RNA-binding protein regulates plant DNA methylation by controlling mRNA processing at the intronic heterochromatin-containing gene IBM1 Xingang Wang a,1 , Cheng-Guo Duan a,1 , Kai Tang a , Bangshing Wang a , Huiming Zhang a , Mingguang Lei a , Kun Lu a,b , Satendra K. Mangrauthia a,c , Pengcheng Wang a , Guohui Zhu a,d , Yang Zhao a,e , and Jian-Kang Zhu a,e,2 a Department of Horticulture and Landscape Architecture, Purdue University, West Lafayette, IN 47907; b College of Agronomy and Biotechnology, Southwest University, Chongqing 400715, China; c Biotechnology Section, Directorate of Rice Research, Hyderabad 500030, India; d Guangdong Provincial Key Laboratory of Protein Function and Regulation in Agricultural Organisms, College of Life Sciences, South China Agricultural University, Guangzhou 510642, China; and e Shanghai Center for Plant Stress Biology, Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences, Shanghai 200032, China Contributed by Jian-Kang Zhu, August 14, 2013 (sent for review July 7, 2013) DNA methylation-dependent heterochromatin formation is a con- served mechanism of epigenetic silencing of transposons and other repeat elements in many higher eukaryotes. Genes adjacent to repetitive elements are often also subjected to this epigenetic silencing. Consequently, plants have evolved antisilencing mecha- nisms such as active DNA demethylation mediated by the REPRESSOR OF SILENCING 1 (ROS1) family of 5-methylcytosine DNA glycosylases to protect these genes from silencing. Some transposons and other repeat elements have found residence in the introns of genes. It is unclear how these intronic repeat elements-containing genes are regulated. We report here the identification of ANTI-SILENCING 1 (ASI1), a bromo-adjacent homology domain and RNA recognition motif-containing protein, from a forward genetic screen for cellular antisilencing factors in Arabidopsis thaliana. ASI1 is required to pre- vent promoter DNA hypermethylation and transcriptional silencing of some transgenes. Genome-wide DNA methylation analysis reveals that ASI1 has a similar role to that of the histone H3K9 demethylase INCREASE IN BONSAI METHYLATION 1 (IBM1) in preventing CHG methylation in the bodies of thousands of genes. We found that ASI1 is an RNA-binding protein and ensures the proper expression of IBM1 full-length transcript by associating with an intronic hetero- chromatic repeat element of IBM1. Through mRNA sequencing, we identified many genes containing intronic transposon elements that require ASI1 for proper expression. Our results suggest that ASI1 associates with intronic heterochromatin and binds the gene tran- scripts to promote their 3′ distal polyadenylation. The study thus reveals a unique mechanism by which higher eukaryotes deal with the collateral effect of silencing intronic repeat elements. DNA methylome | ChIP | gene expression I n higher eukaryotes including plants, DNA methylation is an important epigenetic mark that silences transposons and other repetitive elements. In Arabidopsis thaliana, DOMAINS REARRANGED METHYLASE 2 (DRM2) catalyzes de novo DNA methylation in all cytosine contexts including CG, CHG, and CHH (H represents A, T, or G) (1), through the RNA- directed DNA methylation pathway (RdDM) (2–9). At the same time, preexisting DNA methylation in plants can be pruned by enzymatic excision that is catalyzed by a subfamily of bifunc- tional DNA glycosylases represented by REPRESSOR OF SI- LENCING 1 (ROS1) and DEMETER (DME) (10–14). Following the enzymatic removal of 5-methylcytosine, the resultant single- nucleotide gap is filled with an unmodified cytosine through the DNA base excision repair pathway (15, 16). Cytosine methylation and demethylation are both tightly linked with histone modifications. Increased DNA methylation was observed in an A. thaliana mutant defective in INCREASED DNA METHYLATION 1 (IDM1), an acetyltransferase that catalyzes acetylation of histone H3 lysine 18 (H3K18) and lysine 23 (H3K23) necessary for subsequent DNA demethylation and prevention of transcriptional silencing (17). In A. thaliana, the repressive histone mark H3K9me2 is established mainly by the histone methyltransferase KRYPTONITE (KYP) (18). KYP and CHROMOMETHYLASE 3 (CMT3) bind to CHG methylation and H3K9me2, respectively, thereby forming a self-reinforcing loop of DNA methylation and H3K9me2 (4, 19). In addition, the binding of SAWADEE HOMEODOMAIN HOMOLOG 1 (SHH1) to methylated H3K9 helps recruit the DNA-DEPENDENT RNA POLYMERASE IV (Pol IV) for generation of heterochromatic siRNAs that mediate RdDM (20). Accumulation of H3K9 meth- ylation is negatively regulated by the histone demethylase IBM1, the mutation of which results in elevated levels of H3K9me2 and con- comitant CHG hypermethylation in thousands of genic loci (21, 22). Although DNA methylation at promoters and intergenic transposon elements (TEs) generally correlates with transcrip- tional repression, the role of gene body methylation is mostly unclear. On a whole-genome scale, A. thaliana and human cy- tosine methylation show a preference for nucleosome-bound DNA over flanking sequences (23), indicating an interplay be- tween DNA methylation and RNA biogenesis because nucleo- somes are highly enriched on exons and are preferentially positioned at intron–exon and exon–intron boundaries (23). It has been proposed that exonic cytosine methylation may serve as Significance Transposons in higher eukaryotes are subject to epigenetic silencing. Some transposons have found residence in introns of genes. It is unclear how intronic transposon-containing genes are regulated. This paper reports a cellular factor, ANTI- SILENCING 1 (ASI1), which is required for proper expression of intronic transposon-containing genes. ASI1 plays an impor- tant role in regulating genome DNA methylation patterns. The work represents a major advancement in cellular anti- silencing mechanisms. A previously undescribed mechanism for higher eukaryotes to cope with the collateral effects of silencing intronic transposon elements is discovered. This work has implications beyond the plant epigenetics field be- cause studies in mammalian systems have suggested important roles of intronic heterochromatin on 3′ distal polyadenylation, although the underlying mechanism is not known. Author contributions: X.W., C.-G.D., and J.-K.Z. designed research; X.W., C.-G.D., B.W., M.L., K.L., S.K.M., P.W., G.Z., and Y.Z. performed research; K.T. analyzed data; and C.-G.D., H.Z., and J.-K.Z. wrote the paper. The authors declare no conflict of interest. Data deposition: The raw data of mRNA sequencing of asi1-1, WT, and bisulfite sequenc- ing of asi1-1; and the bisulfite sequencing of ibm1-4 have been deposited in the Gene Expression Omnibus (GEO) database, www.ncbi.nlm.nih.gov/geo (accession nos. GSE48026 and GSE48053). 1 X.W. and C.-G.D contributed equally to this work. 2 To whom correspondence should be addressed. E-mail: [email protected]. This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10. 1073/pnas.1315399110/-/DCSupplemental. www.pnas.org/cgi/doi/10.1073/pnas.1315399110 PNAS Early Edition | 1 of 6 PLANT BIOLOGY

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

RNA-binding protein regulates plant DNA methylationby controlling mRNA processing at the intronicheterochromatin-containing gene IBM1Xingang Wanga,1, Cheng-Guo Duana,1, Kai Tanga, Bangshing Wanga, Huiming Zhanga, Mingguang Leia, Kun Lua,b,Satendra K. Mangrauthiaa,c, Pengcheng Wanga, Guohui Zhua,d, Yang Zhaoa,e, and Jian-Kang Zhua,e,2
aDepartment of Horticulture and Landscape Architecture, Purdue University, West Lafayette, IN 47907; bCollege of Agronomy and Biotechnology, SouthwestUniversity, Chongqing 400715, China; cBiotechnology Section, Directorate of Rice Research, Hyderabad 500030, India; dGuangdong Provincial Key Laboratoryof Protein Function and Regulation in Agricultural Organisms, College of Life Sciences, South China Agricultural University, Guangzhou 510642, China; andeShanghai Center for Plant Stress Biology, Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences, Shanghai 200032, China
Contributed by Jian-Kang Zhu, August 14, 2013 (sent for review July 7, 2013)
DNA methylation-dependent heterochromatin formation is a con-served mechanism of epigenetic silencing of transposons andother repeat elements in many higher eukaryotes. Genes adjacentto repetitive elements are often also subjected to this epigeneticsilencing. Consequently, plants have evolved antisilencing mecha-nisms such as active DNA demethylationmediated by the REPRESSOROF SILENCING 1 (ROS1) family of 5-methylcytosine DNA glycosylasesto protect these genes from silencing. Some transposons and otherrepeat elements have found residence in the introns of genes. It isunclear how these intronic repeat elements-containing genes areregulated. We report here the identification of ANTI-SILENCING 1(ASI1), a bromo-adjacent homology domain and RNA recognitionmotif-containing protein, from a forward genetic screen for cellularantisilencing factors in Arabidopsis thaliana. ASI1 is required to pre-vent promoter DNA hypermethylation and transcriptional silencingof some transgenes. Genome-wide DNAmethylation analysis revealsthat ASI1 has a similar role to that of the histone H3K9 demethylaseINCREASE IN BONSAI METHYLATION 1 (IBM1) in preventing CHGmethylation in the bodies of thousands of genes. We found thatASI1 is an RNA-binding protein and ensures the proper expressionof IBM1 full-length transcript by associating with an intronic hetero-chromatic repeat element of IBM1. Through mRNA sequencing, weidentified many genes containing intronic transposon elements thatrequire ASI1 for proper expression. Our results suggest that ASI1associates with intronic heterochromatin and binds the gene tran-scripts to promote their 3′ distal polyadenylation. The study thusreveals a unique mechanism by which higher eukaryotes deal withthe collateral effect of silencing intronic repeat elements.
DNA methylome | ChIP | gene expression
In higher eukaryotes including plants, DNA methylation isan important epigenetic mark that silences transposons and
other repetitive elements. In Arabidopsis thaliana, DOMAINSREARRANGED METHYLASE 2 (DRM2) catalyzes de novoDNA methylation in all cytosine contexts including CG, CHG,and CHH (H represents A, T, or G) (1), through the RNA-directed DNA methylation pathway (RdDM) (2–9). At thesame time, preexisting DNA methylation in plants can be prunedby enzymatic excision that is catalyzed by a subfamily of bifunc-tional DNA glycosylases represented by REPRESSOR OF SI-LENCING 1 (ROS1) and DEMETER (DME) (10–14). Followingthe enzymatic removal of 5-methylcytosine, the resultant single-nucleotide gap is filled with an unmodified cytosine through theDNA base excision repair pathway (15, 16).Cytosine methylation and demethylation are both tightly
linked with histone modifications. Increased DNA methylationwas observed in an A. thaliana mutant defective in INCREASEDDNA METHYLATION 1 (IDM1), an acetyltransferase thatcatalyzes acetylation of histone H3 lysine 18 (H3K18) and lysine23 (H3K23) necessary for subsequent DNA demethylation andprevention of transcriptional silencing (17). In A. thaliana, the
repressive histone mark H3K9me2 is established mainly by thehistone methyltransferase KRYPTONITE (KYP) (18). KYP andCHROMOMETHYLASE 3 (CMT3) bind to CHG methylationand H3K9me2, respectively, thereby forming a self-reinforcing loopof DNAmethylation and H3K9me2 (4, 19). In addition, the bindingof SAWADEE HOMEODOMAIN HOMOLOG 1 (SHH1) tomethylated H3K9 helps recruit the DNA-DEPENDENT RNAPOLYMERASE IV (Pol IV) for generation of heterochromaticsiRNAs that mediate RdDM (20). Accumulation of H3K9 meth-ylation is negatively regulated by the histone demethylase IBM1, themutation of which results in elevated levels of H3K9me2 and con-comitant CHG hypermethylation in thousands of genic loci (21, 22).Although DNA methylation at promoters and intergenic
transposon elements (TEs) generally correlates with transcrip-tional repression, the role of gene body methylation is mostlyunclear. On a whole-genome scale, A. thaliana and human cy-tosine methylation show a preference for nucleosome-boundDNA over flanking sequences (23), indicating an interplay be-tween DNA methylation and RNA biogenesis because nucleo-somes are highly enriched on exons and are preferentiallypositioned at intron–exon and exon–intron boundaries (23). Ithas been proposed that exonic cytosine methylation may serve as
Significance
Transposons in higher eukaryotes are subject to epigeneticsilencing. Some transposons have found residence in intronsof genes. It is unclear how intronic transposon-containinggenes are regulated. This paper reports a cellular factor, ANTI-SILENCING 1 (ASI1), which is required for proper expression ofintronic transposon-containing genes. ASI1 plays an impor-tant role in regulating genome DNA methylation patterns.The work represents a major advancement in cellular anti-silencing mechanisms. A previously undescribed mechanismfor higher eukaryotes to cope with the collateral effects ofsilencing intronic transposon elements is discovered. Thiswork has implications beyond the plant epigenetics field be-cause studies in mammalian systems have suggested importantroles of intronic heterochromatin on 3′ distal polyadenylation,although the underlying mechanism is not known.
Author contributions: X.W., C.-G.D., and J.-K.Z. designed research; X.W., C.-G.D., B.W.,M.L., K.L., S.K.M., P.W., G.Z., and Y.Z. performed research; K.T. analyzed data; and C.-G.D.,H.Z., and J.-K.Z. wrote the paper.
The authors declare no conflict of interest.
Data deposition: The raw data of mRNA sequencing of asi1-1, WT, and bisulfite sequenc-ing of asi1-1; and the bisulfite sequencing of ibm1-4 have been deposited in the GeneExpression Omnibus (GEO) database, www.ncbi.nlm.nih.gov/geo (accession nos.GSE48026 and GSE48053).1X.W. and C.-G.D contributed equally to this work.2To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1315399110/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1315399110 PNAS Early Edition | 1 of 6
PLANTBIOLO
GY

a physiological barrier that slows Pol II during transcriptionelongation, thereby influencing cotranscriptional pre-mRNAsplicing (24, 25). Indeed, a positive correlation between DNAmethylation and inclusion of alternative exons was observed inhuman and honeybee (26, 27). DNA methylation can also befound in introns and show, in some cases, positive regulation ofgene expression by unknown mechanisms. Gene expression ofEARLY GROWTHRESPONSE 2 (EGR2), a mammalian tumorsuppressor, is enhanced by intronic DNA methylation (28). Sim-ilarly, DNA methylation of an intronic repeat element was shownto be necessary for generating full-length transcripts of IBM1 inA. thaliana (29). Alternative polyadenylation was observed inmouse imprinted histocompatibility 13 (H13) and HECT ANDRLD DOMAIN CONTAINING PROTEIN 3 (Herc3) genes, inwhich differential DNA methylation in an intron dictates RNApolyadenylation sites (30, 31). It follows that intronic DNAmethylation can be a cis-element that affects alternative poly-adenylation and regulates the expression levels of functional full-length transcripts. The transacting molecular factors involved insuch epigenetic regulation, however, remain to be identified.In this study, we identified and characterized ANTI-SILENCING
1 (ASI1), a previously undescribed epigenetic regulator that isimportant for genome DNAmethylation patterns and preventionof gene silencing inA. thaliana. Through a forward genetic screen,we isolated the asi1-1 mutant that exhibited transcriptional si-lencing of reporter genes resulting from promoter DNA hyper-methylation. We found that ASI1 is necessary for preventing genebody CHG hypermethylation across the genome. Our resultsshow that a defect in the expression of IBM1 underlies the DNAhypermethylation patterns observed in the asi1-1 mutant. AtIBM1 and other genic loci that contain heterochromatic elementsin introns, ASI1 is required for the production of full-lengthtranscripts by promoting distal polyadenylation downstream of
the intronic heterochromatic elements. ASI1 associates specificallywith heterochromatic sequences in introns and exhibits RNA-binding activity in vitro. Our research has thus uncovered a trans-acting factor that is critical for the proper expression of genes withintronic heterochromatic elements. These results increase ourunderstanding of the mechanisms used by higher eukaryotes todeal with the collateral effect of silencing intronic repeat elements.
ResultsASI1 Is an Antisilencing Factor. To identify antisilencing factorsrequired for prevention of transcriptional gene silencing, wedeveloped a unique transgene-based forward genetic screen inA. thaliana. In this system, the SUCROSE-PROTON SYM-PORTER 2 (SUC2) gene encoding a sucrose transporter isoverexpressed under the Cauliflower Mosaic Virus 35S promoter(32). When grown on a sucrose-containing culture medium, the35S-SUC2 transgenic plants overaccumulate sucrose because ofthe high expression of SUC2, resulting in severe inhibition ofroot growth (Fig. 1A). The transgenic plants also contain two 35Spromoter-driven marker genes, HYGROMYCIN PHOSPHO-TRANSFERASE II (HPTII) and NEOMYCIN PHOSPHO-TRANSFERASE II (NPTII). From an ethyl methanesulfonate-mutagenized 35S-SUC2 [hereinafter referred to as the wild type(WT)] population, we isolated a previously undescribed mutant,asi1-1, and mutant alleles of the known antisilencing factors ROS1and IDM1 (10, 17), which show a derepression of root growth andsilencing of the 35S-SUC2, 35S-HPTII, and 35S-NPTII transgenes(Fig. 1B). Map-based cloning revealed that asi1-1 is a recessivemutant allele of AT5G11470 that encodes a bromo-adjacenthomology (BAH) domain and an RNA recognition motif (RRM)-containing protein (Fig. 1C). The asi1-1 mutation changed gluta-mine-321 to a premature stop codon (Fig. 1 and Fig. S1). In-troduction of WT AT5G11470 genomic DNA rescued the short
Fig. 1. Characterization of the transgene si-lencing phenotypes of the asi1-1 mutant. (A)The root growth phenotype of asi1-1, ros1-13,and idm1-9 on 1/2 MS media supplementedwith 1% sucrose or 1% glucose (Top andMiddle) and of two complemented transgeniclines on medium containing 1% sucrose (Bot-tom). (B) Relative expression levels of SUC2,NPTII, and HPTII in the WT, asi1-1, ros1-13, andidm1-9. (C) Diagram of ASI1 (AT5G11470) pro-tein domains. The red asterisk indicates theposition of the asi1-1 mutation. (D) Relativeexpression of SUC2, NPTII, and HPTII in the WT,asi1-1, and FLAG-ASI1/asi1-1 complementationplants treated with DMSO (negative control) or5′Aza. (E) A snapshot of IGB (33) shows themethylation level of the 35S promoter. MethylCsequencing data were mapped to the 35S-SUC2transgene sequence to retrieve methylation in-formation.
2 of 6 | www.pnas.org/cgi/doi/10.1073/pnas.1315399110 Wang et al.

root and transgene silencing defects of the asi1-1 mutant (Fig. 1Aand Fig. S2), suggesting that ASI1 is a previously undescribedantisilencing factor.To test whether DNA methylation is associated with transgene
silencing, we treated the asi1-1 mutant, the WT, and comple-mentation plants with 5-Aza-2′-deoxycytidine (5′Aza), a DNAmethylation inhibitor. We found that the silencing of transgenesin asi1-1 was suppressed by 5′Aza treatment (Fig. 1D), suggestingthat the silencing in asi1-1 was associated with DNA methylation.We then determined the DNA methylation levels of the 35Spromoter in asi1-1 by genome bisulfite sequencing. The datashowed clear increases in CHG (H is A, T, or C) and CHHmethylation in the 35S promoter in asi1-1 (Fig. 1E). Theseresults suggested that the transgene silencing in asi1-1 wascaused by hypermethylation of promoter DNA. Real-time PCRassays showed that the expression of ROS1 and IDM1 was notreduced in asi1-1 (Fig. S3), indicating that ASI1 may be a pre-viously undescribed regulator of active DNA demethylation.
ASI1 Prevents CHG Hypermethylation in Gene Bodies Like IBM1.Based on the whole-genome bisulfite sequencing data, we found3,767 differentially methylated regions (DMRs) in asi1-1 mutantplants, among which 3,272 DMRs are hypermethylated and 495are hypomethylated (Fig. 2A and Fig. S4). We compared theDMRs in asi1-1 with those in rdd (ros1, dml2, and dml3 triplemutant) and ibm1-4. ROS1, DEMETER-LIKE 2 (DML2), andDML3 are 5-methylcytosine DNA glycosylases required for pro-tecting transposon elements (TEs) and other repeat sequencesfrom hypermethylation (12–14). IBM1 is a histone H3K9 deme-thylase that protects genes from CHG hypermethylation by CMT3(21). As expected, most of the hyper-DMRs in rdd correspondedto TEs and intergenic sequences rather than to genes. However,most of the hyper-DMRs in asi1-1 were mapped to genes, whichwas very similar to the case with ibm1-4 (Fig. 2A). Approximately81% (2,652/3,272) of the hyper-DMRs in asi1-1 overlapped withthose in ibm1-4, whereas only 10% overlapped with those in rdd(Fig. 2A and Fig. S4). In DMRs that overlapped between asi1-1and ibm1-4, CHG methylation was dramatically increased (Fig. 2B and C). In contrast, the DMRs that were shared by asi1-1 andrdd or by asi1-1, ibm1, and rdd showed increases not only in CHGmethylation but also in CG and CHH methylation (Fig. 2B).To further characterize the DNA methylation profiles, we
analyzed the distribution of DNA methylation along the genebodies, TEs, and their 2-kb upstream and downstream flankingsequences in the three mutants. There was a significant increasein CHG methylation and a slight increase in CHH methylationbut no change in CG methylation in gene bodies in asi1-1 and inibm1-4 (Fig. 2 B–D). In contrast, the CHG and CHH methyla-tion of TEs was slightly reduced in asi1-1 and ibm1-4mutants butnot in the rdd mutant (Fig. S5). Like the genic CHG hyper-methylation in ibm1-4, the genic CHG hypermethylation in asi1-1was strongly associated with gene length. Longer genes were moreheavily hypermethylated than shorter ones in asi1-1 and ibm1-4(Fig. S6). These results suggest that ASI1 may act in the samepathway as IBM1 in preventing genic CHG methylation.
ASI1 Controls the Expression of Full-Length IBM1 Transcript. IBM1 isknown to produce two transcripts (29), a long form (IBM1-L)containing the jmjC histone demethylase domain and a shortform (IBM1-S) lacking the domain (Fig. 3A). Real-time RT-PCRassays showed that the IBM1-L transcript was substantially re-duced in asi1-1 compared with the WT, whereas the IBM1-Stranscript was slightly increased (Fig. 3B). The total amount ofIBM1 transcripts (IBM1-5′) containing both IBM1-L and IBM1-Swas slightly reduced in asi1-1 (Fig. 3B). Northern analysis sup-ported these conclusions (Fig. 3C). Consistent with these find-ings, mRNA sequencing (mRNA-seq) experiments showed thatthe expression of the IBM1-3′ region was reduced (Fig. 3D). The3′Rapid Amplification of cDNA Ends (RACE) assay revealedthe presence of two major polyadenylated IBM1 transcripts (Fig.3D). Polyadenylation occurred predominantly at the 3′ distal endin the WT but mainly in the seventh intron of the IBM1 gene in
the asi1-1 mutant (Fig. 3D). These results showed that ASI1 isrequired for accumulation of the IBM1-L transcript.The IBM1 gene contains a very large (1,974-bp) seventh intron,
in which a ∼500-bp repeat sequence (referred to as IBM1-IH) ismethylated and enriched in H3K9 dimethylation (Fig. 3D) (29).The accumulation of IBM1-L transcript depends on CG and CHGmethylation of this region (29). Our whole-genome bisulfite se-quencing confirmed that the IBM-IH region was indeed heavilymethylated and that the asi1-1 mutation did not affect thismethylation (Fig. 3D). To determine the potential importance ofIBM1-IH in ASI1 regulation of IBM1 expression, we introduceda WT IBM1 genomic DNA clone driven by its native promoter(pIBM1::IBM1) as well as a corresponding construct lacking onlythe IBM1-IH region (pIBM1::IBM1ΔIH) into the asi1-1 mutant.The results showed that IBM1-L expression was restored to theWT level in asi1-1 by pIBM1::IBM1ΔIH but not by pIBM1::IBM
Fig. 2. Whole-genome methylation analysis of the asi1-1 mutant and com-parison of the asi1-1 mutant with the ibm1-4 and rdd mutants. (A) Analysis ofhyper-DMRs. (Left) Composition of the hyper-DMRs in asi1-1, ibm1-4, and rddmutants. Gene, DMRs in protein-codinggenesbut not TEs; IG, intergenic regions;Other, DMRs that overlapwith other features like pseudogenes, miRNAs, rRNAs,etc.; TE, DMRs in TEs but not protein-coding genes; TE +Gene, DMRs in both TEsand protein-coding genes. (Right) Numbers of hyper-DMRs that are overlappingamong or unique to the asi1-1, ibm1-4, and rdd mutants. In a few cases whereone region in one mutant overlaps with two regions in another mutant, wecalculated the number of overlapping regions from the perspective of asi1-1,ibm1-4, and rdd, in that order. (B) Box plots displaying the distribution of aver-age CG, CHG, or CHH methylation levels calculated for hyper-DMRs in the in-dicated subgroups. (C) Snapshots of two representative DNA hypermethylationregions that show dramatic increases in CHGmethylation and slight increases inCHH methylation in asi1-1 and ibm1-4. (D) Average methylation levels in genebodies and flanking 2-kb regions. Each gene was aligned from start to end anddivided into 20 equal bins. Upstream and downstream 2-kb regions were eachalso divided into 20 equal bins. Weighted methylation level was calculated foreach of the 60 bins across all of the corresponding regions and plotted. Onlycytosines with fourfold or more coverage were used for this analysis.
Wang et al. PNAS Early Edition | 3 of 6
PLANTBIOLO
GY

(Fig. 3E). DNA methylation-sensitive PCR assays showed thatCHG hypermethylation in tested loci, including BONSAI, wassuppressed in pIBM1::IBM1ΔIH but not in pIBM1::IBM1 trans-formants (Fig. 3F). The transgene silencing and root growth pheno-types of asi1-1 were also suppressed in pIBM1::IBM1ΔIH but not inpIBM1::IBM1 transformants (Fig. 3E). Furthermore, developmentaldefects of asi1-1 such as dwarf stature and short siliques, which werealso observed in ibm1 mutant plants, were rescued by expression ofpIBM1::IBM1ΔIH (Fig. S7). These results demonstrated that thefunction of ASI1 in preventing transgene silencing and CHG hyper-methylation is explained by its regulation of IBM1 and that this reg-ulation requires the intronic heterochromatin region.
ASI1 Controls the Alternative Processing of Intronic Heterochromatin-Containing Genes by Promoting 3′ Distal Polyadenylation. To in-vestigate whether ASI1 plays a general role in regulating geneswith a large and heterochromatic intron like that in IBM1, wesearched and found 683 genes with intronic TEs; 45 of thempossibly had a reduced expression in the 3′ region downstream ofthe TEs according to our cutoff (Table S1). The three candidategenes with the largest intronic TEs were selected for further analysis(Fig. 4 and Table S1). Our methylome data indicated the presence
of a heavily methylated TE in a large intron in two of these threegenes:COPIA8B inAT1G58602 andATLINE2 inAT3G05410 (Fig.4). The third gene, AT1G11270, contains a COPIA78 family TE ina large intron, and the TE is methylated near its two ends (Fig. 4).For all three of these genes, Northern blot and RT-qPCR analysesshowed that there were lower levels of full-length transcripts with 3′distal polyadenylation in asi1-1 than in the WT. The 3′ RACEassays showed the presence of multiple shorter transcripts withpolyadenylation inside the TE or broader heterochromatic regionfor the three candidates and the absence of full-length transcriptswith 3′ distal polyadenylation for two of the three genes in the asi1-1mutant (Fig. 4). These results suggest that ASI1 is required for theproper expression of genes with heterochromatic introns.
ASI1 Is Enriched in the Intronic Heterochromatin Region of TargetGenes. To investigate the underlying mechanism of ASI1-medi-ated mRNA processing of intronic heterochromatin-containinggenes, we first performed chromatin immunoprecipitation (ChIP)assays to determine whether ASI1 affects Pol II elongationthrough the heterochromatic introns. The results show thatthere were no substantial differences between asi1-1 and theWT in Pol II occupancy in the heterochromatic intron or in the
Fig. 3. Regulation of IBM1 expression by ASI1.(A) Schematic representation of IBM1-L andIBM1-S and positions of primers used for quan-titative real-time PCR. (B) Relative expression ofIBM1-5′, IBM1-3′, IBM1-L, and IBM1-S in the WTand the asi1-1 mutant. (C) Detection of the longand short forms of IBM1 transcripts in WT, asi1-1,and complemented asi1-1 by Northern blot anal-ysis. (D) mRNA-seq 3′ RACE data. (Top) A snapshotof IGB (33) showing the read coverage of IBM1transcripts and methylation levels. (Bottom) The 3′RACE PCR and sequencing results. The uppercaseblue letters and lowercase orange letters in-dicate the IBM1 transcripts in the WT and asi1-1,respectively. The green box in the IBM1 genestructure indicates the intronic heterochromatinregion (IH). (E) Root growth and relative ex-pression of SUC2 and IBM1-L transcript in theWT, asi1-1, 3FLAG-ASI1/asi1-1, IBM1Δ1H/asi1-1,and IBM1/asi1-1. (F) DNA methylation-sensitivePCR analysis of the WT, asi1-1, FLAG-ASI1/asi1-1,IBM1Δ1H/asi1-1, and IBM1/asi1-1 at the testedloci including BONSAI (BNS).
Fig. 4. Regulation of the expression of AT3G05410, AT1G11270, and AT1G58602 by ASI1. (Top) mRNA-seq data showing reduced transcript levels at the3′ regions, genome bisulfite sequencing data showing DNA methylation levels, gene structures, and 3′ RACE results of (A) AT3G05410, (B) AT1G11270, and (C)AT1G58602. The green boxes indicate intronic TEs. (Bottom) Photographs of 3′ RACE PCR product gels, and real-time RT-PCR analysis of 5′ and 3′ transcriptlevels, and Northern blot analysis of the alternative transcripts in the WT, asi1-1, and complemented asi1-1.
4 of 6 | www.pnas.org/cgi/doi/10.1073/pnas.1315399110 Wang et al.
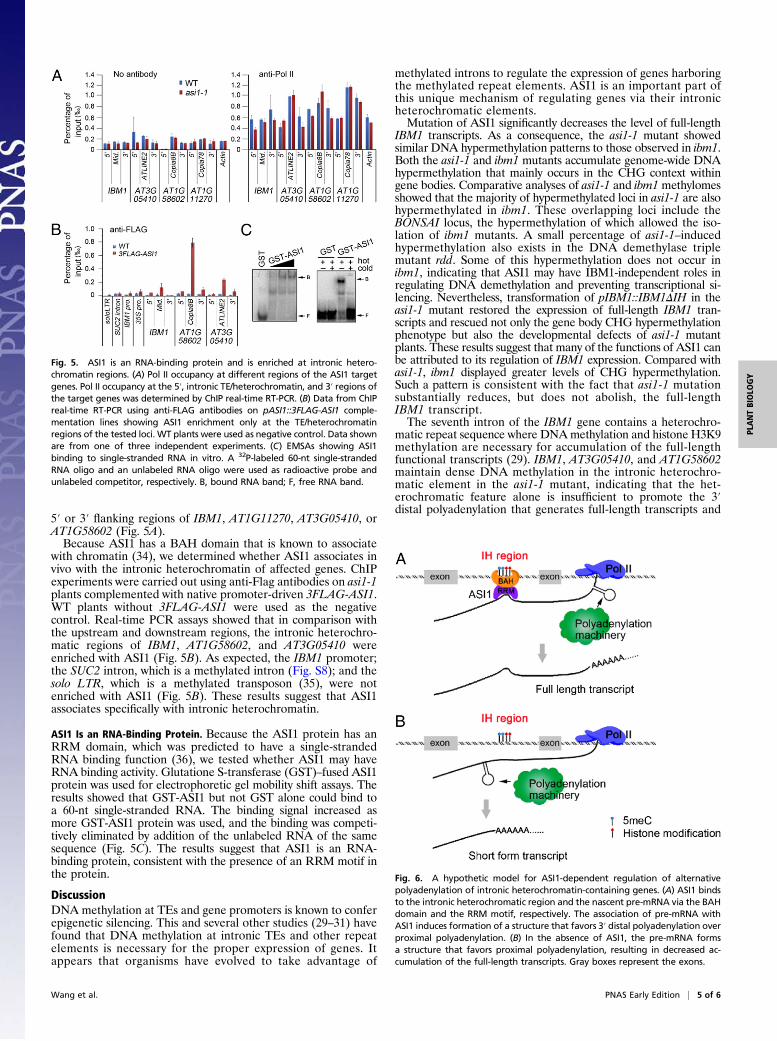
5′ or 3′ flanking regions of IBM1, AT1G11270, AT3G05410, orAT1G58602 (Fig. 5A).Because ASI1 has a BAH domain that is known to associate
with chromatin (34), we determined whether ASI1 associates invivo with the intronic heterochromatin of affected genes. ChIPexperiments were carried out using anti-Flag antibodies on asi1-1plants complemented with native promoter-driven 3FLAG-ASI1.WT plants without 3FLAG-ASI1 were used as the negativecontrol. Real-time PCR assays showed that in comparison withthe upstream and downstream regions, the intronic heterochro-matic regions of IBM1, AT1G58602, and AT3G05410 wereenriched with ASI1 (Fig. 5B). As expected, the IBM1 promoter;the SUC2 intron, which is a methylated intron (Fig. S8); and thesolo LTR, which is a methylated transposon (35), were notenriched with ASI1 (Fig. 5B). These results suggest that ASI1associates specifically with intronic heterochromatin.
ASI1 Is an RNA-Binding Protein. Because the ASI1 protein has anRRM domain, which was predicted to have a single-strandedRNA binding function (36), we tested whether ASI1 may haveRNA binding activity. Glutatione S-transferase (GST)–fused ASI1protein was used for electrophoretic gel mobility shift assays. Theresults showed that GST-ASI1 but not GST alone could bind toa 60-nt single-stranded RNA. The binding signal increased asmore GST-ASI1 protein was used, and the binding was competi-tively eliminated by addition of the unlabeled RNA of the samesequence (Fig. 5C). The results suggest that ASI1 is an RNA-binding protein, consistent with the presence of an RRM motif inthe protein.
DiscussionDNA methylation at TEs and gene promoters is known to conferepigenetic silencing. This and several other studies (29–31) havefound that DNA methylation at intronic TEs and other repeatelements is necessary for the proper expression of genes. Itappears that organisms have evolved to take advantage of
methylated introns to regulate the expression of genes harboringthe methylated repeat elements. ASI1 is an important part ofthis unique mechanism of regulating genes via their intronicheterochromatic elements.Mutation of ASI1 significantly decreases the level of full-length
IBM1 transcripts. As a consequence, the asi1-1 mutant showedsimilar DNA hypermethylation patterns to those observed in ibm1.Both the asi1-1 and ibm1 mutants accumulate genome-wide DNAhypermethylation that mainly occurs in the CHG context withingene bodies. Comparative analyses of asi1-1 and ibm1 methylomesshowed that the majority of hypermethylated loci in asi1-1 are alsohypermethylated in ibm1. These overlapping loci include theBONSAI locus, the hypermethylation of which allowed the iso-lation of ibm1 mutants. A small percentage of asi1-1–inducedhypermethylation also exists in the DNA demethylase triplemutant rdd. Some of this hypermethylation does not occur inibm1, indicating that ASI1 may have IBM1-independent roles inregulating DNA demethylation and preventing transcriptional si-lencing. Nevertheless, transformation of pIBM1::IBM1ΔIH in theasi1-1 mutant restored the expression of full-length IBM1 tran-scripts and rescued not only the gene body CHG hypermethylationphenotype but also the developmental defects of asi1-1 mutantplants. These results suggest that many of the functions of ASI1 canbe attributed to its regulation of IBM1 expression. Compared withasi1-1, ibm1 displayed greater levels of CHG hypermethylation.Such a pattern is consistent with the fact that asi1-1 mutationsubstantially reduces, but does not abolish, the full-lengthIBM1 transcript.The seventh intron of the IBM1 gene contains a heterochro-
matic repeat sequence where DNAmethylation and histone H3K9methylation are necessary for accumulation of the full-lengthfunctional transcripts (29). IBM1, AT3G05410, and AT1G58602maintain dense DNA methylation in the intronic heterochro-matic element in the asi1-1 mutant, indicating that the het-erochromatic feature alone is insufficient to promote the 3′distal polyadenylation that generates full-length transcripts and
Fig. 5. ASI1 is an RNA-binding protein and is enriched at intronic hetero-chromatin regions. (A) Pol II occupancy at different regions of the ASI1 targetgenes. Pol II occupancy at the 5′, intronic TE/heterochromatin, and 3′ regions ofthe target genes was determined by ChIP real-time RT-PCR. (B) Data from ChIPreal-time RT-PCR using anti-FLAG antibodies on pASI1::3FLAG-ASI1 comple-mentation lines showing ASI1 enrichment only at the TE/heterochromatinregions of the tested loci. WT plants were used as negative control. Data shownare from one of three independent experiments. (C) EMSAs showing ASI1binding to single-stranded RNA in vitro. A 32P-labeled 60-nt single-strandedRNA oligo and an unlabeled RNA oligo were used as radioactive probe andunlabeled competitor, respectively. B, bound RNA band; F, free RNA band.
Fig. 6. A hypothetic model for ASI1-dependent regulation of alternativepolyadenylation of intronic heterochromatin-containing genes. (A) ASI1 bindsto the intronic heterochromatic region and the nascent pre-mRNA via the BAHdomain and the RRM motif, respectively. The association of pre-mRNA withASI1 induces formation of a structure that favors 3′ distal polyadenylation overproximal polyadenylation. (B) In the absence of ASI1, the pre-mRNA formsa structure that favors proximal polyadenylation, resulting in decreased ac-cumulation of the full-length transcripts. Gray boxes represent the exons.
Wang et al. PNAS Early Edition | 5 of 6
PLANTBIOLO
GY

that the transacting factor ASI1 is necessary to ensure the pro-duction of full-length transcripts.At IBM1 and several other genes with intronic heterochro-
matic elements, ChIP assays showed that occupancy of the elon-gating Pol II is not affected by the asi1-1mutation, suggesting thattranscription elongation is independent of ASI1 function. In ad-dition, Pol II does not seem to exhibit greater occupancy at theintronic heterochromatic regions than at their corresponding5′ and 3′ portions, indicating that ASI1 function does not involvealteration in transcriptional elongation kinetics. mRNA-seq andindividual 3′-RACE analyses demonstrated that ASI1 facilitatesdistal polyadenylation downstream of the intronic heterochromaticelements or inhibits proximal polyadenylation, thereby pro-moting the generation of full-length transcripts. Highly usedpolyadenylation sites are positively associated with an mRNAstructure that is energetically favorable and one that effectivelyexposes a critical polyadenylation cis-element (37). ASI1 con-tains a chromatin-binding BAH domain and an RNA-bindingRRM motif. ChIP assays revealed that ASI1 associates withheterochromatic elements that reside within introns but notwithin gene promoters or intergenic regions, although it remainsunclear how such specificity is achieved. One possibility is thatunlike heterochromatic elements in gene promoters or inter-genic regions, intronic heterochromatic elements are activelytranscribed and thus must have various active chromatin marksin addition to their heterochromatic marks. Given that ASI1 alsoshowed RNA-binding activities in vitro, we hypothesize that theintronic heterochromatin-associated ASI1 may bind to the pre-mRNAs in vivo and the RNA-binding subsequently induces anRNA structure that favors 3′ distal polyadenylation over proximalpolyadenylation. Alternatively, ASI1 may impede the formationof an RNA structure that favors proximal polyadenylation. Ineither case, ASI1 promotes distal polyadenylation that generatesfull-length transcripts (Fig. 6).
Our mRNA-seq analyses identified dozens of intronic het-erochromatin-containing genes that require ASI1 for properexpression. Other intronic heterochromatin-containing genes mayrequire the ASI1 paralog, AT3G15605, for proper expression. Forsome of these intronic heterochromatin-containing genes, proximalpolyadenylation-generated shorter transcripts could be functional,and the alternative polyadenylation may even be subjected to reg-ulation by developmental or environmental cues. It appears that thecontrol of gene expression by intronic heterochromatic elements isa common biological feature and that plants have acquired anASI1-dependent mechanism to ensure the proper expression ofthis type of genes. Alternative polyadenylation regulated by intronicDNA methylation has also been observed in the mouse imprintedgenes H13 and Herc3, in which the methylated and unmethylatedalleles use downstream and upstream polyadenylation sites, re-spectively (30, 31), supporting the conclusion that DNAmethylationplays a positive role in determining the use of polyadenylation sites.It is likely that mammalian cells require transacting factors likeASI1 for the epigenetic regulation of alternative polyadenylation.
Materials and MethodsDetails are provided in SI Materials and Methods, including plant materialsand growth conditions, map-based cloning and identification of the ASI1locus, plasmid construction and mutant complementation, DNA methylation-sensitive PCR, quantitative real-time PCR, Northern blot analysis, 3′RACE, ChIPassays, protein purification and EMSAs, whole-genome bisulfite sequencing andanalysis, and mRNA-seq data analysis. Primers used in this study are listed inTable S2. The raw data of mRNA-seq of asi1-1, WT, and bisulfite sequencing ofasi1-1 have been deposited in the GEO database (accession no. GSE48026). Thebisulfite sequencing of ibm1-4 has also been deposited in the GEO database(accession no. GSE48053).
ACKNOWLEDGMENTS. This work was supported by National Institutes ofHealth Grants R01GM070795 and R01GM059138 (to J.-K.Z.) and by theChinese Academy of Sciences.
1. Cao X, Jacobsen SE (2002) Role of the arabidopsis DRM methyltransferases in de novoDNA methylation and gene silencing. Curr Biol 12(13):1138–1144.
2. Zhang X, et al. (2006) Genome-wide high-resolution mapping and functional analysisof DNA methylation in arabidopsis. Cell 126(6):1189–1201.
3. Henderson IR, Jacobsen SE (2007) Epigenetic inheritance in plants. Nature 447(7143):418–424.
4. Law JA, Jacobsen SE (2010) Establishing, maintaining and modifying DNAmethylationpatterns in plants and animals. Nat Rev Genet 11(3):204–220.
5. Matzke M, Kanno T, Daxinger L, Huettel B, Matzke AJ (2009) RNA-mediated chro-matin-based silencing in plants. Curr Opin Cell Biol 21(3):367–376.
6. Haag JR, Pikaard CS (2011) Multisubunit RNA polymerases IV and V: Purveyors of non-coding RNA for plant gene silencing. Nat Rev Mol Cell Biol 12(8):483–492.
7. Zhang H, Zhu JK (2011) RNA-directed DNA methylation. Curr Opin Plant Biol 14(2):142–147.8. Wierzbicki AT (2012) The role of long non-coding RNA in transcriptional gene si-
lencing. Curr Opin Plant Biol 15(5):517–522.9. Pikaard CS, Haag JR, Pontes OM, Blevins T, Cocklin R (2012) A transcription fork model
for Pol IV and Pol V-dependent RNA-directed DNA methylation. Cold Spring HarbSymp Quant Biol 77:205–212.
10. Gong Z, et al. (2002) ROS1, a repressor of transcriptional gene silencing in Arabi-dopsis, encodes a DNA glycosylase/lyase. Cell 111(6):803–814.
11. Gehring M, et al. (2006) DEMETER DNA glycosylase establishes MEDEA polycombgene self-imprinting by allele-specific demethylation. Cell 124(3):495–506.
12. Agius F, Kapoor A, Zhu JK (2006) Role of the Arabidopsis DNA glycosylase/lyase ROS1in active DNA demethylation. Proc Natl Acad Sci USA 103(31):11796–11801.
13. Ortega-Galisteo AP, Morales-Ruiz T, Ariza RR, Roldán-Arjona T (2008) ArabidopsisDEMETER-LIKE proteins DML2 and DML3 are required for appropriate distribution ofDNA methylation marks. Plant Mol Biol 67(6):671–681.
14. Penterman J, et al. (2007) DNA demethylation in the Arabidopsis genome. Proc NatlAcad Sci USA 104(16):6752–6757.
15. Zhu JK (2009) Active DNA demethylation mediated by DNA glycosylases. Annu RevGenet 43:143–166.
16. Zhang H, Zhu JK (2012) Active DNA demethylation in plants and animals. Cold SpringHarb Symp Quant Biol 77:161–173.
17. Qian W, et al. (2012) A histone acetyltransferase regulates active DNA demethylationin Arabidopsis. Science 336(6087):1445–1448.
18. Liu C, Lu F, Cui X, Cao X (2010) Histone methylation in higher plants. Annu Rev PlantBiol 61:395–420.
19. Du J, et al. (2012) Dual binding of chromomethylase domains to H3K9me2-containingnucleosomes directs DNA methylation in plants. Cell 151(1):167–180.
20. Law JA, et al. (2013) Polymerase IV occupancy at RNA-directed DNA methylation sitesrequires SHH1. Nature 498(7454):385–389.
21. Saze H, Shiraishi A, Miura A, Kakutani T (2008) Control of genic DNA methylation bya jmjC domain-containing protein in Arabidopsis thaliana. Science 319(5862):462–465.
22. Miura A, et al. (2009) An Arabidopsis jmjC domain protein protects transcribed genesfrom DNA methylation at CHG sites. EMBO J 28(8):1078–1086.
23. Chodavarapu RK, et al. (2010) Relationship between nucleosome positioning andDNA methylation. Nature 466(7304):388–392.
24. Oberdoerffer S (2012) A conserved role for intragenic DNA methylation in alternativepre-mRNA splicing. Transcription 3(3):106–109.
25. Shukla S, Oberdoerffer S (2012) Co-transcriptional regulation of alternative pre-mRNA splicing. Biochim Biophys Acta 1819(7):673–683.
26. Lyko F, et al. (2010) The honey bee epigenomes: Differential methylation of brainDNA in queens and workers. PLoS Biol 8(11):e1000506.
27. Shukla S, et al. (2011) CTCF-promoted RNA polymerase II pausing links DNA methyl-ation to splicing. Nature 479(7371):74–79.
28. Unoki M, Nakamura Y (2003) Methylation at CpG islands in intron 1 of EGR2 confersenhancer-like activity. FEBS Lett 554(1–2):67–72.
29. Rigal M, Kevei Z, Pélissier T, Mathieu O (2012) DNA methylation in an intron of theIBM1 histone demethylase gene stabilizes chromatin modification patterns. EMBO J31(13):2981–2993.
30. Cowley M, Wood AJ, Böhm S, Schulz R, Oakey RJ (2012) Epigenetic control of alternativemRNA processing at the imprinted Herc3/Nap1l5 locus. Nucleic Acids Res 40(18):8917–8926.
31. Wood AJ, et al. (2008) Regulation of alternative polyadenylation by genomic im-printing. Genes Dev 22(9):1141–1146.
32. Lei M, et al. (2011) Genetic and genomic evidence that sucrose is a global regulator ofplant responses to phosphate starvation in Arabidopsis. Plant Physiol 156(3):1116–1130.
33. Nicol JW, Helt GA, Blanchard SG, Jr., Raja A, Loraine AE (2009) The Integrated Ge-nome Browser: Free software for distribution and exploration of genome-scale da-tasets. Bioinformatics 25(20):2730–2731.
34. Callebaut I, Courvalin JC, Mornon JP (1999) The BAH (bromo-adjacent homology)domain: A link between DNA methylation, replication and transcriptional regulation.FEBS Lett 446(1):189–193.
35. Huettel B, et al. (2006) Endogenous targets of RNA-directed DNA methylation and PolIV in Arabidopsis. EMBO J 25(12):2828–2836.
36. Cléry A, Blatter M, Allain FH (2008) RNA recognition motifs: Boring? Not quite. CurrOpin Struct Biol 18(3):290–298.
37. Khaladkar M, Smyda M, Hannenhalli S (2011) Epigenomic and RNA structural corre-lates of polyadenylation. RNA Biol 8(3):529–537.
6 of 6 | www.pnas.org/cgi/doi/10.1073/pnas.1315399110 Wang et al.
Related Documents











