culture plates gas-treated for adherent cells. After 10–15 days, con-tamination with MSCs was removed by trypsin digestion, and the MPC density was then evaluated by AlamarBlueTM (Invitrogen) reduction assay performed on undetached cells according to the manufacturer’s instructions.
Effect of culture volume. Cells from the same donors were seeded at 8 × 105/cm2 in six-well culture plates fi lled with 1 mL (100 μL/cm2), 2 mL (200 μL/cm2), or 3 mL (300 μL/cm2) of culture media (DMEM-10% PhABS). The MPC density was evaluated by AlamarBlueTM reduction assay after detaching MSCs by Trypsin/EDTA (Invitrogen) digestion.
Effect of uncoated different plastics. To evaluate the adhe-sion properties of the MPCs, BMMNCs from the same fi ve donors were plated at 8 × 105/cm2 in DMEM-10% PhABS (Lonza) in gas-treated six-well plates for adherent cells (Becton Dickinson) or in hydrophobic untreated six-well plates for suspension cultures (Greiner Bio-one, Monroe, NC, USA) and microscopically scored for MPCs and MSCs. Subsequently, 12 bone marrow samples were cultured in DMEM-10% PhABS (Lonza) on hydrophobic plastics at 4 or 8 × 105 cells/cm2. To evaluate the frequency of MPCs, iso-lated by optimal culture conditions, the cells were exposed to TrypLE Select® (Invitrogen) for at least 30 min at 37°C and then gently scraped to detach them. MPCs were counted in a Bürker hemocytometer chamber and characterized as described below.
Characterization of isolated MPCs
Flow cytometry. Cells cultured in DMEM supplemented with 10% hAS, FBS, or PhABS were digested by TrypLE
13-SCD-2009_0054.indd 1228 10/8/2009 3:20:02 PM

SELECTION OF MESODERMAL PROGENITOR CELLS 1229
Results
Optimal culture conditions for MPCs
BMMNCs from fi ve donors (two females and three males) with a median age of 76 years (range 60–77) were plated in low-glucose DMEM supplemented with different sera: hAS, FBS, and four batches of commercial AB-type pooled sera (two from Lonza, one from LiStarFish, one from Euroclone) at different percentages. As previously described, when we cultured the cells in hAS, we obtained a mixed culture of typical MSCs and large round MPCs [1]. After cultur-ing the BMMNCs in the presence of FBS, we only detected MSCs. Conversely, when we used commercial AB sera, we obtained different amounts of MPCs. In detail, we detected few MPCs (1–10 MPCs/microscopical fi eld) after culturing the BMMNCs in the presence of 5 and 10% of AB serum from LiStarFish, while MPCs were absent from the cultures con-taining 5 and 10% of AB serum from Euroclone. Interestingly, we always obtained a large amount of MPCs (>10 MPCs/microscopical fi eld) in cultures with the AB serum from Lonza, with better recovery at 10%, regardless of the batch. Because we detected MPCs in all the samples cultured with AB serum from Lonza, it became possible to obtain a uni-form evaluation of the samples and, subsequently, to stan-dardize the culture conditions. To try to increase the MPC recovery, we seeded BMMNCs at three different cell den-sities (2, 4, 8 × 105/cm2) in DMEM-10% PhABS from Lonza. After removing the MSCs with trypsin, AlamarBlueTM assay showed a signifi cant enhancement of MPCs after overseed-ing mononuclear cells (P < 0.05) (Fig. 1A). The maximum recovery of MPCs was reached after seeding mononuclear cells at 8 × 105 cells/cm2 in all experiments except one, in which an even higher recovery was obtained after seeding BMMNCs at 4 × 105 cell/cm2. Moreover, we tested the same samples, seeded at 4 × 105 cells/cm2, in different volumes of culture medium (1, 2, 3 mL of DMEM-10% PhABS from Lonza). The MPC density turned out to be higher at 3 mL (P < 0.05) (Fig. 1B).
Furthermore, BMMNCs were cultured both in six-well TC plates for adherent cells and in hydrophobic untreated six-well plates for suspension cultures. We obtained a highly purifi ed MPC population when culturing the cells in untreated plastic, while MSCs were almost completely absent.
Thus, the optimal culture conditions for obtaining highly purifi ed MPCs included the use of (1) 10% PhABS from Lonza, (2) overseeding at 8 × 105 cells/cm2, and (3) hydro-phobic untreated plates. These parameters identify the “selective culture” of MPCs and were adopted in the sub-sequent experiments. By culturing samples from 12 donors, seven females and fi ve males (median age 70 years, range 39–82) at 8 × 105 cells/cm2 in T75 fl asks, we were able to har-vest 5.40 ± 0.48 × 105 MPCs.
Characterization and differentiation potential of MPCs cultured under selective conditions
Morphological and cytofl uorimetric characterization. Under phase-contrast microscope, MPCs isolated as above reported were morphologically identical to ones previously de-scribed in hAS cultures (Fig. 1C.E), large round cells with a rifrangent central core region, and characteristic peripheral
Endothelial differentiation. Selected MPCs were detached and re-plated at 10,000 cells/cm2 in EndoCult® liquid media (StemCell) on fi bronectin BioCoatTM (Becton Dickinson) six-well plates. Parallel cultures were performed on double-chamber slides previously coated with human fi bronectin (Becton Dickinson) at 5 μg/cm2. Cultures were maintained until 80%–90% of confl uence, and then pre-differentiated cells were detached by trypsin digestion to avoid contam-ination of undifferentiated MPCs (trypsin-resistant) and re-plated at 10,000 cells/cm2 in fi bronectin-coated double-chamber slides to perform terminal differentiation. They were then cultured in EGM-2 media (Lonza) supplemented by 50 ng/mL of VEGF (PeproTech, Rocky Hill, NJ, USA) for 5 days. At confl uence, slides from pre- and terminal differen-tiation were fi xed for 15 min in PLP and made permeable by Triton X-100 0.05% 30 min before being processed for indirect immunofl uorescence staining by a Goat anti-mouse SFX kit (Invitrogen), according to the manufacturer’s instructions, using Alexa Fluor® 488 anti-mouse IgG to reveal the expres-sion of CD31 (Becton Dickinson), CD146 (Becton Dickinson), CD133 (Miltenyi), and VEGFR2/KDR (Santa Cruz).
Effect of adding FBS to PhABS selective culture conditions on MPC isolation. We defi ned selective culture conditions to isolate highly purifi ed MPCs by seeding 8 × 105/cm2 in DMEM-10% PhABS (Lonza) in untreated plastics for suspen-sion cultures. Cells from marrow aspirates were cultured in untreated six-well plates under selective culture conditions adding 0, 2, 5, 10, or 15% of FBS (Invitrogen). Further wells were set up by substituting 10% FBS for PhABS. The cells were digested by trypsin/EDTA, and the detached MSCs were counted in a Bürker hemocytometer. The density of the undetached MPCs was evaluated by AlamarBlueTM reduc-tion assay. To normalize the results, the data were expressed as the ratio (R) of the percentage of reduced AlamarBlueTM (%ABred) detected in the sample (2% to 15% FBS or 10% FBS without PhABS) to ones detected in control cells (cultured in PhABS without any FBS). The MSC ratio was calculated by dividing the number of detached MSCs in the sample by the ones in control cultures.
We also performed the reverse experiment, adding 0, 2, 5, 10, 15% of PhABS to cells cultured in 10% FBS cultured on the plastics for adherent cells (optimal conditions to culture MSCs).
MPC isolation from umbilical cord blood. Umbilical cord blood (UCB) samples were collected from the umbilical vein of fi ve-term deliveries with informed maternal consent. A 16-gauge needle was inserted into the umbilical vein, and UCB was allowed to fl ow into Vacutainer® K2EDTA Tubes (Becton Dickinson). Mononuclear cells were isolated by den-sity centrifugation as reported earlier and cultured under MPC selective conditions. Control experiments were per-formed by substituting FBS for PhABS. After being mor-phologically evaluated using phase-contrast microscopy, the cells were detached by TrypLE Select® after 10 days of culture and characterized by fl ow cytometry for the same specifi cities described earlier.
Statistical analysis
The data are presented as mean ± standard error (SE). Statistical signifi cance (P < 0.05) between treatment groups was evaluated by a single factor analysis of variance (ANOVA) for multiple groups or unpaired t-test for two groups.
13-SCD-2009_0054.indd 1229 10/8/2009 3:20:02 PM

TROMBI ET AL.1230
stain [7] (data not shown). NANOG is totally absent in stan-dard cultured MSCs but highly expressed on MPCs.
MPC differentiation potential. By culturing the MPCs (derived from selective culture) for 8–10 days in plates for adherent cells with DMEM-10% FBS, we obtained a confl u-ence of new MSCs in the presence of few residual undif-ferentiated MPCs (pre-differentiation step) (Fig. 4A.1). Subsequently, culturing the cells in differentiating media from Miltenyi Biotech (NH AdipoDiff, NH OsteoDiff, and NH ChondroDiff), the terminal differentiation of MSCs into adipocytes (Fig. 4A.2), osteoblasts (Fig. 4A.3), and chondro-cytes (Fig. 4A.4) was confi rmed. MPCs, pre-differentiated on fi bronectin in EndoCult® medium for at least 2 weeks, turned out CD90- and VEGFR2/KDR-positive (Fig. 4B.1–2) but were still negative for CD31 and CD146. When we detached pre-differentiated cells and cultured them in EGM-2 with VEGF for 5 days, the cells maintained positivity for CD90 and VEGFR2/KDR, becoming positive for CD31, and thus sug-gesting terminal endothelial differentiation (Fig. 4B.3–4).
Effect of adding FBS to PhABS selective culture conditions on MPC isolation
The addition of increasing concentrations of FBS to selec-tive cell culture conditions induced a progressively increased number of MSCs (P < 0.05; Fig. 5A–B). Nevertheless, the addition of FBS at the selective culture did not signifi cantly
lamellipodia. Moreover, cytospins of detached MPCs showed intense basophilic cytoplasm and membrane extrusions (Fig. 2D). Flow cytometry confi rmed the MPC phenotype, re-vealing weak reactivity for CD105 and peculiar CD90 nega-tivity of high SSC events (Fig. 2C). Interestingly, as indicated by Figure 2, standard FBS cultures produced highly pure MSCs (CD90brightCD105bright), while selective cultures allowed the recovery of MPCs (CD90negCD105dim) with no MSC con-tamination. As expected, culturing cells in treated plastics with hAS media gave rise to mixed populations of MSCs and MPCs.
MPCs expressed intense ALDH activity (Fig. 2E) and did not incorporate BrdU (data not shown), demonstrating that they were in a state of quiescence. All these results con-fi rmed that MPCs, isolated under selective conditions, were identical to the cells previously described.
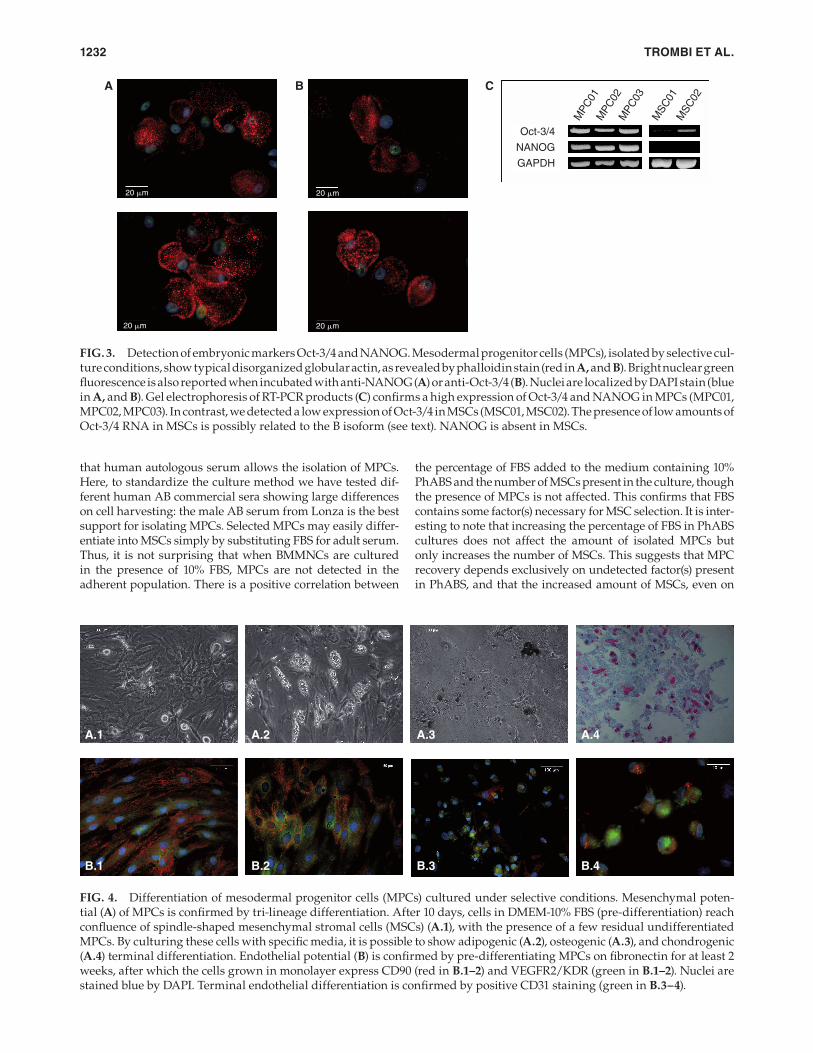
Detection of embryonic markers Oct-3/4 and NANOG. Immunofl uorescence studies, performed on MPCs cul-tured on Permanox® slides, showed a mainly disorganized actin disposition (Fig. 3A,B) and bright green nuclear stain for NANOG (Fig. 3A) or Oct-3/4 (Fig. 3B), confi rming that these cells strongly expressed genuine embryonic markers. RT-PCR performed both on MPC samples and MSC samples confi rmed this data (Fig. 3C). A high expression of Oct-3/4 and NANOG in MPCs was reported, while MSCs express low levels of Oct-3/4, probably due to isoform B detection [6], as confi rmed by weak cytoplasmatic Oct-3/4-positive
10
20
40
60
%A
B R
ed
80
100
120
2
*
*
*
*
mL of Culture Media
3
B
C
D
E
0.20
10
20
30
40
50%
AB
Red 60
70
80
90
100A
0.4
×106 Cells/cm2
0.8
*
*
*
FIG. 1. Effect of bone marrow mononuclear cell (BMMNC) density and amount of culture media on isolating mesodermal progenitor cells (MPCs). Overseeding signifi cantly enhances recovery of MPCs (A, *P < 0.05). Increased volume of culture medium modifi es the MPC recovery from 8 × 105/cm2 BMMNCs (B). MPCs from BMMNCs seeded at 2 × 105/cm2 (C), 4 × 105/cm2 (D), and 8 × 105/cm2 (E) show similar fried egg-shaped morphology.
13-SCD-2009_0054.indd 1230 10/8/2009 3:20:02 PM

SELECTION OF MESODERMAL PROGENITOR CELLS 1231
able to count >10 MPCs/microscopic fi eld with morphology identical to bone marrow-derived MPCs. Cytofl uorimetric analysis of the cells recovered (from PhABS cultures in un-treated plastics) revealed a phenotype CD90-negative and CD105 weakly positive, demonstrating that they were MPCs. In contrast, in FBS cultures on plates for adherent cells, we obtained only a few spindle-shaped cells.
Discussion
The seminal fi nding of this article is the identifi cation of optimal culture conditions for selectively isolating MPCs [1] from bone marrow and cord blood. This article demonstrates that making methodological modifi cations to the cell culture conditions allows us to avoid the strong heterogeneity peculiar to mesenchymal cell cultures [8,9]. We had previously shown
affect the MPC recovery, as documented by AlamarBlueTM reduction (Fig. 5B). Conversely, adding PhABS (2, 5, 10, 15%) to cultures supplemented with 10% FBS in plastic for adher-ent cells (optimal conditions for MSCs), MPCs were detect-able regardless of the percentage of PhABS. Under these conditions, PhABS did not affect MSC growth (Fig. 6). These fi ndings suggest that MPC isolation is related to PhABS sup-plementation, while plastic hydrophobicity contributes to purity, making ex vivo MSC adhesion diffi cult.
MPC isolation from UCB
Mononuclear cells were isolated from UCB and cultured at 8 × 105 cells/cm2, both in TC plastics and untreated ones, in medium supplemented with 10% of FBS or 10% of PhABS. After 8–10 days, in PhABS cultures (selective culture), we were
FSC-Height
0
A
B
C
D
E
01023
SS
C-H
eig
ht
1023
FSC-Height
0
01023
SS
C-H
eig
ht
1023
FSC-Height CD105 FITC
0 100
100
101
102
CD
90 P
C5
103
104
100
101
102
CD
90 P
C5
103
104
100
101
102
CD
90 P
C5 1
03
104
101 102 103
100
0256
101
ALDH Activity
102 103
104
CD105 FITC
100 101
R2
R1
R1
R2
R1
R2
102 103 104
CD105 FITC
100 101 102 103 104
01023
SS
C-H
eig
ht
1023
FIG. 2. Characterization of mesodermal progenitor cells (MPCs) cultured in selective conditions. Mesenchymal stromal cells (red dots, SSClowCD105brightCD90bright) are detected when culturing bone marrow mononuclear cells (BMMNCs) in 10% FBS (A) or 10% hAS (B) on TC culture plates. However, under the latter conditions, an MPC population is also detected (blue dots, SSChighCD105dimCD90neg). Culturing in DMEM-10% PhABS on hydrophobic untreated plates (C) represents a stringent selective condition that allows the isolation of almost pure MPCs from adult bone marrow. May Grünwald–Giemsa stain-ing of cytospins of PhABS-cultured cells (D) shows cells with intense basophilic cytoplasm and membrane extrusions. ALDEFLUOR® assay reveals ALDH activity on MPCs (E).
13-SCD-2009_0054.indd 1231 10/8/2009 3:20:03 PM
TROMBI ET AL.1232
the percentage of FBS added to the medium containing 10% PhABS and the number of MSCs present in the culture, though the presence of MPCs is not affected. This confi rms that FBS contains some factor(s) necessary for MSC selection. It is inter-esting to note that increasing the percentage of FBS in PhABS cultures does not affect the amount of isolated MPCs but only increases the number of MSCs. This suggests that MPC recovery depends exclusively on undetected factor(s) present in PhABS, and that the increased amount of MSCs, even on
that human autologous serum allows the isolation of MPCs. Here, to standardize the culture method we have tested dif-ferent human AB commercial sera showing large differences on cell harvesting: the male AB serum from Lonza is the best support for isolating MPCs. Selected MPCs may easily differ-entiate into MSCs simply by substituting FBS for adult serum. Thus, it is not surprising that when BMMNCs are cultured in the presence of 10% FBS, MPCs are not detected in the adherent population. There is a positive correlation between
A B
Oct-3/4
MPC
01M
PC
02M
PC
03
MSC
01M
SC
02
NANOG
GAPDH
C
20 μm20 μm
20 μm 20 μm
FIG. 3. Detection of embryonic markers Oct-3/4 and NANOG. Mesodermal progenitor cells (MPCs), isolated by selective cul-ture conditions, show typical disorganized globular actin, as revealed by phalloidin stain (red in A, and B). Bright nuclear green fl uorescence is also reported when incubated with anti-NANOG (A) or anti-Oct-3/4 (B). Nuclei are localized by DAPI stain (blue in A, and B). Gel electrophoresis of RT-PCR products (C) confi rms a high expression of Oct-3/4 and NANOG in MPCs (MPC01, MPC02, MPC03). In contrast, we detected a low expression of Oct-3/4 in MSCs (MSC01, MSC02). The presence of low amounts ofOct-3/4 RNA in MSCs is possibly related to the B isoform (see text). NANOG is absent in MSCs.
A.1 A.2 A.3 A.4
B.1 B.2 B.3 B.4
FIG. 4. Differentiation of mesodermal progenitor cells (MPCs) cultured under selective conditions. Mesenchymal poten-tial (A) of MPCs is confi rmed by tri-lineage differentiation. After 10 days, cells in DMEM-10% FBS (pre-differentiation) reach confl uence of spindle-shaped mesenchymal stromal cells (MSCs) (A.1), with the presence of a few residual undifferentiated MPCs. By culturing these cells with specifi c media, it is possible to show adipogenic (A.2), osteogenic (A.3), and chondrogenic (A.4) terminal differentiation. Endothelial potential (B) is confi rmed by pre-differentiating MPCs on fi bronectin for at least 2 weeks, after which the cells grown in monolayer express CD90 (red in B.1–2) and VEGFR2/KDR (green in B.1–2). Nuclei are stained blue by DAPI. Terminal endothelial differentiation is confi rmed by positive CD31 staining (green in B.3–4).
13-SCD-2009_0054.indd 1232 10/8/2009 3:20:04 PM

SELECTION OF MESODERMAL PROGENITOR CELLS 1233
a semiconservative proliferation of MPCs: further studies will help clarify this point.
MPCs are thus selected in the presence of human adult serum by their capacity to strongly adhere to (hydrophobic) plastic and to be resistant to trypsin digestion. The peculiar properties of MPCs allow their adhesion to the untreated, hydrophobic plastics usually used for culturing nonadher-ent cells. MSCs do not adhere to this plastic and, thus, are washed out when nonadherent cells are removed from the culture. The infl uence of the hydrophobicity of the culture surface on proliferation and differentiation has been previ-ously described for MSCs [10], osteoblasts and fi broblasts [11], and even on embryoid body formation [12].
Moreover, the trypsin resistance of MPCs suggests that the adhesion molecules involved are different from those of MSCs. This property allows us to easily select progen-itor cells even from UCB, thus confi rming the presence of MPCs in this tissue and the ability of our standardized method to isolate these cells not exclusively from adult bone marrow. It is relevant to underline that our purifi -cation method allows MPC purifi cation even from UCB, where it is usual for many different cell populations to contaminate the MSC preparations (ie, the adherent osteo-clast-like cells) [13].
The currently reported cells are identical to those previ-ously described [1]: MPCs cells are large, round, with intense basophilic cytoplasm and membrane extrusions. These cells are CD90neg, CD105dim, and ALDH+, as previ-ously reported [1]. The present results confi rm that MPCs do not incorporate BrdU and do express some putative embry-onic markers such as Oct-3/4 or NANOG. When cultured in the presence of FBS, the MPCs were able to differentiate into typical MSCs capable of undergoing several cell cycles. MPC-derived MSCs may be induced to further differentiate
the hydrophobic plastic, when FBS is present may possibly be related to increased MSC proliferation or to an enhanced adhesion of these cells to the plastics.
The addition of PhABS, even in small percentages, to FBS primary cultures allows MPC recovery, thus confi rming the presence of undetected agents able to select MPCs. Together, these results do not suggest the presence in FBS of factors able to interfere with MPC isolation. Obviously, the appear-ance of MSCs in the MPC cultures when FBS is added with-out any signifi cant reduction in the number of MPCs could be explained by assuming that this sera combination allows
A
A.1 +0% FBS (CTRL) +2% FBS
+5% FBS +10% FBS
+15% FBS 10% FBS only
A.3
A.2
A.4
A.5 A.6
B
0+2% FBS +5% FBS +10% FBS +15% FBS FBS only
MPCsMSCs
2
4*
*
*
*
6
R (
Ce
lls/C
ells
of
CT
RL
)
8
10
12
FIG. 5. Effects of FBS addition to mesodermal progenitor cell (MPC) cultures in PhABS on hydrophobic plates. (A) Increased percentage of FBS to MPC cultures clearly shows an increased mesenchymal stromal cell (MSC) number (from A.1 to A.5). Cells cultured in FBS without PhABS are almost exclusively MSCs. Obviously, the plastic type does not allow the recovery of high quantities of MPCs, even under these conditions (A.6). (B) Cell counting confi rms a signifi cant increase in the MSC fraction (black bars, *P < 0.05). The num-ber of residual MPCs, after trypsin digestion, is not signifi -cantly modifi ed, as evaluated by the percentage of reduced AlamarBlueTM. Dotted bars express the ratio between cells cultured in PhABS plus increasing amounts of added FBS versus control (cells cultured in 10% PhABS without any addition of FBS).
+2% PhABS0
2
4**
** **
**
**6
R (
Ce
lls/C
ells
of C
TR
L)
8
10
12MPCs
MSCs
+5% PhABS +10% PhABS +15% PhABS PhABS only
FIG. 6. Effects of PhABS addition to standard mesenchy-mal stromal cell (MSC) cultures. Adding PhABS to standard MSC cultures (10% of FBS on gas-treated plastic) allows the isolation of mesodermal progenitor cells (MPCs) (**P < 0.05), even at low percentages of human serum, without affecting MSC recovery. Black bars express the ratio between MSCs detected in cultures supplemented with increasing concen-trations of PhABS versus control (MSCs cultured with 10% FBS only). Dotted bars express the ratio of MPC detected cells, cultured as earlier.
13-SCD-2009_0054.indd 1233 10/8/2009 3:20:19 PM

TROMBI ET AL.1234
7. Mueller T, J Luetzkendorf, K Nerger, HJ Schmoll and LP Mueller.
(2009). Analysis of OCT4 expression in an extended panel of
human tumor cell lines from multiple entities and in human
mesenchymal stem cells. Cell Mol Life Sci 66:495–503.
8. Wagner W and AD Ho. (2007). Mesenchymal stem cell prepara-
tions, comparing apples and oranges. Stem Cell Rev 3:239–248.
9. Ho AD, W Wagner and W Franke. (2008). Heterogeneity of mesen-
chymal stromal cell preparations. Cytotherapy 10(4): 320–330.
10. Kim MS, YN Shin, MH Cho, SH Kim, SK Kim, YH Cho, G Khang,
IW Lee and HB Lee. (2007). Adhesion behavior of human bone
marrow stromal cells on differentially wettable polymer sur-
faces. Tissue Eng 13(8):2095–2103.
11. Schweikl H, R Müller, C Englert, KA Hiller, R Kujat, M Nerlich
and G Schmalz. (2007). Proliferation of osteoblasts and fi bro-
blasts on model surfaces of varying roughness and surface
chemistry. J Mater Sci Mater Med 18(10):1895–1905.
12. Luo TJ, ON Witte, X Liu, B Dunn and H Wu. (2008).
Hydrophobic surfaces for enhanced differentiation of embry-
onic stem cell-derived embryoid bodies. Proc Natl Acad Sci
USA 105(38):14459–14464.
13. Barachini S, L Trombi, S Danti, D D’Alessandro, B Battolla, A
Legitimo, C Nesti, I Mucci, M D’Acunto, MG Cascone, L Lazzeri,
L Mattii, R Consolini and M Petrini. (2009). Morpho-functional
characterization of human mesenchymal stem cells from umbil-
ical cord blood for potential applications in regenerative medi-
cine Stem Cells Dev 18(2):293–305.
Address correspondence to:Prof. Mario Petrini
Hematology DivisionDepartment of Oncology, Transplants,
and New Advances in MedicineUniversity of Pisa
Via Roma 55Pisa PI 56100
Italy
E-mail: [email protected]
Received for publication February 19, 2009Accepted after revision March 30, 2009
Prepublished on Liebert Instant Online March 30, 2009
into osteoblasts, chondrocytes, and adipocytes when cul-tured under appropriate conditions. Endothelial differentia-tion was also demonstrated from MPCs.
Further studies are ongoing in order to identify any putative factors responsible for both the maintenance of the precursor stage or differentiation toward spindle-shaped mesenchymal-like cells.
We believe that the inexpensive method to purify pro-genitors of mesodermal cells reported here will allow an expansion of studies of the biological properties of these cells and, possibly, their use in regenerative medicine.
Acknowledgments
The authors would like to thank Prof. A. Mussi, Dr. F. Verunelli, Dr. P. Di Vecchio, and surgery staff from the Cardiac and Thoracic Department of the University of Pisa for their support.
References
1. Petrini M, S Pacini, L Trombi, R Fazzi, M Montali, S Ikehara and
Abraham NG. (2008). Identifi cation and purifi cation of meso-
dermal progenitor cells (MPCs) from human adult bone mar-
row. Stem Cells Dev Nov 7. [Epub ahead of print]
2. Bruder SP, Jaiswal N and SE Haynesworth. (1997). Growth
kinetics, self-renewal, and the osteogenic potential of purifi ed
human mesenchymal stem cells during extensive subcultivation
and following cryopreservation. J Cell Biochem 64:278–294.
3. Caplan AI. (1991). Mesenchymal stem cells. J Orthop Res
9:641–650.
4. Pittenger MF, AM Mackay, SC Beck, RK Jaiswal, R Douglas, JD
Mosca, MA Moorman, DW Simonetti, S Craig and DR Marshak.
(1999). Multilineage potential of adult human mesenchymal
stem cells. Science 284:143–147.
5. Levenberg S, JS Golub, M Amit, J Itskovitz-Eldor and R Langer.
(2002). Endothelial cells derived from human embryonic stem
cells. Proc Natl Acad Sci USA 99:4391–4396.
6. Kotoula V, SI Papamichos and AF Lambropoulos. (2008).
Revisiting OCT4 expression in peripheral blood mononuclear
cells. Stem Cells 26:290–291.
13-SCD-2009_0054.indd 1234 10/8/2009 3:20:20 PM

This article has been cited by:
1. Karen Bieback, Andrea Hecker, Asli Kocaömer, Heinrich Lannert, Katharina Schallmoser, Dirk Strunk, Harald Klüter. 2009.Human Alternatives to Fetal Bovine Serum for the Expansion of Mesenchymal Stromal Cells from Bone Marrow. Stem Cells27:9, 2331-2341. [CrossRef]
Related Documents











