CHAPTER 12 TOXIC RESPONSES OF THE IMMUNE SYSTEM Leigh Ann Burns-Naas, B. Jean Meade, and Albert E. Munson THE IMMUNE SYSTEM Innate Immunity General Considerations Cellular Components: NK, PMN, Macrophage Soluble Factors: Acute-Phase Proteins and Complement Acquired (Adaptive) Immunity General Considerations Cellular Components: APCs, T Cells, B Cells Humoral and Cell-Mediated Immunity Neuroendocrine Immunology ASSESSMENT OF IMMUNOLOGIC INTEGRITY Methods to Assess Immunocompetence General Assessment Functional Assessment Regulatory Approaches to the Assessment of Immunotoxicity The NTP Tier Approach Health Effects Test Guidelines Immunotoxicity Testing of Medical Devices Animal Models in Immunotoxicology Relationship between Immunotoxicity Data in Animals and Humans Evaluation of Mechanism of Action IMMUNOMODULATION BY XENOBIOTICS Immunosuppression Halogenated Aromatic Hydrocarbons Polycyclic Aromatic Hydrocarbons Nitrosamines Pesticides Metals Inhaled Substances Organic Solvents and Related Chemicals Mycotoxins Natural and Synthetic Hormones Therapeutic Agents Drugs of Abuse Electromagnetic Fields Ultraviolet Radiation Food Additives Silicon-Based Materials Immune-Mediated Disease Hypersensitivity Autoimmunity NEW FRONTIERS AND CHALLENGES Molecular Biology Methods: Proteomics and Genomics Animal Models: Transgenics and SCID Developmental Immunotoxicology Systemic Hypersensitivity Computational Toxicology Biomarkers Risk Assessment CONCLUDING COMMENT AND FUTURE DIRECTIONS Immunity, by definition, is a homeostatic condition in which the body maintains protection from infectious disease. It is a series of delicately balanced, complex, multicellular, and physiologic mech- anisms that allow an individual to distinguish foreign material from “self” and to neutralize and/or eliminate the foreign matter. It is characterized by a virtually infinite repertoire of specificities, highly specialized effectors, complex regulatory mechanisms, and an ability to travel throughout the body. The immune system pro- vides the means to initiate rapid and highly specific responses against a myriad of potentially pathogenic organisms. Indeed, the conditions of genetically determined immunodeficiency and of ac- quired immunodeficiency syndrome (AIDS) graphically highlight the importance of the immune system in the host’s defense against microbial infection. In addition, evidence is rapidly building that the immune system plays a role in tumor identification and rejec- tion (immune surveillance). In light of the central role that the immune system plays in the maintenance of the health of the individual, the interaction of xenobiotics (pharmacologic agents, environmental contaminants, and other chemicals) with the various components of the immune system has become an area of profound interest. Indeed, in some instances, the immune system has been shown to be compromised (decreased lymphoid cellularity, alterations in lymphocyte sub- populations, decreased host resistance, altered specific immune function responses) in the absence of observed toxicity in other or- gan systems. Decreased immunocompetence (immunosuppression) may result in repeated, more severe, or prolonged infections as well as the development of cancer. Immunoenhancement may lead to immune-mediated diseases such as hypersensitivity responses or autoimmune disease (Fig. 12-1). Because of the potentially pro- found effects resulting from disruption of the delicately balanced immune system, there is a need to understand the cellular, bio- chemical, and molecular mechanisms of xenobiotic-induced im- munomodulation. With the availability of sensitive, reproducible, and predictive tests, it is now apparent that the inclusion of im- munotoxicity testing may represent a significant adjunct to routine 419 Copyrighted Material Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

S4 ch12 toxic_responsesoftheimmunesystem
Aug 20, 2015
Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

CHAPTER 12
TOXIC RESPONSES OF THE IMMUNE SYSTEM
Leigh Ann Burns-Naas, B. Jean Meade, and Albert E. Munson
THE IMMUNE SYSTEM
Innate ImmunityGeneral ConsiderationsCellular Components: NK, PMN, MacrophageSoluble Factors: Acute-Phase Proteins
and ComplementAcquired (Adaptive) Immunity
General ConsiderationsCellular Components: APCs, T Cells, B Cells
Humoral and Cell-Mediated ImmunityNeuroendocrine Immunology
ASSESSMENT OF IMMUNOLOGIC INTEGRITY
Methods to Assess ImmunocompetenceGeneral AssessmentFunctional Assessment
Regulatory Approaches to the Assessment of ImmunotoxicityThe NTP Tier ApproachHealth Effects Test GuidelinesImmunotoxicity Testing of Medical Devices
Animal Models in ImmunotoxicologyRelationship between Immunotoxicity Data
in Animals and HumansEvaluation of Mechanism of Action
IMMUNOMODULATION BY XENOBIOTICS
ImmunosuppressionHalogenated Aromatic Hydrocarbons
Polycyclic Aromatic HydrocarbonsNitrosaminesPesticidesMetalsInhaled SubstancesOrganic Solvents and Related ChemicalsMycotoxinsNatural and Synthetic HormonesTherapeutic AgentsDrugs of AbuseElectromagnetic FieldsUltraviolet RadiationFood AdditivesSilicon-Based Materials
Immune-Mediated DiseaseHypersensitivityAutoimmunity
NEW FRONTIERS AND CHALLENGES
Molecular Biology Methods: Proteomics and Genomics
Animal Models: Transgenics and SCIDDevelopmental ImmunotoxicologySystemic HypersensitivityComputational ToxicologyBiomarkersRisk Assessment
CONCLUDING COMMENT AND FUTURE DIRECTIONS
Immunity, by definition, is a homeostatic condition in which thebody maintains protection from infectious disease. It is a series ofdelicately balanced, complex, multicellular, and physiologic mech-anisms that allow an individual to distinguish foreign material from“self” and to neutralize and/or eliminate the foreign matter. It ischaracterized by a virtually infinite repertoire of specificities,highly specialized effectors, complex regulatory mechanisms, andan ability to travel throughout the body. The immune system pro-vides the means to initiate rapid and highly specific responsesagainst a myriad of potentially pathogenic organisms. Indeed, theconditions of genetically determined immunodeficiency and of ac-quired immunodeficiency syndrome (AIDS) graphically highlightthe importance of the immune system in the host’s defense againstmicrobial infection. In addition, evidence is rapidly building thatthe immune system plays a role in tumor identification and rejec-tion (immune surveillance).
In light of the central role that the immune system plays inthe maintenance of the health of the individual, the interaction of
xenobiotics (pharmacologic agents, environmental contaminants,and other chemicals) with the various components of the immunesystem has become an area of profound interest. Indeed, in someinstances, the immune system has been shown to be compromised(decreased lymphoid cellularity, alterations in lymphocyte sub-populations, decreased host resistance, altered specific immunefunction responses) in the absence of observed toxicity in other or-gan systems. Decreased immunocompetence (immunosuppression)may result in repeated, more severe, or prolonged infections as wellas the development of cancer. Immunoenhancement may lead toimmune-mediated diseases such as hypersensitivity responses orautoimmune disease (Fig. 12-1). Because of the potentially pro-found effects resulting from disruption of the delicately balancedimmune system, there is a need to understand the cellular, bio-chemical, and molecular mechanisms of xenobiotic-induced im-munomodulation. With the availability of sensitive, reproducible,and predictive tests, it is now apparent that the inclusion of im-munotoxicity testing may represent a significant adjunct to routine
419
2996R_ch12_419-470 4/27/01 10:01 AM Page 419
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

420 UNIT 4 TARGET ORGAN TOXICITY
safety evaluations for therapeutic agents, biological agents, andchemicals now in development.
This chapter provides (1) an overview of basic concepts inimmunology (structure, components, and functions), which are important to the understanding of the impact xenobiotics may have on the exposed individual; (2) a summary of selected cur-rent methods utilized to assess immune function; and (3) a briefreview of current information on the immunomodulation (im-munosuppression, hypersensitivity, and autoimmunity) induced by a variety of xenobiotics. This chapter is not meant to be an immunology textbook nor an exhaustive review of the mechan-isms of immunotoxicity of a myriad of xenobiotics. For detailedinformation on immunology, the reader is referred to two texts, thefirst edited by Paul (1999), the second edited by Roitt and col-leagues (1993). For a more comprehensive review of immunotox-icology, the reader is referred to texts edited by Dean and col-leagues (1994), Smialowicz and Holsapple (1996), and Lawrence(1997).
THE IMMUNE SYSTEM
Unlike most organ systems, the immune system has the uniquequality of not being confined to a single site within the body. Itcomprises numerous lymphoid organs (Table 12-1) and numerousdifferent cellular populations with a variety of functions. The bonemarrow and the thymus contain microenvironments capable of sup-porting the production of mature T and B lymphocytes and myeloidcells, such as macrophages and polymorphonuclear cells from non-functional precursors (stem cells), and are thus referred to as pri-mary lymphoid organs. With regard to T and B cells, key eventsthat occur in both primary and secondary organs are (1) acquisitionof the ability to recirculate and become localized in appropriateplaces in the periphery (homing capacity), (2) the ability to rec-ognize antigen (rearrangement of the T-cell receptor and the B-cellantigen receptor germline genes), and (3) the ability to interact withaccessory cells (through the expression of various cell surfacemolecules and the development of biochemical signaling pathways)to allow differentiation into both effector and memory cellpopulations.
The bone marrow is the site of origin of the pluripotent stemcell, a self-renewing cell from which all other hematopoietic cellsare derived (Figs. 12-2 and 12-6). During gestation, this cell is
found in the embryonic yolk sac and fetal liver; subsequently, itmigrates to the bone marrow. Within the bone marrow, the cells ofthe immune system developmentally “commit” to either the lym-phoid or myeloid lineages. Cells of the lymphoid lineage make afurther commitment to become either T cells or B cells. Becauseof their critical role in initiation and regulation of immune re-sponses, T cell precursors are programmed to leave the bone mar-row and migrate to the thymus, where they undergo “thymic edu-cation” for recognition of self and nonself. This is discussed inmore detail under “Cellular Components” in the section below ti-tled “Acquired (Adaptive) Immunity.”
Mature naive or virgin lymphocytes (those T and B cells thathave never undergone antigenic stimulation) are first brought intocontact with exogenously derived antigens within the highly or-ganized microenvironment of the spleen and lymph nodes, other-wise known as the secondary lymphoid organs. These organs canbe thought of as biological sieves. The spleen serves as a filter forthe blood, removing both foreign antigens and any circulating deadcells and cellular debris. The lymph nodes are part of a networkof lymphatic veins that filter antigens from the fluid surroundingthe tissues of the body. Key events that occur within the second-ary lymphoid organs are (1) specific antigen recognition in the con-text of the major histocompatibility complex (MHC) class II, (2)clonal expansion (proliferation) of antigen-specific cells, and (3)
Figure 12-1. Potential consequences of immunomodulation.
Table 12-1Organization of the Immune System: Lymphoid Tissue
CLASSIFICATION LYMPHOID ORGANS
Primary Bone marrowThymus
Secondary SpleenLymph nodes
Tertiary Skin-associated lymphoid tissue (SALT)Mucosal lamina propria (MALT)Gut-associated lymphoid tissue (GALT)Bronchial-associated lymphoid
tissue (BALT)Cells lining the genitourinary tract
2996R_ch12_419-470 4/27/01 10:01 AM Page 420
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

CHAPTER 12 TOXIC RESPONSES OF THE IMMUNE SYSTEM 421
differentiation of antigen-stimulated lymphocytes into effector andmemory cells.
Lymphoid tissues associated with the skin (skin-associatedlymphoid tissue, or SALT) and mucosal lamina propria (mucosa-associated lymphoid tissue, or MALT) can be classified as tertiarylymphoid tissues. Tertiary lymphoid tissues are primarily effectorsites where memory and effector cells exert immunologic and im-munoregulatory functions. Although in a broad interpretation thiswould include essentially all tissues of the body, tertiary lymphoidtissues are associated primarily with the surfaces lining the intes-tines (gut-associated lymphoid tissue, or GALT), respiratory tract(bronchial-associated lymphoid tissue, or BALT), and the geni-
tourinary tract, since these tissues have access directly to the ex-ternal environment. For extensive reviews of respiratory, mucosal,and dermal immunology and immunotoxicology, the reader is re-ferred to chapters in Smialowicz and Holsapple (1996) and inLawrence (1997).
Innate Immunity
General Considerations Mammalian immunity can be classifiedinto two functional divisions: innate immunity and acquired (adap-tive) immunity (Table 12-2). Innate immunity acts as a first line ofdefense against infectious agents, eliminating most potential
Figure 12-2. Development of the cellular components of the immune system.
Table 12-2Innate verses Acquired Immunity
CHARACTERISTIC INNATE IMMUNITY ACQUIRED IMMUNITY
Cells involved Polymorphonuclear cells T cells(PMN) B cells
Monocyte/macrophage MacrophagesNK cells NK cells
Primary soluble mediators Complement AntibodyLysozyme CytokinesAcute phase proteinsInterferon-�/�Other cytokines
Specificity of response None Yes (very high specificity)
Response enhanced by No Yesrepeated antigenchallenge
2996R_ch12_419-470 4/27/01 10:01 AM Page 421
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

422 UNIT 4 TARGET ORGAN TOXICITY
pathogens before significant infection occurs. It is characterized bybeing nonspecific and includes physical and biochemical barriersboth inside and outside of the body as well as immune cells de-signed for specific responses. Unlike acquired immunity, there isno immunologic memory associated with innate immunity. There-fore, in a normal healthy adult, the magnitude of the immune re-sponse to a foreign organism is the same for a secondary or terti-ary challenge as it is for the primary exposure.
Externally, the skin provides an effective barrier, as most or-ganisms cannot penetrate intact skin. Most infectious agents enterthe body through the respiratory system, gut, or genitourinary tract.Innate defenses present to combat infection from pathogens enter-ing through the respiratory system include mucus secreted alongthe nasopharynx, the presence of lysozyme in most secretions, andcilia lining the trachea and main bronchi. In addition, reflexes suchas coughing, sneezing, and elevation in body temperature are alsoa part of innate immunity. Pathogens that enter the body via the digestive tract are met with severe changes in pH (acid) with-in the stomach and a host of microorganisms living in the intes-tines.
Cellular Components: NK, PMN, Macrophage Two generaltypes of cells are involved in nonspecific (innate) host resistance:natural killer (NK) cells and professional phagocytes (Table 12-3).Like other immune cells, NK cells are derived from the bonemarrow stem cell. It is not yet clear exactly how the NK lineage
progresses; however, NK cells do possess several surface markerswhich have been used to define T cells, suggesting that the NKcell is a derivative of a lymphoid precursor cell. The vast majorityof NK cells express CD16 (Fc receptor for IgG) on their surface.Although apparently derived from a similar lineage as the T cell,NK cells do not express cell surface CD3 (T cell receptor-associated protein complex) or either chain of the T-cell receptor(TCR). NK cells are located primarily in the spleen, blood, andperitoneal exudate, although they are occasionally found in lymphnode tissue as well. For their part in innate immunity, NK cells canrecognize virally infected and malignant changes on the surface ofcells as well as the Fc portion of IgG on an antibody-coated tar-get cell. The latter recognition is utilized in cell-mediated immu-nity. Using surface receptors, the NK cell binds and undergoes cy-toplasmic reorientation so that cytolytic granules (perforins andenzymatic proteins) are localized near the target cell. These gran-ules are then expelled onto the surface of the target cell. The resultof this process is the induction of apoptosis (DNA fragmentation,membrane blebbing, and cellular disintegration) of the target cell.
Phagocytic cells include polymorphonuclear cells (PMN; neu-trophil) and the monocyte/macrophage. The precursors of themacrophage and PMN develop from pluripotent stem cells thathave become committed to the myeloid lineage (Fig. 12-2). Evi-dence exists that there are bipotentiating reactive precursors forPMN and macrophage and that differentiation into one or the other
Table 12-3Characteristics of Selected Immune Cells
MONOCYTE/PROPERTIES MACROPHAGE T CELLS B CELLS NK CELLS
Phagocytosis Yes No No NoAdherence Yes No No No
Surface receptors:Antigen receptors No Yes Yes NoComplement Yes No Yes YesFc region of Ig Yes Some Yes Yes
Surface markers CD64 CD4 Ig CD16CD11b CD8 Asialo-GM1
CD3 (mouse)Thy-1(mouse) CD11b
Proliferation in response to:Allogeneic cells (MLR) No Yes No NoLipopolysaccharide (LPS) No No Yes NoPhytohemagglutinin (PHA) No Yes No NoConcanavalin A (Con A) No Yes No NoAnti-Ig � IL-4 No No Yes NoAnti-CD3 � IL-2 No Yes No No
Effector functions:Antibody production No No Yes NoCytokine production Yes Yes Yes YesBactericidal activity Yes No No NoTumor cell cytotoxicity Yes Yes No YesImmunologic memory No Yes Yes No
SOURCE: Modified from Dean JH, Murray MJ: Toxic responses of the immune system, in Amdur MO, Doull J, Klaassen CD(eds): Casarett and Doull’s Toxicology: The Basic Science of Poisons, 4th ed. New York: Pergammon Press, 1991, p 286.
2996R_ch12_419-470 4/27/01 10:01 AM Page 422
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

CHAPTER 12 TOXIC RESPONSES OF THE IMMUNE SYSTEM 423
is dependent upon the interaction with specific colony-stimulatingfactors (CSFs) such as macrophage-CSF (M-CSF), granulocyte-CSF (G-CSF), granulocyte-macrophage-CSF (GM-CSF), inter-leukin-3 (IL-3), and others (Unanue, 1993). Within the bone marrow, both cell types undergo several rounds of replication be-fore entering the bloodstream where they circulate for about 10 hand then enter the tissues where they perform effector functionsfor about 1 to 2 days. PMNs are capable of passing through thecell membrane of the blood vessels and thereby represent a pri-mary line of defense against infectious agents. They are excellentphagocytic cells and can eliminate most microorganisms. Theirphagocytic activity is greatly enhanced by the presence of com-plement and antibody deposited on the surface of the foreign tar-get. They are also important in the induction of an inflammatoryresponse.
Macrophages are terminally differentiated monocytes. Uponexiting the bone marrow, monocytes circulate within the blood-stream for about 1 day. At that time, they begin to distribute to thevarious tissues where they can then differentiate into macrophages.Macrophages can be found in all tissues, most notably in the liver,lung, spleen, kidney, and brain. Within different tissues, mac-rophages have distinct properties and vary in extent of surface re-ceptors, oxidative metabolism, and expression of MHC class II.This is likely due to the factors present within the microenviron-ment in which the monocyte differentiates. The liver macrophages,or Kupffer cells, are primarily responsible for particulate and mi-crobial clearance from the blood. They express high levels of MHCclass II, are actively phagocytic, and release several soluble medi-ators. Thus, they are the primary cells responsible for the acutephase response. Alveolar macrophages remove foreign particulatematter from the alveolar space. They are self-renewing and have aparticularly long lifespan. These cells can be harvested by bron-cheoalveolar lavage and actively secrete proteases and bactericidalenzymes such as lysozyme. Splenic macrophages also phagocytoseparticulate material and polysaccharides from the blood and tissue.However, unlike other tissue macrophages, they are more diversewithin the tissue and their level of expression of MHC class II andtheir stage of differentiation appears to be dependent upon wherewithin the splenic architecture the macrophages are located.Mononuclear phagocytes within the central nervous system (CNS)are known as microglia and are responsible for antigen presenta-tion in immunologic diseases of the CNS. Microglia have a veryslow turnover time, and thus recruitment of monocytes to areas ofinflammation within the CNS is also slow.
Should PMNs be unable to contain an infection, macrophagesare then recruited to the site of infection. Although macrophagesare phagocytic by nature, their bactericidal activity can be aug-mented by lymphokines produced by T cells that recognize a spe-cific microbial antigen. Macrophages are unique cells within theimmune system because they play roles in both the innate arm ofimmunity (as phagocytic cells) and the acquired arm (as antigen-presenting cells). They adhere well to glass or plastic, are recruitedto sites of inflammation by chemotactic factors, can be activatedby cytokines to become more effective killers, and can produce cy-tokines, such as IL (interleukin)-1, IL-6, and TNF (tumor necrosisfactor), that act in a paracrine and autocrine fashion. Macrophagesalso play critical roles as scavengers in the daily turnover of senes-cent tissues such as red cell nuclei from maturing red cells, PMNs,and plasma cells. The importance of phagocytic cells to the or-ganism can be seen in individuals with spontaneous or induced re-duction in the numbers or activity of these cells. This condition is
associated with repeated and sometimes fatal bacterial and fungalinfections.
Soluble Factors: Acute-Phase Proteins and Complement Inaddition to the cellular components of innate immunity, there areseveral soluble components (Table 12-2). These are the acute-phaseresponse and the complement cascade. Upon infection, mac-rophages (Kupffer cells, in particular) become activated and se-crete a variety of cytokines, which are carried by the bloodstreamto distant sites. This global response to foreign agents is termedthe acute-phase response and consists of fever and large shifts inthe types of serum proteins synthesized by hepatocytes, such asserum amyloid A, serum amyloid P, and C-reactive protein. Theseproteins increase rapidly to concentrations up to 100 times the nor-mal concentration and stay elevated through the course of infec-tion. These proteins can bind to bacteria and facilitate the bindingof complement and the subsequent uptake of the bacteria by phago-cytic cells. This process of protein coating to enhance phagocyto-sis is termed opsonization.
The complement system is a series of about 30 serum pro-teins whose primary functions are the modification of membranesof infectious agents and the promotion of an inflammatory re-sponse. The components of the complement cascade interact witheach other and with other elements of both the innate and acquiredarms of immunity. Complement activation occurs with each com-ponent sequentially, acting on others in a manner similar to theblood clotting cascade (Fig. 12-3). Early components of the cas-cade are often modified serine proteases, which activate the sys-tem but have limited substrate specificity. Several components arecapable of binding to microbial membranes and serve as ligandsfor complement receptors associated with the membrane. The fi-nal components, which are related structurally, are also membrane-binding proteins that can enter into the membrane and disrupt mem-brane integrity (membrane attack complex). And finally, there areseveral regulatory complement proteins designed to protect the hostfrom inadvertent damage.
Figure 12-3. The complement cascade.
2996R_ch12_419-470 4/27/01 10:01 AM Page 423
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

424 UNIT 4 TARGET ORGAN TOXICITY
Two pathways have been identified in the complement cas-cade. The classic pathway is involved when antibody binds to themicroorganism. Because specific antibody defines the target, thisis a mechanism by which complement aids effectors of the acquiredside of immunity. The second, or alternative pathway, is used toassist the innate arm of immunity. For this cascade, it is not nec-essary for the host to have prior contact with the pathogen, sinceseveral microbial proteins can alone initiate this pathway. What-ever the mechanism of activation, the results are the same. Thecomplement-coated material is targeted for elimination by interac-tion with complement receptors on the surface of circulating im-mune cells.
Acquired (Adaptive) Immunity
General Considerations If the primary defenses against infec-tion (innate immunity) are breached, the acquired arm of the im-mune system is activated and produces a specific immune responseto each infectious agent, which usually eliminates the infection.This branch of immunity is also capable of remembering thepathogen and can protect the host from future infection by the sameagent. Therefore, the two key features which distinguish acquiredimmunity are specificity and memory. This means that in a normalhealthy adult, the speed and magnitude of the immune response toa foreign organism is greater for a secondary challenge than it isfor the primary challenge (Table 12-2). This is the principleexploited in vaccination.
Acquired immunity may be further subdivided into cell-mediated immunity (CMI) and humoral immunity. CMI, in itsbroadest sense, includes all immunologic activity in which anti-body plays a minimal role. Humoral immunity is directly depend-ent upon the production of antigen-specific antibody by B cells andinvolves the coordinated interaction of antigen-presenting cells, Tcells, and B cells. A more detailed discussion of both CMI and hu-moral immunity appears later.
Essential to the development of specific immunity is the recog-nition of antigen and the generation of an antibody that can bind toit. An antigen (sometimes referred to as an immunogen or allergen)is defined functionally as a substance that can elicit the productionof a specific antibody and can be specifically bound by that anti-body. Antigens are usually (but not absolutely) biological moleculesthat can be cleaved and rearranged for presentation. They may beeither proteins, carbohydrates (often bacterial), lipids, nucleic acids,or human-engineered substances, and they must be foreign (nonself)or occult (hidden, sequestered). Generally, antigens are about 10 kDaor larger in size. Smaller antigens are termed haptens and must beconjugated with carrier molecules (larger antigens) in order to elicita specific response. However, once a response is made, the haptencan interact with the specific antibody in the absence of the carrier.
Antibodies are produced by B cells and are also defined func-tionally by the antigen with which they react (i.e., anti-sheep redblood cell IgM, or anti-sRBC IgM). Because the immune systemgenerates antibody to thousands of antigens with which the hostmay or may not ever come into contact, general antibody of un-known specificity is referred to as immunoglobulin (e.g., serum im-munoglobulin or serum IgM) until it can be defined by its specificantigen (e.g., anti-sRBC IgM). A simple way to view this point isthat an antibody is an immunoglobulin, but immunoglobulin is notnecessarily antibody. There are five types of immunoglobulin thatare related structurally (Table 12-4): IgM, IgG (and subsets), IgE,IgD, and IgA. All immunoglobulins are made up of heavy and lightchains and of constant and variable regions. It is the variable re-gions that determine antibody specificity (Fig. 12-4). It is the vari-able region that interacts with antigen, while the Fc region medi-ates effector functions such as complement fixation (IgM and someIgG subclasses) and phagocyte binding (via Fc receptors). Anti-bodies subserve several functions in acquired immunity: (1) op-sonization (coating of a pathogen with antibody to enhance Fc re-ceptor-mediated endocytosis by phagocytic cells); (2) initiation ofthe classic pathway of complement-mediated lysis; (3) neutraliza-
Table 12-4Properties of Immunoglobulin Classes and Subclasses
MEAN SERUM HUMAN
CONCENTRATION, HALF-LIFE
CLASS mg/mL DAYS BIOLOGICAL PROPERTIES
IgG Complement fixation(selected subclasses)
Crosses placentaHeterocytotropic antibody
SubclassesIgG1 9 21IgG2 3 20IgG3 1 7IgG4 1 21
IgA 3 6 Secretory antibody
IgM 1.5 10 Complement fixationEfficient agglutination
IgD 0.03 3 Possible role in antigen-triggeredlymphocyte differentiation
IgE 0.0001 2 Allergic responses (mast celldegranulation)
2996R_ch12_419-470 4/27/01 10:01 AM Page 424
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

CHAPTER 12 TOXIC RESPONSES OF THE IMMUNE SYSTEM 425
tion of viral infection by binding to viral particles and preventingfurther infection; and (4) enhancement of the specificity of effec-tors of CMI by binding to specific antigens on target cells, whichare then recognized and eliminated by effector cells such as NKor cytotoxic T lymphocytes (CTL).
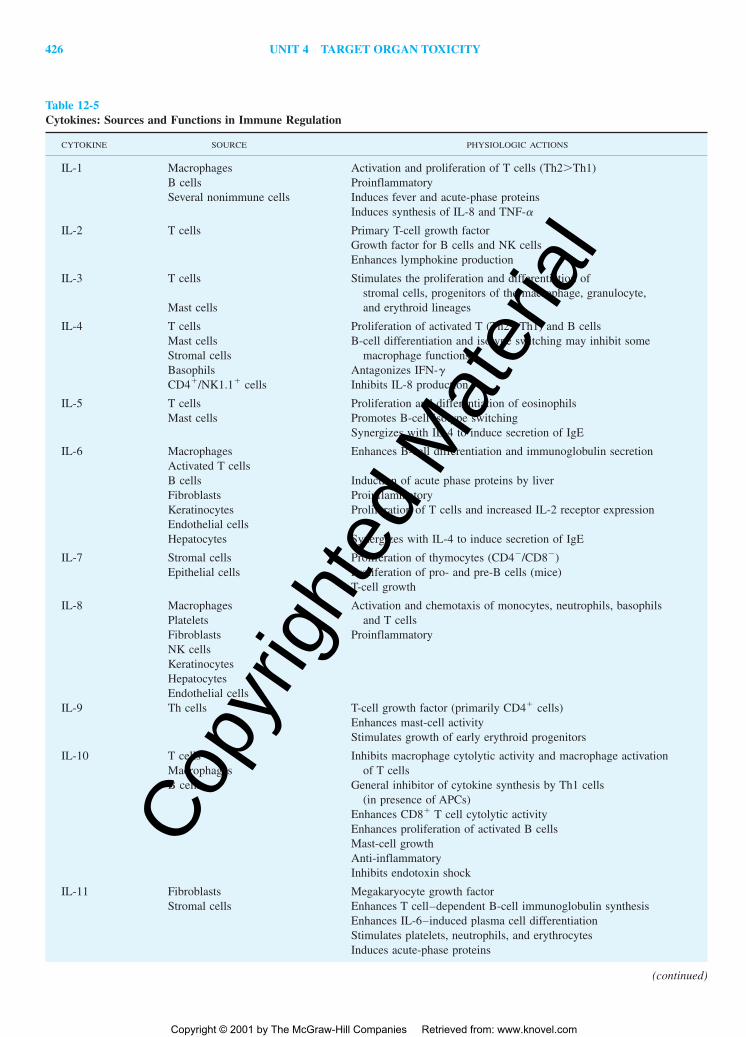
During an immune response, the cells of the immune systemmust be able to communicate to coordinate all the activities thatoccur during the recognition and elimination of foreign antigens.Connecting all the cells of the immune system with each other aswell as with other non-immune cell types within the body is a vastnetwork of soluble mediators: the cytokines. Nearly all immunecells secrete cytokines, which may have local or systemic effects.Table 12-5 provides a brief summary of the sources and functionsof cytokines of interest in the immune system. Although it wouldappear that many cytokines have related functions, these functionsare not often identical, and a single cytokine may have multiple ef-fects on a variety of cell types. Since cytokines work to tightly reg-ulate immune responses, some induce synthesis of other cytokinesand inflammatory mediators, while others inhibit this process. Al-though the actual number of cytokines (lymphokines, monokines,chemokines, etc.) may not be altogether that large, the complexityof the network is magnified severalfold by the multitude of bio-logical actions of each cytokine and the diversity of cells secret-ing each mediator.
Cellular Components: APCs, T Cells, B Cells In order to elicita specific immune response to a particular antigen, that antigenmust be taken up and processed by accessory cells for presenta-tion to lymphocytes. Accessory cells that perform this function are termed antigen-presenting cells (APCs) and include themacrophage, follicular dendritic cell (FDC), Langerhans dendriticcell, and B cells. A description of the macrophage is found in thesection above titled “Cellular Components of Innate Immunity”;however, the macrophage also plays a critical role as an APC in
acquired immunity. Unique among the APCs is the FDC. Unlikehematopoietic cells, the FDC is not derived from the bone marrowstem cell. It is found in secondary lymphoid organs and bindsantigen-antibody complexes, but it does not internalize and processthe antigen. Instead, the primary function of the FDC is in the per-sistence of antigen within the secondary lymphoid tissues and thepresentation of antigen to B cells. This is believed to be critical forthe maintenance of memory for B cells and the induction of high-affinity B cell clones. Although thought of more for its ability toproduce immunoglobulin, the B cell can also serve as an APC, andin low antigen concentrations this cell is equally as competent asthe macrophage in serving this function. The Langerhans dendriticcell is also a bone marrow–derived cell, but its lineage is distinctfrom that of the macrophage. It is found primarily in the epider-mis, mucosal epithelium, and lymphoid tissues. The Langerhansdendritic cell can migrate into the lymphatic system, where it servesas an APC in the lymph nodes. This cell plays a primary role incontact sensitization.
The interaction of APCs and lymphocytes is critical for thedevelopment of an immune response. With the exception of theFDC, APCs internalize the antigen either by phagocytosis, pinocy-tosis, or receptor-mediated endocytosis (via antigen, Fc, or com-plement receptors). Following internalization, antigen is processed(intracellular denaturation and catabolism) through several cyto-plasmic compartments, and a piece of the antigen (peptide frag-ments about 20 amino acids in length) becomes physicallyassociated with major histocompatibility complex (MHC) class II(Fig. 12-5). This MHC class II-peptide complex is then transportedto the surface of the cell and can interact in a specific manner withlymphocytes. For most APCs, an immunogenic determinant is ex-pressed on the surface of the APC within an hour after internal-ization, although this is slightly longer for B cells (3 to 4 h). In ad-dition to processing and presentation, pieces of processed antigenmay be expelled into the extracellular space. These pieces ofprocessed antigen can then bind in the peptide groove of emptyMHC class II on the surface of other APCs for the presentationof that peptide fragment to lymphocytes.
Not only are B lymphocytes capable of serving as APCs, butthey are also the effector cells of humoral immunity, producing anumber of isotypes of immunoglobulin (Ig) with varying speci-ficities and affinities. Like other immune cells, the B cell developsin the bone marrow from the pluripotent stem cell and becomescommitted to the B-cell lineage when the cell begins to rearrangeits Ig genes (Fig. 12-6). If, after several attempts, the cell is un-
Figure 12-4. Immunoglobulin structure.
Figure 12-5. General schematic of antigen processing and presentation.
2996R_ch12_419-470 4/27/01 10:01 AM Page 425
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com
426 UNIT 4 TARGET ORGAN TOXICITY
Table 12-5Cytokines: Sources and Functions in Immune Regulation
CYTOKINE SOURCE PHYSIOLOGIC ACTIONS
IL-1 Macrophages Activation and proliferation of T cells (Th2�Th1)B cells ProinflammatorySeveral nonimmune cells Induces fever and acute-phase proteins
Induces synthesis of IL-8 and TNF-�
IL-2 T cells Primary T-cell growth factorGrowth factor for B cells and NK cellsEnhances lymphokine production
IL-3 T cells Stimulates the proliferation and differentiation ofstromal cells, progenitors of the macrophage, granulocyte,
Mast cells and erythroid lineages
IL-4 T cells Proliferation of activated T (Th2�Th1) and B cellsMast cells B-cell differentiation and isotype switching may inhibit someStromal cells macrophage functionsBasophils Antagonizes IFN-�CD4�/NK1.1� cells Inhibits IL-8 production
IL-5 T cells Proliferation and differentiation of eosinophilsMast cells Promotes B-cell isotype switching
Synergizes with IL-4 to induce secretion of IgE
IL-6 Macrophages Enhances B-cell differentiation and immunoglobulin secretionActivated T cellsB cells Induction of acute phase proteins by liverFibroblasts ProinflammatoryKeratinocytes Proliferation of T cells and increased IL-2 receptor expressionEndothelial cellsHepatocytes Synergizes with IL-4 to induce secretion of IgE
IL-7 Stromal cells Proliferation of thymocytes (CD4�/CD8�)Epithelial cells Proliferation of pro- and pre-B cells (mice)
T-cell growth
IL-8 Macrophages Activation and chemotaxis of monocytes, neutrophils, basophils Platelets and T cellsFibroblasts ProinflammatoryNK cellsKeratinocytesHepatocytesEndothelial cells
IL-9 Th cells T-cell growth factor (primarily CD4� cells)Enhances mast-cell activityStimulates growth of early erythroid progenitors
IL-10 T cells Inhibits macrophage cytolytic activity and macrophage activationMacrophages of T cellsB cells General inhibitor of cytokine synthesis by Th1 cells
(in presence of APCs)Enhances CD8� T cell cytolytic activityEnhances proliferation of activated B cellsMast-cell growthAnti-inflammatoryInhibits endotoxin shock
IL-11 Fibroblasts Megakaryocyte growth factorStromal cells Enhances T cell–dependent B-cell immunoglobulin synthesis
Enhances IL-6–induced plasma cell differentiationStimulates platelets, neutrophils, and erythrocytesInduces acute-phase proteins
(continued)
2996R_ch12_419-470 4/27/01 10:01 AM Page 426
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

CHAPTER 12 TOXIC RESPONSES OF THE IMMUNE SYSTEM 427
(continued)
Table 12-5Cytokines: Sources and Functions in Immune Regulation (continued)
CYTOKINE SOURCE PHYSIOLOGIC ACTIONS
IL-12 Macrophages Proliferation and cytolytic action of NK cellsB cells Activation, proliferation, and cytolytic action of CTL
Stimulates production of IFN-�Proliferation of activated T cellsDecreases IgG1 and IgE primary response
IL-13 T cells Stimulates class II expression on APCEnhances antigen processing by APCEnhances B-cell differentiation and isotype switchingAnti-inflammatory (inhibits synthesis of proinflammatory
cytokines)Inhibits antibody-dependent cellular cytotoxicity (ADCC)
IL-14 T cells Enhances B-cell proliferationSome malignant B cells Inhibition of immunoglobulin secretion
Selective expansion of some B-cell subpopulations
IL-15 Activated monocytes NK-cell activationMacrophages T-cell proliferationSeveral nonimmune cells Mast-cell growth
IL-16 T cells Chemoattractant for T cells, eosinophils, and monocytesMast cells Promotes CD4� T-cell adhesionEosinophils Increases expression of IL-2 receptor
Promotes synthesis of IL3, GM-CSF, and IFN-�ProinflammatoryMay exacerbate allergic reactions
IL-17 CD4� memory T cells Induced production of IL-6, IL-8, G-CSF, and PGE2
Enhances proliferation of activated T cellsInducer of stromal cell–derived proinflammatory cytokinesInducer of stromal cell–derived hematopoietic cytokines
IL-18 Hepatocytes Synergizes with IL-12 to enhance the activity of Th1 cellsEnhances production of IFN-�
Interferon-�/� Leukocytes Induction of class I expression(IFN-�/�) Epithelial cells Antiviral activity(Type 1 IFN) Fibroblasts Stimulation of NK cells
Interferon-� T cells Induction of class I and II(IFN-�) NK cells Activates macrophages (as APC and cytolytic cells)
Epithelial cells Improves CTL recognition of virally infected cellsFibroblasts
Tumor necrosis Macrophages Induces inflammatory cytokinesfactor (TNF-�) Lymphocytes Increases vascular permeabilityand Mast cells Activates macrophages and neutrophilslymphotoxin Tumor necrosis (direct action)
(TNF-�) Primary mediator of septic shockInterferes with lipid metabolism (result is cachexia)Induction of acute phase proteins
Transforming Macrophages Enhances monocyte/macrophage chemotaxisgrowth factor-� Megakaryocytes Enhances wound healing: angiogenesis, fibroblast
(TGF-�) Chondrocytes proliferation, deposition of extracellular matrixInhibits T- and B-cell proliferationInhibits macrophage cytokine synthesisInhibits antibody secretionPrimary inducer of isotype switch to IgA
GM-CSF T cells Stimulates growth and differentiation of monocytes andMacrophages granulocytesEndothelial cellsFibroblasts
2996R_ch12_419-470 4/27/01 10:01 AM Page 427
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

428 UNIT 4 TARGET ORGAN TOXICITY
Table 12-5Cytokines: Sources and Functions in Immune Regulation (continued)
CYTOKINE SOURCE PHYSIOLOGIC ACTIONS
Migration T cells Inhibits macrophage migrationinhibitory Anterior pituitary cells Proinflammatory (induces TNF-� production by macrophages)factor (MIF) Monocytes Appears to play a role in delayed hypersensitivity responses
May be a counterregulator of glucocorticoid activityErythropoietin Endothelial cells Stimulates maturation of erythrocyte precursors
(EPO) Fibroblasts
SOURCE: Information on selected cytokines taken from Ruddle (1992), Quesniaux (1992), Paul and Seder (1994), Zurawski and de Vries (1994), Lawrence (1997), and Paul(1999).
successful at rearranging its Ig genes, it dies. Following Ig re-arrangement, these cells express heavy chains in their cytoplasmand are termed pre-B cells. Expression of surface IgM and IgD in-dicates a mature B cell. Mature B cells are found in the lymphnodes, spleen, and peripheral blood. Upon antigen binding to sur-face Ig, the mature B cell becomes activated and, after prolifera-tion, undergoes differentiation into either a memory B cell or anantibody-forming cell (AFC; plasma cell), actively secretingantigen-specific antibody. A broad description of several B-cellcharacteristics can be found in Table 12-3.
At a specified time following their commitment to the T-celllineage, pre-T cells migrate from the bone marrow to the thymuswhere, in a manner analogous to their B-cell cousins, they beginto rearrange their TCRs (Fig. 12-6). This receptor consists of twochains (� and �, or � and �) and is critical for the recognition ofMHC � peptide on APCs. At this time, the T cells begin to ex-press the surface marker CD8. CD8 (and CD4) are coreceptors ex-pressed by T cells and are involved in the interaction of the T cellwith the APC. T cells bearing the ��� TCR subsequently lose ex-pression of CD8 and proceed to the periphery. T cells with the���TCR gain surface expression of both the TCR and CD4 andare termed immature double-positive cells (CD4�/CD8�). Theseimmature cells then undergo positive selection to eliminate cellsthat cannot interact with MHC. Following this interaction,TCR expression increases. Any of these T cells that interact withMHC � self peptide are then eliminated (negative selection). Thedouble-positive cells then undergo another selection processwhereby they lose expression of either CD4 or CD8 and then pro-ceed to the periphery as mature single-positive cells (CD4� orCD8�) with a high level of TCR expression. This rigorous selec-tion process produces T cells that can recognize MHC � foreignpeptides and eliminates autoreactive T cells. Generally, T cells thatexpress CD8 mediate cell killing (CTL) or suppressor activity (Tsuppressor cells). Lymphocytes that participate in delayed hyper-sensitivity reaction (DHR) or that provide “B-cell help” in humoralresponses (helper T cells; Th1 and Th2) express CD4 on their sur-face. A broad description of several T-cell characteristics can befound in Table 12-3.
Humoral and Cell-Mediated Immunity
The activation of antigen-specific T cells begins with the interac-tion of the T-cell receptor with MHC class II � peptide. This in-teraction is strengthened by the presence of co-receptors such as
CD4, LFA-3, CD2, LFA-1, and ICAM-1 and involves the bilateralexchange of information, triggering a cascade of biochemicalevents that ultimately leads to the activation of not only the T cellbut the APC as well. Although the macrophage or dendritic cell istraditionally thought of as the APC involved in humoral responses,B cells can also subserve this function. In fact, many believe that,in low antigen concentrations, the B cell serves as the primary APCbecause of the presence of the high-affinity Ig receptor on the sur-face of the B cell.
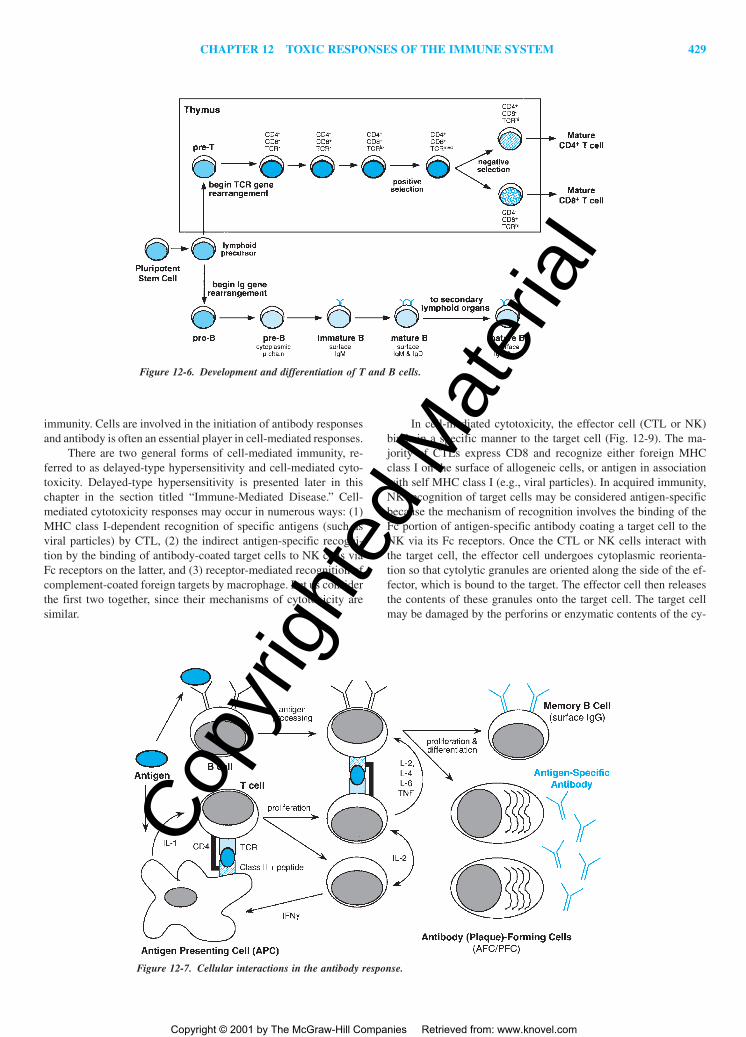
Upon activation and in the presence of IL-1 secreted by theAPC, T cells begin to express high-affinity receptors for the ma-jor T-cell growth factor, IL-2. In addition, T cells begin to produceIL-2, which can act in an autocrine fashion (on IL-2 receptors onthe same T cell) or paracrine fashion (IL-2 receptors on other Tcells or on B cells). As T cells begin to undergo clonal expansion(proliferation), they secrete numerous lymphokines (cytokines se-creted from lymphocytes; Table 12-5) which can influence (1) thestrength of an immune response, (2) the down-regulation of the im-mune response, (3) the isotype of antibody secreted by the AFC,(4) the activation of cells involved in cell-mediated immunity, and(5) the modulation of activities of numerous immune and nonim-mune cells. The next step in the generation of the humoral responseis the interaction of activated T cells with B cells. This may be adirect interaction of the T cell with B cell (antigen-specific) or maysimply involve the production of lymphokines (such as IL-2,IL-4, IL-6, and TNF-� and TNF-�), which lead to B-cell growthand differentiation into AFCs or memory B cells. A general dia-gram of the cellular interactions involved in the humoral immuneresponse is given in Fig. 12-7. The production of antigen-specificIgM requires 3 to 5 days after the primary (initial) exposure to anti-gen (Fig. 12-8). Upon secondary antigen challenge, the B cells un-dergo isotype switching, producing primarily IgG antibody, whichis of higher affinity. In addition, there is a higher serum antibodytiter associated with a secondary antibody response.
Cell-mediated immunity (CMI), in its broadest sense, includesall immunologic activity in which antibody plays a minimal role.However, for purposes of discussion here, CMI is more specifi-cally defined as the T-cell–mediated response such as DHR or CTLactivity, antibody-dependent cellular cytotoxicity (ADCC) medi-ated by NK cells, and soluble factor–mediated macrophage cyto-toxic responses. Whether an antigen will elicit a primarily cell-mediated or humoral response (or a combination of both) is de-pendent upon numerous factors. However, it should be noted thatthere is often an interplay between these two branches of acquired
2996R_ch12_419-470 4/27/01 10:01 AM Page 428
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com
CHAPTER 12 TOXIC RESPONSES OF THE IMMUNE SYSTEM 429
immunity. Cells are involved in the initiation of antibody responsesand antibody is often an essential player in cell-mediated responses.
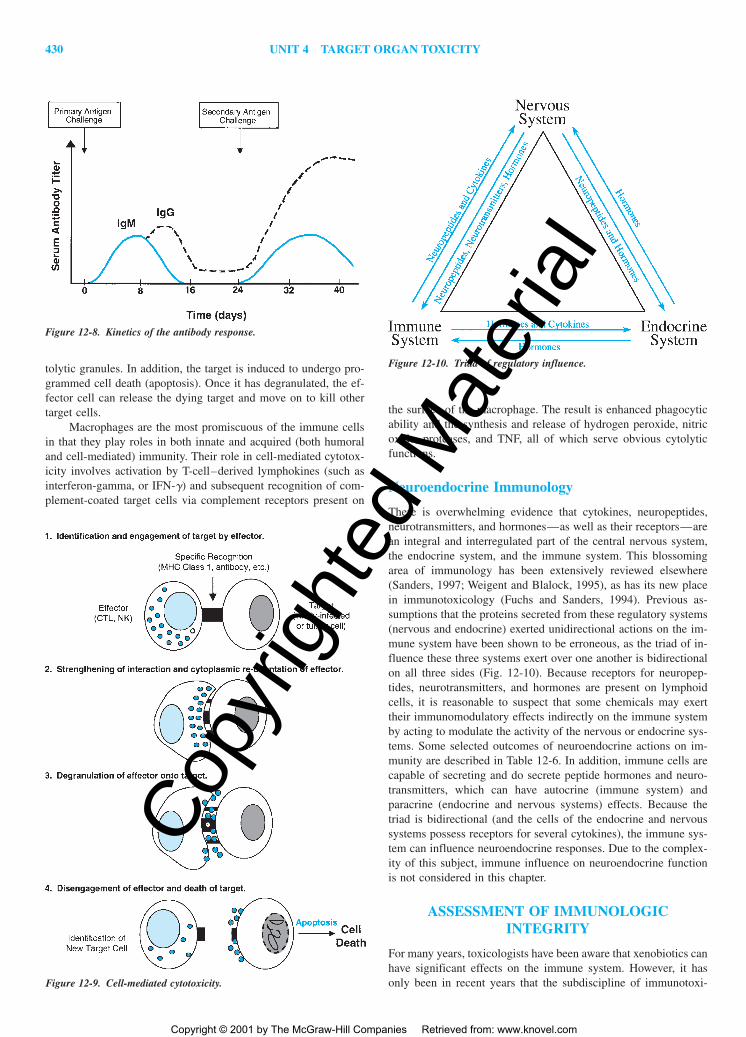
There are two general forms of cell-mediated immunity, re-ferred to as delayed-type hypersensitivity and cell-mediated cyto-toxicity. Delayed-type hypersensitivity is presented later in thischapter in the section titled “Immune-Mediated Disease.” Cell-mediated cytotoxicity responses may occur in numerous ways: (1)MHC class I-dependent recognition of specific antigens (such asviral particles) by CTL, (2) the indirect antigen-specific recogni-tion by the binding of antibody-coated target cells to NK cells viaFc receptors on the latter, and (3) receptor-mediated recognition ofcomplement-coated foreign targets by macrophage. Let us considerthe first two together, since their mechanisms of cytotoxicity aresimilar.
In cell-mediated cytotoxicity, the effector cell (CTL or NK)binds in a specific manner to the target cell (Fig. 12-9). The ma-jority of CTLs express CD8 and recognize either foreign MHCclass I on the surface of allogeneic cells, or antigen in associationwith self MHC class I (e.g., viral particles). In acquired immunity,NK recognition of target cells may be considered antigen-specificbecause the mechanism of recognition involves the binding of theFc portion of antigen-specific antibody coating a target cell to theNK via its Fc receptors. Once the CTL or NK cells interact withthe target cell, the effector cell undergoes cytoplasmic reorienta-tion so that cytolytic granules are oriented along the side of the ef-fector, which is bound to the target. The effector cell then releasesthe contents of these granules onto the target cell. The target cellmay be damaged by the perforins or enzymatic contents of the cy-
Figure 12-6. Development and differentiation of T and B cells.
Figure 12-7. Cellular interactions in the antibody response.
2996R_ch12_419-470 4/27/01 10:01 AM Page 429
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com
430 UNIT 4 TARGET ORGAN TOXICITY
tolytic granules. In addition, the target is induced to undergo pro-grammed cell death (apoptosis). Once it has degranulated, the ef-fector cell can release the dying target and move on to kill othertarget cells.
Macrophages are the most promiscuous of the immune cellsin that they play roles in both innate and acquired (both humoraland cell-mediated) immunity. Their role in cell-mediated cytotox-icity involves activation by T-cell–derived lymphokines (such asinterferon-gamma, or IFN-�) and subsequent recognition of com-plement-coated target cells via complement receptors present on
Figure 12-8. Kinetics of the antibody response.
Figure 12-9. Cell-mediated cytotoxicity.
Figure 12-10. Triad of regulatory influence.
the surface of the macrophage. The result is enhanced phagocyticability and the synthesis and release of hydrogen peroxide, nitricoxide, proteases, and TNF, all of which serve obvious cytolyticfunctions.
Neuroendocrine Immunology
There is overwhelming evidence that cytokines, neuropeptides,neurotransmitters, and hormones—as well as their receptors—arean integral and interregulated part of the central nervous system,the endocrine system, and the immune system. This blossomingarea of immunology has been extensively reviewed elsewhere(Sanders, 1997; Weigent and Blalock, 1995), as has its new placein immunotoxicology (Fuchs and Sanders, 1994). Previous as-sumptions that the proteins secreted from these regulatory systems(nervous and endocrine) exerted unidirectional actions on the im-mune system have been shown to be erroneous, as the triad of in-fluence these three systems exert over one another is bidirectionalon all three sides (Fig. 12-10). Because receptors for neuropep-tides, neurotransmitters, and hormones are present on lymphoidcells, it is reasonable to suspect that some chemicals may exerttheir immunomodulatory effects indirectly on the immune systemby acting to modulate the activity of the nervous or endocrine sys-tems. Some selected outcomes of neuroendocrine actions on im-munity are described in Table 12-6. In addition, immune cells arecapable of secreting and do secrete peptide hormones and neuro-transmitters, which can have autocrine (immune system) andparacrine (endocrine and nervous systems) effects. Because thetriad is bidirectional (and the cells of the endocrine and nervoussystems possess receptors for several cytokines), the immune sys-tem can influence neuroendocrine responses. Due to the complex-ity of this subject, immune influence on neuroendocrine functionis not considered in this chapter.
ASSESSMENT OF IMMUNOLOGICINTEGRITY
For many years, toxicologists have been aware that xenobiotics canhave significant effects on the immune system. However, it hasonly been in recent years that the subdiscipline of immunotoxi-
2996R_ch12_419-470 4/27/01 10:01 AM Page 430
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

CHAPTER 12 TOXIC RESPONSES OF THE IMMUNE SYSTEM 431
cology has come into its own, with a battery of tests to evaluateimmunocompetence. Among the unique features of the immunesystem is the ability of immune cells to be removed from the bodyand to function in vitro. This unique quality offers the toxicologistan opportunity to comprehensively evaluate the actions of xenobi-otics on the immune system by providing an excellent system fordissecting the cellular, biochemical, and molecular mechanisms ofaction of multitudes of xenobiotics. While standard toxicologicendpoints such as organ weights, cellularity, and enumeration ofcell subpopulations are important components in assessing immuneinjury, by far the most sensitive indicators of immunotoxicity arethe tests that challenge the various immune cells to respond func-tionally to exogenous stimuli (reviewed in White, 1992). This sec-tion focuses on selected in vivo and in vitro tests currently usedfor evaluating immunotoxicity.
Methods to Assess Immunocompetence
General Assessment Central to any series of studies evaluatingimmunocompetence is the inclusion of standard toxicologic stud-ies, because any immunologic findings should be interpreted inconjunction with effects observed on other target organs. Standardtoxicologic studies that are usually evaluated include body and se-lected organ weights, general observations of overall animal health,selected serum chemistries, hematologic parameters, and status ofthe bone marrow (ability to generate specific colony-formingunits). In addition, histopathology of lymphoid organs—such asthe spleen, thymus, and lymph nodes—may provide insight intopotential immunotoxicants. Because of the unique nature of the im-mune system, there are several experimental approaches that maybe taken to assess immunotoxicity and to evaluate the mechanismsof action of xenobiotics. These are depicted in Fig. 12-11 and varywith respect to in vivo or in vitro exposure, immunologic chal-lenge, or immunologic evaluation (immune assay). As an example,the plaque-forming cell assay [Fig. 12-11 (2)] is an ex vivo assaywhere xenobiotic exposure and antigen challenge occur in vivo andthe immune response is evaluated in vitro. In contrast [as depictedin Fig. 12-11 (4)], splenocytes can be removed from a naive ani-mal, exposed to xenobiotic and antigen in vitro, and evaluated in
vitro. An example of this would be the in vitro–generated PFC re-sponse (Mishell-Dutton assay; Mishell and Dutton, 1967).
Using fluorescently labeled monoclonal antibodies to cellsurface markers (Table 12-3) in conjunction with a flow cytome-ter, it is now possible to accurately enumerate lymphocyte sub-sets. Antibodies are available to the T-cell surface markers CD4,CD8, and CD3 (among others). Dual colored fluorochromes al-low cells to be stained for two markers simultaneously. In thismanner, the number of CD4� and CD8� cells can be determinedsimultaneously on a single sample of cells. In the thymus, thisdual staining also helps determine the number of CD4�/CD8�
(double positive) and CD4�/CD8� (double negative) cells resid-ing in this organ (Fig. 12-12). This gives the researcher insightinto which specific T-cell subsets are targeted and whether thexenobiotic may affect T-cell maturation. Antibodies available tosurface immunoglobulin (Ig) and to B220 (the CD45 phosphataseon B cells) help determine the numbers of B cells. Surface mark-ers can reveal significant alterations in lymphocyte subpopula-tions, and in many instances this is indicative of alterations inimmunologic integrity. Indeed, an indicator of AIDS is the changesobserved in CD4� T-cell numbers. Luster and coworkers (1992)have reported that, in conjunction with two or three functionaltests, the enumeration of lymphocyte subsets can greatly enhancethe detection of immunotoxic chemicals. However, it is importantto keep in mind that although surface marker analysis can indi-cate shifts in lymphocyte populations, functional analysis of theimmune system offers greater sensitivity for the detection of im-munotoxicity.
Flow cytometry can be useful in immunotoxicology as a toolto assess mechanism of action beyond evaluation of surface mark-ers. These include evaluation of the effects of chemicals in the cellcycle, intracellular free calcium, cellular viability assessment, in-duction of apoptosis, evaluation of p53, DNA strand breaks(TUNEL assay), membrane potential, intracellular pH, oxidativestress, and membrane lipophilicity (reviewed in Burchiel et al.,1997 and 1999). Flow cytometry has applications in cell sortingand high throughput screening, and there is interest in the utilityof this tool in the refinement of certain cellular immune and hypersensitivity assays. A key to acceptance of flow cytometric
Table 12-6Reported Influences of Neuroendocrine Factors on Immunity*
CYTOKINE NK MACROPHAGE T CELL HUMORAL
PRODUCTION ACTIVITY ACTIVITY ACTIVITY IMMUNITY
ACTH � � � � �Prolactin � ? � � �Growth Hormone � � � � �� Endorphins � � � � �� Endorphins � � � � �Enkephalins � � ? ? �Substance P ? � � � �hCG ? � ? � ?Chemical sympathectomy � ? ? �� �Norepinephrine ? ? ? � ��Epinephrine ? ? ? � ��
*Data from Madden et al. (1995); Blalock (1989); Weigent and Blalock (1995); Sanders (1997); and Lawrence and Kim (2000). KEY: ACTH, adrenocorticotropin; hCG, human chorionic gonadotropin; �, generally enhanced responses; �, generally decreased responses; ��, both enhanced and sup-
pressed responses have been reported and may depend on receptor types or subclass of chemical or time of exposure relative to antigen challenge; ?, generally unknownor not reported in the references utilized.
2996R_ch12_431 5/21/01 4:35 PM Page 431
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

432 UNIT 4 TARGET ORGAN TOXICITY
methods for immunotoxicity assessment is assay validation. Thisconcept is supported by one of the conclusions of a workshop focusing on the application of flow cytometry in immunotoxicityassessments. The workshop concluded that while immunopheno-typing certainly has a place in the field of immunotoxicology (e.g.,used in conjunction with functional tests to identify immunotoxicchemicals; as a method to assess mechanism of action), more re-search is needed in both the human and the animal before im-
munophenotyping alone is sufficiently validated for use in pre-dicting chemical-induced effects on human health (ILSI, 1999).
Functional Assessment Innate Immunity As described ear-lier, innate immunity encompasses all those immunologic re-sponses that do not require prior exposure to an antigen and thatare nonspecific in nature. These responses include recognition oftumor cells by NK cells, phagocytosis of pathogens by
Figure 12-11. Approaches to assessing the immunotoxicity of xenobiotics.
Figure 12-12. Flow cytometry.
In this example, cells from the thymus are stained simultaneously with a fluorescent PE-conjugated (orange)antibody to CD8 and a fluorescent FITC-conjugated (green) antibody. When analyzed on a flow cytometer, theinstrument is requested to display a four-quadrant analysis (left). Increasing fluorescent intensity (brightness) isindicated by the arrows on each axis. The key to this analysis is displayed on the right. Cells which possess onlyCD8 fluoresce orange (CD8�) and are displayed in the upper left quadrant (light blue). Cells that possess onlyCD4 fluoresce green (CD4�) and are displayed in the lower right quadrant (medium blue). Cells that possessboth CD8 and CD4 fluoresce both orange and green (CD4�/CD8� ; double positives) and are displayed in theupper right quadrant (dark blue). Cells that do not possess either CD8 or CD4 do not fluoresce (CD4�/CD8�;double negatives) and are displayed in the lower left quadrant (black). The instrument can then be requested todetermine the percentage of cells in each quadrant. In a typical mouse thymus, there are approximately 8 to 13percent CD4�, 2 to 5 percent CD8�, 80- to 85 percent CD4�/CD8�, and 2 to 5 percent CD4�/CD8� cells.
2996R_ch12_419-470 4/27/01 10:01 AM Page 432
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

CHAPTER 12 TOXIC RESPONSES OF THE IMMUNE SYSTEM 433
macrophages, and the lytic activity of the components of the com-plement cascade.
To evaluate phagocytic activity, macrophages are harvestedfrom the peritoneal cavity [peritoneal exudate (PE) cells] and areallowed to adhere in 24-well tissue culture plates. The cells are then incubated with chromated chicken red blood cells (51Cr-cRBCs). Following incubation, the supernatant, containing 51Cr-cRBCs that have not been bound by macrophages, is removed.The cRBCs which are bound to the macrophages, but which havenot been phagocytized, are removed by a brief incubation with am-monium chloride. Finally, macrophages are lysed with NaOH andradioactivity in the lysate is counted to determine the amount ofphagocytosis that occurred. A set of control wells is needed to de-termine DNA content for each set of wells. Data are presented asa specific activity for adherence and phagocytosis (adhered orphagocytized cpm/DNA content) since xenobiotics altering adher-ence will have a significant effect on the results.
Another method to evaluate phagocytosis, but which does notrequire radioactivity, begins similarly to the 51Cr-cRBC assay.Peritoneal macrophages are allowed to adhere to each chamber ofa tissue culture slide. After adherence, macrophages are washedand incubated with latex covaspheres. At the end of incubation,cells are fixed in methanol and stained in methylene chloride.Macrophages containing five covaspheres or more are counted aspositive and data are expressed as a percentage of phagocytosis(the ratio of macrophages with 5 covaspheres to totalmacrophages counted).
The previous macrophage assays are conducted in vitro afterchemical exposure either in vivo or in vitro. If an in vivo assay ofthe ability of tissue macrophages to phagocytose a foreign antigenis required, the functional activity of the reticuloendothelial sys-tem can be evaluated. Intravenously injected radiolabeled sheep redblood cells (51Cr-sRBCs) are removed by the tissue macrophagefrom the circulation and sequestered for degradation in organs suchas the liver, spleen, lymph nodes, lung, and thymus. Clearance ofthe 51Cr-sRBCs is monitored by sampling of the peripheral blood.When steady state has been attained, animals are euthanized andorgans are removed and counted in a gamma counter to assess up-take of the 51Cr-sRBCs.
Evaluation of the ability of NK cells to lyse tumor cells isachieved using the YAC-1 cell line as a tumor target for an in vitrocytotoxicity assay. YAC-1 cells are radiolabeled with 51Cr and in-cubated (in 96-well microtiter plates) in specific effector-to-targetratios with splenocytes from xenobiotic-exposed and nonexposedanimals. During an incubation step, splenic NK cells (effectors)lyse the 51Cr-YAC-1 cells, releasing 51Cr into the supernatant. Atthe end of the incubation, plates are centrifuged and the supernatantis removed and counted on a gamma counter. After correcting forspontaneous release (which should be 10 percent), specific re-lease of 51Cr is calculated for each effector-to-target ratio and com-pared to the specific release from control animals.Acquired Immunity—Humoral The plaque-(antibody)formingcell (PFC or AFC) assay is a sensitive indicator of immunologicintegrity for several reasons. It is a test of the ability of the host tomount an antibody response to a specific antigen. When the par-ticulate T-dependent antigen (an antigen that requires T cells tohelp B cells make antibody) sheep erythrocytes (sRBCs) is used,this response requires the coordinated interaction of several dif-ferent immune cells: macrophages, T cells, and B cells. Therefore,an effect on any of these cells (e.g., antigen processing and pre-
sentation, cytokine production, proliferation, or differentiation) canhave a profound impact on the ability of B cells to produce antigen-specific antibody. Other antigens, termed T cell-independent anti-gens, such as DNP-Ficoll or TNP-LPS (lipopolysaccharide), canbe used that bypass the requirement for T cells in eliciting anti-body production by B cells.
A standard PFC assay involves immunizing control andxenobiotic-exposed mice either intravenously or intraperitoneallywith the sRBC. The antigen is taken up in the spleen and anantibody response occurs. Four days after immunization, spleensare removed and splenocytes are mixed with sRBC, complement,and agar. This mixture is plated onto petri dishes and covered witha cover slip. After the agar hardens the plates are incubated for 3 h at 37°C. During this time, B cells secrete anti-sRBC IgMantibody. When the IgM and complement coat the surrounding sRBCs, areas of hemolysis (plaques) appear which can be enu-merated (Fig. 12-13). At the center of each plaque is a single Bcell (antibody- or plaque-forming cell; AFC or PFC). Data are usu-ally presented as IgM PFC (or AFC) per million splenocytes. IgGPFC can also be enumerated by slight modifications of this sameassay. This isotype switching (from IgM to IgG) is important insecondary responses in which memory B cells respond morequickly to an antigen.
More recently, it has become evident that the PFC assay canbe evaluated in vivo using serum from peripheral blood of immu-nized mice and an enzyme-linked immunosorbent assay (ELISA;Fig. 12-14). Although the optimal response is delayed by 1 to 2days (compared to the PFC assay), this assay takes into accountantigen-specific antibody secreted by B cells in the spleen as wellas B cells residing in the bone marrow. Like the PFC assay, mice(or other experimental animals) are immunized with sRBCs and 6days later peripheral blood is collected. Serum from each sampleis serially diluted and incubated in microtiter plates that have beencoated with sRBC membranes. The membranes serve as the anti-gen to which sRBC-specific IgM or IgG will bind. After incuba-tion of the test sera and a wash step, an enzyme-conjugated mon-oclonal antibody (the secondary antibody) against IgM (or IgG) isadded. This antibody recognizes the IgM (or IgG) and binds specif-ically to that antibody. After incubation and a wash step, the en-zyme substrate (chromogen) is added. When the substrate comesinto contact with the enzyme on the secondary antibody, a colorchange occurs which can be detected by measuring absorbancewith a plate reader. Since this is a kinetic assay (color developsover time and is dependent upon concentration of anti-sRBCantibody in the test sera), it is important to establish controlconcentration–response curves so that data can be evaluated in thelinear range of the curve. Data are usually expressed in arbitraryoptical density (OD) units. Advantages of the ELISA over the PFCassay are the ability to conduct in vivo analyses and to attain agreater degree of flexibility, since serum samples can be storedfrozen for analysis at a later date.
One final assay measures the ability of B cells to undergoblastogenesis and proliferation, which are critical steps in the generation of an antibody response. This is achieved in micro-titer plates by stimulating splenocytes with a monoclonal anti-body to surface Ig (anti-Ig) in the presence of IL-4, or with the B-cell mitogen LPS. Proliferation is evaluated 2 to 3 days af-ter stimulation by measuring uptake of 3H-thymidine into the DNA of the cultured cells. Data are usually expressed as meancounts per minute for each treatment group. These studies are usu-
2996R_ch12_419-470 4/27/01 10:01 AM Page 433
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

434 UNIT 4 TARGET ORGAN TOXICITY
ally done in conjunction with T-cell proliferative responses de-scribed below.Humoral Immunity—Cell-Mediated While there are numerousassays used to assess cell-mediated immunity, three primary testsare used routinely in the National Toxicology Program (NTP) testbattery: the cytotoxic T-lymphocyte (CTL) assay, the delayed hypersensitivity response (DHR), and the T-cell proliferativeresponses to antigens (anti-CD3 � IL-2), mitogens (PHA andCon A), and allogeneic cell antigens (mixed lymphocyte responses;MLR).
The CTL assay measures the in vitro ability of splenic T cellsto recognize allogeneic target cells by evaluating the ability of theCTLs to proliferate and then lyse the target cells. Splenocytes areincubated with P815 mastocytoma cells, which serve as target cells.These target cells are pretreated with mitomycin C so that they can-not proliferate themselves. During this sensitization phase, theCTLs recognize the targets and undergo proliferation. Five daysafter sensitization, the CTLs are harvested and incubated in mi-crotiter plates with radiolabeled (51Cr) P815 mastocytoma cells.During this elicitation phase, the CTLs that have acquired mem-ory recognize the foreign MHC class I on the P815 cells and lysethe targets. At the end of the incubation, plates are centrifuged, thesupernatant is removed, and radioactivity released into the super-natant is counted on a gamma counter. After correcting for spon-taneous release, the percent cytotoxicity is calculated for eacheffector-to-target ratio and compared to that from control animals.
The DHR evaluates the ability of memory T cells to recog-nize foreign antigen, proliferate and migrate to the site of the anti-gen, and secrete cytokines which result in the influx of other in-flammatory cells. Like the PFC response, this assay is conductedcompletely in vivo. The assay itself quantitates the influx of radio-labeled monocytes into the sensitization site. During xenobiotic ex-posure, mice are sensitized twice with keyhole limpet hemocyanin(KLH) subcutaneously between the shoulders. On the last day ofexposure, mononuclear cells are labeled in vivo with an IV injec-tion of 125I-5-iododeoxyuridine (IUdR). One day later, mice arechallenged intradermally in one ear with KLH. Twenty-four hours
Figure 12-13. The plaque-forming cell (PFC) assay.
A. Demonstration of plaques (areas of hemolysis) that have formed within the lawn of sheep red blood cells,310 magnification. B. 3100 magnification of a plaque from panel A showing the B cell evident in the center ofthe plaque. (From photos by Dr. Tracey L. Spriggs, with permission.)
BA
Figure 12-14. Schematic diagram of a standard enzyme-linked im-munosorbent assay (ELISA).
2996R_ch12_419-470 4/27/01 10:01 AM Page 434
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

CHAPTER 12 TOXIC RESPONSES OF THE IMMUNE SYSTEM 435
after challenge, animals are euthanized, the ears are biopsied, andradiolabeled cells are counted in a gamma counter. Data are ex-pressed as a stimulation index which represents the cpm in the chal-lenged ear divided by the cpm in the unchallenged ear.
T cells play a central role in cell-mediated immunity and theability of T cells to undergo blastogenesis and proliferation is crit-ical to this role. Several mechanisms exist to evaluate proliferativecapacity. The mixed lymphocyte response (MLR) measures theability of T cells to recognize foreign MHC class I on splenocytesfrom an MHC-incompatible mouse (allogeneic cells) and undergoproliferation. For example, splenocytes from B6C3F1 mice (re-sponders) are incubated with splenocytes from mitomycin C-treated DBA/2 mice (stimulators). Proliferation is evaluated 4 to 5days after stimulation by measuring uptake of 3H-thymidine intothe DNA of the cultured responder cells. Cells are collected fromeach well using a cell harvester and counted in a scintillationcounter. Data may be expressed as either the mean cpm for eachtreatment group or as a stimulation index where the index is cal-culated by dividing the cpm of wells containing responders andstimulators by the cpm of wells containing responders alone.
General T cell proliferation can be evaluated in a manner sim-ilar to that described above for B cells (Table 12-3). Splenocytesare stimulated in microtiter plates with a monoclonal antibody tothe CD3 complex of the T-cell receptor (anti-CD3) in the presenceof IL-2, or with the T-cell mitogens concanavalin A (Con A) andphytohemagglutinin (PHA). Proliferation is evaluated 2 to 3 daysafter stimulation by measuring uptake of 3H-thymidine into theDNA of the cultured T cells. Data are usually expressed as meancpm for each treatment group. These studies are usually done inconjunction with B-cell proliferative responses described above.Host Resistance Assays Host resistance assays represent a wayof assaying how xenobiotic exposure affects the ability of the hostto handle infection by a variety of pathogens. Although host re-sistance studies provide significant insight into the mechanisms bywhich an immunotoxicant is acting, these assays should not be afirst or only choice for evaluating immunocompetence. An exam-ple of why this is true is the actions of the semiconductor materialgallium arsenide (GaAs) on the immune system. Although GaAsproduces profound immunosuppression of nearly all cell types eval-uated, this compound was observed to confer varying degrees ofprotection to challenge with Listeria monocytogenes and Strepto-coccus pneumoniae. It was subsequently determined that the cir-culating blood arsenic concentrations were sufficient to inhibitgrowth of both of these organisms. In host resistance studies, it isalso important to consider the following: (1) strain, route of ad-ministration, and challenge size of the pathogen; (2) strain, age,
and sex of the host; (3) physiologic state of the host and thepathogen; and (4) time of challenge with the pathogen (prior to,during, or after xenobiotic exposure). All of these can have signif-icant effects on the results from any individual study.
As with other immune function tests, no single host resistancemodel can predict overall immunocompetence of the host, prima-rily because each model uses different mechanisms for eliminationof various pathogens. A representative list of host resistance mod-els is shown in Table 12-7 as well as some of the cells involved inthe immune response to these pathogens. Typically, three challengelevels of pathogen (approximating the LD20, LD50, and LD80) foreach concentration of xenobiotic are used in order to be able to de-tect both increases and decreases in resistance. Endpoint analysesare lethality (for bacterial and viral pathogens), changes in tumorburden, and increased or decreased parasitemia.
Regulatory Approaches to theAssessment of Immunotoxicity
The NTP Tier Approach Luster and colleagues (1988) have de-scribed the selection of a battery of tests used by the National Tox-icology Program to screen for potential immunotoxic agents. Theresult was a tier approach to assessing immunotoxicity and is sum-marized in Table 12-8. Tier I provides assessment of general tox-icity (immunopathology, hematology, body and organ weights) aswell as endline functional assays (proliferative responses, PFC as-say, and NK assay). It was designed to detect potential immuno-toxic compounds at concentrations that do not produce overt tox-icity. Tier II was designed to further define an immunotoxic effectand includes tests for cell-mediated immunity (CTL and DHR),secondary antibody responses, enumeration of lymphocyte popu-lations, and host resistance models. Subsequently, several testingconfigurations were defined that would minimize the number ofimmune tests needed, yet still provide a high degree of sensitivityfor detecting potential immunotoxicants. These configurations aredepicted in Table 12-9. The FDA has adopted a tier approach inits assessment of the immunotoxicity of food and color additives(Redbook I).
Health Effects Test Guidelines After several years of interna-tional debate regarding inclusion of functional immunotoxicity as-sessments in regulatory studies (as opposed to relying onhistopathology as an indicator of further testing needs), the Envi-ronmental Protection Agency published health effects test guide-lines for immunotoxicity testing: TSCA 799.9780 (1997) and
Table 12-7Models of Host Resistance
PRIMARY FACTORS INVOLVED
IN CHALLENGE MODEL PATHOGEN HOST RESISTANCE
Bacterial Listeria monocytogenes Macrophage, T cell, NK cellStreptococcus pneumoniae Complement, PMN, macrophage,
B cellParasite Plasmodium yoelii T cellViral Influenza A2 Cytotoxic T cell, antibody, complementTumor B16F10 melanoma NK cell, macrophage
SOURCE: From Bradley and Morahan (1982), with permission. See also for an extensive review of host resistance models.
2996R_ch12_419-470 4/27/01 10:01 AM Page 435
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

436 UNIT 4 TARGET ORGAN TOXICITY
OPPTS 870.7800 (1998). These guidelines flow from the config-urations depicted in Table 12-8, although conduct of 3 tests is notrequired by law. Assessment of immunotoxicity begins by expo-sure for a minimum of 28 days to the chemical followed by as-sessment of humoral immunity (PFC assay or anti-sRBC ELISA).If the chemical produces significant suppression of the humoral re-sponse, surface marker assessment by flow cytometry may be per-formed. If the chemical produces no suppression of the humoralresponse, an assessment of innate immunity (NK assay) may beperformed. The tests do not represent a comprehensive assessmentof immune function but are intended to complement assessmentmade in routine toxicity testing (hematological assessments, lym-phoid organ weights, and histopathology). A phagocytosis assayon pulmonary macrophages obtained by broncheoalveolar lavagewas included in TSCA 799.9135 (acute inhalation toxicity withhistopathology) to address potential immunological changes in thelung associated with hazardous air pollutants. Although additionaltests are not required, these guidelines should be viewed as justthat–“guidelines,” with both sound science and responsible man-agement of chemicals being the larger guide to determiningwhether additional tests are needed. These documents are availableon the EPA OPPTS Internet web site. In Europe, the OECD hasnot as yet adopted specific guidelines for immunotoxicity assess-ment. Instead, functional assessments are included in standard tox-icology studies when desired or when suggested by expandedhistopathological results on other standard toxicology studies.
Immunotoxicity Testing of Medical Devices Concern over theinfluence of medical devices on the immune system has grown overthe past 10 years (reviewed in Rodgers et al., 1997). Many of thesedevices may have intimate and prolonged contact with the body.Possible immunologic consequences of this contact could be en-visioned to include immunosuppression, immune stimulation, in-flammation, and sensitization. In 1999, the Office for Device Eval-uation in the Center for Devices and Radiological Health (CDRH)published a guidance document (Immunotoxicity Testing Guid-ance) to provide FDA reviewers and device manufacturers a sys-
tematic approach for evaluating potential adverse immunologicaleffects of medical devices and constituent materials. Because ofthe complexity of the guidance, the reader is referred to the docu-ment itself, which may be located on the FDA CDRH Internet website.
Animal Models in Immunotoxicology
The mouse has been the animal of choice for studying the actionsof xenobiotics on the immune system for several reasons: (1)because there is a vast database available on the immune systemof the mouse, (2) mice are less expensive to maintain than largeranimals, and (3) a wider variety of reagents (cytokines, antibodies,etc.) are available for the mouse (Vos et al., 1994). Because of theneed in the industrial setting to integrate immunotoxicologic as-sessments with routine toxicology testing, a worldwide effort hasbeen under way to validate the rat as a model for immunotoxicol-ogy testing. With the exception of a few functional studies, the ratprovides a near equal model to the mouse for assessing immuno-competence. Many reagents that are available for studying the hu-man immune system can also be used in rhesus and cynomolgusmonkeys.
Other experimental animals including the chicken and fish arebeing used to evaluate the immunotoxicity of xenobiotics (reviewedin IPCS, 1996). Development of these animal models and the re-finement in methods used to assess immunomodulation in thesespecies is anticipated to grow as pressure continues to build re-garding alternative animal models and environmental conscious-ness, and as the consumer market for poultry and farm-raised fishcontinues to expand. An understanding of the chicken immune sys-tem is well established. Because of the ease in handling the de-veloping embryo, chickens may serve to aid the advancement ofthe field of developmental immunotoxicology.
There has been significant interest in the teleost immune sys-tem over the past 15 years, and equivalents of T, B, NK, and ac-cessory cells have been identified. Fish B cells possess many bio-chemical and molecular qualities of their mammalian counterparts,and recently T-cell receptor alpha and beta genes have been char-acterized (reviewed in Miller et al., 1998). The immune systemsof several species of fish have been reported to be sensitive to en-vironmental stressors, including chemicals, which produce im-munomodulation in mammalian species (reviewed in Zelikoff,1994; Bly et al., 1997). As a result, teleosts may serve as early in-dicators of environmental exposure to immunotoxicants (Zelikoff,1998; Zelikoff et al., 2000). Whether or not fish will eventually
Table 12-8Tier Approach for Immunotoxicology Testing
TESTING LEVEL PROCEDURES
Tier I HematologyBody weightOrgan weights (spleen, thymus,
kidney, liver)Spleen cellularityBone marrow cellularity and CFUImmunopathologyPFC assayProliferative responsesNK assay
Tier II Surface marker analysisSecondary (IgG) PFC assayCTL assayDHR assayHost resistance studies
Table 12-9Suggested Testing Configurations: Three Tests with 100 percent Concordance
PFC DHR Surface markersPFC NK DHRPFC NK Thymus:body weightPFC DHR Thymus:body weightSurface markers NK DHRSurface markers DHR T-cell mitogensSurface markers DHR Thymus:body weightSurface markers DHR LPS response
SOURCE: Luster et al. (1988), modified, with permission.
2996R_ch12_419-470 4/27/01 10:02 AM Page 436
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

CHAPTER 12 TOXIC RESPONSES OF THE IMMUNE SYSTEM 437
have efficacy as models in immunotoxicology risk assessment forthe human population remains uncertain, and more effort will berequired before the teleost models are at the current level of therodent (reviewed in Gogal et al., 1999).
Relationship between ImmunotoxicityData in Animals and Humans
There is a clear association between suppression of immune func-tion and an increased incidence of infectious and neoplastic dis-ease in humans (reviewed in Biagini, 1998). Chemicals that pro-duce immunotoxicity in animals have the potential to produceimmune effects in the human population, and these effects may oc-cur in the absence of observable disease (reviewed in Luster et al.,1994). While it would appear relatively straightforward, “the as-sessment [epidemiologically] of immunotoxicity in humans ex-posed to potentially immunotoxic chemicals is much more com-plicated than in experimental animals” (reviewed in IPCS, 1996).Because of this, there is a need and desire to extrapolate from an-imal data to human health effects. A paralleogram approach hasbeen used to assess relationships between animal data and humandata (Fig. 12-15; Selgrade et al., 1999; Van Loveren et al. 1995).In this approach, filled circles represent data which can be readilyobtained in vivo or in vitro. Unfilled circles represent data whichcannot be obtained experimentally. Data that may occasionally beobtained but must sometimes be extrapolated are represented bycircles filled with blue lines. This approach has been used to ex-trapolate animal to human data in an initial quantitative assessmentof risk for deleterious effects of UV radiation (Van Loveren et al.1995). A dual parallelogram approach has also been used to pro-pose that data for ozone suggest that effects of in vivo human ex-posure to phosgene on alveolar macrophage phagocytosis may bepredicted based on effects of in vitro exposure and in vivo animaldata (reviewed in Selgrade et al., 1995).
Evaluation of Mechanisms of Action
When we think of the ways chemicals can act on various organsystems we most often think of the direct effects of chemical ex-posure. Direct effects on the immune system may include chemi-cal effects on immune function, structural alterations in lymphoidorgans or on immune cell surfaces, or compositional changes inlymphoid organs or in serum (Table 12-10). Xenobiotics may ex-ert an indirect action on the immune system as well. They may bemetabolically activated to their toxic metabolites, may also haveeffects on other organ systems (e.g., liver damage) which then im-pacts the immune system, or may induce alterations in hormonalhomeostasis.
As mentioned previously, a unique quality of the immune sys-tem is the ability to remove immune cells and have them functionin vitro. This is particularly important in investigating the mecha-nisms of action of xenobiotics. For example, it can be determinedwhether a compound acts either directly or indirectly on immunecells by comparing in vivo to in vitro chemical exposure (refer toFig. 12-11). Immunotoxic compounds that act indirectly will haveno effect on an in vitro—generated immune response. Compoundsthat require metabolism to reactive metabolites will also have noeffect on in vitro—generated immune responses following in vitroexposure. However, this metabolic requirement can be mimickedin vitro by incubating the chemical with a microsomal S9 prepa-
ration prior to in vitro exposure of splenocytes. The metabolicallyactivated compound may then be capable of suppressing in vitro—generated immune responses.
Numerous methodologies are available to evaluate cellular andmolecular mechanisms of action. Xenobiotic-induced effects onspecific cell types of the antibody response can be determined us-ing antigens that require several cell types, such as macrophage, T,and B cells (sRBC), macrophages, and B cells only (DNP-Ficoll),or B cells alone (LPS) for the production of antigen-specific anti-body. In addition, splenocytes can be separated into the various cellpopulations such as adherent cells (primarily macrophages) andnonadherent cells (T and B cells), that can be individually exposedand then reconstituted in cell culture to yield an in vitro—gener-ated immune response equal to that of unseparated cells. This sep-aration/reconstitution analysis is an excellent way to determine spe-cific cell targets of xenobiotic action. In addition, supernatants fromin vitro—generated antibody responses can be transferred amongthemselves in an effort to assess xenobiotic action on soluble fac-tors such as the cytokines, and ELISA methodology and quantita-tive PCR can be used to quantitate in vitro cytokine production andcytokine gene transcription, respectively, in response to variousstimuli. Much of the progress in immunotoxicology research hasbeen the result of the application of advances made in immunol-ogy and molecular biology and it is expected that this will con-tinue as these areas of basic science progress by leaps and bounds.
Figure 12-15. Parallelogram approach in immunotoxicology for relatinganimal data to human data. [Modified from Selgrade (1999), with per-mission.]
2996R_ch12_419-470 4/27/01 10:02 AM Page 437
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

438 UNIT 4 TARGET ORGAN TOXICITY
IMMUNOMODULATION BY XENOBIOTICS
Immunosuppression
Halogenated Aromatic Hydrocarbons Few classes of xenobi-otics have been as extensively studied for immunotoxicity as thehalogenated aromatic hydrocarbons (HAHs; reviewed by Kerkvlietand Burleson, 1994, and Holsapple, 1996). The prototypical andmost biologically potent member of this family of chemicals, whichincludes the polychlorinated biphenyls (PCBs), the polybrominatedbiphenyls (PBBs), the polychlorinated dibenzofurans (PCDFs), andthe polychlorinated dibenzodioxins (PCDDs), is 2,3,7,8-tetra-chlorodibenzo-p-dioxin (TCDD; dioxin). Substantial evidence hasaccumulated that demonstrates the immune system to be a targetfor toxicity for these chemicals. Derived from a variety of animalmodels, primarily rodents, this evidence includes thymic atrophy,pancytopenia, cachexia, immunosuppression, and tumor promo-tion. There is also epidemiological evidence suggesting immuno-toxicity by the HAHs can also occur in humans; however, signif-icant immunosuppression has not been associated conclusivelywith specific alterations of human immune function.
Quite possibly the most important advance in recent years inthe study of the HAHs has been the determination of a genetic ba-
sis for sensitivity to the toxic effects of this family of chemicals.Many of the biochemical and toxic effects of the HAHs appear tobe mediated via HAH binding to an intracellular heterodimericcomplex between the aryl hydrocarbon receptor (Ah-R) and thearomatic receptor nuclear transporter (ARNT; Hoffman et al.,1991). The Ah-R–ARNT complex translocates to the nucleus, bindsto dioxin-responsive elements (DREs), and directs transcriptionalactivation (e.g., CYP1A1, PAI-2, fos/jun) and mRNA stabilization(e.g., TGF�, IL-1�) (Sutter et al., 1991; Sutter and Greenlee, 1992).In mice, allelic variation at the Ah locus has been described. Thesealleles code for Ah-Rs with differential binding affinities forTCDD. For example, the C57BI/6 mouse represents a strain ofmice (Ahbb) which is exquisitely sensitive to TCDD (TCDD re-sponsive), while the DBA/2 strain of mice (Ahdd) is much less sen-sitive to the toxic effects of TCDD (TCDD nonresponsive or TCDDlow-responsive). These allelic differences may ultimately explainthe controversial differences in observed toxic responses betweenanimal species and even between individual tissues within the samespecies.Polychlorinated Biphenyls PCBs have seen extensive commer-cial use for over half a century. Their unique physical and chemi-cal properties make PCB mixtures ideal for use as plasticizers, ad-hesives, and as dielectric fluids in capacitors and transformers.Mixtures of PCBs (e.g., Aroclors) have been commonly used to
Table 12-10Possible Mechanisms of Chemically Induced Immune Modulation
TYPE OF
EFFECT MECHANISM EXAMPLES
Direct Functional Altered antibody-mediated responseschanges Altered cell-mediated responses
Altered release of preformed mediatorsAltered host resistanceInability of one or more cell types to perform a required activity, e.g.,
Production of antibodyRelease of cytokinesProcessing and presentation of antigenProliferation and differentiationReceptor-mediated signal transduction
Structural Alterations in surface receptors or ligandschanges Alteration in expression of receptors or ligands
Histopathologic changes in lymphoid organs
Compositional Alterations in CD3�, CD4�, CD8�, B220�, and/or Ig� in spleenchanges Alterations in CD4�, CD8�, CD4�/CD8�, and/or CD4�/CD8� in thymus
Changes in hematologic cellular parametersAlterations in circulating IgAlterations in CFU profile in bone marrow
Indirect Metabolic Conversion to a toxic metaboliteActivation
Effects Induction of acute-phase proteins as a result of liver injurysecondary toother targetorgan toxicity
Hormonal Increased corticosteroid release from the adrenal glandchanges Alteration in neuroendocrine regulation
Alteration in autonomic output from the CNSAltered release of steroids from sex organs
2996R_ch12_419-470 4/27/01 10:02 AM Page 438
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

CHAPTER 12 TOXIC RESPONSES OF THE IMMUNE SYSTEM 439
evaluate the immunotoxicity of PCBs and have been reported tosuppress immune responses and decrease host resistance (reviewedby Kerkvliet, 1984). The first indication that PCBs produced im-munotoxic effects was the observation of severe atrophy of the pri-mary and secondary lymphoid organs in general toxicity tests andthe subsequent demonstration of the reduction in numbers of cir-culating lymphocytes. Studies to characterize the immunotoxic ac-tion of the PCBs have primarily focused on the antibody response.This parameter is by far the one most consistently affected by PCBexposure and effects on antibody response have been demonstratedin guinea pigs, rabbits, mice, and rhesus monkeys. PCB-exposedmonkeys exhibit chloracne, alopecia, and facial edema, all classi-cal symptoms of HAH toxicity. In an extensive characterization ofthe effects of PCBs on nonhuman primates, Tryphonas and col-leagues (1991a,b) exposed rhesus monkeys to Aroclor 1254 for 23to 55 months. The only immune parameter consistently suppressedwas the PFC response to sRBC (both IgM and IgG). In addition,after 55 months of exposure, lymphoproliferative responses weredose-dependently suppressed and serum complement levels weresignificantly elevated. The observed elevation in serum comple-ment has also been reported in PCDD-exposed children fromSeveso, Italy (Tognoni and Bonaccorsi, 1982) and in B6C3F1-exposed mice (White et al., 1986).
The effects of PCBs on cell-mediated immunity (CMI) are farless clear and both suppression and enhancement have been reported. Exposure to Aroclor 1260 has been demonstrated to suppress delayed-type hypersensitivity (DTH) responses in guineapigs, whereas exposure to Aroclor 1254 was reported to enhancelymphoproliferative responses in rats. In a similar study in Fischer344 rats (Aroclor 1254), thymic weight was decreased, NK cell activity was suppressed, PHA-induced proliferative responses wereenhanced, and there was no effect on the MLR proliferative response or CTL activity. Other investigators (Silkworth and Loose,1978, 1979) have reported enhancement of graft –versus host reactivity and the MLR proliferative response. The augmentationof selected CMI assays may reflect a PCB-induced change in T cell subsets (as described above) and thus in immunoregula-tion.
Studies on host resistance following exposure to PCBs indi-cate that the host defenses against hepatitis virus (ducks) and toherpes simplex virus, Plasmodium berghei, L. monocytogenes, andSalmonella typhimurium (mice) are suppressed (reviewed by Deanet al., 1985). PCB-induced changes in tumor defenses have notbeen well defined and both augmentation and suppression havebeen reported. This probably reflects the variability in observed re-sponses in CMI.Polybrominated Biphenyls The polybrominated biphenyls(PBBs) have been used primarily as flame retardants (FiremasterBP-6 and FF-1). While it is assumed that their profile of activityis similar to that of the PCBs, few studies have actually evaluatedthe action of the PBBs on immunocompetence. In Michigan (in1973) Firemaster BP-6 was inadvertently substituted for a nutrientadditive in cattle feed, resulting in widespread exposure of animalsand humans to PBBs. Studies conducted on livestock following theincident indicated little if any PBB-induced alterations in im-munocompetence (Kately and Bazzell, 1978; Vos and Luster,1989). Like CMI observations involving PCBs, CMI responses inPBB-exposed individuals are not conclusive, showing both a re-duction in circulating numbers of T and B cells and a suppressionof selected CMI parameters or no effect on CMI at all.Polychlorinated Dibenzodioxins By far the majority of the in-
vestigations into the immunotoxic potential and mechanisms of ac-tion of the HAHs have focused on TCDD, primarily because thischemical is the most potent of the HAHs, binding the Ah-R withthe highest affinity. The effects of TCDD on immune function havebeen demonstrated to be among the earliest and most sensitive in-dicators of TCDD-induced toxicity (reviewed by Holsapple et al.,1991a,b). TCDD is not produced commercially, except in smallamounts for research purposes. Rather, it is an environmental con-taminant formed primarily as a by-product of the manufacturingprocess that uses chlorinated phenols or during the combustion ofchlorinated materials. It is usually associated with the productionof herbicides such as 2,4,5-trichlorophenoxyacetic acid (2,4,5-T),and Agent Orange [a 1:1 combination of 2,4-dichlorophenoxyaceticacid (2,4-D) and 2,4,5-T]. Other sources include pulp and papermanufacturing (chlorine bleaching), automobile exhaust (leadedgasoline), combustion of municipal and industrial waste, and theproduction of PCBs.
Like other HAHs, exposure to TCDD results in severe lym-phoid atrophy. Because thymus-derived cells play an integral rolein tumor surveillance and host resistance, the earliest studies onTCDD-induced immunotoxicity focused on changes in cell-mediated immunity. Studies on CMI have shown that this branchof acquired immunity is sensitive to the toxic effects of TCDD.CTL development and activity has been shown by numerous in-vestigators to be significantly decreased after exposure to TCDD,an effect which appeared to be age dependent (e.g., the youngerthe mice when exposed, the greater the sensitivity to TCDD). Inaddition to suppression of CTL function, TCDD exposure also re-sults in decreases in PHA- and Con A–induced proliferative re-sponses, DHR, and graft-versus-host responses. Enhanced prolif-erative responses in juvenile mice have also been observed(Lundberg et al., 1990).
Consistent with the observation that mice exposed perinatallyor postnatally (developmentally younger animals) are more sensi-tive to the effects of TCDD, it has recently been determined thatthymic involution is a result of TCDD-induced terminal differen-tiation of the thymic epithelium and, thus, T cells do not have aproper nutrient-filled microenvironment in which to develop(Greenlee et al., 1984, 1985). This conclusion is supported by pre-vious observations that TCDD significantly decreased the numberof immature T cells (CD4�/CD8�) in the thymus (Kerkvliet andBrauner, 1990).
Numerous investigations have demonstrated the PFC responseto be exquisitely sensitive to the toxic effects of TCDD. This ef-fect segregates with the Ah locus (Vecchi et al., 1983) and appearsto be dependent upon duration and conditions of exposure (Hols-apple et al., 1991c). Although TCDD induces profound changes inthe PFC assay, no changes have been observed in splenic cellular-ity (numbers of Ig�, CD4�, or Thy-1� cells) either before or af-ter antigen challenge. In addition, in spite of suppression of anti-gen-induced antibody production, TCDD has been reported toenhance total serum immunoglobulin concentrations.
Although many labs have defined Th cells as unaffected byTCDD exposure, Kerkvliet has reported suppression of T-cell reg-ulation of immune responses (Kerkvliet and Brauner, 1987;Kerkvliet et al., 1990). However, studies using separation/recon-stitution or in vitro exposure techniques have elucidated the B cellas the primary immune cell target for TCDD. In addition, B-celldifferentiation was identified as the stage affected. Consistent withthese findings, TCDD has a selectively greater effect on immatureB cells than on mature B cells. Additionally, the selective effect on
2996R_ch12_419-470 4/27/01 10:02 AM Page 439
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

440 UNIT 4 TARGET ORGAN TOXICITY
B cells cannot be accounted for by induction of Ts (suppressor Tcells).
The information presented so far implicates an effect of TCDDon lymphocyte development/maturation either in the bone marrow,thymus, or after antigenic challenge. In support of this conclusion,TCDD exposure has been shown to suppress bone marrow cellu-larity and stem cell proliferation in rodent neonates exposed inutero. TCDD also appears to alter homing patterns of lymphocytesafter perinatal or postnatal exposure in Fischer 344 rats and can al-ter development of granulocyte-macrophage colony-forming units(CFU-GM) at concentrations below those that induce thymic atro-phy. Using fetal thymic or bone marrow equivalent cells from birds,it has been demonstrated that TCDD severely attenuates prolifer-ation and development of CD4�/CD8� cells and of B cells. Thesedata support the conclusions of Greenlee and colleagues(1984,1985) that TCDD alters the microenvironment in which lym-phocytes develop.
The effects of TCDD on innate immunity are less well stud-ied. TCDD has been shown to inhibit some functions of polymor-phonuclear leukocytes (PMNs), including cytolytic and cytostaticactivities. This inhibition has been postulated to be related to PMNdevelopment in the bone marrow. Results by several investigatorshave shown TCDD-induced alterations in serum C3, indicating sol-uble mediators of innate immunity may also be targeted (Whiteand Anderson, 1985; White et al., 1986). There have been no ob-served effects on macrophage-mediated cytotoxicity, NK function,or interferon production. In host resistance models, TCDD expo-sure has been shown to increase susceptibility to several bacterial,viral, and tumor models.
There is little doubt that TCDD and related PCDDs are im-munotoxic, particularly in mice. However, extrapolation to humanexposure has proven to be difficult. There are a few instances inwhich accidental human exposure to TCDD and related congenershas afforded the opportunity to study exposure-related human im-munologic responses. In children exposed to PCDDs in Seveso,Italy (1976), nearly half of the exposed study group exhibited chlo-racne (a hallmark of human exposure to PCDDs) 3 years after theaccident. Immune parameters measured at that time were unaf-fected. In a second study conducted 6 years later on different sub-jects, there was an increase in complement, which correlated withthe incidence of chloracne, an increase in circulating T and B cells,and an increase in peripheral blood lymphocyte (PBL) mitogenicresponses. A second incident occurred in 1971 in Times Beach,Missouri, when wastes containing TCDD were sprayed on roadsto prevent dust formation. Both low-risk and high-risk individualsfrom this area were examined for DHR responses. Slight, but sta-tistically nonsignificant alterations were observed in high-risk com-pared to low-risk individuals. In addition, there was a low-level in-crease in mitogenic responsiveness in high-risk persons. In a secondstudy conducted 12 years later, no alterations were observed inDHR or mitogenic responses between exposed or control individ-uals. More recently, studies have been undertaken to evaluate thein vitro effects of TCDD on human cells. TCDD suppressed IgMsecretion by human B cells in response to the superantigen toxicshock syndrome toxin-1 (TSST-1) and the proliferation and IgGsecretion of human tonsillar B cells in response to LPS and cy-tokines.Polychlorinated Dibenzofurans Like the PCDDs, polychlori-nated dibenzofurans (PCDFs) are not produced commercially butare true environmental contaminants associated with the produc-tion of chlorophenoxy acids, pentachlorophenol, and other PCB
mixtures. Although higher concentrations are required to achieveobservable effects, the immunotoxic profile of the PCDFs is sim-ilar in nature to that described for TCDD. In fact, most of what isknown regarding the immunotoxicity of the PCDFs in animal mod-els has been learned during structure–activity relationship studiescomparing TCDD to congeners of the dibenzofurans. TCDF (tetra-chlorodibenzofuran) exposure in most species is associated withthymic atrophy and in guinea pigs it has been shown to suppressthe DHR and lymphoproliferative responses to PHA and LPS. Sup-pression of the PFC response to SRBC after exposure to severalPCDF congeners has also been reported.
Two important case studies of human immunotoxicology in-volved populations accidentally exposed to HAHs. There is evi-dence that the PCDFs were the primary contributors to the ob-served toxic effects. Greater than 1850 individuals in Japan (in1968) and in excess of 2000 people in Taiwan (in 1979) were af-fected when commercial rice oil was found to be contaminatedwith HAHs. PCDFs were observed in the tissues of the exposedpopulations and subsequent studies on immune status revealed adecrease in total circulating T cells, decreased DHR, and enhancedlymphoproliferative responses to PHA and pokeweed mitogen(PWM). In addition, many of the exposed individuals suffered fromrecurring respiratory infections, suggesting that host resistancemechanisms had been compromised.
Polycyclic Aromatic Hydrocarbons The polycyclic aromatichydrocarbons (PAHs) are a ubiquitous class of environmental con-taminants. They enter the environment through many routes in-cluding the burning of fossil fuels and forest fires. In addition tobeing carcinogenic and mutagenic, the PAHs have been found tobe potent immunosuppressants. Effects have been documented onhumoral immunity, cell-mediated immunity, and on host resistance(reviewed by Ward et al., 1985, and White et al., 1986). The mostextensively studied PAHs are 7,12-dimethylbenz[a]anthracene(DMBA) and benzo[a]pyrene (BaP).
The PAHs suppress the antibody response to a variety of Tcell–dependent and T cell–independent antigens. In addition, miceexposed to BaP exhibit suppressed lymphoproliferative responsesto mitogens but not alloantigens (Dean et al., 1983). In Dean’s stud-ies, host resistance to the PYB6 tumor and to L. monocytogeneswere unaffected by BaP exposure, as was the DHR response andallograft rejection, suggesting that the T cell (and CMI) was onlyminimally affected by BaP. Since the PFC response to T cell–dependent and T cell–independent antigens is markedly suppressedby BaP exposure, it would appear that BaP may target themacrophage or the B cell.
In contrast to BaP, DMBA (the more potent PAH) significantlysuppresses not only PFC responses but also NK activity, CTL re-sponses, DHR responses, and alloantigen-induced lymphoprolifer-ative responses as well. Therefore DMBA exposure seems to re-sult in long-lasting immunosuppression of humoral immunity (HI),CMI, and tumor resistance mechanisms in mice. Suppression ofthe immunologic mechanism of tumor resistance by PAHs tendsto correlate with their carcinogenic properties and may contributeto their carcinogenicity.
Although it is well established that the PAHs are immuno-suppressive in nature, the mechanism or mechanisms by which theyelicit this action have remained elusive until recently (reviewed byWhite et al., 1994). It is generally recognized that PAHs exert car-cinogenic and mutagenic effects after being metabolized by P450enzymes to more toxic metabolites. It has recently been shown that
2996R_ch12_419-470 4/27/01 10:02 AM Page 440
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

CHAPTER 12 TOXIC RESPONSES OF THE IMMUNE SYSTEM 441
splenocytes from unexposed mice can metabolize exogenouslyadded DMBA via P450 enzymes (Ladics et al., 1991). In addition,Ladics and colleagues (1992a,b) demonstrated that macrophageswere the primary cell capable of metabolizing PAHs, and that thesecells were capable of generating 7,8-dihydroxy-9,10-epoxy-7,8,9,10-benzo[a]pyrene (BPDE), the reactive metabolite proposedto be the ultimate carcinogenic form of BaP. These data are con-sistent with other studies demonstrating the presence and in-ducibility of aryl hydrocarbon hydrolase (AHH) in cells of themacrophage lineage and with other data suggesting that themacrophage is the primary target and is functionally compromised(with respect to accessory cell help) following exposure to thePAHs.
Nitrosamines The nitrosamine family comprises the ni-trosamines, nitrosamides, and C-nitroso compounds. Exposure tonitrosamines, especially N-nitrosodimethylamine (DMN, the mostprevalent nitrosamine) comes primarily through industrial and di-etary means, and minimally through environmental exposure. Thetoxicity and immunotoxicity of DMN have been extensively re-viewed (Myers and Shook, 1996). Single or repeated exposure toDMN inhibits T-dependent humoral immune responses (IgM andIgG), but not T-independent responses. Other symmetrical ni-trosamines, such as diethylnitrosamine (DEN), dipropylnitro-samine (DPN), and dibutylnitrosamine (DBN), demonstrated sim-ilar effects on humoral immunity but were not as potent as DMN.In fact, as the length of the aliphatic chain increased, the dose re-quired to suppress the anti-sRBC PFC response by 50 percent(ED50) also increased. In contrast, nonsymmetrical nitrosaminessuppressed humoral immunity at comparable concentrations. Over-all, the rank order of ED50 values paralleled their LD50 values. Tcell–mediated lyphoproliferative responses (mitogens or mixedlymphocyte response) and delayed hypersensitivity response arealso suppressed following DMN exposure. In vivo exposure toDMN followed by challenge with several pathogens did not pro-duce a pattern of effects that was consistent (decreased resistanceto Streptococcus zooepidemicus and influenza, no effects on re-sistance to herpes simplex types 1 or 2 or Trichinella spiralis, in-creased resistance to L. monocytogenes). In contrast, anti-tumor ac-tivity in DMN-exposed animals was consistently enhanced.DMN-exposed animals also have altered development ofhematopoietic cells (increased macrophage precursors). Togetherthese data suggest the macrophage (or its developmental precur-sors) as a primary target. Mechanistic studies have demonstratedthat DMN alterations in cell-mediated immunity are associatedwith enhanced macrophage activity, increased myelopoietic activ-ity, and alterations in TNF-� transcriptional activity. It has beenpostulated that DMN may cause the enhanced production of GM-CSF, which can have autocrine (enhanced tumoricidal and bacte-ricidal activity) and paracrine (induced secretion of T cell–sup-pressing cytokines by macrophages) activities. Further work isneeded to clarify the apparent selectivity of DMN for alteration inmacrophage function.
Pesticides Pesticides include all xenobiotics whose specific pur-pose is to kill another form of life, usually insects or small rodents.These compounds can be divided into four classes: the organophos-phates, organotins, carbamates, and organochlorines. While thereis increasing evidence that certain pesticides can produce alter-ations in immune function in animal models (reviewed by Pen-ninks et al., 1990; Barnett and Rodgers, 1994; and Voccia et al.,
1999), studies following human exposure are limited and reveal noconclusive results (Thomas et al., 1990).Organophosphates Occupational exposure to organophosphateshas been linked to decreased PMN chemotaxis and increased up-per respiratory infection. Overall, relatively little is known aboutthe immunotoxic effects of organophosphates on the immune sys-tem. The most extensively studied of these are malathion,parathion, and methyl parathion. Prolonged exposure to low dosesof malathion (a cumulative high dose) results in decreased humoralimmunity. High doses, which have direct cholinergic effects, alsosuppress humoral immunity, but whether this is a direct effect ofthe chemical on the immune system or a stress response elicitedby cholinergic effects is unclear. In contrast, acute oral exposurehas been shown to enhance humoral immunity and mitogenic pro-liferative responses with no other immune-related effects. In vitroexposure of either human mononuclear cells or murine splenocytesto malathion results in decreased lymphoproliferative responses,suppressed CTL generation, and a decrease in the stimulus-inducedrespiratory burst in peritoneal cells. Moreover, after metabolism ofmalathion by liver S9 preparations, the metabolites of malathionwere not immunosuppressive. In addition, separation and recon-stitution studies revealed that the adherent population (i.e., themacrophage) is the primary cellular target for malathion. Malathionexposure also induces peritoneal mast cell degranulation and en-hances macrophage phagocytosis. In light of the above findingsand the fact that mast cell degranulation products can modulateleukocyte activity, it has been suggested that peritoneal mast celldegranulation following acute malathion exposure may subse-quently lead to augmentation of leukocyte functions, thereby non-specifically enhancing the generation of an immune response.
Parathion has attracted more attention than malathion, prob-ably because it is more acutely toxic. This pesticide suppressesboth humoral and cell-mediated immunity. Following exposure tomethyl parathion, decreased germinal centers after antigen chal-lenge, thymic atrophy, and suppressed DHR responses have beenreported. Other experiments have shown suppression of lympho-proliferative responses as well as increased susceptibility topathogens. In vitro exposure to parathion or paraoxon suppressesCMI in murine splenocytes, IL-2 production in rat splenocytes, andproliferative responses in human lymphocytes. Finally, exposure ofhuman bone marrow cells to organophosphates may result in inhi-bition of CFU formation.Organochlorines The organochlorines include chemicals such aschlordane, dichlorodiphenyltrichloroethane (DDT), Mirex, pen-tachlorophenol, aldrin, dieldrin, and hexachlorobenzene. These areamong the longer-lived pesticides and they have an increasedpropensity for contamination of soil and ground water. The hu-moral immune response to both T cell–dependent and T cell–in-dependent antigens is suppressed following exposure to dieldrin,and macrophage functions from dieldrin-exposed animals are de-pressed. The apparent effect of dieldrin on macrophages correlateswith the increased susceptibility of dieldrin-exposed animals tomurine hepatitis virus, which targets macrophages (Krzystyniak etal., 1985).
Definitive immunosuppression produced by chlordane wasfirst reported in 1982 by Spyker-Cranmer and colleagues. In uteroexposure resulted in decreased DHR responses in mice with nodeficit in antibody production to sRBC. This correlated with an in-crease in resistance to influenza infection because the DHR con-tributes to the pathology of the infection (Menna et al., 1985). Asreported for dieldrin, the primary cellular target for chlordane ap-
2996R_ch12_419-470 4/27/01 10:02 AM Page 441
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

442 UNIT 4 TARGET ORGAN TOXICITY
pears to be the macrophage. Although peritoneal exudate cells frommice exposed to chlordane in utero showed normal cytotoxic re-sponses, the response is delayed by 24 to 48 h. Preliminary evi-dence suggests that prenatal exposure inhibits myeloid progenitordevelopment in bone marrow, but no cause–effect relationship be-tween this and macrophage deficits has been determined. In con-trast to observations from mice exposed in utero, exposure of adultmice to chlordane does not result in any changes to several im-mune parameters, including PFC response to sRBC, MLR, DHR,or mitogenic lymphoproliferation.
DDT is one of the oldest pesticides in use today and one ofthe first studied for its immunotoxic potential. DDT inhibited an-tiovalbumin serum antibody titers in rats exposed via the drinkingwater (Wasserman et al., 1969). In contrast, both rats and guineapigs fed DDT exhibited no alterations in antitoxin antibody (Gab-liks et al., 1973, 1975). These animals did, however, have a sup-pressed anaphylactic reaction as a result of decreased numbers ofmast cells. Studies by Street (1981) indicated that chickens exposedto DDT or Mirex had suppressed levels of circulating IgM and IgG,although specific antibody titers were normal. In addition, DDTexposure resulted in decreased antigen-induced germinal centers,thymic atrophy, and suppressed CMI. While most studies on DDThave focused on humoral immunity, the effects of DDT on CMI,host resistance, and particularly macrophage function remain rel-atively unexplored.Organotins Trisubstituted organotins such as TBTO are widelyused as biocides and have recently been recognized as producingsome immunotoxic effects. The action of these compounds on lym-phoid tissue and immunity has been extensively reviewed (Pen-ninks et al., 1990). The most outstanding action of TBTO is theinduction of profound but reversible thymic atrophy. In addition,the developing immune system appears to be more sensitive to theeffects of TBTO than does the immune system of the adult animal.Studies by Vos et al. (1984) demonstrated a decrease in cellularityin the spleen, bone marrow, and thymus. The decrease in spleniccellularity was associated with a concomitant loss of T lympho-cytes. More specifically, oral TBTO exposure resulted in decreasedserum IgG, increased serum IgM, and suppression of DHR re-sponses to tuberculin and ovalbumin. In those studies, host resist-ance to L. monocytogenes was diminished. Cytotoxicity by adher-ent peritoneal cells was suppressed but there was no observed effecton NK cytotoxicity. In contrast, van Loveren et al. (1990) observedsuppressed lung NK cytotoxicity in rats exposed orally to TBTO.In addition, the lymphoproliferative response of thymocytes toPHA, Con A, and PWM was significantly suppressed.Carbamates Carbamate insecticides such as carbaryl (Sevin) andaldicarb have frequently been studied as immunotoxicants. Stud-ies involving oral exposure of chickens to Sevin resulted in acuteand sometimes prolonged suppression of germinal centers and an-tibody production. In addition, carbaryl exposure causes suppres-sion of granulocyte phagocytosis, which may last for up to 9months. However, other studies have found no indication of im-munotoxicity except at near lethal concentrations. In an evaluationof humoral immunity following a 2-week exposure to carbaryl inrats, suppression of the IgM PFC response to sRBC was observedfollowing inhalation exposure but not oral or dermal exposure(Ladics et al., 1994). Conflicting results have also been observedin animals exposed to aldicarb or methyl isocyanate, an interme-diate in carbamate pesticide production. Deo and colleagues (1987)reported alterations in T cells and lymphoproliferative responsesin humans accidentally exposed to methyl isocyanate. In contrast,
mice exposed to the same compound showed no significant alter-ations in immune status (Luster et al., 1986). More recently, Pruettand coworkers (1992a) evaluated the immunotoxicity of sodiummethyldithiocarbamate, a chemical widely used for the control ofweeds, fungi, and nematodes in soil. These investigators observeddecreased thymus weight, depletion of the CD4�/CD8� popula-tion of thymocytes, and profound suppression of NK activity fol-lowing both oral and dermal exposure. Given the number of con-flicting reports, currently there is insufficient evidence in eitherhumans or animal models to indicate that carbamate pesticides posea significant risk to the human population. However, the data byLadics and colleagues (1994) suggest that immunotoxicologic stud-ies on pesticides should consider relevant exposure routes, as car-baryl was not immunosuppressive by routes through which contactby the general population typically occurs (oral and dermal viaresidues in water and on food).Pyrethroids Supermethrin has been demonstrated to decrease theplaque-forming cell response after a single oral administration.However, following repeated exposure (up to 3 mg/kg/day), onlyslight changes in circulating leukocytes and in nucleated cell num-bers in the bone marrow were observed (Siroki et al., 1994). Rab-bits administered cypermethrin orally for 7 weeks demonstrateddecreased tuberculin skin reactions and a decrease in the antibodytiter to S. typhimurium (Desi et al., 1985).Other Pesticides Naphthalene is a bicyclic aromatic hydrocarbonthat is used, among other things, as an insect repellent, insecticide,and vermicide. To date, no evidence for immunotoxicity has beendemonstrated despite prolonged exposure (Shopp et al., 1984). Re-cently it has been suggested that this lack of effect may be relatedto the inability of splenocytes to metabolize naphthalene and/or torelatively low concentrations of metabolites that may be generatedin the liver and diffuse to the spleen (Kawabata and White, 1990).Paraquat has been shown to decrease the percentage of circulatingneutrophils as a result of their migration to the lungs and propanilcan alter immune cell development (myeloid and erythroid pro-genitors). Thymic atrophy in response to propanil exposure has alsobeen observed, although the contribution of induced glucocorti-coids to this effect has not been ascertained (Voccia et al., 1999).
Metals Generally speaking, metals target multiple organ systemsand exert their toxic effects via an interaction of the free metal withthe target: enzyme systems, membranes, or cellular organelles.Although specific immunotoxic consequences of metal exposureare well documented in the literature (see reviews by Lawrence,1985; McCabe, 1994; Burns et al., 1994e; and a text edited byZelikoff and Thomas, 1998), this section focuses on the four best-studied immunotoxic metals: lead, arsenic, mercury, and cadmium.In considering the immunotoxicity of most metals, it is importantto remember that at high concentrations, metals usually exert im-munosuppressive effects; however, at lower concentrations, im-munoenhancement is often observed (Koller, 1980; Vos, 1977).Lead By far the most consistent finding in studies evaluating theeffects of metals on immune responses is increased susceptibilityto pathogens. For lead (Pb), decreased resistance to the bacterialpathogens S. typhimurium, Escherichia coli, and L. monocytogeneshas been observed. Enhanced susceptibility to viral challenge hasalso been reported. Other investigators found no change in virallyinduced IFN production in Pb-exposed animals.
Studies on the specific effects of Pb on functional cell-mediated immunity have yielded no conclusive results, as reportsrange from significant suppression to no effect. Currently, these
2996R_ch12_419-470 4/27/01 10:02 AM Page 442
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

CHAPTER 12 TOXIC RESPONSES OF THE IMMUNE SYSTEM 443
differences cannot be explained by differences in routes of expo-sure or dose. Suppression of humoral immunity, however, has beendemonstrated. In rodents exposed to Pb, lower antibody titers havebeen observed. In addition, children environmentally exposed toPb and infected naturally with Shigella dysenteriae had prolongeddiarrhea, and occupationally exposed persons reported more coldsand influenza and exhibited suppressed secretory IgA levels, sug-gesting Pb-induced suppression of humoral immunity. Pb-inducedeffects on myeloid cells include an increase in the number ofmyeloid progenitors in the bone marrow (CFU-GM) with a sub-sequent decrease in more mature cells. Following in vivo exposureto Pb, splenocytes displayed consistently suppressed IgM PFC re-sponses to sRBC. Separation and reconstitution experiments indi-cated that this suppression is likely due to an effect on macrophagefunction.
In recent mechanistic studies (reviewed by McCabe, 1994),an alteration in the ability of the macrophage to process and pres-ent antigen to antigen-primed T cells confirmed the previous ob-servation and suggested that Pb alters immune recognition. In con-trast to other reports concerning the immunosuppressive action ofPb on PFC responses, enhancement of the in vitro–generated PFCresponse (in vitro exposure in Mishell-Dutton-type culture) hasbeen reported which appeared to be the result of enhancement ofB-cell differentiation. This effect may occur at the level of B-cellactivation or cytokine responsiveness. And finally, in vitro additionstudies indicate that Pb may have differential effects on Th1 andTh2 cells and can inhibit the production of IL-2. If the productionor activity of other cytokines is observed to be suppressed or en-hanced by metal exposure, it may be that metals can exert signif-icant effects on immune regulation, which can result in either im-munoenhancement or immunosuppression.Arsenic The literature concerning arsenic (As)-induced im-munomodulation is fraught with inconsistencies due to differencesin speciation of As (which plays a significant role in arsenic toxi-city), the route of administration, the concentrations used, and thevarious species and strains of animals utilized. As with many othermetals, exposure to low concentrations of As often leads to en-hanced immune responses while exposure to higher concentrationsresults in immunosuppression (reviewed by Burns et al., 1994e).Exposure of mice to sodium arsenite (NaAsO2) in the drinking wa-ter or subcutaneously was shown to decrease resistance to viralpathogens. Other investigators have shown that exposure to ar-senicals offers some degree of protection against tumor incidence,although tumors that did develop grew at a much faster rate. Noalterations in CMI were observed in those investigations. Interest-ingly, host resistance studies, conducted after exposure to the semi-conductor material gallium arsenide (GaAs), revealed that GaAsafforded modest protection against infection with both S. pneumo-niae and L. monocytogenes, although resistance to the B16F10melanoma was reduced. It was subsequently determined that theAs concentrations in the blood of these animals was high enoughto offer a chemotherapeutic effect against the bacterial pathogens(arsenicals were once widely used as chemotherapeutic agents be-fore the development of drugs with higher efficacy and lower tox-icity). These studies are important because they are among the firstto demonstrate the intricate interplay between the host, thepathogen, and the xenobiotic.
In addition to these holistic immune alterations, As exposurehas been shown to inhibit both the PFC response in animal mod-els and peripheral blood lymphocyte proliferation in humans. Also,substantial mechanistic information exists regarding the immuno-
toxicity of intratracheally instilled GaAs. Exposure results in sup-pression of the PFC, CTL, DHR, and MLR responses. Followinginstillation, both arsenic and gallium can be detected in the bloodand tissues for as long as 30 days, suggesting that the lung acts asa depot for prolonged systemic exposure to dissociated gallium andarsenic. Mechanistic studies revealed that all cell types involved inthe generation of an antibody response (macrophage, T, and B cells)are affected by GaAs exposure. Decreased expression of Ia andability to process and present the particulate antigen, sRBC, rep-resent functional deficits of the macrophage, while inhibition ofmitogen- or receptor-driven proliferation, expression of the IL-2receptor, and production of cytokines during the antibody responserepresent functional deficits of the T cell. A criticism of the stud-ies using GaAs has been that instillation of particulate matter causesa stress response resulting in increased levels of circulating corti-costeroids, known to have potent immunosuppressive activity.Studies utilizing the glucocorticoid antagonist RU-486 showed thatalthough GaAs in the lung did increase circulating corticosteronelevels, elevated corticosterone was not responsible for suppressionof the AFC response. Rather, GaAs exerted direct immunosup-pressive effects independent of its ability to increase serum corti-costeroid levels.Mercury Both organic and inorganic mercury (Hg) has beenshown to decrease immunologic responses. Specifically, Hg expo-sure suppresses the PFC response and increases susceptibility toencephalomyocarditis (EMC) virus in addition to decreasing poly-clonal activation of lymphocytes by T-cell mitogens. It has alsobeen reported that Hg can activate B cells and augment anaphy-laxis by enhancing IgE production. Recently, interest in Hg has fo-cused on the ability of this metal to induce type III hypersensitiv-ity. Hg administration is used to induce glomerulonephritis inbrown Norway rats (a model for induction of autoimmune disease;Sapin et al., 1981).Cadmium Like other metals, cadmium (Cd) exposure increasessusceptibility to both bacterial and viral pathogens, although en-hanced resistance to tumor and EMC virus has been reported. Ex-posure to Cd has also been demonstrated to modulate lymphocyteproliferative responses to mitogens and allogeneic cells. Greenspanand Morrow (1984) reported decreased macrophage phagocyticability, which correlates with changes in host resistance. Humoralimmunity (PFC response and serum antibody titer) and NK func-tion have also been demonstrated to be suppressed by Cd expo-sure, while CTL activity appears to be enhanced.Other Metals Organotin compounds are used primarily as heatstabilizers and catalytic agents (dialkyltin compounds) and as bio-cides (trisubstituted organotins). The immunotoxicity of the organ-otins has been extensively reviewed (Penninks et al., 1990). Sincethe trisubstituted organotins are examined elsewhere (see “Pesti-cides,” above) discussion here is limited to the dialkyltins, di-n-octyltin dichloride (DOTC) and di-n-dibutyltin dichloride (DBTC).As in the case of tributyltin oxide (TBTO), the most outstandingaction of the dibutyltins is the induction of profound but reversiblethymic atrophy. Additionally, there is a preferential loss of CD4�
cells observed in the peripheral blood. The dialkylorganotins havealso been observed to decrease resistance to L. monocytogenes andto suppress the DHR and allograft rejection responses. Suppres-sion of the PFC response to sRBC and inhibition of T-cell mito-gen responses was also observed, while no effect on B-cell mito-genesis or the PFC response to LPS occurred. These data suggestthat the T cell may be a primary target for compounds like DOTCand DBTC. Like the trisubstituted organotins and the HAHs, the
2996R_ch12_419-470 4/27/01 10:02 AM Page 443
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

444 UNIT 4 TARGET ORGAN TOXICITY
developing immune system appears to be more sensitive to the ef-fects of these compounds than does the immune system of the adult.
Beryllium is known primarily for its ability to produce beryl-lium lung disease, a chronic granulomatous inflammation of thelung often observed in persons with occupational contact or envi-ronmental exposure to beryllium compounds. This metal producesa T-cell–mediated hypersensitivity and causes the in vitro trans-formation of PBL (from exposed but not unexposed patients) intolarge lymphoblasts (Hanifin et al., 1970). In addition, lymphocytesfrom beryllium oxide–exposed individuals produce migration in-hibitor factor (MIF) that inhibits the migration of macrophages(Henderson et al., 1972).
Platinum compounds have been used in cancer chemotherapyand have been shown to suppress macrophage chemotaxis, to in-hibit humoral immunity and lymphoproliferation, and to induce hy-persensitivity responses. Gold salts, used therapeutically in rheu-matic disease, may cause immune complex hypersensitivity andenhance allergic reactions. While nickel has been reported to en-hance anaphylaxis, it also inhibits humoral immunity, NK activity,and impairs resistance to pathogenic challenge. Chromium at lowdoses enhances phagocytic ability and PFC responses but appearsto suppress these responses at higher concentrations. Cobalt, a con-stituent of vitamin B12, has been demonstrated to suppress PMNchemotaxis and host resistance to streptococcal infection and to in-hibit the PFC response. Vanadium impairs the activities ofmacrophages and results in increased susceptibility to disease andbacterial challenge with L. monocytogenes.
Inhaled Substances Pulmonary defenses against inhaled gasesand particulates are dependent upon both physical and immuno-logic mechanisms. Immune mechanisms primarily involve thecomplex interactions between PMNs and alveolar macrophages andtheir abilities to phagocytize foreign material and produce cy-tokines, which not only act as local inflammatory mediators butalso serve to attract other cells into the airways.Urethane Urethane (ethyl carbamate) was once widely used asa veterinary anesthetic until its carcinogenic potential was definedin 1948. Exposure to urethane produces severe myelotoxicity, re-sulting in suppression of NK-cell activity and antibody responsesto sRBC (Luster et al., 1982; Gorelik and Heberman, 1981). In ad-dition, urethane exposure leads to increased frequency of sponta-neous lung adenomas in susceptible mouse strains and impairedresistance to B16F10 melanoma cells and metastatic tumor growthin the lungs.Tobacco Smoke Cigarette smoke has been implicated in acuterespiratory illness and chronic obstructive lung disease, but theeffect of exposure to mainstream cigarette smoke has yielded am-biguous results in humans and in animal models (reviewed by So-pori et al., 1994). In humans, the number of alveolar macrophagesis increased three- to fivefold in smokers compared to nonsmok-ers. This may be a result of increased production of IL-1 by theresident alveolar macrophages, resulting in enhanced influx ofother inflammatory cells (PMNs and peripheral blood mononuclearcells) into the lung. In addition to the increased numbers ofmacrophages, the macrophages present appear to be in an activatedstate, as evidenced by an increase in cytoplasmic inclusions, in-creased enzyme levels, altered surface morphology, and enhancedproduction of oxygen radicals. However, despite their apparent ac-tivated state, these macrophages seem to have decreased phago-cytic and bactericidal activity. Although the primary site of expo-sure of the immune system to cigarette smoke is the lung, selected
immune parameters have been shown to be altered in smokers. De-creased serum immunoglobulin levels and decreased NK-cell ac-tivity have been reported. Concentration-dependent leukocytosis(increased numbers of T and B cells) is well-defined in smokerswhen compared to nonsmokers. However, the question of whetherthere is a relationship between smoking and lymphocyte functionis debatable.
Numerous immunologic studies conducted in animals exposedto cigarette smoke demonstrate suppression of antibody responses,biphasic lymphoproliferative capacity (enhanced, then suppressedwith continued exposure), and enhanced susceptibility to murinesarcoma virus and influenza virus. Animal studies cannot preciselyreplicate human exposure conditions because of the route of ex-posure and the rapid chemical changes that occur in the compo-nents of tobacco smoke upon its generation.Particles: Asbestos and Silica It is believed that alterations inboth humoral and cell-mediated immunity occur in individuals ex-posed to asbestos and exhibiting asbestosis. Decreased DHR andfewer T cells circulating in the periphery as well as decreased T-cell proliferative responses have been reported to be associated withasbestosis (reviewed by Miller and Brown, 1985, and Warheit andHesterberg, 1994). Autoantibodies and increased serum im-munoglobulin levels have also been observed. Within the lung,alveolar macrophage activity has been implicated as playing a sig-nificant role in asbestos-induced changes in immunocompetence.Fibers of asbestos that are deposited in the lung are phagocytizedby macrophages, resulting in macrophage lysis and release of lyso-somal enzymes and subsequent activation of other macrophages.Recently it has been hypothesized that the development of as-bestosis in animal models occurs by the following mechanism.Fibers of asbestos deposited in the alveolar space recruitmacrophages to the site of deposition. Some fibers may migrate tothe interstitial space where the complement cascade becomes ac-tivated, releasing C5a, a potent macrophage activator and chemoat-tractant for other inflammatory cells. Recruited interstitial and res-ident alveolar macrophages phagocytize the fibers and releasecytokines, which cause the proliferation of cells within the lungand the release of collagen. A sustained inflammatory responsecould then contribute to the progressive pattern of fibrosis whichis associated with asbestos exposure.
The primary adverse consequence of silica exposure, like thatto asbestos, is the induction of lung fibrosis (silicosis). However,several immune alterations have been associated with silica expo-sure in experimental animals, including decreased antibody- andcell-mediated immune parameters (reviewed in IPCS, 1996). Al-terations in both T- and B-cell parameters have been reported, al-though T cell–dependent responses appear to be more affected thanB cell–dependent responses. Dose and route of antigen exposureappear to be important factors in determining silica-induced im-munomodulation. Silica is toxic to macrophages and PMNs, andexposure is correlated with increased susceptibility to infectiouspathogens. The significance of these immunologic alterations forthe pathogenesis of silicosis remains to be determined. The asso-ciation of this disease with the induction of autoantibodies is cov-ered elsewhere in this chapter.Pulmonary Irritants Chemicals such as formaldehyde, silica,and ethylenediamine have been classified as pulmonary irritantsand may produce hypersensitivity-like reactions. Macrophagesfrom mice exposed to formaldehyde vapor exhibit increased syn-thesis of hydroperoxide (Dean et al., 1984). This may contributeto enhanced bactericidal activity and potential damage to local tis-
2996R_ch12_419-470 4/27/01 10:02 AM Page 444
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

CHAPTER 12 TOXIC RESPONSES OF THE IMMUNE SYSTEM 445
sues. Although silica is usually thought of for its potential to in-duce silicosis in the lung (a condition similar to asbestosis), its im-munomodulatory effects have also been documented (Levy andWheelock, 1975). Silica decreased reticuloendothelial system(RES) clearance and suppressed both humoral immunity (PFC re-sponse) and the cell-mediated response (CTL) against allogeneicfibroblasts. Both local and serum factors were found to play a rolein silica-induced alterations in T-cell proliferation. Silica exposuremay also inhibit phagocytosis of bacterial antigens (related to RESclearance) and inhibit tumoricidal activity (Thurmond and Dean,1988).Oxidant Gases It is becoming increasingly clear that exposure tooxidant gases—such as ozone (O3), sulfur dioxide (SO2), nitrogendioxide (NO2), and phosgene—alters pulmonary immunologic re-sponses and may increase the susceptibility of the host to bacter-ial infections (reviewed by Selgrade and Gilmour, 1994). Infiltra-tion of both PMNs and macrophages has been observed, resultingin the release of cellular enzyme components and free radicals,which contribute to pulmonary inflammation, edema, and vascularchanges. Exposure to O3 has been demonstrated to impair thephagocytic function of alveolar macrophages and to inhibit theclearance of bacteria from the lung. This correlated with decreasedresistance to S. zooepidemicus and suggests that other extracellu-lar bacteriostatic factors may be impaired following exposure tothese oxidant gases. Short-term NO2 exposure decreases killing ofseveral bacterial pathogens and, like O3, this decreased resistanceis probably related to changes in pulmonary macrophage function.A role for the products of aracadonic acid metabolism (specifically,the prostaglandins) has recently been implied. This is supported bythe facts that decreased macrophage functions are associated withincreased PGE2 production and that pretreatment with in-domethacin inhibits O3-induced pulmonary hyperresponsivenessand related inflammatory responses.
It is clear that exposure to oxidant gases can also augmentpulmonary allergic reactions. This may be a result of increasedlung permeability (leading to greater dispersion of the antigen) andto the enhanced influx of antigen-specific IgE-producing cells inthe lungs. In studies involving O3 exposure and challenge with L.monocytogenes, decreased resistance to the pathogen correlated notonly with changes in macrophage activity, but with alterations inT cell–derived cytokine production (which enhances phagocyto-sis) as well. In support of an effect on T cells, other cell-mediatedchanges were observed: changes in the T- to B-cell ratio in thelung, decreased DHR response, enhanced allergic responses, andchanges in T-cell proliferative responses. Together, these data sug-gest that in addition to altering macrophage functions, oxidantgases may also produce an imbalance in the Th1 and Th2 cell pop-ulations. Given the different patterns of lymphokine secretion bythese T-cell subpopulations (Mosmann and Coffman, 1989) this isa very plausible explanation for some of the observed immune al-terations.
Organic Solvents and Related Chemicals Aromatic Hydrocar-bons There is limited but substantive evidence that exposure toorganic solvents and their related compounds can produce im-munosuppression (reviewed by Snyder, 1994). By far the best-characterized immunotoxic effects are those produced by benzene.In animal models, benzene induces anemia, lymphocytopenia, andhypoplastic bone marrow. In addition, it has recently been sug-gested that this myelotoxicity may be a result of altered differen-tiative capacity in bone marrow–derived lymphoid cells. Benzene
(oral and inhaled) exposure alters both humoral and cell-mediatedimmune parameters including suppression of the anti-sRBC anti-body response, decreased T- and B-cell lymphoproliferative re-sponses (mitogens and alloantigens), and inhibition of CTL activ-ity. Benzene exposure also appears to increase the production ofboth IL-1 and TNF-� and to inhibit the production of IL-2. Withthese dramatic effects on immune responses, it is not surprisingthat animals exposed to benzene exhibit reduced resistance to a va-riety of pathogens. More recently, nitrobenzene (an oxidizing agentused in the synthesis of aniline and benzene compounds) has beenreported to also produce immunotoxic effects (Burns et al., 1994a),with the primary targets being the peripheral blood erythrocyte andthe bone marrow.
Immunomodulating activity has also been observed fortoluene, although most effects occur at significantly high concen-trations. When compared with benzene, toluene has little to no ef-fect on immunocompetence. However, it should be noted thattoluene exposure effectively attenuates the immunotoxic effects ofbenzene (probably because of competition for metabolic enzymes).
In contrast to the parent toluene, the monosubstituted nitro-toluenes (para- and meta-nitrotoluene) do significantly alter theimmune system (Burns et al., 1994b,c). Exposure to p-nitrotoluenehas been demonstrated to suppress the antibody response to sRBC,to decrease the number of CD4� splenic T cells, and to inhibit theDHR to keyhole limpet hemocyanin (KLH). In addition, host re-sistance to L. monocytogenes was impaired, suggesting the T cellas a primary target. Similarly, m-nitrotoluene suppresses the anti-body response to sRBC, the DHR to KLH, T-cell mitogenesis, andhost resistance to L. monocytogenes, again suggesting the T cell asthe cellular target. The di-substituted nitrotoluene (2,4-dinitro-toluene; DAT) is also immunosuppressive (Burns et al., 1994d),with exposure resulting in suppressed humoral immunity, NK ac-tivity, and phagocytosis by splenic macrophages. Host resistanceto bacterial challenge was also impaired. It would appear that DATmay perturb the differentiation and maturation of leukocytes.Haloalkanes and Haloalkenes Carbon tetrachloride (CCl4) iswidely recognized as hepatotoxic. Recent studies have revealed thatCCl4 is also immunotoxic. Mice exposed for 7 to 30 days to CCl4(orally or intraperitoneally) exhibit a decreased T cell–dependentantibody response (sRBC), suppressed mixed lymphocyte re-sponses (allogeneic cells), and lower lymphoproliferative capacity(T and B cells). This change in immune status, which primarily af-fects helper T-cell function, is associated with the serum of treatedanimals, suggesting specific cytokine involvement. In fact, theCCl4-dependent induction and release of TGF-�1 from the liver re-sults in the indirect suppression of these T cell–mediated im-munologic responses (Delaney et al, 1994; Jeon et al., 1997). In-duction or inhibition of liver P450 activity augmented and blocked,respectively, the immunotoxic actions of CCl4, suggesting a re-quirement for metabolism in order for CCl4 to be immunosup-pressive. In contrast, Fischer 344 rats exposed orally for 10 daysexhibited no immunotoxic effects, despite signs of liver toxicity.These differences may represent differences in the metabolic ca-pabilities between these two species.
There is relatively little information on other solvents and re-lated chemicals. Exposure to dichloroethylene (in drinking waterfor 90 days) has been reported to suppress the anti-sRBC antibodyresponse in male CD-1 mice an to inhibit macrophage function intheir female counterparts (Shopp et al., 1985). Similarly, exposureto trichloroethylene (in the drinking water for 4 to 6 months) wasreported to inhibit both humoral and cell-mediated immunity and
2996R_ch12_419-470 4/27/01 10:02 AM Page 445
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

446 UNIT 4 TARGET ORGAN TOXICITY
bone marrow colony-forming activity (Sanders et al., 1982). Inthose experiments, females were more sensitive than males. Ex-posure to 1,1,2-trichloroethane results in suppression of humoralimmunity in both sexes. In addition, macrophage function was in-hibited (males only; Sanders et al., 1985). Finally, inhalation ofdichloroethane, dichloromethane, tetrachloroethane, and trichloro-ethene has been reported to suppress pulmonary host resistance toKlebsiella pneumoniae (Aranyi et al., 1986; Sherwood et al., 1987),suggesting that alveolar macrophages may be affected.Glycols and Glycol Ethers Exposure to glycol ethers has beenassociated with adverse effects in laboratory animals, includingthymic atrophy and mild leukopenia. Oral administration of ethyl-ene glycol monomethyl ether (EGME) for 1 to 2 weeks (House etal., 1985; Kayama et al., 1991) or its meatabolite methoxyaceticacid (MAA) for 2 weeks (House et al., 1985), produced decreasedthymic weight, thymic atrophy, and a selective depletion of im-mature thymocytes in mice. No alterations in humoral immunity,cell-mediated immunity, macrophage function, or host resistanceto L. monocytogenes were observed (House et al., 1985). More re-cently, it has been suggested that perinatal exposure to EGME mayproduce thymic hypocellularity and inhibition of thymocyte mat-uration and that it may affect pro-lymphocytes in fetal liver (Hol-laday et al., 1994).
Oral studies (5 to 10 days) on the glycol ether 2-methoxyethanol (ME) have consistently shown a decrease in thy-mus weight in the rat (Williams et al., 1995; Smialowicz et al.,1991a). This decrease is often accompanied by alterations in lym-phoproliferative responses, although suppression is seen in somecases and stimulation in others, with no clear reason for the dif-ferences in response. Alterations in spleen weight and splenic cellpopulations have also been observed, as well as suppression ofTNP-LPS and anti-sRBC plaque-forming-cell responses. Similarresults have been obtained following dermal exposure to 2-methoxyethanol (Williams et al., 1995). A decrease in IL-2 pro-duction has also been reported (Smialowicz et al., 1991a). Studiesusing the metabolites of 2-methoxyethanol (methoxyacetaldehydeand methoxyacetic acid) or specific metabolic pathway inhibitorshave shown that methoxyacetaldehyde (MAAD) and methoxy-acetic acid (MAA) are more immunotoxic than 2-methoxyethanolalone (MAAD�MAA�ME) (Smialowicz et al., 1991a,b; Kim andSmialowicz, 1997), suggesting a role for metabolism in the ob-served alterations in immunocompetence. Although there was noeffect following 10-day oral exposures to 2-methoxyethanol (50 to200 mg/kg/day) (Smialowicz et al., 1991a), subchronic exposurefor 21 days to 2000 to 6000 ppm (males) or 1600 to 4800 ppm (fe-males) did produce an enhanced NK response (Exon et al., 1991)in addition to suppression of the PFC response and a decrease inIFN-� production. In that study, it was also determined that 2-methoxyethanol produced greater immunotoxic effects than 2-butoxyethanol. 2-Butoxyethanol was observed to enhance NK ac-tivity, but only at the low doses.
Mycotoxins The immunotoxicity of mycotoxins, structurally di-verse secondary metabolites of fungi that grow on feed, has beenreviewed (IPCS, 1996). This class of chemicals comprises suchtoxins as aflatoxin, ochratoxin, and the tricothecenes, notably T-2toxin and vomitoxin. As a class, these toxins can produce cellulardepletion in lymphoid organs, alterations in T- and B-lymphocytefunction, suppression of antibody responses, suppression of NKactivity, decreased delayed hypersensitivity responses, and an ap-parent increase in susceptibility to infectious disease. T-2 toxin has
also been implicated as a developmental immunotoxicant, target-ing fetal lymphocyte progenitors leading the thymic atrophy oftenobserved with these mycotoxins ( Holladay et al., 1993). For ochra-toxin, at least, the dose, the route of administration, and the speciesappear to be critical factors in results obtained in immunotoxicitystudies. For the extensively studied tricothecenes, the mechanismof immunoimpairment may be related to inhibition of protein syn-thesis. The tricothecenes are currently considered among the mostpotent small-molecule inhibitors of protein synthesis in eukaryoticcells (IPCS, 1996).
Natural and Synthetic Hormones It is well established that asexual dimorphism exists in the immune system. Females havehigher levels of circulating immunoglobulins, a greater antibodyresponse, and a higher incidences of autoimmune disease than domales. Males appear to be more susceptible to the development ofsepsis and mortality associated with this following soft tissuetrauma and hemorrhagic shock. Specific natural sex hormones inthis dichotomy have been implicated. Immune effects of androgensand estrogens appear to be very tightly controlled within the phys-iologic range of concentrations, and profound changes in immuneactivity can result for very slight changes in concentrations of hor-mones.Estrogens Diethylstilbestrol (DES) is a synthetic nonsteroidalcompound possessing estrogenic activity. DES was used in men totreat prostatic cancer and in women to prevent threatened abor-tions, as an estrogen replacement, and as a contraceptive drug.Extensive functional and host resistance studies on DES (mg/kg/day range) have indicated that exposure to this chemicalresults in alterations in cell-mediated immunity and/or macrophagefunction and are believed to be mediated by the presence of the es-trogen receptor on immune cells (Kalland, 1980; Luster et al., 1980,1984a,b; Holsapple et al., 1983). Targeted sites of action includethe thymus (thymic depletion, alteration in T-cell maturationprocess), T cells (decreased MLR, DTH, lymphoproliferative re-sponses), and macrophage (enhanced phagocytic, anti-tumor, andsuppressor function). Pre- and neonatal exposures (�g/kg/day doserange) have also demonstrated immunotoxic effects related to T-cell dysfunction. Delayed hypersensitivity and inflammatory re-sponses associated with DES exposure in adult mice have beenshown to be reversible upon cessation of exposure (Luster et al.,1980; Holsapple et al., 1983). However, effects from in utero andneonatal exposures appear to have more lasting, possibly perma-nent effects on immune responses (Luster et al., 1979; Kalland etal., 1979; Ways, 1980).
Exposure to 17�-estradiol in male rats (63 days of age) in-traperitoneally for 15 days (1 to 50 �g/kg/day) did not alter spleenweight, spleen cellularity, or the humoral immune response tosRBC (Ladics et al., 1998). As observed with other estrogenicagents, thymic weight was decreased following exposure. Serumandrogens and luteinizing hormone and male accessory organweights were depressed, while serum estradiol and prolactin wereincreased. Dietary exposure (2.5 to 50 ppm) of male and femalerats for 90 days resulted in decreased spleen weights and alter-ations in hematologic elements suggestive of bone marrow effects.Body weights were also affected. No histological alterations werenoted. Decreases in splenic T- and B-cell populations were ob-served at the higher concentrations. These data suggest the possi-bility that exposure to 17�-estradiol may have resulted in alterednormal immune cell trafficking and distribution, the mechanism ofwhich is not clear. This hypothesis is supported by recent data in-
2996R_ch12_419-470 4/27/01 10:02 AM Page 446
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

CHAPTER 12 TOXIC RESPONSES OF THE IMMUNE SYSTEM 447
dicating the observed anti-inflammatory effects of estrogens maybe related to combination of alterations in homing and the acti-vation of inflammatory cells and their production of TNF-� andIFN-� (Salem et al., 2000).
While it appears that estrogens can affect the maturation andfunction of the thymus and its components, it has recently beenobserved that estrogen receptor knockout mice have significantlysmaller thymi than do their wild-type littermates, apparently dueto the lack of the estrogen receptor-alpha (ER�) (Staples et al.,1999). In addition, it has been suggested that the effects of estro-gens on the thymus appear to be mediated not only through ER�but also through another pathway. Androgens Oxymetholone is a synthetic androgen structurallyrelated to testosterone and used in the past in the treatment ofpituitary dwarfism and as an adjunctive therapy in osteoporosis. Itscurrent use is limited to treatment of certain anemias.Oxymetholone was administered orally to male mice daily for 14consecutive days (50 to 300 mg/kg/day). In male mice,oxymetholone exposure resulted in a minimal decrease in cell-mediated immunity (MLR and CTL response) but did not alter theability of the animals to resist infection in host resistance assays.In contrast, anabolic androgenic steroids have been shown to sig-nificantly inhibit the sRBC PFC response and to increase the pro-duction of pro-inflammatory cytokines from human peripheralblood lymphocytes.
Exposure to flutamide, an androgen receptor antagonist, inmale rats (63 days of age) intraperitoneally for 15 days (0.25 to20 mg/kg/day) did not alter spleen weight or the humoral immuneresponse to sRBC (Ladics et al., 1998). Relative thymic weight andtotal spleen cellularity were minimally increased. Serum andro-gens, estrogens, luteinizing hormone, and follicle-stimulating hor-mone were increased, while male accessory organ weights weredepressed.
No comprehensive studies evaluating the effects of testos-terone on immune parameters have been conducted. However, it isclear that testosterone is capable of contributing to the suppressionof immune function—in particular, cell-mediated responses andmacrophage activity. There are numerous reports in the clinical lit-erature that males are more susceptible than females to infectionfollowing soft tissue trauma and hemorrhagic shock (reviewed inCatania and Chaudry, 1999). Treatment of males with agents thatblock testosterone (e.g., flutamide) can prevent the trauma- andhemorrhage-induced depression of immunity. Similarly, treatmentof females with dihydrotestosterone prior to trauma-hemorrhageresults in depression of CMI similar to that of males. Furthermore,gonadectomized mice of either sex have elevated immune re-sponses to endotoxin, which can be attenuated in either sex by theadministration of testosterone. The mechanisms in these cases, in-cluding influences of the neuroendocrine system, are not clear.Other investigators have reported that, like estrogenic agents,testosterone and other androgens are capable of influencing hostdefense by altering lymphocyte trafficking in the body and alter-ing the ability of the macrophage to participate in immune re-sponses.Glucocorticoids The immunosuppressive actions of corticos-teroids have been known for years. Following binding to an intra-cellular receptor, these agents produce profound lymphoid cell de-pletion in rodent models. In nonhuman primates and humans,lymphopenia associated with decreased monocytes and eosinophilsand increased PMNs are seen. Corticosteroids induce apoptosis andT cells are particularly sensitive. In addition, these agents inhibit
macrophage accessory cell function, the production of IL-1 fromthe macrophages, and the subsequent synthesis of IL-2 by T cells.In general, corticosteroids suppress the generation of CTL re-sponses, MLR, NK activity, and lymphoproliferation. While it isclear that these drugs inhibit T-cell function, their effects on B cellsare not completely clear. Corticosteroids inhibit humoral responses,but this appears to be due to effects on T cells, as antigen-specificantibody production by B cells to T-independent antigens does notappear to be affected by corticosteroid treatment.
Mifepristone (RU-486) is a potent competitive antagonist ofboth progesterone receptor and glucocorticoid type 1 receptor bind-ing. While use of mifepristone has focused on termination of preg-nancy, it may find use as a contraceptive, anti-cancer agent, or in-ducer of labor. No significant studies could be located examiningthe immunotoxicity of mifepristone. However, in studies utilizingthis compound as a glucocorticoid type 1 receptor antagonist todemonstrate the role of corticosteroids in chemical-induced im-munomodulation, some insights can be gleaned. Acute oral ad-ministration of mifepristone to mice at 100 mg/kg did elevate cir-culating levels of corticosterone, presumably by preventing bindingto the type 1 glucocorticpoid receptor and thus altering the feed-back loop. However, the chemical did not alter humoral immunityor the weights and cellularity of lymphoid organs. Similarly, no ef-fects on thymic or splenocyte cellular subpopulations have beenobserved.
Therapeutic Agents Historically speaking, very few drugs usedtoday as immunosuppressive agents were actually developed forthat purpose. In fact, if one looks closely enough, nearly all ther-apeutic agents possess some degree of immunomodulatory activ-ity (Descotes, 1986). The recent explosion of knowledge regard-ing the function and regulation of the immune system (at thecellular, biochemical, and molecular levels) has provided investi-gators with a relatively new avenue for specific drug development.The following discussion focuses on those drugs used primarily formodulating the immune system: the immunosuppressants (corti-costeroids are considered above), AIDS therapeutics, and the re-combinant cytokines. Extensive reviews of these drugs can befound elsewhere (Spreafico et al., 1985; Rosenthal and Kowolenko,1994; Talmadge and Dean, 1994).Immunosuppressive Drugs Originally developed as an antineo-plastic agent, cyclophosphamide (Cytoxan, CYP) is the prototyp-ical member of a class of drugs known as alkylating agents. Uponentering the cell, the inactive drug is cleaved into phosphoramidemustard, a powerful DNA alkylating agent that leads to blockadeof cell replication. Clinically, CYP has found use in reducing symp-toms of autoimmune disease and in the pretreatment of bone mar-row transplant recipients. Experimentally, this drug is often usedas a positive immunosuppressive control in immunotoxicologystudies because it can suppress both humoral and cell-mediatedimmune responses. There appears to be preferential inhibition ofB-cell responses, possibly due to decreased production and surfaceexpression of immunoglobulins. CMI activities that are suppressedinclude the DHR, CTL, graft-versus-host (GVH) disease, and theMLR.
Azathioprine (AZA), one of the antimetabolite drugs, is apurine analog that is more potent than the prototype, 6-mercap-topurine, as an inhibitor of cell replication. Immunosuppressionlikely occurs because of the ability of the drug to inhibit purinebiosynthesis. It has found widespread use in the inhibition of al-lograft rejection, although it is relatively ineffective in attenuating
2996R_ch12_419-470 4/27/01 10:02 AM Page 447
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

448 UNIT 4 TARGET ORGAN TOXICITY
acute rejection reactions. It can also act as an anti-inflamma-tory drug and can reduce the number of PMNs and monocytes.Clinical use of the drug is limited by bone marrow suppression and leukopenia. AZA inhibits humoral immunity, but secondary responses (IgG) appear more sensitive than primary respons-es (IgM). A large range of CMI reactivities are also reduced by AZA treatment, including DHR, MLR, and GVH disease. Al-though T-cell functions are the primary targets for this drug, inhi-bition of NK function and macrophage activities has also been re-ported.
Cyclosporin A (Sandimmune, CsA) is a cyclic undecapeptideisolated from fungal organisms found in the soil. Important to itsuse as an immunosuppressant is the relative lack of secondary tox-icity (e.g., myelotoxicity) at therapeutic concentrations (Calne etal., 1981). However, hepatotoxicity and nephrotoxicity are limit-ing side effects. CsA acts preferentially on T cells by inhibiting thebiochemical signaling pathway emanating from the T-cell receptor(TCR). The result is inhibition of IL-2 gene transcription and sub-sequent inhibition of T-cell proliferation. More specifically, CsAinteracts with the intracellular molecule cyclophillin, an intracel-lular protein with peptidyl proline isomerase activity (although thisenzymatic activity probably has nothing to do with the immuno-suppressive effect of CsA). The CsA–cyclophillin complex inhibitsthe serine/threonine phosphatase activity of a third molecule, cal-cineurin. Calcineurin is proposed to dephosphorylate the cytoplas-mic subunit of NF-AT (nuclear factor of activated T cells) and al-low the transport of NF-AT into the nucleus, where it can couplewith nuclear components and induce the transcription of the IL-2gene. Inhibition of calcineurin phosphatase activity by the CsA–cyclophillin complex then prevents nuclear translocation of NF-ATand the resulting IL-2 gene transcription.
FK506 is a cyclic macrolide which is structurally distinct fromCsA, but which possesses a nearly identical mechanism of action.Like CsA, FK506 binds intracellularly to proteins with peptidylproline isomerase activity, the most abundant of which is FK506binding protein-12 (FKBP12). The FK506–FKBP12 complex alsobinds to and inhibits calcineurin activity, thereby inhibiting IL-2gene transcription. Clinically, FK506 inhibits T-cell proliferation,lacks myelotoxicity (although, like CsA, it does cause nephrotox-icity), and induces transplantation tolerance. In addition, the min-imum effective dose appears to be approximately tenfold lowerthan that of CsA.
Rapamycin (RAP) is also a cyclic macrolide which is struc-turally related to FK506. However, the mechanism by which it pro-duces inhibition of proliferation is strikingly distinct. Unlike CsAand FK506, RAP does not inhibit TCR-dependent signaling eventsand IL-2 gene transcription. Rather, this compound inhibits IL-2–stimulated T-cell proliferation by blocking cell-cycle progressionfrom late G1 into S phase (Morice et al., 1993; Terada et al., 1993).Like FK506, RAP binds to the intracellular protein FKBP12. Butthis RAP–FKBP12 complex does not bind calcineurin. Moreover,until very recently, the actual target protein or proteins of this com-plex have remained elusive. Now it is clear that the RAP–FKBP12complex binds to the mammalian target of rapamycin, mTOR(Sabers et al., 1995), also referred to as FRAP-1 and RAFT-1(Brown et al., 1994; Sabatini et al., 1994). This protein, originallyidentified as two proteins (TOR-1 and TOR-2) in rapamycin-re-sistant yeast mutants (Kunz et al., 1993), has homology to the lipidkinase domain of the p110 catalytic subunit of phosphatidylinosi-tol 3-kinase (PI3K, a biochemical signaling molecule) and VPS34(a yeast PI3K). The function of the TOR proteins in cellular reg-
ulation (specifically cell-cycle progression) remains unknown atthis time. Unlike both CsA and FK506, RAP does not appear tobe nephrotoxic.
Leflunomide, an isoxazole derivative, is a relatively new drugthat has shown promise as an immunosuppressive agent in thetreatment of rheumatic disease and transplantation (Xiao et al.,1994). Experimentally, this agent can block the generation of allo-specific antibodies, decrease the mononuclear infiltrate in graftsundergoing rejection, and reverse acute graft rejections. It has beenfound to be equal to or better than CsA in its ability to inhibit Bcell–mediated autoimmune disease. Early mechanistic studies in-dicate that leflunomide can directly inhibit B-cell proliferation(IC50 � 20 �M), and this may account for the drug’s ability to in-hibit both T-cell–dependent and T-cell–independent specific anti-body production. Leflunomide also can inhibit T-cell proliferation(IC50 50 to 75 �M) induced by mitogens or antibody directedagainst CD3 or IL-2. IL-2 production is also attenuated, but ex-pression of the IL-2 receptor (CD25) is not altered. Biochemicalanalyses indicate that this drug can inhibit IL-2–dependent proteintyrosine kinase activity and suggest that the mechanism of T-cellinhibition may be at the level of T-cell responsiveness to IL-2. Al-though similar in broad terms, this mechanism of action is dis-tinctly different from the mechanism of action of RAP.Aids Therapeutics Traditionally, antiviral therapies have not beenextremely successful in their attempt to rid the host of viral infec-tion. This may be due to the fact that these organisms target theDNA of the host. Thus, eradication of the infection means killinginfected cells. Although numerous strategies have been developedto combat the AIDS virus (primarily targeting viral reverse tran-scriptase or viral protease and up-regulation of other immune re-sponses), no one drug has produced any significant advance. Thisis possibly because the very nature of the infection has significantimmunosuppressive consequences. Without doubt, more basic sci-entific knowledge about the physiology and biochemistry of thevirus is required before rational drug design will yield an effectivetherapeutic agent.
Zidovudine (3�-azido-3�-deoxythymidine; AZT) is a pyrimi-dine analog that inhibits viral reverse transcriptase. It was the firstdrug shown to have any clinical efficacy in the treatment of HIV-1infection. Unfortunately, its use is limited by myelotoxicity (macro-cytic anemia and granulocytopenia). Animal studies have confirmedthat the primary action of AZT is on innate immunity, althoughchanges in both humoral and cell-mediated immunity have also beenobserved. Clinically, AZT increases the number of circulating CD4�
cells and can transiently stimulate cell-mediated immune responses(lymphoproliferation, NK activity, and IFN-� production).
Stavudine (2�,3�-didehydro-2�,3�-dideoxythymidine; d4T) isanother pyrimidine analog currently in clinical trials. Unlike its sis-ter drug AZT, the limiting toxicity appears to be peripheral neu-ropathy rather than myelotoxicity. In addition, d4T also appears toincrease the number of circulating CD4� cells. Animal studies sug-gest that d4T does not modulate generation of CTL, NK activity,PFC responses, mitogenicity of lymphocytes, or lymphocyte sub-sets.
Zalcitabine (2�,3�-dideoxycytidine; ddC) is a third pyrimidineanalog which has recently been approved for use. Clinically, thereappears to be an increase in circulating CD4� cells and somerestoration of CMI in HIV-infected persons. There also appears tobe no significant myelotoxicity and, like d4T, the limiting toxic ef-fect of ddC is peripheral neuropathy. Investigations in animals re-vealed no significant effect on immune status.
2996R_ch12_419-470 4/27/01 10:02 AM Page 448
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

CHAPTER 12 TOXIC RESPONSES OF THE IMMUNE SYSTEM 449
Videx (2�,3�-dideoxyinosine; ddI) is the first purine analog ap-proved for use in HIV infection. In clinical trials, the dose-limit-ing toxicities were shown to be peripheral neuropathy and pancre-atitis. There appears to also be an increase in circulating CD4�
cells, some restoration of CMI, and a reversal of HIV-inducedmyelotoxicity. Although ddI is converted into ddA-TP, the use ofddA as an antiviral agent was ruled out due to severe nephrotoxiceffects. In animal models, both ddI and ddA produce suppressionof humoral immunity.Recombinant DNA–Derived Proteins The development of ther-apeutic proteins for clinical use is an ever expanding arena for bothlarge and small pharmaceutical and biotechnology companies (re-viewed in Warner and Haggerty, 1997). In general, biologics (e.g.,blood or vaccine products) and recombinant DNA–derived pro-teins are derived in some manner from living organisms. Becauseof the very nature of many of the recombinant DNA–derived pro-teins, the immune system is often the target not only of therapybut also of toxicity. Manifestations of toxicity include exaggeratedpharmacology, effects due to receptor-biochemical cross-talk, anddisruptions in immune regulation by cytokine networks. Mono-clonal antibodies can bind normal as well as targeted tissues, andany foreign protein may elicit the production of neutralizing anti-bodies against the therapeutic protein (i.e., the therapeutic proteinmay be immunogenic). The effects of neutralizing antibodies mayalso lead to hypersensitivity reactions via either IgE production orimmune complex disease, resulting from the activation of the com-plement cascade and the subsequent release of vasoactive aminesthat produce anaphylaxis. The majority of recombinant DNA–derived proteins have been used as immunostimulants; these in-clude IFN-�, IFN-�, GM-CSF, and erythropoietin (EPO). Their im-munopharmacology (and toxicity) has been reviewed extensivelyelsewhere (Talmadge and Dean, 1994). An excellent review of thetoxicity associated with CTLA4Ig, anti-CD3, and anti-CD4 (forimmunosuppression, transplantation, autoimmunity), and IL-12 (asa cancer therapeutic and an immuno-stimulant), along with otherimmune related issues in the development of therapeutic proteinsis provided in Warner and Haggerty (1997).
Drugs of Abuse Drug abuse is a social issue with far-reachingeffects on the abuser as well as on friends and family. While drugparaphernalia has been directly associated with the spread of theAIDS virus, in recent years the actual abuse of some drugs hasbeen linked to the progression, and possibly the onset, of AIDS.Although definitive scientific proof of the hypothesis is lacking,drugs which are often abused have been shown to alter immuno-competence.Cannabinoids Much attention has been focused on the im-munomodulatory effects of the cannabinoids (�9-tetrahydro-cannabinol; THC) owing to the therapeutic potential of this drugin the treatment of glaucoma and as an antiemetic in patients un-dergoing cancer chemotherapy. Early studies showed that exposureto THC decreases host resistance to bacterial and viral pathogens(reviewed by Kaminski, 1994). In addition, cannabinoids alter bothhumoral and cell-mediated immune responses. Suppression of NKand CTL activity by THC appears to be related to an effect oc-curring subsequent to target-cell binding. THC exposure also al-ters macrophage morphology and some nonspecific functions, butthe effects on accessory cell activities (e.g., antigen processing andpresentation) are only beginning to be elucidated. Recently, it hasbeen shown that THC increases aspartyl cathepsin D proteolyticactivity and impairs lysosomal processing in the macrophage
(Matveyeva et al., 2000). What is clear, however, is that the sup-pression of humoral immunity is exquisitely dependent upon thetemporal association between exposure and antigen sensitization.Oral exposure to THC during the sensitization process (in vivo anti-gen administration) suppresses the PFC response to sRBC. In con-trast, exposure to THC prior to sensitization (but not during thesensitization time) resulted in no observable effects on the PFC re-sponse. This may be one of the most critical factors influencingthe reported effects of THC on immune responsiveness.
As in the in vivo situation, for in vitro THC exposure (Mishell-Dutton cultures), the drug must be added within 2 h of the addi-tion of antigen in order to suppress the PFC response (Schatz etal., 1992). Also, humoral responses to T-cell–dependent antigensbut not T-cell–independent antigens are suppressed by THC ex-posure. Together with the fact that T-cell proliferative responsesare suppressed after THC exposure in vivo, this suggests that THCaffects primarily T cells and may alter early T-cell activation events(e.g., biochemical signaling). Most recently, cannabinoid receptortranscripts have been identified in human spleen, tonsils, periph-eral blood lymphocytes, and macrophages (Bouaboula et al., 1993;Munro et al., 1993). Additionally, murine splenocytes exhibit a highdegree of saturable, specific binding of THC with a Kd approxi-mating 1 nM and a Bmax of about 1000 receptors per cell (Kamin-ski et al., 1992). An understanding of a potential role of this re-ceptor in immune responses awaits identification of the endogenousligand.Cocaine Cocaine is a potent local anesthetic and central nervoussystem (CNS) stimulant. This drug and its derivatives have beenshown to alter several measures of immunocompetence, includinghumoral and cell-mediated immune responses and host resistance(Watson et al., 1983; Ou et al., 1989; Starec et al., 1991). Func-tions of PMNs—including superoxide production and cell-surfacereceptor expression as well as inhibition of macrophage killing abil-ity by decreasing the production of reactive oxygen intermedi-ates—have been reported (Haines et al., 1990; Lefkowitz et al.,1993). Cocaine also induces the secretion of TGF-�, which hasbeen linked to the observation that cocaine exposure enhancesreplication of the HIV-1 virus in human peripheral blood mononu-clear cells (PBMC) (Chao et al., 1991; Peterson et al., 1991). Hols-apple et al. (1993) evaluated the effect of in vitro cocaine expo-sure on the generation of an antibody response against sRBCs andfound effects only at concentrations that were not clinically rele-vant (100 �M; lethal blood concentrations are estimated to bearound 6 �M). These investigators postulated that the immuno-suppressive effects of cocaine in vivo were mediated by P450-generated reactive intermediates; they subsequently demonstratedsex and strain differences in cocaine immunosuppressive activity,which correlated with well-characterized differences in metaboliccapability in mice. Male B6C3F1 mice are more sensitive than fe-males and DBA/2 females are more sensitive than B6C3F1 females.Opioids: Heroin and Morphine Chronic morphine exposure hasbeen associated with increased susceptibility to both bacterial andviral antigens (Arora et al., 1990; Chao et al., 1990), and it is clearthat exposure to opioids can suppress immune responses. What isnot clear, however, is whether this action is a direct effect of thedrug on immune cells or an indirect effect resulting from drug-induced increases in circulating corticosteroids. Evaluation of theimmunocompetence of heroin addicts revealed a decrease in totalT cells and E-rosette capability (McDonough et al., 1980). In thatstudy, treatment with naloxone reversed these effects, suggestinga role for an opioid receptor in mediating immune suppression.
2996R_ch12_419-470 4/27/01 10:02 AM Page 449
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

450 UNIT 4 TARGET ORGAN TOXICITY
LeVier and coworkers (1994) reported that chronic morphine ex-posure decreased serum C3, NK activity, total leukocyte counts,the PFC response to sRBC, and RES clearance. However, becausemany of these effects were not dose-related (i.e., the dose–responsecurve was flat), the investigators concluded that these effects werenot receptor-mediated but were the result of increased circulatingcorticosteroids (which were significantly elevated in those ani-mals). This conclusion is supported by the findings of other in-vestigators as well (Pruett et al., 1992b). Morphine-induced sup-pression of macrophage phagocytosis and cytokine production hasalso been reported (Eisenstein et al., 1993; Tubaro et al., 1987;LeVier et al., 1993). In the study by LeVier and colleagues (1993),the glucocorticoid antagonist RU-486 was utilized to demonstratethat while suppression of hepatic macrophage function may be duein part to a receptor-mediated event, inhibition of splenicmacrophage activity was wholly receptor-independent. A reviewby Giorgio and colleagues (1996) eloquently describes a likely hy-pothesis (involving neuroendocrine immunologic mechanisms) forthe dual effects observed with opioids as well as a few other com-pounds with similar effects.Ethanol Until recently (reviewed by Jerrells and Pruett, 1994),data concerning the immunomodulatory effects of ethanol (EtOH)exposure have largely been based on clinical observations of alco-holic patients. A prime reason for this is that rodents (the animalmodel of choice for extensive immunologic evaluation) do not vol-untarily consume intoxicating quantities of EtOH. Thus, the crite-ria for the development of animal models for EtOH exposure needto be refined to assure that clinically relevant blood levels are at-tained and long-term exposure can be assessed. In addition, the ef-fect of acute exposure (binge drinking) needs to be further assessed.
In humans, alcoholism is associated with an increased inci-dence of pulmonary infection and mortality from it. There is alsoan increased incidence of bacterial infection and spontaneous bac-teremia in alcoholics with cirrhosis of the liver. A consistent find-ing in abusers of EtOH is the significant change in the mononu-clear cells of the peripheral blood. In animal models, this isobserved as depletion of T and B cells in the spleen and the T cellsin the thymus, particularly CD4�/CD8� cells. The latter effect maybe related in part to increased levels of corticosteroids.
There are also numerous indications that acute EtOH exposurecan have profound immunodepressive consequences: decreasedPMN chemotaxis, decreased host resistance, and inhibition of thePFC response. EtOH administration also inhibits mitogen-drivenT-cell proliferation and T-cell responsiveness to IL-2. The actionsof EtOH exposure on B-cell antibody production and NK-cell ac-tivity are still controversial.
Electromagnetic Fields Several epidemiologic studies have sug-gested an association (albeit very small) between low-frequency(LF) (300 Hz) electromagnetic fields (EMF) and cancer. Otherepidemiologic studies have found no association. In response togrowing concern over the effects of LF EMF (300 Hz) on hu-man health, many studies have been conducted to assess their abil-ity to alter immunocompetence. While some studies, using in vitroexposures of animal or human immune cells, have shown mixedresults, a comprehensive evaluation of immunocompetence in mice(using the NTP tier approach) has demonstrated that exposure toLF EMF (60 Hz) for 28 or 90 days does not alter immunocompe-tence (House et al., 1996).
Ultraviolet Radiation The immunomodulatory effects of ultra-violet radiation (UVR) have been reviewed (IPCS, 1996). UVR has
been demonstrated to suppress delayed hypersensitivity responsesin both animals and humans and to result in decreased host resist-ance to infection. The dose of UVR required to suppress the im-mune response depends on the strain of mouse and the antigen be-ing used. Interestingly, however, it appears that the dose of UVRis not as important to the observed immunosuppression as is theinterval between irradiation and antigen exposure (Noonan and DeFabo, 1990). The mechanism of immunosuppression by UVR isnot completely clear. Induction of suppressor T cells or alterationsin homing patterns have been suggested as possibilities. One plau-sible explanation is that UVR induces a switch from a predomi-nantly Th1 response (favoring delayed hypersensitivity responses)to a Th2 response (favoring antibody responses). This hypothesisis supported by findings of altered cytokine secretion patterns in-dicative of a Th1 to Th2 switch (Araneo et al., 1989; Simon et al.,1990). This switch may explain decreased resistance to some in-fectious pathogens. It may also be related to effects of UVR onLangerhans cells in the skin.
Food Additives Thirty-five food flavoring ingredients (generallyrecognized as safe) have been screened for potential immunotoxi-city. Compounds were administered to mice by oral gavage for 5consecutive days at multiple dose levels (Gaworski et al., 1994).In addition to body weights, lymphoid organ weights, and cellu-larity, humoral immunity (sRBC PFC response) and host resistanceto L. monocytogenes were evaluated. Only two materials gave re-sults suggestive of enhanced susceptibility to infection (peppermintoil and citral dimethyl acetal). Only one material exhibited sup-pression of humoral immunity (hexanoic acid), and suppressionwas observed only in the presence of overt toxicity.
Silicon-Based Materials Silicon-based materials have knownuses in consumer products such as cosmetics, toiletries, food stuffs,household products, and paints as well as in the medical field (e.g.,as lubricants in tubing and syringes and as components in numer-ous implantable devices). In recent years, significant interest hasfocused on the biocompatibility of certain silicon-based materials(silicones) and the potential for these products to produce im-munotoxic effects. Despite the fact that there has been persistent,unsubstantiated speculation that breast implants made with siliconematerials may provoke connective tissue disease, no link betweenexposure to silicones and human disease has been established. Re-cently, a committee formed by the Institute of Medicine has con-cluded that “a review of the toxicology studies of silicones andother substances known to be in breast implant does not provide abasis for health concerns.” An extensive report by the IOM Com-mittee, including additional conclusions and recommendations forresearch, is publicly available (IOM, 2000).
Numerous studies have been conducted that both support andrefute the specific actions of various silicon-based materials on theimmune system. Two studies have been reported that extensivelyevaluated immune status following exposure to di-methylpolysiloxanes used in medical practice (Bradley et al.,1944a,b). In the first study, mice were implanted for 10 days withdimethylpolysiloxane fluid, gel, and elastomer as well aspolyurethane as a control. There were no observable alterations ininnate or acquired immune function. In fact, the materials testedafforded modest protection to an approximate LD50 challenge withL. monocytogenes. Implantation of the same materials for 180 daysresulted in a modest suppression of NK-cell activity that did notcorrelate with altered susceptibility to challenge with B16F10melanoma. No alterations in host resistance have been observed.
2996R_ch12_419-470 4/27/01 10:02 AM Page 450
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

CHAPTER 12 TOXIC RESPONSES OF THE IMMUNE SYSTEM 451
Studies have also been conducted on two-low molecular-weight cyclic siloxanes: octamethylcyclotetrasiloxane anddecamethylcyclotetrasiloxane. One-month inhalation exposures ofrats to high concentrations of octamethylcyclotetrasiloxane (up to540 ppm) and decamethylcyclotetrasiloxane (up to a maximum 160ppm) did not result in alterations in humoral immunity (Burns-Naas et al., 1998; Klykken et al., 1999). Inhalation exposure of hu-man volunteers to octamethylcyclotetrasiloxane at 10 ppm for 1 hrevealed no effects on several immune parameters (Looney et al.,1998). Repeated oral exposure of rodents to octamethylcyclote-trasiloxane at high concentrations has been observed to produceimmunomodulatory activity in both humoral and innate immunity;however, as with other chemicals, there are concentrations belowwhich no immunomodulation is observed (LeVier et al., 1995; Wil-son et al., 1995; Le and Munson, 1997; Munson et al., 1997; Mun-son, 1998).
Finally, under highly specific experimental conditions that donot mimic human exposure, a few silicon-based materials havebeen observed to act as immunologic adjuvants (reviewed in Potterand Rose, 1996, and IOM, 2000; Woolhiser et al., 1995). Undertypical exposure conditions, neither octamethylcyclotetrasiloxane,a combination of octamethylcyclotetrasiloxane and decamethylcy-clotetrasiloxane, nor dimethylpolysiloxane acts as an immunologicadjuvant (Bradley et al., 1994a,b; Klykken and White, 1996;Klykken et al., 1999; Vohr and Bomhard, 2000).
Immune-Mediated Disease
As stated earlier, the purpose of the immune system is to protectthe individual from disease states, whether infectious, parasitic, orcancerous—through both cellular and humoral mechanisms. In sodoing, the ability to distinguish “self” from “nonself” plays a pre-dominant role. However, situations arise in which the individual’simmune system responds in a manner producing tissue damage,resulting in a self-induced disease. These disease states fall intotwo categories (1) hypersensitivity, or allergy, and (2) autoimmu-nity. Figure 12-16 is a schematic delineating the possible cascadeof effects that can occur when a chemical produces an immune-mediated disease. Hypersensitivity reactions result from the im-
mune system responding in an exaggerated or inappropriate man-ner. These reactions have been subdivided by Coombs and Gell(1975) into four types, which represent four different mechanismsleading to tissue damage. In the case of autoimmunity, mechanismsof self-recognition break down and immunoglobulins and T-cellreceptors react with self-antigens, resulting in tissue damage anddisease.
Hypersensitivity Classification of Hypersensitivity Reactions Onecharacteristic common to all four types of hypersensitivity reac-tions is the necessity of prior exposure leading to sensitization inorder to elicit a reaction upon subsequent challenge. In the case oftypes I, II and III, prior exposure to antigen leads to the produc-tion of specific antibody, IgE, IgM, or IgG, and, in the case of typeIV, to the generation of memory T cells. Figure 12-17 illustratesthe mechanisms of hypersensitivity reactions as classified byCoombs and Gell. Although not completely understood, regulationof immunoglobulin production is dependent in part on the charac-teristics of the antigen, the genetics of the individual, and envi-ronmental factors. The mechanisms of antibody production in hy-persensitivity reactions are identical to those described earlier inthe chapter (Fig. 12-7). A brief description of the four types of hy-persensitivity reactions is presented below.
Type I (Immediate Hypersensitivity) Using penicillin as anexample, Fig. 12-18 depicts the major events involved in a type Ihypersensitivity reaction. Sensitization occurs as the result of ex-posure to appropriate antigens through the respiratory tract, der-mally, or by exposure through the gastrointestinal tract. IgE pro-duction is highest in lymphatic tissues that drain sites of exposure(i.e., tonsils, bronchial lymph nodes, and intestinal lymphatic tis-sues, including Peyer’s patches). It is low in the spleen. Serum con-centration of IgE is low compared to other immunoglobulins, andserum half-life is short (Table 12-4). Once produced, IgE binds tolocal tissue mast cells before entering the circulation, where it bindsto circulating mast cells, basophils, and tissue mast cells at distantsites. Once an individual is sensitized, reexposure to the antigenresults in degranulation of the mast cells with the release of pre-formed mediators and cytokines typical of Th2 cells. Synthesis ofleukotrienes and thromboxanes is also induced. These mediators
Figure 12-16. Schematic diagram of chemical interaction leading to hypersensitivity reactions or autoim-munity.
2996R_ch12_419-470 4/27/01 10:02 AM Page 451
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

452 UNIT 4 TARGET ORGAN TOXICITY
promote vasodilation, bronchial constriction, and inflammation.Clinical manifestations can vary from urticarial skin reactions(wheals and flares) to signs of hay fever, including rhinitis and con-juctivitis, to more serious diseases, such as asthma and potentiallylife-threatening anaphylaxis. These responses may begin with-in minutes of reexposure to the offending antigen; therefore, typeI hypersensitivity is often referred to as immediate hypersensitiv-ity.
Type II (Antibody-Dependent Cytotoxic Hypersensitivity)Type II hypersensitivity is IgG-mediated. Figure 12-19 shows themechanisms of action of a complement-independent cytotoxic re-action and complement-dependent lysis. Tissue damage may resultfrom the direct action of cytotoxic cells—such as macrophages,neutrophils, or eosinophils—linked to immunoglobulin-coated tar-get cells through the Fc receptor on the antibody or by antibodyactivation of the classic complement pathway. Complement acti-vation may result in C3b or C3d binding to the target cell surface.This acts as a recognition site for effector cells. Alternatively, theC5b-9 membrane attack complex may be bound to the target cellsurface, resulting in cell lysis (Fig. 12-3).
Type III (Immune Complex – Mediated Hypersensitivity)Type III hypersensitivity reactions also involve IgG immunoglob-ulins. The distinguishing feature of type III is that, unlike type II,in which immunoglobulin production is against specific tissue-associated antigen, immunoglobulin production is against solubleantigen in the serum (Fig. 12-20). This allows for the formation ofcirculating immune complexes composed of a lattice of antigenand immunoglobulin, which may result in widely distributed tis-sue damage in areas where immune complexes are deposited. Themost common location is the vascular endothelium in the lung,joints, and kidneys. The skin and circulatory systems may also beinvolved. Pathology results from the inflammatory response initi-ated by the activation of complement. Macrophages, neutrophils,and platelets attracted to the deposition site contribute to the tis-sue damage.
Type IV (Cell-Mediated Hypersensitivity) Type IV, ordelayed-type hypersensitivity (DTH) responses, can be divided intotwo classes: contact hypersensitivity and tuberculin-type hyper-sensitivity. Contact hypersensitivity is initiated by topical exposure,and the associated pathology is primarily epidermal. It is charac-
Figure 12-17. Schematic of classification of hypersensitivity reactions.
Figure 12-18. Schematic of type I hypersensitivity reaction. Figure 12-19. Schematic of type II hypersensitivity reactions.
2996R_ch12_419-470 4/27/01 10:02 AM Page 452
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

CHAPTER 12 TOXIC RESPONSES OF THE IMMUNE SYSTEM 453
terized clinically by an eczematous reaction at the site of allergencontact and, like type I through III responses, consists of twophases: sensitization and elicitation. However, in this case sensiti-zation is the result of the development of activated and memory Tcells as opposed to antibody production (Figs. 12-21 and 12-22).Sensitization occurs when the hapten penetrates the epidermis andforms a complex with a protein carrier. The hapten–carrier com-plex is processed by Langerhans-dendritic cells that migrate out ofthe epidermis to the local lymph nodes. There, the APC presentsthe processed antigen to CD4� T cells, leading to clonal expan-sion and the generation of memory T cells.
Upon second contact, Langerhans-dendritic cells present theprocessed hapten–carrier complex to memory T cells in either theskin or the lymph nodes. These activated T cells then secrete cy-tokines that bring about further proliferation of T cells and inducethe expression of adhesion molecules on the surface of ker-atinocytes and endothelial cells in the dermis. Both the expressionof adhesion molecules and the secretion of proinflammatory cy-tokines by T cells and keratinocytes facilitate the movement of in-flammatory cells into the skin, resulting in erythema and the for-
mation of papules and vesicles. CD8� cells may play a role in tis-sue damage. In cases where chemicals are lipid-soluble and cantherefore readily cross the cell membrane, they can modify intra-cellular proteins. These cells then present modified peptides ontheir cell surface in conjunction with MCH class I molecules.CD8� cells recognize these foreign peptides and cause tissue dam-age by either direct cytotoxic action or the secretion of cytokinesthat further promote the inflammatory response. The descriptiongiven above has been the accepted dogma for the mechanism ofdelayed hypersensitivity for years. This is currently an area of in-tense research; a developing hypothesis for the elicitation phase ofcontact hypersensitivity involves a greater role for a non-specificinflammatory signal, CD8� cells, and APCs other than Langerhanscells (Grabbe and Schwarz, 1998).
Tuberculin-type hypersensitivity is primarily a dermal reac-tion and begins following the intradermal injection of a specificantigen to which the individual has been previously exposed (suchas a microbial antigen). Within hours, a cellular infiltrate (prima-rily CD4� T cells) begins to appear. This infiltration continues asmacrophages and Langerhans-dendritic cells begin to migrate intothe area of injection. Circulation of immune cells to and from thelocal lymph nodes is thought to be like that in contact hypersensi-tivity. Also, like the contact hypersensitivity response, CD4� Tcells then secrete lymphokines that cause the expression of MHCclass II on the surfaces of macrophages and keratinocytes. The re-sult is activation of these cells, the release of proinflammatory me-diators, and the generation of an area of firm red swelling in thedermal tissue.
Separation of hypersensitivity responses into the types I to IVas in the classification of Coombs and Gell is helpful in under-standing mechanisms involved. It is important, however, to realizethat this is a simplification and that often pathology is the resultof a combination of these mechanisms. In addition, associated in-flammation may be a non-immune acute response and/or the re-sult of an immune-mediated event. For example, if one looks at thepathophysiology of respiratory disease induced by the acid anhy-
Figure 12-20. Schematic of type III hypersensitivity reaction.
Figure 12-21. Schematic of sensitization phase of type IV hypersensitivity reaction.
2996R_ch12_453 5/21/01 2:52 PM Page 453
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

454 UNIT 4 TARGET ORGAN TOXICITY
drides, a mixture of immune and nonimmune events is found to beimportant. Direct toxic effects of the chemicals may lead tobronchial epithelial damage, causing cells to release cytokines thatinduce a nonimmune inflammatory response. Damage to epithelialcells may also expose underlying lamina propia, allowing the chem-ical to exert direct effects on inflammatory cells and to stimulatesensory vagal afferents, leading to reflex bronchoconstriction andhyperresponsiveness. Along with these non-immune mechanisms,all four classes of immune-mediated hypersensitivity responses toacid anhydrides have been shown to occur (Bernstein and Bern-stein, 1994).Assessment of Hypersensitivity Responses One of the most im-portant and challenging problems in the field of immunotoxicol-ogy is determining the potential for chemicals to induce im-munomodulation and, in the current context, to promotehypersensitivity reactions. Thus, it becomes essential to have val-idated predictive animal models and to understand the underlyingmechanisms of action. The following is a review of the currentlyused methods of predicting types I and IV, the most frequently oc-curring hypersensitivity reactions to chemicals.
Assessment of Respiratory Hypersensitivity in ExperimentalAnimals Methods for detecting pulmonary hypersensitivity havebeen reviewed by Sarlo and Karol (1994) and can be divided intotwo types: (1) those for detecting immunologic sensitization and(2) those for detecting pulmonary sensitization. In some cases themethodologies may overlap. In the case of types I to III, im-munologic sensitization occurs when antigen-specific im-munoglobulin is produced in response to exposure to an antigenor, in the case of type IV, when a population of sensitized T lym-phocytes is produced. Pulmonary sensitization is determined by achange in respiratory function subsequent to the challenge of a sen-sitized animal or patient. In certain cases, immunologic sensitiza-tion may be confirmed by the detection of antigen-specific anti-body; however, subsequent challenge does not produce clinicalsigns of respiratory distress. It is also possible to detect pulmonarysensitization in animal models where there is no detectable anti-gen-specific antibody production. In these cases, cell-mediated orother mechanisms may be involved or there may be a difficulty inantibody detection.
Guinea pig models have been most frequently used for de-tection of pulmonary reactions to chemicals. In the guinea pig, asin the human, the lung is the major shock organ for anaphylacticresponse. Like humans, the guinea pig also demonstrates immedi-ate- and late-onset allergic reactions as well as bronchial hyperre-activity. The major difference in the mechanism of pulmonary re-sponses between humans and guinea pigs is that the antibodyinvolved in type I reactions in the former is IgE and in the latteris predominantly IgG1. Murine models are becoming more fre-quently utilized in the evaluation of respiratory hypersensitivity. Asin the human, IgE is a major anaphylactogenic antibody in themouse, and more murine immunologic reagents are available, al-lowing for more detailed mechanistic studies.
Methods utilized for respiratory exposure to chemicals are in-halation or either intranasal or intratracheal administration. Thereare advantages and disadvantages to each. Inhalation more closelyrepresents environmental exposure by allowing for chemical con-tact with the upper as well as the lower respiratory tract. However,the equipment required is expensive and difficult to maintain. Ex-posure via the intranasal route is easily accomplished and allowsfor distribution of antigen to the upper and lower respiratory tract;however, studies have shown that a large proportion of the mate-rial can be recovered from the stomach (Robinson et al., 1996). Incontrast, intratracheal instillation results in exposure to the lowerrespiratory tract only, and this procedure requires the use of anes-thesia.
Immunologic sensitization may be determined by obtainingsequential blood samples throughout the induction period andmeasuring antibody titer. Pulmonary sensitization is evaluated bydetecting the presence of pulmonary reactivity following challenge.This may be accomplished by visual inspection of the animals’ res-piratory pattern or more quantitatively by plethysmography. Withplethysmography, changes in the respiratory rate, tidal volume, andplethysmographic pressure can be measured.
Inhalation models are generally used for low-molecular-weight compounds, whereas intratracheal and intranasal models arefrequently used with high-molecular-weight compounds. One ofthe drawbacks of low-molecular-weight models is that often thesecompounds must conjugate with body proteins to become anti-
Figure 12-22. Schematic of elicitation phase of type IV hypersensitivity reaction.
2996R_ch12_419-470 4/27/01 10:02 AM Page 454
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

CHAPTER 12 TOXIC RESPONSES OF THE IMMUNE SYSTEM 455
genic. Often, a challenge with the conjugated chemical is neces-sary to induce a pulmonary response. Adding this variable can makethe analysis of test results more difficult. False negative results mayoccur due to variability in test article conjugation. Chemical con-jugates are also necessary to measure immunologic response.
Assessment of IgE-Mediated Hypersensitivity Responses inHumans Described below are methods of human type I hyper-sensitivity testing. These test results, in conjunction with a relevanthistory and physical exam, can be diagnostic of IgE-mediated pul-monary disease. Two skin tests are available for immediate hyper-sensitivity testing. In both, the measured endpoint is a “wheal andflare” reaction—is the result of edema and erythema subsequentto the release of preformed mediators. The prick-puncture test in-troduces very small amounts of antigen under the skin and, owingto the reduced chance of systemic reaction, is recommended as ascreening test. For test compounds not eliciting a reaction in theless sensitive test, the intradermal test using dilute concentrationsof antigen may be used, but there is a higher risk of systemicreactions. For a more detailed description of testing methods seeDemoly et al. 1998.
In vitro serologic tests, enzyme-linked immunosorbent assays(ELISAs), and radioallergosorbent tests (RASTs) may also be usedto detect the presence of antigen-specific antibody in the patient’sserum. These tests do not pose a risk of adverse reactions and maybe used in situations where standardized reagents for skin testingare not available. Serologic testing is often used in population-based epidemiologic studies.
Bronchial provocation tests may be performed by having thepatient inhale an antigen into the bronchial tree and evaluating hisor her pulmonary response. In some cases this may be the onlyway to demonstrate that a test article is capable of producing anasthmatic response. Care must be taken in these test situations inthat it is possible to produce severe asthmatic reactions or ana-phylaxis in sensitized individuals.
Assessment of Contact Hypersensitivity in Experimental Ani-mals Classically, the potential for a chemical to produce contacthypersensitivity has been assessed by the use of guinea pig mod-els. These tests vary in their method of application of the test ar-ticle, in the dosing schedule, and in the utilization of adjuvants.For a description of methods employed in representative tests, seeKlecak (1987). The two most commonly utilized guinea pig mod-els, the Büehler test (Büehler, 1965) and the guinea pig maxi-mization test (Magnusson and Kligman, 1969), are describedbriefly below. In the Büehler test, the test article is applied to theshaven flank and covered with an occlusive bandage for 6 h. Thisprocedure is repeated on days 7 and 14. On day 28, a challengedose of the test article is applied to a shaven area on the oppositeflank and covered with an occlusive dressing for 24 h. At 24 and48 h after the patch is removed, test animals are compared withvehicle-treated controls for signs of edema and erythema. Theguinea pig maximization test differs in that the test article is ad-ministered by intradermal injection, an adjuvant is employed, andirritating concentrations are used. Animals are given pairs ofintradermal injections at a shaven area on the shoulders. One pairof injections contains adjuvant alone, one pair contains test articlealone, and one pair contains the test article mixed with adjuvant.Seven days following injection, after the area is reshaven, the testarticle is applied topically and an occluded patch is applied for 48h. In cases where the test article at the given concentration is non-irritating, the area is pretreated with 10% sodium lauryl sulfate 24h before the patch is applied to produce a mild inflammatory re-
sponse. Two weeks following topical application, the animals arechallenged on the shaven flank with a non-irritating concentrationof the test article, which remains under an occluded patch for 24h. Then, after 24 and 48 h, the test site is examined for signs oferythema and edema. The endpoints for evaluation in the guineapig assays are subjective and it is difficult to assess irritating orcolored compounds using these models.
Over the past 15 years, efforts have been made to develop andvalidate more quantitative and immunologically based assay meth-ods in other species, focusing mainly on the mouse and in vitrosystems. Gad and coworkers (1986) developed the mouse ear-swelling test, which uses a quantitative measurement of ear thick-ness as an endpoint. Animals are sensitized by topical applicationof the test article for 4 consecutive days to abdominal skin that hasbeen prepared by intradermal injection of adjuvant and tape strip-ping. On day 10, the animals are challenged by topical applicationof the test article to one ear and vehicle to the contralateral ear.Measurements are made of ear thickness 24 and 48 h later. A pos-itive response is considered anything above a 20 percent increasein thickness of the treated ear over the control ear. Thorne and col-leagues (1991) showed that dietary supplementation with vitaminA enhanced the mouse ear-swelling assay in the absence of adju-vants, injections, or occlusive patches.
The assays described above evaluate the elicitation phase ofthe response in previously sensitized animals. The mouse locallymph node assay has recently undergone peer review coordinatedby the Interagency Coordinating Committee on the Validation ofAlternative Methods and has been accepted by government agen-cies as a stand-alone alternative to the guinea pig assays for use inhazard identification of chemical sensitizers. In this assay, the in-duction phase of contact sensitization is measured by the incorpo-ration of 3H-thymidine into proliferating lymphocytes in lymphnodes draining the site where the test article has been applied.Animals are dosed by topical application of the test article to theears for 3 consecutive days. The animals are rested for 2 days andthen injected intravenously with 20 �Ci of 3H-thymidine. Fivehours later, animals are sacrificed, the draining lymph nodes aredissected out, and single-cell suspensions are made and radioas-sayed. With consideration of dose response and statistical signifi-cance, a threefold increase in 3H-thymidine counts in chemicallyexposed animals over vehicle control animals is considered to bea positive response. This assay offers several advantages over theguinea pig assays in that (1) has the potential to reduce the num-ber of animals required and reduces animal distress; (2) it providesquantitative data that allow for statistical analysis; (3) and it pro-vides dose–response data. Additionally, the assay evaluates the in-duction phase of the immune response, making it more applicableto mechanistic studies (NIH publication no. 99-44940). As an ex-ample, some compounds capable of producing contact sensitiza-tion also induce IgE production and subsequent respiratory hyper-sensitivity. Using three known allergenic diisocyanates—diphenylmethane-4,4�,-diisocyanate (MDI); dicyclohexylmethane-4,4�-diisocyanate (HMDI); and isophorone diisocyanate (IPDI)—Dearman and coworkers (1992) showed that all three known con-tact sensitizers induced lymphocyte proliferation in the draininglymph node but that only MDI, a known respiratory sensitizer, in-duced elevated levels of serum IgE and IgG2b. Attempts have beenmade to correlate cytokine levels produced by draining lymph nodecells with contact and respiratory sensitizing potential.
Antigens, once processed, are presented on the surface of theAPC in conjunction with the MHC II antigen. Activation of either
2996R_ch12_419-470 4/27/01 10:02 AM Page 455
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

456 UNIT 4 TARGET ORGAN TOXICITY
Th1 or Th2 cells stimulates the production of cytokines, which areinstrumental in driving the system toward immunoglobulin pro-duction or the activation and proliferation of sensitized T cells(delayed-type hypersensitivity). The specific cytokines involved areshown in Fig. 12-23. IL-2, TNF-�, and IFN-� are produced by Th1cells and lead to the development of delayed-type hypersensitivity,whereas IL-4, IL-5, IL-6, IL-10, and IL-13 are produced by Th2cells and lead to the production of IgE (Mosmann et al., 1991).IFN-� not only promotes the induction of delayed hypersensitiv-ity but also appears to have an inhibitory effect on IgE production(Mosmann and Coffman, 1989). Likewise, IL-10, which promotesIgE production, inhibits the delayed-type hypersensitivity response(Enk et al., 1993; Schwarz et al., 1994). Dearman and Kimber(1999) have demonstrated an elevation in INF-� but little IL-4 orIL-10 production in draining lymph nodes of animals exposed tocontact allergens and, conversely, elevated production of IL-4 orIL-10 and little INF-� following exposure to chemical respiratoryallergens. Measurement of cytokine levels may prove to be an im-portant predictive tool for assessing the potential of chemicals toelicit hypersensitivity reactions.
Assessment of Contact Hypersensitivity in Humans Humantesting for contact hypersensitivity reactions is by skin patch test-ing. Patch testing allows for the diagnostic production of acute le-sions of contact hypersensitivity by the application of a suspectedallergen to the skin. Patches containing specified concentrations ofthe allergen in the appropriate vehicle are applied under an occlu-sive patch for 48 h in most test protocols. Once the patch is re-moved and enough time elapses for the signs of mechanical irrita-tion to resolve—approximately 30 min—the area is read for signsof erythema, papules, vesicles, and edema. Generally, the test isread again at 72 h and in some cases signs may not appear for upto 1 week or more. For more detailed information on patch test-ing, the reader is referred to Mydlarski et al. (1998).
Human repeat insult patch tests (HRIPT) are available as pre-dictive tests in humans. Like predictive testing in animal models,there are many variations in attempts to increase the sensitivity ofthese procedures. These include preparation of the induction siteby either stripping, the application of an irritating concentration ofsodium lauryl sulfate, or use of high concentrations of the test ar-ticle for induction of sensitization. In general, the application ofmultiple occlusive patches, up to ten for 48 h each at the same site,is followed by a rest period and then challenge under an occlusivepatch at a different site. Positive reactions are scored in the samemanner as for diagnostic patch tests.Hypersensitivity Reactions to Xenobiotics PolyisocyanatesPolyisocyanates have a widespread use in industry and are re-sponsible for more cases of occupationally related lung diseasethan any other class of low-molecular-weight (LMW) compounds.These chemicals are used in the production of adhesives, paint
hardeners, elastomers, and coatings. Occupational exposure is byinhalation and skin contact. Members of the group are known toinduce the full spectrum of hypersensitivity responses, types I toIV, as well as nonimmune inflammatory and neuroreflex reactionsin the lung (Bernstein and Bernstein, 1994; Grammer, 1985). Sen-sitized individuals have shown cross-reactivity between com-pounds in this group.
Toluene diisocyanate (TDI) is among the most widely usedand most studied members of this group. Pulmonary sensitizationto this compound can occur through either topical or inhalation ex-posure. It is a highly reactive compound that readily conjugateswith endogenous protein. Laminin, a 70,000-kDa protein, has beenidentified as the protein that TDI conjugates in the airways. Stud-ies with guinea pigs have confirmed the need for a threshold levelof exposure to be reached in order to obtain pulmonary sensitiza-tion. This finding supports the human data in which pulmonarysensitization is frequently the result of exposure to a spill, whereasworkers exposed to low levels of vapors for long periods of timefail to develop pulmonary sensitization. Unlike the case in manyhypersensitivity reactions, where removal of the antigen allevi-ates the symptoms of disease, symptoms may persist for as longas years after cessation of exposure in many TDI-induced asthmapatients.
Acid Anhydrides The acid anhydrides make up anothergroup of compounds for which nonimmune and IgE, cytotoxic, im-mune complex, and cell-mediated reactions have been reported(Bernstein and Bernstein, 1994; Grammer, 1985). These reactiveorganic compounds are used in the manufacturing of paints, var-nishes, coating materials, adhesives, and casting and sealing ma-terials. Trimellitic acid anhydride (TMA) is one of the most widelyused compounds in this group. Inhaled TMA fumes may conjugatewith serum albumin or erythrocytes leading, to type I (TMA-asthma), type II (pulmonary disease–anemia), or type III (hyper-sensitivity pneumonitis) hypersensitivity reactions upon subse-quent exposure. Topical exposure to TMA may lead to type IVhypersensitivity reactions, resulting in contact dermatitis. Also, re-exposure by inhalation may lead to a cell-mediated immune re-sponse in the lung, which plays a role in the pathology seen in con-junction with type II and III pulmonary disease. Human and animaltesting has supported the clinical findings in TMA-exposed work-ers. Levels of serum IgE can be measured in exposed workers andare predictive of the occurrence of type I pulmonary reactions.Serum titers of IgA, IgG, and IgM have been detected in patientswith high levels of exposure to TMA. Similar findings have beenreported in studies with rhesus monkeys, in which exposed ani-mals showed IgA, IgG, and IgM titers to TMA-haptenized ery-throcytes. Inhalation studies with rats have produced a model cor-responding to human TMA-induced pulmonary pneumonitis. Otheranhydrides known to induce immune-mediated pulmonary disease
Figure 12-23. Schematic of cytokines involved in hypersensitivity reactions.
2996R_ch12_419-470 4/27/01 10:02 AM Page 456
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

CHAPTER 12 TOXIC RESPONSES OF THE IMMUNE SYSTEM 457
include phthalic anhydride, himic anhydride, and hexahydroph-thalic anhydride.
Metals Metals and metallic substances, including metallicsalts, are responsible for producing contact and pulmonary hyper-sensitivity reactions. Metallic salts have been implicated in nu-merous immunologic and nonimmunologic pulmonary diseases.Exposure to these compounds may occur via inhalation or due totheir solubility in aqueous media (they can be dissociated and trans-ported into the lungs, where damage due to sensitization ornonimmunologic events takes place). Platinum, nickel, chromium,and cobalt are the most commonly implicated salts. For details thereader is referred to the reviews by Bernstein and Bernstein (1994),Menné (1987), and Marzulli and Maibach (1987).
PLATINUM Chlorplatinate salts are highly allergenic in com-parison to other metalic salts. Exposure may occur in the miningand metallurgic industries, in chemical industries where platinumis used as a catalyst, and in the production of catalytic converters.Exposed workers are at risk of developing allergic rhinitis andasthma secondary to IgE production. Sensitized workers show pos-itive skin tests and antigen-specific IgE by RAST testing.
COBALT Five percent of workers exposed to cobalt developoccupational asthma. These patients exhibit antigen-specific IgE,and their lymphocytes proliferate in response to free cobalt orcobalt conjugated to human serum albumin.
NICKEL Although nickel is a common contact sensitizer, pul-monary hypersensitivity reactions to nickel salts are rare. Occupa-tional exposure to nickel is most common in the mining, milling,smelting, and refinishing industries. When they occur, pulmonaryreactions are most frequently due to a direct toxic effect on theCNS and lung tissues. In addition to industrial contact, exposureoccurs in the form of jewelry, coins, and fasteners on clothing,making nickel one of the most frequently contacted sensitizers forthe general population. Studies have shown nickel sulfate to be themost frequent sensitizer when standard tray sensitizers are used,with positive results being between 6.7 and 11 percent. Nickel ap-pears to require a long sensitization period, and studies have shownthat patients may lose their hypersensitivity to nickel after long pe-riods of avoidance.
CHROMIUM Chromium is another metal often associated withdermatoses and less frequently with respiratory disease. Occupa-tional exposure to chromium is most frequent in industries involvedin electroplating processes, leather tanning, and paint, cement, andpaper pulp production. Chromium eczema (type IV hypersensitiv-ity) is among the most common occupationally associated skin dis-eases. Predictive tests on normal human subjects have shown sen-sitization rates to chromium sulfate as high as 48 percent.Occupational asthma from chromium exposure is less well docu-mented, and skin-prick tests have been negative. Evidence of IgE-mediated disease has been supported by immediate bronchial hy-perreactivity after challenge and the identification ofantigen-specific IgE antibodies. Cell-mediated (type IV) reactionshave been postulated to play a role, since late asthmatic reactionsfollowing bronchial challenge have been seen.
BERYLLIUM Beryllium is a metal capable of producing bothcontact and tuberculin type IV hypersensitivity reactions. The roleof CMI in beryllium-induced disease has been reviewed byNewman (1994). Beryllium exposure occurs most frequently in theaerospace industry, in high-technology ceramics and dental alloymanufacturing, and in the electronics, nuclear weapons, and nu-clear reactors industries. A major source of exposure was in theproduction of fluorescent light bulbs until the discontinuance of its
use for this purpose. Skin contact has been found to produce le-sions of contact hypersensitivity, whereas lesions produced by pen-etration of splinters of beryllium under the skin are granulomatousin nature. Inhalation of beryllium can result in disease ranging fromacute pneumonitis, tracheobronchitis, and chronic beryllium dis-ease to an increase in the risk of lung cancer. Environmentally in-duced berylliosis was evidenced by the incidence of disease in non-factory workers in communities around beryllium extraction plants.Exposure resulted from emissions from plants and contact withberyllium-contaminated family members’ clothing.
In cases of chronic beryllium disease, there is often a latentperiod of up to 10 years following first exposure. Lung pathologyconsists of multiple granulomas with mononuclear cell infiltrates—primarily macrophages, lymphocytes, and plasma cells—and fibrosis. Although lesions are usually localized in the lungs and associated lymph nodes, granulomatous involvement of other or-gans has been seen. As pulmonary disease progresses, effects onpulmonary circulation may lead to right-sided heart failure. Deathdue to berylliosis may then be due to respiratory or cardiac failure.
Unlike most hypersensitivity reactions in which removal fromexposure to the offending agent usually abates the disease, removalfrom beryllium exposure does little to alter the course of the dis-ease. Although the majority of beryllium is eliminated from thelung soon after inhalation, small amounts of retained beryllium aresufficient to induce and sustain the ongoing cellular immune re-sponse. Years after the last exposure, mass absorption data haveshown beryllium to be present in lung granulomas.
Owing to the similarities in clinical symptoms and pathologybetween berylliosis and other granulomatous lung diseases, im-munologic testing is important in definitive diagnosis. Patients withberyllium disease tested positive to patch testing with berylliumsalt and often showed granulomatous lesions at the patch-test sitewithin 3 weeks. However, these test procedures proved to be unsafe.Patch tests were found to induce sensitization in some patients andoften caused exacerbation of lung disease. The beryllium-specificlymphocyte proliferation test (BeLT) has been utilized to detectberyllium sensitization. This test has proven to be a more sensitiveindicator of early disease than patient history, physical exam, chestradiographs, or lung function test. Although this disease is not cur-able, progression of the disease process can be slowed by corti-costeriod therapy. BeLT allows for earlier detection of sensitiza-tion. This results in improved patient monitoring and permits earlierinstitution of treatment. In industry, BeLT provides a means of de-tecting jobs with a high risk of exposure.
Drugs Hypersensitivity responses to drugs are among themajor types of unpredictable drug reactions, accounting for up to10 percent of all adverse effects. Drugs are designed to be reactivein the body and multiple treatments are common. This type of ex-posure is conducive to producing an immunologic reaction. Im-munologic mechanisms of hypersensitivity reactions to drugs in-clude types I through IV. For a detailed review of drug allergy seeDeSwarte (1985). Penicillin is the most common agent involved indrug allergy and is discussed here as an example. Exposure to peni-cillin is responsible for 75 percent of the deaths due to anaphylaxisin the United States. The route of administration, dosage, and lengthof treatment all appear to play a role in the type and severity ofhypersensitivity reaction elicited. Severe reactions are less likelyfollowing oral administration as compared to parenteral, and pro-longed treatment with high doses increases the risk of acute inter-stitial nephritis and immune hemolytic anemia. The high incidence
2996R_ch12_419-470 4/27/01 10:02 AM Page 457
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

458 UNIT 4 TARGET ORGAN TOXICITY
of allergic reaction to penicillin is in part due to widespread ex-posure to the compound. Not only has there been indiscriminantuse of the drug, but exposure occurs through food products in-cluding milk from treated animals and the use of penicillin as anantimicrobial in the production of vaccines. Efforts have been madeto reduce unnecessary exposure.
Reactions to penicillin are varied and may include any of thefour types of hypersensitivity reactions. The most commonly seenclinical manifestation of type I reactions is urticaria; however, ana-phylactic reactions occur in about 10 to 40 of every 100,000 pa-tients receiving injections. Clinical signs of rhinitis and asthma aremuch less frequently observed. Blood dyscrasias can occur due tothe production of IgG against penicillin metabolites bound to thesurface of red blood cells (type II reaction). Penicillin has also beenimplicated in type III reactions leading to serum-sickness-likesymptoms. Owing to the high frequency of type IV reactions whenpenicillin is applied topically, especially to inflamed or abradedskin, products are no longer available for topical application. TypeIV reactions generally result in an eczematous skin reaction, but—infrequently—a life-threatening form of dermal necrosis may re-sult. In these cases there is severe erythema and a separation of theepidermis at the basal layer. This reaction, which gives the clini-cal appearance of severe scalding, is thought to be a severe delayedreaction.
Pesticides Pesticides have been implicated as causal agentsin both contact and immediate hypersensitivity reactions. Defini-tive diagnosis is often difficult or lacking in reported cases and an-imal and human predictive data often do not correlate well. Pesti-cide hypersensitivity responses have been reviewed by Thomas andcoworkers (1990) and are described briefly below.
One of the difficulties in obtaining good epidemiologic datato document reactions to pesticides is the nature of exposure. Agri-cultural workers are among those most commonly exposed, andthe fact that workers are exposed to multiple chemicals as well asharsh environmental factors makes diagnosis difficult. Further-more, diagnostic follow-up among this group is infrequent.
In the case of barban, a carbamate insecticide, the reportedincidence of contact sensitivity due to exposure is rare; however,predictive testing with the guinea pig maximization test (GPMT)and the diagnostic human patch test indicates that this pesticide isa potent sensitizer. Likewise, malathion, captan, benomyl, maneb,and naled have been identified as strong to extreme sensitizersusing the GPMT. Human predictive data, diagnostic patch testing,and the reported incidence in the literature of toxicity with the useof these compounds are often not in agreement with the animaldata.
Pesticides have been implicated in cases of immediate hy-persensitivity, including rhinitis, conjunctivitis, asthma, and ana-phylaxis. However, there has been no definitive proof of the asso-ciation. It is possible that observed reactions are of an irritant naturerather than being an immunologic response. It has been shown thatthe asthmatic response to organophosphate insecticides is not dueto the acetylcholinesterase activity, since the administration of at-ropine, a cholinergic antagonist, failed to block the response. An-imal studies show some evidence to support the role of immediatehypersensitivity responses to pesticides. Mice injected intraperi-toneally with malathion or 2,4-dichlorophenoxyacetic acid conju-gates developed IgE antibody. However, antibody was not detectedto these chemicals when they were applied topically. More epi-demiologic and mechanistic studies are needed in the area of pes-ticide hypersensitivity to further define these relationships.
Others NATURAL RUBBER LATEX PRODUCTS Allergic reac-tions to natural rubber latex products have become an importantoccupational health concern over the past decade. Natural rubberlatex is derived from the rubber tree Hevea brasiliensis and is usedin the manufacture of over 40,000 products including examinationand surgical gloves, among other medical products. Dermatologicreactions to latex include irritant dermatitis due to chemical addi-tives or mechanical abrasion and the occlusive conditions causedby wearing gloves; contact dermatitis (which represents approxi-mately 80 percent of the allergic responses) due to the chemicaladditives used in the glove manufacturing (e.g., thiurams, carba-mates, mercapto compounds, and phenylenediamines), and poten-tially more serious IgE-mediated responses due to residual latexproteins that remained in the finished products. The IgE responsesmay manifest as urticaria, asthma, or life-threatening anaphylaxis.For a review of latex allergy, see Germolec (1999).
COSMETICS AND PERSONAL HYGIENE PRODUCTS Contact der-matitis and dermatoconjuctivitis may result from exposure to manycosmetic and personal hygiene products, including makeup,deodorants, hair sprays, hair dyes and permanent-waving solutions,nail polish, soaps, face creams, and shampoos (Liberman et al.,1985). These agents contain coloring agents, lanolin, paraffin,petrolatum, vehicles, perfumes, and antimicrobials such as parabenesters, sorbic acid, phenolics, organic mercurials, quaternary am-monium compounds, EDTA, and formaldehyde. The devices usedto apply these products may also induce allergic reactions. Diag-nosis may be accomplished by patch testing; however, in patch test-ing, it is often necessary to employ products used by the patient inaddition to those on a standard test tray. In cases of dermatocon-junctivitis, false-negative testing may occur. The skin of the eye-lids may be more sensitive to agents than that of the forearm orback, making patch testing unreliable. Elimination-provocation procedures may be helpful in the diagnosis of difficultcases. All suspect offending agents must be removed from the pa-tient’s environment. Once clinical signs have resolved, agents maybe reintroduced one at a time while the patient is observed for therecurrence of signs.
ENZYMES Enzymes are another group capable of elicitingtype I hypersensitivity responses (Gutman, 1985). Subtilin, a pro-teolytic enzyme derived from Bacillus subtilis, is used in laundrydetergents to enhance their cleaning ability. Both individuals work-ing in the environment where the product is made and those usingthe product may become sensitized. Subsequent exposure may pro-duce signs of rhinitis, conjunctivitis, and asthma. An alveolarhypersensitivity reaction associated with precipitation antibodiesand an Arthus-type reaction from skin testing has also been seen.Papain is another enzyme known to induce IgE-mediated disease.It is a high-molecular-weight sulfhydryl protease obtained from thefruit of the papaya tree and most commonly used as a meat ten-derizer and a clearing agent in the production of beer. However, itis also used in the production of tooth powders, laxatives, andcontact-lens cleaning solutions.
FORMALDEHYDE Formaldehyde was discussed above as oneof the components of cosmetics capable of causing a contact hy-persensitivity reaction. Formaldehyde exposure also occurs in thetextile industry, where it is used to improve wrinkle resistance, andin the furniture, auto upholstery, and resins industries. The generalpublic may be exposed to low levels of formaldehyde in productsas ubiquitous as newspaper dyes and photographic films and pa-per. This low-molecular-weight compound is extremely soluble inwater and haptenates human proteins easily (Maibach, 1983). Hu-
2996R_ch12_419-470 4/27/01 10:02 AM Page 458
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

CHAPTER 12 TOXIC RESPONSES OF THE IMMUNE SYSTEM 459
man predictive testing with 1 to 10% formalin (formalin is 37%formaldehyde) for induction and 1% formalin for challenge showedsensitization rates of 4.5 to 7.8 percent (Marzulli and Maibach,1974). Occupational exposure to formaldehyde has been associ-ated with the occurrence of asthma, although it has been difficultto demonstrate antibodies to formaldehyde in the affected individ-uals (Hendrick et al., 1982).
For further information and a listing of chemicals known tocause hypersensitivity reactions affecting the respiratory systemand skin, see Chaps. 15 and 19.
Autoimmunity In the section on hypersensitivity presentedabove, we discussed two mechanisms, types II and III, by whichhost tissues are damaged by the host’s own immune system, cre-ating autoimmune-like disease. In these situations, unaltered selfantigens are not the target of the immune mechanisms but damageoccurs to cells bearing hapten on membranes or to innocent by-stander cells in close proximity to antigen-antibody complexes. Forexample, damage produced in autoimmune Goodpasture’s diseaseis similar to that seen in type III hypersensitivity reactions in thelung due to TMA. Although the resulting pathology may be thesame for autoimmune reactions and hypersensitivity, mechanismsof true autoimmune disease are distinguished from hypersensitiv-ity. In cases of autoimmunity, self antigens are the target, and inthe case of chemical-induced autoimmunity, is the disease state isinduced by a modification of host tissues or immune cells by thechemical and not the chemical acting as an antigen/hapten.Mechanisms of Autoimmunity The immunopathogenesis of au-toimmune disease has been reviewed by Rose (1994) and is de-scribed briefly below as background information for understand-ing how chemicals may induce autoimmunity. Three types ofmolecules are involved in the process of self-recognition: im-munoglobulins (Igs), T-cell receptors (TCRs), and the products ofmajor histocompatibility complex (MHC). Igs and TCRs are ex-pressed clonally on B and T cells, respectively, whereas MHC mol-ecules are present on all nucleated cells. The ability of lympho-cytes to distinguish one molecule from another stems from theantigen binding a specific lymphocyte receptor. In B-cell lines,through rearrangement of heavy and light Ig chains, tremendousdiversity (106 to 107 specificities), occurs among Ig recognitionstructures. Likewise, a similar number of specific TCRs are pro-duced as the result of gene rearrangement in T cells induced bypeptide hormones produced by thymic epithelial cells. Two majortypes of B and T cells are produced. B cells expressing CD5 pre-dominate in embryonic life and are later found mostly in the in-testinal mucosa. These cells produce high levels of IgM, and muchof it is autoantibody. Although most B cells do not express CD5prior to class switching, they do express high levels of IgM. In-fluenced by cytokines produced by interacting T cells followingantigen stimulation, these cells produce primarily IgG, IgA, or IgE.Similarly, T cells develop from one of two lineages—those with��TCRs and those with ��TCRs. Although most mature T cellsexpress ��TCRs, ��TCRs are predominant on mucosal surfaces.As described earlier in the chapter, ��TCRs continue differentia-tion into CD4� or CD8� T cells. CD4� cells have primarily helperand inducer functions and recognize antigens in the context ofMHC class II molecules. CD8� T lymphocytes are mainly cyto-toxic cells and recognize antigenic determinants in conjunctionwith MHC class I molecules.
The process of negative selection against autoreactive T cellsin the thymus is important in the prevention of autoimmune dis-
ease. T cells expressing ��TCRs that fit self MHC molecules withhigh affinity undergo apoptosis (programmed cell death) at an ac-celerated rate, whereas those with a low affinity for self antigenand a high affinity for foreign antigen undergo positive selectionand proliferate in the thymus, eventually migrating to the periph-eral lymphatics. Although negative selection greatly reduces thenumbers of self-reactive T cells, some of these cells do leave thethymus and remain in circulation in a state of anergy. These cellsare able to bind their designated antigen but do not undergo pro-liferation owing to a lack of necessary second signal. This secondsignal is generally provided by an APC in the form of a cytokine,IL-2, or a cell surface receptor that interacts with the T cell.
Reactive CD4� T cells recognize only processed antigen pre-sented by APCs—generally macrophages, B cells, or dendriticcells—in conjunction with MHC class II molecules. These APCstake up exogenous antigens, cleave them with proteolytic enzymes,and express them on their cell surfaces. In contrast, intracellularantigens are processed and presented on cell surfaces in conjunc-tion with MHC class I molecules. These antigens may be the prod-ucts of malignant or normal cells or may result from infection withbacterial, viral, or other intracellular pathogens. The processedantigen–MHC class I complex is recognized by a specific TCR ona CD8� lymphocyte.
Several mechanisms are available that may break down self-tolerance, leading to autoimmunity. The first is exposure to anti-gens not available in the thymus during embryonic development.Therefore, the antigen-specific T cell–reactive lymphocytes notsubjected to negative selection could induce an autoimmune reac-tion. Examples include myelin and organ-specific antigens such asthyroglobulin. Breakdown of self-tolerance to these antigens maybe induced by exposure to adjuvants or to another antigenically re-lated protein. The second is the overcoming of T-cell anergy bychronic lymphocyte stimulation. Finally, there is interference withnormal immunoregulation by CD8� T cell suppressor cells, whichmay create an environment conducive to the development of au-toimmune disease.
Effector mechanisms involved in autoimmune disease can bethe same as those described earlier for types II and III hypersen-sitivity or, in the case of pathology associated with solid tissues,including organs, they may involve CD8� cytotoxic T cells. Tis-sue damage associated with CD8� cells may be the result of di-rect cell-membrane damage and lysis induced by binding or the re-sults of cytokines produced and released by the T cell. TN-� hasthe ability to kill susceptible cells and IFN-� may increase the ex-pression of MHC class I on cell surfaces, making them more sus-ceptible to CD8� cells. Cytokines may also be chemotactic formacrophages, which can cause tissue damage directly or indirectlythrough the release of proinflammatory cytokines. As is the casewith hypersensitivity reactions, autoimmune disease is often theresult of more than one mechanism working simultaneously. There-fore, pathology may be the result of antibody-dependent cytotox-icity, complement-dependent antibody-mediated lysis, or direct orindirect effects of cytotoxic T cells.
Genetic and environmental factors appear to affect the sus-ceptibility of individuals to autoimmune disease. Familial predis-position to autoimmune disease has been found, as well as a sim-ilarity in MHC genetic traits among individuals involved. Certainchemicals and drugs are known to induce autoimmune disease ingenetically predisposed individuals, and examples of these are dis-cussed below. The role of environmental pollutants is uncertain;more study is needed in this area. One point of interest is that in
2996R_ch12_419-470 4/27/01 10:02 AM Page 459
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

460 UNIT 4 TARGET ORGAN TOXICITY
all known cases of drug-induced autoimmunity, the disease hasabated once the offending chemical was removed.Assessment of Autoimmune Responses Few models are avail-able for accessing the potential of a chemical to induce autoim-mune disease, and these have major limitations. The most com-monly used models include graft-versus-host disease (GVHD), thepopliteal lymph node (PLN) model in mice, and human lympho-cyte transformation assays. Although these models may have somepredictive value, at this point immunohistopathology is the onlydefinitive diagnostic tool.
There are numerous reports of chemicals that have been as-sociated with autoimmunity. These relationships may be causativethrough direct mechanisms or they may be indirect, acting as anadjuvant. They may also serve to exacerbate a preexisting autoim-mune state (reviewed by Kilburn and Warshaw, 1994; Coleman andSim, 1994). In the area of autoimmunity, exact mechanisms of ac-tion are not always known. Table 12-11 lists chemicals known tobe associated with autoimmunity, showing the proposed self-antigenic determinant or stating adjuvancy as the mechanism ofaction. A brief discussion of selected drug and nondrug chemicalsis provided.Autoimmune Reactions to Xenobiotics Methyldopa Methyl-dopa is a centrally acting sympatholytic drug that has been widelyused for the treatment of essential hypertension. With the adventof newer antihypertensive drugs, the use of methyldopa has de-clined. Platelets and erythrocytes are targeted by the immune sys-tem in individuals treated with this drug. In the case of thrombo-cytopenia, antibodies are detected against platelets, which isindicative of immune recognition of a self- or altered self-antigen.Hemolytic anemia occurs in at least 1 percent of individuals treatedwith methyldopa, and up to 30 percent of these individuals developantibodies to erythrocytes as manifest in a positive Coombs test.The antibodies are not directed against the chemical or a chemicalmembrane conjugate.
Hydralazine, Isoniazid, and Procainamide Hydralazine, iso-niazid, and procainamide produce autoimmunity, which is mani-fest as a sytemic lupus erythematosus (SLE)-like syndrome. Anti-bodies to DNA have been detected in individuals showing thissyndrome. Hydralazine is a direct-acting vasodilator drug used in
the treatment of hypertension. Isoniazid is an antimicrobial drugused in the treatment of tuberculosis. Procainamide is a drug thatselectively blocks Na� channels in myocardial membranes, mak-ing it useful in the treatment of cardiac arrhythmias. Studies withhydralazine and isoniazid indicate that the antigenic determinantis myeloperoxidase. Immunoglobulins are produced againstmyeloperoxidase in individuals treated with these drugs. DNA isthe apparent antigenic determinant for procainamide. For thesethree drugs, there is no evidence indicating that the immune sys-tem is recognizing the chemical or a chemical conjugate. In addi-tion, these drugs have also been shown to produce hypersensitiv-ity responses not associated with the SLE syndrome.
Halothane Halothane, one of the most widely studied of thedrugs inducing autoimmunity, is an inhalation anesthetic that caninduce autoimmune hepatitis. The incidence of this iatrogenic dis-ease in humans is about 1 in 20,000. The pathogenesis of the hep-atitis results from the chemical altering self (a specific liver pro-tein) to such a degree that the immune system recognizes the alteredself and antibodies are produced. Studies using rat microsomesshow that halothane has to be oxidized by cytochrome P450 en-zymes to trifluoroacetylhalide before it binds to the protein. In-vestigations indicate that in affected individuals antibodies to spe-cific microsomal proteins are produced.
Vinyl Chloride Vinyl chloride, which is used in the plasticsindustry as a refrigerant and in the synthesis of organic chemicals,is a known carcinogen and is also associated with a scleroderma-like syndrome. The disease affects multisystemic collagenous tis-sues, manifesting itself as pulmonary fibrosis, skin sclerosis, and/orfibrosis of the liver and spleen. Ward and coworkers (1976) re-ported on 320 exposed workers, showing that 58 (18 percent) hada scleroderma-like syndrome. The individuals who showed the dis-ease were in a group genetically similar (i.e., HLA-DR5) to pa-tients with classic idiopathic scleroderma patients. Although theexact mechanism whereby this chemical produces autoimmunityis unclear, it is presumed that vinyl chloride acts as an amino acidand is incorporated into protein. Because this would produce astructurally abnormal protein, which would be antigenic, an im-mune response would be directed against tissues with the modi-fied protein present.
Table 12-11Chemical Agents Known to be Associated with Autoimmunity
PROPOSED ANTIGENIC
CHEMICAL CLINICAL MANIFESTATIONS DEPARTMENT REFRENCE
DrugsMethyl dopa Hemolytic anemia Rhesus antigens Murphy and Kelton (1991)Hydralazine SLE-like syndrome Myeloperoxidase Cambridge et al. (1994)Isoniazid SLE-like syndrome Myeloperoxidase Jiang et al. (1994)Procainamide SLE-like syndrome DNA Totoritis et al. (1988)Halothane Autoimmune hepatitis Liver microsomal Kenna et al. (1987)
proteinsNon-drug chemicals
Vinyl Chloride Scleroderma-like Abnormal protein Ward et al. (1976)syndrome synthesized in liver
Mercury Glomerular neuropathy Glomerular basement Pelletier et al. (1994)membrane protein
Silica Scleroderma Most likely acts as Pernis and Paronetto (1962)an adjuvant
2996R_ch12_419-470 4/27/01 10:02 AM Page 460
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

CHAPTER 12 TOXIC RESPONSES OF THE IMMUNE SYSTEM 461
Mercury This widely used metal is now known to have sev-eral target systems, including CNS and renal system. Mercury alsohas two different actions with respect to the immune system. Thefirst action is direct injury, described previously. Mercury also pro-duces an autoimmune disease demonstrated as glomerularnephropathy. Antibodies produced to laminin are believed to be re-sponsible for damage to the basement membrane of the kidney.Mice and rats exposed to mercury also show antinuclear antibod-ies. The role of these antibodies in the autoimmune disease is notclear. However, they represent a known biomarker of autoimmu-nity. Studies in the brown Norway rat point to a mercury-inducedautoreactive CD4� cell as being responsible for the polyclonal an-tibody response. Mercury chloride induces an increase in the ex-pression of MHC class II molecules on B lymphocytes as well asshifting the T helper cell population along the Th2 line. It is theTh2 cell that promotes antibody production. The imbalance be-tween Th1 and Th2 cells is believed to be caused by the depletionof cysteine and the reduced form of gluthathione in Th1 cells. Thesechemical groups are known to be important in the synthesis of andresponsiveness to IL-2 in T cells. Thus, Th1 cells that synthesizeand respond to IL-2 would be at a greater risk than Th2 cells.
Mercury-induced autoimmunity has a strong genetic compo-nent. This has been extensively studied in the rat. Some strains ofrats, such as the Lewis rat, are completely resistant, while others,such as the brown Norway, are exquisitely sensitive. Susceptibil-ity appears to be linked to three or four genes, one of which is themajor histocompatibility complex. An excellent review of mercuryand autoimmunity is provided by Pelletier and coworkers. (1994).
Silica Crystalline silica (silicon dioxide) is a primary sourceof elemental silicon and is used commercially in large quantitiesas a constituent of building materials, ceramics, concretes, andglasses. Experimental animals as well as humans exposed to silicamay have perturbations in the immune system. Depending on thelength of exposure, dose, and route of administration of silica, itmay kill macrophages or may act as an immunostimulant. Silicahas been shown to be associated with an increase in sclerodermain silica-exposed workers (reviewed by Kilburn and Warshaw,1994). This effect is believed to be mediated via an adjuvant mech-anism. Adjuvancy as a mechanism of causing autoimmunity hasbeen implicated with a number of other chemicals, including paraf-fin and silicones. Inherent in adjuvancy as a mechanism of pro-ducing autoimmunity is that the population affected by these chem-icals must already be at risk for the autoimmune disease. This is
supported by the data indicating a genetic component to many au-toimmune diseases.
Table 12-12 shows chemicals that have been implicated in au-toimmune reactions, but in these cases the mechanism of autoim-munity has not been as clearly defined or confirmed. The list in-cludes both drug and nondrug chemicals. The heterogeneity ofthese structures and biological activities illustrate the breadth ofpotential for the induction of chemically mediated autoimmunedisease.Multiple Chemical Sensitivity Syndrome Multiple chemical sen-sitivity syndrome (MCS) has been associated with hypersensitiv-ity responses to chemicals. The disease associated with MCS ischaracterized by multiple subjective symptoms related to more thanone system. The more common symptoms are nasal congestion,headaches, lack of concentration, fatigue, and memory loss. Manymechanisms have been suggested to explain how chemicals causethese symptoms; however, there remains considerable controversyas to a cause–effect relationship. Clinical ecologists, the major pro-ponents of MCS, have focused on immunologic mechanisms to ex-plain the etiology. They hypothesize that MCS occurs when chem-ical exposure sensitizes certain individuals, and, upon subsequentexposure to exceedingly small amounts of these or unrelated chem-icals, the individual exhibits an adverse response. Controlled stud-ies on the immunologic status of individuals with MCS have shownno alterations in their immune system or any indication that MCSresults from impairment of the immunity, including inappropriateimmune response to chemicals. The search for a theoretical basisfor MCS is now being focused on the nervous system. Two untestedhypotheses have emerged. The first involves a nonspecific inflam-matory response to low-level irritants known as “neurogenic in-flammation.” The second involves induction of lasting changes inlimbic and neuronal activity (via kindling) that alter a broad spec-trum of behavioral and physiologic functions. The reader is referredto a review by Sikorski and colleagues (1995) for details and ref-erences concerning MCS.
NEW FRONTIERSAND CHALLENGES
Immunotoxicology has grown significantly since its beginnings asa recognized subdiscipline of toxicology in the late 1970s. Earlywork focused on the development of methods that were sensitiveand predictive and on an understanding of xenobiotic-induced im-
Table 12-12Chemicals Implicated in Autoimmunity
MANIFESTATION IMPLICATED CHEMICAL REFERENCE
Scleroderma Solvents (toluene, xylene) Walder (1983)Tryptophan Silver et al. (1990)Silicones Fock et al. (1984)
Systemic lupus Phenothiazines Canoso et al. (1990)erythrematosus Penicillamine Harpey et al. (1971)
Propylthiouracil DeSwarte (1985)Quinidine Jiang et al. (1994)L-dopa DeSwarte (1985)Lithium carbonate Ananth et al. (1989)Trichloroethylene Kilburn and Washaw (1992)Silicones Fock et al. (1984)
2996R_ch12_419-470 4/27/01 10:02 AM Page 461
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

462 UNIT 4 TARGET ORGAN TOXICITY
ing certain receptors, transcription factors, cytokines, etc., can beused for similar mechanistic studies. Severe combined immuno-deficient mice (SCID) can easily be engrafted with human or ratimmune cells (SCID/hu and SCID/ra) and have been used to studyimmune regulation, hematopoiesis, hypersensitivity, and autoim-munity. Uses of reconstituted SCID mice have been consideredelsewhere (IPCS, 1996), and their use in mechanistic studies is ob-vious. As with the use of molecular biological techniques, how-ever, there are several questions regarding the broader utility oftransgenics and SCID animals in immunotoxicology. Because oftheir altered biological status, how do they compare with standardanimal models with respect to time course of action, dose response,pharmacokinetics, and other factors of chemical toxicity? Are thehoming patterns of engrafted cells identical to those in the stan-dard animal (i.e., to the extent possible, how identical are the im-mune systems of these mice compared to standard mice)? Can theseanimal models be used to identify new or suspected immunotoxi-cants, and if so, are they as or more sensitive, predictive, and/orcost-effective as traditional animal models?
Developmental Immunotoxicology
The development (ontogeny) of the mammalian immune systemoccurs primarily in utero, though some postnatal development ormaturation does occur. Developmental immunotoxicology involvesinvestigation into the effects that xenobiotics have on the ontogenyof the immune system and includes prenatal (in utero), perinatal(36 h of age), and neonatal periods of exposure (reviewed in Bar-nett, 1996). Recent studies suggest that immune development inhumans and other species may be altered after perinatal exposureto immunotoxic chemicals, including chemotherapeutics, corticos-teroids, polycyclic hydrocarbons, and polyhalogenated hydrocar-bons (reviewed in Barnett, 1996, and in Holladay, 1999). It hasalso been suggested that these effects may be more dramatic orpersistent than those following exposure during adult life. With thepassage of the Food Quality Protection Act (FQPA), more atten-tion has focused on children’s health and thus on this emergingarea of immunotoxicology. Specifically, an understanding of thedevelopment and functioning of the juvenile immune system andsubsequent validated methods (similar to those currently validatedin adult animal models) are needed.
Systemic Hypersensitivity
An adverse immune response, in the form of systemic hypersensi-tivity, is among the most frequent causes for withdrawal of drugsthat have made it to the market; such a response can account forapproximately 15 percent of adverse reactions to xenobiotics (deWeck, 1983; Guzzie, 1995). These findings are generally unex-pected in that they were not predicted in preclinical toxicology andimmunotoxicology studies. A primary reason for this is the lack ofgood preclinical models for predicting systemic hypersensitivityresponses, particularly to orally administered chemicals. This con-cern applies not only to the pharmaceutical industry but also to thefood industry (proteins and genetically modified foods). It has beennoted that a positive guinea pig hypersensitivity test (any test) hasless than a 25 percent chance of predicting human clinical re-sponses with respect to systemic hypersensitivity of drugs (Weaveret al., 1999). Assays are needed that are more predictive of drug
munomodulation—in particular, immunosuppression. With un-precedented growth in the understanding of immune regulation(brought about in the 1980s as a result of the emergence of AIDS)came investigations into mechanisms of action (including cellularinteractions, modulation of transcription factors, and alterations ingene regulation) and in the use of in vitro methods to study thesemechanisms and to attempt to predict effects in the whole animal.While these types of investigations continued to grow, the 1990ssaw a surge in investigations into the understanding of immune-mediated disease: hypersensitivity and autoimmunity. As under-standing of the human genome grows, pharmaceutical companiesmove faster and deeper into biotechnology, public concern and sci-entific debate over genetically modified foods increases, concernfor the effects of chemicals on children’s health grows, concernand debate over the potential impact of chemically induced en-docrine disruption continues, the push to use kinetic and mecha-nistic data in human health risk assessments continues, and in-creased pressure to reduce the use of animals in testing increases,there is no doubt that immunotoxicology will continue to grow andexpand in many ways. The following are a few of the new fron-tiers and challenges facing the immunotoxicologist of today andtomorrow.
Molecular Biology Methods: Proteomicsand Genomics
The use of molecular biology methods in immunotoxicology in thepast has been primarily in the understanding of the mechanism ofaction of identified immunotoxicants. Much research is currentlyfocused on investigating induction of certain mRNAs (e.g., cy-tokines, transcription factors) in elucidating mechanism or as pos-sible biomarkers of exposure. Proteomics (the study of all ex-pressed proteins in a particular cell, and thus the functionalexpression of the genome) and genomics (the study of all genesencoded by an organism’s DNA), combined with bioinformatics,are making it possible to evaluate chemically induced alterationsin entire pathways and signaling networks. The utility of molecu-lar biology tools such as proteomics and genomics in eludicatingmechanism of action is obvious. However, can these powerful toolsbe used to identify new or suspected immunotoxicants? Is it pos-sible that common profiles of gene expression will emerge forclasses of structurally related or structurally dissimilar known orsuspected immunotoxicants? How is the profile or gene expressionrelated to administered dose, tissue dose, and time course of action of a chemical? Is human gene expression in response tochemical exposure the same as that in the animal or cell culture?If it is determined that these methods are useful tools in the iden-tification of possible immunotoxicants, how should they be vali-dated?
Animal Models: Transgenics and SCID
The developments in molecular biology have not only permittedthe evaluation of specific genes or arrays of genes but have alsoallowed for the manipulation of the embryonic genome, creatingtransgenic and knockout mice (reviewed in IPCS, 1996). As a con-sequence of transgenic technology, complex immune responses canbe dissected into their components. In this way, the mechanismsby which immunotoxicants act can be better understood. Mice lack-
2996R_ch12_419-470 4/27/01 10:02 AM Page 462
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

CHAPTER 12 TOXIC RESPONSES OF THE IMMUNE SYSTEM 463
antigenicity or hypersensitivity in humans. In the area of food al-lergy, again there are no validated animal models available, al-though the brown Norway rat and the Balb/c mouse have beenshown to have some promise (Kimber et al., 1999, 2000; Knippelset al., 1999, 2000).
Computational Toxicology
The possibility of ascertaining certain molecular properties of amolecule even before it is synthesized is a highly beneficial use ofcomputational chemistry. In fact, computational chemistry has beena rapidly growing and valued field, particularly in the fast-pacedand highly competitive pharmaceutical industry. As a result of theusefulness of this tool, interest has grown in the use of computa-tional toxicology methods to predict the potential biological/toxi-cologic activity of chemicals. The premise is that the structure ofa chemical determines the physiochemical properties and reactiv-ities that underlie its biological and toxicologic properties [quan-titative structure-activity relationships (QSAR)]. Being able to pre-dict potential adverse effects cannot only aid in the designeddevelopment of new chemicals but also has the potential to reducethe need for animal testing. It may ultimately or potentially leadto better health and environmental protection through the strategicapplication of limited testing resources and existing informationassets to help sort out or identify the most hazardous chemicals.Computational toxicology can also help in screening large num-bers of chemicals more efficiently for a variety of toxicologic end-points. Computational methods will not assure 95 to 100 percentaccuracy in the prediction of toxicity, and they will not eliminatethe need for testing, but these methods will help reduce and prior-itize the testing that is required. The use of computational toxi-cology cannot replace the need for toxicologists. These individu-als will still be needed for their understanding of the relationshipbetween the predicted endpoints and product or component ADMEprofiles (absorption, distribution, metabolism, and elimination) andthe true potential for human exposure as determined by the process,method of application, and end use of the product. Immunotoxi-cologists studying hypersensitivity have begun to investigatewhether computational methods can be used to screen for poten-tial chemical sensitizers (Karol, et al., 1996, 1999; Graham et al.,1997; Gerberick, 1999). Thus, computational toxicology is a toolthat may offer promise in the identification/prediction of chemicalsensitization. The use of computational methods to predict otheraspects of immunotoxicology has not been explored.
Biomarkers
True biomarkers indicate exposure to a specific chemical as wellas susceptibility to adverse effect; and/or are predictive of diseaseassociated with chemical exposure. They must be sensitive, spe-cific, relevant, reproducible, and measurable in the population. Themost sensitive (and desirable) biomarkers would be those that in-dicate exposure in the absence of immediate adverse effect. Bio-markers of effect would indicate subclinical effects of chemical ex-posure. Various avenues are currently being explored as potentialbiomarkers, including cytokine gene expression patterns, flow cy-tometry, and immunomodulation in alternative species such as fish,just to name a few. The International Programme on ChemicalSafety (IPCS, 1996) has concluded that, as with other systems in
toxicology, few biomarkers are available for immunotoxicity, par-ticularly for assessing immunotoxicity of individual susceptibility.More epidemiologic studies are needed to obtain a better view ofthe utility of biomarkers for detecting immunotoxic events and, ul-timately, possible health risks that may be associated with expo-sure to chemicals that modulate the immune system.
Risk Assessment
With the demonstration that (1) chemicals can perturb the immunesystem of animals, (2) perturbation of immune function is corre-lated with an increased risk of infectious disease, and (3) pertur-bations in immune function can occur in the absence of any clin-ically observable effect, attention has focused on the risk to thehuman population following exposure to chemicals that can alterimmune function in animals. However, these initial assessments ofthe use of immunotoxicology data in animals as predictors of riskfor human clinical effects have limitations (reviewed in Luster etal., 1994, and IPCS, 1996). These include the fact that though manywere good indicators, no single immune test has been observed tobe highly predictive of altered host resistance. Also, to demonstratechemical-related clinical immune effects in the human population,a significant number of individuals may be required. Variability inthe virulence of infectious agents in the human population, the com-plexity of the immune system, and the redundancy (“immune re-serve”; multiple components capable of responding to a foreignchallenge) in the immune system may all contribute to the diffi-culty in quantifying relationships between chemical-induced alter-ations in immune status and alterations in host resistance in hu-mans. Finally, there is the question of whether the relationshipbetween functional immune changes and susceptibility to diseasefollows a linear (i.e., any change in immune function may increasethe susceptibility to disease) or threshold-like model (i.e., smallchanges in immune function may be without appreciable changesin host resistance). Although there are suggestions in the literaturethat this relationship may follow a linear model, the answer to thisquestion in the context of a broad array of chemicals is not clear.Further investigation in this area is warranted.
CONCLUDING COMMENTAND FUTURE DIRECTIONS
Our understanding of the immune system as a target for toxicity,whether it be via xenobiotic-induced immune injury or immune-mediated disease, continues to progress in concert with our knowl-edge of the biochemistry and physiology of the immune system.The balance between immune recognition and destruction of for-eign invaders and the proliferation of these microbes and/or can-cer cells can be a precarious one. Xenobiotics that alter the im-mune system can upset this balance, giving the edge to the invader.Furthermore, new xenobiotics continuously being introduced rep-resent the potential for increased hypersensitivity and/or autoim-mune responses. Validated methods are in place to detect xenobi-otics that produce adverse effects related to the immune system.Once these are identified, fundamental principles of toxicology canbe applied leading to risk assessment and determination of “no-effect” levels. These methods must continually be improved usingthe latest knowledge and technologies in order to provide a safeenvironment.
2996R_ch12_419-470 4/27/01 10:02 AM Page 463
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

464 UNIT 4 TARGET ORGAN TOXICITY
REFERENCES
Burns LA, LeVier DG, Munson AE: Immunotoxicology of Arsenic, in DeanJH, Luster MI, Munson AE, Kimber I (eds): Immunotoxicology andImmunopharmacology, 2d ed. New York: Raven Press, 1994, pp 213–225.
Burns-Naas LA, Mast RW, Klykken PC, et al: Toxicology and humoral im-munity assessment of decamethylcyclopentasiloxane (D5) following a1-month whole-body inhalation exposure in Fischer 344 rats. ToxicolSci 43:28–38, 1998.
Calne RY, Rolles K, White DJ, et al: Cyclosporin A in clinical organ graft-ing. Transplant Proc 13:349–358, 1981.
Cambridge G, Wallace H, Bernstein RM, Leaker B: Autoantibodies tomyeloperoxidase in idiopathic and drug-induced systemic lupus ery-thematosus and vasculitis. Br J Rheumatol 33(2):109–114, 1994.
Canoso RT, de Oliveira RM, Nixon RA: Neuroleptic-associated autoanti-bodies. A prevalence study. Biol Psychiatry 27(8):863–870, 1990.
Catania RA, Chaudry IH: Immunological consequences of trauma andshock. Ann Acad Med Singapore 28(1):120–132, 1999.
Chao CC, Molitor TW, Gekker G, et al: Cocaine-mediated suppression ofsuperoxide production by human peripheral blood mononuclear cells.J Pharmacol Exp Ther 256:255–258, 1991.
Chao CC, Sharp BM, Pomeroy C, et al: Lethality of morphine in mice in-fected with Toxoplasma gondii. J Pharmacol Exp Ther 252:605–609,1990.
Coleman JW, Sim E: Autoallergic responses to drugs: Mechanistic aspects,in Dean JH, Luster MI, Munson AE, Kimber I (eds): Immunotoxicol-ogy and Immunopharmacology, 2d ed. New York: Raven Press, 1994,pp 553–572.
Coombs RRA, Gell PGH: Classification of allergic reactions responsiblefor clinical hypersensitivity and disease, in Gell PGH, Coombs RRA,Lachmann PJ (eds): Clinical Aspects of Immunology, Oxford, England:Oxford University Press, 1975, p 761.
Dean JH, Luster MI, Boorman GA: Immunotoxicology, in Sirois P, Rola-Pleszcynski M (eds): Immunopharmacology. Amsterdam: Elsevier,1982, pp 349–397.
Dean JH, Luster MI, Boorman GA, et al: Selective immunosuppression re-sulting from exposure to the carcinogenic congener of benzopyrene inB6C3F1 mice. Clin Exp Immunol 52:199–206, 1983.
Dean JH, Lauer LD, House RV, et al: Studies of immune function and hostresistance in B6C3F1 mice exposed to formaldehyde. Toxicol ApplPharmacol 72:519–529, 1984.
Dean JH, Luster MI, Munson AE, Amos H (eds): Immunotoxicology andImmunopharmacology. New York: Raven Press, 1985.
Dean JH, Luster MI, Munson AE, Kimber I (eds): Immunotoxicology andImmunopharmacology, 2d ed. New York: Raven Press, 1994.
Dearman RJ, Kimber I: Cytokine fingerpriniting: Characterization of chem-ical allergens. Methods 79(1):56–63, 1999.
Dearman RJ, Kimber I: Differential stimulation of immune function by res-piratory and contact chemical allergens. Immunology 72:563–570,1991.
Dearman RJ, Spence LM, Kimber I: Characterization of murine immuneresponses to allergenic diisocyanates. Toxicol Appl Pharmacol112:190–197, 1992.
Delaney B, Strom SC, Collins S, Kaminski NE: Carbon tetrachloride suppresses T-cell-dependent immune responses by induction to trans-forming growth factor-�1. Toxicol Appl Pharmacol 126:98–107,1994.
Demoly P, Michel F, Bousquet J: In vivo methods for study of allergy skintests, techniques and interpretation, in Middleton E Jr, Reed C, EllisE, et al (eds): Allergy Principles and Practice. St. Louis: Mosby, 1998,pp 430–439.
Deo MG, Gangal S, Bhisey AN, et al: Immunological, mutagenic, and geno-toxic investigations in gas-exposed population of Bhopal. Indian J MedRes 86:63–76, 1987.
Ananth J, Johnson R, Kataria P, et al: Immune dysfunctions in psychiatricpatients. Psychiatr J 14(4):542–546, 1989.
Araneo BAT, Dowell T, Moon HB, Daynes RA: Regulation of murine lym-phokine production in vivo: Ultraviolet radiation exposure depressesIL-2 and enhances IL-4 production through an IL-1 independent mech-anism. J Immunol 143:1737–1745, 1989.
Aranyi C, O’Shea W, Graham J, Miller F: The effects of inhalation of or-ganic chemical air contaminants on murine lung host defenses. Fun-dam Appl Toxicol 6:713–720, 1986.
Arora PK, Fride E, Petitto J, et al: Morphine-induced immune alterationsin vivo. Cell Immunol 126:343–353, 1990.
Barnett JB: Developmental immunotoxicology, in Smialowicz RJ, Holsap-ple MP (eds): Experimental Immunotoxicology. Boca Raton, FL: CRCPress, 1996, pp 47–62.
Barnett JB, Rodgers KE: Pesticides, in Dean JH, Luster MI, Munson AE,Kimber I (eds): Immunotoxicology and Immunopharmacology, 2d ed.New York: Raven Press, 1994, pp 191–212.
Bernstein JA, Bernstein IL: Clinical aspects of respiratory hypersensitivityto chemicals, in Dean JH, Luster MI, Munson AE, Kimber I (eds):Immunotoxicology and Immunopharmacology, 2d ed. New York:Raven Press, 1994, pp 617–642.
Biagini RE: Epidemiology studies in immunotoxicity evaluations. Toxicol-ogy 129:37–54, 1998.
Biegel LB, Flaws JA, Hirshfield AN, et al: 90-day feeding and one-generation reproduction study in Crl:CD BR rats with 17�-estradiol.Toxicol Sci 44:116–142, 1998.
Blalock JE: A molecular basis for bidirectional communication betweenthe immune and neuroendocrine systems. Physiol Rev 69:1–32, 1989.
Bly JE, Quiniou SM, Clem LW: Environmental effects on fish immunemechanisms. Dev Biol Stand 90:33–43, 1997.
Bouaboula M, Rinaldi M, Carayon P, et al: Cannabinoid receptor expres-sion in human leukocytes. Eur J Biochem 214:173–180, 1993.
Bradley SG, Morahan PS, Approaches to assessing host resistance. Envi-ron Health Perspect 43:61–69, 1982.
Bradley SG, Munson AE, McCay JA, et al: Subchronic 10 day immuno-toxicity of polydimethylsiloxane (silicone) fluid, gel and elastomerand polyurethane disks in female B6C3F1 mice. Drug Chem Toxicol17(3):175–220, 1994a.
Bradley SG, White KL Jr, McCay JA, et al: Immunotoxicity of 180 dayexposure to polydimethylsiloxane (silicone) fluid, gel and elastomerand polyurethane disks in female B6C3F1 mice. Drug Chem Toxicol17(3):221–269, 1994b.
Brown EJ, Albers MW, Shin TB, et al: A mammalian protein targeted byG1-arresting rapamycin receptor complex. Nature 369:756–758, 1994.
Buehler EV: Delayed contact hypersensitivity in the guinea pig. Arch Der-matol 91:171–177, 1965.
Burchiel SW, Krekvliet NI, Gerberick GF, et al: Workshop overview: As-sessment of immunotoxicity by multiparameter flow cytometry. Fun-dam Appl Toxicol 38:38–54, 1997.
Burchiel SW, Lauer FT, Gurule, D, et al: Uses and applications of flow cy-tometry in immunotoxicity testing. Methods 19:28–35, 1999.
Burns LA, Bradley SG, White KL, et al: Immunotoxicology of nitroben-zene in female B6C3F1 mice. Drug Chem Toxicol 17(3):271–315,1994a.
Burns LA, Bradley SG, White KL, et al: Immunotoxicology of nitrotoluenesin female B6C3F1 mice. I. Para-nitrotoluene. Drug Chem Toxicol17(3):359–399, 1994b.
Burns LA, Bradley SG, White KL, et al: Immunotoxicology of nitrotoluenesin female B6C3F1 mice. II. Meta-nitrotoluene. Drug Chem Toxicol17(3):401–436, 1994c.
Burns LA, Bradley SG, White KL, et al: Immunotoxicology of 2,4-di-aminotoluene in female B6C3F1 mice. Drug Chem Toxicol 17(3):317–358, 1994d.
2996R_ch12_419-470 4/27/01 10:02 AM Page 464
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

CHAPTER 12 TOXIC RESPONSES OF THE IMMUNE SYSTEM 465
Descotes J (ed): Immunotoxicology and Drugs and Chemicals. Amsterdam:Elsevier, 1986.
Desi I, Varga L, Dobronyi L, Szklenarik G: Immunotoxicological investi-gation of the effects of a pesticide: Cypermethrin. Arch Toxicol (suppl8):305–309, 1985.
DeSwarte RD: Drug allergy, in Patterson R (ed): Allergic Diseases, Diag-nosis and Management, 3d ed. Philadelphia: Lippincott, 1985, pp 505–661.
deWeck AL: Immunopathological mechanisms and clinical aspects of al-lergic reactions to drugs, in deWeck AL, Bundgaard H (eds): Hand-book of Experimental Pharmacology: Allergic Reactions to Drugs.New York: Springer-Verlag, 1983, pp 75–133.
Eisenstein TK, Bussiere JL, Rogers TJ, Adler MW: Immunosuppressive ef-fects of morphine on immune responses in mice. Adv Exp Med Biol335:41–52, 1993.
Enk AH, Angeloni VL, Udey MC, Katz SI: Inhibition of Langerhans cellantigen-presenting function by IL-10. J Immunol 151:2390–2398,1993.
Exon JH, Mather GG, Bussiere JL, et al: Effects of subchronic exposureof rats to 2-methoxyethanol or 2-butoxyethanol: Thymic atrophy andimmunotoxicity. Fundam Appl Toxicol 16(4):830–840, 1991.
Fock KM, Feng PH, Tey BH: Autoimmune disease developing after aug-mentation mammoplasty: Report of 3 cases. J Rheumatol 11:98–100,1984.
Fuchs BA, Sanders VM: The role of brain-immune interactions in im-munotoxicology. Crit Rev Toxicol 24(2):151–176, 1994.
Gabliks J, Al-zubaidy T, Askari E: DDT and immunological responses. 3.Reduced anaphylaxis and mast cell population in rats fed DDT. ArchEnviron Health 30:81–84, 1975.
Gabliks J, Askari EM, Yolen N: DDT and immunological responses. I.Serum antibodies and anaphylactic shock in guinea pig. Arch EnvironHealth 26:305–309, 1973.
Gad SC, Dunn BJ, Dobbs DW, et al: Development and validation of an al-ternative dermal sensitization test: The mouse ear swelling test(MEST). Toxicol Appl Pharmacol 84:93–114, 1986.
Gaworski CL, Vollmuth TA, Dozier MM, et al: An immunotoxicity as-sessment of food flavouring ingredients. Fundam Chem Toxicol32(5):409–415, 1994.
Gerberick GF: Development, validation, and application of expert systemsfor predicting skin sensitization. Society of Toxicology Continuing Ed-ucation course, The Practice of Structure-Activity Relationships (SAR)in Toxicology, March, 1999.
Germolec DR, Woolhiser MR, Meade BJ: Allergy to natural rubber latex.Chem Health Saf July: 44–48, 1999.
Giorgio RL, Bongiorno L, Trani E, et al: Review paper: Positive and neg-ative immunomodulation by opioid peptides. Int J Immunopharmacol18(1):1–16, 1996.
Gogal Jr RM, Ahmed SA, Smith SA, Holladay SD: Commentary. Man-dates to develop non-mammalian models for chemical immunotoxic-ity evaluation: Are fish a viable alternate to rodents? Toxicol Lett106:89–92, 1999.
Gorelik E, Heberman R: Susceptibility of various strains of mice to ure-thane-induced lung tumors and depressed natural killer activity. J NatlCancer Inst 67:1317–1322, 1981.
Grabbe S, Schwarz T: Immunoregulatory mechanisms involved in elicita-tion of allergic contact hypersensitivity. Immunol Today 19:37–44,1998.
Graham C, Rosenkrantz HS, Karol MH: Structure-activity model of chem-icals that cause human respiratory sensitization. Reg Toxicol Pharma-col 26:296–306, 1997.
Grammer LC: Occupational immunologic lung disease, in Patterson R (ed):Allergic Diseases, Diagnosis and Management, 3d ed. Philadelphia:Lippincott, 1985, pp 691–708.
Greenlee WF, Dold KM, Irons RD, Osborne R: Evidence for a direct ac-tion of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) on thymic ep-ithelium. Toxicol Appl Pharmacol 79:112–120, 1985.
Greenlee WF, Dold KM, Osborne R: A proposed model for the actions ofTCDD on epidermal and thymic epithelial target cells, in Poland A,Kimbrough RD (eds): Banbury Report 18: Biological Mechanisms ofDioxin Action. Cold Spring Harbor, NY: Cold Spring Harbor Labora-tory, 1984, pp 435ff.
Greenspan BJ, Morrow PE: The effects of in vitro and aerosol exposuresto cadmium on phagocytosis by rat pulmonary macrophages. FundamAppl Toxicol 4:48–57, 1984.
Gutman AA: Allergens and other factors important in atopic disease, in Pat-terson R (ed): Allergic Diseases, Diagnosis and Management, 3d ed.Philadelphia: Lippincott, 1985, pp 123–175.
Guzzie PJ: Immunotoxicology in pharmaceutical development. in Gad SC(ed): Safety Assessment for Pharmaceuticals. New York: Wiley, 1995,pp 325–385.
Haines KA, Reibman J, Callegari PE, et al: Cocaine and its derivativesblunt neutrophil functions without influencing phosphorylation of a47-kilodalton component of the reduced nicotinamide-adenine dinu-cleotide phosphate oxidase. J Immunol 144:4757–4764, 1990.
Hanifin JM, Epstein WL, Cline MJ: In vitro studies of granulomatous hy-persensitivity to beryllium. J Invest Dermatol 55:284–288, 1970.
Harpey JP, Caille B, Moulias R, Goust JM: Lupus-linked syndrome inducedby D-penicillamine in Wilson’s Disease. Lancet 1:292, 1971.
Henderson WR, Fukuyama K, Epstein WL, Spitler LE: In vitro demon-stration of delayed hypersensitivity in patients with beryliosis. J In-vest Dermatol 58:5–8, 1972.
Hendrick DJ, Rando RJ, Lane DJ, Morris MJ: Formaldhyde asthma: Chal-lenge exposure levels and fate after five years. J Occup Med 24:893–897, 1982.
Hjorth N: Diagnostic patch testing, in Marzulli FN, Maibach HI (eds): Der-matotoxicology, 3d ed. Washington, DC: Hemisphere, 1987, pp 307–317.
Hoffman EC, Reyes H, Chu F-F, et al: Cloning of a factor required for ac-tivity of the Ah (dioxin) receptor. Science 252:954, 1991.
Holladay SD: Prenatal immunotoxicant exposure and postnatal autoim-mune disease. Environ Health Perspect 107(suppl 5):687–691, 1999.
Holladay SD, Blaylock BL, Comment CE, et al: Fetal thymic atrophy af-ter exposure to T-2 toxin: Selectivity for lymphoid progenitor cells.Toxicol Appl Pharmacol 121(1):8–14, 1993.
Holladay SD, Comment CE, Kwon J, Luster MI: Fetal hematopoietic al-terations after maternal exposure to ethylene glycol monomethyl ether:Prolymphoid cell targeting. Toxicol Appl Pharmacol 129(1):53–60,1994.
Holsapple MP: Immunotoxicity of halogenated aromatic hydrocarbons, inSmialowicz R, Holsapple MP (eds): Experimental Immunotoxicology.Boca Raton, FL: CRC Press, 1996, pp 257–297.
Holsapple MP, Matulka RA, Stanulis ED, Jordan SD: Cocaine and im-munocompetence: Possible role of reactive metabolites. Adv Exp MedBiol 335:121–126, 1993.
Holsapple MP, Morris DL, Wood SC, Snyder NK: 2,3,7,8-tetra-chlorodibenzo-p-dioxin-induced changes in immunocompetence: Pos-sible mechanisms. Annu Rev Pharmacol Toxicol 31:73–100, 1991a.
Holsapple MP, Munson AE, Munson JA, Bick PH: Suppression of cell-mediated immunocompetence after subchronic exposure to diethyl-stilbestrol in female B6C3F1 mice. J Pharmacol Exp Ther 227:130–138, 1983.
Holsapple MP, Snyder NK, Gokani V, et al: Role of Ah receptor in sup-pression of in vivo antibody response by 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) is dependent on exposure conditions. Fed Am SocExp Biol J 5(4):A508, 1991c.
Holsapple MP, Snyder NK, Wood SC, Morris DL: A review of 2,3,7,8-tetrachlorodibenzo-p-dioxin-induced changes in immunocompetence.Toxicology 69:219–255, 1991b.
House RV, Laurer LD, Murray MJ, et al: Immunological studies in B6C3F1mice following exposure to ethylene glycol monomethyl ether and itsprincipal metabolite methoxyacetic acid. Toxicol Appl Pharmacol77(2):358–362, 1985.
2996R_ch12_419-470 4/27/01 10:02 AM Page 465
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

466 UNIT 4 TARGET ORGAN TOXICITY
House RV, Ratajczak HV, Gauger JR, et al: Immune function and host de-fense in rodents exposed to 60-Hz magnetic fields. Fundam Appl Tox-icol 34(2):228–239, 1996.
Institute of Medicine (IOM): in Bondurant S, Ernster V, Herdman R (eds):Safety of Silicone Breast Implants. Washington, DC: National Acad-emy Press, 2000.
International Life Sciences Institute (ILSI): Application of Flow Cytome-try to Immunotoxicity Testing: Summary of a Workshop. Washington,DC: ILSI Press, 1999.
International Programme on Chemical Safety (IPCS): EnvironmentalHealth Criteria 180: Principles and Methods for Assessing DirectImmunotoxicity Associated with Exposure to Chemicals. Geneva,Switzerland: World Health Organization, 1996.
Jeon YJ, Han SH, Yang KH, Kaminski NE: Induction of liver-associatedtransforming growth factor �1 (TGF-�1) mRNA expression by car-bon tetrachloride leads to the inhibition of T helper 2 cell-associatedlymphokines. Toxicol Appl Pharmacol 144:27–35, 1997.
Jerrells TR, Pruett SB: Immunotoxic effects of ethanol, in Dean JH, Lus-ter MI, Munson AE, Kimber I (eds): Immunotoxicology and Im-munopharmacology, 2d ed. New York: Raven Press, 1994, pp 323–347.
Jiang X, Khursigara G, Rubin RL: Transformation of lupus-inducing drugsto cytotoxic products by activated neutrophils. Science266(5186):810–813, 1994.
Kalland T: Alterations of antibody responses in female mice after neona-tal exposure to diethylstilbestrol. J Immunol 124:194–198, 1980.
Kalland T, Strand O, Forsberg JG: Long-term effects of neonatal estrogentreatment on mitogen responsiveness of mouse spleen lymphocytes. JNatl Cancer Inst 63:413–421, 1979.
Kaminski NE: Mechanisms of immune modulation by cannabinoids, inDean JH, Luster MI, Munson AE, Kimber I (eds): Immunotoxicologyand Immunopharmacology, 2d ed. New York: Raven Press, 1994, pp349–362.
Kaminski NE, Abood ME, Kessler FK, et al: Identification of a function-ally relevant cannabinoid receptor on mouse spleen cells involved incannabinoid-mediated immune modulation. Mol Pharmacol 42:736–742, 1992.
Karol MH: Models of contact and respiratory sensitivity and structure-activity relationships, continuing education course, Chemical Hyper-sensitivity. New Orleans: Society of Toxicology, March 1999.
Karol MH, Graham C, Gealy R, et al: Structure-activity relationships andcomputer-assisted analysis of respiratory sensitization potential. Tox-icol Lett 86:187–191, 1996.
Kately JR, Bazzell SJ: Immunological studies in cattle exposed to poly-brominated biphenyl. Environ Health Perspect 23:750, 1978.
Kawabata TT, White KL Jr: Effects of naphthlene metabolites on the invitro humoral immune response. J Toxicol Environ Health 30:53–67,1990.
Kayama F, Yamashita U, Kawamoto T, Kodama Y: Selective depletion ofimmature thymocytes by oral administration of ethylene glycolmonomethyl ether. Int J Immunopharmacol 13(5):531–540, 1991.
Kenna JG, Neuberger J, Williams R: Identification by immunoblotting ofthree halothane-induced microsomal polypeptide antigens recognizedby antibodies in sera from patients with halothane-associated hepati-tis. J Pharmacol Exp Ther 242:733–740, 1987.
Kerkvliet NI: Halogenated aromatic hydrocarbons (HAH) as immunotox-icants, in Kende M, Gainer J, Chirigos M (eds): Chemical Regulationof Immunity in Veterinary Medicine. New York: Liss, 1984, pp 369–387.
Kerkvliet NI, Brauner JA: Flow cytometric analysis of lymphocyte sub-populations in the spleen and thymus of mice exposed to an acuteimmunosuppressive dose of 2,3,7,8-tetrachlorodibenzo-p-dioxin(TCDD). Environ Res 52:146–164, 1990.
Kerkvliet NI, Brauner JA: Mechanisms of 1,2,3,4,6,7,8-Hepta-chlorodibenzo-p-dioxin (HpCDD)-induced humoral immune suppres-sion: Evidence of primary defect in T cell regulation. Toxicol ApplPharmacol 87:18–31, 1987.
Kerkvliet NI, Burleson GR: Immunotoxicity of TCDD and related halo-genated aromatic hydrocarbons, in Dean JH, Luster MI, Munson AE,Kimber I (eds): Immunotoxicology and Immunopharmacology, 2d ed.New York: Raven Press, 1994, pp 97–121.
Kerkvliet NI, Steppan LB, Brauner JA, et al: Influence of the Ah locus onthe humoral immunotoxicity of 2,3,7,8-tetrachlorodibenzo-p-dioxin:Evidence for Ah receptor-dependent and Ah receptor-independentmechanisms of immunosuppression. Toxicol Appl Pharmacol 105:26–36, 1990.
Kilburn KH, Warshaw RH: Chemical-induced autoimmunity, in Dean JH,Luster MI, Munson AE, Kimber I (eds): Immunotoxicology and Im-munopharmacology, 2d ed. New York: Raven Press, 1994, pp 523–538.
Kilburn KH, Warshaw RH: Prevalence of symptoms of systemic lupus ery-thematosus (SLE) and of fluorescent antinuclear antibodies associatedwith chronic exposure to trichlorethylene and other chemicals in wellwater. Environ Res 57:1–9, 1992.
Kim BS, Smialowicz RJ: The role of metabolism in 2-methoxyethanol-induced suppression of in vitro polyclonal antibody responses by ratand mouse lymphocytes. Toxicology 123(3):227–239, 1997.
Kimber I, Kerkvliet NI, Taylor SL, et al: Toxicology of protein allergenic-ity: Prediction and characterization. Toxicol Sci 48:157–162, 1999.
Kimber I, Basketter DA, Dearman RJ: Divergent antibody responses in-duced in mice by food proteins. Toxicol Sci 54(1): A1162, 2000.
Kimber IA, Hilton J, Botham PA, et al: The murine local lymph node assay: Results of an inter-laboratory trial. Toxicol Lett 55:203–213,1991.
Kimber IA, Hilton J, Weisenberger C: The murine local lymph node assayfor identification of contact allergens: A preliminary evaluation of insitu measurement of lymphocyte proliferation. Contact Dermatitis21:215–220, 1989.
Klecak G: Identification of contact allergens: Predicitive tests in animals,in Marzulli FN, Maibach HI (eds): Dermatotoxicology, 3d ed.Washington, DC: Hemisphere, 1987, pp 227–290.
Klykken PC, Galbraith TM, Kolesar GB, et al: Toxicology and humoralimmunity assessment of octamethylcyclotetrasiloxane (D4) followinga 28-day whole-body vapor inhalation exposure in Fischer 344 rats.Drug Chem Toxicol 22(4):655–677, 1999.
Klykken PC, White KL Jr: The adjuvancy of silicones: Dependency oncompartmentalization, in Potter M, Rose NR (eds): Current Topics inMicrobiology and Immunology. Vol 210. New York: Springer-Verlag,1996, pp 113–121.
Knippels LMJ, Spanhaak S, Pennicks AH: Oral sensitization to peanut pro-teins: A brown Norway rat food allergy model. Toxicol Sci 54(1):A1161, 2000.
Knippels LMJ, Penninks AH, Smit JJ, Houben GF: Immune-mediated ef-fects upon oral challenge of ovalbumin-sensitized brown Norway rats:Further characterization of a rat food allergy model. Toxicol Appl Phar-macol 156:161–169, 1999.
Koller LD: Immunotoxicology of heavy metals. Int J Immunopharmacol2:269–279, 1980.
Krzystyniak K, Hugo P, Flipo D, Fournier M: Increased susceptibility tomouse hepatitis virus 3 of peritoneal macrophages exposed to diel-drin. Toxicol Appl Pharmacol 80:397–408, 1985.
Kunz J, Henriquez R, Schneider U, et al: Target of rapamycin in yeast,TOR2, is an essential phosphatidylinositol kinase homolog requiredfor G1 progression. Cell 73:585–596, 1993.
Ladics GS, Kawabata TT, Munson AE, White KL Jr: Metabolism ofbenzo[a]pyrene by murine splenic cell types. Toxicol Appl Pharma-col 116:248–257, 1992a.
Ladics GS, Kawabata TT, Munson AE, White KL Jr: Generation of 7,8-dihydroxy-9,10-epoxy-7,8,9,10-tetrahydrobenzo[a]pyrene by murinesplenic macrophages. Toxicol Appl Pharmacol 115:72–79, 1992b.
Ladics GS, Kawabata TT, White KL Jr: Suppression of the in vitro humoralimmune response of mouse splenocytes by 7,12-dimethylbenz[a]an-thracene metabolites and inhibition of suppression by a-naph-thoflavone. Toxicol Appl Pharmacol 110:31–44, 1991.
2996R_ch12_419-470 4/27/01 10:02 AM Page 466
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

CHAPTER 12 TOXIC RESPONSES OF THE IMMUNE SYSTEM 467
Ladics GS, Smith C, Heaps K, Loveless SE: Evaluation of the humoral im-mune response to CD rats following a 2-week exposure to the pesti-cide carbaryl by the oral, dermal, and inhalation routes. J Toxicol En-viron Health 42:143–156, 1994.
Ladics GS, Smith C, Nicastro S, et al: Evaluation of the primary humoralimmune response following exposure of male rats to 17� estradiol orflutamide for 15 days. Toxicol Sci 46:75–82, 1998.
Lawrence DA: Immunotoxicity of heavy metals, in Dean JH, Luster MI,Munson AE, Amos H (eds): Immunotoxicology and Immunopharma-cology. New York: Raven Press, 1985, pp. 341–353.
Lawrence DA (ed), in Sipes IG, McQueen, CA, Gandolfi AJ (eds-in-chief):Comprehensive Toxicology. Vol 5. Toxicology of the Immune System.New York: Elsevier, 1997.
Lawrence DA, Kim D: Central/peripheral nervous system and immune re-sponses. Toxicology 142:189–291, 2000.
Le UL, Munson AE: Differential effects of octamethylcyclotetrasiloxane(D4) on CD4� and CD8�-T cell functions in B6C3F1 mice. FundamAppl Toxicol Suppl 36(1):A1346, 1997.
Lefkowitz SS, Vaz A, Lefkowitz DL: Cocaine reduces macrophage killingby inhibiting reactive oxygen intermediates. Int J Immunopharmacol15:717–721, 1993.
LeVier DG, Brown RD, McCay JA, et al: Hepatic and splenic phagocyto-sis in female B6C3F1 mice implanted with morphine sulfate pellets.J Pharmacol Exp Ther 367:357–363, 1993.
LeVier DG, McCay JA, Stern ML, et al: Immunotoxicological profile ofmorphine sulfate in B6C3F1 mice. Fundam Appl Toxicol 22:525–542,1994.
LeVier DG, Musgrove DL, Munson AE: The effect of octamethylcyclote-trasiloxane (D4) on the humoral immune response of B6C3F1 mice.Fundam Appl Toxicol Suppl 15(1):A1199, 1995.
Levy MH, Wheelock EF: Effects of intravenous silica on immune and non-immune functions of the murine host. J Immunol 115:41–48,1975.
Liberman P, Crawford L, Drewry RD Jr, Tuberville A: Allergic diseases ofthe eye and ear, in Patterson R (ed): Allergic Diseases, Diagnosis andManagement, 3d ed. Philadelphia: Lippincott, 1985, pp 374–407.
Looney RJ, Frampton MW, Byam J, et al: Acute respiratory exposure ofhuman volunteers to octamethylcyclotetrasiloxane (D4): Absence ofimmunological effects. Toxicol Sci 44:214–220, 1998.
Lundberg K, Grovnick K, Goldschmidt TJ, et al: 2,3,7,8-tetra-chlorodibenzo-p-dioxin (TCDD) alters intrathymic T cell developmentin mice. Chem Biol Interact 74:179, 1990.
Luster MI, Boorman GA, Dean JH, et al: Effect of in utero exposure of di-ethylstilbestrol on the immune response in mice. Toxicol Appl Phar-macol 47:287–293, 1979.
Luster MI, Boorman GA, Dean JH, et al: The effect of adult exposure todiethylstilbestrol in the mouse: Alterations in immunological func-tions. J Retic Soc 28:561–569, 1980.
Luster MI, Boorman GA, Korach KS, et al: Mechanisms of estrogen-induced myelotoxicity: Evidence of thymic regulation. Int J Im-munopharmacol 6:287–297, 1984a.
Luster MI, Dean JH, Boorman GA, et al: Host resistance and immune func-tions in methyl and ethyl carbamate treated mice. Clin Exp Immunol50:223–230, 1982.
Luster MI, Hayes HT, Korach K, et al: Estrogen immunosuppression is reg-ulated through estrogenic responses in the thymus. J Immunol133:110–116, 1984b.
Luster MI, Munson AE, Thomas PT, et al: Development of a testing bat-tery to assess chemical-induced immunotoxicity: National ToxicologyProgram’s guidelines for immunotoxicity evaluation in mice. FundamAppl Toxicol 10:2–9, 1988.
Luster MI, Portier C, Pait DG, et al: Risk assessment in immunotoxicol-ogy. I. Sensitivity and predictability of immune tests. Fundam ApplToxicol 18:200–210, 1992.
Luster MI, Portier C, Pait DG, and Germolec DR: Use of animal studiesin risk assessment for immunotoxicology. Toxicology 92:229–243,1994.
Luster MI, Tucker JA, Germolec DR, et al: Immunotoxicity studies in miceexposed to methyl isocyanate. Toxicol Appl Pharmacol 86:140–144,1986.
Madden KS, Sanders VM, Felten DL: Catecholamine influences and sym-pathetic neural modulation of immune responsiveness. Annu Rev Phar-macol Toxicol 35:417–448, 1995.
Magnusson B, Kligman AM: The identification of contact allergens by an-imal assay. The guinea pig maximization test. J Invest Dermatol52:268–276, 1969.
Maibach H: Formaldehyde: Effects on animal and human skin, in GibsonJ (ed): Formaldehyde Toxicity. Washinton, DC: Hemisphere, 1983, pp166–174.
Marzulli FN, Maibach HI: Contact allergy: Predictive testing in humans,in Marzulli FN, Maibach HI (eds): Dermatotoxicology, 3d ed. Wash-ington, DC: Hemisphere, 1987, pp 319–340.
Marzulli FN, Maibach HI: The use of graded concentrations in studyingskin sensitizers: Experimental contact sensitization in man. Food Cos-met Toxicol 12:219–227, 1974.
Matveyeva M, Hartmann CB, Harrison MT et al: Delta(9)-tetrahydro-cannabinol selectively increases aspartyl cathepsin D proteolytic ac-tivity and impairs lysozomal processing by macrophages. Int J Im-munopharmacol 22(5):373–381, 2000.
McCabe MJ Jr: Mechanisms and consequences of immunomodulation bylead, in Dean JH, Luster MI, Munson AE, Kimber I (eds): Immuno-toxicology and Immunopharmacology, 2d ed. New York: Raven Press,1994, pp 143–162.
McDonough RJ, Madden JJ, Falck A, et al: Alteration of T and null lym-phocyte frequencies in the peripheral blood of human opiate addicts:In vivo evidence for opiate receptor sites on T lymphocytes. J Im-munol 125:2539–2543, 1980.
Menna JH, Barnett JB, Soderberg LSF: Influenze type A infection of miceexposed in utero to chlordane: Survival and antibody studies. ToxicolLett 24:45–52, 1985.
Menné T: Reactions to systemic exposure to contact allergens, in MarzulliFN, Maibach HI (eds): Dermatotoxicology, 3d ed. Washington, DC:Hemisphere, 1987, pp 535–552.
Miller K, Brown RC: The immune system and asbestos-associated disease,in Dean JH, Luster MI, Munson AE, Amos H (eds): Immunotoxicol-ogy and Immunopharmacology. New York: Raven Press, 1985, pp429–440.
Miller N, Wilson M, Bengten E, et al: Functional and molecular charac-terization of teleost leukocytes. Immunol Rev 166:187–197, 1998.
Mishell RI, Dutton RW: Immunization of dissociated mouse spleen cellculture from normal mice. J Exp Med 126:423–442, 1967.
Morice WG, Brunn GJ, Wiederrecht G, et al: Rapamycin-induced inhibi-tion of p34cdc2 kinase activation is associated with G1/S phase growtharrest in T lymphocytes. J Biol Chem 268:3734–3738, 1993.
Mosmann TR, Coffman RL: Heterogeneity of cytokine secretion patternsand function of helper T cells. Adv Immunol 46:111–147, 1989.
Mosmann TR, Schumacher JH, Street NF, et al: Diversity of cytokine syn-thesis and function of mouse CD4� T cells. Immunol Rev 123:209–229, 1991.
Munro S, Thomas KL, Abu-Shaar M: Molecular characterization of pe-ripheral receptor for cannabinoids. Nature 365:61–65, 1993.
Munson AE: Immunological evaluation of octamethylcyclotetrasiloxane(D4) using a twenty-eight day exposure in male and female Fischer344 rats. Document Control Number 86980000072. Washington, DC:USEPA-OPPT, TSCA Document Processing Center, 1998.
Munson AE, McCay JA, Brown RD, et al: The immune status of Fischer344 rats administered octamethylcyclotetrasiloxane (D4) by oral gav-age. Fundam Appl Toxicol Suppl 36(1):A1347, 1997.
Murphy WG, Kelton JG: Immune haemolytic anaemia and thrombocy-topaenia with drugs and antibodies. Biochem Soc Trans 19:183–186,1991.
Myers MJ, Schook LB: Immunotoxicity of nitrosamines, in Smialowicz R,Holsapple MP (eds): Experimental Immunotoxicology. Boca Raton,FL: CRC Press, 1996, pp 351–366.
2996R_ch12_419-470 4/27/01 10:02 AM Page 467
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

468 UNIT 4 TARGET ORGAN TOXICITY
Mydlarski P, Katz A, Sauder D: Contact dermatitis, in Middleton E Jr, ReedC, Ellis E, et al (eds:) Allergy Principles and Practice. St. Louis:Mosby, 1998, pp 1141–1143.
Newman LS: Beryllium lung disease: The role of cell-mediated immunityin pathogenesis, in Dean JH, Luster MI, Munson AE, Kimber I (eds):Immunotoxicology and Immunopharmacology, 2d ed. New York:Raven Press, 1994, pp 377–394.
NIH Publication No. 99-4494: The Murine Local Lymph Node Assay: ATest Method for Assessing the Allergic Contact Dermatitis Potentialof Chemicals/Compounds. Washington, DC: National Institute of Environmental Health Sciences, National Institutes of Health, U.S.Public Health Service, Department of Health and Human Services,1999.
Noonan FP, De Fabo EC: Ultraviolet-B-dose response curves for local andsystemic immunosuppression are identical. Photochem Photobiol52:801–810, 1990.
Ou D, Shen ML, Luo YD: Effects of cocaine on the immune system ofBALB/c mice. Clin Immunol Immunopathol 52:305–312, 1989.
Paul WE (ed): Fundamental Immunology, 4d ed. New York: Raven Press,1999.
Paul WE, Seder RA: Lymphocyte responses and cytokines. Cell 76:241–251, 1994.
Pelletier L, Castedo M, Bellon B, Druet P: Mercury and autoimmunity, inDean JH, Luster MI, Munson AE, Kimber I (eds): Immunotoxicologyand Immunopharmacology, 2d ed. New York: Raven Press, 1994, pp539–552.
Penninks AH, Snoeij NJ, Pieters RHH, Seinen W: Effect of organotin com-pounds on lymphoid organs and lymphoid functions: An overview, inDayan AD, Hentel RF, Heseltine E, et al (eds): Immunotoxicity of Met-als and Immunotoxicology. New York: Plenum Press, 1990, pp 191–207.
Pernis B, Paronetto F: Adjuvant effect of silica (tridymite) on antibody pro-duction. Proc Soc Exp Biol Med 110:390–392, 1962.
Peterson PK, Gekker G, Chao CC, et al: Cocaine potentiates HIV-1 repli-cation in human peripheral blood mononuclear cell cultures. Involve-ment of transforming growth factor-b. J Immunol 146:81–84, 1991.
Potter M, Rose NR (eds): Immunology of Silicones, New York: Springer-Verlag, 1996.
Pruett SB, Barnes DB, Han YC, Munson AE: Immunotoxicological char-acteristics of sodium methyldithiocarbamate. Fundam Appl Toxicol18:40–47, 1992a.
Pruett SB, Han Y, Fuchs BA: Morphine suppresses primary humoral im-mune responses by a predominantly indirect mechanism. J Pharma-col Exp Ther 262:923–928, 1992b.
Quesniaux VFJ: Interleukins 9, 10, 11, and 12, and kit ligand: A briefoverview. Res Immunol 143:385–400, 1992.
Robinson MK, Babcock LS, Horn PA, Kawabata TT: Specific antibody re-sponses to subtilisin Carlsberg (alcalase) in mice: Development of anintranasal exposure model. Fundam Appl Toxicol 34:15–24, 1996.
Rodgers K, Klykken P, Jacobs J, et al: Symposium overview: Immunotox-icity of medical devices. Fundam Appl Toxicol 36:1–14, 1997.
Roitt I, Brostoff J, Male D (eds): Immunology, 3d ed. London: Mosby,1993.
Rose RN: Immunopathogenesis of autoimmune diseases, in Dean JH, Lus-ter MI, Munson AE, Kimber I (eds): Immunotoxicology and Im-munopharmacology, 2d ed. New York: Raven Press, 1994, pp 513–522.
Rosenthal GJ, Kowolenko M: Immunotoxicologic manifestations of AIDStherapeutics, in Dean JH, Luster MI, Munson AE, Kimber I (eds): Im-munotoxicology and Immunopharmacology, 2d ed. New York: RavenPress, 1994, pp 227–247.
Ruddle NH: Tumor necrosis factor (TNFa) and lymphotoxin (TFNb). CurrOpin Immunol 4:327–332, 1994.
Sabatini DM, Erdjument-Bromage H, Lui M, et al: A mammalian proteinthat binds to FKBP12 in a rapamycin-dependent fashion and is ho-mologous to yeast TORs. Cell 78:35–43, 1994.
Sabers CJ, Martin MM, Brunn GJ, et al: Isolation of a protein target of theFKBP12–rapamycin complex in mammalian cells. J Biol Chem270(2):815–822, 1995.
Salem ML, Hossain MS, Nomoto K: Mediation of the immunomodulatoryeffect of b-estradiol on inflammatory responses by inhibition of re-cruitment and activation of inflammatory cells and their gene expres-sion of TNF� and IFN�. Int Arch Allergy Immunol 121(3):235–245,2000.
Sanders VM: Neuroimmunology, in Sipes IG, McQueen CA, and GandolfiAJ (eds-in-chief): Comprehensive Toxicology. Vol 5. Lawrence DA(ed): Toxicology of the Immune System. New York: Elsevier, 1997, pp175–199.
Sanders VM, Tucker AN, White KL Jr, et al: Humoral and cell-mediatedimmune status in mice exposed to trichloroethylene in the drinkingwater. Toxicol Appl Pharmacol 62:358–368, 1982.
Sanders VM, White KL Jr, Shopp GM, Munson AE: Humoral and cell-mediated immune status of mice exposed to 1,1,2-trichloroethane.Drug Chem Toxicol 8(5):357–372, 1985.
Sapin C, Mandet C, Druet E, et al: Immune complex type disease inducedby HgCl2: Genetic control of susceptibility. Transplant Proc 13:1404–1406, 1981.
Sarlo K, Karol MH: Guinea pig predictive tests for respiratory allergy, inDean JH, Luster MI, Munson AE, Kimber I (eds): Immunotoxicologyand Immunopharmacology, 2d ed. New York: Raven Press, 1994, pp703–720.
Schatz AR, Kessler FK, Kaminski NE: Inhibition of adenylate cyclase byD9-tetrahydrocannabinol in mouse spleen cells: A potential mecha-nism for cannabinoid-mediated immunosuppression. Life Sci 51:25–30, 1992.
Schwarz A, Grabbe S, Reimann H, et al: In vivo effects of interleukin-10on contact hypersensitivity and delayed-type hypersensitivity reac-tions. J Invest Dematol 103:211–216, 1994.
Selgrade MJK: Use of immunotoxicity data in health risk assessments: Un-certainties and research to improve the process. Toxicology 133:59–72, 1999.
Selgrade MJK, Cooper KD, Devlin RB, et al: Symposium overview: Im-munotoxicity—bridging the gap between animal research and humanhealth effects. Fundam Appl Toxicol 24:13–32, 1995.
Selgrade MJK, Gilmour MI: Effects of gaseous air pollutants on immuneresponses and susceptibility to infectious and allergic diseases, in DeanJH, Luster MI, Munson AE, Kimber I (eds): Immunotoxicology andImmunopharmacology, 2d ed. New York: Raven Press, 1994, pp 395–411.
Sherwood R, O’Shea W, Thomas P, et al: Effects of inhalation of ethylenedichloride on pulmonary defenses of mice and rats. Toxicol Appl Phar-macol 91:491–496, 1987.
Shopp GM, Sanders VM, Munson AE: Humoral and cell-mediated immunestatus of mice exposed to trans-1,2-dichloroethylene. Drug Chem Tox-icol 8(5):393–407, 1985.
Shopp GM, White KL Jr, Holsapple MP, et al: Naphthlene toxicity in CD-1mice: General toxicology and immunotoxicology. Fundam Appl Tox-icol 4:406–419, 1984.
Sikorski EE, Kipen HM, Selner JC, et al: Roundtable summary: The ques-tion of multiple chemical sensitivity. Fundam Appl Toxicol 24:22–28,1995.
Silkworth JB, Loose LD: Cell-mediated immunity in mice fed either Aro-clor 1016 or hexachlorobenzene. Toxicol Appl Pharmacol 45:326–327, 1978.
Silkworth JB, Loose LD: PCB and HCB induced alteration of lymphocyteblastogenesis. Toxicol Appl Pharmacol 49:86, 1979.
Silver RM, Heyes MP, Maize JC, et al: Scleroderma, fasciitis andeosinophilia associated with the ingestion of tryptophan. N Engl J Med322:874–881, 1990.
Simon JC, Cruz PD, Bergstresser PR, Tigelaar RE: Low dose ultravioletB-irradiated Langerhans cells preferentially activate CD4� cells of theT helper 2 subset. J Immunol 145:2087–2094, 1990.
2996R_ch12_419-470 4/27/01 10:02 AM Page 468
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

CHAPTER 12 TOXIC RESPONSES OF THE IMMUNE SYSTEM 469
Siroki O, Institoris L, Tatar F, Desi I: Immunotoxicological investigationof SCMF, a new pyrethroid pesticide in mice. Hum Exp Toxicol13:337–343, 1994.
Smialowicz RJ, Holsapple MP: Experimental Immunotoxicology BocaRaton, FL: CRC Press, 1996.
Smialowicz RJ, Riddle MM, Lubke RW, et al: Immunotoxicity of 2-methoxyethanol following oral adminstration in Fischer 344 rats. Tox-icol Appl Pharmacol 109(3):494–506, 1991a.
Smialowicz RJ, Riddle MM, Rogers RR, et al: Evaluation of the immuno-toxicity of orally administered 2-methoxyacetic acid in Fischer 344rats. Fundam Appl Toxicol 17(4):771–781, 1991b.
Snyder CA: Organic solvents, in Dean JH, Luster MI, Munson AE, KimberI (eds): Immunotoxicology and Immunopharmacology, 2d ed. NewYork: Raven Press, 1994, pp 183–190.
Sopori ML, Goud NS, Kaplan AM: Effects of tobacco smoke on the im-mune system, in Dean JH, Luster MI, Munson AE, Kimber I (eds):Immunotoxicology and Immunopharmacology, 2d ed. New York:Raven Press, 1994, pp 413–434.
Spreafico F, Allegrucci M, Merendino A, Luini W: Chemical immunode-pressive drugs: Their action on the cells of the immune system andimmune mediators, in Dean JH, Luster MI, Munson AE, Amos H (eds):Immunotoxicology and Immunopharmacology. New York: RavenPress, 1985, pp 179–192.
Spyker-Cranmer JM, Barnett JB, Avery DL, Cranmer MF: Immunotera-tology of chlordane: Cell-mediated and humoral immune responses inadult mice exposed in utero. Toxicol Appl Pharmacol 62:402–408,1982.
Staples JE, Gasiewicz TA, Fiore NC et al: Estrogen receptor alpha is nec-essary in thymic development and estradiol-induced thymic alter-ations. J Immunol 162(8):4168–4174, 1999.
Starec M, Rouveix B, Sinet M, et al: Immune status and survival of opiate-and cocaine-treated mice infected with Friend virus. J Pharmacol ExpTher 259:745–750, 1991.
Street JC: Pesticides and the immune system, in Sharma RP (ed): Im-munologic Considerations in Toxicology. Boca Raton, FL: CRC Press,1981, pp 46–66.
Sutter TR, Greenlee WF: Classification of members of the Ah gene bat-tery. Chemosphere 25:223, 1992.
Sutter TR, Guzman K, Dold KM, Greenlee WF: Targets for dioxin: Genesfor plasminogen activator inhibitor-2 and interleukin-1b. Science254:415, 1991.
Talmadge JE, Dean JH: Immunopharmacology of recombinant cytokines,in Dean JH, Luster MI, Munson AE, Kimber I (eds): Immunotoxicol-ogy and Immunopharmacology, 2d ed. New York: Raven Press, 1994,pp 227–247.
Terada N, Lucas JJ, Szepesi A, et al: Rapamycin blocks cell cycle pro-gression of activated T cells prior to events characteristic of the mid-dle to late G1 phase of the cycle. J Cell Physiol 154:7–15, 1993.
Thomas PT, Busse WW, Kerkvliet NI, et al: Immunologic effects of pesti-cides, in Baker SR, Wilkinson CF (eds): The Effects of Pesticides onHuman Health. Vol 18. New York: Princeton Scientific Publishers,1990, pp 261–295.
Thorne PS, Hawk C, Kaliszewski SD, Guiney PD: The noninvasive mouseear swelling assay. I. Refinements for detecting weak contact sensi-tizers. Fundam Appl Toxicol 17:790–806, 1991.
Thurmond LM, Dean JH: Immunological responses following inhalationexposure to chemical hazards, in Gardner EE, Crapo JD, Massaro EJ(eds): Toxicology of the Lung. Raven Press, New York, 1988, pp 375–406.
Tognoni G, Boniccorsi A: Epidemiological problems with TCDD. A criti-cal review. Drug Metab Rev 13:447–469, 1982.
Totoritis MC, Tan EM, McNally EM, Rubin RL: Association of antibodyto histone complex H2A-H2B with symptomatic procainamide-induced lupus. N Engl J Med 318(22):1431–1436, 1988.
Tryphonas H, Luster MI, Schiffman G, et al: Effect of chronic exposure ofPCB (Aroclor 1254) on specific and nonspecific immune parameters
in the rhesus (Macaca mulatta) monkey. Fundam Appl Toxicol 16:773–786, 1991a.
Tryphonas H, Luster MI, White KL Jr, et al: Effect of chronic exposure ofPCB (Aroclor 1254) on specific and nonspecific immune parametersin the rhesus (Macaca mulatta) monkey. Int J Immunopharmacol13:639–648, 1991b.
Tubaro E, Santiangeli C, Belogi L, et al: Methadone vs morphine: Com-parison of their effect on phagocytic functions. Int J Immunopharma-col 9:79–88, 1987.
Unanue E: Macrophages, antigen-presenting cells, and the phenomena ofantigen handling and presentation, in Paul WE (ed): Fundamental Im-munology, 3d ed. New York: Raven Press, 1993, pp 111–144.
Van Loveren H, Goettsch W, Slob W, Garssen J: Risk assessment of theharmful effects of UVB radiation on the immunological resistance toinfectious diseases. Arch Toxicol 18:21–28, 1995.
Van Loveren H, Krajnc E, Rombout PJA, et al: Effects of ozone, hexa-chlorobenzene and bis(tri-n-butyltin)oxide on natural killer activity inthe rat lung. Toxicol Appl Pharmacol 102:21–33, 1990.
Vecchi A, Sironi M, Canegrati MA, et al: Immunosuppressive effects of2,3,7,8-tetrachlorodibenzo-p-dioxin in strains of mice with differentsusceptibility to induction of aryl hydrocarbon hydroxylase. ToxicolAppl Pharmacol 68:434–441, 1983.
Voccia I, Blakley B, Brousseau P, Fournier M: Immunotoxicity of pesti-cides: A review. Toxicol Ind Health 15:119–132, 1999.
Vohr HW, Bomhard EM: Investigations on the immunostimulating poten-tial of a combination of octamethylcyclotetra and decamethylcy-cloptentasiloxane in rats and mice. Toxicol Sci Suppl 54(1):A734,2000.
Vos JG: Immune suppression as related to toxicology. CRC Crit Rev Tox-icol 5:67–101, 1977.
Vos JG, de Klerk A, Krajnc FI, et al: Toxicity of bis(tri-n-butyltin)oxide inthe rat. II. Suppression of thymus-dependent immune responses andof parameters of non-specific resistance after short-term exposure. Tox-icol Appl Pharmacol 75:387–408, 1984.
Vos JG, Luster MI: Immune alterations, in Kimbrough RD, Jensen AA(eds): Halogenated Biphenyls, Terphenyls, Napthalenes, Dibenzodi-oxins, and Related Products. Amsterdam: Elsevier, 1989, pp 295–322.
Vos JG, Smialowicz RJ, Van Loveren H: Animal models for assessment, inDean JH, Luster MI, Munson AE, Kimber I (eds): Immunotoxicologyand Immunopharmacology, 2d ed. New York: Raven Press, 1994, pp19–30.
Walder BK: Do solvents cause scleroderma? Int J Dermatol 22:157–158,1983.
Ward AM, Udnoon S, Watkins J, et al: Immunological mechanisms in thepathogenesis of vinyl chloride disease. Br Med J 1:936–938, 1976.
Ward EC, Murray MJ, Dean JH: Immunotoxicity of non-halogenated poly-cyclic aromatic hydrocarbons, in Dean JH, Luster MI, Munson AE,Amos H (eds): Immunotoxicology and Immunopharmacology. NewYork: Raven Press, 1985, pp 291–303.
Warheit DB, Hesterberg TW: Asbestos and other fibers in the lung, in DeanJH, Luster MI, Munson AE, Kimber I (eds): Immunotoxicology andImmunopharmacology, 2d ed. New York: Raven Press, 1994, pp 363–376.
Warner GL, Haggerty HG: Immunotoxicology of recombinant DNA-derived therapeutic proteins, in Sipes IG, McQueen CA, Gandolfi AJ(eds-in-chief): Comprehensive Toxicology. Vol 5. Lawrence DA (ed):Toxicology of the Immune System. New York: Elsevier, 1997, pp 435–450.
Wasserman M, Wasserman D, Gershon Z, Zellermayer L: Effects oforganochlorine insecticides on body defense systems. Ann NY AcadSci 160:393–401, 1969.
Watson ES, Murphy JC, El Sohly HN, et al: Effect of the administrationof coca alkaloids on the primary immune responses of mice: Interac-tion with D9-tetrahydrocannabinol and ethanol. Toxicol Appl Phar-macol 71:1–13, 1983.
Ways SC, Blair PB, Bern HA, Staskawicz MO: Immune responsiveness of
2996R_ch12_419-470 4/27/01 10:02 AM Page 469
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com

470 UNIT 4 TARGET ORGAN TOXICITY
adult mice exposed neonatally to diethylstilbestrol, steroid hormones,or vitamin A. J Environ Pathol Toxicol 3:207–220, 1980.
Weigent DA, Blalock JE: Associations between the neuroendocrine and im-mune systems. J Leukocyte Biol 58:137–150, 1995.
Wester PW, Vethaak AD, van Muiswinkel WB: Fish as biomarkers in im-munotoxicology. Toxicology 86:213–232, 1994.
White KL Jr: Specific immune function assays, in Miller K, Turk JL, Nick-lin S (eds): Principles and Practice of Immunotoxicology. Boston:Blackwell, 1992, pp 304–323.
White KL Jr, Anderson AC: Suppression of mouse complement activity bycontaminants of technical grade pentachlorophenol. Agents Actions16(5):385, 1985.
White KL Jr, Kawabata TT, Ladics GS: Mechanisms of polycyclic aromatichydrocarbon immunotoxicity, in Dean JH, Luster MI, Munson AE,Kimber I (eds): Immunotoxicology and Immunopharmacology, 2d ed.New York: Raven Press, 1994, pp 123–142.
White KL Jr, Lysy HH, McCay JA, Anderson AC: Modulation of serumcomplement levels following exposure to polychlorinated dibenzo-p-dioxins. Toxicol Appl Pharmacol 84:209–219, 1986.
Williams WC, Riddle MM, Copeland CB, et al: Immunological effects of2-methoxyethanol administered dermally or orally to Fischer 344 rats.Toxicology 98(1–3):215–223, 1995.
Wilson SD, LeVier DG, Butterworth LF, Munson AE: Natural killer cell
activity is enhanced following exposure to octamethylcyclotetrasilox-ane (D4). Fund Appl Toxicol Suppl 15(1):A1198, 1995.
Woolhiser MR, Galbraith TW, Mast RW, Klykken PC: A comparative hu-moral adjuvancy study of low molecular weight cyclosiloxanes (D4,D5, D6, and D7) in rats. Fund Appl Toxicol Suppl 15(1):A1197, 1995.
Xiao F, Chong AS-F, Bartlett RR, Williams JW: Leflunomide: A promis-ing immunosuppressant in transplantation, in Thomas AW, Starzyl TE(eds): Immunosuppressive Drugs: Developments in Anti-RejectionTherapy. Boston: Edward Arnold, 1994, pp 203–212.
Zelikoff JT: Biomarkers of immunotoxicity in fish and other non-mammalian sentinel species: Predictive value for mammals? Toxicol-ogy 129(1):63–71, 1998.
Zelikoff JT: Fish immunotoxicology. in Dean JH, Luster MI, Munson AE,Kimber I (eds): Immunotoxicology and Immunopharmacology, 2d ed.New York: Raven Press, 1994, pp 71–95.
Zelikoff JT, Thomas P (eds): Immunotoxicology of Environmental and Oc-cupational Metals. Bristol, PA: Taylor & Francis, 1998.
Zelikoff JT, Raymond A, Carlson E, et al: Biomarkers of immunotoxicityin fish: From the lab to the ocean. Toxicol Lett 112–113:325–331,2000.
Zurawski G, de Vries JE: Interleukin 13, an interleukin 4-like cytokine thatacts on monocytes and B cells, but not T cells. Immunol Today15(1):19–26, 1994.
2996R_ch12_419-470 4/27/01 10:02 AM Page 470
Copy
right
ed M
ater
ial
Copyright © 2001 by The McGraw-Hill Companies Retrieved from: www.knovel.com
Related Documents











