The FASEB Journal express article10.1096/fj.02-0937fje. Published online July 18, 2003. Mechanisms of ouabain toxicity Raphael C. Valente,* Luiz S. Capella, † Robson Q. Monteiro,* Vivian M. Rumjanek,* Aníbal G. Lopes, ‡ and Márcia A.M. Capella* ,‡ *Departamento de Bioquímica Médica, Instituto de Ciências Biomédicas, Universidade Federal do Rio de Janeiro (UFRJ); † Departamento de Clínica Médica - Universidade do Rio de Janeiro (UNI-RIO); ‡ Instituto de Biofísica Carlos Chagas Filho, Universidade Federal do Rio de Janeiro (UFRJ) Rio de Janeiro, Brazil Corresponding author: Márcia A.M. Capella, Instituto de Biofísica Carlos Chagas Filho - Universidade Federal do Rio de Janeiro, CCS - Bloco G 21949-900 Rio de Janeiro, RJ, Brazil. E- mails: [email protected]; [email protected] ABSTRACT The suggested involvement of ouabain in hypertension raised the need for a better understanding of its cellular action, but the mechanisms of ouabain toxicity are only now being uncovered. In the present study, we show that reduced glutathione (GSH) protected ouabain-sensitive (OS) cells from ouabain-induced toxicity and that the inhibition of GSH synthesis by D,L-buthionine- (S,R)-sulfoximine (BSO) sensitized ouabain-resistant (OR) cells. We could not observe formation of • OH or H 2 O 2 , but there was an increase in O 2 • ¯ only in OS cells. Unexpectedly, an increased number of OR cells depolarized after treatment with ouabain, and BSO blocked this depolarization. Moreover, GSH increased ouabain-induced depolarization in OS cells. A sustained increase in tyrosine phosphorylation (P-Tyr) and Ras expression was observed after treatment of OS cells, and GSH prevented it. Conversely, BSO induced P-Tyr and Ras expression in ouabain-treated OR cells. The results obtained have three major implications: There is no direct correlation between membrane depolarization and ouabain-induced cell death; ouabain toxicity is not directly related to its classical action as a Na + , K + -ATPase inhibitor but seems to be associated to signal transduction, and GSH plays a major role in preventing ouabain- induced cell death. Key words: hypertension • glutathione • reactive oxygen species • apoptosis • necrosis a + , K + -ATPase is essential for the maintenance of the membrane potential of living cells and for the transport of organic solutes by sodium symporters in epithelial cells. Also, it indirectly participates in the regulation of intracellular pH and Ca 2+ concentration. The prolonged inhibition of this pump may lead to several alterations in ion transport and cell volume, compromising cell homeostasis. One of the most potent inhibitors of the Na + , K + -ATPase is the cardiotonic steroid ouabain. The partial inhibition of cardiac myocyte Na + , K + -ATPases by ouabain and other related glycosides regulates contractility, and this is the basis of the major therapeutic use of these drugs in the management of congestive heart failure (1). However, the same nontoxic concentrations of ouabain that increase contractility also stimulate the nonproliferative growth (hypertrophy) of N

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

The FASEB Journal express article10.1096/fj.02-0937fje. Published online July 18, 2003.
Mechanisms of ouabain toxicity Raphael C. Valente,* Luiz S. Capella,† Robson Q. Monteiro,* Vivian M. Rumjanek,* Aníbal G. Lopes,‡ and Márcia A.M. Capella*,‡
*Departamento de Bioquímica Médica, Instituto de Ciências Biomédicas, Universidade Federal do Rio de Janeiro (UFRJ); †Departamento de Clínica Médica - Universidade do Rio de Janeiro (UNI-RIO); ‡Instituto de Biofísica Carlos Chagas Filho, Universidade Federal do Rio de Janeiro (UFRJ) Rio de Janeiro, Brazil
Corresponding author: Márcia A.M. Capella, Instituto de Biofísica Carlos Chagas Filho - Universidade Federal do Rio de Janeiro, CCS - Bloco G 21949-900 Rio de Janeiro, RJ, Brazil. E-mails: [email protected]; [email protected]
ABSTRACT
The suggested involvement of ouabain in hypertension raised the need for a better understanding of its cellular action, but the mechanisms of ouabain toxicity are only now being uncovered. In the present study, we show that reduced glutathione (GSH) protected ouabain-sensitive (OS) cells from ouabain-induced toxicity and that the inhibition of GSH synthesis by D,L-buthionine-(S,R)-sulfoximine (BSO) sensitized ouabain-resistant (OR) cells. We could not observe formation of •OH or H2O2, but there was an increase in O2
•¯ only in OS cells. Unexpectedly, an increased number of OR cells depolarized after treatment with ouabain, and BSO blocked this depolarization. Moreover, GSH increased ouabain-induced depolarization in OS cells. A sustained increase in tyrosine phosphorylation (P-Tyr) and Ras expression was observed after treatment of OS cells, and GSH prevented it. Conversely, BSO induced P-Tyr and Ras expression in ouabain-treated OR cells. The results obtained have three major implications: There is no direct correlation between membrane depolarization and ouabain-induced cell death; ouabain toxicity is not directly related to its classical action as a Na+, K+-ATPase inhibitor but seems to be associated to signal transduction, and GSH plays a major role in preventing ouabain-induced cell death.
Key words: hypertension • glutathione • reactive oxygen species • apoptosis • necrosis
a+, K+-ATPase is essential for the maintenance of the membrane potential of living cells and for the transport of organic solutes by sodium symporters in epithelial cells. Also, it indirectly participates in the regulation of intracellular pH and Ca2+ concentration. The
prolonged inhibition of this pump may lead to several alterations in ion transport and cell volume, compromising cell homeostasis.
One of the most potent inhibitors of the Na+, K+-ATPase is the cardiotonic steroid ouabain. The partial inhibition of cardiac myocyte Na+, K+-ATPases by ouabain and other related glycosides regulates contractility, and this is the basis of the major therapeutic use of these drugs in the management of congestive heart failure (1). However, the same nontoxic concentrations of ouabain that increase contractility also stimulate the nonproliferative growth (hypertrophy) of
N

cardiac myocytes (2). Ouabain may also lead to adverse consequences in the kidney, where the chronic inhibition of Na+, K+-ATPase impairs renal hemodynamics and pressure natriuresis (3).
Although plasma membrane Na+, K+-ATPases are the only recognized receptors for ouabain, recent evidence suggests the existence of other functional receptors. A mammalian analog of ouabain, namely endogenous ouabain or ouabain-like factor, has been described and shown to circulate in elevated concentrations in many patients with essential hypertension (4, 5). Prolonged infusions with ouabain, but not digoxin, induce hypertension in rats, even though both drugs are near equipotent inhibitors of rat Na+, K+-ATPase (6). The same authors observed that the hypertensinogenic potency of a series of ouabain analogs is inversely correlated with their efficacy as Na+, K+-ATPase inhibitors (7). More recently, Ward et al. (8) showed, for the first time, that adrenocortical cells probably possess ouabain receptors that are distinct from Na+, K+-ATPase and suggested that these novel receptors may be involved in the regulation and/or secretion of endogenous ouabain.
In cultured cells, a prolonged ouabain blockade of Na+, K+-ATPase leads cells to loose K+, depolarize, detach from each other and from the substrate, and die (9, 10). Although the sensitivity to ouabain is cell specific, it was commonly believed that cellular resistance to ouabain was due to the presence of Na+, K+-ATPase isoforms with low affinity for this glycoside. However, in a series of related papers, Contreras et al. (10–12) showed that Ma104, an epithelial kidney cell line, is resistant to ouabain, although its Na+, K+-ATPase has high affinity for this drug, thus suggesting that ouabain toxicity is not directly related to Na+, K+-ATPase inhibition. Other recent studies (13–15) demonstrated that ouabain induces the formation of reactive oxygen species (ROS) in cardiomyocytes, pancreatic islets cells, and several cell lines, and that this induction probably occurs by a pathway parallel to the inhibition of the Na+, K+-ATPase transport activity. These findings led us to hypothesize that differences in sensitivity to ouabain could be related to differences in cellular sensitivity to ROS. In a recent paper, we showed that reduced glutathione (GSH) protected ouabain-sensitive MDCK cells from ouabain-induced toxicity and that the inhibition of GSH synthesis by BSO sensitized the ouabain-resistant Ma104 cells (16). These results suggest that at least part of ouabain toxicity may be related to ROS. However, the mechanisms of the ouabain-induced cytotoxicity, as well as the mechanisms of GSH protection, are not yet understood, and the comprehension of these phenomena may contribute to the understanding of the pathogenesis of ouabain-induced hypertension.
The purpose of the present work was to study the mechanisms underlying ouabain toxicity in two kidney cell lines previously shown to have different susceptibilities to ouabain: MDCK, an ouabain-sensitive cell line, and the ouabain-resistant Ma104 cells (10–12, 16). For that, ouabain-induced plasma membrane depolarization, increase in tyrosine phosphorylation, and induction of Ras expression were analyzed. The role of ROS and GSH in the ouabain toxicity was also analyzed.
MATERIALS AND METHODS
Cells and culture conditions
The epithelial cell lines Ma104 (obtained from Rhesus monkey kidney) and MDCK (derived from canine kidney) were grown in Dulbecco’s modified eagle medium (DMEM) (Gibco-BRL,

Gaithersburg, MD) with penicillin and streptomycin (Gibco) and supplemented with 10% fetal bovine serum (Gibco) in disposable plastic bottles, at 37°C in a CO2 incubator, until reaching confluence.
Treatment with ouabain and analysis of cellular viability
Ma104 and MDCK cells were seeded in 24-well plates, at a concentration of 1×105 cells/ml (1 ml/well). All experiments were performed 24 h after seeding, to ensure uniform attachment of cells at the onset of the experiments. Cells were then incubated for 24 h with ouabain concentrations varying from 10−8 M to 10−3 M. After incubation, the cells were washed twice with phosphate-buffered saline (PBS) to remove detached cells. The remaining attached cells were harvested by trypsinization, and the cellular viability was measured by trypan blue exclusion. In some experiments, reduced GSH (Calbiochem, San Diego, CA) and D,L-buthionine-(S,R)-sulfoximine (BSO) (Sigma, St. Louis, MO) were added at a final concentration of 2 mM.
Induction of apoptosis by ouabain
The number of apoptotic cells was assessed by AnnexinV-FITC kit (PharMingen). MDCK cells were treated with ouabain at different concentrations and incubation times, or left untreated. After treatments, cells were washed twice with PBS, harvested by trypsinization, centrifuged, and suspended in 100 µl of a solution containing Annexin V-FITC and propidium iodide (PI) (PharMingen). After incubation for 15 min at room temperature in the dark, 400 µl of binding buffer were added. The two-color flow cytometry analysis was performed on a FACSCalibur device (Becton Dickinson, Franklin Lakes, NJ).
Analysis of plasma membrane potential
Changes in plasma membrane potential from either Ma104 or MDCK cells were measured by flow cytometry, using a bis-oxonol dye, DiBAC4(3) (Molecular Probes, Eugene, OR), as described by Bortner et al. (17). Cells were preincubated with 2 µM DiBAC4(3) for 15 min to reach equilibrium. MDCK cells were subsequently incubated with 10µM ouabain in the presence or absence of 2 mM GSH. Ma104 cells were treated with the same ouabain concentration in the presence or absence of 2 mM BSO, which was added to the medium 24 h before the beginning of the experiment and was maintained in the medium during the incubation time. Both cell lines were incubated with ouabain for 30 min at 37°C. After treatments, cells were washed twice with PBS, harvested by trypsinization, and kept on ice until flow cytometry analysis. Fluorescence intensities were measured through a 530 nm long pass filter. After acquisition of 104 living cells, a limit of fluorescence intensity (M1, Fig. 1) was established. Cells within M1 region, presenting higher fluorescence intensity than controls, were considered depolarized.
Immunofluorescence techniques
Cells grown on coverslips were treated with ouabain (1–100 µM) for 3, 6, or 24 h in the presence or absence of 2 mM GSH (MDCK) or 2 mM BSO (Ma104).

After incubation, cells were washed three times with PBS, fixed, and permeabilized with cold methanol for 3 min, followed by incubation with 10% fetal bovine serum for 30 min at room temperature. Next, cells were incubated overnight in a humidified chamber with one of the following primary mouse monoclonal antibodies: pan Ras (F132), which recognizes antigenic determinants common to H-Ras, K-Ras, and N-Ras, or P-Tyr (PY20), which specifically detects phosphotyrosine-containing proteins and does not cross-react with phosphoserine or phosphothreonine. Both antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA). After washing with PBS, cells were incubated with Alexa Fluor 546 anti-mouse secondary antibody (Molecular Probes) for 1 h at room temperature in the dark. Cells were then washed three times with PBS and once with water, and the coverslips were mounted in glass slides with 50% glycerol in PBS. The fluorescence was visualized under an Axiovert 100 microscope (Zeiss, Munich, Germany), using a 40× fluar oil objective (Zeiss). Once collected, the images were analyzed using the software VIDAS (Kontron Elektronic GMBH). Three to five images obtained from different fields of the same coverslip were used and the average of pixels above background was measured. Four different experiments were performed for each cell line and for each antibody to obtain the mean and standard deviations presented in each figure.
Fluorescence measurements of ouabain-induced ROS generation
The leuco probe dihydrorhodamine 123 (DHR) (Molecular Probes), which preferentially reacts with H2O2 and is oxidized into the fluorescent dye rhodamine 123, was used to assess H2O2 production. Both cell lines were preincubated for 15 min with 1 µM DHR and then treated with 1 mM H2O2 or ouabain for 30 min at 37°C. Next, cells were washed twice in PBS, harvested by trypsinization, and kept on ice. Flow cytometry analyses were performed as described previously for plasma membrane potential measurements.
Detection of O2•¯ was evaluated using the probe proxyl-fluorescamine (P-Fluo) (Molecular
Probes), which reacts directly with O2•¯, resulting in an increase of its fluorescence. This probe
also reacts with the methyl radical generated by the interaction of •OH with dimethyl sulfoxide (DMSO) (18). Hence, for •OH measurements, DMSO was added to the medium at a concentration of 5% (v/v). P-Fluo was preincubated for 15 min, and the reactions were initiated by the addition of 10 µM ouabain in the presence or absence of DMSO, at a final volume of 200 µl, and continued for up to 30 min at 37°C. At each incubation time, the cells were harvested by adding 800 µl of cold PBS and streaming vigorously. The suspension was then analyzed in a Hitachi F4500 fluorescence spectrophotometer. Excitation and emission wavelengths were set at 393 nm and 482 nm, respectively.
Statistical analysis
Each experiment was repeated three to five times. Data are expressed as means ±SD of the mean and were analyzed using Student’s t test for comparison of the differences. Values of P<0.05 were considered statistically significant.

RESULTS
Sensitivity of MDCK and Ma104 cells to ouabain
It has been shown that Ma104 cells are resistant to ouabain and that the viability of MDCK cells falls acutely after incubation with 1 µM ouabain (10, 16). Therefore, we tried to verify whether incubation of cells with this glycoside would lead to similar results under our experimental conditions. As shown in Figure 1, ouabain changed the morphology of MDCK and Ma104 cells. A massive detachment and the appearance of rounded, swelled, and floating MDCK cells could be observed, while Ma104 cells remained attached even with a ouabain concentration 10 times greater, although undergoing some morphologic alterations. We also observed that the growth of Ma104 cells was inhibited by ouabain concentrations ranging from 1 to 100 µM, but the majority of the cells remained viable, because when the medium was replaced by ouabain-free medium, the growth restarted (data not shown), confirming the results previously reported (10, 11).
The viability of ouabain-treated Ma104 and MDCK cells can be seen in Figure 2. Less than 20% of MDCK cells remained viable when treated with 1 µM ouabain, a concentration 1000 times greater was necessary to lead Ma104 cells to this same viability. Therefore, to compare the effects of ouabain in these two cell lines, ouabain concentrations of 1 and 10 µM were used in the majority of the subsequent experiments, and, in some cases, a concentration of 100 µM was used for Ma104 cells.
Induction of apoptosis and necrosis
It has been observed that ouabain-induced MDCK cell death occurred by necrosis (12, 19). To verify whether ouabain induced necrosis and/or apoptosis under our experimental conditions, MDCK cells were treated with various concentrations of ouabain for different incubation times and then labeled with the probes Annexin V-FITC and PI, which evaluate apoptosis and necrosis, respectively. Cells entering apoptosis expose outward their plasma membrane phosphatidylserine residues, which in normal conditions are only found in the inner plasma membrane layer. The fluorescent dye Annexin-V FITC binds these residues, increasing cellular fluorescence. However, PI enters only cells that lack membrane integrity, a characteristic of necrosis.
Confirming previous observations (12, 19), ouabain increased retention of PI, but did not enhance Annexin V-FITC labeling of MDCK cells, suggesting that cell death occurred by necrosis (Fig. 3). Experiments were also performed with other ouabain concentrations and incubation times, and there were still no significant increase in Annexin V-FITC labeling (Table 1).
Generation of ROS
It has been demonstrated that ouabain induces the formation of ROS in cardiomyocytes and other cell types, which reaches a plateau in ∼30 min (13–15). This observation points to a possible relationship between ROS and the toxic effects of ouabain. Nevertheless, due to the lack of specificity of the probe used by those authors, it was not possible to recognize which radical was being produced. Thus, in the present report, the fluorescent probes DHR and P-Fluo were used to assess ROS generation. DHR is a leuco probe that reacts with H2O2 and also with peroxinitrite,

converting into the fluorescent dye rhodamine 123. P-Fluo reacts directly with O2•¯ and
indirectly with •OH, the latter needing the presence of DMSO in the medium.
No ROS production could be seen in Ma104 cells treated with 10 µM ouabain (data not shown). In contrast, as shown in Figure 4, a significant increase in O2
•¯ (14%) was noticed in MDCK cells treated with 10 µM ouabain for 30 min, although there is no enhanced formation of •OH. The generation of O2
•¯ could be seen in the first 5 min of ouabain treatment (data not shown) but was greater at 30 min. Although, according to the manufacturer, this probe is impermeable to cell membrane, we noticed a cellular fluorescence in MDCK cells after ouabain treatment (Fig. 4, inset). No fluorescence was detected in control cells, suggesting that alterations in cell membrane permeability due to ouabain treatment cause leaking of the dye into the cell.
No production of H2O2 could be seen in 30 min of incubation with 10 µM ouabain in MDCK cells (Fig. 5). This was also verified for incubation times varying from 5 to 60 min (data not shown). In addition, Ma104 cells did not present any increase in H2O2 production (data not shown).
Effect of SOD, catalase and GSH, on the sensitivity of MDCK cells to ouabain
The previous results suggest that at least superoxide anion is being produced in MDCK cells by ouabain treatment. Therefore, we tested whether the main cellular antioxidant enzymes catalase and superoxide dismutase (SOD) and the major nonenzymatic cellular antioxidant GSH would be able to protect MDCK cells from ouabain toxicity. As shown in Figure 6, GSH protected MDCK cells from ouabain-induced toxicity. Moreover, BSO, an inhibitor of GSH synthesis, sensitized the ouabain-resistant Ma104 cells. On the other hand, SOD (in concentrations ranging from 10 to 40 µg/ml) and catalase (varying from 10 to 200 µg/ml) did not significantly alter the sensitivity of MDCK cells to ouabain.
Effect of GSH and BSO on ouabain-induced plasma membrane depolarization
It is well known that the blockage of Na+, K+-ATPase by ouabain leads to plasma membrane depolarization (PMD), and this, in turn, has been associated with cell death (17, 20–22). Thus, PMD was evaluated in MDCK and Ma104 cells treated with ouabain in the presence or absence of GSH and BSO, respectively. To assess this information, cells were treated for 30 min with ouabain in the presence of DiBAC4(3) and GSH (MDCK) or after pretreatment for 24 h with BSO (Ma104). The anionic oxonol dye DiBAC4(3) enters cells upon PMD, with a consequent increase in cellular fluorescence intensity. Preliminary experiments showed no alteration in DiBAC4(3) fluorescence in cells treated with CCCP, which alter the membrane potential of mitochondria, indicating the specificity of DiBAC4(3) in measuring only changes in the plasma membrane potential (data not shown).
Representative experiments of ouabain-induced PMD in MDCK (upper panel) and Ma104 (lower panel) cells are shown in Figure 7. Insets show the means and standard deviations of the percentage of cells gated in region M1 of three independent experiments for each cell line. Ma104 cells seem to be slightly hyperpolarized in relation to MDCK cells, but we cannot exclude the possibility of individual differences in dye uptake. Ouabain treatment induced significant PMD in both cell lines (28.1% of MDCK cells and 47.9% of Ma104 cells were gated

in region M1 vs. 11.8% and 14.6% of the respective controls), but, unexpectedly, this effect was more evident for Ma104 cells. Preincubation of Ma104 cells with BSO did not significantly alter its membrane polarization (8.6% of cells gated in region M1) but completely blocked the ouabain-induced depolarization (12.2% of cells gated in region M1). Surprisingly, GSH alone depolarized the plasma membrane of MDCK cells (42.4% of cells gated in region M1) and further increased the number of depolarized cells after ouabain treatment (71.0%).
Effect of GSH and BSO on ouabain-induced tyrosine phosphorylation (P-Tyr)
Plasma membrane depolarization may lead to an increase in P-Tyr of several proteins and to the activation of the Ras-regulated kinase pathway (23, 24). Contreras et al. (12) showed that ouabain causes a sustained increase in P-Tyr in MDCK but not in Ma104 cells. However, the relationship between P-Tyr and ouabain toxicity is not well determined. To address this issue, we examined the effects of GSH and BSO in sustained ouabain-induced P-Tyr in these two cell lines. Preliminary experiments showed that 1 and 10 µM ouabain increased tyrosine phosphorylation in MDCK cells, but these concentrations failed to increase P-Tyr in Ma104 cells (data not shown).
The effects of GSH and BSO on ouabain-induced P-Tyr are shown in Figure 8. Insets illustrate representative photographs of each treatment. Ouabain (10 µM) induced a sustained increase in P-Tyr in MDCK cells, and GSH almost completely reversed this effect (upper panel). Conversely, even 100 µM ouabain was insufficient to substantially increase P-Tyr in Ma104 cells, but the preincubation with BSO for 24 h amplified ouabain-induced P-Tyr (lower panel).
Effect of GSH and BSO on ouabain-induced Ras expression
The p21 Ras proteins transduce signals from certain protein tyrosine kinase (PTK) receptors to the mitogen-activated protein kinase (MAPK) cascade. It is known that ouabain activates Ras and MAPK upon phosphorylation of Src and EGFR (25, 26), and Contreras et al. showed that ouabain induced activation of MAPK in MDCK, but not in Ma104 cells (12), suggesting that this activation is related to ouabain toxicity.
Although ouabain activates Ras, it is not known whether it also increases the expression of this protein. We have previously shown that the induction of Ras expression by vanadate was related to cell death and that GSH was able to revert both (27). Moreover, others have observed that increased Ras expression is associated with cell death (28, 29). Therefore, we investigated whether ouabain induced Ras expression in the cell lines studied and whether GSH and BSO were able to modify this effect.
Preliminary experiments revealed that 100 nM, a dose that did not alter MDCK cell viability, did not induce Ras expression, even after incubation for 24 h. However, Ras expression was induced by ouabain concentrations of 1 µM or above (data not shown). Because these cells detach from the substrate after incubation with ouabain concentrations higher than 1 µM for periods longer than 6 h, this time was used to study the effects of BSO and GSH in ouabain-induced Ras expression in Ma104 and MDCK cells.

As shown in Figure 9, 10 µM ouabain, a dose that greatly affects MDCK cell viability, increased the expression of Ras protein, and GSH completely avoided this increase (upper panel). On the other hand, no significant induction of Ras expression could be detected in Ma104 cells treated with 100 µM ouabain (lower panel). However, similar to what was observed for P-Tyr, the preincubation with BSO led to the induction of Ras expression following ouabain treatment.
DISCUSSION
Ouabain-induced cell death: apoptosis and necrosis
The sustained blockage of Na+, K+-ATPase by ouabain is thought to lead to cell death as a result of hydroelectrolytic imbalance. However, recent reports suggest that ouabain-induced cell death may occur for other reasons. Contreras et al. (10) showed, for the first time, that Ma104 cells were resistant to the toxic effects of ouabain, even though their pumps were inhibited and the cells lost K+. Ho et al. (30) observed that neither the number of ouabain binding sites nor their affinity for ouabain could fully account for the ouabain resistance of a human trophoblast cell line, and, more recently, Pchejetski et al. (19) demonstrated that ouabain-induced MDCK cell death is independent of the inhibition of Na+, K+-ATPase-mediated ion fluxes.
In the present study, we observed that Ma104 cells are ∼1000 times more resistant to ouabain than MDCK cells, confirming the results of Contreras et al. (10). The trypan blue exclusion method allows measurement of cells that have lost membrane integrity, which is characteristic of necrosis. However, it does not discriminate from late apoptotic cells, which also lose membrane integrity. By labeling cells with Annexin V-FITC, we were not able to observe apoptosis in any of the ouabain concentrations or incubation times tested, suggesting that ouabain-induced MDCK cell death occurs primarily by necrosis. Corroborating our results, Contreras et al. (12) could not detect apoptosis in Ma104 or MDCK cells, measuring DNA degradation. More recently, Pchejetski et al. (19) performed a panel of experiments to test apoptosis (nucleus condensation, caspase-3 activation) and necrosis (cell swelling, LDH release, and a smear-like pattern of DNA degradation) in MDCK cells treated with ouabain, also concluding that cell death was occurring by necrosis.
Interestingly, ouabain is known to induce apoptosis in a variety of cells, such as human lymphocytes (31), human prostatic smooth muscle cells (32), and murine cortical neurons (33). In the latter study, the authors also observed that ouabain could induce apoptosis and necrosis simultaneously in individual neurons.
Contrasting its toxic effects, ouabain can also protect vascular smooth muscle cells from apoptosis induced by serum withdrawal (34, 35). Thus, it seems that the cellular effects of ouabain are a complex issue involving different pathways, which can lead to necrosis, apoptosis, or even the prevention of them, depending on the cell type and experimental conditions.
Ouabain-induced ROS and the effect of antioxidants on ouabain-induced MDCK cell death
There are presently two mechanisms described for the cellular actions of ouabain: i) the classical inhibition of Na+, K+-ATPase, which leads to an increase in cytosolic calcium, and ii) the signal transduction pathway, leading to Ras activation and production of ROS. The first mechanism is

known to regulate contractility in cardiomyocytes, and the second is related to the nonproliferative growth (hypertrophy) of these cells (2, 12–14, 25). Although two different groups reported the production of ROS by ouabain (13–15), those authors could not identify the type of ROS formed, due to insufficient selectivity of the probe used. Moreover, none of these studies focused on the possible relationship between ouabain-induced ROS and ouabain toxicity.
In the present study, two probes were used to detect ROS. The results of the experiments with DHR, a probe used to detect H2O2 and peroxinitrite, suggest that these ROS are not generated in ouabain-treated cells. However, by using P-Fluo, a probe specific for the detection of O2
•¯ and •OH, we were able to identify superoxide anion as the ROS formed by ouabain treatment. It has been shown that Ras activation leads to O2
•¯ formation (36, 37). Thus, the O2•¯ generated in
MDCK cells by ouabain may be a product of Ras activation.
The induction of O2•¯ only in MDCK cells suggests that this anion, or at least the pathway that
lead to its production, is related to ouabain toxicity. Therefore, we tested whether the cellular antioxidant enzymes catalase and superoxide dismutase, and the major nonenzymatic cellular antioxidant GSH, would be able to protect MDCK cells from ouabain toxicity.
It is known that most cell types metabolize extracellular GSH through the ectoenzyme g-glutamyl transpeptidase (GGT), which transfers glutamate to an amino acid acceptor. The cysteinylglycine is then hydrolyzed, and the amino acids are internalized independently. However, there is growing evidence suggesting the existence of both Na+-dependent and Na+-independent GSH transporters in monocytes and in brain, kidney, and lung cells (38–41). It has also been shown that cells can uptake superoxide dismutase added to the culture medium, increasing their defense against oxidative agents (42–44). Although, to our knowledge, there is no data on cellular uptake of catalase, several authors reported protection from oxidative stress by exogenous catalase (27, 43, 45, 46).
We did not notice any significant protection against ouabain toxicity by catalase; moreover, no increase in H2O2 was seen using the probe DHR, suggesting that ouabain does not generate H2O2. However, although GSH protected MDCK cells from ouabain toxicity, SOD did not alter cell survival. This result may be due to at least two possibilities: SOD does not penetrate in MDCK cells, or, although ouabain induces the formation of O2
•¯, this is not directly related to ouabain-induced cell death. This last hypothesis is supported by the fact that we observed only a marginal increase in O2
•¯ production (14%) in MDCK cells treated with 10 µM ouabain, a concentration that virtually destroys all the cells. Conversely, Xie et al. (13) observed a much greater increase in ROS production (approximately twofold) in cardiomyocytes treated with ouabain in concentrations that did not alter cellular viability, also suggesting that ROS are not directly related to ouabain-induced cell death. However, more studies are necessary in order to answer this issue fully.
Ouabain-induced plasma membrane depolarization: relation to cell death?
It is known that the blockage of Na+, K+-ATPase by ouabain leads to membrane depolarization, and this effect has been associated with cell death (17, 20–22). Unexpectedly, our results suggest that ouabain-induced depolarization is not harmful per se: First, because an increased number of ouabain-resistant Ma104 cells depolarize after treatment with ouabain, in comparison to MDCK

cells; second, because GSH increased the number of MDCK cells depolarizing after ouabain treatment, while BSO blocked the ouabain-induced depolarization in Ma104 cells. Surprisingly, GSH alone was able to depolarize MDCK cells. Although the mechanisms for this effect are not presently understood, it is known that GSH depolarizes neurons (47).
Providing GSH protected MDCK cells and BSO sensitized Ma104 cells to ouabain, these results suggest that the ability of cells to depolarize their plasma membrane after treatment with ouabain may be indeed a protection against cell death. Confirming this assumption, there is at least one more evidence that PMD can protect cells from death: Soler et al. (48) showed that elevation of extracellular K+, which induces chronic neuronal PMD in vitro, promotes the survival of chick spinal cord motoneurons deprived of any neurotrophic support.
Sustained increase in P-Tyr and induction of Ras expression: relation to ouabain-induced cell death
Contreras et al. (12) showed that incubation of MDCK cells with ouabain led to a sustained increase in P-Tyr, which reaches a maximum at 2 h and persists even after 8 h of incubation; however, they could not detect any increase in P-Tyr in Ma104 cells. In the present report, we established that ouabain-induced P-Tyr increase in MDCK cells could be modulated by GSH. These results were further confirmed by the fact that BSO led to an increase in ouabain-induced P-Tyr in Ma104 cells. Given that GSH protected MDCK and BSO sensitized Ma104 cells to ouabain, these results suggest that the sustained increase in P-Tyr is related to ouabain cytotoxicity.
It is known that ouabain induces a rapid increase in protein P-Tyr in a number of cell lines (12, 14, 25). Thus, phosphorylation of tyrosine might be part of a physiological signaling cascade induced by ouabain, returning to normal levels within few minutes, whereas a sustained increase in P-Tyr may lead to cell death. However, we could not visualize any increase in P-Tyr in Ma104 cells treated with 10 µM ouabain at shorter times (5 and 15 min; data not shown), suggesting that their resistance is related to a “failure” on increasing protein phosphorylation when treated with ouabain. This “failure” is not a general feature of Ma104 cells, because we observed previously an intense protein P-Tyr when these cells were treated with vanadate (27).
Relationship between plasma membrane depolarization, P-Tyr, and Ras protein
Plasma membrane depolarization may lead to an increase in tyrosine phosphorylation of several proteins and to the activation of the Ras-regulated kinase pathway (23, 24). However, our results suggest that this is not an obligatory pathway, because Ma104 cells depolarize even more than MDCK cells, but the increase in P-Tyr and Ras was seen only in the latter. The p21 Ras proteins transduce signals from certain PTK receptors to the MAPK cascade and are related both to cell growth and differentiation as well as to cell death. Although it is not known how they can lead to these opposite effects, there is presently a growing literature relating increased expression of Ras to cell death (28, 29, 49, 50). Interesting, in all those studies, it was noticed that Ras-induced cell death was related to autophagy, a type of programmed cell death distinct from apoptosis but sharing features of both apoptosis and necrosis. Thus, it is possible that the ouabain-induced MDCK cell death may be occurring by autophagy, but more studies are necessary to address this issue.

Relationship between ouabain-induced cell death and Na+, K+-ATPase inhibition
Based on their results, Contreras et al. (10, 12) suggested that there is a mechanism that transduces a blockade of the Na+, K+-ATPase in a detachment of the cells from each other and from the substrate and that Ma104 cells have lost this mechanism. Our results do not support this hypothesis, because we were able to modulate the cellular response to ouabain simply by adding GSH or BSO to the medium. Moreover, Pchejetski et al. (19) showed that the detachment occurs subsequently to cell death.
Contreras et al. (10, 12) also showed that the Na+, K+-ATPases of MDCK and Ma104 cells have essentially the same affinity for ouabain. However, the toxic effects occur only with a ouabain concentration 10 times greater (1 µM) than that necessary to completely inhibit the Na+, K+-ATPase of MDCK cells, suggesting that ouabain toxicity is not directly related to the inhibition of the pump. Ho et al. (30) also showed that the resistance of a human trophoblast cell line to ouabain was not due to a lack of response of its Na+, K+-ATPase and suggested that the cellular response to ouabain could be mediated through different pathways, resulting in different phenotypes. More recently, Pchejetski et al. (19) proposed that the death signal might be triggered by a conformational change of the Na+, K+-ATPase α-subunit, induced by ouabain.
In a recent paper, Ward et al. (8) showed, for the first time, the existence of two different ouabain binding sites in adrenocortical cells, probably due to the existence of another receptor distinct from Na+, K+-ATPase. Thus, the differences in cellular responses to ouabain may be related to the presence of other ouabain-receptor(s) or even distinct binding sites in Na+, K+-ATPase itself. However, this assumption needs further confirmation and may not answer how GSH protects against ouabain cytotoxicity.
Role of GSH on ouabain-induced signal transduction
The action of GSH in P-Tyr and Ras expression does not seem to be due to its very well known antioxidant and free radical scavenger function, because ouabain-induced ROS formation is subsequent to the increase in P-Tyr (14), and Ma104 was not phosphorylated at any incubation time. Thus, it is probable that GSH may have another function in the events induced by ouabain.
Novel actions of GSH have been described in the past few years, suggesting a direct role of GSH in signal transduction. It has been shown that GSH acts both as a neuromodulator and as a neurotransmitter in the nervous system, binding to its own receptor (for a review, see ref 47), that GSH stimulates the autophosphorylation of the EGFR and the insulin receptor (51), and that the platelet-derived growth factor receptor autophosphorylation is severely impaired at low cellular GSH levels (52). These authors suggest that the interaction of GSH with this growth-factor receptor is complex and may involve both cytosolic and extracellular receptor domains. However, Wilhelm et al. (53) observed in cells treated with methyl methanesulfonate, which activates the Jun N-terminal kinases/stress-activated protein kinases (JNK/SAPK), that GSH or N-acetylcysteine (NAC), a precursor of GSH synthesis, inhibits this activation, whereas depletion of the cellular GSH pool increased it. They suggest that GSH and NAC interact directly or through enzyme-catalyzed reactions with electrophylic compounds, reducing the number of reactions of alkylating agents with critical target molecules. Bhunia et al. (54) showed that the activation of Ras and p44 MAPK and the induction of c-fos were inhibited by NAC and

GSH and that depletion of GSH by BSO up-regulated this signaling cascade. Thus, because many signaling molecules contain redox-sensitive cysteine residues, which regulate enzyme activity, the effects of GSH on signal transduction may be due to its ability to alter the intracellular reducing environment.
Another possible explanation for the effect of GSH in P-Tyr is the regulation of phosphotyrosine phosphatases. The cyclic cascades, with reversible P-Tyr, are essential for the regulation of cell cycles and signal transduction. The elevation of P-Tyr may be achieved either by the activation of PTKs and/or inactivation of protein tyrosine phosphatases (PTPs), the latter being of particular importance due to a much higher specific activity relative to that of PTKs (55). It has been shown that GSH participates in the regulation of PTP-1B, a highly abundant PTP, by the conversion of an unstable sulfenic derivative to a glutathionylated PTP-1B, which can be further reactivated by thioltransferases (56). Thus, GSH may maintain PTPs in their active form.
Final considerations
Although it has long been known that ouabain may be cytotoxic, the mechanisms of its toxicity are only now being uncovered. The suggested involvement of ouabain in hypertension raised the need for a better understanding of its cellular action. The results obtained in the present study have three major implications: First, there is no direct correlation between membrane depolarization and ouabain-induced cell death; second, ouabain toxicity is related to signal transduction, more specifically to the Ras-ROS pathway, and is not directly related to its classical action in Na+, K+-ATPase inhibition; and third, GSH plays a major role in ouabain-induced signal transduction and cell death. This role of GSH needs better investigation, because it is known that plasma levels of GSH are reduced and that plasma levels of endogenous ouabain are increased in animal and human hypertension (4–7, 57, 58).
ACKNOWLEDGMENTS
The present work was supported by grants from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ), Programa de Apoio aos Núcleos de Excelência (PRONEX), and Programa de Apoio ao Desenvolvimento Científico e Tecnológico (PADCT). We are indebted to Drs. Ottilia Affonso-Mitidieri, Fernanda Mello de Queiroz, and Álvaro Augusto da Costa Leitão for the generous gift of reagents and for helpful discussions.
REFERENCES
1. Kelly, R. A., and Smith, T. W. (1993) Digoxin in heart failure: implications of recent trials. J. Am. Coll. Cardiol. 4(Suppl A), 107A–112A
2. Kometiani, P., Li, J., Gnudi, L., Khan, B. B., Askari, A., and Xie, Z. (1998) Multiple signal transduction pathways link Na+/K+-ATPase to growth-related genes in cardiac myocytes. The roles of Ras and mitogen-activated protein kinases. J. Biol. Chem. 273, 15249–15256
3. Kurashina, T., Kirchner, K. A., Granger, J. P., and Patel, A. R. (1996) Chronic sodium-potassium-ATPase inhibition with ouabain impairs renal haemodynamics and pressure natriuresis in the rat. Clin. Sci. 91, 497–502

4. Hamlyn, J. M., Hamilton, B. P., and Manunta, P. (1996) Endogenous ouabain, sodium balance and blood pressure: a review and a hypothesis. J. Hypertens. 14, 151–167
5. Manunta, P., Stella, P., Rivera, R., Ciurlino, D., Cusi, D., Ferrandi, M., Hamlyn, J. M., and Bianchi, G. (1999) Left ventricular mass, stroke volume, and ouabain-like factor in essential hypertension. Hypertension 34, 450–456
6. Manunta, P., Hamilton, J., Rogowski, A. C., Hamilton, B. P., and Hamlyn, J. M. (2000) Chronic hypertension induced by ouabain but not digoxin in the rat: antihypertensive effect of digoxin and digitoxin. Hypertens. Res. 23, S77–S85
7. Manunta, P., Hamilton, B. P., and Hamlyn, J. M. (2001) Structure-activity relationships for the hypertensinogenic activity of ouabain: role of the sugar and lactone ring. Hypertension 37, 472–477
8. Ward, S. C., Hamilton, B. P., and Hamlyn, J. M. (2002) Novel receptors for ouabain: studies in adrenocortical cells and membranes. Hypertension 39, 536–542
9. Bolivar, J. J., Lazaro, A., Fernandez, S., Stefani, E., Pena-Cruz, V., Lechene, C., and Cereijido, M. (1987) Rescue of a wild-type MDCK cell by a ouabain-resistant mutant. Am. J. Physiol. 253, C151–C161
10. Contreras, R. G., Lazaro, A., Mujica, A., Gonzalez-Mariscal, L., Valdes, J., Garcia-Villegas, M. R., and Cereijido, M. (1995) Ouabain resistance of the epithelial cell line (Ma104) is not due to lack of affinity of its pumps for the drug. J. Membr. Biol. 145, 295–303
11. Contreras, R. G., Lazaro, A., Bolivar, J. J., Flores-Maldonado, C., Sanchez, S. H., Gonzalez-Mariscal, L., Garcia-Villegas, M. R., Valdes, J., and Cereijido, M. (1995) A novel type of cell-cell cooperation between epithelial cells. J. Membr. Biol. 145, 305–310
12. Contreras, R. G., Shoshani, L., Flores-Maldonado, C., Lazaro, A., and Cereijido, M. (1999) Relationship between Na(+),K(+)-ATPase and cell attachment. J. Cell Sci. 112, 4223–4232
13. Xie, Z., Kometiani, P., Liu, J., Li, J., Shapiro, J. I., and Askari, A. (1999) Intracellular reactive oxygen species mediate the linkage of Na+/K+-ATPase to hypertrophy and its marker genes in cardiac myocytes. J. Biol. Chem. 274, 19323–19328
14. Liu, J., Tian, J., Haas, M., Shapiro, J. I., Askari, A., and Xie, Z. (2000) Ouabain interaction with cardiac Na+/K+-ATPase initiates signal cascades independent of changes in intracellular Na+ and Ca2+ concentrations. J. Biol. Chem. 275, 27838–27844
15. Kajikawa, M., Fujimoto, S., Tsuura, Y., Mukai, E., Takeda, T., Hamamoto, Y., Takehiro, M., Fujita, J., Yamada, Y., and Seino, Y. (2002) Ouabain suppresses glucose-induced mitochondrial ATP production and insulin release by generating reactive oxygen species in pancreatic islets. Diabetes 51, 2522–2529

16. Capella, L. S., Gefe, M., Silva, E. F., Morales, M. M., Affonso-Mitidieri, O., Lopes, A. G., Rumjanek, V. M., and Capella, M. A. M. (2001) Reduced glutathione protect cells from ouabain toxicity. Biochim. Biophys. Acta 1526, 293–300
17. Bortner, C. D., Gomez-Angelats, M., and Cidlowski, J. A. (2001) Plasma membrane depolarization without repolarization is an early molecular event in anti-Fas-induced apoptosis. J. Biol. Chem. 276, 4304–4314
18. Pou, S., Huang, Y. I., Bhan, A., Bhadti, V. S., Hosmane, R. S., Wu, S. Y., Cao, G. L., and Rosen, G. M. (1993) A fluorophore-containing nitroxide as a probe to detect superoxide and hydroxyl radical generated by stimulated neutrophils. Anal. Biochem. 212, 85–90
19. Pchejetski, D., Taurin, S., Sarkissian, S. D., Lopina, O. D., Pshezhetsky, A. V., Tremblay, J., deBlois, D., Hamet, P., and Orlova, S. N. (2003) Inhibition of Na+,K+-ATPase by ouabain triggers epithelial cell death independently of inversion of the [Na+]i/[K+]i ratio. Biochem. Biophys. Res. Commun. 301, 735–744
20. Balestrino, M., Young, J., and Aitken, P. (1999) Block of (Na+,K+)ATPase with ouabain induces spreading depression-like depolarization in hippocampal slices. Brain Res. 838(1-2), 37–44
21. Potapova, T.V., Aslanidi, K.B., and Boitzova, L.Ju. (1990) Energy transfer via cell-to-cell junctions. Ouabain-resistant cells maintain a membrane potential in ouabain-sensitive cells. FEBS Lett. 262(1), 69–71
22. Mann, C. L., Bortner, C. D., Jewell, C. M., and Cidlowski, J. A. (2001) Glucocorticoid-induced plasma membrane depolarization during thymocyte apoptosis: association with cell shrinkage and degradation of the Na(+)/K(+)-adenosine triphosphatase. Endocrinology 142, 5059–5068
23. Zwick, E., Wallasch, C., Daub, H., and Ullrich, A. (1999) Distinct calcium-dependent pathways of epidermal growth factor receptor transactivation and PYK2 tyrosine phosphorylation in PC12 cells. J. Biol. Chem. 274, 20989–20996
24. Sohn, H. Y., Keller, M., Gloe, T., Morawietz, H., Rueckschloss, U., and Pohl, U. (2000) The small G-protein Rac mediates depolarization-induced superoxide formation in human endothelial cells. J. Biol. Chem. 275, 18745–18750
25. Haas, M., Askari, A., and Xie, Z. (2000) Involvement of Src and epidermal growth factor receptor in the signal-transducing function of Na+/K+-ATPase. J. Biol. Chem. 275, 27832–27837
26. Haas, M., Wang, H., Tian, J., and Xie, Z. (2002) Src-mediated Inter-receptor cross-talk between the Na+/K+-ATPase and the epidermal growth factor receptor relays the signal from ouabain to mitogen-activated protein kinases J. Biol. Chem. 277, 18694–18702

27. Capella, L. S., Gefe, M., Silva, E. F., Affonso-Mitidieri, O., Lopes, A. G., Rumjanek, V. M., and Capella, M. A. M. (2002) Mechanisms of vanadate-induced cellular toxicity: role of cellular glutathione and NADPH. Arch. Biochem. Biophys. 406, 65–72
28. Botella, J. A., Kretzschmar, D., Kiermayer, C., Feldmann, P., Hughes, D. A., and Schneuwly, S. (2003) Deregulation of the EGFR/Ras Signaling Pathway Induces Age-related Brain Degeneration in the Drosophila Mutant vap. Mol. Biol. Cell 14, 241–250
29. Kitanaka, C., Kato, K., Ijiri, R., Sakurada, K., Tomiyama, A., Noguchi, K., Nagashima, Y., Nakagawara, A., Momoi, T., Toyoda, Y., et al. (2002) Increased Ras Expression and Caspase-Independent Neuroblastoma Cell Death: Possible Mechanism of Spontaneous Neuroblastoma Regression. J. Natl. Cancer Inst. 94, 358–368
30. Ho, C. K., Chen, S. M., and Wang, S. Y. (2000) Ouabain resistance of a human trophoblast cell line is not related to its reactivity to ouabain. Eur. J. Cell Biol. 79, 765–770
31. Olej, B., dos Santos, N. F., Leal, L., and Rumjanek, V. M. (1998) Ouabain induces apoptosis on PHA-activated lymphocytes. Biosci. Rep. 18, 1–7
32. Chueh, S. C., Guh, J. H., Chen, J., Lai, M. K., and Teng, C. M. (2001) Dual effects of ouabain on the regulation of proliferation and apoptosis in human prostatic smooth muscle cells. Urol. 166, 347–353
33. Xiao, A. Y., Wei, L., Xia, S., Rothman, S., and Yu, S. P. (2002) Ionic mechanism of ouabain-induced concurrent apoptosis and necrosis in individual cultured cortical neurons. J. Neurosci. 22, 1350–1362
34. Orlov, S. N., Thorin-Trescases, N., Kotelevtsev, S. V., Tremblay, J., and Hamet, P. (1999) Inversion of the Intracellular Na+/K+ Ratio Blocks Apoptosis in Vascular Smooth Muscle at a Site Upstream of Caspase-3. J. Biol. Chem. 274, 16545–16552
35. Orlov, S. N., Taurin, S., Thorin-Trescases, N., Dulin, N. O., Tremblay, J., and Hamet, P. (2000) Inversion of the Intracellular Na+/K+ Ratio Blocks Apoptosis in Vascular Smooth Muscle Cells by Induction of RNA Synthesis. Hypertension 35, 1062–1068
36. Yang, J. Q., Zhao, W., Duan, H., Robbins, M. E., Buettner, G. R., Oberley, L. W., and Domann, F. E. (2001) v-Ha-RaS oncogene upregulates the 92-kDa type IV collagenase (MMP-9) gene by increasing cellular superoxide production and activating NF-kappaB. Free Radic. Biol. Med. 31, 520–529
37. Yang, J. Q., Buettner, G. R., Domann, F. E., Li, Q., Engelhardt, J. F., Weydert, C. D., and Oberley, L. W. (2002) v-Ha-ras mitogenic signaling through superoxide and derived reactive oxygen species. Mol. Carcinog. 33, 206–218
38. Seres, T., Knickelbein, R. G., Warshaw, J. B., and Johnston, J. B., Jr. (2000) The Phagocytosis-Associated Respiratory Burst in Human Monocytes Is Associated with Increased Uptake of Glutathione. J. Immunol. 165, 3333–3340

39. Hagen, T. M., Aw, T. Y., and Jones, D. P. (1988) Glutathione uptake and protection against oxidative injury in isolated kidney cells. Kidney Int. 34, 74–81
40. Kannan, R., Yi, J. R., Tang, D., Li, Y., Zlokovic, B. V., and Kaplowitz, N. (1996) Evidence for the existence of a sodium-dependent glutathione (GSH) transporter. J. Biol. Chem. 271, 9754–9758
41. Kannan, R., Mittur, A., Bao, Y., Tsuruo, T., and Kaplowitz, N. (1999) GSH transport in immortalized mouse brain endothelial cells: evidence for apical localization of a sodium-dependent GSH transporter. J. Neurochem. 73, 390–399
42. Emerit, I., Garban, F., Vassy, J., Levy, A., Filipe, P., and Freitas, J. (1996) Superoxide-mediated clastogenesis and anticlastogenic effects of exogenous superoxide dismutase. Proc. Natl. Acad. Sci. USA 93, 12799–12804
43. Thamilselvan, S., Khan, S. R., and Menon, M. (2003) Oxalate and calcium oxalate mediated free radical toxicity in renal epithelial cells: effect of antioxidants. Urol. Res. 31, 3–9
44. Filipe, P., Emerit, I., Vassy, J., Levy, A., Huang, V., and Freitas, J. (1999) Cellular penetration of fluorescently labeled superoxide dismutase of various origins. Mol. Med. 5, 517–525
45. Chen, C. Y., Wang, Y. F., Lin, Y. H., and Yen, S. F. (2003) Nickel-induced oxidative stress and effect of antioxidants in human lymphocytes. Arch. Toxicol. 77, 123–130
46. Zhang, Z., Huang, C., Li, J., Leonard, S. S., Lanciotti, R., Butterworth, L., and Shi, X. (2001) Vanadate-induced cell growth regulation and the role of reactive oxygen species. Arch. Biochem. Biophys. 392, 311–320
47. Janaky, R., Ogita, K., Pasqualotto, B. A., Bains, J. S., Oja, S. S., Yoneda, Y., and Shaw, C. A. (1999) Glutathione and signal transduction in the mammalian CNS. J. Neurochem. 73, 889–902
48. Soler, R. M., Egea, J., Mintenig, G. M., Sanz-Rodriguez, C., Iglesias, M., and Comella, J. X. (1998) Calmodulin is involved in membrane depolarization-mediated survival of motoneurons by phosphatidylinositol-3 kinase and MAPK-independent pathways. J. Neurosc. 18, 1230–1239
49. Luth, H. J., Holzer, M., Gertz, H.-J., and Arendt, T. (2000) Aberrant expression of nNOS in pyramidal neurons in Alzheimer’s disease is highly co-localized with p21ras and p16INK4a. Brain Res. 852, 45–55
50. Gartner, U., Holzer, M., and Arendt, T. (1999) Elevated Expression of P21Ras is an Early Event in Alzheimer’s Disease and Precedes Neurofibrillary Degeneration. Neuroscience 91, 1–5

51. Clark, S., and Konstantopoulos, N. (1993) Sulphydryl agents modulate insulin- and epidermal growth factor (EGF)-receptor kinase via reaction with intracellular receptor domains: differential effects on basal versus activated receptors. Biochem. J. 292, 217–223
52. Rigacci, S., Iantomasi, T., Marraccini, P., Berti, A., Vincenzini, M. T., and Ramponi, G. (1997) Evidence for glutathione involvement in platelet-derived growth-factor-mediated signal transduction. Biochem. J. 324, 791–796
53. Wilhelm, D., Bender, K., Knebel, A., and Angel, P. (1997) The level of intracellular glutathione is a key regulator for the induction of stress-activated signal transduction pathways including Jun N-Terminal protein kinases and p38 kinase by alkylating agents. Mol. Cell. Biol. 17, 4792–4800
54. Bhunia, A. K., Han, H., Snowden, A., and Chatterjee, S. (1997) Redox-regulated Signaling by Lactosylceramide in the Proliferation of Human Aortic Smooth Muscle Cells. J. Biol. Chem. 272, 15642–15649
55. Fischer, E. H., Charbonneau, H., and Tonks, N. K. (1991) Protein tyrosine phosphatases: a diverse family of intracellular and transmembrane enzymes. Science 253, 401–406
56. Barrett, W. C., DeGnore, J. P., Keng, Y. F., Zhang, Z. Y., Yim, M. B., and Chock, P. B. (1999) Roles of superoxide radical anion in signal transduction mediated by reversible regulation of protein-tyrosine phosphatase 1B. J. Biol. Chem. 274, 34543–34546
57. Nemeth, I., Orvos, H., and Boda, D. (2001) Blood glutathione redox status in gestational hypertension. Free Radic. Biol. Med. 30, 715–721
58. Vaziri, N. D., Wang, X. Q., Oveisi, F., and Rad, B. (2000) Induction of oxidative stress by glutathione depletion causes severe hypertension in normal rats. Hypertension 36, 142–146
Received November 14, 2002; accepted June 10, 2003.

Table 1 Detection of ouabain-induced apoptosis in ouabain-treated MDCK cellsa Annexin V-Positive Cells Control 5.2±2.5 100 nM � 3 h 10.0±3.2 10 µM � 3 h 8.2±3.2 1 µM � 3 h 8.7±4.5 1 µM � 6 h 1.8±0.6 1 µM � 12 h 0.8±0.6
aThe methodology is described in Materials and Methods. Values are the percentage of cells labeled with Annexin-V FITC. Means ±SD were obtained from three experiments.
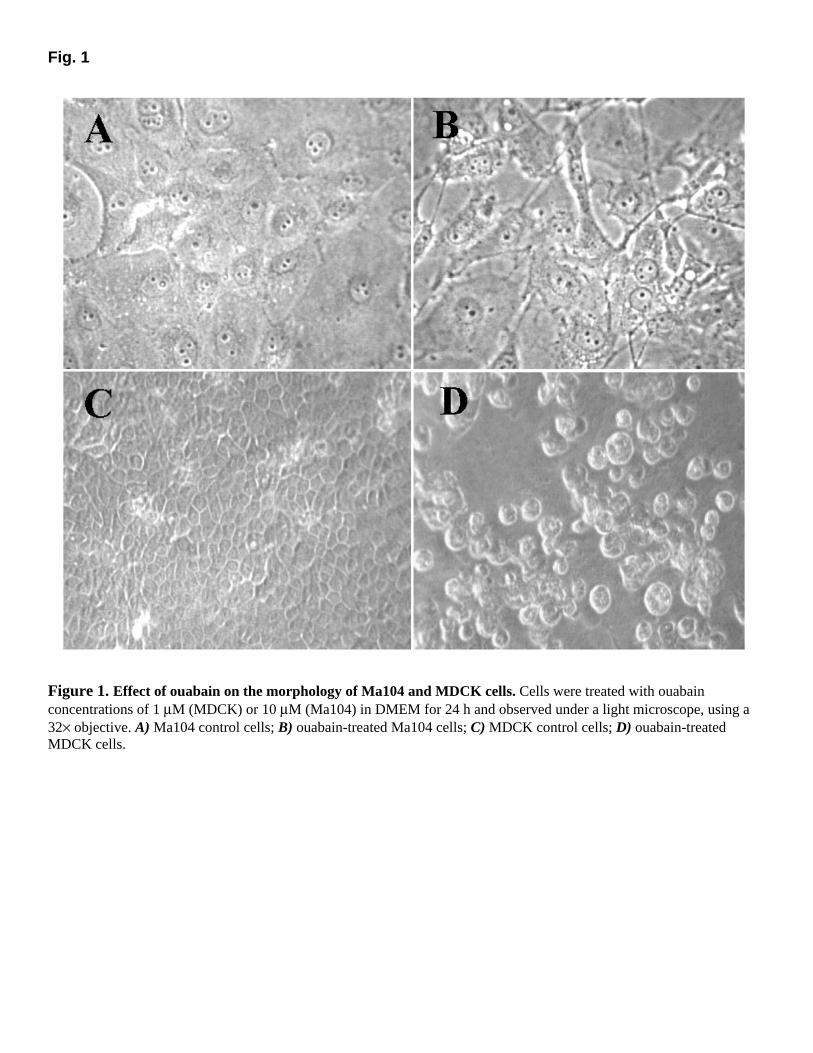
Fig. 1
Figure 1. Effect of ouabain on the morphology of Ma104 and MDCK cells. Cells were treated with ouabain concentrations of 1 µM (MDCK) or 10 µM (Ma104) in DMEM for 24 h and observed under a light microscope, using a 32× objective. A) Ma104 control cells; B) ouabain-treated Ma104 cells; C) MDCK control cells; D) ouabain-treated MDCK cells.
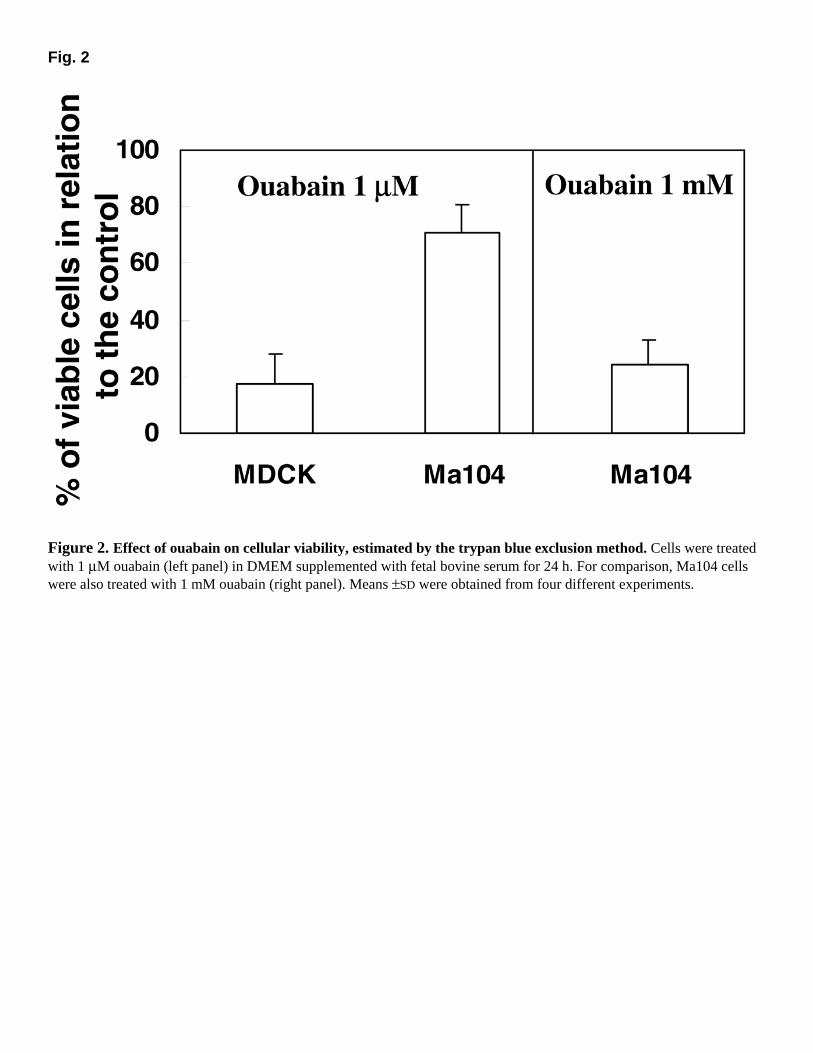
Fig. 2
Figure 2. Effect of ouabain on cellular viability, estimated by the trypan blue exclusion method. Cells were treated with 1 µM ouabain (left panel) in DMEM supplemented with fetal bovine serum for 24 h. For comparison, Ma104 cells were also treated with 1 mM ouabain (right panel). Means ±SD were obtained from four different experiments.
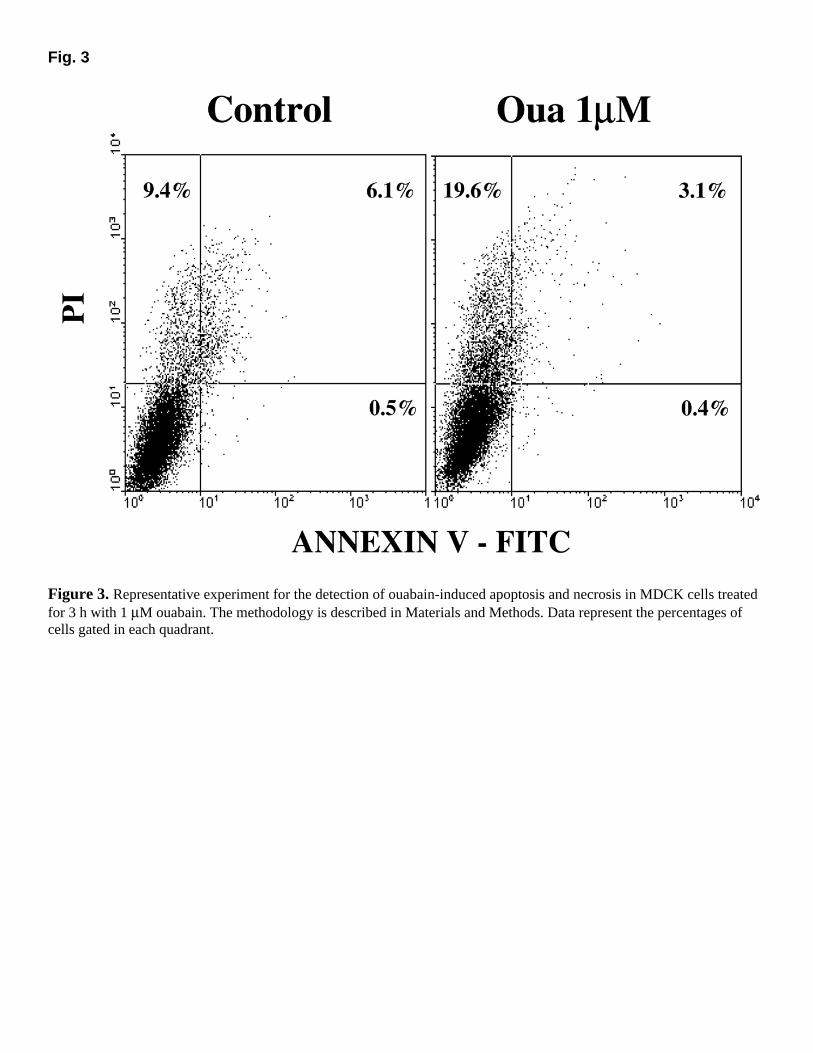
Fig. 3
Figure 3. Representative experiment for the detection of ouabain-induced apoptosis and necrosis in MDCK cells treated for 3 h with 1 µM ouabain. The methodology is described in Materials and Methods. Data represent the percentages of cells gated in each quadrant.
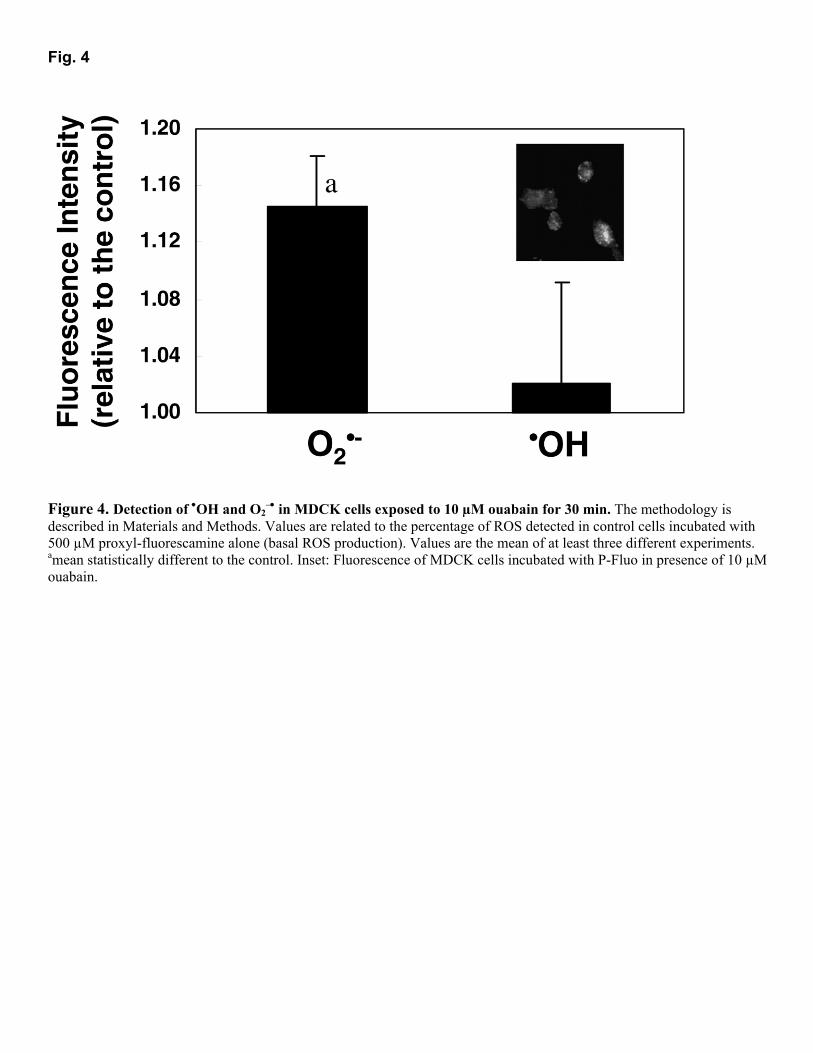
Fig. 4
Figure 4. Detection of •OH and O2
�• in MDCK cells exposed to 10 µM ouabain for 30 min. The methodology is described in Materials and Methods. Values are related to the percentage of ROS detected in control cells incubated with 500 µM proxyl-fluorescamine alone (basal ROS production). Values are the mean of at least three different experiments. amean statistically different to the control. Inset: Fluorescence of MDCK cells incubated with P-Fluo in presence of 10 µM ouabain.
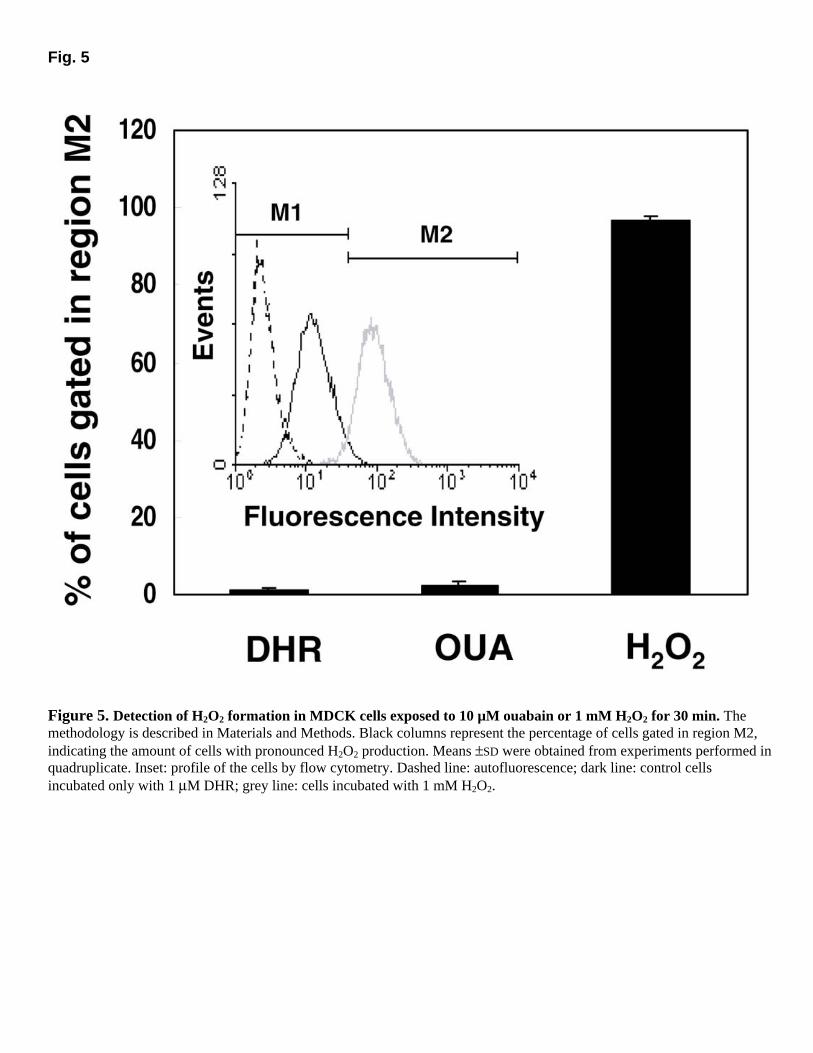
Fig. 5
Figure 5. Detection of H2O2 formation in MDCK cells exposed to 10 µM ouabain or 1 mM H2O2 for 30 min. The methodology is described in Materials and Methods. Black columns represent the percentage of cells gated in region M2, indicating the amount of cells with pronounced H2O2 production. Means ±SD were obtained from experiments performed in quadruplicate. Inset: profile of the cells by flow cytometry. Dashed line: autofluorescence; dark line: control cells incubated only with 1 µM DHR; grey line: cells incubated with 1 mM H2O2.
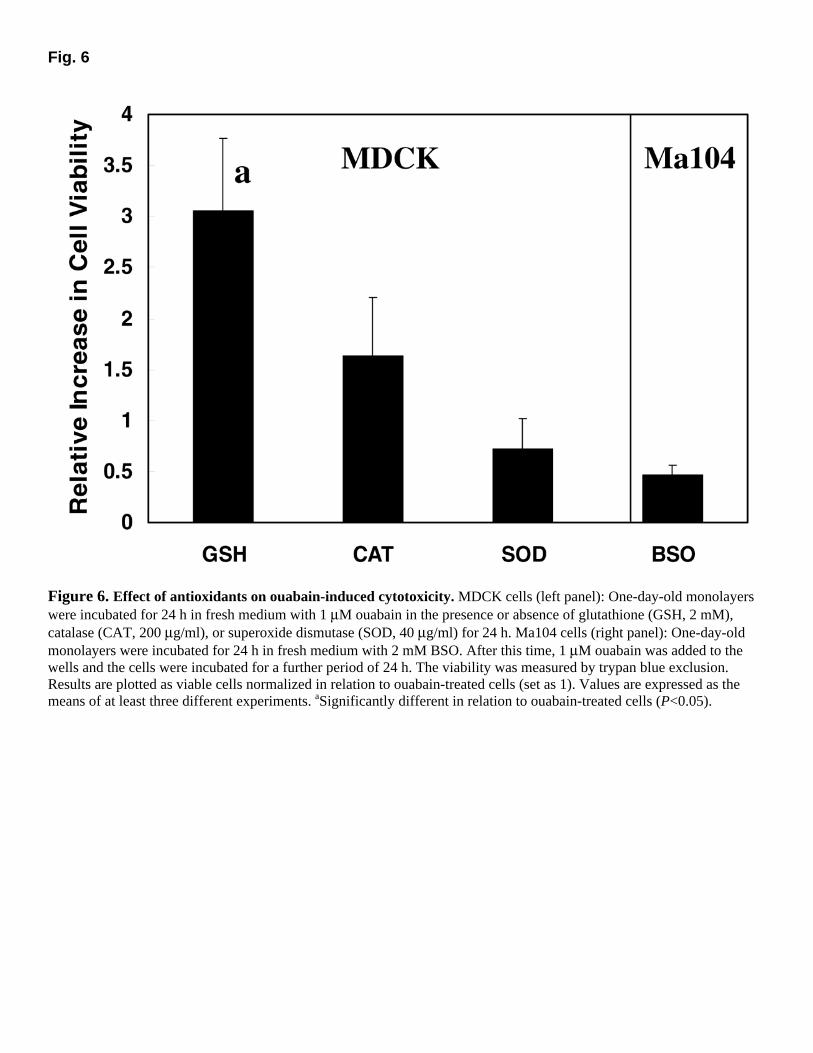
Fig. 6
Figure 6. Effect of antioxidants on ouabain-induced cytotoxicity. MDCK cells (left panel): One-day-old monolayers were incubated for 24 h in fresh medium with 1 µM ouabain in the presence or absence of glutathione (GSH, 2 mM), catalase (CAT, 200 µg/ml), or superoxide dismutase (SOD, 40 µg/ml) for 24 h. Ma104 cells (right panel): One-day-old monolayers were incubated for 24 h in fresh medium with 2 mM BSO. After this time, 1 µM ouabain was added to the wells and the cells were incubated for a further period of 24 h. The viability was measured by trypan blue exclusion. Results are plotted as viable cells normalized in relation to ouabain-treated cells (set as 1). Values are expressed as the means of at least three different experiments. aSignificantly different in relation to ouabain-treated cells (P<0.05).
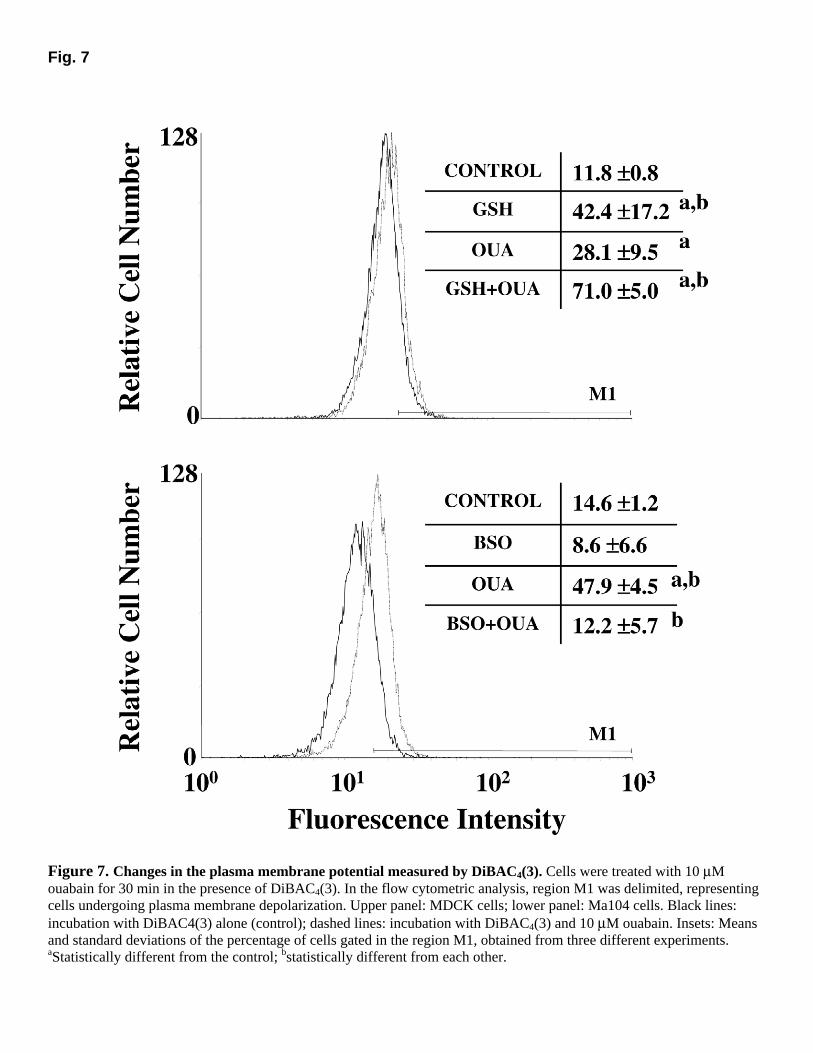
Fig. 7
Figure 7. Changes in the plasma membrane potential measured by DiBAC4(3). Cells were treated with 10 µM ouabain for 30 min in the presence of DiBAC4(3). In the flow cytometric analysis, region M1 was delimited, representing cells undergoing plasma membrane depolarization. Upper panel: MDCK cells; lower panel: Ma104 cells. Black lines: incubation with DiBAC4(3) alone (control); dashed lines: incubation with DiBAC4(3) and 10 µM ouabain. Insets: Means and standard deviations of the percentage of cells gated in the region M1, obtained from three different experiments. aStatistically different from the control; bstatistically different from each other.
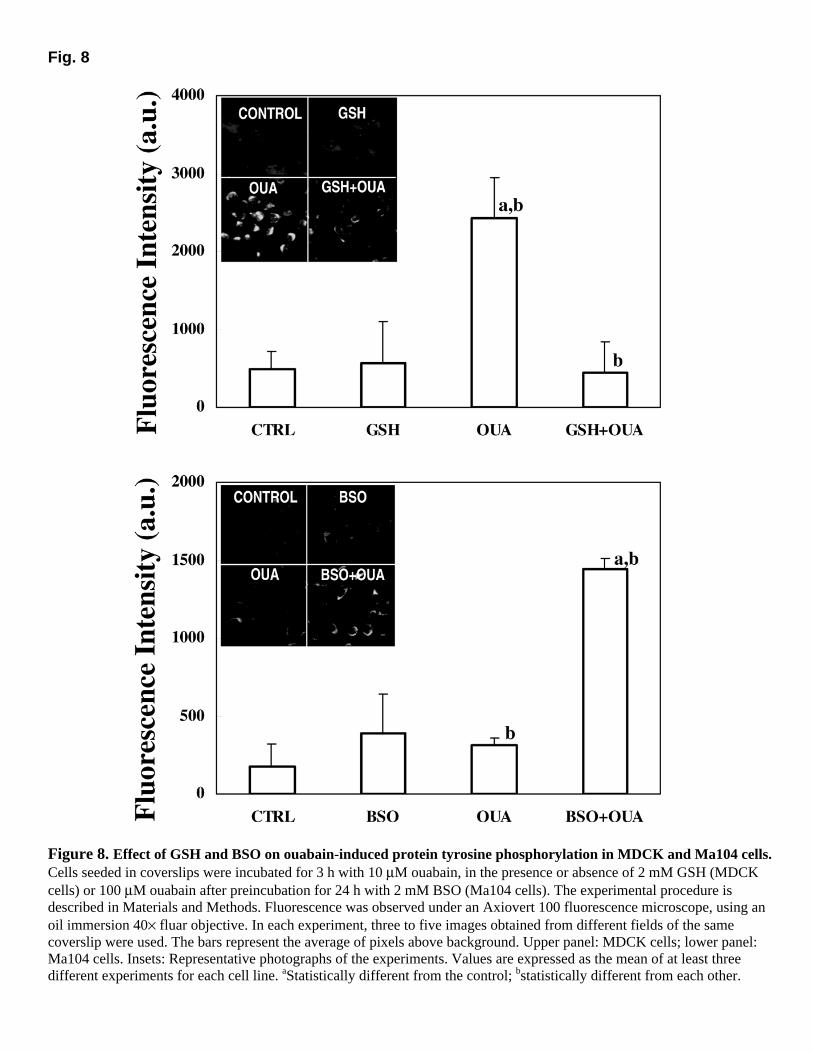
Fig. 8
Figure 8. Effect of GSH and BSO on ouabain-induced protein tyrosine phosphorylation in MDCK and Ma104 cells. Cells seeded in coverslips were incubated for 3 h with 10 µM ouabain, in the presence or absence of 2 mM GSH (MDCK cells) or 100 µM ouabain after preincubation for 24 h with 2 mM BSO (Ma104 cells). The experimental procedure is described in Materials and Methods. Fluorescence was observed under an Axiovert 100 fluorescence microscope, using an oil immersion 40× fluar objective. In each experiment, three to five images obtained from different fields of the same coverslip were used. The bars represent the average of pixels above background. Upper panel: MDCK cells; lower panel: Ma104 cells. Insets: Representative photographs of the experiments. Values are expressed as the mean of at least three different experiments for each cell line. aStatistically different from the control; bstatistically different from each other.
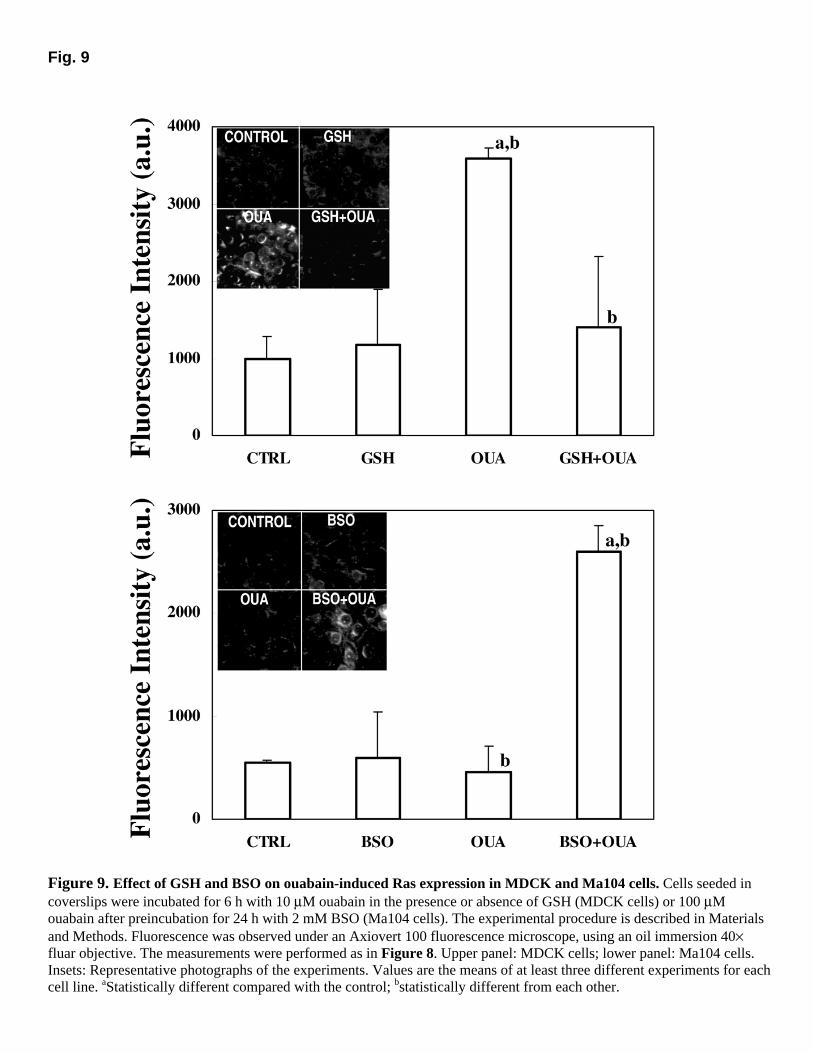
Fig. 9
Figure 9. Effect of GSH and BSO on ouabain-induced Ras expression in MDCK and Ma104 cells. Cells seeded in coverslips were incubated for 6 h with 10 µM ouabain in the presence or absence of GSH (MDCK cells) or 100 µM ouabain after preincubation for 24 h with 2 mM BSO (Ma104 cells). The experimental procedure is described in Materials and Methods. Fluorescence was observed under an Axiovert 100 fluorescence microscope, using an oil immersion 40× fluar objective. The measurements were performed as in Figure 8. Upper panel: MDCK cells; lower panel: Ma104 cells. Insets: Representative photographs of the experiments. Values are the means of at least three different experiments for each cell line. aStatistically different compared with the control; bstatistically different from each other.
Related Documents











