Original article Soluble factors released by endothelial progenitor cells promote migration of endothelial cells and cardiac resident progenitor cells Carmen Urbich a , Alexandra Aicher a , Christopher Heeschen a , Elisabeth Dernbach a , Wolf K. Hofmann b , Andreas M. Zeiher a , Stefanie Dimmeler a, * a Molecular Cardiology, Department of Internal Medicine III, University of Frankfurt, Theodor-Stern-Kai 7, 60590 Frankfurt, Germany b Department of Hematology and Oncology, Internal Medicine II, University of Frankfurt, Theodor-Stern-Kai 7, Frankfurt, Germany Received 12 October 2004; received in revised form 7 June 2005; accepted 11 July 2005 Available online 29 September 2005 Abstract Circulating endothelial progenitor cells (EPC) are incorporated into newly formed capillaries, enhance neovascularization after hind limb ischemia and improve cardiac function after ischemic injury. Incorporated progenitor cells may also promote neovascularization and cardiac regeneration by releasing factors, which act in a paracrine manner to support local angiogenesis and mobilize tissue residing progenitor cells. Therefore, we analyzed the expression profile of cytokines in human peripheral blood-derived EPC as opposed to human umbilical vein endothelial cells (HUVEC), human microvascular endothelial cells (HMVEC), and CD14 + monocytes by microarray technology. A gene tree analysis revealed a distinct expression pattern of angiogenic growth factors in EPC, mature endothelial cells, and CD14 + monocytes. VEGF-A, VEGF-B, SDF-1, and IGF-1 mRNA levels were higher in EPC as compared to HUVEC or HMVEC. The enhanced mRNA expression was paralleled by a significant release ofVEGF, SDF-1, and IGF-1 protein into the cell culture supernatant of EPC. Moreover, immunohistological analysis of ischemic limbs from nude rats revealed that VEGF is also released from recruited human EPC in vivo. As a functional conse- quence, conditioned medium of EPC induced a strong migratory response of mature endothelial cells, which was significantly inhibited by VEGF and SDF-1 neutralizing antibodies. Finally, conditioned medium of EPC significantly stimulated the migration of cardiac resident c-kit + progenitor cells in vitro. Taken together, EPC exhibit a high expression of angiogenic growth factors, which enhanced migration of mature endothelial cells and tissue resident cardiac progenitor cells. In addition to the physical contribution of EPC to newly formed vessels, the enhanced expression of cytokines may be a supportive mechanism to improve blood vessel formation and cardiac regeneration after cell therapy. © 2005 Elsevier Ltd. All rights reserved. Keywords: Growth factors; Endothelial progenitor cells; Angiogenesis 1. Introduction Postnatal neovascularization is an important process to res- cue tissue from critical ischemia [1]. Circulating bone marrow-derived endothelial progenitor cells (EPC) signifi- cantly contribute to adult blood vessel formation [2–4]. These endothelial progenitor cells are positive for endothelial marker proteins such as VEGF receptor 2 (KDR), von Willebrand factor (vWF), VE-cadherin, eNOS, take-up of diacetylated LDL, and bind lectin [5–7]. Injected EPC were shown to inte- grate into blood vessels and improve neovascularization of ischemic hind limbs and ischemic hearts in animal models [8–11]. In accordance, initial clinical trials indicate that bone marrow-derived or circulating blood-derived progenitor cells may be therapeutically useful to improve blood flow of ischemic tissue and improve heart function [12,13]. At present it is unclear, whether the improved heart function seen in patients after progenitor cell therapy is exclusively mediated by an improvement of neovascularization or also may reflect regeneration of cardiac tissue. Indeed, EPC have been shown to acquire a cardiac phenotype in vitro after co-cultivation * Corresponding author. Tel.: +49 69 6301 7440/5789; fax: +49 69 6301 7113/6374. E-mail address: [email protected] (S. Dimmeler). Journal of Molecular and Cellular Cardiology 39 (2005) 733–742 www.elsevier.com/locate/yjmcc 0022-2828/$ - see front matter © 2005 Elsevier Ltd. All rights reserved. doi:10.1016/j.yjmcc.2005.07.003

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Original article
Soluble factors released by endothelial progenitor cells promote migrationof endothelial cells and cardiac resident progenitor cells
Carmen Urbich a, Alexandra Aicher a, Christopher Heeschen a, Elisabeth Dernbach a,Wolf K. Hofmann b, Andreas M. Zeiher a, Stefanie Dimmeler a,*
a Molecular Cardiology, Department of Internal Medicine III, University of Frankfurt, Theodor-Stern-Kai 7, 60590 Frankfurt, Germanyb Department of Hematology and Oncology, Internal Medicine II, University of Frankfurt, Theodor-Stern-Kai 7, Frankfurt, Germany
Received 12 October 2004; received in revised form 7 June 2005; accepted 11 July 2005
Available online 29 September 2005
Abstract
Circulating endothelial progenitor cells (EPC) are incorporated into newly formed capillaries, enhance neovascularization after hind limbischemia and improve cardiac function after ischemic injury. Incorporated progenitor cells may also promote neovascularization and cardiacregeneration by releasing factors, which act in a paracrine manner to support local angiogenesis and mobilize tissue residing progenitor cells.Therefore, we analyzed the expression profile of cytokines in human peripheral blood-derived EPC as opposed to human umbilical veinendothelial cells (HUVEC), human microvascular endothelial cells (HMVEC), and CD14+ monocytes by microarray technology. A gene treeanalysis revealed a distinct expression pattern of angiogenic growth factors in EPC, mature endothelial cells, and CD14+ monocytes. VEGF-A,VEGF-B, SDF-1, and IGF-1 mRNA levels were higher in EPC as compared to HUVEC or HMVEC. The enhanced mRNA expression wasparalleled by a significant release of VEGF, SDF-1, and IGF-1 protein into the cell culture supernatant of EPC. Moreover, immunohistologicalanalysis of ischemic limbs from nude rats revealed that VEGF is also released from recruited human EPC in vivo. As a functional conse-quence, conditioned medium of EPC induced a strong migratory response of mature endothelial cells, which was significantly inhibited byVEGF and SDF-1 neutralizing antibodies. Finally, conditioned medium of EPC significantly stimulated the migration of cardiac residentc-kit+ progenitor cells in vitro. Taken together, EPC exhibit a high expression of angiogenic growth factors, which enhanced migration ofmature endothelial cells and tissue resident cardiac progenitor cells. In addition to the physical contribution of EPC to newly formed vessels,the enhanced expression of cytokines may be a supportive mechanism to improve blood vessel formation and cardiac regeneration after celltherapy.© 2005 Elsevier Ltd. All rights reserved.
Keywords: Growth factors; Endothelial progenitor cells; Angiogenesis
1. Introduction
Postnatal neovascularization is an important process to res-cue tissue from critical ischemia [1]. Circulating bonemarrow-derived endothelial progenitor cells (EPC) signifi-cantly contribute to adult blood vessel formation [2–4]. Theseendothelial progenitor cells are positive for endothelial markerproteins such as VEGF receptor 2 (KDR), von Willebrand
factor (vWF), VE-cadherin, eNOS, take-up of diacetylatedLDL, and bind lectin [5–7]. Injected EPC were shown to inte-grate into blood vessels and improve neovascularization ofischemic hind limbs and ischemic hearts in animal models[8–11]. In accordance, initial clinical trials indicate that bonemarrow-derived or circulating blood-derived progenitor cellsmay be therapeutically useful to improve blood flow ofischemic tissue and improve heart function [12,13].At presentit is unclear, whether the improved heart function seen inpatients after progenitor cell therapy is exclusively mediatedby an improvement of neovascularization or also may reflectregeneration of cardiac tissue. Indeed, EPC have been shownto acquire a cardiac phenotype in vitro after co-cultivation
* Corresponding author. Tel.: +49 69 6301 7440/5789; fax: +49 69 63017113/6374.
E-mail address: [email protected] (S. Dimmeler).
Journal of Molecular and Cellular Cardiology 39 (2005) 733–742
www.elsevier.com/locate/yjmcc
0022-2828/$ - see front matter © 2005 Elsevier Ltd. All rights reserved.doi:10.1016/j.yjmcc.2005.07.003

with neonatal rat cardiac myocytes [14]. However, cardiacregeneration may also be mediated by mobilization of endog-enous tissue residing cardiac progenitor cells [15–17].
EPC are incorporated to a significant extent into the cap-illaries after ischemia and, thereby, physically contribute tothe formation of new capillaries in the ischemic tissue. How-ever, the lower number of incorporated cells with an endot-helial phenotype in some of the experimental studies may notentirely explain the strong therapeutic effect observed aftercell therapy in experimental models [7,9,18]. Thus, the effi-ciency of EPC-induced neovascularization may not solely bedue to the incorporation of EPC into newly formed vessels,but may also be influenced by the release of pro-angiogenicfactors in a paracrine manner. In line with this hypothesis,the deletion of Tie-2-positive bone marrow-derived cellsthrough activation of a suicide gene blocked tumor angiogen-esis, even though in this study, cells were not found to beintegrated into the endothelial lining of the tumor vessels butrather located adjacent to the vessel wall [19]. Moreover,recent studies suggest that monocytic cells as well as mesen-chymal stem cells augment arteriogenesis via the release ofangiogenic growth factors [20–22]. The local release ofgrowth factors by progenitor cells may additionally protectcardiac myocytes from apoptosis and potentially mobilize car-diac progenitor cells [23–25]. For example, HGF and IGF-1 were shown to exert a potent anti-apoptotic activity andmobilize cardiac progenitor cells [24–27].
In the present study, we analyzed the expression of poten-tial angiogenic growth factors in EPC as opposed to humanumbilical vein endothelial cells, human microvascular endot-helial cells, and CD14+ monocytes. Our results indicate thatEPC express higher levels of growth factors such as the vas-cular endothelial growth factor (VEGF) A, stromal cell-derived factor-1 (SDF-1), insulin-like growth factor-1 (IGF-1), and hepatocyte growth factor (HGF) as compared tomature endothelial cells. All factors have been described aspro-angiogenic growth factors (for review see [28]).As a func-tional consequence, conditioned medium of EPC promotedmigration of mature endothelial cells and cardiac resident pro-genitor cells. The local EPC-mediated release of growth fac-tors may contribute to neovascularization and cardiac regen-eration after ischemia.
2. Material and methods
2.1. EPC culture assay
Mononuclear cells (MNCs) were isolated by density gra-dient centrifugation with Ficoll from peripheral blood ofhealthy human volunteers as previously described [6]. Imme-diately after isolation, 8 × 106 MNC/ml were plated on cul-ture dishes coated with human fibronectin (Sigma) and main-tained in endothelial basal medium (EBM; Cambrex,Verviers,Belgium) supplemented with hydrocortisone, bovine brain
extract, gentamicin, amphotericin B, epidermal growth fac-tor and 20% fetal calf serum (FCS). After 3 days in culture,nonadherent cells were removed by thorough washing withPBS and adherent cells were incubated in fresh medium for24 h before starting the experiments. EPC were characterizedby dual-staining for 1,1′-dioctadecyl-3,3,3′,3′-tetramethyl-indocarbocyanine-labeled acetylated low-density lipopro-tein (DiLDL) and lectin and by the expression of endothelialmarker proteins KDR, VE-cadherin, eNOS and von Will-ebrand factor [9,29].
2.2. Cell culture
Pooled human umbilical venous endothelial cells (HU-VEC) were purchased from Cambrex and cultured in endot-helial basal medium (Cambrex) supplemented with hydrocor-tisone, bovine brain extract, gentamicin, amphotericin B,epidermal growth factor and 10% fetal calf serum until thethird passage. After detachment with trypsin, cells (4.0 × 105
cells) were grown on 6 cm cell culture dishes for at least 18 h.Human dermal microvascular endothelial cells (HMVEC)were purchased from Cambrex and cultured according to theinstructions of the manufacturer in endothelial basalmedium-2 (EBM-2; Cambrex) supplemented with epidermalgrowth factor, hydrocortisone, fibroblast growth factor, vas-cular endothelial growth factor, insulin-like growth factor,ascorbic acid, gentamicin, amphotericin-B and 5% FCS untilthe fifth passage.
2.3. Isolation of CD14+ monocytes
Human mononuclear cells were obtained from peripheralblood of healthy human volunteers by density gradient cen-trifugation using Ficoll separating solution (density 1.077;Biochrom, Berlin, Germany). CD14+ monocytes were puri-fied from mononuclear cells by positive selection with anti-CD14-microbeads (Miltenyi Biotec, Bergisch-Gladbach, Ger-many) using a magnetic cell sorter device (Miltenyi Biotec).Purity assessed by FACS analysis was > 95%.
2.4. Oligonucleotide microarray analysis
Total RNA was isolated with the RNeasy Kit according toQiagen (Hilden, Germany). Ten micrograms of total RNAwas hybridized to the HG-U95Av2 microarray (Affymetrix,Santa Clara, CA, USA). The standard protocol used for samplepreparation and microarray processing is available fromAffymetrix. Expression data were analyzed using Microar-ray Suite version 5.0 (Affymetrix) and the software Gene-Spring version 4.2 (Silicon Genetics, San Carlos, CA, USA)as previously described [30].
2.5. Western blot analysis
Cells were incubated with lysis buffer (20 mmol/l Tris(pH 7.4), 150 mmol/l NaCl, 1 mmol/l EDTA, 1 mmol/l EGTA,
734 C. Urbich et al. / Journal of Molecular and Cellular Cardiology 39 (2005) 733–742

1% Triton, 2.5 mmol/l sodium pyrophosphate, 1 mmol/lb-glycerolphosphate, 1 mmol/l Na3VO4, 1 µg/ml leupeptinand 1 mmol/l phenylmethylsulfonyl fluoride) for 15 min onice. After centrifugation for 15 min at 20,000 × g (4 °C), theprotein content of the samples was determined according toBradford. Proteins (50 µg per lane) were loaded onto SDS-polyacrylamide gels and blotted onto PVDF membranes.Western blots were performed using antibodies directedagainst VEGF (R&D systems). Enhanced chemilumines-cence was performed according to the instructions of themanufacturer (Amersham, Germany). Blots were reprobedwith actin (1:2000; Sigma). The autoradiographies werescanned and semiquantitatively analyzed.
2.6. Hind limb ischemia model in athymic rats
For transplantation of human EPC into a xenogeneic ratmodel, immunodeficient athymic rnu:rnu rats (5–7 week oldfemales; Harlan, Borchen, Germany) were required. Thesuperficial femoral artery was ligated. CM-Dil (MolecularProbes; Göttingen, Germany)-labeled cells (2 × 106 cells perrat) were intravenously injected 24 h after induction of uni-lateral hind limb ischemia.
2.7. Immunohistochemistry
In nude rats, the gastrocnemius muscles were harvested48 h after injection of human CM-Dil-labeled EPC. Frozentissue sections (5 µm) were then cut and fixed with ice coldacetone. Incorporated CM-Dil-labeled EPC were detected bythe red fluorescence of the cell tracker. Myocytes were stainedwith laminin (Abcam) followed by goat-anti-rabbit AlexaFluor 647 (Molecular Probes). VEGF was detected using anantibody against VEGF (R&D systems, Wiesbaden, Ger-many), which reacts with human VEGF and, to a lesser extent,with rat VEGF, followed by goat-anti-mouse Alexa Fluor 488(Molecular Probes). Nuclei were stained with Sytox (Molecu-lar Probes). Tissue sections were examined using confocalmicroscopy (LSM 510, Zeiss, Germany).
2.8. Preparation of conditioned media and ELISA
EPC and HUVEC were incubated with equal amounts ofbasal EBM medium without supplements and FCS for 24 h.The medium was collected and 10 × concentrated by centrifu-gation for 20 min at 5000 × g at 4 °C using Ultrafree-4 centrifugal filter tubes with Biomax-5 membrane (Milli-pore). Release of VEGF, SDF-1, HGF, and IGF-1 wasmeasured in concentrated (10×) cell culture supernatants byELISA according to the instructions of the manufacturer(R&D systems).
2.9. Cell migration
Migration of HUVEC was detected using a “scratchedwound assay”. Therefore, in vitro “scratched” wounds were
created by scraping cell monolayers [31,32]. Cells were grownon 6 cm wells, which were previously labeled with a tracedline. After injury, the cells were gently washed with PBS andincubated with the concentrated conditioned media of EPCor HUVEC in the presence or absence of a neutralizing anti-body against VEGF (R&D systems) or SDF-1 (R&D sys-tems). EC migration from the edge of the injured monolayerwas quantified by measuring the distance between the woundedges at the time of injury and after 24 h of incubation usinga computer-assisted microscope (Zeiss, Jena, Germany) at fivedistinct positions (every 5 mm).
2.10. Isolation of cardiac progenitor cells
Cardiac progenitor cells were isolated from femaleC57BL/6 mice at 2 months of age. Mice hearts were mincedand incubated with a mixture of 0.1% dispase (Gibco,Karlsruhe, Germany) and 0.2% pronase E (Merck, Darms-tadt, Germany) in PBS for 20 min at 37 °C, then minced again,and mixed with pure fetal calf serum to stop the digestion.After centrifugation, cells were washed with PBS and counted.c-kit (CD117)+ cells were purified using positive selectionwith anti-CD117-microbeads (Miltenyi Biotec, Bergisch-Gladbach, Germany). Purity of sorted cells was determinedby FACS.
2.11. Statistical analysis
Data are expressed as mean ± S.E.M. from at least threeindependent experiments. Statistical analysis was performedby t-test. ANOVA was performed for serial analyses.
3. Results
3.1. Expression of pro-angiogenic factors in EPC
The endothelial phenotype of human peripheral blood-derived EPC was confirmed by expression of KDR, von Will-ebrand factor, CD146, VE-cadherin, and eNOS in previousstudies [9,29]. A gene tree analysis revealed a distinct geneexpression pattern of growth factors (99 different oligonucle-otides) in EPC as compared to other cell types. As shown inFig. 1, the expression of growth factors in EPC was differentas compared to HUVEC and HMVEC, and was also distin-guishable from those of CD14+ monocytes. The expressionpattern of HUVEC resembled that of HMVEC (Fig. 1). Indetail, VEGF A and VEGF B are highly expressed in EPC ascompared to HUVEC and HMVEC (Figs. 1 and 2A). In con-trast, VEGF C mRNA was exclusively expressed in HUVECand HMVEC (Figs. 1 and 2B). VEGF D was similarlyexpressed in EPC, HUVEC, and CD14+ monocytes and acidicFGF mRNA was similar in EPC, HMVEC, and CD14+ mono-cytes (Fig. 2B, C). Basic FGF revealed an enhanced expres-sion in HUVEC and HMVEC (Fig. 2C). The chemokinesSDF-1, IGF-1, HGF, and to a minor extent granulocyte colony
735C. Urbich et al. / Journal of Molecular and Cellular Cardiology 39 (2005) 733–742

stimulating factor (G-CSF) were increased in EPC as com-pared to HUVEC (Fig. 2D–G). In contrast, no differenceswere observed for the isoforms SDF-2 and IGF-2(Figs. 2D,2E).
3.2. EPC-mediated release of growth factors in vitroand in vivo
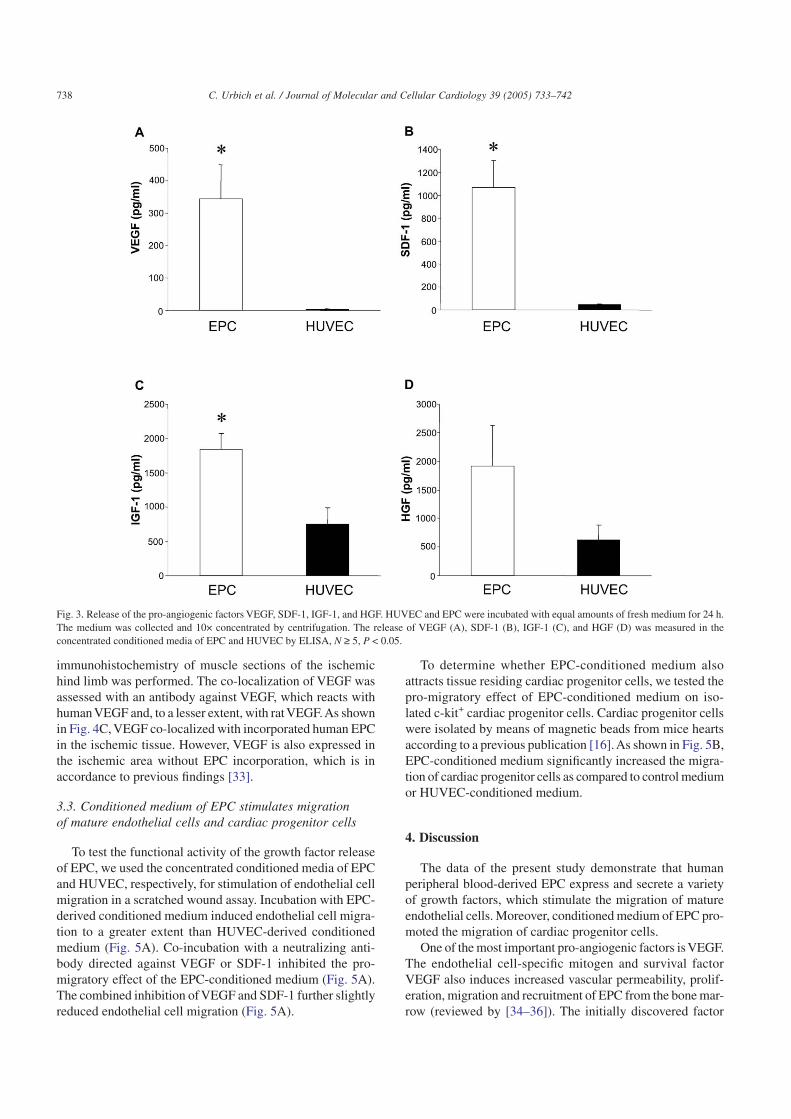
Growth factors are secreted to stimulate neighboring cells.Therefore, we measured the release of different factors intocell culture supernatants. As shown in Fig. 3, the release of
VEGF, SDF-1, IGF-1, and HGF was profoundly increased inEPC cell culture supernatants as compared to HUVEC.
Because VEGF is a predominant growth factor involvedin angiogenesis as well as vasculogenesis, we focused on theexpression and function of VEGF in EPC. Consistent withthe mRNA expression and the release into supernatants, theprotein expression ofVEGFA was also significantly increasedin EPC as compared to HUVEC and HMVEC (Figs. 4A, B).In contrast to the mRNA expression, the protein expressionof VEGF was lower in CD14+ monocytes as compared toEPC. However, the release of VEGF from cell culture super-
Fig. 1. Gene expression analysis of growth factors in EPC, HMVEC, HUVEC and CD14+ monocytes. Total RNA of EPC, HMVEC, HUVEC and CD14+
monocytes (each N = 3) were isolated and gene expression profiles were assessed with the Affymetrix gene chip expression assay. A gene tree analysis ofgrowth factors is shown. The blue color indicates low expression, the red color indicates high expression. The brightness indicates the reliability of hybridiza-tion signal. The expression of prominent clusters is marked with boxes and the corresponding genes are listed. Of note, some genes (e.g. VEGF A) arerepresented by two or more different oligonucleotides within the clusters, emphasizing the reproducibility of the assay.
736 C. Urbich et al. / Journal of Molecular and Cellular Cardiology 39 (2005) 733–742
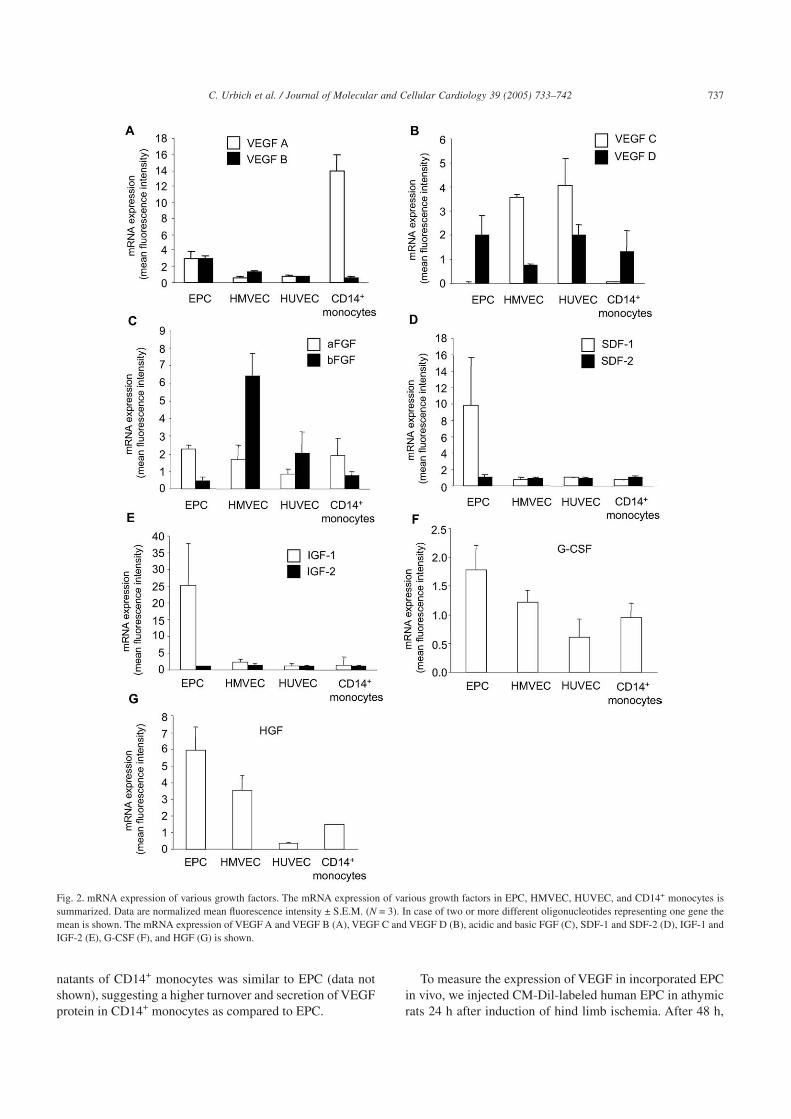
natants of CD14+ monocytes was similar to EPC (data notshown), suggesting a higher turnover and secretion of VEGFprotein in CD14+ monocytes as compared to EPC.
To measure the expression of VEGF in incorporated EPCin vivo, we injected CM-Dil-labeled human EPC in athymicrats 24 h after induction of hind limb ischemia. After 48 h,
Fig. 2. mRNA expression of various growth factors. The mRNA expression of various growth factors in EPC, HMVEC, HUVEC, and CD14+ monocytes issummarized. Data are normalized mean fluorescence intensity ± S.E.M. (N = 3). In case of two or more different oligonucleotides representing one gene themean is shown. The mRNA expression of VEGF A and VEGF B (A), VEGF C and VEGF D (B), acidic and basic FGF (C), SDF-1 and SDF-2 (D), IGF-1 andIGF-2 (E), G-CSF (F), and HGF (G) is shown.
737C. Urbich et al. / Journal of Molecular and Cellular Cardiology 39 (2005) 733–742
immunohistochemistry of muscle sections of the ischemichind limb was performed. The co-localization of VEGF wasassessed with an antibody against VEGF, which reacts withhumanVEGF and, to a lesser extent, with ratVEGF.As shownin Fig. 4C, VEGF co-localized with incorporated human EPCin the ischemic tissue. However, VEGF is also expressed inthe ischemic area without EPC incorporation, which is inaccordance to previous findings [33].
3.3. Conditioned medium of EPC stimulates migrationof mature endothelial cells and cardiac progenitor cells
To test the functional activity of the growth factor releaseof EPC, we used the concentrated conditioned media of EPCand HUVEC, respectively, for stimulation of endothelial cellmigration in a scratched wound assay. Incubation with EPC-derived conditioned medium induced endothelial cell migra-tion to a greater extent than HUVEC-derived conditionedmedium (Fig. 5A). Co-incubation with a neutralizing anti-body directed against VEGF or SDF-1 inhibited the pro-migratory effect of the EPC-conditioned medium (Fig. 5A).The combined inhibition of VEGF and SDF-1 further slightlyreduced endothelial cell migration (Fig. 5A).
To determine whether EPC-conditioned medium alsoattracts tissue residing cardiac progenitor cells, we tested thepro-migratory effect of EPC-conditioned medium on iso-lated c-kit+ cardiac progenitor cells. Cardiac progenitor cellswere isolated by means of magnetic beads from mice heartsaccording to a previous publication [16].As shown in Fig. 5B,EPC-conditioned medium significantly increased the migra-tion of cardiac progenitor cells as compared to control mediumor HUVEC-conditioned medium.
4. Discussion
The data of the present study demonstrate that humanperipheral blood-derived EPC express and secrete a varietyof growth factors, which stimulate the migration of matureendothelial cells. Moreover, conditioned medium of EPC pro-moted the migration of cardiac progenitor cells.
One of the most important pro-angiogenic factors is VEGF.The endothelial cell-specific mitogen and survival factorVEGF also induces increased vascular permeability, prolif-eration, migration and recruitment of EPC from the bone mar-row (reviewed by [34–36]). The initially discovered factor
Fig. 3. Release of the pro-angiogenic factors VEGF, SDF-1, IGF-1, and HGF. HUVEC and EPC were incubated with equal amounts of fresh medium for 24 h.The medium was collected and 10× concentrated by centrifugation. The release of VEGF (A), SDF-1 (B), IGF-1 (C), and HGF (D) was measured in theconcentrated conditioned media of EPC and HUVEC by ELISA, N ≥ 5, P < 0.05.
738 C. Urbich et al. / Journal of Molecular and Cellular Cardiology 39 (2005) 733–742

VEGF A consists of five splice variants with 121, 145, 165,189, 206 amino acids, whereby VEGF 165 is the most pre-dominant protein (reviewed by [37]). Meanwhile, severalother members of the VEGF family have been identifiedincluding VEGF B, C and D, which differ in the activation ofVEGF receptor subtypes [37]. The biological effects ofVEGFare mediated by two tyrosine kinase receptors, namely Flt-1(VEGFR-1) and KDR (VEGFR-2). Interestingly, membersof the VEGF gene family were differentially regulated in thedifferent cell types. Whereas VEGF D mRNA expression was
similar in EPC, HUVEC, and CD14+ monocytes, VEGF CmRNA was highest in mature endothelial cells. In contrast,the most predominant VEGF isoform, VEGF A, was signifi-cantly higher expressed in EPC as compared to mature endot-helial cells and was secreted and produced in vitro and invivo by EPC. However, monocytes showed a fourfold highermRNA expression level of VEGF A as compared to EPC.This is consistent with previous findings demonstrating thepreferential expression of VEGF A in monocytes [38,39].Interestingly, despite the high expression of VEGF A in
Fig. 4. Increased protein expression of VEGF in EPC as compared to mature endothelial cells. EPC, HMVEC, HUVEC, and CD14+ cells were lyzed and theexpression of VEGF A was analyzed by Western blot. Actin serves as loading control. A) A representative blot out of 3 independent experiments is shown. B)Blots were scanned and protein expression was quantified by densitometric analysis. The ratio for VEGF A/loading is shown. Data are mean ± S.E.M., N ≥ 3;*P < 0.05 versus HUVEC. C) Human CM-Dil-labeled EPC were intravenously injected into rats 24 h after induction of hind limb ischemia. Histologicalsections were obtained 48 h after injection of EPC. Human EPC were identified by CM-Dil (red fluorescence). VEGF was stained with an antibody againsthuman and rat VEGF (green fluorescence). Laminin staining is given in white. Nuclei were stained with Sytox (blue fluorescence). Representative confocalmicrographs are shown.
739C. Urbich et al. / Journal of Molecular and Cellular Cardiology 39 (2005) 733–742

CD14+ monocytic cells, the infusion of these cells did notaugment neovascularization after hind limb ischemia [9]. Thismight be explained by a lower protein expression as deter-mined by Western blot (Fig. 4). However, one may also specu-late that the release of pro-angiogenic factors may not be theonly prerequisite to improve neovascularization. This assump-tion is also supported by a recent publication of Hur andco-workers, who compared two different types of EPC, earlyspindle-shaped and late cobblestone-shaped EPC. Althoughlate EPC produced significantly lower levels of pro-angiogenic factors (6–10-fold) as compared to early EPC, thevasculogenic potential of early and late EPC in vivo was simi-lar [40]. Thus, although the release of pro-angiogenic factorsmay play a crucial role for augmenting neovascularization[20–22,41], distinct mechanisms may underlie the observedimprovement in blood flow after ischemia for different pro-genitor cell subsets. Supplementary to the improvement ofneovascularization, injection of EPC also significantlyimproved cardiac function in animal models [10,11]. Consis-tently, initial clinical trials indicate that bone marrow-derived or circulating blood-derived progenitor cells are thera-peutically useful to improve heart function [13,42]. However,the relative contributions of different potential mechanisms
(differentiation versus paracrine effects) are difficult to dis-sect in vivo, and, remain to be elucidated.
Other factors such as IGF-1, HGF and SDF-1 were alsohighly expressed in EPC as compared to mature endothelialcells or monocytes. IGF-1 and HGF are potent anti-apoptoticproteins and promote angiogenesis in different models[43,44]. In addition, IGF-1 plays a crucial role in muscleregeneration by promoting the proliferation and differentia-tion of satellite cells in the muscle, enabling them to fuse toexisting muscle fibers and repair damaged regions and sub-sequently to enhance muscle regenerative capacity [45,46].Recent studies provide further evidence that HGF contrib-utes to cardiac regeneration by affecting tissue resident pro-genitor cells. HGF stimulated the mobilization of cardiac pro-genitor cells from their niche and improved cardiacregeneration and heart function [26]. Interestingly, the super-natant of cultured EPC increased the migration of isolatedcardiac progenitor cells. SDF-1 represents an additional cytok-ine involved in the attraction of hematopoietic and endothe-lial progenitor cells. Together with VEGF, SDF-1 not onlystimulates the migration of mature endothelial cells but alsoacts as a chemoattractant to promote migration and tissue inva-sion of endothelial and hematopoietic progenitor cells [18,47].
Fig. 5. Conditioned medium of EPC enhances migration of endothelial cells and cardiac progenitor cells. A) HUVEC and EPC were incubated with equalamounts of fresh medium for 24 h. The medium was collected and 10× concentrated by centrifugation. The concentrated conditioned media of EPC andHUVEC was used to incubate endothelial cells to measure cell migration by the use of a scratched wound assay in the presence or absence of neutralizingantibodies against VEGF (1 µg/ml) and/or SDF-1 (100 µg/ml). IgG isotype antibody was used as control. Data are mean ± S.E.M., N = 4; *P < 0.05 versusconditioned medium of EPC. B) c-kit+ cardiac progenitor cells were isolated from minced mice hearts. Migration of c-kit+ cells was analyzed using conditionedmedia of HUVEC or EPC as stimulus in a modified Boyden chamber. Data are mean ± S.E.M. (% of control medium), N ≥ 3; *P < 0.05 vs. no cells, #P < 0.05 vs.EPC.
740 C. Urbich et al. / Journal of Molecular and Cellular Cardiology 39 (2005) 733–742

Consistently, overexpression of SDF-1 promoted neovascu-larization of ischemic tissues [18]. Thus, SDF-1 and HGFreleased from EPC that have already been recruited into theischemic tissue may further promote the mobilization andrecruitment of circulating and tissue residing progenitor cellsinto the ischemic tissue and, thereby, further accelerate therevascularization and regeneration process [36,47,48].
Interestingly, other members of the IGF and SDF family,namely IGF-2 and SDF-2, whose functions are not welldefined yet, are not differentially expressed in EPC as com-pared to other cell types studied. Moreover, the pro-angiogenicprotein bFGF was even higher in microvascular endothelialcells as compared to EPC. Obviously, the release of factorsfrom EPC involved in neovascularization is a dynamic pro-cess and it is very likely that the expression pattern of angio-genic factors in EPC may change during homing and in vivoincorporation in the ischemic tissue.
Taken together, the present study demonstrates that endot-helial progenitor cells express and release a variety of growthfactors, which in turn can support the survival and functionof tissue residing cells (e.g. mature endothelial cells or car-diac progenitors) in a paracrine manner, thereby, accelerat-ing the process of new blood vessel formation and regenera-tion of ischemic tissues.
Acknowledgments
We would like to thank Melanie Näher, Andrea Knau andMarion Muhly-Reinholz for their excellent technical help.This study was supported by the Deutsche Forschungsge-meinschaft (FOR501 Di600/6-1 and He 3044/2-1).
References
[1] Isner JM, Asahara T. Angiogenesis and vasculogenesis as therapeuticstrategies for postnatal neovascularization. J Clin Invest 1999;103:1231–6.
[2] Asahara T, Murohara T, Sullivan A, Silver M, van der Zee R, Li T,et al. Isolation of putative progenitor endothelial cells for angiogen-esis. Science 1997;275:964–7.
[3] Takahashi T, Kalka C, Masuda H, Chen D, Silver M, Kearney M, et al.Ischemia- and cytokine-induced mobilization of bone marrow-derived endothelial progenitor cells for neovascularization. Nat Med1999;5:434–8.
[4] Asahara T, Masuda H, Takahashi T, Kalka C, Pastore C, Silver M,et al. Bone marrow origin of endothelial progenitor cells responsiblefor postnatal vasculogenesis in physiological and pathologicalneovascularization. Circ Res 1999;85:221–8.
[5] Dimmeler S, Aicher A, Vasa M, Mildner-Rihm C, Adler K, Tie-mann M, et al. HMG-CoA reductase inhibitors (statins) increaseendothelial progenitor cells via the PI 3-kinase/Akt pathway. J ClinInvest 2001;108:391–7.
[6] Vasa M, Fichtlscherer S, Adler K, Aicher A, Martin H, Zeiher AM,et al. Increase in circulating endothelial progenitor cells by statintherapy in patients with stable coronary artery disease. Circulation2001;103:2885–90.
[7] Kalka C, Masuda H, Takahashi T, Kalka-Moll WM, Silver M, Kear-ney M, et al. Transplantation of ex vivo expanded endothelial progeni-tor cells for therapeutic neovascularization. Proc Natl Acad Sci USA2000;97:3422–7.
[8] Murohara T, Ikeda H, Duan J, Shintani S, Sasaki K, Eguchi H, et al.Transplanted cord blood-derived endothelial precursor cells augmentpostnatal neovascularization. J Clin Invest 2000;105:1527–36.
[9] Urbich C, Heeschen C, Aicher A, Dernbach E, Zeiher AM, Dim-meler S. Relevance of monocytic features for neovascularizationcapacity of circulating endothelial progenitor cells. Circulation 2003;108:2511–6.
[10] Kawamoto A, Gwon HC, Iwaguro H, Yamaguchi JI, Uchida S,Masuda H, et al. Therapeutic potential of ex vivo expanded endothe-lial progenitor cells for myocardial ischemia. Circulation 2001;103:634–7.
[11] Kocher AA, Schuster MD, Szabolcs MJ, Takuma S, Burkhoff D,Wang J, et al. Neovascularization of ischemic myocardium by humanbone-marrow-derived angioblasts prevents cardiomyocyte apoptosis,reduces remodeling and improves cardiac function. Nat Med 2001;7:430–6.
[12] Tateishi-Yuyama E, Matsubara H, Murohara T, Ikeda U, Shintani S,Masaki H, et al. Therapeutic angiogenesis for patients with limbischaemia by autologous transplantation of bone-marrow cells: a pilotstudy and a randomised controlled trial. Lancet 2002;360:427–35.
[13] Assmus B, Schachinger V, Teupe C, Britten M, Lehmann R, Dobert N,et al. Transplantation of progenitor cells and regeneration enhance-ment in acute myocardial infarction (TOPCARE-AMI). Circulation2002;106:3009–17.
[14] Badorff C, Brandes RP, Popp R, Rupp S, Urbich C, Aicher A, et al.Transdifferentiation of blood-derived human adult endothelial pro-genitor cells into functionally active cardiomyocytes. Circulation2003;107:1024–32.
[15] Hierlihy AM, Seale P, Lobe CG, Rudnicki MA, Megeney LA. Thepost-natal heart contains a myocardial stem cell population. FEBSLett 2002;530:239–43.
[16] Beltrami AP, Barlucchi L, Torella D, Baker M, Limana F, Chimenti S,et al. Adult cardiac stem cells are multipotent and support myocardialregeneration. Cell 2003;114:763–76.
[17] Oh H, Bradfute SB, Gallardo TD, Nakamura T, Gaussin V, MishinaY,Pocius J, Michael LH, Behringer RR, Garry DJ, Entman ML,Schneider MD. Cardiac progenitor cells from adult myocardium:homing, differentiation, and fusion after infarction. Proc Natl AcadSci USA 2003;100:12313–8.
[18] Yamaguchi J, Kusano KF, Masuo O, Kawamoto A, Silver M,Murasawa S, et al. Stromal cell-derived factor-1 effects on ex vivoexpanded endothelial progenitor cell recruitment for ischemic neovas-cularization. Circulation 2003;107:1322–8.
[19] De Palma M, Venneri MA, Roca C, Naldini L. Targeting exogenousgenes to tumor angiogenesis by transplantation of genetically modi-fied hematopoietic stem cells. Nat Med 2003;9:789–95.
[20] Kinnaird T, Stabile E, Burnett MS, Shou M, Lee CW, Barr S, Fuchs S,Epstein SE. Local delivery of marrow-derived stromal cells augmentscollateral perfusion through paracrine mechanisms. Circulation 2004;109:1543–9.
[21] Kinnaird T, Stabile E, Burnett MS, Lee CW, Barr S, Fuchs S,Epstein SE. Marrow-derived stromal cells express genes encoding abroad spectrum of arteriogenic cytokines and promote in vitro and invivo arteriogenesis through paracrine mechanisms. Circ Res 2004;94:678–85.
[22] Rehman J, Li J, Orschell CM, March KL. Peripheral blood “endothe-lial progenitor cells” are derived from monocyte/macrophages andsecrete angiogenic growth factors. Circulation 2003;107:1164–9.
[23] Anversa P, Sussman MA, Bolli R. Molecular genetic advances incardiovascular medicine: focus on the myocyte. Circulation 2004;109:2832–8.
741C. Urbich et al. / Journal of Molecular and Cellular Cardiology 39 (2005) 733–742

[24] Li Q, Li B, Wang X, Leri A, Jana KP, Liu Y, et al. Overexpression ofinsulin-like growth factor-1 in mice protects from myocyte death afterinfarction, attenuating ventricular dilation, wall stress, and cardiachypertrophy. J Clin Invest 1997;100:1991–9.
[25] Nakamura T, Mizuno S, Matsumoto K, Sawa Y, Matsuda H. Myocar-dial protection from ischemia/reperfusion injury by endogenous andexogenous HGF. J Clin Invest 2000;106:1511–9.
[26] Linke A, Muller P, Nurzynska D, Casarsa C, Torella D, Nascim-bene A, et al. Stem cells in the dog heart are self-renewing, clono-genic, and multipotent and regenerate infracted myocardium, improv-ing cardiac function. Proc Natl Acad Sci USA 2005;102(25):8944–8971.
[27] Torella D, Rota M, Nurzynska D, Musso E, Monsen A, Shiraishi I,Zias E, Walsh K, Rosenzweig A, Sussman MA, Urbanek K, Nadal-Ginard B, Kajstura J, Anversa P, Leri A. Cardiac stem cell andmyocyte aging, heart failure, and insulin-like growth factor-1 overex-pression. Circ Res 2004;94:514–24.
[28] Carmeliet P, Jain RK. Angiogenesis in cancer and other diseases.Nature 2000;407:249–57.
[29] Vasa M, Fichtlscherer S, Aicher A, Adler K, Urbich C, Martin H, et al.Number and migratory activity of circulating endothelial progenitorcells inversely correlate with risk factors for coronary artery disease.Circ Res 2001;89:E1–E7.
[30] Hofmann WK, de Vos S, Tsukasaki K, Wachsman W, Pinkus GS,Said JW, et al. Altered apoptosis pathways in mantle cell lymphomadetected by oligonucleotide microarray. Blood 2001;98:787–94.
[31] Urbich C, Reissner A, Chavakis E, Dernbach E, Haendeler J, Flem-ing I, et al. Dephosphorylation of endothelial nitric oxide synthasecontributes to the anti-angiogenic effects of endostatin. FASEB J2002;16:706–8.
[32] Tamura M, Gu J, Matsumoto K, Aota S, Parson R, Yamada KM.Inhibition of cell migration, spreading and focal adhesions by tumorsuppressor PTEN. Science 1998;280:1614–7.
[33] Lee SH, Wolf PL, Escudero R, Deutsch R, Jamieson SW, Thistleth-waite PA. Early expression of angiogenesis factors in acute myocar-dial ischemia and infarction. N Engl J Med 2000;342:626–33.
[34] Ferrara N, Alitalo K. Clinical applications of angiogenic growthfactors and their inhibitors. Nat Med 1999;5:1359–64.
[35] Ferrara N, Gerber HP, LeCouter J. The biology of VEGF and itsreceptors. Nat Med 2003;9:669–76.
[36] Asahara T, Takahashi T, Masuda H, Kalka C, Chen D, Iwaguro H,et al. VEGF contributes to postnatal neovascularization by mobilizingbone marrow-derived endothelial progenitor cells. EMBO J 1999;18:3964–72.
[37] Veikkola T, Alitalo K. VEGFs, receptors and angiogenesis. SeminCancer Biol 1999;9:211–20.
[38] Berse B, Brown LF, Van de Water L, Dvorak HF, Senger DR. Vascularpermeability factor (vascular endothelial growth factor) gene isexpressed differentially in normal tissues, macrophages, and tumors.Mol Biol Cell 1992;3:211–20.
[39] Polverini PJ, Cotran PS, Gimbrone Jr. MA, Unanue ER. Activatedmacrophages induce vascular proliferation. Nature 1977;269:804–6.
[40] Hur J, Yoon CH, Kim HS, Choi JH, Kang HJ, Hwang KK, Oh BH,Lee MM, Park YB. Characterization of two types of endothelialprogenitor cells and their different contributions to neovasculogen-esis. Arterioscler Thromb Vasc Biol 2004;24:288–93.
[41] Kamihata H, Matsubara H, Nishiue T, Fujiyama S, Tsutsumi Y,Ozono R, et al. Implantation of bone marrow mononuclear cells intoischemic myocardium enhances collateral perfusion and regionalfunction via side supply of angioblasts, angiogenic ligands, and cytok-ines. Circulation 2001;104:1046–52.
[42] Wollert KC, Meyer GP, Lotz J, Ringes-Lichtenberg S, Lippolt P,Breidenbach C, et al. Intracoronary autologous bone-marrow celltransfer after myocardial infarction: the BOOST randomised con-trolled clinical trial. Lancet 2004;364:141–8.
[43] Delafontaine P, SongYH, LiY. Expression, regulation, and function ofIGF-1, IGF-1R, and IGF-1 binding proteins in blood vessels. Arterio-scler Thromb Vasc Biol 2004;24:435–44.
[44] Morishita R, Aoki M, Hashiya N, Yamasaki K, Kurinami H,Shimizu S, et al. Therapeutic angiogenesis using hepatocyte growthfactor (HGF). Curr Gene Ther 2004;4:199–206.
[45] Barton-Davis ER, Shoturma DI, Musaro A, Rosenthal N,Sweeney HL. Viral mediated expression of insulin-like growth factorI blocks the aging-related loss of skeletal muscle function. Proc NatlAcad Sci USA 1998;95:15603–7.
[46] Musaro A, McCullagh K, Paul A, Houghton L, Dobrowolny G,Molinaro M, et al. Localized Igf-1 transgene expression sustainshypertrophy and regeneration in senescent skeletal muscle. Nat Genet2001;27:195–200.
[47] Kalka C, Masuda H, Takahashi T, Gordon R, Tepper O, Gravereaux E,et al. Vascular endothelial growth factor(165) gene transfer augmentscirculating endothelial progenitor cells in human subjects. Circ Res2000;86:1198–202.
[48] Takano H, Ohtsuka M, Akazawa H, Toko H, Harada M, Hasegawa H,et al. Pleiotropic effects of cytokines on acute myocardial infarction:G-CSF as a novel therapy for acute myocardial infarction. Curr PharmDes 2003;9:1121–7.
742 C. Urbich et al. / Journal of Molecular and Cellular Cardiology 39 (2005) 733–742
Related Documents





![Effect of vitamin D on endothelial progenitor cells function · vitamin D on EPCs function. Aim ... immune cells and endothelial cells [16]). Additional studies suggest a favorable](https://static.cupdf.com/doc/110x72/60c10a1fa60e3e04a118fdb0/effect-of-vitamin-d-on-endothelial-progenitor-cells-function-vitamin-d-on-epcs-function.jpg)





