Tohoku J. Exp. Med., 2012, 227, 13-22 13 Received February 3, 2012; revision accepted for publication April 2, 2012. doi: 10.1620/tjem.227.13 Correspondence: Akira Ando, M.D., Ph.D., Department of Orthopaedic Surgery, Tohoku University School of Medicine, 1-1 Seiryomachi, Aoba-ku, Sendai, Miyagi 980-8574, Japan. e-mail: andoakir@yahoo.co.jp Remobilization Does Not Restore Immobilization-Induced Adhesion of Capsule and Restricted Joint Motion in Rat Knee Joints Akira Ando, 1 Hideaki Suda, 1 Yoshihiro Hagiwara, 1 Yoshito Onoda, 1 Eiichi Chimoto 1 and Eiji Itoi 1 1 Department of Orthopaedic Surgery, Tohoku University School of Medicine, Sendai, Japan Joint immobilization, which is used in orthopaedic treatments and observed in bedridden people, usually causes restricted joint motion. Decreased joint motion diminishes activities of daily living and increases burden of nursing-care. The purpose of this study was to clarify the reversibility of immobilization-induced capsular changes and restricted joint motion in rat knee joints. The unilateral knee joints of adult male rats were immobilized with an internal fixator for 1, 2, 4, 8, and 16 weeks as a model of immobilization after surgery or disuse of the joint. After the fixation devices were removed, the rats were allowed to move freely for 16 weeks. Sham-operated rats were used as controls. Sagittal sections at medial midcondylar regions were made and assessed with histological, histomorphometric, and immunohistochemical methods. Joint motion was measured using a custom-made device under x-ray control after removal of the periarticular muscles. In the 1/16-week and 2/16-week immobilization-remobilization (Im-Rm) groups, cord-like structures connecting the superior and inferior portions of the posterior capsule (partial adhesion) were observed without restricted joint motion. In the 4/16-, 8/16-, and 16/16-week Im-Rm groups, global adhesion of the posterior capsule and restricted joint motion were observed. The restricted joint motion was not completely restored after incision of the posterior capsule. These data indicate that immobilization alone causes irreversible capsular changes and arthrogenic restricted joint motion. Besides the joint capsule, other arthrogenic factors such as ligaments might influence the restricted joint motion. Prolonged immobilization over 4 weeks should be avoided to prevent irreversible joint contracture. Keywords: adhesion; immobilization; joint contracture; remobilization; synovial membrane Tohoku J. Exp. Med., 2012, 227 (1), 13-22. © 2012 Tohoku University Medical Press Joint immobilization is frequently used in the treat- ment of musculoskeletal disorders for pain relief and to decrease inflammation, but it also causes unfavorable out- comes such as joint contracture (Akeson et al. 1987). Joint contracture is defined as a limitation in the active and pas- sive range of motion on the affected joints, and it causes permanent impairment and disability to the affected patients (Trudel et al. 2003). The factors responsible for joint con- tracture are mainly divided into the muscles (myogenic) and the periarticular structures (arthrogenic) (Evans et al. 1960; Enneking and Horowitz 1972; Trudel and Uhthoff 2000). Recent studies indicated that the arthrogenic factors, especially the joint capsule, played an important role in the progression of limited range of motion after immobilization (Trudel and Uhthoff 2000; Moriyama et al. 2006; Lee et al. 2010). We revealed in the previous report that the arthro- genic limitation occurred after a 4-week immobilization in a rat immobilized knee model and the restricted motion was restored after incision of the capsule, which indicated that the joint capsule was one of the most responsible factors of limited range of motion after immobilization (Chimoto et al. 2007). The joint capsule is composed of two distinct layers: an external dense fibrous layer of the capsule and internal thin synovial membrane (Roy and Ghadially 1967). Though the importance of the joint capsule to the pathogen- esis of the joint contracture is well accepted, the underlying changes within the joint capsule are not fully understood. Adhesions and shortening of the synovial membrane, atro- phy and fibrosis of the capsule are known as structural changes (Trudel and Uhthoff 2000; Trudel et al. 2000; Moriyama et al. 2007), whereas decreased glycosaminogly- cans and water content, and possibly accumulation of advanced glycation endproducts such as pentosidine are known as biochemical changes (Akeson et al. 1977; Amiel et al. 1985; Lee et al. 2010). We reported in our previous

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Reversibility of Immobilization-Induced Joint Contracture 13Tohoku J. Exp. Med., 2012, 227, 13-22
13
Received February 3, 2012; revision accepted for publication April 2, 2012. doi: 10.1620/tjem.227.13Correspondence: Akira Ando, M.D., Ph.D., Department of Orthopaedic Surgery, Tohoku University School of Medicine, 1-1
Seiryomachi, Aoba-ku, Sendai, Miyagi 980-8574, Japan.e-mail: [email protected]
Remobilization Does Not Restore Immobilization-Induced Adhesion of Capsule and Restricted Joint Motion in Rat Knee Joints
Akira Ando,1 Hideaki Suda,1 Yoshihiro Hagiwara,1 Yoshito Onoda,1 Eiichi Chimoto1 and Eiji Itoi1
1Department of Orthopaedic Surgery, Tohoku University School of Medicine, Sendai, Japan
Joint immobilization, which is used in orthopaedic treatments and observed in bedridden people, usually causes restricted joint motion. Decreased joint motion diminishes activities of daily living and increases burden of nursing-care. The purpose of this study was to clarify the reversibility of immobilization-induced capsular changes and restricted joint motion in rat knee joints. The unilateral knee joints of adult male rats were immobilized with an internal fixator for 1, 2, 4, 8, and 16 weeks as a model of immobilization after surgery or disuse of the joint. After the fixation devices were removed, the rats were allowed to move freely for 16 weeks. Sham-operated rats were used as controls. Sagittal sections at medial midcondylar regions were made and assessed with histological, histomorphometric, and immunohistochemical methods. Joint motion was measured using a custom-made device under x-ray control after removal of the periarticular muscles. In the 1/16-week and 2/16-week immobilization-remobilization (Im-Rm) groups, cord-like structures connecting the superior and inferior portions of the posterior capsule (partial adhesion) were observed without restricted joint motion. In the 4/16-, 8/16-, and 16/16-week Im-Rm groups, global adhesion of the posterior capsule and restricted joint motion were observed. The restricted joint motion was not completely restored after incision of the posterior capsule. These data indicate that immobilization alone causes irreversible capsular changes and arthrogenic restricted joint motion. Besides the joint capsule, other arthrogenic factors such as ligaments might influence the restricted joint motion. Prolonged immobilization over 4 weeks should be avoided to prevent irreversible joint contracture.
Keywords: adhesion; immobilization; joint contracture; remobilization; synovial membraneTohoku J. Exp. Med., 2012, 227 (1), 13-22. © 2012 Tohoku University Medical Press
Joint immobilization is frequently used in the treat-ment of musculoskeletal disorders for pain relief and to decrease inflammation, but it also causes unfavorable out-comes such as joint contracture (Akeson et al. 1987). Joint contracture is defined as a limitation in the active and pas-sive range of motion on the affected joints, and it causes permanent impairment and disability to the affected patients (Trudel et al. 2003). The factors responsible for joint con-tracture are mainly divided into the muscles (myogenic) and the periarticular structures (arthrogenic) (Evans et al. 1960; Enneking and Horowitz 1972; Trudel and Uhthoff 2000). Recent studies indicated that the arthrogenic factors, especially the joint capsule, played an important role in the progression of limited range of motion after immobilization (Trudel and Uhthoff 2000; Moriyama et al. 2006; Lee et al. 2010). We revealed in the previous report that the arthro-genic limitation occurred after a 4-week immobilization in a rat immobilized knee model and the restricted motion was
restored after incision of the capsule, which indicated that the joint capsule was one of the most responsible factors of limited range of motion after immobilization (Chimoto et al. 2007).
The joint capsule is composed of two distinct layers: an external dense fibrous layer of the capsule and internal thin synovial membrane (Roy and Ghadially 1967). Though the importance of the joint capsule to the pathogen-esis of the joint contracture is well accepted, the underlying changes within the joint capsule are not fully understood. Adhesions and shortening of the synovial membrane, atro-phy and fibrosis of the capsule are known as structural changes (Trudel and Uhthoff 2000; Trudel et al. 2000; Moriyama et al. 2007), whereas decreased glycosaminogly-cans and water content, and possibly accumulation of advanced glycation endproducts such as pentosidine are known as biochemical changes (Akeson et al. 1977; Amiel et al. 1985; Lee et al. 2010). We reported in our previous

A. Ando et al.14
study that immobilization of rat knee joints induced focal adhesion of the capsule to the opposing capsule and the underlying articular cartilage even at 2 weeks, and global adhesion of the capsule to the opposing capsule and obliter-ation of the joint space by loose connective tissues after 4 weeks of immobilization (Ando et al. 2010). These focal and global adhesions of the capsule caused not only a short-ening of the synovial membrane but also a mechanical obstacle to the joint motion.
An inflammatory or a fibrotic process is proposed for the etiology of joint contracture (Neviaser 1945). The major structural collagens of the capsule are type I and III collagens (Kleftogiannis et al. 1994) and they are associated with tissue fibrosis (Matsumoto et al. 2002). Some researchers reported high levels of type I collagen and low levels of type III collagen in immobilized joints like ours (Matsumoto et al. 2002). Others reported neither type I nor type III collagen changed in immobilized knee joints induced by spinal cord injury in immunohistochemical and biochemical studies (Moriyama et al. 2007). In our previ-ous study, despite the decreased expression levels of type I and III collagen mRNAs as judged by PCR, the protein lev-els of type I and III collagens remained unchanged after immobilization by immunohistochemistry and western blot-ting (Hagiwara et al. 2010).
Reversibility of the articular cartilage after immobili-zation is well described in literature (Finsterbush and Friedman 1975; Palmoski et al. 1979; Behrens et al. 1989; Jurvelin et al. 1989; Schollmeier et al. 1996; Haapala et al. 1999; Ando et al. 2011). However, there were only a few studies that investigated the reversibility of the joint capsule (van Harreveld et al. 2002; Hildebrand et al. 2004; Trudel et al. 2008). Some investigators believed that immobiliza-tion alone did not cause a permanent contracture and the trauma or inflammation within the joint was essential to induce an irreversible restriction of the joint motion (Schollmeier et al. 1996; Hildebrand et al. 2004). Other conflicting data suggested that the restricted joint motion induced by immobilization alone was permanent after dis-continuation of immobilization, or even after discontinua-tion and extended exercises (Cronan 1986; van Harreveld et al. 2002; Trudel et al. 2008). It appears that duration and method of immobilization and remobilization affect the final outcome of the joint motion. Trudel et al. (2008) reported the reversibility of the joint contracture after 4 weeks of remobilization in immobilized knees for 8 weeks in a rat model and concluded that the contracture induced by joint immobilization alone was irreversible after remobi-lization. However, 4 weeks of remobilization was thought to be insufficient to clarify the reversibility of the joint motion in clinical situations, because in case of patients with joint contracture, it takes more than 4 weeks of reha-bilitation to acquire sufficient joint motion. Therefore, we set 16 weeks as the duration of remobilization. The pur-pose of this study was to clarify the reversibility of immo-bilization-induced capsular changes and restricted joint
motion in rat knee joints. We applied histological, histo-morphometric, and immunohistochemical techniques to localize the changes. Furthermore, we measured the joint angle under x-ray control after incision of the periarticular muscles of the joint, which allowed us to provide more accurate information about the arthrogenic restriction of the joint motion.
Materials and MethodsExperimental design and surgical procedure
Skeletally mature Sprague-Dawley male rats aged 12 weeks were used (CLEA Japan Inc., Tokyo, Japan). Institutional Animal Research Committee approval of Tohoku University was obtained prior to the animal experiments. Eighty and sixty-four rats were pre-pared for histology and joint angle measurement, respectively. Under anesthesia with intra-peritoneal administration of sodium pentobarbi-tal (50 mg/kg), the unilateral knee joints of the rats were rigidly immobilized at 150 degrees flexion with a plastic plate and two metal screws for 1, 2, 4, 8, and 16 weeks (n = 8/each period) according to a previously described method (Hagiwara et al. 2006; Hagiwara et al. 2010). Sham-operated rats had metal screws in the femur and the tibia, but the plate was not inserted. Buprenorphine (0.05 mg/kg) was injected subcutaneously for postoperative analgesia. After the experi-mental periods, a second anesthetic was administered to remove the plastic plate and metal screws of the immobilized rats and screws of the sham-operated rats, and the rats were allowed to move freely for 16 weeks (Ando et al. 2011). The immobilized and sham-operated rats made up the immobilized-remobilized (Im-Rm) group and the control group, respectively. Because limitation in the range of motion was almost accomplished at 8-weeks of immobilization in our previ-ous study (Chimoto et al. 2007), we set 1, 2, 4, and 8 weeks for joint angle measurement. Histological changes and joint angle measure-ment after immobilization alone were described in our previous reports (Chimoto et al. 2007; Ando et al. 2010); therefore, they were not included in this study.
Tissue preparationAfter the remobilization period, the rats were euthanized with
an overdose of sodium pentobarbital injection and fixed with 4% paraformaldehyde in 0.1 M phosphate buffered saline (PBS) by per-fusion through the ascending aorta. The knee joints were manually flexed at 150 degrees and kept in the position during the fixation. The resected knee joints were kept in the same fixative for 24 hours and decalcified in 10% ethylenediaminetetraacetic acid in 0.01 M PBS for 2 months at 4°C. The decalcified specimens were then embedded in paraffin. Five-μm sections were obtained at the medial midcondylar region of the knee in the sagittal plane.
Histology and histomorphometryThe histological examination was carried out with sections
stained by Elastica-Masson. Morphological changes of the capsule were observed in the anterior and posterior capsule. The length of the anterior and posterior synovial membrane divided by the anterior or posterior meniscus, which was called the antero-superior, antero-infe-rior, postero-superior, and postero-inferior length, was separately measured using a previously reported method (Trudel et al. 2000; Ando et al. 2010). In brief, the lengths were measured from the fem-oral synoviocartilage junctions to the upper meniscus and from the lower meniscus to the tibial synoviocartilage junctions in the anterior

Reversibility of Immobilization-Induced Joint Contracture 15
and posterior capsule. In the case of adhesions between the upper synovial membrane and the lower synovial membrane, the length of the residual joint space was also included in the total length. If syn-oviocytes were not observed on the surface of the joint space, it was not included in the total length. Light microscopic images were cap-tured (DMLB 100 HC light microscope; LEICA, Wetzlar, Germany) and the synovial membrane length was measured blindly by another observer using free image analysis software (Image J 1.44, Bethesda, MD).
ImmunohistochemistryThe immunohistochemical studies followed protocols estab-
lished in our laboratory (Ando et al. 2010; Hagiwara et al. 2010). Synovial membrane contains macrophage-like cells (type A synovio-cytes) and fibroblast-like cells (type B synoviocytes). CD68 was used as a marker of macrophages and type A synoviocytes (Damoiseaux et al. 1994; Ando et al. 2010). Deparaffinized sections were predigested with 0.1% trypsin (WAKO Chemicals, Tokyo, Japan) in 0.01 M PBS for 30 minutes at 37°C for CD 68 as an unmasking procedure. After the inhibition of endogenous peroxidase with 1.5% H2O2 in distilled water for 15 minutes, nonspecific binding was eliminated with 10% normal goat serum (Nichirei, Tokyo, Japan) for 30 minutes at room temperature. The sections were then incubated with mouse anti-rat CD68 monoclonal antibody (MCA341R, Serotec, Oxford, U.K., dilu-tion 1:100), rabbit anti-rat type I collagen polyclonal antibody (LB-1102, LSL, Tokyo, Japan, dilution 1:800), or rabbit anti-rat type III collagen polyclonal antibody (LB-1393, LSL, dilution 1:800) for 24 hours at 4°C. The secondary antibody, horseradish peroxidase conju-gated goat anti-mouse or goat anti-rabbit immunoglobulin (Nichirei) was applied for 1 hour at room temperature. The final detection step was carried out using 3,3′-diaminobenzidine tetrahydrochloride (Sigma, St. Louis, MO) in 0.1 M imidazole and 0.03% H2O2 as the chromogen for 10 minutes. Counterstaining was made with Carazzi’s hematoxylin. A negative control was performed using normal mouse or rabbit IgG (Dako, Copenhagen, Denmark) as a primary antibody. All slides were stained in one session.
Joint angle measurementsAfter injection of an overdose of sodium pentobarbital, the hind
limb of the rats was resected and periarticular skins and muscles were removed with special care not to break arthrogenic components. The angle between the longitudinal axis of the femur and the tibia was measured using a custom-made device under x-ray control (Chimoto et al. 2007). The joint angles in extension were measured at two torque values: 225 g-cm (Torque 1) and 450 g-cm (Torque 2). In order to evaluate the influence of the posterior capsule and the other arthrogenic components, the angle was measured again at torque 2 and torque 3 (1,350 g-cm) after incision of the posterior capsule. The posterior capsule was released from the femoral condyle with special care not to break cruciate and collateral ligaments (Chimoto et al. 2007). The x-ray pictures were scanned and the joint angle was mea-sured with the aid of free image analysis software (Image J 1.44).
Statistical analysisAll data were expressed as mean ± standard deviation.
Differences between the Im-Rm and control groups were compared at each time point by unpaired t-test. Significance was set at p < 0.05.
ResultsHistological changes of the capsule
The overall appearance of the posterior capsule in the control groups and Im-Rm groups was shown in Fig. 1. In the control groups, deeply folded joint capsule, postero-superior and postero-inferior synovial folds were observed and the apparent differences among the groups were not found (Fig. 1A-E). Cord-like structures were observed between the upper capsule and the lower capsule, and between the postero-superior synovial fold and the oppos-ing capsule in the 1/16-week and 2/16-week Im-Rm groups (Fig. 1F and G). In the 4/16-week and 8/16-week Im-Rm groups, the capsule adhered to the opposing capsule and femoral articular cartilage, and the residual joint space was obliterated by loose connective tissues and hardly visible (Fig. 1H and I). In the 16/16-week Im-Rm group, the folded capsule was totally disorganized and the joint space was not observed (Fig. 1J).
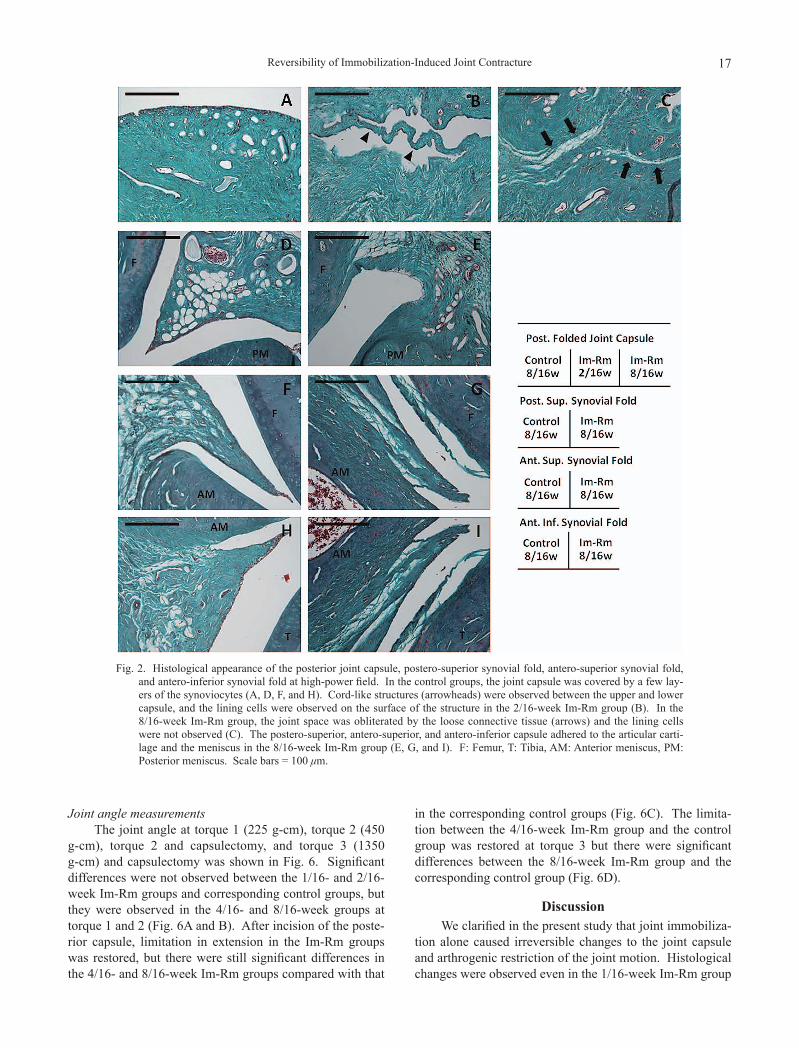
The posterior joint capsule, postero-superior synovial fold, antero-superior synovial fold, and antero-inferior synovial fold in the control and the Im-Rm groups at high power field were shown in Fig. 2. In the control groups, the capsule was covered with a few layers of the lining synovi-ocytes (Fig. 2A, D, F, and H). The joint space between the capsule and the articular cartilage, the capsule and the meniscus was maintained in each area. Cord-like structure was observed between the upper and the lower capsule in the 2/16-week Im-Rm group, where the lining cells were observed (Fig. 2B). In the 8/16-week Im-Rm group, the joint space between the upper and the lower capsule was obliterated by loose connective tissues and the lining cells were not observed (Fig. 2C). The postero-superior synovial fold adhered to the opposing articular cartilage and the meniscus in the 8/16-week Im-Rm group (Fig. 2E). In the antero-superior and antero-inferior synovial fold, the adhered connective tissue was observed on the surface of the articular cartilage, and the synovial cleft between the capsule and the meniscus was obliterated (Fig. 2G and I).
Length of the synovial membraneThe length of the antero-superior, antero-inferior, pos-
tero-superior, and postero-inferior synovial membrane was shown in Fig. 3. There were no statistical differences in the antero-superior synovial membrane length between the Im-Rm and control groups (Fig. 3A). The length of the antero-inferior synovial membrane in the 4/16-, 8/16-, and 16/16-week Im-Rm groups was significantly shorter com-pared with that in the corresponding control groups (Fig. 3B). The length of the postero-superior synovial membrane was significantly shorter in the 2/16-, 4/16-, 8/16-, and 16/16-week Im-Rm groups compared with the correspond-ing control groups (Fig. 3C). The length of the postero-inferior synovial membrane was significantly shorter in the 8/16- and 16/16-week Im-Rm groups compared with the corresponding control groups (Fig. 3D).

A. Ando et al.16
Expression of type I and III collagens, and CD68 in the capsule
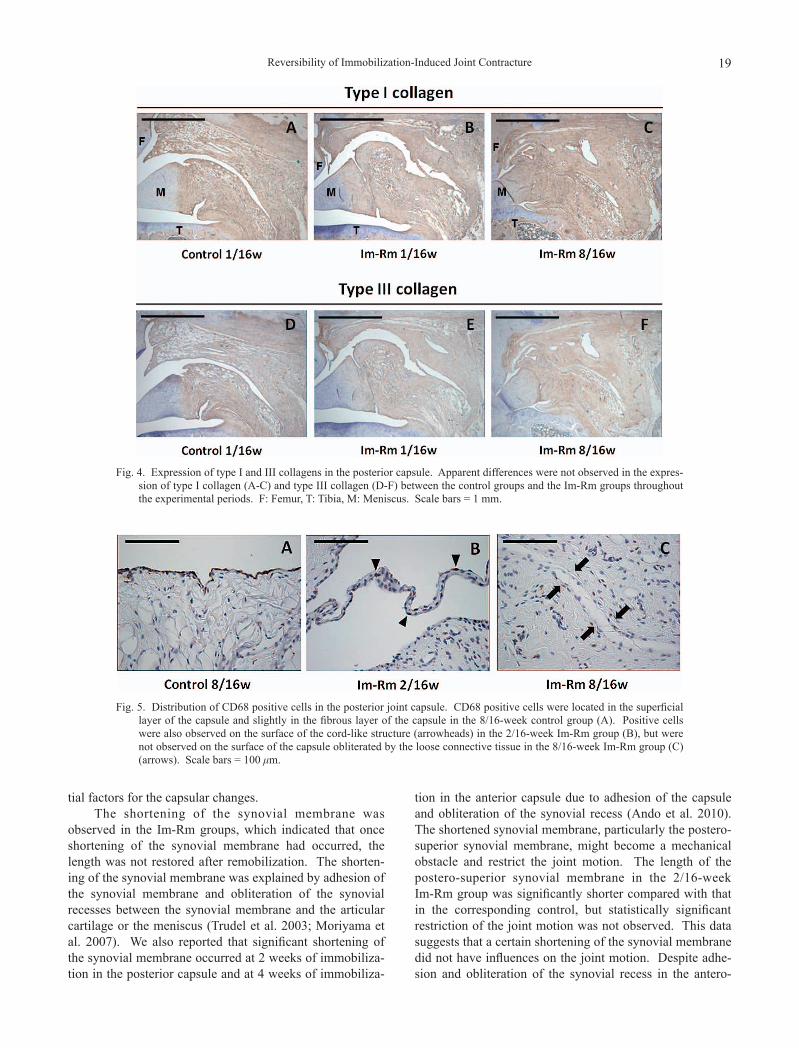
Expression of type I and III collagens in the posterior capsule was shown in Fig. 4. There were no differences in immunostaining intensity of type I (Fig. 4A-C) and type III collagens (Fig. 4D-F) between the Im-Rm groups and the control groups throughout the experimental periods. Apparent differences were not observed in the staining intensity of type I and III collagens in the adhesion area of the Im-Rm groups compared with that in the overall capsule (Fig. 4).
Distribution of CD68 positive cells (type A synovio-
cytes and macrophages) in the posterior folded joint capsule was shown in Fig. 5. CD68 positive cells were located in the superficial layer of the capsule and slightly in the fibrous layer of the capsule in the control groups (Fig. 5A). The positive cells were also observed in the fibrous layer of the capsule in the Im-Rm groups and on the surface of the cord-like structure between the upper and lower capsule in the 2/16-week Im-Rm group (Fig. 5B). The positive cells were not observed on the surface of the capsule obliterated by the loose connective tissues in the 8/16-week Im-Rm group (Fig. 5C).
Fig. 1. Histological appearance of the overall posterior capsule in the control groups and immobilized-remobilized (Im-Rm) groups. In the control groups, deeply folded joint capsule and postero-superior and postero-inferior synovial folds (asterisks) were observed (A-E). Cord-like structures (arrowheads) connecting the upper and lower capsule were observed in the 1/16- and 2/16-week Im-Rm groups (F, G). In the 4/16- and 8/16-week Im-Rm groups, the capsule adhered to the opposing capsule and the residual joint space was hardly visible (arrows) (H, I). In the 16/16-week Im-Rm group, the capsule was totally disorganized and the joint space was not visible (J). F: Femur, T: Tibia, M: Meniscus, JS: Joint space. Scale bars = 1 mm.
Reversibility of Immobilization-Induced Joint Contracture 17
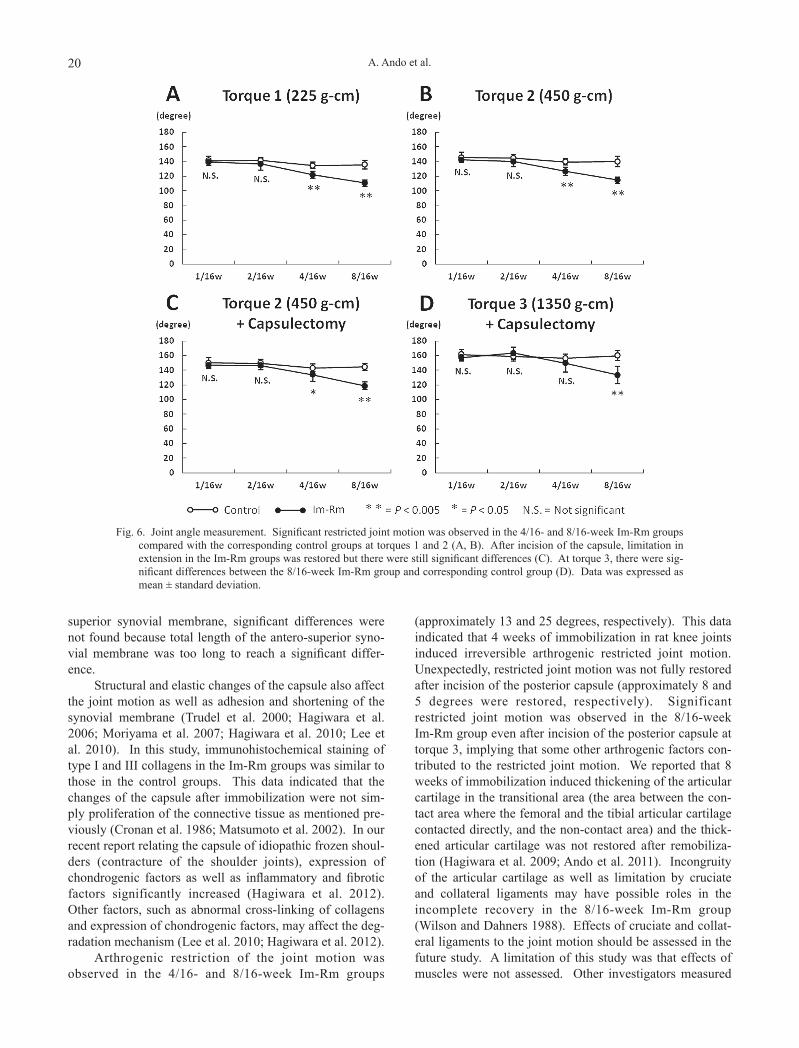
Joint angle measurementsThe joint angle at torque 1 (225 g-cm), torque 2 (450
g-cm), torque 2 and capsulectomy, and torque 3 (1350 g-cm) and capsulectomy was shown in Fig. 6. Significant differences were not observed between the 1/16- and 2/16-week Im-Rm groups and corresponding control groups, but they were observed in the 4/16- and 8/16-week groups at torque 1 and 2 (Fig. 6A and B). After incision of the poste-rior capsule, limitation in extension in the Im-Rm groups was restored, but there were still significant differences in the 4/16- and 8/16-week Im-Rm groups compared with that
in the corresponding control groups (Fig. 6C). The limita-tion between the 4/16-week Im-Rm group and the control group was restored at torque 3 but there were significant differences between the 8/16-week Im-Rm group and the corresponding control group (Fig. 6D).
DiscussionWe clarified in the present study that joint immobiliza-
tion alone caused irreversible changes to the joint capsule and arthrogenic restriction of the joint motion. Histological changes were observed even in the 1/16-week Im-Rm group
Fig. 2. Histological appearance of the posterior joint capsule, postero-superior synovial fold, antero-superior synovial fold, and antero-inferior synovial fold at high-power field. In the control groups, the joint capsule was covered by a few lay-ers of the synoviocytes (A, D, F, and H). Cord-like structures (arrowheads) were observed between the upper and lower capsule, and the lining cells were observed on the surface of the structure in the 2/16-week Im-Rm group (B). In the 8/16-week Im-Rm group, the joint space was obliterated by the loose connective tissue (arrows) and the lining cells were not observed (C). The postero-superior, antero-superior, and antero-inferior capsule adhered to the articular carti-lage and the meniscus in the 8/16-week Im-Rm group (E, G, and I). F: Femur, T: Tibia, AM: Anterior meniscus, PM: Posterior meniscus. Scale bars = 100 μm.

A. Ando et al.18
and significant restriction of the joint motion was observed in the 4/16- and 8/16-week Im-Rm groups. Akeson et al. (1977) reported that the contracture resulting from 9 weeks of immobilization in rabbit knee joints with a steel pin recovered rapidly after remobilization. Schollmeier et al. (1996) showed that the changes induced by 12 weeks of immobilization in dog forelegs with a shoulder spica were completely reversed after 12 weeks of remobilization. These results were inconsistent with ours, which may be explained by differences in joint immobilization techniques and animal species. We immobilized rat knee joints with a plastic plate and metal screws, which did not allow motion compared with the other methods such as casting or pin-ning. Even a small amount of motion may be advantageous to prevent irreversible restricted joint motion.
Cord-like structures were observed in the posterior capsule in the 1/16- and 2/16-week Im-Rm groups. The structure was observed in 4 out of 8 specimens in the 1/16-week Im-Rm group, and in all specimens in the 2/16-week Im-Rm group. Evans et al. (1960) mentioned the cord-like structures as elongated adhesion formed by remobilization. We previously reported that 1-week immobilization induced hypertrophy of the synovial membrane and accumulation of
cells in the subsynovial layer, and 2-week immobilization induced focal adhesion of the capsule to the opposing cap-sule (Ando et al. 2010). The focal adhesion area formed by 1- and 2-week immobilization was stretched by remobiliza-tion, which might make the cord-like structures. Arthrogenic restriction of the joint motion was not observed in the 1/16- and 2/16-week Im-Rm groups compared with that in the control groups, which indicated that the effects of the cord-like structures to the joint motion were not prominent.
Global adhesion of the posterior capsule and oblitera-tion of the joint space was observed in the 4/16-, 8/16-, and 16/16-week Im-Rm groups and the lining synovial cells were not observed on the surface of the adhesion area. In our previous report, over 4-week of immobilization induced global adhesion of the posterior capsule (Ando et al. 2010). This data indicated that once the capsule globally adhered and the lining synovial cells disappeared, restoration was not obtained even after prolonged remobilization. Adhesion of the capsule to the opposing capsule or to the articular cartilage and the meniscus was observed not only in the posterior capsule but also in the anterior capsule, which indicated that the flexed position was not the only influen-
Fig. 3. The length of the antero-superior (A-S), antero-inferior (A-I), postero-superior (P-S), and postero-inferior (P-I) syno-vial membrane. Statistical differences were not found in the length of the A-S synovial membrane (A). The lengths of the A-I, P-S, and P-I synovial membrane in the Im-Rm groups were significantly shorter in the 4/16-, 8/16-, and 16/16-week, in the 2/16-, 4/16-, 8/16-, and 16/16-week, and in the 8/16- and 16/16-week compared with that in the corre-sponding controls (B-D). Data was expressed as mean ± standard deviation.
Reversibility of Immobilization-Induced Joint Contracture 19
tial factors for the capsular changes.The shortening of the synovial membrane was
observed in the Im-Rm groups, which indicated that once shortening of the synovial membrane had occurred, the length was not restored after remobilization. The shorten-ing of the synovial membrane was explained by adhesion of the synovial membrane and obliteration of the synovial recesses between the synovial membrane and the articular cartilage or the meniscus (Trudel et al. 2003; Moriyama et al. 2007). We also reported that significant shortening of the synovial membrane occurred at 2 weeks of immobiliza-tion in the posterior capsule and at 4 weeks of immobiliza-
tion in the anterior capsule due to adhesion of the capsule and obliteration of the synovial recess (Ando et al. 2010). The shortened synovial membrane, particularly the postero-superior synovial membrane, might become a mechanical obstacle and restrict the joint motion. The length of the postero-superior synovial membrane in the 2/16-week Im-Rm group was significantly shorter compared with that in the corresponding control, but statistically significant restriction of the joint motion was not observed. This data suggests that a certain shortening of the synovial membrane did not have influences on the joint motion. Despite adhe-sion and obliteration of the synovial recess in the antero-
Fig. 4. Expression of type I and III collagens in the posterior capsule. Apparent differences were not observed in the expres-sion of type I collagen (A-C) and type III collagen (D-F) between the control groups and the Im-Rm groups throughout the experimental periods. F: Femur, T: Tibia, M: Meniscus. Scale bars = 1 mm.
Fig. 5. Distribution of CD68 positive cells in the posterior joint capsule. CD68 positive cells were located in the superficial layer of the capsule and slightly in the fibrous layer of the capsule in the 8/16-week control group (A). Positive cells were also observed on the surface of the cord-like structure (arrowheads) in the 2/16-week Im-Rm group (B), but were not observed on the surface of the capsule obliterated by the loose connective tissue in the 8/16-week Im-Rm group (C) (arrows). Scale bars = 100 μm.
A. Ando et al.20
superior synovial membrane, significant differences were not found because total length of the antero-superior syno-vial membrane was too long to reach a significant differ-ence.
Structural and elastic changes of the capsule also affect the joint motion as well as adhesion and shortening of the synovial membrane (Trudel et al. 2000; Hagiwara et al. 2006; Moriyama et al. 2007; Hagiwara et al. 2010; Lee et al. 2010). In this study, immunohistochemical staining of type I and III collagens in the Im-Rm groups was similar to those in the control groups. This data indicated that the changes of the capsule after immobilization were not sim-ply proliferation of the connective tissue as mentioned pre-viously (Cronan et al. 1986; Matsumoto et al. 2002). In our recent report relating the capsule of idiopathic frozen shoul-ders (contracture of the shoulder joints), expression of chondrogenic factors as well as inflammatory and fibrotic factors significantly increased (Hagiwara et al. 2012). Other factors, such as abnormal cross-linking of collagens and expression of chondrogenic factors, may affect the deg-radation mechanism (Lee et al. 2010; Hagiwara et al. 2012).
Arthrogenic restriction of the joint motion was observed in the 4/16- and 8/16-week Im-Rm groups
(approximately 13 and 25 degrees, respectively). This data indicated that 4 weeks of immobilization in rat knee joints induced irreversible arthrogenic restricted joint motion. Unexpectedly, restricted joint motion was not fully restored after incision of the posterior capsule (approximately 8 and 5 degrees were restored, respectively). Significant restricted joint motion was observed in the 8/16-week Im-Rm group even after incision of the posterior capsule at torque 3, implying that some other arthrogenic factors con-tributed to the restricted joint motion. We reported that 8 weeks of immobilization induced thickening of the articular cartilage in the transitional area (the area between the con-tact area where the femoral and the tibial articular cartilage contacted directly, and the non-contact area) and the thick-ened articular cartilage was not restored after remobiliza-tion (Hagiwara et al. 2009; Ando et al. 2011). Incongruity of the articular cartilage as well as limitation by cruciate and collateral ligaments may have possible roles in the incomplete recovery in the 8/16-week Im-Rm group (Wilson and Dahners 1988). Effects of cruciate and collat-eral ligaments to the joint motion should be assessed in the future study. A limitation of this study was that effects of muscles were not assessed. Other investigators measured
Fig. 6. Joint angle measurement. Significant restricted joint motion was observed in the 4/16- and 8/16-week Im-Rm groups compared with the corresponding control groups at torques 1 and 2 (A, B). After incision of the capsule, limitation in extension in the Im-Rm groups was restored but there were still significant differences (C). At torque 3, there were sig-nificant differences between the 8/16-week Im-Rm group and corresponding control group (D). Data was expressed as mean ± standard deviation.

Reversibility of Immobilization-Induced Joint Contracture 21
combined arthrogenic and myogenic restriction at first mea-surement, and then, arthrogenic restriction was measured after resection of the periarticular muscles (Trudel and Uhthoff 2000; Moriyama et al. 2006). In these methods, arthrogenic structures may be damaged at the first measure-ment, which influenced the second measurement of the arthrogenic component (Trudel et al. 2008). Therefore, we evaluated the arthrogenic restriction alone in this study.
ConclusionWe have shown in the present study that joint immobi-
lization alone induces irreversible histological changes and restricted joint motion in rat knee joints. Though 1 and 2 weeks of immobilization induced irreversible histological changes to the joint capsule, it did not affect the joint motion. Four weeks of immobilization induced irreversible arthrogenic restriction of the joint motion, which indicated that immobilization over 4 weeks should be avoided to pre-vent irreversible joint contracture.
AcknowledgmentsWe wish to thank Mr. Katsuyoshi Shoji and Mrs. Michiko
Fukuyama, Department of Orthopaedic Surgery, Tohoku Univer-sity School of Medicine, for their technical assistance.
Conflict of InterestThe authors have no conflict of interest to declare.
ReferencesAkeson, W.H., Woo, S.L., Amiel, D. & Doty, D.H. (1977) Rapid
recovery from contracture in rabbit hindlimb: a correlative biomechanical and biochemical study. Clin. Orthop. Relat. Res., 122, 359-365.
Akeson, W.H., Amiel, D., Abel, M.F., Garfin, S.R. & Woo, S.L. (1987) Effects of immobilization on joints. Clin. Orthop. Relat. Res., 219, 28-37.
Amiel, D., Ing, D., Frey, C., Woo, S.L., Harwood, F. & Akeson, W. (1985) Value of hyaluronic acid in the prevention of contrac-ture formation. Clin. Orthop. Relat. Res., 196, 306-311.
Ando, A., Hagiwara, Y., Onoda, Y., Hatori, K., Suda, H., Chimoto, E. & Itoi, E. (2010) Distribution of type A and B synoviocytes in the adhesive and shortened synovial membrane during immobilization of the knee joint in rats. Tohoku J. Exp. Med., 221, 161-168.
Ando, A., Suda, H., Hagiwara, Y., Onoda, Y., Chimoto, E., Saijo, Y. & Itoi, E. (2011) Reversibility of immobilization-induced articular cartilage degeneration after remobilization in rat knee joints. Tohoku J. Exp. Med., 224, 77-85.
Behrens, F., Kraft, E.L. & Oegema, T.R.Jr. (1989) Biochemical changes in articular cartilage after joint immobilization by casting or external fixation. J. Orthop. Res., 7, 335-343.
Chimoto, E., Hagiwara, Y., Ando, A. & Itoi, E. (2007) Progression of an arthrogenic motion restriction after immobilization in a rat experimental knee model. Ups. J. Med. Sci., 112, 347-355.
Cronan, T. (1986) Effects of immobilization and mobilization on cartilaginous, bony, and soft-tissue structure: review of the literature. J. Burn Care Rehabil., 7, 54-57.
Damoiseaux, J.G., Döpp, E.A., Calame, W., Chao, D., MacPherson, G.G. & Dijkstra, C.D. (1994) Rat macrophage lysosomal membrane antigen recognized by monoclonal antibody ED1. Immunology, 83, 140-147.
Enneking, W.F. & Horowitz, M. (1972) The intra-articular effects
of immobilization on the human knee. J. Bone Joint Surg. Am., 54, 973-985.
Evans, E.B., Eggers, G.W., Butler, J.K. & Blumel, J. (1960) Experimental immobilization and remobilization of rat knee joints. J. Bone Joint Surg. Am., 42, 737-758.
Finsterbush, A. & Friedman, B. (1975) Reversibility of joint changes produced by immobilization in rabbits. Clin. Orthop. Relat. Res., 111, 290-298.
Haapala, J., Arokoski, J.P., Hyttinen, M.M., Lammi, M., Tammi, M., Kovanen, V., Helminen, H.J. & Kiviranta, I. (1999) Remobilization does not fully restore immobilization induced articular cartilage atrophy. Clin. Orthop. Relat. Res., 362, 218-229.
Hagiwara, Y., Saijo, Y., Chimoto, E., Akita, H., Sasano, Y., Matsumoto, F. & Kokubun, S. (2006) Increased elasticity of capsule after immobilization in a rat knee experimental model assessed by scanning acoustic microscopy. Ups. J. Med. Sci., 111, 303-313.
Hagiwara, Y., Ando, A., Chimoto, E., Saijo, Y., Ohmori-Matsuda, K. & Itoi, E. (2009) Changes of articular cartilage after immobilization in a rat knee contracture model. J. Orthop. Res., 27, 236-242.
Hagiwara, Y., Ando, A., Onoda, Y., Matsui, H., Chimoto, E., Suda, H. & Itoi, E. (2010) Expression patterns of collagen types I and III in the capsule of a rat knee contracture model. J. Orthop. Res., 28, 315-321.
Hagiwara, Y., Ando, A., Onoda, Y., Takemura, T., Minowa, T., Hanagata, N., Tsuchiya, M., Watanabe, T., Chimoto, E., Suda, H., Takahashi, N., Sugaya, H., Saijo, Y. & Itoi, E. (2012) Coexistence of fibrotic and chondrogenic process in the capsule of idiopathic frozen shoulders. Osteoarthritis Cart., 20, 241-249.
Hildebrand, K.A., Sutherland, C. & Zhang, M. (2004) Rabbit knee model of post-traumatic joint contractures: the long-term natural history of motion loss and myofibroblasts. J. Orthop. Res., 22, 313-320.
Jurvelin, J., Kiviranta, I., Säämänen, A.M., Tammi, M. & Helminen, H.J. (1989) Partial restoration of immobilization-induced softening of canine articular cartilage after remobili-zation of the knee (stifle) joint. J. Orthop. Res., 7, 352-358.
Kleftogiannis, F., Handley, C.J. & Campbell, M.A. (1994) Charac-terization of extracellular matrix macromolecules from bovine synovial capsule. J. Orthop. Res., 12, 365-374.
Lee, S., Sakurai, T., Ohsako, M., Saura, R., Hatta, H. & Atomi, Y. (2010) Tissue stiffness induced by prolonged immobilization of the rat knee joint and relevance of AGEs (pentosidine). Connect. Tissue Res., 51, 467-477.
Matsumoto, F., Trudel, G. & Uhthoff, H.K. (2002) High collagen type I and low collagen type III levels in rat knee joint contracture: an immunohistochemical study with histological correlate. Acta. Orthop. Scand., 73, 335-343.
Moriyama, H., Yoshimura, O., Sunahori, H. & Tobimatsu, Y. (2006) Comparison of muscular and articular factors in the progression of contractures after spinal cord injury in rats. Spinal Cord, 44, 174-181.
Moriyama, H., Yoshimura, O., Kawamata, S., Takemoto, H., Saka, Y. & Tobimatsu, Y. (2007) Alteration of knee joint connective tissues during contracture formation in spastic rats after an experimentally induced spinal cord injury. Connect. Tissue Res., 48, 180-187.
Neviaser, J.S. (1945) Adhesive capsulitis of the shoulder. J. Bone Joint Surg. Am., 27, 211-222.
Palmoski, M.J., Perricone, E. & Brandt, K.D. (1979) Development and reversal of a proteoglycan aggregation defect in normal canine knee cartilage after immobilization. Arthritis Rheum., 22, 508-517.
Roy, S. & Ghadially, F.N. (1967) Ultrastructure of normal rat synovial membrane. Ann. Rheum. Dis., 26, 26-38.
Schollmeier, G., Sarkar, K., Fukuhara, K. & Uhthoff, H.K. (1996)

A. Ando et al.22
Structural and functional changes in the canine shoulder after cessation of immobilization. Clin. Orthop. Relat. Res., 323, 310-315.
Trudel, G. & Uhthoff, H.K. (2000) Contractures secondary to immobility: is the restriction articular or muscular? An exper-imental longitudinal study in the rat knee. Arch. Phys. Med. Rehabil., 81, 6-13.
Trudel, G., Seki, M. & Uhthoff, H.K. (2000) Synovial adhesions are more important than pannus proliferation in the pathogen-esis of knee joint contracture after immobilization: an experi-mental investigation in the rat. J. Rheumatol., 27, 351-357.
Trudel, G., Jabi, M. & Uhthoff, H.K. (2003) Localized and adap-tive synoviocyte proliferation characteristics in rat knee joint contractures secondary to immobility. Arch. Phys. Med.
Rehabil., 84, 1350-1356.Trudel, G., Zhou, J. & Uhthoff, H.K. (2008) Four weeks of
mobility after 8 weeks of immobility fails to restore normal motion: a preliminary study. Clin. Orthop. Relat. Res., 466, 1239-1244.
van Harreveld, P.D., Lillich, J.D., Kawcak, C.E., Gaughan, E.M., McLaughlin, R.M. & DeBowes, R.M. (2002) Clinical evalua-tion of the effects of immobilization followed by remobiliza-tion and exercise on the metacarpophalangeal joint in horses. Am. J. Vet. Res., 63, 282-288.
Wilson, C.J. & Dahners, L.E. (1988) An examination of the mech-anism of ligament contracture. Clin. Orthop. Relat. Res., 227, 286-291.
Related Documents











