Transgenic mice expressing Na -K -ATPase in smooth muscle decreases blood pressure Tracy J. Pritchard, 1 Michelle Parvatiyar, 1 Daniel P. Bullard, 1 Ronald M. Lynch, 2 John N. Lorenz, 1 and Richard J. Paul 1 1 Department of Molecular and Cellular Physiology, University of Cincinnati College of Medicine, Cincinnati, Ohio; and 2 Department of Physiology, University of Arizona Health Sciences Center, Tucson, Arizona Submitted 6 March 2007; accepted in final form 25 April 2007 Pritchard TJ, Parvatiyar M, Bullard DP, Lynch RM, Lorenz JN, Paul RJ. Transgenic mice expressing Na -K -ATPase in smooth muscle decreases blood pressure. Am J Physiol Heart Circ Physiol 293: H1172–H1182, 2007. First published April 27, 2007; doi:10.1152/ajpheart.00279.2007.—The Na -K -ATPase (NKA) is a transmembrane protein that sets and maintains the electrochemical gradient by extruding three Na in exchange for two K . An impor- tant physiological role proposed for vascular smooth muscle NKA is the regulation of blood pressure via modulation of vascular smooth muscle contractility (5). To investigate the relations between the level of NKA in smooth muscle and blood pressure, we developed mice carrying a transgene for either the NKA 1- or 2-isoform (1sm or 2sm mice) driven by the smooth muscle-specific -actin promoter SMP8. Interestingly, both -isoforms, the one contained in the trans- gene and the one not contained, were increased to a similar degree at both protein and mRNA levels. The total -isoform protein was increased from 1.5-fold ( 1sm mice) to 7-fold (2sm mice). The increase in total NKA -isoform protein was accompanied by a 2.5-fold increase in NKA activity in 2sm gastric antrum. Immuno- cytochemistry of the 1- and 2-isoforms in 2sm aortic smooth muscle cells indicated that -isoform distributions were similar to those shown in wild-type cells. 2sm Mice (high expression) were hypotensive (109.9 1.6 vs. 121.3 1.4 mmHg; n 13 and 11, respectively), whereas 1sm mice (low expression) were normoten- sive (122.7 2.5 vs. 117.4 2.3; n 11 or 12). 2sm Aorta, but not 1sm aorta, relaxed faster from a KCl-induced contraction than wild-type aorta. Our results show that smooth muscle displays unique coordinate expression of the -isoforms. Increasing smooth muscle NKA decreases blood pressure and is dependent on the degree of increased -isoform expression. vascular contractility; hypertension; Na -K -ATPase -isoforms THE Na -K -ATPase (NKA) is a transmembrane protein that sets and maintains the electrochemical gradient by extruding three Na in exchange for two K , which underlies numerous Na -coupled transport processes. An important physiological role proposed for NKA is the regulation of blood pressure via modulation of vascular smooth muscle contractility (5). With hypertension, heart failure, and diabetes, conditions known to have a vascular involvement, cardiac NKA expression has been reported to be changed (18, 32, 34). NKA expression can be induced by insulin, thyroid hormone, and aldosterone (8), hormones whose concentrations are altered in disease states. However, there are limited data regarding smooth muscle- specific changes in NKA expression with disease. Furthermore, diseases are multifactorial, thereby complicating the under- standing of changes in smooth muscle phenotypes solely in terms of NKA expression. In the case of salt-sensitive hypertension, NKA is hypothe- sized (5, 13) to participate, via modulation of intracellular Ca 2 concentration ([Ca 2 ] i ) and contractility, by coupling to the Na /Ca 2 exchanger (NCX). A variety of studies have demonstrated that decreases in NKA activity are associated with an elevated blood pressure. These include pharmacolog- ical manipulation with cardiac glycosides as well as genetically induced decreases in NKA expression (5, 7, 11). However, to validate causality rather than correlation, gain of function studies are needed to demonstrate whether increases in NKA activity can produce decreases in blood pressure. NKA is a heterodimer composed of an -subunit, responsi- ble for catalytic function, and a -subunit (2, 4, 7–9, 16). In aorta, two ( 1 and 2 ) (30) of the four known -isoforms (2, 4, 7) are present; 70% of the total NKA is composed of the 1 -isoform and the remaining 30% is the 2 -isoform (30). To investigate a gain of NKA function on blood pressure, we developed mice that carry transgenes for either the NKA 1 - or 2 -isoform, using the smooth muscle-specific -actin promoter (6) SMP8. The -isoforms in smooth muscle from the 1sm mouse line were increased by 1.5-fold and up to 7-fold in the 2sm mouse line. An important, but surprising observation was that the -isoform not contained in the transgene was upregulated to a similar degree. We show that an increase in NKA -isoforms leads to a decreased blood pressure, which is dependent on the degree of increased vascular NKA expres- sion. Thus increased expression of vascular NKA can lead to hypotension and perhaps provides a novel pathway for treat- ment of hypertension. METHODS Generation of transgenic mice. 1- or 2-isoform NKA rat cDNA (gift from Dr. Jerry Lingrel, University of Cincinnati) was inserted via HindIII restriction sites into transfer vector pSAHSK GSSS (Fig. 1, A and B) containing AscI and SnabI restriction sites (gener- ated by Genomatix, Cincinnati, OH). Resultant clones were screened for cDNA orientation using XbaI and NotI restriction endonucleases for vectors containing the 1-isoform or EcoRI and NotI enzymes for vectors with the 2-isoform. The 1- or 2-isoform NKA cDNA was then cloned via AscI and SnabI sites into target vector pSMP8 RPNKAA human growth hormone poly(A) (generated by Geno- matix) containing the -actin smooth muscle-specific promoter SMP8 (6) and a human growth hormone polyadenylation sequence (Fig. 1C). Proper construction of the transgenes was verified by sequencing the Address for reprint requests and other correspondence: R. J. Paul, Molecular and Cellular Physiology, Univ. of Cincinnati College of Medicine, 231 Albert Sabin Way, Cincinnati, OH 45267-0576 (e-mail: [email protected]). The costs of publication of this article were defrayed in part by the payment of page charges. The article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. Am J Physiol Heart Circ Physiol 293: H1172–H1182, 2007. First published April 27, 2007; doi:10.1152/ajpheart.00279.2007. 0363-6135/07 $8.00 Copyright © 2007 the American Physiological Society http://www.ajpheart.org H1172

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Transgenic mice expressing Na�-K�-ATPase in smooth muscle decreasesblood pressure
Tracy J. Pritchard,1 Michelle Parvatiyar,1 Daniel P. Bullard,1 Ronald M. Lynch,2
John N. Lorenz,1 and Richard J. Paul11Department of Molecular and Cellular Physiology, University of Cincinnati College of Medicine, Cincinnati, Ohio;and 2Department of Physiology, University of Arizona Health Sciences Center, Tucson, Arizona
Submitted 6 March 2007; accepted in final form 25 April 2007
Pritchard TJ, Parvatiyar M, Bullard DP, Lynch RM, LorenzJN, Paul RJ. Transgenic mice expressing Na�-K�-ATPase insmooth muscle decreases blood pressure. Am J Physiol Heart CircPhysiol 293: H1172–H1182, 2007. First published April 27, 2007;doi:10.1152/ajpheart.00279.2007.—The Na�-K�-ATPase (NKA) is atransmembrane protein that sets and maintains the electrochemicalgradient by extruding three Na� in exchange for two K�. An impor-tant physiological role proposed for vascular smooth muscle NKA isthe regulation of blood pressure via modulation of vascular smoothmuscle contractility (5). To investigate the relations between the levelof NKA in smooth muscle and blood pressure, we developed micecarrying a transgene for either the NKA �1- or �2-isoform (�1sm� or�2sm� mice) driven by the smooth muscle-specific �-actin promoterSMP8. Interestingly, both �-isoforms, the one contained in the trans-gene and the one not contained, were increased to a similar degree atboth protein and mRNA levels. The total �-isoform protein wasincreased from 1.5-fold (�1sm� mice) to 7-fold (�2sm� mice). Theincrease in total NKA �-isoform protein was accompanied by a2.5-fold increase in NKA activity in �2sm� gastric antrum. Immuno-cytochemistry of the �1- and �2-isoforms in �2sm� aortic smoothmuscle cells indicated that �-isoform distributions were similar tothose shown in wild-type cells. �2sm� Mice (high expression) werehypotensive (109.9 � 1.6 vs. 121.3 � 1.4 mmHg; n � 13 and 11,respectively), whereas �1sm� mice (low expression) were normoten-sive (122.7 � 2.5 vs. 117.4 � 2.3; n � 11 or 12). �2sm� Aorta, butnot �1sm� aorta, relaxed faster from a KCl-induced contraction thanwild-type aorta. Our results show that smooth muscle displays uniquecoordinate expression of the �-isoforms. Increasing smooth muscleNKA decreases blood pressure and is dependent on the degree ofincreased �-isoform expression.
vascular contractility; hypertension; Na�-K�-ATPase �-isoforms
THE Na�-K�-ATPase (NKA) is a transmembrane protein thatsets and maintains the electrochemical gradient by extrudingthree Na� in exchange for two K�, which underlies numerousNa�-coupled transport processes. An important physiologicalrole proposed for NKA is the regulation of blood pressure viamodulation of vascular smooth muscle contractility (5). Withhypertension, heart failure, and diabetes, conditions known tohave a vascular involvement, cardiac NKA expression hasbeen reported to be changed (18, 32, 34). NKA expression canbe induced by insulin, thyroid hormone, and aldosterone (8),hormones whose concentrations are altered in disease states.However, there are limited data regarding smooth muscle-specific changes in NKA expression with disease. Furthermore,diseases are multifactorial, thereby complicating the under-
standing of changes in smooth muscle phenotypes solely interms of NKA expression.
In the case of salt-sensitive hypertension, NKA is hypothe-sized (5, 13) to participate, via modulation of intracellularCa2� concentration ([Ca2�]i) and contractility, by coupling tothe Na�/Ca2� exchanger (NCX). A variety of studies havedemonstrated that decreases in NKA activity are associatedwith an elevated blood pressure. These include pharmacolog-ical manipulation with cardiac glycosides as well as geneticallyinduced decreases in NKA expression (5, 7, 11). However, tovalidate causality rather than correlation, gain of functionstudies are needed to demonstrate whether increases in NKAactivity can produce decreases in blood pressure.
NKA is a heterodimer composed of an �-subunit, responsi-ble for catalytic function, and a �-subunit (2, 4, 7–9, 16). Inaorta, two (�1 and �2) (30) of the four known �-isoforms (2, 4,7) are present; 70% of the total NKA is composed of the�1-isoform and the remaining 30% is the �2-isoform (30). Toinvestigate a gain of NKA function on blood pressure, wedeveloped mice that carry transgenes for either the NKA �1- or�2-isoform, using the smooth muscle-specific �-actin promoter(6) SMP8. The �-isoforms in smooth muscle from the �1sm�
mouse line were increased by �1.5-fold and up to 7-fold in the�2sm� mouse line. An important, but surprising observationwas that the �-isoform not contained in the transgene wasupregulated to a similar degree. We show that an increase inNKA �-isoforms leads to a decreased blood pressure, which isdependent on the degree of increased vascular NKA expres-sion. Thus increased expression of vascular NKA can lead tohypotension and perhaps provides a novel pathway for treat-ment of hypertension.
METHODS
Generation of transgenic mice. �1- or �2-isoform NKA rat cDNA(gift from Dr. Jerry Lingrel, University of Cincinnati) was insertedvia HindIII restriction sites into transfer vector pSAHSK � GSSS(Fig. 1, A and B) containing AscI and SnabI restriction sites (gener-ated by Genomatix, Cincinnati, OH). Resultant clones were screenedfor cDNA orientation using XbaI and NotI restriction endonucleasesfor vectors containing the �1-isoform or EcoRI and NotI enzymes forvectors with the �2-isoform. The �1- or �2-isoform NKA cDNA wasthen cloned via AscI and SnabI sites into target vector pSMP8 �RPNKAA � human growth hormone poly(A) (generated by Geno-matix) containing the �-actin smooth muscle-specific promoter SMP8(6) and a human growth hormone polyadenylation sequence (Fig. 1C).Proper construction of the transgenes was verified by sequencing the
Address for reprint requests and other correspondence: R. J. Paul, Molecularand Cellular Physiology, Univ. of Cincinnati College of Medicine, 231 AlbertSabin Way, Cincinnati, OH 45267-0576 (e-mail: [email protected]).
The costs of publication of this article were defrayed in part by the paymentof page charges. The article must therefore be hereby marked “advertisement”in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Am J Physiol Heart Circ Physiol 293: H1172–H1182, 2007.First published April 27, 2007; doi:10.1152/ajpheart.00279.2007.
0363-6135/07 $8.00 Copyright © 2007 the American Physiological Society http://www.ajpheart.orgH1172

5� and 3� ends of the cDNA using primers targeted to the plasmid atthe 3� end of the promoter (5�-GTGTTAGTTGAGAACTGTGG-3�)and the 5� end of the polyadenylation sequence (5�-GAAGGACAC-CTAGTCAGACA-3�). Sequencing was performed by the Universityof Cincinnati DNA Core. The �1- or �2-isoform transgenes werelinearized (Fig. 1D) via two NotI sites, purified, and microinjected intothe pronucleus of fertilized mouse eggs (FVB/n) that were thenimplanted into pseudopregnant females by the University of Cincin-nati Transgenic Mouse Core (Cincinnati, OH). Resultant founder miceproduced from this procedure were identified by PCR and Southernblot analyses and then subsequently bred to wild-type (WT) FVB/nmice.
Animal use and tissue preparation. The treatment of animalsfollowed experimental protocols that were approved by the Universityof Cincinnati and University of Arizona Institutional Animal Care andUse Committees. Animals were given normal rodent chow (HarlanTeklad, Madison, WI) and water ad libitum. All animals used in thesubsequent experiments were gender-matched littermate pairs of12–15 wk old. Adult mice were euthanized in a precharged CO2
chamber followed by immediate removal of thoracic aorta, gastricantrum (lower portion of the stomach), and heart. Tissues weredissected and rinsed with cold physiological saline solution (PSS; inmM: 118 NaCl, 4.7 KCl, 25 NaHCO3, 2.5 CaCl2, 1.2 MgSO4, 1.2NaH2PO4, 0.026 EDTA, and 11 glucose).
Genotyping of transgenic mice. Founder mice and subsequenttransgenic offspring were identified by PCR. DNA from tail biopsies(�1 cm) was isolated using ethanol precipitation. Primers targetingthe 5� end of the �1-isoform cDNA (5�-ATGTCCCTTTCCTTCT-TCGC-3�) and the 3� end of the SMP8 promoter (5�-GGATCAT-CAAAGGCTTTACAGC-3�) or the 5� end of the human growthhormone poly(A) tail (5�-CAGGTTGTCTTCCCAACTTG-3�) andthe 3� end of the �2-isoform cDNA (5�-GGGTGGAGAAG-GAGACGTAC-3�) were generated for PCR analysis of �1sm� and�2sm� mice, respectively. PCR conditions included denaturation at94°C for 3 min, 30 cycles at 94°C for 1 min, 57°C for 1 min, 72°C for1 min, and elongation at 72°C for 10 min. PCR products wereelectrophoresed on 1% agarose gels containing ethidium bromide forvisualization.
Southern blot analysis. For Southern blot, tail DNA (extracted thesame as for genotyping) was digested with HindIII restriction enzymeand separated on a 1% agarose gel. DNA was transferred to a nylonmembrane and probed with a random primed 32P-labeled (randomprimer DNA labeling kit; Invitrogen, Carlsbad, CA) 1,360-bp frag-ment of the �1-isoform cDNA (isolated using BamHI and XbaIrestriction enzymes) or 1,965-bp fragment of the �2-isoform cDNA(isolated using BamHI and EcoRI restriction enzymes). The probeshybridized to a 4-kb product of the HindIII-digested tail DNA.
Western blot analysis. Aorta was dissected free from connectivetissue and fat, and the endothelium was removed. Gastric antrum wasrinsed free of digestive debris, and the mucosa was removed. Heartswere removed of any fat and connective tissue and rinsed thoroughlyof blood. Tissues were frozen in liquid N2. Aorta was homogenizedwith a dental amalgamator (Caulk, Milford, DE) and antrum and heartwith a PowerGen 125 homogenizer (Fisher Scientific, Pittsburgh, PA)in buffer containing 50 mM Tris �HCl, 1% Triton X-100, 0.5%deoxycholic acid, 0.1% SDS, 500 mM NaCl, and 10 mM MgCl2. Thehomogenization buffer was supplemented with the following inhibi-tors used at 1:100: phosphatase inhibitor cocktails I and II andprotease inhibitor cocktail (Sigma, St. Louis, MO). Protein homoge-nates were then assayed to determine concentration, with BSA used asa standard (Bio-Rad protein assay, Hercules, CA).
Protein from aorta (10–40 �g) and protein from antrum or heart(2.5–20 �g) were incubated at 37°C for 30 min in Laemmli buffersupplemented with 5% �-mercaptoethanol followed by separation bySDS-PAGE (Genemate 4–20% express gels; ISC BioExpress,Kaysville, UT) at 100 V for 3 h and then electrotransferred at 220 mAto polyvinylidene difluoride membranes (Millipore, Bedford, MA)overnight at 4°C. The membranes were incubated in 1% blockingreagent (Roche Diagnostics, Indianapolis, IN) in TBST (50 mMTris �HCl, pH 7.4, 150 mM NaCl, and 0.05% Tween 20) for 1 h atroom temperature. Blots were then incubated with monoclonal anti-�1-antibody that binds to the NH2 terminus of the �1-isoform (�6F;Developmental Studies Hybridoma Bank, University of Iowa), poly-clonal anti-�2-antibody that recognizes an intracellular loop region ofthe �2-isoform [affinity purified from �2-antisera using the syntheticHERED peptide (gift from Dr. Thomas Pressley)] (26), or polyclonal
Fig. 1. Linear map of the Na�-K�-ATPase (NKA) �-isoform transgenes. HindIII-bound NKA �1- or �2-isoform rat cDNAs (A) were cloned into transfer vectorpSAHSK � GSSS (Genomatix) (B). The �-isoform cDNA was then inserted into target vector pSMP8 � RPNKAA � human growth hormone (hGH) poly(A)(Genomatix) via AscI and SnabI sites (C). The transgene was released at NotI sites for pronuclear microinjection (D).
H1173SMOOTH MUSCLE NKA AND BLOOD PRESSURE
AJP-Heart Circ Physiol • VOL 293 • AUGUST 2007 • www.ajpheart.org

KETYY antibody that recognizes the COOH terminus of NKA�-isoforms (kindly provided by Dr. Jack Kyte) in TBST for 1.5 h atroom temperature. Equal loading was verified with the monoclonalanti-actin C4 antibody (gift from Dr. James Lessard) or Ponceau Sstaining of the actin band. Peroxidase-conjugated goat anti-mouse(Bio-Rad) or anti-rabbit (Calbiochem, La Jolla, CA) secondary anti-bodies were used.
Blots were developed with an enhanced chemiluminescence kit(Amersham Biosciences, Piscataway, NJ) and exposed with BlueBasic autorad film (ISC BioExpress). Multiple exposures were takento ensure linearity of the signal. Relative protein expression intransgenic mice compared with paired WT mice was quantitated bydensitometry using ImageQuant 5.2 software (Molecular Dynamics,Amersham Biosciences). All summarized results consist of data fromat least two Western blots representing two different protein samples.For each genotype, antrum from an average of three mice and aortafrom an average of seven mice were pooled for each sample.
Quantitative real-time PCR. Mouse antrum was dissected in ice-cold PSS and rinsed of all debris; the tissue was immediately frozenin liquid N2. Total RNA was isolated using Tri Reagent (MolecularResearch Center, Cincinnati, OH) according to manufacturer’s in-structions. Total RNA concentration and purity were measured spec-trophotometrically. Total RNA (2 �g for each sample) was treatedwith RQ1 RNase-free DNase I (Promega, Madison, WI) before cDNAsynthesis. cDNA synthesis was performed with random hexamerprimers using the SuperScript III first-strand synthesis system (In-vitrogen). cDNA was used in real-time PCR reactions to quantitaterelative mRNA levels of the NKA �-isoforms using published primersequences for the �1- and �2-isoforms (22) and GAPDH as an internalcontrol (12). iQ SYBR green Supermix (Bio-Rad) was used forreal-time PCR reactions according to manufacturer’s instructions.Reactions were run for 50 cycles at 95°C for 15 s, 58°C for 30 s, and72°C for 30 s using the iCycler real-time PCR machine and iCyclersoftware (Bio-Rad). cDNA reactions in which reverse transcriptasewas omitted were used as negative controls. Threshold cycles (auto-matically calculated by iCycler software) for GAPDH mRNA werecompared between transgenic and paired WT samples for normaliza-tion. Each antrum sample was assayed in triplicate, and each real-timePCR reaction was performed at least twice per sample. Three pairedantrum samples per transgenic line were used in the analysis. Relative�-isoform expression (in fold) in transgenic lines compared withpaired WT mice was calculated using the delta-delta threshold cyclemethod (25).
Western blot analysis of the NKA �-subunits. Gastric antrum from�1sm�, �2sm�, and paired WT mice were isolated and homogenized asdescribed in Western blot analysis above. Crude protein homogenatesremained either untreated or denatured and digested with glycosidasepeptide N-glycosidase F (New England Biolabs, Beverly, MA) ac-cording to manufacturer’s instructions. Protein samples were resolvedby SDS-PAGE and transferred to polyvinylidene difluoride mem-branes as described in Western blot analysis above. Blots were probedwith the polyclonal anti-�1- or anti-�2-antibodies spETB1 or spETB2(kindly provided by Dr. Pablo Martin-Vasallo). Three different an-trum samples per genotype were used for analysis where each samplecontained three pooled antrum. Relative NKA �-isoform proteinquantitation was performed as described in Western blot analysisabove.
Fluorescent, enzyme-linked measurements of NKA activity. Gastricantrum from �1sm�, �2sm�, and paired WT mice were dissected andhomogenized as described in Western blot analysis above, with theexception of the homogenization buffer. The homogenization buffercontained 0.25 M sucrose, 30 mM imidazole, and 1 mM Na�-freeEDTA and was not supplemented with any protease or phosphataseinhibitors.
NKA activities were determined with an enzyme-linked assaymeasuring the rate of ADP production as linked to the rate of NADHfluorescence decrease in the absence or presence of 1 mM ouabain.
Crude protein (50 �g) was incubated for 20 min at 37°C in solutionwith pH 7.4 containing 100 mM NaCl, 20 mM KCl, 8 mM MgCl2, 40mM Tris, 1 mM EGTA, 1 mM Na� azide, 25 mM choline chloride,1 U/ml lactate dehydrogenase, 1 U/ml pyruvate kinase, 1 mM phos-phoenolpyruvate, and 80 �M NADH. The reaction was initiated withthe addition of 1 mM ATP. NADH fluorescence was continuouslyfollowed (excitation � 340 nm, emission � 460 nm) for 20 min withthe use of a spectrofluorometer (PTI Delta Scan-1; Photon Technol-ogy International, South Brunswick, NJ). Rates were calculated asnanomoles ATP hydrolyzed per minute per milligram of protein.Three �1sm� or �2sm� and their paired WT antrum samples were usedin the NKA activity measurements, where each protein sample con-sisted of three pooled antrum. NKA activity was measured for eachsample at least twice.
Aorta and cell isolation. Adult mice were euthanized in a pre-charged CO2 chamber followed by removal of the entire thoracic aorta(exposed from aortic arch to diaphragm). The aorta was dissected freefrom all associated fatty tissue and removed by cutting the intercostalarteries. Blood was immediately flushed from the isolated vessel withPBS � 0.1% BSA. To produce cell cultures, aortae were incubated ina PBS solution containing collagenase B (6 mg/ml) and BSA (6mg/ml) and then incubated at 37°C for 40 min. Cells were dispersedfrom the digested aorta by trituration, diluted in PBS, and centrifugedat �1,000 g for 3 min. The cell pellet was suspended in 250 �l ofDMEM supplemented with 5% FBS and 1% penicillin-streptomycin.Cell suspension from one aorta (250 �l) was plated as 50-�l aliquotsonto the center of six glass coverslips housed in individual wells of asix-well plate. The six-well plate was then placed into a 37°Cincubator equilibrated with 5% CO2. After �16 h, 5 ml of freshDMEM were added to each well. The PBS contained (in mM) 2.7KCl, 1.5 KH2PO4, 138 NaCl, and 8 Na2HPO4, at pH 7.2.
Immunocytochemistry. At least 96 h after isolation, cells on cov-erslips were fixed with 3% paraformaldehyde, rinsed with 25 mMglycine, and permeabilized with 0.1% Triton X-100 (17). The cellswere incubated with rabbit polyclonal antibody raised against theNKA �2-isoform (26) for 2 h at 25°C. For �1-isoform labeling, cellswere incubated with the �1-isoform-specific antibody NASE (26) for12 h. After labeling with �-isoform-specific antibodies was com-pleted, cells were washed three times (5 min each) in PBS to removeunbound primary antibody and then incubated with a secondaryanti-rabbit IgG labeled with Texas red (Jackson ImmunoResearch,West Grove, PA) for 45 min at 25°C. Specificity was determined byincubating the antibody in the presence of its peptide antigen (26)before incubation with the sample. Under this condition, significantlabeling was not observed. Coverslips were mounted onto glass slidesusing a 50% glycerol-saline solution containing the antibleach agentparaphenylendiamine (0.1%).
For standard wide-field imaging, slides were mounted on the stageof an Olympus IX-70 microscope equipped with a 60 1.4 numericalobjective. Illumination was provided by a 100-W Hg lamp, andimages were acquired by a liquid-cooled charge-coupled device cam-era (Roper Scientific, Tucson, AZ) equipped with a Kodak charge-coupled device array (KAF1401E). Image analysis was performed ona Silicon Graphics INDY2 workstation using customized software.
Smooth muscle contractility measurements. Thoracic aorta wasdissected from male littermate mice. After removal of fatty tissue, theaorta was cut into two pieces (one piece with the endothelium intactand the other half with the endothelium removed by gently rubbingthe tissue between the thumb and index finger). Weight and calculatedthickness (15) of each aortic segment were not different between�1sm� or �2sm� mice and their respective WT pairs, averaging 0.81 �0.02 mg and 114 � 3 �m (n � 36), respectively. The aortic stripswere threaded between two steel triangles 100 �m in diameter. Thetriangles were mounted in an organ bath system between a fixed postand a Harvard Apparatus differential capacitor force transducer(South Natick, MA). Isometric force was recorded continuously(Biopac Instruments and Acqknowledge software, Goleta, CA). Tis-
H1174 SMOOTH MUSCLE NKA AND BLOOD PRESSURE
AJP-Heart Circ Physiol • VOL 293 • AUGUST 2007 • www.ajpheart.org

sues were maintained in PSS and bubbled with 95% O2-5% CO2 forpH 7.4 at 37°C. Resting tension of the aorta was adjusted to 30 mN toset an initial length in the range of maximum isometric force gener-ation. The tissues were then challenged with at least two contraction-relaxation cycles with 50 mM KCl (added from concentrate) to ensurereproducible forces. The tissues were then exposed to increasingcumulative concentrations of KCl or phenylephrine (PE). ACh (105
M) was used to verify the presence of endothelium in aorta. All forceswere analyzed and normalized to cross-sectional area as previouslydescribed (15).
Smooth muscle kinetics measurements. Contraction was elicited viadepolarization by raising medium KCl to 50 mM, and then relaxationwas initiated by rinsing with PSS. Relaxation half-times were calcu-lated as the time (in s) for aorta to relax to 50% of its peak force. Forcedevelopment half-times were calculated by measuring the elapsedtime required for each vessel to reach 50% of its peak force frombaseline in response to 50 mM KCl. Similar measurements wereperformed for challenges with 1 �M PE.
Blood pressure measurements. Systolic blood pressure (SBP) wasmeasured in male littermate mice using a computerized tail-cuffsystem (BP-2000 blood pressure analysis system; Visitech Systems,Apex, NC). The mice were allowed to acclimate to the apparatus for5 min before the start of measurement. Ten preliminary and 10experimental pressures were acquired for 5 consecutive days/wk.Pressures were taken on the same days each week and at the sametime each day. Data were accepted if the mouse had valid readings forat least 5 of the 10 experimental pressures taken. Blood pressures wereobtained during a 2-wk period. The daily blood pressure averages permouse were averaged to determine the mean SBP for that mouse.
Statistical analysis. The data are presented as means � SE, wheren is equal to the number of samples. Significance was determined with
an unpaired Student’s t-test, with a P � 0.05 taken as significantwhere only two variables were compared. For force-concentrationrelations, multivariate analysis using ANOVA and Holm-Sidak posthoc analysis was applied using SigmaStat 3.1.
RESULTS
Generation of NKA �-isoform transgenic mice. Transgenicmice were generated by inserting �1- or �2-isoform cDNA(31) between the smooth muscle-specific �-actin promoter,SMP8 (6) and a polyadenylation sequence in the SMP8 �RPNKAA � human growth hormone poly(A) plasmid(Fig. 1). After pronuclear microinjection with the linearizedtransgene, one NKA transgenic founder for the �1-isoform(line 09) screened from 14 mice by PCR and Southern blotanalyses was identified (Fig. 2). Two NKA transgenic foundersfor the �2-isoform (line 79 and line 84) were identified from 35resultant mice (Fig. 2). Transmission of the transgenes fromfounder mice to offspring was verified by Southern blot (Fig.2B). Both �1sm� line 09 mice and �2sm� line 84 mice havenormal litter sizes of eight and seven pups, respectively,although the frequency of litters born is less in �1sm� line 09than in WT mice. As calculated from PCR analysis, thetransgene was passed onto �44% of offspring in �1sm� line 09mice and to �51% of �2sm� line 84 mice. The �2sm� line 79mice have small litter sizes, averaging only 3.6 mice, and a lowfrequency of births; transmittance of the transgene appearednormal with �53% of the offspring carrying the transgene. Inall transgenic lines, Mendelian ratios of male to female mice
Table 1. Summary of body and wet tissue weights
Genotype
�1 WT �1sm� �2 WT �2sm�
Body weight, g 26.86�0.57 (51) 28.09�0.67 (43) 27.37�0.72 (57) 28.96�0.55 (85)Aorta, mg 1.90�0.09 (39) 2.00�0.14 (36) 1.87�0.08 (55) 2.01�0.07 (86)Antrum, mg 118.79�3.84 (36) 122.90�5.42 (36) 116.40�4.42 (48) 121.78�4.36 (63)Bladder, mg 15.73�2.19 (23) 19.53�2.67 (23) 16.45�2.08 (22) 21.04�2.66 (31)Fundus, mg 36.00�1.62 (15) 38.01�2.43 (21) 36.14�1.96 (20) 39.38�2.54 (23)Heart, mg 113.05�3.70 (13) 114.61�3.62 (16) 122.34�4.28 (15) 125.66�5.96 (12)
Values are means � SE, where n (no. of samples) is shown in parentheses. WT, wild type. There were no significant differences of weights between pairedgenotypes. Wet tissue weights normalized to body weight also showed no significant differences between paired genotypes (data not shown).
Fig. 2. Identification of NKA �-isoform transgenic mice.A: PCR analysis. Primers targeting the �1-isoform transgeneyielded a 354-bp product, and primers targeting the �2-isoformtransgene yielded a 345-bp product. Linearized constructs of the�-isoform transgenes served as positive controls. H2O andwild-type (WT) mouse DNA served as negative controls (datanot shown). B: Southern blot. Probes containing �1- or �2-isoform cDNA hybridized to a 4-kb fragment of tail DNA thatwas digested with HindIII. The linearized �-isoform transgeneconstructs served as positive controls (data not shown), and WTmouse DNA served as negative controls.
H1175SMOOTH MUSCLE NKA AND BLOOD PRESSURE
AJP-Heart Circ Physiol • VOL 293 • AUGUST 2007 • www.ajpheart.org

were observed. Because of the small litter sizes in line 79�2sm� mice, studies in �2sm� mice were confined to line 84.There were no gross anatomic abnormalities in NKA trans-genic mice as measured by body and wet tissue weights of bothsmooth and nonsmooth muscle tissues (Table 1).
Coordinate expression of NKA �-isoforms. We next deter-mined the degree of NKA �-isoform expression at the proteinlevel using Western blot analysis (Table 2 and Fig. 3). Ourinitial focus was on vascular function; however, limitationsposed by tissue mass (aorta �1–2 mg) and fecundity of the�1-isoform line necessitated various approaches with respect toWestern blot analysis. We used gastric antrum (100–150 mg)for overall assessment of NKA and aorta where possible. The�1-isoform was increased by �1.4-fold in gastric antrum from�1sm� mice. Interestingly, the �2-isoform, which is not con-tained in the transgene in �1sm� mice, was also increased by1.6-fold. In the �2sm� mice, the �2-isoform was increased2.4-fold in aorta and 5.8-fold in gastric antrum compared withpaired WT mice. Similar to the �1sm� mice, we measured acoordinate increase in the �-isoform not contained in thetransgene in �2sm� mice, as the �1-isoform was elevated bytwofold in aorta and sevenfold in gastric antrum. Using anantibody recognizing all NKA �-isoforms, we obtained in-creases in total NKA of 1.5- and 3.7-fold in the �1sm� and
�2sm� lines, respectively. These differences in increases werenot statistically significant within each tissue type. The greaterdegree of increased �-isoform expression in the antrum than inthe aorta is consistent with previously reported findings that theSMP8 promoter has a greater expression in stomach than aorta(33). Therefore, both NKA transgenic lines (�1sm� and �2sm�
mice) exhibited increases in both the NKA �1- and �2-isoformsin smooth muscle with the �2sm� mouse line displaying agreater degree of increased NKA �-isoform expression thanthe �1sm� line.
Both NKA �1- and �2-isoforms, as well as total NKA(detected with an antibody recognizing all �-isoforms), weremeasured in hearts from �1sm� or �2sm� mice and theirrespective WT pairs. There were no statistically significantdifferences in �1- or �2-isoform protein levels in NKA trans-genic hearts compared with WT hearts (Table 2). These datasupport previous reports (6, 33) detailing the specificity of theSMP8 promoter to smooth muscle.
Because upregulation of the �-isoform not contained in thetransgene was observed at the protein level in NKA transgenicmice, we wanted to verify whether the coordinate upregulationoccurred at the transcriptional level as well. We measuredmRNA levels of the �-isoforms in gastric antrum from �1sm�
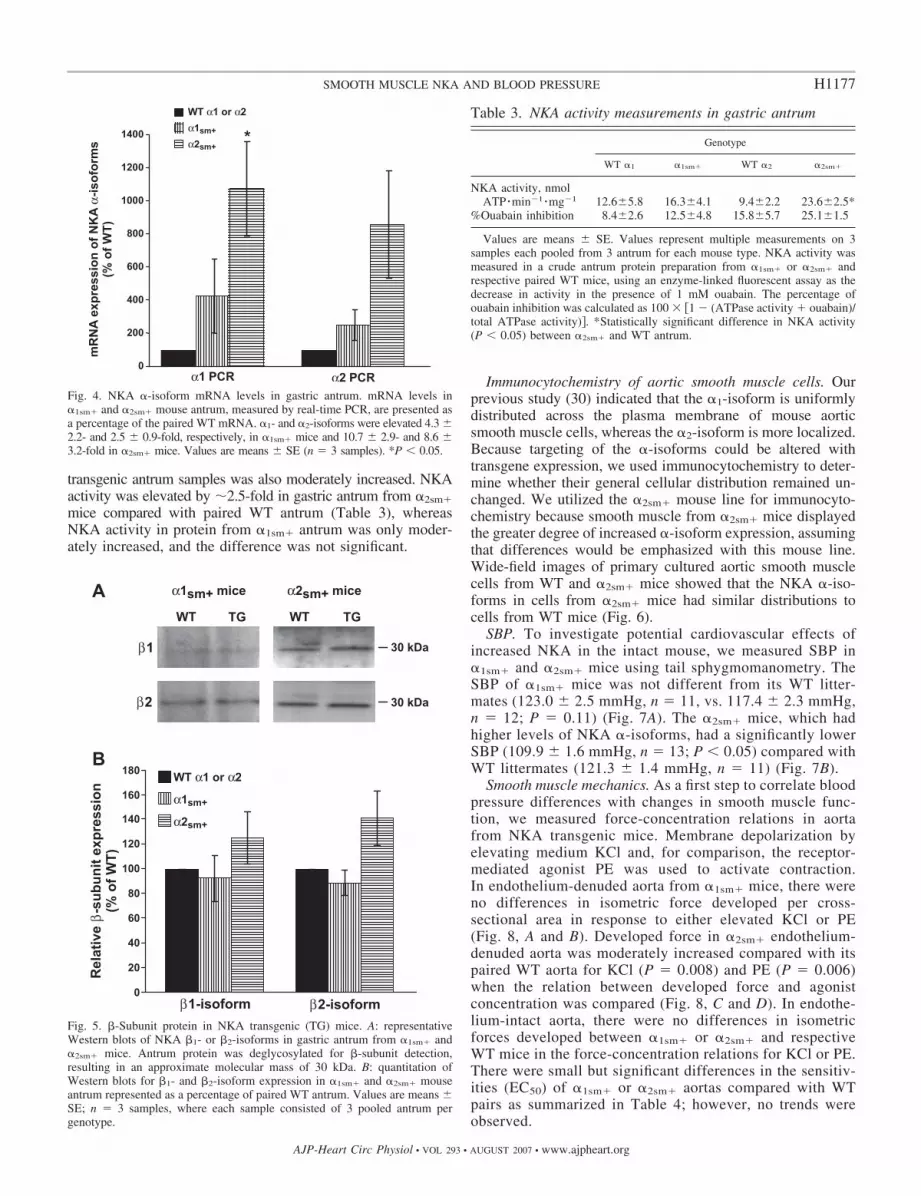
or �2sm� mice relative to respective paired WT antrum usingreal-time PCR analysis. Coordinate upregulation of the �-iso-forms was also evident at the mRNA level in NKA transgenicmice (Fig. 4). The mRNA for �1- and �2-isoforms was in-creased 4.3- and 2.5-fold, respectively, in �1sm� mice and10.7- and 8.6-fold in �2sm� mice.
Relative �-isoform levels. NKA is composed of a catalytic�-subunit (�112 kDa) and a glycosylated �-subunit (40–60kDa) (2, 4, 8, 16), which is required to make a functional NKAheterodimer (9). We measured relative protein levels of theNKA �1- and �2-isoforms in gastric antrum from NKA trans-genic mice (Fig. 5). In �1sm� mice, little differences in �1- or�2-isoforms were seen, with expression being 0.9 of WTlevels. In �2sm� mice, which expressed increased �-isoformsto a greater extent, the �1- and �2-isoforms were increased by1.3- and 1.4-fold, respectively.
NKA activity. To measure a functional increase in NKA in ourtransgenic mice, we measured NKA activity. The total enzymeactivity was increased in gastric antrum from both �1sm� and�2sm� mice, and the percentage of ouabain inhibition {100 [1 (ATPase activity � ouabain)/total ATPase activity]} measured in
Table 2. Summary of Western blot data of NKA �-isoforms
Tissue Genotype
Antibody
Anti-�1-isoform Anti-�2-isoform Anti-NKA
�1sm� antrum 1.4�0.1* (6) 1.6�0.3 (2) 1.5�0.2* (3)�2sm� antrum 7.0�1.6* (5) 5.8�1.2* (8) 3.7�1.5 (4)�1sm� heart 1.2�0.4 (4) 0.9�0.3 (4) 0.8�0.1 (3)�2sm� heart 1.0�0.2 (4) 1.0�0.2 (4) 1.2�0.1 (3)�2sm� aorta 2.0�0.1* (2) 2.4�0.4* (4) 3.8�1.5 (4)
Values are means � SE. Values in parentheses refer to no. of blots andhence different samples included in the analyses (see Western blot analysis inMETHODS). Western blot data are presented as fold-increases in �-isoformprotein expression in transgenic mouse smooth muscle tissues compared withpaired WT tissues with heart tissue as a nonsmooth muscle control. NKA,Na�-K�-ATPase. Analysis of antrum by 2-way ANOVA indicated that thevalues for the �1sm� and �2sm� mice were significantly different (P � 0.002).There were no differences between fold-increases for �1-, �2-, or total�-isoform. Similar 1-way analysis of aorta also indicated no statistical differ-ences between isoforms. There were no differences in between any cardiacmuscle class. *Fold difference from 1 (P � 0.05), which was assigned to WTgel density.
Fig. 3. NKA �-isoform protein in mouse antrum andaorta. Representative Western blots for the relativeNKA �-isoform contents in �1sm� (A) and �2sm� (B)mouse smooth muscle tissues compared with pairedWT tissue samples are shown. Actin was used as aloading control. Western blot data are summarized inTable 2.
H1176 SMOOTH MUSCLE NKA AND BLOOD PRESSURE
AJP-Heart Circ Physiol • VOL 293 • AUGUST 2007 • www.ajpheart.org
transgenic antrum samples was also moderately increased. NKAactivity was elevated by �2.5-fold in gastric antrum from �2sm�
mice compared with paired WT antrum (Table 3), whereasNKA activity in protein from �1sm� antrum was only moder-ately increased, and the difference was not significant.
Immunocytochemistry of aortic smooth muscle cells. Ourprevious study (30) indicated that the �1-isoform is uniformlydistributed across the plasma membrane of mouse aorticsmooth muscle cells, whereas the �2-isoform is more localized.Because targeting of the �-isoforms could be altered withtransgene expression, we used immunocytochemistry to deter-mine whether their general cellular distribution remained un-changed. We utilized the �2sm� mouse line for immunocyto-chemistry because smooth muscle from �2sm� mice displayedthe greater degree of increased �-isoform expression, assumingthat differences would be emphasized with this mouse line.Wide-field images of primary cultured aortic smooth musclecells from WT and �2sm� mice showed that the NKA �-iso-forms in cells from �2sm� mice had similar distributions tocells from WT mice (Fig. 6).
SBP. To investigate potential cardiovascular effects ofincreased NKA in the intact mouse, we measured SBP in�1sm� and �2sm� mice using tail sphygmomanometry. TheSBP of �1sm� mice was not different from its WT litter-mates (123.0 � 2.5 mmHg, n � 11, vs. 117.4 � 2.3 mmHg,n � 12; P � 0.11) (Fig. 7A). The �2sm� mice, which hadhigher levels of NKA �-isoforms, had a significantly lowerSBP (109.9 � 1.6 mmHg, n � 13; P � 0.05) compared withWT littermates (121.3 � 1.4 mmHg, n � 11) (Fig. 7B).
Smooth muscle mechanics. As a first step to correlate bloodpressure differences with changes in smooth muscle func-tion, we measured force-concentration relations in aortafrom NKA transgenic mice. Membrane depolarization byelevating medium KCl and, for comparison, the receptor-mediated agonist PE was used to activate contraction.In endothelium-denuded aorta from �1sm� mice, there wereno differences in isometric force developed per cross-sectional area in response to either elevated KCl or PE(Fig. 8, A and B). Developed force in �2sm� endothelium-denuded aorta was moderately increased compared with itspaired WT aorta for KCl (P � 0.008) and PE (P � 0.006)when the relation between developed force and agonistconcentration was compared (Fig. 8, C and D). In endothe-lium-intact aorta, there were no differences in isometricforces developed between �1sm� or �2sm� and respectiveWT mice in the force-concentration relations for KCl or PE.There were small but significant differences in the sensitiv-ities (EC50) of �1sm� or �2sm� aortas compared with WTpairs as summarized in Table 4; however, no trends wereobserved.
Fig. 4. NKA �-isoform mRNA levels in gastric antrum. mRNA levels in�1sm� and �2sm� mouse antrum, measured by real-time PCR, are presented asa percentage of the paired WT mRNA. �1- and �2-isoforms were elevated 4.3 �2.2- and 2.5 � 0.9-fold, respectively, in �1sm� mice and 10.7 � 2.9- and 8.6 �3.2-fold in �2sm� mice. Values are means � SE (n � 3 samples). *P � 0.05.
Fig. 5. �-Subunit protein in NKA transgenic (TG) mice. A: representativeWestern blots of NKA �1- or �2-isoforms in gastric antrum from �1sm� and�2sm� mice. Antrum protein was deglycosylated for �-subunit detection,resulting in an approximate molecular mass of 30 kDa. B: quantitation ofWestern blots for �1- and �2-isoform expression in �1sm� and �2sm� mouseantrum represented as a percentage of paired WT antrum. Values are means �SE; n � 3 samples, where each sample consisted of 3 pooled antrum pergenotype.
Table 3. NKA activity measurements in gastric antrum
Genotype
WT �1 �1sm� WT �2 �2sm�
NKA activity, nmolATP �min1 �mg1 12.6�5.8 16.3�4.1 9.4�2.2 23.6�2.5*
%Ouabain inhibition 8.4�2.6 12.5�4.8 15.8�5.7 25.1�1.5
Values are means � SE. Values represent multiple measurements on 3samples each pooled from 3 antrum for each mouse type. NKA activity wasmeasured in a crude antrum protein preparation from �1sm� or �2sm� andrespective paired WT mice, using an enzyme-linked fluorescent assay as thedecrease in activity in the presence of 1 mM ouabain. The percentage ofouabain inhibition was calculated as 100 �1 (ATPase activity � ouabain)/total ATPase activity) . *Statistically significant difference in NKA activity(P � 0.05) between �2sm� and WT antrum.
H1177SMOOTH MUSCLE NKA AND BLOOD PRESSURE
AJP-Heart Circ Physiol • VOL 293 • AUGUST 2007 • www.ajpheart.org

We evaluated the potential role of increased NKA �-isoformlevels on relaxation. Endothelium-dependent relaxation to AChin aorta from �1sm� or �2sm� mice was not different comparedwith respective WT mice (Fig. 9). We also tested �-adrenergicrelaxation in response to isoproterenol or the exogenous nitric
oxide donor, sodium nitroprusside, and found no differences inrelaxation-concentration relations in aorta from �2sm� micecompared with paired WT mice (data not shown). However,aorta from �2sm� mice had a small but significant decrease insensitivity (EC50) in response to ACh (Table 4).
Fig. 6. Immunocytochemical micrographs showing NKA�-isoforms in primary cultured aortic smooth muscle cells.Wide-field microscopic images were taken of cultured aorticsmooth muscle cells incubated with fluorescently labeledanti-�1- or anti-�2-antibodies. Left: cells from WT mice.Right: cells from �2sm� mice. The �2-isoform has a reticulardistribution in cells from �2sm� similar to those from WTaorta, whereas the �1-isoform maintained a wide-spreaddistribution in �2sm� cells similar to WT. Scale bars � 20�m.
Fig. 7. Systolic blood pressure (SBP) in NKA transgenic mice. SBP was measured using tail cuff sphygmomanometry (see METHODS). �1sm� and �2sm� micewere compared with their WT littermates (labeled WT �1 and WT �2, respectively). A: histogram of the daily SBP measurements in WT �1 and �1sm� mice.B: WT �2 and �2sm� mice fitted to Gaussian distributions. Black bars and lines represent transgenic data, and gray bars and lines represent WT data. SBP resultsin �1sm� mice was similar to those in WT �1 mice (122.7 � 2.5 mmHg, n � 11, vs. 117.4 � 2.3 mmHg, n � 12). The �2sm� mice had a significantly lowerSBP than the WT �2 mice (109.9 � 1.6 mmHg, n � 13, vs. 121.3 � 1.4 mmHg, n � 11). *P � 0.05.
H1178 SMOOTH MUSCLE NKA AND BLOOD PRESSURE
AJP-Heart Circ Physiol • VOL 293 • AUGUST 2007 • www.ajpheart.org

Smooth muscle kinetics in NKA transgenic mice. To assessany possible differences in the kinetics of contraction andrelaxation, we measured the half times for the aorta to achieve50% of its maximal contraction (peak) in response to either 50mM KCl or 1 �M PE and the recovery to 50% of the peakvalue after removal of the activating agent. Using a datastructure analogous to Table 5, we found no statisticallysignificant differences in half times of force development inresponse to either KCl or PE in either �1sm� or �2sm� micecompared with their respective WT pairs (n � 9–17; data notshown). In contrast to the half-time of force development, the
half-time for relaxation from a KCl-induced contraction of�2sm� endothelium-denuded aorta was smaller than its WT(P � 0.05, Table 5), indicating that the relaxation was faster.The relaxation half-time from a KCl-induced contraction for�1sm� endothelium-denuded aorta, which has a lesser degreeof increased �-isoform levels than the �2sm� line, was not sig-nificantly different (Table 5). In endothelium-intact aorta, nodifferences in relaxation half-times for KCl or PE contractureswere seen in �1sm� or �2sm� mice. Aortas with an intact endo-thelium relaxed faster than endothelium-denuded aortas, as wouldbe expected because of the endothelial contribution to relaxation.
Fig. 8. Developed force-concentration relations in endothelium-intact (�E) and endothelium-denuded (E) aorta from �1sm� and �2sm� mice. Developedisometric force was measured in aorta from �1sm� and paired WT mice (left) and �2sm� and respective WT aorta (right) in response to KCl (A and C) andphenylephrine (PE; B and D). Symbols are mean values, with bars representing �SE. For �1sm� (A and B, circles), there were no differences from either �Eaorta (solid symbols and lines) or E aorta (open symbols and dashed lines) and their respective WT (squares). For �2sm� (C and D), the �E aorta also didnot differ; however, the E aorta were statistically different from the WT.
Table 4. Summary of sensitivities (EC50) of �1sm� and �2sm� aorta to KCl, PE, or ACh
Genotype
�1 WT �1sm� �2 WT �2sm�
Endothelium intactKCl, mM 22.9�0.1 (7) 24.4�0.2* (7) 23.5�0.2 (10) 22.1�0.3* (11)PE, nM 130.0�7.5 (6) 128.0�6.4 (7) 117.0�8.6 (10) 82.4�6.6* (11)ACh, nM 37.2�9.3 (6) 43.2�5.6 (7) 29.6�5.9 (10) 52.1�6.0* (11)
Endothelium denudedKCl, mM 19.2�0.4 (7) 22.2�0.2* (5) 14.8�0.5 (6) 17.8�0.6* (9)PE, nM 99.3�11.7 (7) 112.0�15.4 (5) 62.3�6.7 (6) 55.9�4.9 (9)
Values for EC50 for aorta (with the endothelium intact or denuded) from �1sm� and �2sm� mice and their respective WT littermates are presented as means �SE, where n (no. of samples) is indicated in parentheses. *Statistically significant difference (P � 0.05, using Student’s t-test) in sensitivity in �1sm� or �2sm�
aorta compared with respective WT pairs.
H1179SMOOTH MUSCLE NKA AND BLOOD PRESSURE
AJP-Heart Circ Physiol • VOL 293 • AUGUST 2007 • www.ajpheart.org

DISCUSSION
The NKA is of known importance in blood pressure regu-lation, and alterations in NKA are hypothesized to underlieseveral disease conditions. However, there is little direct evi-dence on the effects attributable to vascular smooth muscleNKA per se on blood pressure. Thus we developed miceexpressing a transgene for either the NKA �1- or �2-isoformsusing the smooth muscle-specific promoter SMP8 (6). BothNKA �-isoform proteins increased by �1.5-fold in the �1sm�
mice, whereas the �2sm� mice had a greater degree of expres-sion, of up to 7-fold. The bases underlying differences in theextent of �-isoform expression in the transgenic lines is notknown but could be due to differences in transgene insertion orcopy number.
The first novel result was coordinate expression of the �1-and �2-isoforms in smooth muscle; that is, the �-isoformcontained in the transgene and the �-isoform not containedin the transgene were coordinately increased by approximatelythe same percentages. There is little available evidence on howthe levels of NKA are regulated. A coordinate increase in�-isoform expression could be due to a decrease in proteindegradation, transcriptional upregulation, or stabilization of thetranscript. Our real-time PCR data indicate that the coordinate�-isoform increases seen at the protein level are due in part toan increase in the RNA message for both isoforms.
There is substantial evidence indicating that the SMP8promoter is expressed only in smooth muscle (6, 33). This issupported by our results in hearts from NKA transgenic mice,which showed no change in �-isoform protein levels. Thus it isunlikely that the coordinate increase in �-isoforms is due togeneral changes in endocrine status or compensation, as thiswould have affected the �-isoform distribution of the heart.
NKA is reported to participate in several signaling pathwaysvia its modulation of [Ca2�]i (28). Alteration of NKA activityby ouabain can translate to activation of signaling proteins,such as NF-�B, which may lead to activation or deactivation ofexpression of various genes (28). This could underlie thecoordinate upregulation of the opposite �-isoform.
Importantly, the increases in NKA �-isoform levels insmooth muscle from NKA transgenic mice were accompaniedby increases in total NKA activity. Thus the increase in proteintranslated to increased NKA function. The measured increasein total NKA activity was less than the observed increase inprotein expression in �2sm� mice. One explanation is thatWestern blot and NKA activity measurements are two verydifferent parameters that require different sample preparationsand different measurement techniques, each with varying de-grees of sensitivity of detection, extent of nonspecific signals,and amplification, thus making absolute measurements notnecessarily directly comparable. The low NKA density in
Table 5. Relaxation half-times for �1sm� and �2sm� aorta in response to 50 mM KCl or 1 �M PE
Stimulation
Genotype
�1 WT �1sm� �2 WT �2sm�
KCl�Endo 74.8�5.6 (9) 72.2�5.6 (9) 60.8�5.3 (14) 71.5�6.3 (17)Endo 102.1�12.5 (9) 113.3�8.4 (8) 119.4�8.2 (10) 93.3�7.6* (15)
PE�Endo 37.5�3.1 (7) 30.5�4.0 (7) 37.6�6.2 (10) 40.7�3.5 (10)Endo 112.1�22.5 (7) 150.5�20.0 (6) 159.6�35.1 (8) 153.2�27.8 (11)
Values are means � SE, where n (no. of samples) is shown in parentheses. Half times are calculated as the time (in s) for an endothelium-intact (�Endo) oran endothelium-denuded (Endo) aorta to relax to 50% of the peak force after a rinse with physiological saline solution from a KCl- or PE-induced contraction.*Statistically significant difference (P � 0.05) in relaxation half-time between paired WT and �2sm� aorta.
Fig. 9. Force-concentration relations for aorta for the endothelium-dependent vasodilator ACh from �1sm� (A) or �2sm� (B) mice. �E aortas (solid symbols)from �1sm� or �2sm� mice did not display any differences in the relaxation-concentration relations. E aortas (open symbols) only had minimal vasorelaxationwith ACh. Symbols are mean values, with bars representing �SE.
H1180 SMOOTH MUSCLE NKA AND BLOOD PRESSURE
AJP-Heart Circ Physiol • VOL 293 • AUGUST 2007 • www.ajpheart.org

smooth muscle, 100-fold less than cardiac or skeletal muscle(3, 14), and limited tissue mass decrease the precision of NKAmeasurements in smooth muscle. Another possibility is theavailability of the �-subunit, which is required for NKAfunction (9). In smooth muscle from antrum, we measured littlechange in �-subunit levels from �1sm� mice, but �2sm� micehad a 25–40% increase in �-subunit protein levels. There is noavailable evidence on the absolute ratio of � to � in smoothmuscle to our knowledge. There are some indications that the�-subunit is synthesized in excess of the �-subunit (19, 21).The �-subunit could be a limiting factor to increasing the NKAactivity. However, the �-subunit is also reported to be requiredfor targeting of the �-isoform to the plasma membrane (9). Ourimmunomicrographs showed that the distributions of �1-and�2-isoforms in the �2sm� mice were similar to those in WTmice, which would argue against a �-subunit-limited NKAactivity.
Increased expression of NKA had a major impact on SBP.The �2sm� mice were hypotensive compared with WT litter-mates, whereas �1sm� mice were normotensive. Because totalNKA activity is substantially higher in the �2sm� mice than inthe �1sm� mice, the differences in SBP are likely a result of thedegree of NKA expression. A significant decrease in SBP isrelatively unusual in light of the generally small SBP changesin the literature associated with a single gene alteration (29).Our data support the hypothesis that increased NKA activityresults in a decreased SBP and moreover that this can be solelya vascular phenomenon. Thus our gain-of-function experi-ments complement studies of decreasing NKA activity (7, 11,36), which suggest that the vascular NKA alone could be animportant factor in hypertension.
To evaluate the potential changes in vascular smooth musclethat may underlie the decreased blood pressure, we measuredcontractility and relaxation parameters in aorta. Consistentwith the absence of change in blood pressure, there were nodifferences in contractility to KCl or PE stimulation in aortasfrom the �1sm� mice. The �2sm� aortas had moderately in-creased force in response to KCl (15–20%) or PE stimulation(12–15%). Higher forces are seemingly the opposite of thepredicted increased Ca2� extrusion by NKA-NCX coupling.
One might have anticipated that the increase in NKA and fallin blood pressure would both be accompanied by a decrease inaortic smooth muscle contractility. There are several possibil-ities behind the increase in force seen in endothelium-denuded�2sm� aorta. 1) The increased Na� gradient could stimulateNa�/H� exchange, leading to cytosolic alkalinization, whichcan lead to higher levels of contractility (20). 2) The relativefunctional roles of �1 to �2 may be important. A modificationof the hypothesis of coupled NKA-NCX Ca2� extrusion hasbeen proposed based on the differential distribution (Fig. 6) of�-isoforms. Blaustein and colleagues (1, 10) proposed that�2-NKA and NCX are localized in plasma membrane microdo-mains with preferential access to Ca2� loading of the sarco-plasmic reticulum (SR). Increases in the �2-isoform in thiscompartment would lower SR loading, which could outweighthe effects of �1 on global [Ca2�]i. For example, a hypoloadedSR could lead to lower spontaneous SR Ca2� release andconsequently lower KCa channel activity, which would lead todepolarization and greater contractility (35). 3) Another possi-bility to consider is the role of NKA in signaling. NKA ispostulated to lead to possible activation of signal transduction
factors like NF-�B or Src (28). This may lead to the upregu-lation and downregulation of genes related to and/or unrelatedto ion transport and/or contractility, thereby resulting inchanges in aortic contractility, which remains to be elucidated.
One can also envision that either decreases or increases incontractile force, depending on which circulation (renal, cen-tral nervous system, etc.), could lead to reduced blood pressure.In some ways, it would be rather remarkable if changes inaortic smooth muscle reflected every microcirculation. How-ever, this was the simplest hypothesis and important to test.The increases in contractile forces in �2sm� mice were onlymeasured in endothelium-denuded aorta. Endothelium-intactaorta, a perhaps more physiological representative of thein vivo vessel, did not display any differences in contractility,suggesting some basal endothelial compensation. It is possiblethat the compensation is more effective in other microcircula-tions or at the resistance vessel level or that other microvas-culatures simply differ from aorta. Understanding the mecha-nism(s) underlying the decrease in blood pressure will requirefuture studies.
Importantly, mechanical factors other than the developedforce may be in play in the observed hypotension. Of particularinterest, there is a correlation between speed of relaxation andblood pressure. Studies of spontaneously hypertensive ratsreport slower relaxation rates in caudal and mesenteric arteries(23, 24). We measured relaxation half-times in �1sm� and�2sm� mice. The �2sm� aortas had faster rates of relaxation,indicating possible increased Ca2� extrusion. A decreasedblood pressure associated with faster aortic relaxation wasrecently reported for diabetic rats (27). We would suggest thatthe observed faster rates of aortic relaxation may be a largerfactor leading to the observed hypotension rather than theeffects of the moderately increased maximum force.
In summary, our results indicate that a genetically engi-neered increase in NKA activity is associated with a decreasedSBP, which is dependent on the degree of vascular NKAexpression. Thus focusing on vascular NKA per se may pro-vide novel insights for treatment of hypertension.
ACKNOWLEDGMENTS
We thank Dr. Jerry Lingrel for kindly providing us with the rat Na�-K�-ATPase �1- and �2-isoform cDNAs and Dr. Arthur Strauch for the SMP8promoter.
GRANTS
This work was supported by National Heart, Lung, and Blood InstituteGrant HL-66044 (R. J. Paul and R. M. Lynch).
REFERENCES
1. Arnon A, Hamlyn JM, Blaustein MP. Ouabain augments Ca2� tran-sients in arterial smooth muscle without raising cytosolic Na�. Am JPhysiol Heart Circ Physiol 279: H679–H691, 2000.
2. Aydemir-Koksoy A. Na-ATPase: a review. J Ankara Med Sch 24: 73–82,2002.
3. Aydemir-Koksoy A, Allen JC. Regulation of Na� pump expression byvascular smooth muscle cells. Am J Physiol Heart Circ Physiol 280:H1869–H1874, 2001.
4. Blanco G, Mercer RW. Isozymes of the Na-K-ATPase: heterogeneity instructure, diversity in function. Am J Physiol Renal Physiol 275: F633–F650, 1998.
5. Blaustein MP, Zhang J, Chen L, Hamilton BP. How does salt retentionraise blood pressure? Am J Physiol Regul Integr Comp Physiol 290:R514–R523, 2006.
H1181SMOOTH MUSCLE NKA AND BLOOD PRESSURE
AJP-Heart Circ Physiol • VOL 293 • AUGUST 2007 • www.ajpheart.org

6. Cogan JG, Sun S, Stoflet ES, Schmidt LJ, Getz MJ, Strauch AR.Plasticity of vascular smooth muscle alpha-actin gene transcription. Char-acterization of multiple, single-, and double-strand specific DNA-bindingproteins in myoblasts and fibroblasts. J Biol Chem 270: 11310–11321,1995.
7. Dostanic-Larson I, Lorenz JN, Van Huysse JW, Neumann JC, Mose-ley AE, Lingrel JB. Physiological role of the �1- and �2-isoforms of theNa�-K�-ATPase and biological significance of their cardiac glycosidebinding site. Am J Physiol Regul Integr Comp Physiol 290: R524–R528,2006.
8. Ewart HS, Klip A. Hormonal regulation of the Na�-K�-ATPase: mech-anisms underlying rapid and sustained changes in pump activity. Am JPhysiol Cell Physiol 269: C295–C311, 1995.
9. Fambrough DM, Lemas MV, Hamrick M, Emerick M, Renaud KJ,Inman EM, Hwang B, Takeyasu K. Analysis of subunit assembly of theNa-K-ATPase. Am J Physiol Cell Physiol 266: C579–C589, 1994.
10. Golovina V, Song H, James P, Lingrel J, Blaustein M. Regulation ofCa2� signaling by Na� pump �-2 subunit expression. Ann NY Acad Sci986: 509–513, 2003.
11. Haddy FJ. Role of dietary salt in hypertension. Life Sci 79: 1585–1592,2006.
12. Huddleson JP, Srinivasan S, Ahmad N, Lingrel JB. Fluid shear stressinduces endothelial KLF2 gene expression through a defined promoterregion. Biol Chem 385: 723–729, 2004.
13. Iwamoto T, Kita S. Hypertension, Na�/Ca2� exchanger, and Na-ATPase.Kidney Int 69: 2148–2154, 2006.
14. Kjeldsen K, Bjerregaard P, Richter EA, Thomsen PE, Norgaard A.Na�,K�-ATPase concentration in rodent and human heart and skeletalmuscle: apparent relation to muscle performance. Cardiovasc Res 22:95–100, 1988.
15. Lalli J, Harrer JM, Luo W, Kranias EG, Paul RJ. Targeted ablation ofthe phospholamban gene is associated with a marked decrease in sensi-tivity in aortic smooth muscle. Circ Res 80: 506–513, 1997.
16. Lingrel JB, Kuntzweiler T. Na-ATPase. J Biol Chem 269: 19659–19662, 1994.
17. Lynch RM, Fogarty KE, Fay FS. Modulation of hexokinase associationwith mitochondria analyzed with quantitative three-dimensional confocalmicroscopy. J Cell Biol 112: 385–395, 1991.
18. McDonough AA, Velotta JB, Schwinger RH, Philipson KD, FarleyRA. The cardiac sodium pump: structure and function. Basic Res Cardiol97, Suppl 1: I19–I24, 2002.
19. Mircheff AK, Bowen JW, Yiu SC, McDonough AA. Synthesis andtranslocation of Na�-K�-ATPase �- and �-subunits to plasma membranein MDCK cells. Am J Physiol Cell Physiol 262: C470–C483, 1992.
20. Nagesetty R, Paul RJ. Effects of pHi on isometric force and [Ca2�]i inporcine coronary artery smooth muscle. Circ Res 75: 990–998, 1994.
21. Orlowski J, Lingrel JB. Tissue-specific and developmental regulation ofrat Na,K-ATPase catalytic � isoform and � subunit mRNAs. J Biol Chem263: 10436–10442, 1988.
22. Otsu K, Kuruma A, Yanagida E, Shoji S, Inoue T, Hirayama Y,Uematsu H, Hara Y, Kawano S. Na�/K� ATPase and its functionalcoupling with Na�/Ca2� exchanger in mouse embryonic stem cells duringdifferentiation into cardiomyocytes. Cell Calcium 37: 137–151, 2005.
23. Packer CS, Stephens NL. Mechanics of caudal artery relaxation incontrol and hypertensive rats. Can J Physiol Pharmacol 63: 209–213,1985.
24. Packer CS, Stephens NL. Prolonged isobaric relaxation time in smallmesenteric arteries of the spontaneously hypertensive rat. Can J PhysiolPharmacol 65: 230–235, 1987.
25. Pfaffl MW.A new mathematical model for relative quantification inreal-time RT-PCR. Nucleic Acids Res 29: e45, 2001.
26. Pressley TA. Phylogenetic conservation of isoform-specific regionswithin alpha-subunit of Na�-K�-ATPase. Am J Physiol Cell Physiol 262:C743–C751, 1992.
27. Rinaldi GJ. Blood pressure fall and increased relaxation of aortic smoothmuscle in diabetic rats. Diabetes Metab 31: 487–495, 2005.
28. Scheiner-Bobis G, Schoner W. A fresh facet for ouabain action. Nat Med7: 1288–1289, 2001.
29. Schnermann J, Lorenz JN, Briggs JP. Cardiorenal physiology of themouse: from loss of genome to gain of phenome. In: The Molecular andGenetic Basis of Renal Disease, edited by Pollak M and Mount DB. Inpress.
30. Shelly DA, He S, Moseley A, Weber C, Stegemeyer M, Lynch RM,Lingrel J, Paul RJ. Na� pump �2-isoform specifically couples to con-tractility in vascular smooth muscle: evidence from gene-targeted neonatalmice. Am J Physiol Cell Physiol 286: C813–C820, 2004.
31. Shull GE, Greeb J, Lingrel JB. Molecular cloning of three distinct formsof the Na�,K�-ATPase �-subunit from rat brain. Biochemistry 25: 8125–8132, 1986.
32. Sweeney G, Klip A. Regulation of the Na�/K�-ATPase by insulin: whyand how? Mol Cell Biochem 182: 121–133, 1998.
33. Wang J, Niu W, Nikiforov Y, Naito S, Chernausek S, Witte D,LeRoith D, Strauch A, Fagin JA. Targeted overexpression of IGF-Ievokes distinct patterns of organ remodeling in smooth muscle cell tissuebeds of transgenic mice. J Clin Invest 100: 1425–1439, 1997.
34. Wasserstrom JA, Aistrup GL. Digitalis: new actions for an old drug.Am J Physiol Heart Circ Physiol 289: H1781–H1793, 2005.
35. Wellman GC, Nelson MT. Signaling between SR and plasmalemma insmooth muscle: sparks and the activation of Ca2�-sensitive ion channels.Cell Calcium 34: 211–229, 2003.
36. Zhang J, Lee MY, Cavalli M, Chen L, Berra-Romani R, Balke CW,Bianchi G, Ferrari P, Hamlyn JM, Iwamoto T, Lingrel JB, MattesonDR, Wier WG, Blaustein MP. Sodium pump �2 subunits control myo-genic tone and blood pressure in mice. J Physiol 569: 243–256, 2005.
H1182 SMOOTH MUSCLE NKA AND BLOOD PRESSURE
AJP-Heart Circ Physiol • VOL 293 • AUGUST 2007 • www.ajpheart.org
Related Documents











