Journal of Virological Methods, 42 (1993) 1-12 1 © 1993 Elsevier Science Publishers B.V. / All rights reserved / 0166-0934/93/$06.00 VIRMET 01451 Screening for inhibitors of HIV gpl20-CD4 binding using an enzyme-linked immunoabsorbant assay • a* • • b* Mark Gilbert ' , Luis Brlgldo ' , W.E.G. Mfiller c, J-E. Hansen d, R.A. Ezekowitz e and John Mills f'g aDepartment of Hematology-Oncology, University of Washington, Seattle, WA (USA), bDivision of Infectious Diseases, University of California, San Francisco and San Francisco General Hospital, San Francisco, CA (USA), Clnstitute for Physiological Chemistr.~, University of Mainz, Mainz (Germany), dLaboratory for Infectious Diseases, Hvidovre Hospital, Copenhagen (Denmark), eDepartment of Pediatrics, Harvard Medical School and Children's Hospital, Boston, MA (USA), f Department of Medicine, University of California, San Francisco, CA (USA) and gMacfarlane Burnet Centre for Medical Research, Fairfield, Victoria (Australia) (Accepted 21 September 1992) Summary Binding of the HIV-1 major viral surface glycoprotein, gpl20, to the major cell receptor, CD4, is essential for HIV infection of the target cell and syncytium formation. An enzyme-linked immunoassay using solid phase CD4 was used to quantitate the binding of HIV-1 gpl20 to CD4, and to assess the activity and mechanism of action of putative inhibitors of that reaction. Monoclonal antibodies to the gpl20 binding site on CD4 (e.g., Leu3a) blocked gpl20 binding, while monoclonal antibodies to other portions of CD4 (e.g. OKT4) did not. Both aurintricarboxylic acid and sulfonated potysaccharides (e.g., dextran sulfate) blocked CD4-gpl20 interactions by binding to the CD4 component• Human polyclonal antibodies to gpl20 also blocked gpl20-CD4 binding, but none of the monoclonal antibodies tested (including several with neutralizing activity) were effective. In contrast, several lectins (including mannose binding protein) bound to gpl20 and blocked CD4-gpl20 interactions. Enzymatic deglycosylation of gpl20 only minimally affected its CD4 binding capacity, while non-glycosylated gpl20 (produced in Escherichia coli)-bound CD4 about 10-fold less well than fully-glycosylated material. The results demonstrate that this assay system can be used to measure the activity of inhibitors of CD4-gp 120 binding, and to determine the mechanism of action of those inhibitors. Correspondence to: J. Mills, Macfarlane Burnet Centre, Yarra Bend Road, Fairfield, Vic., Australia 3078. *Who contributed equally to this work.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Virological Methods, 42 (1993) 1-12 1 © 1993 Elsevier Science Publishers B.V. / All rights reserved / 0166-0934/93/$06.00
VIRMET 01451
Screening for inhibitors of HIV gpl20-CD4 binding using an enzyme-linked immunoabsorbant assay
• a * • • b * Mark Gilbert ' , Luis Brlgldo ' , W.E.G. Mfiller c, J-E. Hansen d, R.A. Ezekowitz e and John Mills f'g
aDepartment of Hematology-Oncology, University of Washington, Seattle, WA (USA), bDivision of Infectious Diseases, University of California, San Francisco and San Francisco General Hospital, San Francisco, CA (USA), Clnstitute for Physiological Chemistr.~, University of Mainz,
Mainz (Germany), dLaboratory for Infectious Diseases, Hvidovre Hospital, Copenhagen (Denmark), eDepartment of Pediatrics, Harvard Medical School and Children's Hospital, Boston, MA (USA), f Department of Medicine, University of California, San Francisco, CA (USA) and gMacfarlane
Burnet Centre for Medical Research, Fairfield, Victoria (Australia)
(Accepted 21 September 1992)
Summary
Binding of the HIV-1 major viral surface glycoprotein, gpl20, to the major cell receptor, CD4, is essential for HIV infection of the target cell and syncytium formation. An enzyme-linked immunoassay using solid phase CD4 was used to quantitate the binding of HIV-1 gpl20 to CD4, and to assess the activity and mechanism of action of putative inhibitors of that reaction. Monoclonal antibodies to the gpl20 binding site on CD4 (e.g., Leu3a) blocked gpl20 binding, while monoclonal antibodies to other portions of CD4 (e.g. OKT4) did not. Both aurintricarboxylic acid and sulfonated potysaccharides (e.g., dextran sulfate) blocked CD4-gpl20 interactions by binding to the CD4 component• Human polyclonal antibodies to gpl20 also blocked gpl20-CD4 binding, but none of the monoclonal antibodies tested (including several with neutralizing activity) were effective. In contrast, several lectins (including mannose binding protein) bound to gpl20 and blocked CD4-gpl20 interactions. Enzymatic deglycosylation of gpl20 only minimally affected its CD4 binding capacity, while non-glycosylated gpl20 (produced in Escherichia coli)-bound CD4 about 10-fold less well than fully-glycosylated material. The results demonstrate that this assay system can be used to measure the activity of inhibitors of CD4-gp 120 binding, and to determine the mechanism of action of those inhibitors.
Correspondence to: J. Mills, Macfarlane Burnet Centre, Yarra Bend Road, Fairfield, Vic., Australia 3078. *Who contributed equally to this work.

HIV; p120; CD4; Lectin
Introduction
Human immunodeficiency virus (HIV) infection is initiated when the viral envelope glycoprotein 120 (gpl20) binds to CD4 molecules on the surface of target cells (Klatzmann et al., 1984; Dalgleish et al., 1984; Layne et al., 1990). Viral gp 120 is synthesised as a 160 kDa polyprotein which is hydrolysed to the transmembrane gp41 and extracellular gpl20; the latter is retained on the surface of the cell by non-covalent bonds (McKeating et al., 1989). Approximately 50% of gpl20 is carbohydrate, with the predominantly N- acetyl galactosamine and mannose saccharides being distributed throughout the length of the molecule (Kowalski et al., 1987; Robinson et al., 1987). Specific domains of the gpl20 and CD4 molecules have been identified which are required for binding (Arthos et al., 1989; Ashkenazi et al., 1990; Brodsky et al., 1990; Cordonnier et al., 1989; Lasky et al., 1987; Olshevsky et al., 1990). In addition to being required for initiation of infection, gpl20-CD4 binding is essential for syncytium formation (Arthos et al., 1989; Crowe et al., 1990; Lifson et al., 1986), partially determines viral tropism (Robinson et al., 1990), and may mediate immunosuppression (Diamond et al., 1988; Mann et al., 1987).
Because the gpl20-CD4 interaction is critical to initiation of infection with HIV, substances which block gpl20-CD4 binding have been shown to neutralise virus infectivity and to prevent fusion between virus-infected cells (expressing surface gpl20) and uninfected CD4-bearing lymphoid cells (Lifson et al., 1986; Crowe et al., 1990). For this reason, development of antiretrovirals which block the gpl20-CD4 interaction has had a high priority. Numerous compounds have been discovered which block viral infectivity or inhibit virus- CD4 cell fusion, including sulfated polysaccharides such as sulphovernan and dextran sulfate (Parish et al., 1990; Weiler et al., 1990), lectins (Miiller et al., 1988; Ezekowitz et al., 1989), antibodies to gpl20, (Linsley et al., 1988; Schnittman et al., 1988; Hansen et al., 1990), aurintricarboxylic acid (Schols et al., 1989), and recombinant soluble CD4 (Byrn et al., 1989; Habeshaw et al., 1989). In some instances, these materials have been shown to inhibit CD4- gpl20 binding directly (Schols et al., 1989; Lederman et al., 1989), although in a few cases, compounds that had high neutralising or anti-syncytium activity had little effect on gpl20-CD4 binding (Sun et al., 1989; Parish et al., 1990).
Recently, we developed a solid-phase enzyme immunoassay for HIV gpl20 using recombinant, soluble CD4 (rsCD4) as the capture system (Gilbert et al., 1991). This assay provided high sensitivity and specificity (Gilbert et al., 1991) for detection of a fully glycosylated recombinant gpl20 (rpgl20) (Gilbert, 1991)~ We used this assay to screen compounds for their ability to inhibit gpl20-CD4 binding. The results emphasise that substances which neutralise

3
virus infectivity or block syncytium formation may not inhibit gpl20-CD4 binding.
Materials and Methods
Recombinant HIV envelope glycoprotein 120 (rgpl20) produced in insect cells with a baculovirus vector was a generous gift of Dr. J. Culp, SmithKline Beecham Pharmaceuticals, King of Prussia, PA (Ivey-Hoyle et al., 1991). Dr. Culp also provided rgpl20 deglycosylated enzymatically with endoglycosylases F and H, which had a molecular weight (by denaturing gel electrophoresis) of about 60000 kDa. Non-glycosylated gpl20 prepared in yeast was obtained from the Reference Reagents Branch, DAIDS, NIAID, NIH, Bethesda, MD (Catalogue No. 388). Native gpl20 was obtained from HIV (strain SF33) prepared by infection of an immortalised CD4-negative neuroblastoma cell line (cell line no. 192) and harvested in Eagle's Minimal Essential Medium with 10% foetal calf serum (both the virus and the cell line were graciously provided by Dr. Jay A. Levy). This virus/cell line system was employed because it yields large amounts of free gpl20 (Gilbert et al., 1990). Recombinant, soluble CD4 was generously provided by Drs. Ray Sweet, Christine Debouck and Martin R0senberg, also of SmithKline Beecham (Deen et al., 1988).
The mannose-affinity lectins concanavalin A (Canavalia ensiformis), and lentil (Lens culinaris), fava (Viciafaba), and sweet pea (Lathyrus odoratus) were obtained from Sigma Chemical Co., St. Louis, MO. Recombinant mannose- binding protein was prepared as described (Ezekowitz et al., 1989). Low molecular weight dextran sulfate (Mol wt. 5000) and aurintricarboxylic acid (Schols et al., 1989) were obtained from Aldrich Biochemicals, Milwaukee, WI. Monoclonal antibodies to CD4 (Leu3a, OKT4) were obtained from Becton- Dickinson, Mountain View, CA and Ortho Pharmaceuticals, Raritan, N J, respectively. Thirteen anticarbohydrate monoclonal antibodies, some of which had HIV neutralising and anti-syncytium activities (Hansen et al., 1990) were also tested, as was a tetrasaccharide (Biocarb 45/05). Rabbit antisera to synthetic gpl20 peptides that neutralised HIV infectivity and blocked gpl20- CD4 binding in some systems were kindly provided by Dr. Blair Ardman (Ardman et al., 1990), and a murine monoclonal antibody (15e) which had been reported to have both HIV neutralising and gpl20-CD4 binding inhibitory activity was provided by Dr. David Ho (Ho et al., 1991).
The solid-phase CD4 enzyme immunoassay has been previously described (Gilbert et al., 1991). Briefly, recombinant soluble CD4 immobilised in polystyrene microtitre trays is reacted with known amounts of gpl20 (either recombinant or native viral) with or without the inhibitor under study. Bound gpl20 is quantitated by successive binding of sheep anti-gpl20, biotinylated rabbit anti-sheep IgG, and avidin-alkaline phosphatase (Gilbert et al., 1991). Various concentrations of each agent were incubated with 10 ng/ml recombinant gpl20 or 1000 pfu/ml HIVsF33 for one hour prior to application

of 100 #1 of the mixture to CD4-coated microtiter wells. This amount of gpl20 was selected because it just reached the optical density plateau (Gilbert et al., 1991). Materials applied to the CD4-coated wells prior to gpl20 or HIVsF33 were also incubated for 1 h following a 45-min blocking step previously described (Gilbert et al., 1991). Appropriate controls (gpl20 without inhibitor; inhibitor without gp 120) were included in each experiment.
Results
Validation o f the assay with known inhibitors
We validated this assay as a screen for inhibitors of gpl20-CD4 binding by first studying compounds with known activity and mechanism of action (Table 1). Antibodies to CD4 that bind near at gpl20 binding site (e.g. Leu3a) inhibit stoichiometrically (Fig. 1), while those that bind elsewhere on the molecule (eg. OKT4) did not (Fig. 1). As expected, Leu3a was active when pre-incubated with the solid-phase rsCD4 and binding of Leu3a to rsCD4 that had been pre- loaded with gpl20 was reduced by over 70% while binding of OKT4 was reduced by only 10% (data not shown). Leu3a also blocked binding of native gpl20 (Fig. 1). Aurintricarboxylic acid (ATA) has been shown to bind to CD4, in turn blocking adherence of gpl20 (Schols et al., 1989). In our assay, ATA inhibition of gpl20 binding to rsCD4 exhibited a clear dose-response relationship and about 1.5 ttg of ATA was required to reduce binding of 12 ng of gpl20 to rsCD4 by 50% (Table 1). Preincubation of the solid phase rsCD4 with ATA also blocked gpl20 binding (data not shown).
Recombinant, soluble CD4 inhibited the binding of recombinant gpl20 to CD4 in this assay system in the expected dose-dependent fashion. It required 30
i i i i i i
IO0 ~ ~ x ~ V OKT4
,-'~8~ =0 o ,~ 0 6O
40 LEU3A Ld 0
~ 2 0
0 I L L I
10-2 10 -I 100 101 102 103 104 105 [mob], ng/ml
Fig. 1. Inhibition of gpl20 binding by monoclonal antibodies Leu3a and OKT4.

ng rsCD4 (mol wt. 270 000) to inhibit binding of 12 ng of recombinant gpl20 (mol wt. 120000) by 50% (Table 1), a molar ratio very close to the predicted value of 1:1. Equivalent concentrations of rsCD4 also blocked the binding of native gpl20 from HIVsF33 (Table 1).
Inhibition of binding by antibodies to gpl20
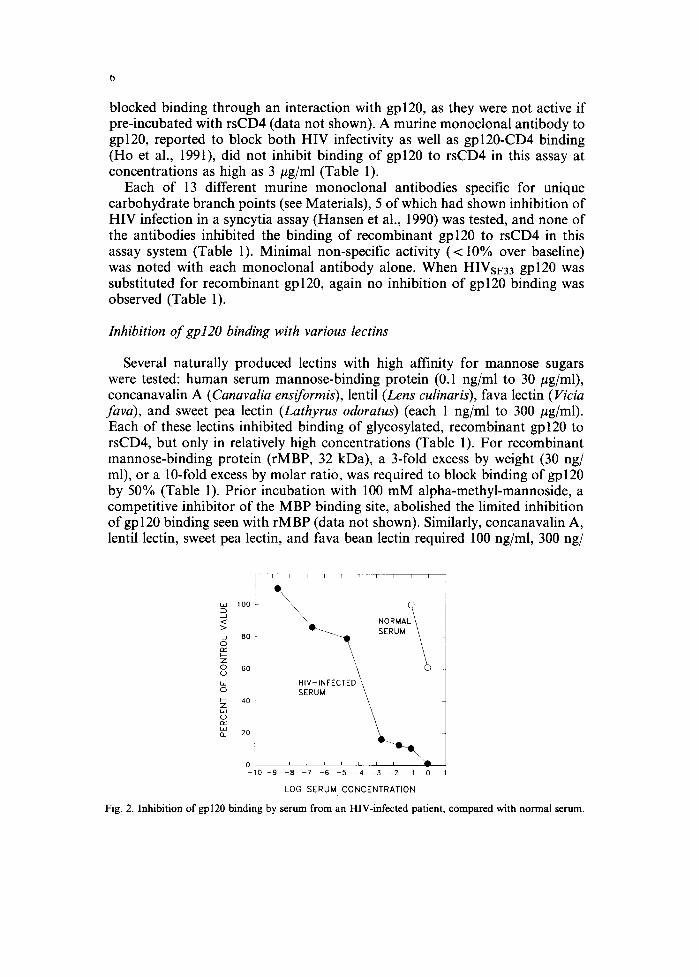
Rabbit antisera raised to peptides synthesised to correspond to the CD4- binding regions of gpl20 (Ardman et al., 1990) failed to inhibit binding of gpl20 to rsCD4 (Table 1). In contrast, each of 20 sera from patients with HIV infection, previously screened by whole-virus ELISA and Western blotting and known to have antibodies to gpl20, inhibited binding of gpl20 to rsCD4 (Table 1 and Fig. 2). When representative HIV-reactive sera were titered in this assay, and compared with sera from healthy subjects that were non-reactive by ELISA or Western blotting, the HIV-reactive sera inhibited binding at concentrations of < 1 0 - 6 , while normal sera inhibited only when used at concentrations of 10-l or higher (Fig. 2). Both normal and HIV-reactive sera
TABLE 1
Relative activity of putative inhibitors of HIV gp 120/CD4 binding
Inhibitor 50% Inhib. concentration (or dilut.) against: gp 120 HIVSF33gp 120
Antibodies to CD4 (Fig. 1) Leu3a OKT4
rsCD4
Aurintricarboxylic acid
Lectins mannose binding protein concanavalin A lentil lectin sweet pea lectin fava bean lectin
45 ng/ml 65 ng/ml > 1 /~g/ml > 1 pg/ml
30 ng/ml 40 ng/ml
1.5 #g/ml n.d.
30 ng/ml > 30/~g/ml 100 ng/ml > 300/~g/ml 300 ng/ml > 300 #g/ml 300 ng/ml > 300 #g/ml 500 ng/ml > 300 itg/ml
Antibodies to gpl20 Monoclonal 15e (35) > 3 #g/ml > 3 pg/ml Rabbit sera (34) > 1:5 > 1:5 Human sera (Fig. 2) 1:10 000 n.d. Oligosacch. mAbs (22) > 5 #g/ml > 5/~g/ml
Sulfated polysaccharides Dextran (MW 5000) (Fig 4) Polyanethol sulfonate Sulphovernan
630 ng/ml 380 ng/ml 2/~g/ml n.d.
150 ng/ml n.d.
See Methods section for description of compounds tested. n.d. Not done. rsCD4: recombinant, soluble CD4. rgpl20: recombinant gpl20.
blocked binding through an interaction with gpl20, as they were not active if pre-incubated with rsCD4 (data not shown). A murine monoclonal antibody to gpl20, reported to block both HIV infectivity as well as gpl20-CD4 binding (Ho et al., 1991), did not inhibit binding of gpl20 to rsCD4 in this assay at concentrations as high as 3 #g/ml (Table 1).
Each of 13 different murine monoclonal antibodies specific for unique carbohydrate branch points (see Materials), 5 of which had shown inhibition of HIV infection in a syncytia assay (Hansen et al., 1990) was tested, and none of the antibodies inhibited the binding of recombinant gpl20 to rsCD4 in this assay system (Table 1). Minimal non-specific activity (< 10% over baseline) was noted with each monoclonal antibody alone. When HIVsF33 gpl20 was substituted for recombinant gpl20, again no inhibition of gpl20 binding was observed (Table 1).
Inhibition of gpl20 binding with various lectins
Several naturally produced lectins with high affinity for mannose sugars were tested: human serum mannose-binding protein (0.1 ng/ml to 30 #g/ml), concanavalin A (Canavalia ensiformis), lentil (Lens culinaris), fava lectin (Vicia fava), and sweet pea lectin (Lathyrus odoratus) (each 1 ng/ml to 300 #g/ml). Each of these lectins inhibited binding of glycosylated, recombinant gpl20 to rsCD4, but only in relatively high concentrations (Table 1). For recombinant mannose-binding protein (rMBP, 32 kDa), a 3-fold excess by weight (30 ng/ ml), or a 10-fold excess by molar ratio, was required to block binding of gpl20 by 50% (Table 1). Prior incubation with 100 mM alpha-methyl-mannoside, a competitive inhibitor of the MBP binding site, abolished the limited inhibition of gp 120 binding seen with rMBP (data not shown). Similarly, concanavalin A, lentil lectin, sweet pea lectin, and fava bean lectin required 100 ng/ml, 300 ng/
i i i i r i i i i i
• ,¢ NORMA
o~Z ~ ~ > 8o 6O ~ SERUM ~ )
L) HIV-INFECTED \ ~ SE
- 1 0 - 9 - 8 - 7 - 6 - 5 - 4 - 3 - 2 -1 0
LOG SERUM CONCENTRATION Fig. 2. Inhibition of gpl20 binding by serum from an HIV-infected patient, compared with normal serum.

ml, 300 ng/ml and 500 ng/ml, respectively, to inhibit binding of 100 ng gpl20/ ml to rsCD4 (Table 1). Increasing concentrations of each produced similar inhibition curves and the relative 50% inhibition concentration corresponded to the relative mannose affinity for each lectin. Again, incubation of each lectin with 100 mM alpha methyl mannoside prior to incubation with gpl20 abrogated inhibition of gpl20 binding. Interestingly, high concentrations of alpha-methyl-mannoside alone did not affect recombinant gpl20 binding to rsCD4 (data not shown). A tetrasaccharide with viral neutralising activity (J.- E. Hansen, unpublished data) also failed to inhibit binding at concentrations of 100 #g/ml.
Since these lectins were relatively ineffective in inhibiting binding of recombinant gpl20 (which although fully glycosylated, has a slightly different oligosaccharide content than native gpl20), HIVsF33 was substituted as a source of native gpl20 with increasing concentrations of each lectin and analysed by EIA. At the highest concentrations tested, these lectins failed to inhibit binding of approximately 100 pfu of HIVsF33 (approximately 10 ng/ml available gp 120) (Table 1).
Effect of sulfated polysaccharides on gpl20-CD4 binding
Low molecular weight (5,000 mol wt.) dextran sulfate (0.005/~g/ml to 500 /~g/ml) (Schols et al., 1990; Balzarini et al., 1991), polyanethol sulfonate (Parish et al., 1990), and sulphovernan (Weiler et al., 1990) were tested for their ability to inhibit gpl20-CD4 binding. All these sulfated polysaccharides inhibited gpl20 binding to rsCD4 in a dose-dependent fashion; a representative experiment with dextran sulfate is shown (Fig. 3). Pretreatment of the solid- phase rsCD4 blocked gpl20 binding as effectively as concurrent incubation with the gpl20-CD4 mixture. In addition, pretreatment of rsCD4 with sulfated
l \ 8 0
\ l
§ 6o \\ ,\ \
c )
~- 20 \,,\ \ ;
o , , L ~ ,) 10 0 101 10 2 10 3 10 4 10 5
[DEXTRAN SULFATE], NG/ML
Fig. 3. Inhibition of gpl20 binding by dextran sulfate.

3000
2500
2000
Z LJ c~ 1500
nF- 1000 0
GLYC. q
gp120~EGLYC. / gp120 5OO
o , 101 102 103 104
[rgp120], PG/ML
Fig. 4. Comparative CD4 binding of recombinant, fully-glycosylated gp 120 ('Glyc. gp 120'), enzymatically- deglycosylated gpl20 ('Deglyc. gpl20'), and recombinant, non-glycosylated gpl20 ('Non-glyc. gpl20').
polysaccharides reduced binding of gpl20 as well as binding of Leu3a and OKT4. For example, while binding of gpl20 was reduced by 50% with dextran sulfate at 630 ng/ml, binding of Leu3a and OKT4 were reduced by 69 and 87%, respectively.
Effect of glycosylation on binding of gpl20 to CD4
Two glycosylation-variant gpl20 molecules were tested. One, kindly provided by Dr. Jeff Culp of Smith Kline Beecham Research Laboratories, was recombinant gpl20 produced in insect cells which had been 80% deglycosylated enzymatically (Ivey-Hoyle et al., 1991) (see Methods). The second gpl20 was non-glycosylated gpl20 produced in yeast (see Methods). Assay showed that binding of the enzymatically-deglycosylated material was equivalent to the fully-glycosylated material on a weight basis, while binding of the non-glycosylated gpl20 was reduced by about 10-fold (Fig. 4).
Discussion
These data show that our microtiter-format assay for gpl20-CD4 binding can be used to rapidly, simply and economically screen for inhibitors of binding, and the assay has the same parameters as more complex systems for measuring binding. Inhibitors with defined mechanism of action, such as Leu3a or rsCD4, all had the same properties in this assay system as reported in more physiologic systems involving binding of whole virus to intact cells (Schols et al., 1990; Balzarini et al., 1991). Similarly, our studies with sulfated polysaccharides confirmed that these compounds block gpl20 binding to

rsCD4 by binding primarily to CD4 (Parish et al., 1990; Lederman et al., 1989; Callahan 1991). Although other investigators have reported some binding of sulfated polysaccharides to gpl20 (Schols et al., 1990; Mbemba et al., 1992), noted by Parish et al., the coating of gpl20 with dextran sulfate does not appear to affect its binding to CD4 (Parish et al., 1990). Our data also confirm previous studies showing that the binding site for sulfated polysaccharides includes more than the Leu3a/OKT4a epitopes of CD4 (Parish et al., 1990).
Several monoclonal antibodies and sera to HIV gpl20 failed to block gpl20- CD4 binding, even though they neutralised infectivity and blocked syncytium formation at high dilutions in vitro (Hansen et al., 1990; Ardman et al., 1990; Ho et al., 1991). This reinforces previous work showing that neutralisating antibodies need not block gpl20 binding to CD4, through strain-specific binding to the V3 loop (Chamat et al., 1992), although all compounds which block the gp 120-CD4 interaction have also been shown to neutralise infectivity (Deen et al., 1988; Schols et al., 1989; Schols et al., 1990; Balzarini et al., 1991; Moore et al., 1992). How such V3-binding antibodies might neutralise virus through a post-binding event is not known, but as these antibodies also block cell-cell fusion, it is thought to be mediated by inhibition of virus-cell fusion (Parish et al., 1990; Healey et al., 1990; Ashorn et al., 1990). Sera from HIV- infected subjects, containing polyclonal human antibodies to HIV, blocked gpl20-binding at dilutions of over 1:10 -6, as Schnittman et al. also reported (Schnittman et al., 1988). The class and isotype of these antibodies involved in binding inhibition, the sequence of development of these antibodies, and their clinical significance, remain to be investigated. Likewise, the identity of the factor(s) in normal serum which prevent binding are not known. Human mannose-binding protein is likely to play a role in first line host defence against HIV and HIV infected cells as previous work has indicated that it can inhibit infectivity of H9 cells with a lymphotrophic strain of HIV (Ezekowitz et al., 1989). Furthermore, mannose-binding protein selectively recognises HIV infected cells by binding to the high mannose glycans present on gpl20. Concentrations of mannose-binding protein that were found to inhibit binding in our assay are within the physiological range that normally occurs within human serum. In this regard, the mannose-binding protein is an acute phase reactant and its precise role in natural defences in HIV infection remains to be elucidated.
The role of gpl20 oligosaccharide residues in CD4-gpl20 binding has been controversial, with some studies showing them to be unimportant (Fenouillet et al., 1989), while others show a marked effect (Matthews et al., 1987). Our studies support previous data suggesting that oligosaccharide residues on gpl20 play a key role in CD4 binding (Matthews et al., 1987). On a molar basis, gpl20 deglycosylated by EndoH/EndoF treatment bound about 50% less well than fully-glycosylated material; non-glycosylated gpl20 bound about 10-fold less than normal, and gpl20-CD4 binding was inhibited by lectins. Recently, elegant studies by Morikawa et al. have shown that removal of a single glycosylation site in HIV-2 gpl20 by mutagenesis resulted in gpl20 with

10
dramatically reduced CD4 binding capacity (Morikawa et al., 1991). The requirement for site-specific glycosylation shown by these studies suggests an explanation for some of our discordant results. Deglycosylation by random enzymatic attack may not remove the critical sugar residues efficiently, and the neutralising antibodies to oligosaccharides which failed to inhibit binding (Hansen et al., 1990) also may not bind these critical sugar residues well. Likewise, drugs which inhibit glycosylation of viral gpl20, such as deoxynojirimycin, may be relatively ineffective because they do not block glycosylation at critical sites (Pal et al., 1989).
In summary, we have developed a rapid, semi-automated assay system which can be used to screen for inhibitors of gpl20-CD4 binding, to quantitate the activity of newly-discovered inhibitors, and to obtain preliminary data on the mechanism of inhibition (binding to CD4 or gpl20). Wider application of this assay system may result in discovery of clinically useful antiretrovirals that act wholly or in part by this mechanism.
References
Ardman, B., Kowalski, M., Bristol, J., Haseltine, W. and Sodroski, J. (1990) Effects on CD4 binding of anti-peptide sera to the fourth and fifth conserved domains of HIV-1 gpl20. J. AIDS 3, 206- 214.
Arthos, J., Deen, K.C., Chaikin, M.A., Fornwald, J.A., Sathe, G., Sattentau, Q.J., Clapham, P.R., Weiss, R.A., McDougall, J.S., Pietropaolo, C., Axel, R., Truneh, A., Maddon, D.J. and Sweet, R.W. (1989) Identification of the residues in human CD4 critical for the binding of HIV. Cell 57, 469-481.
Ashkenazi, A., Presta, L.G., Marsters, S.A., Camerato, T.R., Rosenthal, K.A., Fendly, B.M. and Capon D.J. (1990) Mapping the CD4 binding site for human immunodeficiency virus by alanine- scanning mutagenesis. Proc. Natl. Acad. Sci. USA 87, 7150-7154.
Ashorn, P.A., Berger, E.A. and Moss, B. (1990) Human immunodeficiency virus envelope glycoprotein/CD4-mediated fusion of nonprimate cells with human cells. J. Virol. 64, 2149-2156.
Balzarini, J., Schols, D., Neyts, J., Van, D.E., Peumans, W. and De Clercq, E. (1991) Alpha-(l-3)- and alpha-(1-6)-o-mannose-specific plant lectins are markedly inhibitory to human immuno- deficiency virus and cytomegalovirus infections in vitro. Antimicrob. Agents Chemother. 35, 41ff416.
Brodsky, M.H., Warton, M., Myers, R.M. and Littman, D.R. (1990) Analysis of the site in CD4 that binds to the HIV envelope glycoprotein. J. Immunol. 144, 3078 3086.
Byrn, R.A., Sekigawa, I., Chamow, S.M., Johnson, J.S., Gregory,T.J., Capon, D.J. and Groopman, W.E. (1989) Characterization of in vitro inhibition of human immunodeficiency virus by purified recombinant CD4. J. Virol. 63, 4370-4375.
Callahan, L.N., Phelan, M., Mallinson, M. and Norcross, M.A. (1991) Dextran sulfate blocks antibody binding to the principal neutralizing domain of human immunodeficiency virus type 1 without interfering with gpl20-CD4 interactions. J. Virol. 65, 1543 1550.
Chamat, S., Nara, P., Berquist, L., Whalley, A., Morrow, W.J.W., K6hler, H. and Kang, C-Y. (1992) Two major groups of neutralizing anti-gp 120 antibodies exist in HIV-infected individuals. J. Immunol. 149, 649 654.
Cordonnier, A., Montagnier, L. and Emerman, M. (1989) Single amino-acid changes in HIV envelope affect viral tropism and receptor binding. Nature 340, 571 574.
Crowe, S.M., Mills, J., Kirihara, J., Boothman, J., Marshall, J.A. and McGrath, M.S. (1990) Full- length recombinant CD4 and recombinant gpl20 inhibit fusion between HIV-infected

11
macrophages and uninfected CD4-expressing T-lymphoblastoid cells. AIDS Res. Hum. Retroviruses 6, 1031 1038.
Dalgleish, A.G., Beverley, P.C., Clapham, P.R., Crawford, D.H., Greaves, M.F. and Weiss, R.A. (1984) The CD4 (T4) antigen is an essential component of the receptor for the AIDS retrovirus. Nature 312, 763-767.
Deen, K.C., McDougal, J.S., Inacker, R., Folena-Wasserman, G., Arthos, J., Rosenberg, J., Maddon, P.J., Axel, R. and Sweet, R.W. (1988) A soluble form of CD4 (T4) protein inhibits AIDS virus infection. Nature 331, 82-84.
Diamond, D.C., Steckman, B.P., Gregory, T., Lasky, L.A., Greenstein, J.L. and Burakoff, S.J. (1988) Inhibition ofCD4+ T cell function by the HIV envelope protein, gpl20. J. Immunol. 141, 3715-3717.
Ezekowitz, R.A., Kuhlman, M., Groopman, J.E. and Byrn, R.A. (1989) A human serum mannose- binding protein inhibits in vitro infection by the human immunodeficiency virus. J. Exp. Med. 169, 185-196.
Fenouillet, E., Clerget, R.B., Gluckman, J.C., Guetard, D., Montagnier, L. and Bahraoui, E. (1989) Role of N-linked glycans in the interaction between the envelope glycoprotein of human immunodeficiency virus and its CD4 cellular receptor. Structural enzymatic analysis. J. Exp. Med. 169, 807-822.
Gilbert, M., Kirihara, J., Elbeik, T. and Mills, J. (1990) Human immunodeficiency virus grown in CD4-expressing cells is coated with CD4.30th Interscience Conference on Antimicrobial Agents and Chemotherapy (ICAAC). American Society for Microbiology, Abstract 306, 307.
Gilbert. M., Kirihara, J. and Mills, J. (1991) Enzyme-linked immunoassay for human immunodeficiency virus type 1 envelope glycoprotein 120. J. Clin. Microbiol. 29, 142-147.
Habeshaw, J.A. and Dalgleish, A.G. (1989) The relevance of HIV env/CD4 interactions to the pathogenesis of acquired immune defciency syndrome. J. Acq. Immunol. Syndr. 2, 457-468.
Hansen, J-E., Clausen, H., Nielsen, C., Teglbjaerg, L.S., Hansen, L.L., Nielsen, C.M., Dabelsteen, E., Mathiesen, L., Hakomori, S.I. and Nielsen, J.O. (1990) Inhibition of human immunode- ficiency virus (HIV) infection in vitro by anticarbohydrate monoctonal antibodies: peripheral glycosylation of HIV envelope glycoprotein gpl20 may be a target for virus neutralization. J. Virol. 64, 2833 2840.
Healey, D., Dianda, L., Moore, J.P., McDougal, J.S., Moore, M.J., Estess, P., Buck, D., Kwong, P.D., Beverley, P.C. and Sattentau, Q.J. (1990) Novel anti-CD4 monoclonal antibodies separate human immunodeficiency virus infection and fusion of CD4 + cells from virus binding. J. Exp. Med. 172, 1233-1242.
Ho, D.D., McKeating, J.A., Li, X.L., Moudgil, T., Daar, E.S., Sun, N.C. and Robinson, J. (1991) Conformational epitope on gpl20 important in CD4 binding and human immunodeficiency virus type 1 neutralization identified by a human monoclonal antibody. J. Virol. 65, 489-493.
Ivey-Hoyle, M., Culp, J.S., Chaikin, M.A., Hellmig, B.D., Matthews, T.J., Sweet, R.W. and Rosenberg, M. (1991) Envelope glycoproteins from biologically diverse isolates of immunode- ficiency viruses have widely different affinities for CD4. Proc. Natl. Acad. Sci. USA 88, 512 516.
Klatzmann, D., Champagne, E., Chamaret, S., Gruest, J., Guetard, D., Hercend, T., Gluckman, J.C. and Montagnier, L. (1984) T-lymphocyte T4 molecule behaves as the receptor for human retrovirus LAV. Nature 312, 767-768.
Kowalski, M., Potz, J., Basiripour, L., Dorfman, T., Gob, W.C., Terwilliger, E., Dayton, A., Rosen, C., Haseltine, W. and Sodroski, J. (1987) Functional regions of the envelope glycoprotein of human immunodeficiency virus type 1. Science 237, 1351 1355.
Lasky, L.A., Nakamura, G., Smith, D.H., Fennie, C., Shimasaki, C., Patzer, E., Besman, P., Gregory, T. and Capon, D.J. (1987) Delineation of a region of the human immunodeficiency virus type 1 gpl20 glycoprotein critical for interaction with the CD4 receptor. Cell 50, 975-985.
Layne, S.P., Merges, M.J., Dembo, M., Spouge, J.L. and Nara, P.L. (1990) HIV requires multiple gpl20 molecules for CD4-mediated infection. Nature 346, 277-279.
Lederman, S., Gulick, R. and Chess, L. (1989) Dextran sulfate and heparin interact with CD4 molecules to inhibit the binding of coat protein (gp120) of H IV. J. Immunol. 143, 1149-1154.
Lifson, J.D., Reyes, G.R., McGrath, M.S., Stein, B.S. and Engleman, E.G. (1986) AIDS retrovirus induced cytopathology: giant cell formation and involvement of CD4 antigen. Science 232, 1123-1127.

12
Linsley, P.S., Ledbetter, J.A., Kinney, T.E. and Hu, S-L. (1988) Effects of anti-gpl20 monoclonal antibodies on CD4 receptor binding by the env protein of human immunodeficiency virus type 1. J. Virol. 62, 3695-3702.
Mann, D.L., Lasane, F., Popovic, M., Arthur, L.O., Robey, W.G., Blattner, W.A and Newman, M.J. (1987) HTLV-III Large envelope protein (gpl20) suppresses PHA-induced lymphocyte blastogenesis. J. Immunol. 138, 2640-2644.
Matthews, T.J., Weinhold, K.J., Lyerly, H.K., Langlois, A.J., Wigzell, H. and Bolognesi, D.P. (1987) Interaction between the human T-cell lymphotropic virus type IIIB envelope glycoprotein gpl20 and the surface antigen CD4: role of carbohydrate in binding and cell fusion. Proc. Natl. Acad. Sci. USA 84, 5424-5428.
Mbemba, E., Chams, V., Gluckman, J.C., Klatzmann, D. and Gattegno, L. (1992) Molecular interaction between HIV-I major envelope glycoprotein and dextran sulfate. Biochem. Biophys. Acta 1138, 62~57.
McKeating, J.A. and Willey, R.L. (1989) Structure and function of the HIV envelope. AIDS 3, $35- $39.
Moore, J.P., Simpson, G., McKeating, J.A., Burakoff, S.J., Schreiber, S.L. and Weiss, R.A. (1992) CPF-DD is an inhibitor of infection by human immunodeficiency virus and other enveloped viruses in vitro. Virology 188, 537-544.
Morikawa, Y., Moore, J.P., Wilkinson, A.J. and Jones, I.M. (1991) Reduction in CD4 binding affinity associated with removal of a single glycosylation site in the external glycoprotein of HIV- 2. Virology 180, 853 856.
Miiller, W.E., Renneisen, K., Kreuter, M.H., Schr6der, H.C. and Winkler, I. (1988) The D- mannose-specific lectin from Gerardia savaglia blocks binding of human immunodeficiency virus type I to H9 cells and human lymphocytes in vitro. J. Acq. Immun. Def. Syndr. 1, 453458.
Olshevsky, U., Helseth, E., Furman, C., Li, J., Haseltine, W. and Sodroski, J. (19o0) Identification of individual human immunodeficiency virus type lgpl20 amino acids important for CD4 receptor binding. J. Virol. 64, 5701-5707.
Pal, R., Kalyanaraman, V.S., Hoke, G.M. and Sarngadharan, M.G. (1989) Processing and secretion of envelope glycoproteins of human immunodeficiency virus type 1 in the presence of trimming glucosade inhibitor deoxynojirimycin. Intervirol. 30, 27-35.
Parish, C.R., Low, L., Warren, H.S. and Cunningham, A.L. (1990) A polyanlon binding site on the CD4 molecule. Proximity to the HIV-gpl20 binding region. J. Immunol. 145, 1188-1195.
Robinson, J.E., Holton, D., Pacheco, M.S., Liu, J. and McMurdo, H. (1990) Identification of conserved and variant epitopes of human immunodeficiency virus type 1 (HIV-1) gpl20 by human monoclonal antibodies produced by EBV-transformed cell lines. AIDS Res. Hum. Retroviruses 6, 567-579.
Robinson, W.E., Jr, Montefiori, D.C. and Mitchell, W.M. (1987) Evidence that mannosyl residues are involved in human immunodeficiency virus type 1 (HIV-1) pathogenesis. AIDS Res. Hum. Retroviruses 3, 265-282.
Schnittman, S.M., Lane, H.C., Roth, J., Burrows, A., Folks, T.M., Kehrl, J.H., Koenig, S., Berman, P. and Fauci, A.S. (1988) Characterization of GP 120 binding to CD4 and an assay that measures ability of sera to inhibit this binding. J. Immunol. 141, 41814186.
Schols, D., Baba, M., Pauwels, R., Desmyter, J. and De Clercq, E. (1989) Specific interaction of aurintricarboxylic acid with the human immunodeficiency virus/CD4 cell receptor. Proc. Natl. Acad. Sci. USA 86, 3322-3326.
Schols, D., Pauwels, R., Desmyter, J. and De Clercq, E. (1990) Dextran sulfate and other polyanionic anti-HIV compounds specifically interact with the viral gpl20 glycoprotein expressed by T-cells persistently infected with HIV-1. Virology 175, 556-561.
Sun, N.C., Ho, D.D., Sun. C.R., Liou, R-W., Gordon, W., Fung, M.S.C., Li, X-L., Ting, R.C., Lee, T-H., Chang, N.T. and Chang, T-W. (1989) Generation and characterization of monoclonal antibodies to the putative CD4-binding domain of human immunodeficiency virus type 1 gp 120. J. Virol. 63, 3579-3585.
Weiler, B.E., SchrSder, H.C., Stefanovich, V., Stewart, D., Forrest, J.M., Allen, L.B., Bowden, B.J., Kreuter, M.H., Voth, R. and M~ller, W.E. (1990) Sulphoevernan, a polyanionic polysaccharide, and the Narcissus lectin potently inhibit human immunodeficiency virus infection by binding to viral envelope protein. J. Gen. Virol. 71, 1957-1963.
Related Documents










![Envelope Glycoprotein 120 (gp120) and nef Genes · MMcGrath@php.ucsf.edu [Abstract] The current study provides detailed protocols utilized to amplify the complete HIV-1 gp120 and](https://static.cupdf.com/doc/110x72/5f7f5fb2cb45ef5cc60950cd/envelope-glycoprotein-120-gp120-and-nef-genes-mmcgrathphpucsfedu-abstract.jpg)
