Searching for virulence factors in the non-pathogenic parasite to humans Leishmania tarentolae H. AZIZI 1 #, K. HASSANI 1,2 #, Y. TASLIMI 1 , H. SHATERI NAJAFABADI 3 , B. PAPADOPOULOU 4 and S. RAFATI 1 * 1 Molecular Immunology and Vaccine Research Laboratory, Pasteur Institute of Iran, Tehran, Iran 2 Department of Biotechnology, University College of Science, University of Tehran, Tehran, Iran 3 Institute of Parasitology, McGill University, Montreal, Canada 4 Research Centre in Infectious Diseases, CHUL Research Centre and Department of Medical Biology, Faculty of Medicine, Laval University, 2705 Laurier Blvd., Quebec (QC), Canada G1V 4G2 (Received 15 September 2008; revised 7 November 2008 and 8 January 2009; accepted 8 February 2009; first published online 6 May 2009) SUMMARY Leishmania protozoa are obligate intracellular parasites that reside in the phagolysosome of host macrophages and cause a large spectrum of pathologies to humans known as leishmaniases. The outcome of the disease is highly dependent on the parasite species and on its ascribed virulence factors and the immune status of the host. Characterization of the genome composition of non-pathogenic species could ultimately open new horizons in Leishmania developmental biology and also the disease monitoring. Here, we provide evidence that the lizard non-pathogenic to humans Leishmania tarentolae species expresses an Amastin-like gene, cysteine protease B (CPB), lipophosphoglycan LPG3 and the leishmanolysin GP63, genes well-known for their potential role in the parasite virulence. These genes were expressed at levels comparable to those in L. major and L. infantum both at the level of mRNA and protein. Alignment of the L. tarentolae proteins with their counterparts in the pathogenic species demonstrated that the degree of similarity varied from 59 % and 60 % for Amastin, 89 % for LPG3 and 71 % and 68 % for CPB, in L. major and L. infantum, respectively. Interestingly, the A2 gene, expressed specifically by the L. donovani complex which promotes visceralization, was absent in L. tarentolae. These findings suggest that the lack of pathogenicity in L. tarentolae is not associated with known virulence genes such as LPG3, CPB, GP63 and Amastin, and that other factors either unique to L. tarentolae or missing from this species may be responsible for the non- pathogenic potential of this lizard parasite. Key words : pathogenic, non-pathogenic, Leishmania major, Leishmania infantum, Leishmania tarentolae, virulence factors. INTRODUCTION Protozoan parasites of the Leishmania genus are causative agents of a broad spectrum of diseases generally termed as leishmaniases. Disease manifes- tations range from cutaneous leishmaniasis caused by Leishmania major, Leishmania aethiopica, Leishmania tropica, Leishmania mexicana and Leishmania ama- zonensis, to muco-cutaneous leishmaniasis caused by Leishmania braziliensis and Leishmania guyanensis and to potentially deadly visceral leishmaniasis or Kala-azar caused by the Leishmania donovani com- plex (L. donovani and Leishmania infantum) (Murray et al. 2005). Leishmaniasis is associated with 2 . 4 million disability-adjusted life years and 70 000 deaths per year (Desjeux, 2004). Extracellular flagellated promastigotes of Leishmania live and multiply in the mid-gut of the female Phlebotomine sandfly. Pro- mastigotes enter the skin of the vertebrate host by the bite of the sandfly, are phagocytosed by immune cells, and transform into non-motile ovoid-shaped amastigotes that are able to propagate within the harsh environment of the macrophage phagolyso- some. Not all members of Leishmania genus are parasites of mammals. Certain species of Leishmania including Leishmania tarentolae are lizard parasites. Whether these parasites should be included within the genus Leishmania has been a matter of debate. However, molecular phylogenetic studies have suggested that lizard Leishmania, alternatively named Sauroleish- mania, have evolved from Old World Leishmania after separation of the New World Vianna subgenus (Croan et al. 1997). L. tarentolae is a lizard parasite strain, which has never been found associated with any leishmaniasis in humans and is thus considered as non-pathogenic to humans. L. tarentolae has been used as a model organism for studying unique features of Kinetoplastids such as RNA editing (Landweber and Gilbert, 1994 ; Maslov et al. 1994). However, as Noyes et al. (1998) have also * Corresponding author : Molecular Immunology and Vaccine Research Laboratory, Pasteur Institute of Iran, Tehran, Iran. Tel: +98 21 66953311. Fax: +98 21 66465132. E-mail : [email protected] or sima-rafatisy@ pasteur.ac.ir # These authors contributed equally to this work. 723 Parasitology (2009), 136, 723–735. f Cambridge University Press 2009 doi:10.1017/S0031182009005873 Printed in the United Kingdom

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Searching for virulence factors in the non-pathogenic
parasite to humans Leishmania tarentolae
H. AZIZI1#, K. HASSANI1,2#, Y. TASLIMI1, H. SHATERI NAJAFABADI3,
B. PAPADOPOULOU4 and S. RAFATI1*
1Molecular Immunology and Vaccine Research Laboratory, Pasteur Institute of Iran, Tehran, Iran2Department of Biotechnology, University College of Science, University of Tehran, Tehran, Iran3 Institute of Parasitology, McGill University, Montreal, Canada4Research Centre in Infectious Diseases, CHUL Research Centre and Department of Medical Biology, Faculty of Medicine,Laval University, 2705 Laurier Blvd., Quebec (QC), Canada G1V 4G2
(Received 15 September 2008; revised 7 November 2008 and 8 January 2009; accepted 8 February 2009; first published online 6 May 2009)
SUMMARY
Leishmania protozoa are obligate intracellular parasites that reside in the phagolysosome of host macrophages and cause a
large spectrum of pathologies to humans known as leishmaniases. The outcome of the disease is highly dependent on the
parasite species and on its ascribed virulence factors and the immune status of the host. Characterization of the genome
composition of non-pathogenic species could ultimately open new horizons in Leishmania developmental biology and also
the disease monitoring. Here, we provide evidence that the lizard non-pathogenic to humans Leishmania tarentolae species
expresses anAmastin-like gene, cysteine protease B (CPB), lipophosphoglycan LPG3 and the leishmanolysinGP63, genes
well-known for their potential role in the parasite virulence. These genes were expressed at levels comparable to those in
L. major and L. infantum both at the level of mRNA and protein. Alignment of the L. tarentolae proteins with their
counterparts in the pathogenic species demonstrated that the degree of similarity varied from 59% and 60% for Amastin,
89% for LPG3 and 71% and 68% forCPB, in L. major and L. infantum, respectively. Interestingly, theA2 gene, expressed
specifically by the L. donovani complex which promotes visceralization, was absent in L. tarentolae. These findings suggest
that the lack of pathogenicity in L. tarentolae is not associated with known virulence genes such as LPG3, CPB,GP63 and
Amastin, and that other factors either unique to L. tarentolae or missing from this species may be responsible for the non-
pathogenic potential of this lizard parasite.
Key words: pathogenic, non-pathogenic, Leishmania major, Leishmania infantum, Leishmania tarentolae, virulence factors.
INTRODUCTION
Protozoan parasites of the Leishmania genus are
causative agents of a broad spectrum of diseases
generally termed as leishmaniases. Disease manifes-
tations range from cutaneous leishmaniasis caused by
Leishmania major, Leishmania aethiopica, Leishmania
tropica, Leishmania mexicana and Leishmania ama-
zonensis, to muco-cutaneous leishmaniasis caused by
Leishmania braziliensis and Leishmania guyanensis
and to potentially deadly visceral leishmaniasis or
Kala-azar caused by the Leishmania donovani com-
plex (L. donovani and Leishmania infantum) (Murray
et al. 2005). Leishmaniasis is associated with 2.4
million disability-adjusted life years and 70000 deaths
per year (Desjeux, 2004). Extracellular flagellated
promastigotes of Leishmania live and multiply in the
mid-gut of the female Phlebotomine sandfly. Pro-
mastigotes enter the skin of the vertebrate host by the
bite of the sandfly, are phagocytosed by immune
cells, and transform into non-motile ovoid-shaped
amastigotes that are able to propagate within the
harsh environment of the macrophage phagolyso-
some.
Not all members of Leishmania genus are parasites
of mammals. Certain species ofLeishmania including
Leishmania tarentolae are lizard parasites. Whether
these parasites should be included within the genus
Leishmania has been a matter of debate. However,
molecular phylogenetic studies have suggested that
lizard Leishmania, alternatively named Sauroleish-
mania, have evolved from Old World Leishmania
after separation of the New World Vianna subgenus
(Croan et al. 1997). L. tarentolae is a lizard parasite
strain, which has never been found associated with
any leishmaniasis in humans and is thus considered
as non-pathogenic to humans. L. tarentolae has
been used as a model organism for studying unique
features of Kinetoplastids such as RNA editing
(Landweber and Gilbert, 1994; Maslov et al.
1994). However, as Noyes et al. (1998) have also
* Corresponding author: Molecular Immunology andVaccine Research Laboratory, Pasteur Institute of Iran,Tehran, Iran. Tel: +98 21 66953311. Fax: +98 2166465132. E-mail : [email protected] or [email protected]# These authors contributed equally to this work.
723
Parasitology (2009), 136, 723–735. f Cambridge University Press 2009
doi:10.1017/S0031182009005873 Printed in the United Kingdom

indicated previously, comparative studies of lizard
Leishmania and virulent Leishmania can be of critical
importance towards understanding the pathogenicity
of Leishmania and its relationships with the host.
Leishmania spp. form complex host-parasite inter-
actions and are able to alter the host immune re-
sponse for their benefit. In order to explain the
absence of pathogenicity in L. tarentolae, we tested
whether known virulence factors of pathogenic
Leishmania species were present and or expressed in
L. tarentolae.
A number ofLeishmania’s virulence factors such as
GP63 (Joshi et al. 2002; McGwire et al. 2002; Yao
et al. 2003; Campbell et al. 1992) and cysteine pro-
teases (CPs) (Alexander et al. 1998; Rosenthal, 1999;
Mundodi et al. 2002; Mottram et al. 2004) have been
characterized and their roles have been extensively
studied; some others such asLPG3 (Descoteaux et al.
2002; Ouakad et al. 2007b), Amastins (Salotra et al.
2006) and A2 (Zhang andMatlashewski, 1997, 2001;
Zhang et al. 2003) have also been identified. GP63
(also called Major Surface Protease) is one of the
most extensively studied virulence factors of Leish-
mania. GP63 is a surface endopeptidase with a wide
range of substrate and pH specificity, well suited for
its dual expression in promastigote and amastigote
stages. Targeted gene deletion studies have shown a
crucial role for GP63 in the pathogenicity of Leish-
mania ; gp63nul mutants of L. major showed reduced
pathogenicity and were weakened in forming lesions
in BALB/C mice in comparison to wild types (Joshi
et al. 2002). As another group of virulence factors of
Leishmania, the cysteine proteases have been studied
mostly in L. mexicana due to their crucial role in its
pathogenicity. Most studies on cysteine proteases in
L. mexicana have been done on the most abundant
Cysteine Protease families A, B and C. Similar to
gp63x/x L. major, Dcpb L. mexicana have impaired
infectivity against BALB/c mice (Mottram et al.
1998; Alexander et al. 1998). Furthermore, Williams
et al. (2006) have shown that CPB and CPA are es-
sential for parasite autophagy during promastigote-
amastigote transformation. The dominant surface
molecule of promastigotes is lipophosphoglycan
(LPG). This molecule plays important roles in the
early stages of the infection, wherein the parasite is
still in the promastigote form, namely interaction
with the complement system, phagocytosis and pro-
tection of promastigotes against lysis in the phago-
lysosome (Spath et al. 2003). LPG3, a molecular
chaperone, is part of the metabolic pathway of
synthesis of LPG as well as phosphoglycan residues
that are added to extracellular proteins, such asGP63,
GP46, sAP and GPI-anchored proteins (Descoteaux
et al. 2002). The expression level of surface proteins
such as GP63 and GP46 is greatly reduced in lpg3nul
mutants (Descoteaux et al. 2002). Based on these
findings it was proposed that the role of LPG3 in
virulence should be exerted through its role in the
synthesis of LPG, GPI-anchored proteins (such as
GP63) and other phosphoglycan-bearing (PG)
molecules (Descoteaux et al. 2002). Amastins belong
to a large gene family of surface proteins that are
developmentally regulated in the amastigote stage of
Leishmania (Wu et al. 2000). Although the biological
function of amastins remains still unknown, it is
hypothesized that amastins may play a role in proton
or ion traffic across the membrane to adjust the
cytoplasmic pH under the harsh conditions of the
phagolysosome (Rochette et al. 2005). Like amastins,
A2 is an amastigote-specific protein, originally dis-
covered in L. donovani. The exact function of A2
protein in Leishmania is unknown. However, gene
silencing ofA2 inL. donovani dramatically decreased
its viability in mammalian macrophages and deterio-
rated its pathogenicity (Zhang and Matlashewski,
1997). Interestingly, over-expression of A2 in
L. major resulted in visceralization of the infection
(Zhang and Matlashewski, 1997; Garin et al. 2005).
In this study, we tested whether well-character-
ized virulence factors in the pathogenic Leishmania
species (e.g. L. major, L. infantum and L. braziliensis)
were present and or expressed in the non-pathogenic
L. tarentolae species in order to gain a better insight
into the factors responsible for the lack of patho-
genicity in L. tarentolae. We found that all the well-
known virulence factors that we tested here, with the
only exception of A2, were present and expressed in
L. tarentolae at ratios similar to those of the patho-
genic species.
MATERIALS AND METHODS
Preparation of stationary phase promastigotes
L. major (MRHO/IR75/ER), L. infantum (MCAN/
98/LLM-877) and L. tarentolae (ATCC 30267)
promastigotes were grown at 26 xC in complete
M199medium containing 10% heat-inactivated FCS,
40 mMHEPES, 0.1 mM adenosine, 0.5 mg/ml haemin
and 50 mg/ml gentamicin, and were allowed to mul-
tiply until they reached a density of 2–3r107 fol-
lowing 5 days of growth at 26 xC. The stationary
phase promastigotes were then utilized for macro-
phage cells (MQ) infection and RNA extraction.
Probe preparation
The ORF of cysteine protease type I (GenBank
Accession no. U43706) was digested using BamHI-
HindIII from vector pGEM-cpb (Rafati et al. 2001).
The ORF of GP63 from L. major (GenBank Ac-
cession no. AF039721) was digested usingXbaI from
a vector kindly provided by Dr Mahboudi (Pasteur
Institute of Iran). LPG3 ORF (GeneDB Accession
no. LmjF29.0760) was digested using EcoRI-SalI
from pGEM-lpg3 vector prepared previously in
our lab (unpublished). The A2 ORF (Accession no.
H. Azizi and others 724

S68693) was digested with BamHI from a vector
kindly provided by Dr Greg Matlashewski (McGill
University, Canada). Partial sequences from amas-
tins Lmj24.1270, Lmj28.1390, Lmj30.0870 were
digested by EcoRI from vectors described previously
(Rochette et al. 2005). Following digestion and ex-
traction of fragments from agarose gel, probes were
prepared by random primer extension using Klenow
enzyme (Roche). In certain sections of library
screening, amastin probes were prepared by a non-
radioactive Digoxigenin (DIG) labelling (Roche) in a
similar process.
Total DNA preparation and hybridizations
Total DNA of L. tarentolae (strain ATCC 30267),
L. major (strain MRHO/IR75/ER) and L. infantum
(MCAN/98/LLM-877) was prepared from log-
phase parasites using the standard proteinase K
(Roche) and phenol extraction method (Sambrook
and Russell, 2001) and digested using PstI (Roche).
Southern blotting was done according to standard
methods (Sambrook and Russell, 2001). Resulting
Nylon membranes (Roche) were hybridized to32P-labelled probes in 20 ml of Church buffer (0.5 M
phosphate-buffered saline (PBS) containing 1%
BSA, 1 mM EDTA and 7% SDS) at 65 xC overnight.
Non-stringent and stringent washes were done in
25 ml of NaPi buffer for 15 min at 25 xC and 65 xC,
respectively. When necessary, non-stringent wash-
ing at 42 xC was also performed for confirmation.
Radiofilms (Kodak) were exposed overnight or for 2
days for confirmation.
Library screening
One mg of the previously described cosmid library of
L. tarentolae (Genest et al. 2008) was transformed to
E. coli DH5a by chemical transformation. Colony
lifting was done according to standard methods
(Sambrook and Russell, 2001) and the resulting
nylon membranes were hybridized to the probe for
L. major’s amastin Lmj24.1270 under non-stringent
conditions as stated in the previous section. The
transformation and screening was performed 6 times
in order to achieve high genome coverage. Cosmids
that were hybridized to the amastin probe were
digested using PstI and re-blotted against the probe
and the hybridizing fragment was subcloned in
pGEM-2 vector (Promega) after dephosphorylation
(Shrimp Alkaline Phosphatase, Roche). The cloned
fragment was sequenced using universal T7 and SP6
primers.
Sequence analysis
Orthologues of the sequences acquired from
L. tarentolae were found in the genomes of L. major,
L. infantum and L. braziliensis using GeneDB
BLAST search tool (for each gene, the hit with
the lowest E-value was chosen). Pairwise sequence
alignments were performed on translations of the
acquired sequences at EBI website. Multiple se-
quence alignment was performed using web-based
CLUSTALW at EBI bioinformatics tools. Trans-
membrane helices of L. tarentolae’s amastin were
predicted using TMHMM server version 2 (Krogh
et al. 2001).
Amastigote preparation
Peritoneal exudate cells from naıve BALB/c mice
were used as a source of resident macrophages (MQ).
Briefly, the peritoneal cavity was washed with 5 ml
of cold RPMI 1640 (Sigma) medium, and then
the medium containing peritoneal macrophages was
recovered. Cells were cultured in RPMI medium
1640 supplemented with 5% FCS, 2 mM glutamine,
50 mM 2-mercaptoethanol, 10 mM HEPES and
40 mg/ml gentamicin at a density of 106 cells/well, and
incubated at 37 xC in 5% CO2 for 24 h. In order
to infect MQ, the stationary phase Leishmania pro-
mastigotes were added at the ratio of 10 : 1 (parasites
versus macrophages). The cultures were washed
after 4 h to exclude free parasites and then incubated
for 5 days. Infected attached MQs were collected
using cold PBS 1X (Breton et al. 2005).
RNA extraction
Stationary phase promastigotes and amastigotes of
L. tarentolae, L. major and L. infantum were utilized
for RNA purification using RNeasy Mini kit
(Qiagen). Briefly, after washing the promastigotes
twice with PBS, total RNA was extracted according
to the supplier kit. For total amastigote RNA iso-
lation, the infected murine peritoneal macrophages
were detached from the plate with cold PBS and
lysed according to QIAGEN RNAeasy kit. To pre-
vent co-purifying genomic DNA with RNA, on
column DNase treatment was done using RNase
free DNase set (Qiagen). The quality of the RNA
was assessed by spectrophotometric analysis using
NanoDrop ND-1000. Integrity of the RNA was
checked on 1% formaldehyde agarose gels. To ensure
that the obtained amastigote RNA is free of host
(macrophage) RNA, we looked for the presence of
murine ribosomal RNA on the gel.
cDNA synthesis
One mg of total RNA from each sample was reverse-
transcribed to cDNA using the Omniscript RT kit
(Qiagen) and random hexamer primers as described
by the supplier. After preparation of the mixture,
samples were incubated at 37 xC for 1 h, heat-
inactivated at 70 xC for 6 min and subsequently
stored in x20 xC.
Virulence factors in Leishmania tarentolae 725

Real-time PCR
cDNAs from all samples were utilized as a template
for the real-time polymerase chain reaction. Quanti-
fication of the mRNA corresponding to LPG3,
Amastin and CPB was done in the 7500 real-time
PCR system (Applied Biosystems) using QuantiFast
SYBR Green master mix from Qiagen. Amplifi-
cation of theAmastin fragment (185 bp) fromL.major
(Accession no. LmjF28.1400) and L. infantum (Ac-
cession no. LinJ28.1460) mRNA was carried out
using the same primers (AmastinF, 5k-GCGGTGG-
AGACGTGCTG-3k and AmastinR, 5k-GTGTAG-
GTAGAGTTGTCGCAGTTGT-3k). Amplification
of a 100 bp region from the L. tarentolae Amastin
transcript (GenBank Accession no. EU041954.1)
was accomplished using a primer pair specific to this
parasite (Amastin-tF, 5k-GCTTACCTGATGAT-
GGCGGATA-3k and Amastin-tR, 5k-CCAGTTG-
CGATGTGCAGATC-3k). The primers for LPG3
(LPG3F, 5k-TCTTCATCACGGACGAGTTCC-
3k and LPG3R, 5k-TTGCGCACAAGTTTCTT-
CTT-3k) were designed from a conserved region in
a multiple sequence alignment of L. major, L. in-
fantum, L. donovani and L. braziliensis LPG3
genes (Accession nos LmjF29.0760, LinJ29.0920,
AF369892 and LbrM29-V2.0780, respectively) and
were successfully used in amplifying the LPG3
transcript of L. tarentolae. The primers CPB-F,
5k-AACTTCGAGCGCAACCT-3k and CPB-R,
5k-GGTCAAAGAACTTCGTGATC-3k also servedfor amplifying an 85 bp fragment of L. infantum
(LinJ08.0980) and of L. major (LmjF08.1080).
Likewise, utilizing the primers CPB-LtF: 5k-AGC-
GGCGGGCTGATGT-3k and CPB-LtR: 5k-GAG-
CACTCGGGCGCATAG-3k, a 115 bp region of
L. tarentolae CPB (Accesion. no. EU860161) was
successfully obtained. The primers that served
to amplify a 100 bp rRNA region (LmjF32.3420
and LinJ32.3980 for L. major and L. infantum,
respectively) were rRNA-F: 5k-CGGTTTTTCC-
CTGATGTGGTAA-3k and rRNA-R: 5k-GGAA-
GCGTGGTCGACGTAGA-3k. Finally, GAPDH
from all species was amplified using the 5k-TCG-
CTCTCGTGGACATGAGCA-3k (forward) and
5k- CTTGAGCGACACGCCGTCCA-3k (reverse).PCR reactions were performed at 95 xC for 5 min
followed by 40 cycles of 95 xC for 15 s and 60 xC for
1 min. Melting curve analysis demonstrated specific
single PCR products for all reactions according to
the amplicon melting temperatures. Furthermore,
2 housekeeper genes, GAPDH and rRNA45 were
employed as internal controls for normalization. All
the primers were designed utilizing Primer Express
software v.2 (Applied Biosystems). Relative quantity
of all genes was determined through DDCT Method
(SDS software, Applied Biosystems). RT negative
controls were also performed to check for the pres-
ence of contaminated DNA.
SDS-PAGE and immunoblotting
Stationary phase promastigotes were collected from
in vitro cultures by centrifugation (10 min, 1500 g,
4 xC), washed in PBS and lysed in SDS-PAGE
samplebuffercontaining100 mMTris-HCl,4%SDS,
0.2% bromophenol blue, 20% glycerol and 200 mM
b-mercaptoethanol. The lysate was separated on a
12.5% SDS-PAGE polyacrylamide gel and then
transferred to a nitrocellulose membrane (Bio-Rad
system). In addition, rLPG3 (Rafati et al. unpub-
lished data), rCPB (Rafati et al. 2001) and synthetic
amastin signature (Rafati et al. 2006) were used as a
control. After protein transfer, the membrane was
blocked overnight at 4 xC in TBS (10 mM Tris-HCl,
pH 7.4, 150 mM NaCl) containing 2.5% BSA for
2 h, then incubated overnight with either a rabbit
anti-CPB polyclonal antibody (1 : 500), a mouse anti-
LPG3 polyclonal antibody (1 : 900) or a mouse
anti-amastin polyclonal antibody (1 : 600) in TBS.
The membranes were washed 3 times with TBS,
0.1% Tween 20 and incubated for 1.5 h with either
peroxidase-conjugated anti-rabbit (1 : 4000 Sigma)
or peroxidase-conjugated goat anti-mouse IgG
(1 : 6000, Sigma) as secondary antibodies. Unbound
secondary antibodies were removed by washing
as described above. Diaminobenzidine tetrahydro-
chloride (DAB, Sigma) was used as the substrate to
detect the desired bands of the proteins.
RESULTS
Presence of known virulence factors in the
non-pathogenic Leishmania tarentolae species
First, we looked whether the known virulence factors
of pathogenic Leishmania species CPB, GP63,
LPG3, A2 and Amastin were present in lizard non-
pathogenic L. tarentolae. Total DNA from the
pathogenic parasites L. major and L. infantum and
from the non-pathogenic species L. tarentolae was
digested, transferred to nylon membranes and hy-
bridized with gene-specific probes corresponding to
the coding regions of the above virulence factors.
The results presented in Fig. 1 demonstrate that
the genes for the cysteine peptidase B family, GP63
family, amastin family and LPG3 are present in
L. tarentolae. Interestingly, the band patterns appear
to be similar between the three species especially for
CPB and GP63. The observed multiple banding in
lanes corresponding to CPB and GP63 could be due
to the presence of multiple copies of these genes in
the genomes of these parasites. A similar pattern
in L. tarentolae suggests that these factors are also
present in multiple copies. Detection of the amastin
genes in L. tarentolae required low-stringency con-
ditions for hybridization. Screening of aL. tarentolae
library was performed to seek homologues of the
amastin LmjF24.1270, which is highly conserved
between L. major and L. infantum. Partial screening
H. Azizi and others 726
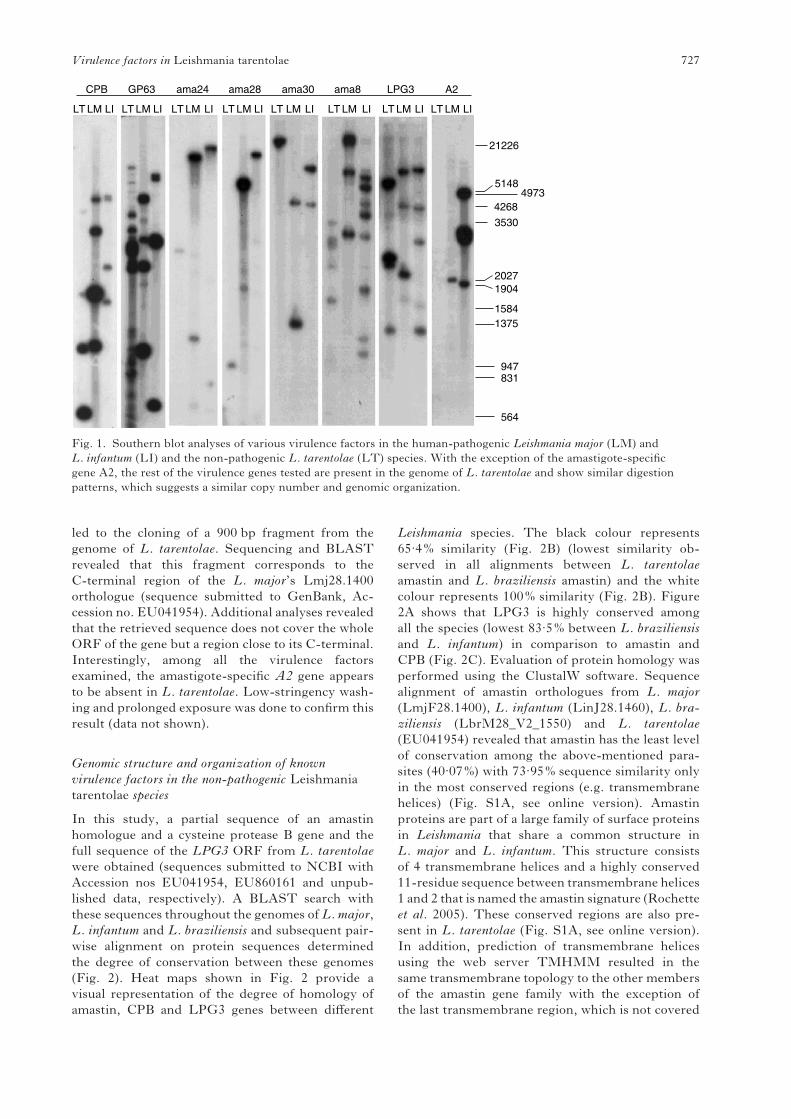
led to the cloning of a 900 bp fragment from the
genome of L. tarentolae. Sequencing and BLAST
revealed that this fragment corresponds to the
C-terminal region of the L. major’s Lmj28.1400
orthologue (sequence submitted to GenBank, Ac-
cession no. EU041954). Additional analyses revealed
that the retrieved sequence does not cover the whole
ORF of the gene but a region close to its C-terminal.
Interestingly, among all the virulence factors
examined, the amastigote-specific A2 gene appears
to be absent in L. tarentolae. Low-stringency wash-
ing and prolonged exposure was done to confirm this
result (data not shown).
Genomic structure and organization of known
virulence factors in the non-pathogenic Leishmania
tarentolae species
In this study, a partial sequence of an amastin
homologue and a cysteine protease B gene and the
full sequence of the LPG3 ORF from L. tarentolae
were obtained (sequences submitted to NCBI with
Accession nos EU041954, EU860161 and unpub-
lished data, respectively). A BLAST search with
these sequences throughout the genomes of L. major,
L. infantum and L. braziliensis and subsequent pair-
wise alignment on protein sequences determined
the degree of conservation between these genomes
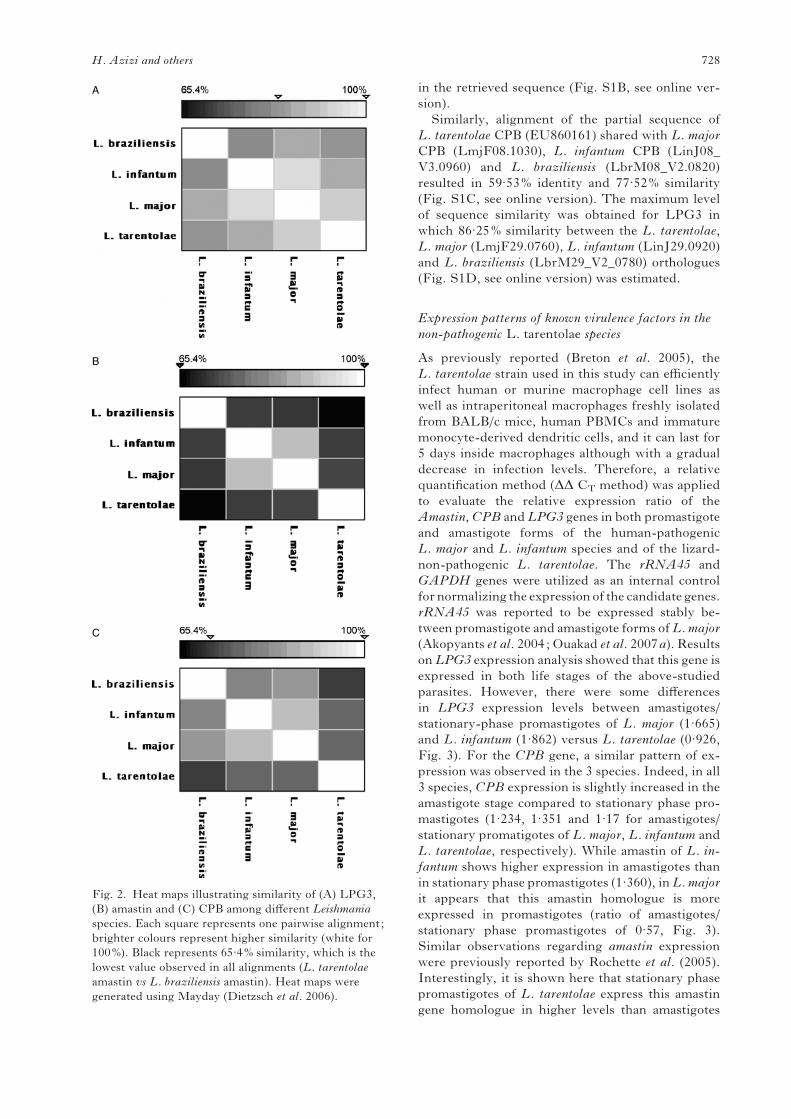
(Fig. 2). Heat maps shown in Fig. 2 provide a
visual representation of the degree of homology of
amastin, CPB and LPG3 genes between different
Leishmania species. The black colour represents
65.4% similarity (Fig. 2B) (lowest similarity ob-
served in all alignments between L. tarentolae
amastin and L. braziliensis amastin) and the white
colour represents 100% similarity (Fig. 2B). Figure
2A shows that LPG3 is highly conserved among
all the species (lowest 83.5% between L. braziliensis
and L. infantum) in comparison to amastin and
CPB (Fig. 2C). Evaluation of protein homology was
performed using the ClustalW software. Sequence
alignment of amastin orthologues from L. major
(LmjF28.1400), L. infantum (LinJ28.1460), L. bra-
ziliensis (LbrM28_V2_1550) and L. tarentolae
(EU041954) revealed that amastin has the least level
of conservation among the above-mentioned para-
sites (40.07%) with 73.95% sequence similarity only
in the most conserved regions (e.g. transmembrane
helices) (Fig. S1A, see online version). Amastin
proteins are part of a large family of surface proteins
in Leishmania that share a common structure in
L. major and L. infantum. This structure consists
of 4 transmembrane helices and a highly conserved
11-residue sequence between transmembrane helices
1 and 2 that is named the amastin signature (Rochette
et al. 2005). These conserved regions are also pre-
sent in L. tarentolae (Fig. S1A, see online version).
In addition, prediction of transmembrane helices
using the web server TMHMM resulted in the
same transmembrane topology to the other members
of the amastin gene family with the exception of
the last transmembrane region, which is not covered
21226
A2LPG3ama8ama30ama28ama24GP63CPB
LT LM LI LT LM LI LT LM LI LT LM LI LT LM LI LT LM LI LT LM LI LT LM LI
51484973
4268
3530
20271904
15841375
947831
564
Fig. 1. Southern blot analyses of various virulence factors in the human-pathogenic Leishmania major (LM) and
L. infantum (LI) and the non-pathogenic L. tarentolae (LT) species. With the exception of the amastigote-specific
gene A2, the rest of the virulence genes tested are present in the genome of L. tarentolae and show similar digestion
patterns, which suggests a similar copy number and genomic organization.
Virulence factors in Leishmania tarentolae 727
in the retrieved sequence (Fig. S1B, see online ver-
sion).
Similarly, alignment of the partial sequence of
L. tarentolae CPB (EU860161) shared with L. major
CPB (LmjF08.1030), L. infantum CPB (LinJ08_
V3.0960) and L. braziliensis (LbrM08_V2.0820)
resulted in 59.53% identity and 77.52% similarity
(Fig. S1C, see online version). The maximum level
of sequence similarity was obtained for LPG3 in
which 86.25% similarity between the L. tarentolae,
L. major (LmjF29.0760), L. infantum (LinJ29.0920)
and L. braziliensis (LbrM29_V2_0780) orthologues
(Fig. S1D, see online version) was estimated.
Expression patterns of known virulence factors in the
non-pathogenic L. tarentolae species
As previously reported (Breton et al. 2005), the
L. tarentolae strain used in this study can efficiently
infect human or murine macrophage cell lines as
well as intraperitoneal macrophages freshly isolated
from BALB/c mice, human PBMCs and immature
monocyte-derived dendritic cells, and it can last for
5 days inside macrophages although with a gradual
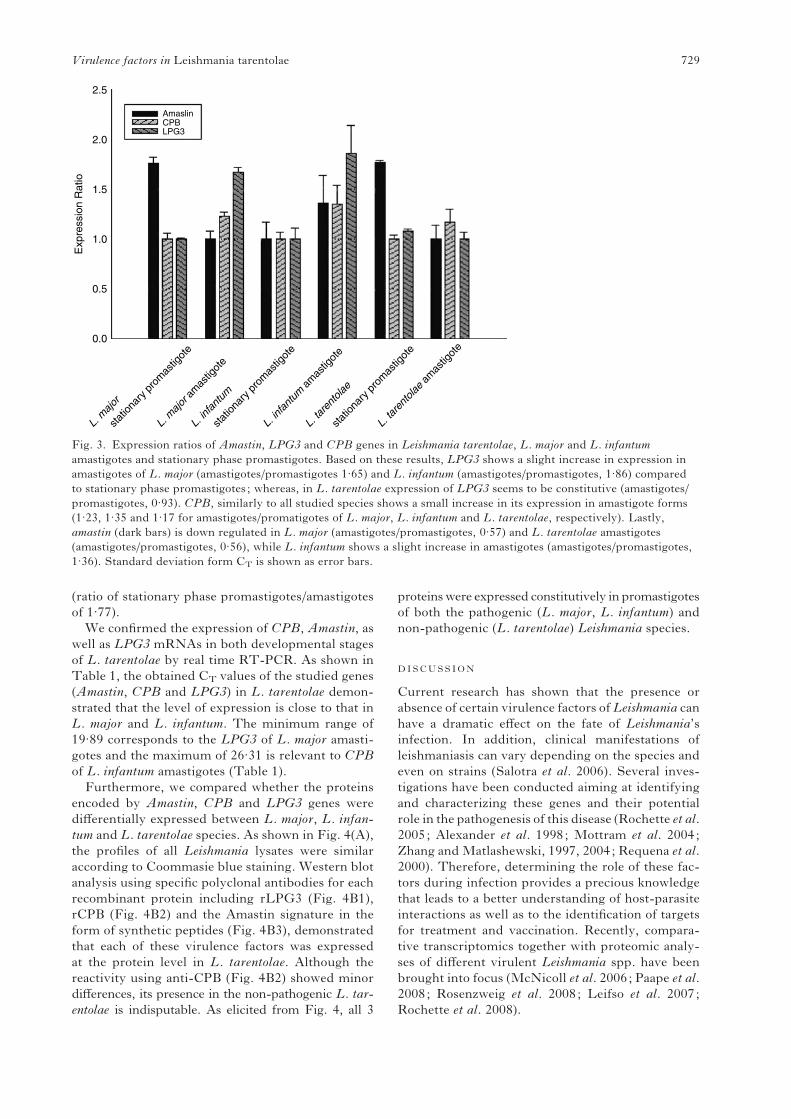
decrease in infection levels. Therefore, a relative
quantification method (DD CT method) was applied
to evaluate the relative expression ratio of the
Amastin,CPB and LPG3 genes in both promastigote
and amastigote forms of the human-pathogenic
L. major and L. infantum species and of the lizard-
non-pathogenic L. tarentolae. The rRNA45 and
GAPDH genes were utilized as an internal control
for normalizing the expression of the candidate genes.
rRNA45 was reported to be expressed stably be-
tween promastigote and amastigote forms ofL. major
(Akopyants et al. 2004; Ouakad et al. 2007a). Results
on LPG3 expression analysis showed that this gene is
expressed in both life stages of the above-studied
parasites. However, there were some differences
in LPG3 expression levels between amastigotes/
stationary-phase promastigotes of L. major (1.665)
and L. infantum (1.862) versus L. tarentolae (0.926,
Fig. 3). For the CPB gene, a similar pattern of ex-
pression was observed in the 3 species. Indeed, in all
3 species, CPB expression is slightly increased in the
amastigote stage compared to stationary phase pro-
mastigotes (1.234, 1.351 and 1.17 for amastigotes/
stationary promatigotes of L. major, L. infantum and
L. tarentolae, respectively). While amastin of L. in-
fantum shows higher expression in amastigotes than
in stationary phase promastigotes (1.360), inL. major
it appears that this amastin homologue is more
expressed in promastigotes (ratio of amastigotes/
stationary phase promastigotes of 0.57, Fig. 3).
Similar observations regarding amastin expression
were previously reported by Rochette et al. (2005).
Interestingly, it is shown here that stationary phase
promastigotes of L. tarentolae express this amastin
gene homologue in higher levels than amastigotes
A
B
C
Fig. 2. Heat maps illustrating similarity of (A) LPG3,
(B) amastin and (C) CPB among different Leishmania
species. Each square represents one pairwise alignment;
brighter colours represent higher similarity (white for
100%). Black represents 65.4% similarity, which is the
lowest value observed in all alignments (L. tarentolae
amastin vs L. braziliensis amastin). Heat maps were
generated using Mayday (Dietzsch et al. 2006).
H. Azizi and others 728
(ratio of stationary phase promastigotes/amastigotes
of 1.77).
We confirmed the expression of CPB, Amastin, as
well as LPG3 mRNAs in both developmental stages
of L. tarentolae by real time RT-PCR. As shown in
Table 1, the obtained CT values of the studied genes
(Amastin, CPB and LPG3) in L. tarentolae demon-
strated that the level of expression is close to that in
L. major and L. infantum. The minimum range of
19.89 corresponds to the LPG3 of L. major amasti-
gotes and the maximum of 26.31 is relevant to CPB
of L. infantum amastigotes (Table 1).
Furthermore, we compared whether the proteins
encoded by Amastin, CPB and LPG3 genes were
differentially expressed between L. major, L. infan-
tum and L. tarentolae species. As shown in Fig. 4(A),
the profiles of all Leishmania lysates were similar
according to Coommasie blue staining. Western blot
analysis using specific polyclonal antibodies for each
recombinant protein including rLPG3 (Fig. 4B1),
rCPB (Fig. 4B2) and the Amastin signature in the
form of synthetic peptides (Fig. 4B3), demonstrated
that each of these virulence factors was expressed
at the protein level in L. tarentolae. Although the
reactivity using anti-CPB (Fig. 4B2) showed minor
differences, its presence in the non-pathogenic L. tar-
entolae is indisputable. As elicited from Fig. 4, all 3
proteinswere expressed constitutively in promastigotes
of both the pathogenic (L. major, L. infantum) and
non-pathogenic (L. tarentolae) Leishmania species.
DISCUSSION
Current research has shown that the presence or
absence of certain virulence factors ofLeishmania can
have a dramatic effect on the fate of Leishmania’s
infection. In addition, clinical manifestations of
leishmaniasis can vary depending on the species and
even on strains (Salotra et al. 2006). Several inves-
tigations have been conducted aiming at identifying
and characterizing these genes and their potential
role in the pathogenesis of this disease (Rochette et al.
2005; Alexander et al. 1998; Mottram et al. 2004;
Zhang andMatlashewski, 1997, 2004; Requena et al.
2000). Therefore, determining the role of these fac-
tors during infection provides a precious knowledge
that leads to a better understanding of host-parasite
interactions as well as to the identification of targets
for treatment and vaccination. Recently, compara-
tive transcriptomics together with proteomic analy-
ses of different virulent Leishmania spp. have been
brought into focus (McNicoll et al. 2006; Paape et al.
2008; Rosenzweig et al. 2008; Leifso et al. 2007;
Rochette et al. 2008).
AmaslinCPBLPG3
2.5
2.0
1.5
1.0
Exp
ress
ion
Rat
io
0.5
0.0
L. majo
r
L. majo
r am
astig
ote
L. inf
antum
amas
tigot
e
L. inf
antum
L. tar
entol
ae am
astig
ote
L. tar
entol
ae
statio
nary
pro
mas
tigot
e
statio
nary
pro
mas
tigot
e
statio
nary
pro
mas
tigot
e
Fig. 3. Expression ratios of Amastin, LPG3 and CPB genes in Leishmania tarentolae, L. major and L. infantum
amastigotes and stationary phase promastigotes. Based on these results, LPG3 shows a slight increase in expression in
amastigotes of L. major (amastigotes/promastigotes 1.65) and L. infantum (amastigotes/promastigotes, 1.86) compared
to stationary phase promastigotes ; whereas, in L. tarentolae expression of LPG3 seems to be constitutive (amastigotes/
promastigotes, 0.93). CPB, similarly to all studied species shows a small increase in its expression in amastigote forms
(1.23, 1.35 and 1.17 for amastigotes/promatigotes of L. major, L. infantum and L. tarentolae, respectively). Lastly,
amastin (dark bars) is down regulated in L. major (amastigotes/promastigotes, 0.57) and L. tarentolae amastigotes
(amastigotes/promastigotes, 0.56), while L. infantum shows a slight increase in amastigotes (amastigotes/promastigotes,
1.36). Standard deviation form CT is shown as error bars.
Virulence factors in Leishmania tarentolae 729

Although Leishmania pathogenicity is most likely
the result of a high complexity with more than one
mechanism involved, it is possible to identify viru-
lence factors by genomic comparisons between
pathogenic and non-pathogenic Leishmania species.
It has been reported that L. tarentolae is likely to be
pathogenic for lizards but the mechanism(s) is un-
known (Dollahon et al. 1973; Orlando et al. 2002).
However, L. tarentolae is unable to cause disease
in mice, even in immunocompromized mice (Breton
et al 2005), and has never been associated with a
Leishmania disease in humans. Here, we investigated
whether known virulence factors of Leishmania
were present and also expressed in the lizard L. tar-
entolae non-pathogenic species to humans. Among
them, we tested the cysteine proteases, GP63, LPG3,
A2 and amastins and found that, with the exception
of A2, all the other proteins were expressed in
L. tarentolae. Using antisense RNA, it was shown
that the L. donovani A2 gene is essential for survival
of amastigotes both in vitro cultured macrophages as
well as in mice (Zhang and Matlashewski, 1997).
Furthermore, using a recombinant A2 protein as
vaccine against L. amazonensis infection in suscep-
tible mice, a robust protection has been obtained
(Coelho et al. 2003). Because A2 protein is truncated
in L. major (Ghedin et al. 1998; Zhang et al. 2003),
it was suggested that A2 might contribute to the
tropism ofL. donovani to visceralization (Zhang et al.
2003). The absence of A2 in L. tarentolae is in-
triguing and it may be associated to some extent with
the loss of pathogenicity of lizard Leishmania in
mammals. This needs to be experimentally vali-
dated, however.
Similar to L. major and L. infantum, L. tarentolae’s
GP63 is a multi-copy gene. The role of GP63 in
pathogenesis has been widely studied (Bordier, 1987;
Medina-Acosta et al. 1993; McMaster et al. 1994;
Yao et al. 2003) although its function in disease
development and also pathogenesis has not been
clearly defined. Chen et al. (2000) have shown that
down regulation ofGP63 expression inL. braziliensis
using antisense RNA strategy led to impaired
internalization of macrophage cells in vitro. On the
contrary, a similar study on the effect of L. major
gp63x/x in disease progression resulted in impaired
macrophage infection, although delayed infectivity
of challenged BALB/cmice was observed (Joshi et al.
2002). On the basis of this finding, it was proposed
that GP63 might contribute to additional pathogenic
pathways such as migration through the extra-
cellular matrix. Interestingly, it has also been shown
(Gregory et al. 2008) that L. tarentolae is able to
perform GP63-dependent cleavages in host proteins
during infection.
In this study we also showed that LPG3,CPBs and
amastins were expressed in L. tarentolae. LPG3 is an
endoplasmic reticulum chaperone and homologue
of human GRP94 (Larreta et al. 2000), which is
believed to be crucial for proper functioning of
LPG synthesis pathway (Descoteaux et al. 2002). In
L. major, it has been shown that mutants lacking this
gene are avirulent (Spath et al. 2000), while in
L. mexicana, it has been demonstrated that LPG is
not essential for macrophage infection as well as
lesion development in mice (Ilg, 2000). It is worth
mentioning that in contrast to the pathogenic
Leishmania, LPG appears to be absent from the
lizard parasite L. tarentolae (Previato et al. 1997).
In this study, we have shown that LPG3 is pref-
erentially expressed in amastigotes of L. major and
L. infantum, which is consistent with similar findings
from L. donovani (Descoteaux et al. 2002). None-
theless, in L. tarentolae, we found that LPG3 gene is
constitutively expressed. Species-specific differences
even between pathogenic Leishmania species have
recently been reported (Rochette et al. 2008). These
differences point to a differential gene regulation
possibly by sequences in the 3kUTR, which generally
are less conserved between species than the protein-
coding sequences (Peacock et al. 2007). Multiple
sequence alignment of the LPG3 protein between
the 4 studied Leishmania species showed a high
level of conservation for the LPG3 orthologues
(86.25%).
Among Leishmania cysteine proteinases (CPA,
CPB and CPC), the role of CPB in pathogenesis has
been extensively highlighted, as L. mexicana de-
ficient in the cpb array genes showed reduced in-
fectivity in experimentally infected BALB/c mice
(Frame et al. 2000; Alexander et al. 1998). It has been
reported that recombinant hybrid proteins encom-
passing CPA and CPB could elicit protection against
L. major in BALB/c mice (Zadeh-Vakili et al. 2004).
Furthermore, the vaccine trial in dogs using cocktails
of CPA and CPB gave satisfactory results (Rafati
et al. 2005), although further study is required.
Alongside these findings, intensive efforts have been
conducted towards identifying the role of CPB in
virulence. Results on L. mexicana revealed that CPB
promotes survival and growth of intracellular para-
sites living in phagolysosomes through inhibition of
Table 1. CT values from two developmental stages
of the three Leishmania species are shown
(According to these results, expression levels of the studiedgenes in L. tarentolae are comparable to those of the otherpathogenic species.)
Species
Obtained CT values
Amastin CPB LPG3
L. major promastigote 22.38 23.31 21.17L. major amastigote 22.98 22.47 19.89L. infantum promastigote 23.96 26.24 22.14L. infantum amastigote 24.02 26.31 21.75L. tarentolae promastigote 25.01 22.73 21.26L. tarentolae amastigote 25.15 22.81 20.69
H. Azizi and others 730

the NF-kB pathway and degradation of MHC class
II molecules thereby preventing IL-12 production
and eliciting IL-4 expression (Mottram et al. 2004).
However, infection of L. major-resistant mouse
strains with L. mexicana resulted in lesion develop-
ment due to the ability of L. mexicana to cleave a
component of NF-kB, a capacity that seems to be
absent in L. major (Cameron et al. 2004; Buxbaum
et al. 2003). Moreover, these studies demonstrated
that infection of macrophages with wild type or
cpbx/x L. mexicana elicited similar levels of IL-12
production, suggesting that other antigen-presenting
cells (APCs) may be involved in vivo, instead of
macrophages or even modulate other Th1-inducing
cytokines (such as IL-18) (Fukao et al. 2000). Given
the above-discussed functions of CPB, it is sur-
prising that the non-pathogenic L. tarentolae does
possess and express CPB in its genome. Besides,
Southern blot analysis suggested thatCPB is amulti-
copy gene in L. tarentolae, similar to the pathogenic
species. Our analysis showed that CPB is expressed
slightly more in the amastigote stage in all 3 parasitic
species. Such a high level of similarity in CPB ex-
pression, even at the protein level, between patho-
genic and non-pathogenic species is a noticeable
finding that needs additional studying, especially at
the level of protein function. Characterization of this
protein in L. tarentolae will be interesting due to the
differences in protein sequence and possibly struc-
ture in comparison to the L. major and L. infantum
CPB. Heat map analysis and alignments of the
obtained partial sequence of CPB from L. tarentolae
with its equivalent in L. major, L. infantum and
L. braziliensis have predicted 71–77% similarity.
Meanwhile, the maximum sequence similarity of
88.4% was estimated between L. major and L. in-
fantum. Based on this fact, one could conclude that
CPB should have a high degree of heterogeneity over
different Leishmania species, which is compatible
with previous findings on its copy number (Mottram
et al. 2004) and variation in the C-terminal extension
(Hide et al. 2007; Nakhaee et al. 2004; Rafati et al.
2003).
Finally, we analysed the presence and organization
of the amastigote-specific amastin gene family in
L. tarentolae. Amastins are the largest family of de-
velopmentally regulated surface proteins in Leish-
mania (Rochette et al. 2005). Although their role has
not yet been characterized, it is suggested that these
proteins may be involved in the transportation of
ions, metals and nutrients by the internalized para-
sites within the phagolysosome (Rochette et al.
2005). As suggested by Southern blot hybridization,
L. tarentolae also has multiple copies of this gene
family. However, the ongoing sequence of the
L. tarentolae genome indicated a significantly smaller
number of amastins in this non-pathogenic species to
humans compared to the pathogenic Leishmania
(Papadopoulou et al., unpublished observations).
Heat map analysis and sequence alignment of the
L. tarentolae amastin homologue revealed that this
putative amastin protein does possess the 11 amino
acid amastin signature sequence, which is charac-
teristic of all amastin proteins. The results of real-
time PCR showed that amastin is differentially
expressed between amastigotes and stationary-phase
promastigotes. Meanwhile, L. major and L. tar-
entolae express this gene preferentially in stationary
phase promastigotes, whereas in L. infantum this
gene is preferentially expressed in amastigotes as
previously reported (Rochette et al. 2005). Western
blot analysis confirmed amastin protein expression
in L. tarentolae promastigotes, as well. It is worth
mentioning that like LPG3 and CPB, expression
levels of amastin in L. tarentolae are comparable to
those of the pathogenic species. The L. tarentolae
A B
B1
B2
B3
Fig. 4. SDS-PAGE and Western blot analysis of LPG3, CPB and amastin proteins in Leishmania tarentolae,
L. infantum and L. major. (A) Lysates of L. tarentolae (lane 2), L. infantum (Lane 3), L. major (lane 4), rLPG3
(Lane 5), rCPB (Lane 6) and amastin signature (Lane 7) were subjected to electrophoresis on 12.5% SDS-PAGE
and stained with Coomasie blue. (B) Same samples were transferred to a nitrocellulose membrane and incubated with
different polyclonal antibodies including anti-LPG3 (B1), anti-CPB (B2) and anti-amastin (B3) as described in the
Materials and Methods section.
Virulence factors in Leishmania tarentolae 731

amastin homologue shows 65–72% sequence hom-
ology to the closest orthologues in L. major, L. in-
fantum and L. braziliensis as compared to 89.8%
sequence identity between L. major and L. infantum.
The lower degree of similarity of the L. tarentolae
amastin could affect interactions with other proteins
or substrates and hence modulate amastin’s function.
Amastin’s putative role in pathogenesis has been
recently suggested by DNA microarray data, in-
dicating that amastin genes were differentially ex-
pressed between differentL. donovani strains isolated
from PKDL and VL patients (Salotra et al. 2006).
However, the exact role of amastin in the disease
progression has not yet been identified.
In summary, to our knowledge, this work provides
the first comparative genomic analysis and ex-
pression profiles of important virulence genes, such
as CPB, LPG3 and Amastin in stationary phase
promastigotes and amastigotes between pathogenic
Leishmania spp. and L. tarentolae, the lizard species
non-pathogenic to humans. Our data confirmed that
these virulence genes were expressed and the
proteins produced in L. tarentolae. At this level, we
cannot exclude that differences in protein sequence
or species-specific differential regulation, mainly at
the post-translational level of these virulence factors,
might alter the interaction of these proteins with
other cellular partners, which could result in changes
in the parasite’s virulence. It is also possible that
differences in the thermophysiology between lizard
and mammalian Leishmania species could account
for the loss of virulence of L. tarentolae in mammals.
It is not known what genes are involved in this pro-
cess but it has been reported that some dehydro-
genases and aldolases showed differential expression
between lizard- and human-infecting Leishmania
species (Janovy et al. 1972, Ghosh et al. 1976). This
interesting aspect requires further investigation,
which is outside of the scope of this study. It is also
likely that other, yet uncharacterized virulence fac-
tors, absent or differentially regulated in the lizard
L. tarentolae could account for the loss of patho-
genicity in this species. Whole-genome comparative
analyses between pathogenic and non-pathogenic
Leishmania species will shed more light in that
direction.
The financial support by the Iran Research Council of theRepublic Presidentship is gratefully acknowledged. Inaddition, this project was supported by Pasteur Institute ofIran grant number 282. We wish to thank Mr Alizadeh fortechnical assistance.
REFERENCES
Akopyants, N. S., Matlib, R. S., Bukanova, E. N.,
Smeds, M. R., Brownstein, B. H., Stormo, G. D.
and Beverley, S. M. (2004). Expression profiling
using random genomic DNA microarrays identifies
differentially expressed genes associated with three
major developmental stages of the protozoan parasite
Leishmania major. Molecular and Biochemical
Parasitology 136, 71–86.
Alexander, J., Coombs, G. H. and Mottram, J. C.
(1998). Leishmania mexicana cysteine proteinase-
deficient mutants have attenuated virulence for mice
and potentiate a Th1 response. Journal of Immunology
161, 6794–6801.
Bordier, C. (1987). The promastigote surface protease
of Leishmania. Parsitology Today 3, 151–153.
Breton, M., Tremblay, M. J., Ouellette, M. and
Papadopoulou, B. (2005). Live nonpathogenic
parasitic vector as a candidate vaccine against visceral
leishmaniasis. Infection and Immunity 73, 6372–6382.
Buxbaum, L. U., Denise, H., Coombs, G. H.,
Alexander, J., Mottram, J. C. and Scott, P. (2003).
Cysteine protease B of Leishmania mexicana inhibits
host Th1 responses and protective immunity. Journal
of Immunology 171, 3711–3717.
Cameron, P., McGachy, A., Anderson, M., Paul, A.,
Coombs, G. H., Mottram, J. C., Alexander, J. and
Plevin, R. (2004). Inhibition of lipopolysaccharide-
induced macrophage IL-12 production by Leishmania
mexicana amastigotes: the role of cysteine peptidases
and the NF-kappaB signaling pathway. Journal of
Immunology 173, 3297–3304.
Campbell, D., Kurath, U. and Fleischmann, J.
(1992). Identification of a gp63 surface glycoprotein
in Leishmania tarentolae. FEMS Microbiology Letters
75, 89–92.
Chen, D. Q., Kolli, B. K., Yadava, N., Lu, H. G.,
Gilman-Sachs, A., Peterson, D. A. and Chang, K-P.
(2000). Episomal expression of specific sense and
antisense mRNAs in Leishmania amazonensis :
modulation of gp63 level in promastigotes and their
infection ofmacrophages in vitro. Infection and Immunity
68, 80–86.
Coelho, E. A., Tavares, C. A., Carvalho, F. A., Chaves,
K. F., Teixeira, K. N., Rodrigues, R. C., Charest, H.,
Matlashewski, G., Gazzinelli, R. T. andFernandes,
A. P. (2003). Immune responses induced by the
Leishmania (Leishmania) donovaniA2 antigen, but not by
the LACK antigen, are protective against experimental
Leishmania (Leishmania) amazonensis infection. Infection
and Immunity 71, 3988–3994.
Croan, D. G., Morrison, D. A. and Ellis, J. T. (1997).
Evolution of the genus Leishmania revealed by
comparison of DNA and RNA polymerase gene
sequences. Molecular and Biochemical Parasitology 89,
149–159.
Descoteaux, A., Avila, H. A., Zhang, K., Turco, S. J.
and Beverley, S. M. (2002). Leishmania LPG3
encodes a GRP94 homolog required for phosphoglycan
synthesis implicated in parasite virulence but not
viability. The EMBO Journal 21, 4458–4469.
Desjeux, P. (2004). Leishmaniasis : current situation
and new perspectives. Comparative Immunology,
Microbiology and Infectious Diseases 27, 305–318.
Dietzsch, J., Gehlenborg, N. and Nieselt, K. (2006).
Mayday – a Microarray Data Analysis Workbench.
Bioinformatics 22, 1010–1012.
Dollahon, N. R. and Janovy, J. (1973). Leismania adleri :
in vitro phagocytosis by lizard leukocytes and peritoneal
cells. Experimental Parasitology 34, 56–61.
H. Azizi and others 732

Frame, M. J., Mottram, J. C. and Coombs, G. H.
(2000). Analysis of the roles of cysteine proteinases of
Leishmania mexicana in the host-parasite interaction.
Parasitology 121, 367–377.
Fukao, T., Matsuda, S. and Koyasu, S. (2000).
Synergistic effects of IL-4 and IL-18 on IL-12-
dependent IFN-gamma production by dendritic cells.
Journal of Immunology 164, 64–71.
Garin, Y. J., Meneceur, P., Pratlong, F., Dedet, J. P.,
Derouin, F. and Lorenzo, F. (2005). A2 gene of Old
World cutaneous Leishmania is a single highly conserved
functional gene. BMC Infectious Disease 28, 18.
Genest, P. A., Haimeur, A., Legare, D., Sereno, D.,
Roy, G., Messier, N., Papadopoulou, B. and
Ouellette, M. (2008). A protein of the leucine-rich
repeats (LRRs) superfamily is implicated in antimony
resistance in Leishmania infantum amastigotes.
Molecular and Biochemical Parasitology 158, 95–99.
Ghedin, E., Charest, H. and Matlashewski, G. (1998).
A2rel : a constitutively expressed Leishmania gene linked
to an amastigote-stage-specific gene. Molecular and
Biochemical Parasitology 93, 23–29.
Ghosh, D. K. and Honigberg, B. M. (1976). Activities
of glucose-6-phosphate, 6-phosphogluconate, and
isocitrate dehydrogenases from Leishmania donovani
cultivated at 25 and 37 C. Journal of Parasitology 23,
450–455.
Gregory, D. J., Godbout, M., Contreras, I., Forget, G.
and Olivier, M. (2008). A novel form of NF-kB is
induced by Leishmania infection: involvement in
macrophage gene expression. European Journal of
Immunology 38, 1071–1081.
Hide,M., Bras-Goncalves, R. andBanuls, A.-L. (2007).
Specific cpb copies within the Leishmania donovani
complex: evolutionary interpretations and potential
clinical implications in humans. Parasitology 134,
379–389.
Ilg, T. (2000). Lipophosphoglycan is not required for
infection of macrophages or mice by Leishmania
mexicana. The EMBO Journal 19, 1953–1962.
Ivens, A. C., Peacock, C. S., Worthey, E. A., Murphy,
L., Aggarwal, G., Berriman, M., Sisk, E.,
Rajandream,M. A., Adlem, E., Aert, R., Anupama,
A., Apostolou, Z., Attipoe, P., Bason, N., Bauser, C.,
Beck, A., Beverley, S. M., Bianchettin, G., Borzym,
K., Bothe, G., Bruschi, C. V., Collins, M., Cadag, E.,
Ciarloni, L., Clayton, C., Coulson, R. M., Cronin, A.,
Cruz, A. K., Davies, R. M., De Gaudenzi, J.,
Dobson, D. E., Duesterhoeft, A., Fazelina, G.,
Fosker, N., Frasch, A. C., Fraser, A., Fuchs, M.,
Gabel, C., Goble, A., Goffeau, A., Harris, D.,
Hertz-Fowler, C., Hilbert, H., Horn, D., Huang, Y.,
Klages, S., Knights, A., Kube, M., Larke, N., Litvin,
L., Lord, A., Louie, T., Marra, M., Masuy, D.,
Matthews, K., Michaeli, S., Mottram, J. C., Muller-
Auer, S., Munden, H., Nelson, S., Norbertczak, H.,
Oliver, K., O’Neil, S., Pentony, M., Pohl, T. M.,
Price, C., Purnelle, B., Quail, M. A.,
Rabbinowitsch, E., Reinhardt, R., Rieger, M.,
Rinta, J., Robben, J., Robertson, L., Ruiz, J. C.,
Rutter, S., Saunders, D., Schafer, M., Schein, J.,
Schwartz, D. C., Seeger, K., Seyler, A., Sharp, S.,
Shin, H., Sivam, D., Squares, R., Squares, S.,
Tosato, V., Vogt, C., Volckaert, G., Wambutt, R.,
Warren, T., Wedler, H., Woodward, J., Zhou, S.,
Zimmermann, W., Smith, D. F., Blackwell, J. M.,
Stuart, K. D., Barrell, B. and Myler, P. J. (2005).
The genome of the kinetoplastid parasite, Leishmania
major. Science 309, 436–442.
Janovy, J. (1972). Temperature and metabolism in
Leishmania. 3. Some dehydrogenases of L. donovani,
L. mexicana, and L. tarentolae. Experimental
Parasitology 32, 196–205.
Joshi, B. P., Kelly, B. L., Kamhawi, H., Sacks, D. L.
and McMaster, W. R. (2002). Targeted gene deletion
in Leishmania major identifies leishmanolysin (GP63)
as a virulence factor. Molecular and Biochemical
Parasitology 120, 33–40.
Krogh, A., Larsson, B., von Heijne, G. and
Sonnhammer, E. L. (2001). Predicting
transmembrane protein topology with a hidden
Markov model: application to complete genomes.
Journal of Molecular Biology 19, 567–580.
Landweber, L. F. and Gilbert, W. (1994). Phylogenetic
analysis of RNA editing: a primitive genetic
phenomenon. Proceedings of the National Academy of
Sciences, USA 91, 918–921.
Larreta, R., Soto, M., Alonso, C. and Requena, J. M.
(2000). Leishmania infantum : gene cloning of the GRP94
homologue, its expression as recombinant protein and
analysis of antigenicity. Experimental Parasitology 96,
108–115.
Leifso, K., Cohen-Freue, G., Dogra, N., Murray, A.
andMcMaster, W. R. (2007). Genomic and proteomic
expression analysis of Leishmania promastigote and
amastigote life stages. The Leishmania genome is
constitutively expressed. Molecular and Biochemical
Parasitology 152, 35–46.
Maslov, D. A., Avila, H. A., Lake, J. A. and Simpson,
L. (1994). Evolution of RNA editing in kinetoplastid
protozoa. Nature, London 368, 345–348.
McGwire, B. S., O’Connell, W. A., Chang, K. P. and
Engman, D. M. (2002). Extracellular release of the
glycosylphosphatidylinositol (GPI)-linked Leishmania
surface metalloprotease, gp63, is independent of GPI
phospholipolysis : implications for parasite virulence.
Journal of Biological Chemistry 277, 8802–8809.
McMaster, W. R., Morrison, C. J., MacDonald, M. H.
and Joshi, P. B. (1994). Mutational and functional
analysis of the Leishmania surface metalloproteinase
GP63: similarities to matrix metalloproteinases.
Parasitology 108, S29–S36.
McNicoll, F., Drummelsmith, J., Muller, M.,
Madore, E., Boilard, N., Ouellette, M. and
Papadopoulou, B. (2006). A combined proteomic and
transcriptomic approach to the study of stage
differentiation in Leishmania infantum. Proteomics 6,
3567–3581.
McNicoll, F., Muller, M., Cloutier, S., Boilard, N.,
Rochette, A., Dube, M. and Papadopoulou, B.
(2005). Distinct 3k-untranslated region elements regulate
stage-specific mRNA accumulation and translation in
Leishmania. Journal of Biological Chemistry 280,
35238–35246.
Medina-Acosta, E., Beverley, S. M. and Russell, D. G.
(1993). Evolution and expression of the Leishmania
surface proteinase (gp63) gene locus. Infectious Agents
and Diseases 2, 25–34
Virulence factors in Leishmania tarentolae 733

Mottram, J. C., Brooks, D. R. and Coombs, G. H.
(1998). Roles of cysteine proteinases of trypanosomes
and Leishmania in host–parasite interactions. Current
Opinion in Microbiology 1, 455–460.
Mottram, J. C., Coombs, G. H. and Alexander, J.
(2004). Cysteine peptidases as virulence factors
of Leishmania. Current Opinion in Microbiology 7,
375–381.
Mundodi, V., Somanna, A., Farrell, P. J. and
Gedamu, L. (2002). Genomic organization and
functional expression of differentially regulated
cysteine protease genes of Leishmania donovani
complex. Gene 282, 257–265.
Murray, H. W., Berman, J. D., Davies, C. R. and
Saravia, N. G. (2005). Advances in leishmaniasis.
Lancet 366, 1561–1577.
Nakhaee, A., Taheri, T., Taghikhani, M., Mohebali,
M., Salmanian, A. H., Fasel, N. and Rafati, S.
(2004). Humoral and cellular immune responses against
type I cysteine proteinase of Leishmania infantum are
higher in asymptomatic than symptomatic dogs selected
from a naturally infected population. Veterinary
Parasitology 119, 107–123.
Noyes, H. A., Chance, M. L., Croan, D. G. and Ellis,
J. T. (1998). Leishmania (Sauroleishmania) : a comment
on classification. Parasitology Today 14, 167.
Orlando, T. C., Rubio, M. A., Sturm, N. R., Campbell,
D. A. and Floeter-Winter, L. M. (2002). Intergenic
and external transcribed spacers of ribosomal RNA
genes in lizard-infecting Leishmania : molecular
structure and phylogenetic relationship to mammal-
infecting Leishmania in the subgenus Leishmania
(Leishmania). Memorias do Instituto Oswaldo Cruz 97,
695–701.
Ouakad, M., Bahi-Jaber, N., Chenik, M., Dellagi, K.
and Louzir, H. (2007a). Selection of endogenous
reference genes for gene expression analysis in
Leishmania major developmental stages. Parasitology
Research 101, 473–477.
Ouakad, M., Chenik, M., Achour-Chenik, Y. B.,
Louzir, H. and Dellagi, K. (2007b). Gene expression
analysis of wild Leishmania major isolates : identification
of genes preferentially expressed in amastigotes.
Parasitology Research 100, 255–264.
Paape, D., Lippuner, C., Schmid, M., Ackermann, R.,
Barrios-Llerena, M. E., Zimny-Arndt, U.,
Brinkmann, V., Arndt, B., Pleissner, K. P.,
Jungblut, P. R. and Aebischer, T. (2008). Transgenic,
fluorescent Leishmania mexicana allow direct analysis of
the proteome of intracellular amastigotes.Molecular and
Cellular Proteomics 7, 1688–1701.
Peacock, C. S., Seeger, K., Harris, D., Murphy, L.,
Ruiz, J. C., Quail, M. A., Peters, N., Adlem, E.,
Tivey, A., Aslett, M., Kerhornou, A., Ivens, A.,
Fraser, A., Rajandream, M. A., Carver, T.,
Norbertczak, H., Chillingworth, T., Hance, Z.,
Jagels, K., Moule, S., Ormond, D., Rutter, S.,
Squares, R., Whitehead, S., Rabbinowitsch, E.,
Arrowsmith, C., White, B., Thurston, S., Bringaud,
F., Baldauf, S. L., Faulconbridge, A., Jeffares, D.,
Depledge, D. P., Oyola, S. O., Hilley, J. D., Brito,
L. O., Tosi, L. R., Barrell, B., Cruz, A. K., Mottram,
J. C., Smith, D. F. and Berriman, M. (2007).
Comparative genomic analysis of three Leishmania
species that cause diverse human disease. Nature
Genetics 39, 839–847.
Previato, J. O., Jones, C., Wait, R., Routier, F.,
Saraiva, E. and Mendonca-Previato, L. (1997).
Leishmania adleri, a lizard parasite, expresses
structurally similar glycoinositolphospholipids to
mammalian Leishmania. Glycobiology 7, 687–695.
Rafati, S., Hassani, N., Taslimi, Y., Movassagh, H.,
Rochette, A. and Papadopoulou, B. (2006). Amastin
peptide-binding antibodies as biomarkers of active
human visceral leishmaniasis. Clinical Vaccine
Immunology 13, 1104–1110.
Rafati, S., Nakhaee, A., Taheri, T., Ghashghaii, A.,
Salmanian, A. H., Jimenez, M., Mohebali, M.,
Masina, S. and Fasel, N. (2003). Expression of
cysteine proteinase type I and II of Leishmania
infantum and their recognition by sera during canine
and human visceral leishmaniasis. Experimental
Parasitology 103, 143–151.
Rafati, S., Nakhaee, A., Taheri, T., Taslimi, Y.,
Darabi, H., Eravani, D., Sanos, S., Kaye, P.,
Taghikhani, M., Jamshidi, S. and Rad, M. A.
(2005). Protective vaccination against experimental
canine visceral leishmaniasis using a combination
of DNA and protein immunization with cysteine
proteinases type I and II of L. infantum. Vaccine 23,
3716–3725.
Rafati, S., Salmanian, A. H., Hashemi, K., Schaff, C.,
Belli, S. and Fasel, N. (2001). Identification of
Leishmania major cysteine proteinases as targets of the
immune response in humans.Molecular and Biochemical
Parasitology 113, 35–43.
Requena, J. M., Alonso, C. and Soto, M. (2000).
Evolutionarily conserved proteins as prominent
immunogens during Leishmania infections. Parasitology
Today 16, 246–250.
Rochette, A., McNicoll, F., Girard, J., Breton, M.,
Leblanc, E., Bergeron, M. G. and Papadopoulou, B.
(2005). Characterization and developmental gene
regulation of a large gene family encoding amastin
surface proteins in Leishmania spp. Molecular and
Biochemical Parasitology 140, 205–220.
Rochette, A., Raymond, F., Ubeda, J. M., Smith, M.,
Messier, N., Boisvert, S., Rigault, P., Corbeil, J.,
Ouellette, M. and Papadopoulou, B. (2008).
Genome-wide gene expression profiling analysis of
Leishmania major and Leishmania infantum
developmental stages reveals substantial differences
between the two species. BMC Genomics 9, 255–281.
Rosenthal, P. J. (1999). Proteases of protozoan parasites.
Advances in Parasitology 43, 105–159.
Rosenzweig, D., Smith, D., Opperdoes, F., Stern, S.,
Olafson, R. W. and Zilberstein, D. (2008). Retooling
Leishmania metabolism: from sand fly gut to human
macrophage. The FASEB Journal 22, 590–602.
Salotra, P., Duncan, R. C., Singh, R., Subba Raju,
B. V., Sreenivas, G. and Nakhasi, H. L. (2006).
Upregulation of surface proteins in Leishmania
donovani isolated from patients of post kala-azar dermal
leishmaniasis. Microbes and Infection 8, 637–644.
Sambrook, J. and Russell, D. (2001). Molecular
Cloning: a Laboratory Manual, 3rd Edn. Cold Spring
Harbor Laboratory Press, Cold Spring Harbor, NY,
USA.
H. Azizi and others 734

Spath, G. F., Epstein, L., Leader, B., Singer, S. M.,
Avila, H. A., Turco, S. J. and Beverley, S. M. (2000).
Lipophosphoglycan is a virulence factor distinct from
related glycoconjugates in the protozoan parasite
Leishmania major. Proceedings of the National Academy
of Sciences, USA 97, 9258–9263.
Spath, G. F., Garraway, L. A., Turco, S. J. and
Beverley, S. M. (2003). The role(s) of
lipophosphoglycan (LPG) in the establishment of
Leishmania major infections in mammalian hosts.
Proceedings of the National Academy of Sciences, USA
100, 9536–9541.
Stiles, J. K., Hicock, P. I., Shah, P. H. and Mead, J. C.
(1999). Genomic organization, transcription, splicing
and gene regulation in Leishmania. Annals of Tropical
Medicine and Parasitology 93, 781–807.
Williams, R. A., Tetley, L., Mottram, J. C. and
Coombs, G. H. (2006).Cysteine peptidases CPA and
CPB are vital for autophagy and differentiation in
Leishmania mexicana. Molecular Microbiology 61,
655–674.
Wu, Y., El-Fakhry, Y., Sereno, D., Tamar, S. and
Papadopoulou, B. (2000). A new developmentally
regulated gene family in Leishmania amastigotes
encoding a homolog of amastin surface proteins.
Molecular and Biochemical Parasitology 110, 345–357.
Yao, C., Donelson, J. E. and Wilson, M. E. (2003).
The major surface protease (MSP or GP63) of
Leishmania sp. Biosynthesis, regulation of expression,
and function. Molecular and Biochemical Parasitology
132, 1–16.
Zadeh-Vakili, A., Taheri, T., Taslimi, Y., Doustdari,
F., Salmanian, A. H. and Rafati, S. (2004).
Immunization with the hybrid protein vaccine,
consisting of Leishmania major cysteine proteinases
Type I (CPB) and Type II (CPA), partially protects
against leishmaniasis. Vaccine 22, 1930–1940.
Zhang, W. W. and Matlashewski, G. (2001).
Characterization of the A2–A2rel gene cluster in
Leishmania donovani : involvement of A2 in
visceralization during infection. Molecular Microbiology
39, 935–948.
Zhang, W. W. and Matlashewski, G. (2004). In vivo
selection for Leishmania donovani miniexon genes that
increase virulence in Leishmania major. Molecular
Microbiology 54, 1051–1062.
Zhang, W. W. and Matlashewski, G. (1997). Loss of
virulence in Leishmania donovani deficient in an
amastigote-specific protein, A2. Proceedings of the
National Academy of Sciences, USA 94, 8807–8811.
Zhang, W. W., Mendez, S., Ghosh, A., Myler, P.,
Ivens, A., Clos, J., Sacks, D. L. and Matlashewski,
G. (2003). Comparison of the A2 gene locus in
Leishmania donovani and Leishmania major and its
control over cutaneous infection. Journal of Biological
Chemistry 278, 35508–35515.
Virulence factors in Leishmania tarentolae 735
Related Documents











