JOURNAL OF BACTERIOLOGY, Apr. 2008, p. 2388–2399 Vol. 190, No. 7 0021-9193/08/$08.000 doi:10.1128/JB.01602-07 Copyright © 2008, American Society for Microbiology. All Rights Reserved. Regulation of Expression and Secretion of NleH, a New Non-Locus of Enterocyte Effacement-Encoded Effector in Citrobacter rodentium Vı ´ctor A. Garcı ´a-Angulo, 1 Wanyin Deng, 2 Nikhil A. Thomas, 2,3 B. Brett Finlay, 2 and Jose L. Puente 1 * Departamento de Microbiologı ´a Molecular, Instituto de Biotecnologı ´a, Universidad Nacional Auto ´noma de Me ´xico, Cuernavaca, Morelos, Mexico 1 ; Michael Smith Laboratories, University of British Columbia, Vancouver, British Columbia, Canada 2 ; and Department of Microbiology and Immunology/Medicine, Dalhousie University, Halifax, Nova Scotia, Canada 3 Received 2 October 2007/Accepted 14 January 2008 Together with enterohemorrhagic Escherichia coli and enteropathogenic Escherichia coli, Citrobacter roden- tium is a member of the attaching-and-effacing (A/E) family of bacterial pathogens. A/E pathogens use a type III secretion system (T3SS) to translocate an assortment of effector proteins, encoded both within and outside the locus of enterocyte effacement (LEE), into the colonized host cell, leading to the formation of A/E lesions and disease. Here we report the identification and characterization of a new non-LEE encoded effector, NleH, in C. rodentium. NleH is conserved among A/E pathogens and shares identity with OspG, a type III secreted effector protein in Shigella flexneri. Downstream of nleH, genes encoding homologues of the non-LEE-encoded effectors EspJ and NleG/NleI are found. NleH secretion and translocation into Caco-2 cells requires a functional T3SS and signals located at its amino-terminal domain. Transcription of nleH is not significantly reduced in mutants lacking the LEE-encoded regulators Ler and GrlA; however, NleH protein levels are highly reduced in these strains, as well as in escN and cesT mutants. Inactivation of Lon, but not of ClpP, protease restores NleH levels even in the absence of CesT. Our results indicate that the efficient engagement of NleH in active secretion is needed for its stability, thus establishing a posttranslational regulatory mechanism that coregulates NleH levels with the expression of LEE-encoded proteins. A C. rodentium nleH mutant shows a moderate defect during the colonization of C57BL/6 mice at early stages of infection. Enteropathogenic Escherichia coli (EPEC) and enterohemor- rhagic E. coli (EHEC) are important causes of infectious di- arrhea. EPEC is the leading cause of severe watery bacterial diarrhea in children under 6 months of age living in developing countries. EHEC infections are associated with outbreaks of bloody diarrhea and hemorrhagic colitis in the developed world and can lead to severe kidney failure due to the devel- opment of the often-fatal hemolytic-uremic syndrome (33). EPEC and EHEC are noninvasive pathogens that infect their hosts by adhering to the surface of the intestinal epithelial cells. These organisms belong to a family of pathogens that are capable of producing a common histopathology called the at- taching-and-effacing (A/E) lesion, which is characterized by the localized destruction of the enterocyte microvilli and the formation of pedestal-shaped actin-rich structures underneath the adherent bacteria (30). In vivo study of the mechanisms underlying EPEC and EHEC pathogenesis has been difficult, mainly due to the inability of these pathogens to efficiently colonize laboratory animals. Because of this, Citrobacter roden- tium, an A/E pathogen that naturally colonizes laboratory mice, causing A/E lesions and disease, has emerged as a suit- able model to study A/E infections (48). The genes required for A/E lesion formation are located within the locus of enterocyte effacement (LEE) and encode an assortment of proteins involved in the assembly of a type III secretion system (T3SS), intimate attachment, effector func- tions, chaperones, and regulation, which to different degrees are all required for virulence (15, 21). The LEE also encodes SepL and SepD, two proteins that form a molecular switch that confers specificity to the T3SS for the secretion of translocator proteins. In addition, this switch acts as a gatekeeper for the secretion of effector proteins, as sepD or sepL mutants hypersecrete them without secreting translocator proteins (14, 15). Proteomic analysis of the se- creted proteins of a C. rodentium sepD mutant led us to the identification of seven new T3SS-secreted proteins, all of which are encoded outside the LEE in C. rodentium and other A/E pathogens. We named these proteins non-LEE-encoded effec- tors (Nle): NleA, NleB, NleC, NleD, NleE, NleF, and NleG (15). Further characterization of NleA (also called EspI), which is widely distributed among EHEC and EPEC strains (9, 10, 49), showed that it is critical for virulence and colonization of the mouse colon and upon translocation into epithelial cells localizes to the Golgi apparatus (27, 49). NleB, which plays a key role in C. rodentium virulence (35, 65), is encoded within O-island 122, which has been proposed to enhance virulence in EHEC O157:H7 strains (34), to be associated with diarrheal disease due to atypical EPEC (1), and to be associated with the LEE, forming a mosaic island in EHEC and atypical EPEC strains of serogroup O26 (4). It has also been demonstrated that NleC and NleD are translocated through the EHEC T3SS. NleC is not required for EHEC colonization of lambs or calves or for C. rodentium colonization of mice (35, 41). An EHEC EDL933 nleD mutant was originally identified by signature- tagged mutagenesis as being attenuated in calves (17); how- ever, it was later reported that no attenuation was observed for nleD deletion mutants of both EHEC 85-170 and EDL933 * Corresponding author. Mailing address: Departamento de Micro- biologı ´a Molecular, Instituto de Biotecnologı ´a, Universidad Nacional Auto ´noma de Me ´xico, Av. Universidad 2001, Cuernavaca, Morelos 62210, Me ´xico. Phone: (52) (777) 329-1621. Fax: (52) (777) 313-8673. E-mail: [email protected]. Published ahead of print on 25 January 2008. 2388 Downloaded from https://journals.asm.org/journal/jb on 25 October 2021 by 119.237.38.221.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

JOURNAL OF BACTERIOLOGY, Apr. 2008, p. 2388–2399 Vol. 190, No. 70021-9193/08/$08.00�0 doi:10.1128/JB.01602-07Copyright © 2008, American Society for Microbiology. All Rights Reserved.
Regulation of Expression and Secretion of NleH, a New Non-Locus ofEnterocyte Effacement-Encoded Effector in Citrobacter rodentium�
Vıctor A. Garcıa-Angulo,1 Wanyin Deng,2 Nikhil A. Thomas,2,3 B. Brett Finlay,2 and Jose L. Puente1*Departamento de Microbiologıa Molecular, Instituto de Biotecnologıa, Universidad Nacional Autonoma de Mexico, Cuernavaca,
Morelos, Mexico1; Michael Smith Laboratories, University of British Columbia, Vancouver, British Columbia, Canada2; andDepartment of Microbiology and Immunology/Medicine, Dalhousie University, Halifax, Nova Scotia, Canada3
Received 2 October 2007/Accepted 14 January 2008
Together with enterohemorrhagic Escherichia coli and enteropathogenic Escherichia coli, Citrobacter roden-tium is a member of the attaching-and-effacing (A/E) family of bacterial pathogens. A/E pathogens use a typeIII secretion system (T3SS) to translocate an assortment of effector proteins, encoded both within and outsidethe locus of enterocyte effacement (LEE), into the colonized host cell, leading to the formation of A/E lesionsand disease. Here we report the identification and characterization of a new non-LEE encoded effector, NleH,in C. rodentium. NleH is conserved among A/E pathogens and shares identity with OspG, a type III secretedeffector protein in Shigella flexneri. Downstream of nleH, genes encoding homologues of the non-LEE-encodedeffectors EspJ and NleG/NleI are found. NleH secretion and translocation into Caco-2 cells requires afunctional T3SS and signals located at its amino-terminal domain. Transcription of nleH is not significantlyreduced in mutants lacking the LEE-encoded regulators Ler and GrlA; however, NleH protein levels are highlyreduced in these strains, as well as in escN and cesT mutants. Inactivation of Lon, but not of ClpP, proteaserestores NleH levels even in the absence of CesT. Our results indicate that the efficient engagement of NleHin active secretion is needed for its stability, thus establishing a posttranslational regulatory mechanism thatcoregulates NleH levels with the expression of LEE-encoded proteins. A C. rodentium nleH mutant shows amoderate defect during the colonization of C57BL/6 mice at early stages of infection.
Enteropathogenic Escherichia coli (EPEC) and enterohemor-rhagic E. coli (EHEC) are important causes of infectious di-arrhea. EPEC is the leading cause of severe watery bacterialdiarrhea in children under 6 months of age living in developingcountries. EHEC infections are associated with outbreaks ofbloody diarrhea and hemorrhagic colitis in the developedworld and can lead to severe kidney failure due to the devel-opment of the often-fatal hemolytic-uremic syndrome (33).EPEC and EHEC are noninvasive pathogens that infect theirhosts by adhering to the surface of the intestinal epithelialcells. These organisms belong to a family of pathogens that arecapable of producing a common histopathology called the at-taching-and-effacing (A/E) lesion, which is characterized bythe localized destruction of the enterocyte microvilli and theformation of pedestal-shaped actin-rich structures underneaththe adherent bacteria (30). In vivo study of the mechanismsunderlying EPEC and EHEC pathogenesis has been difficult,mainly due to the inability of these pathogens to efficientlycolonize laboratory animals. Because of this, Citrobacter roden-tium, an A/E pathogen that naturally colonizes laboratorymice, causing A/E lesions and disease, has emerged as a suit-able model to study A/E infections (48).
The genes required for A/E lesion formation are locatedwithin the locus of enterocyte effacement (LEE) and encodean assortment of proteins involved in the assembly of a type III
secretion system (T3SS), intimate attachment, effector func-tions, chaperones, and regulation, which to different degreesare all required for virulence (15, 21).
The LEE also encodes SepL and SepD, two proteins thatform a molecular switch that confers specificity to the T3SS forthe secretion of translocator proteins. In addition, this switchacts as a gatekeeper for the secretion of effector proteins, assepD or sepL mutants hypersecrete them without secretingtranslocator proteins (14, 15). Proteomic analysis of the se-creted proteins of a C. rodentium sepD mutant led us to theidentification of seven new T3SS-secreted proteins, all of whichare encoded outside the LEE in C. rodentium and other A/Epathogens. We named these proteins non-LEE-encoded effec-tors (Nle): NleA, NleB, NleC, NleD, NleE, NleF, and NleG(15). Further characterization of NleA (also called EspI),which is widely distributed among EHEC and EPEC strains (9,10, 49), showed that it is critical for virulence and colonizationof the mouse colon and upon translocation into epithelial cellslocalizes to the Golgi apparatus (27, 49). NleB, which plays akey role in C. rodentium virulence (35, 65), is encoded withinO-island 122, which has been proposed to enhance virulence inEHEC O157:H7 strains (34), to be associated with diarrhealdisease due to atypical EPEC (1), and to be associated with theLEE, forming a mosaic island in EHEC and atypical EPECstrains of serogroup O26 (4). It has also been demonstratedthat NleC and NleD are translocated through the EHEC T3SS.NleC is not required for EHEC colonization of lambs or calvesor for C. rodentium colonization of mice (35, 41). An EHECEDL933 nleD mutant was originally identified by signature-tagged mutagenesis as being attenuated in calves (17); how-ever, it was later reported that no attenuation was observed fornleD deletion mutants of both EHEC 85-170 and EDL933
* Corresponding author. Mailing address: Departamento de Micro-biologıa Molecular, Instituto de Biotecnologıa, Universidad NacionalAutonoma de Mexico, Av. Universidad 2001, Cuernavaca, Morelos62210, Mexico. Phone: (52) (777) 329-1621. Fax: (52) (777) 313-8673.E-mail: [email protected].
� Published ahead of print on 25 January 2008.
2388
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
25
Oct
ober
202
1 by
119
.237
.38.
221.

(41). The nleE gene is also associated with EHEC O-island 122(34), and C. rodentium nleE mutants are attenuated in mixedinfections and produce delayed mortality in susceptible mice(35, 66). The role of NleG in vivo has not been studied; how-ever, an NleG homologue found in EPEC, named NleI, hasbeen shown to be translocated into epithelial cells in a T3S-dependent manner (39). A recent proteomic and bioinformaticapproach expanded the number of effectors in EHEC to 39,which were classified into 20 families (61). In addition to theNle proteins, other non-LEE-encoded effectors have been de-scribed in A/E pathogens (reviewed in reference 21).
Along with the acquisition of virulence factors, pathogenicbacteria have also acquired or developed regulatory mecha-nisms to ensure the appropriate spatio-temporal expression ofsuch factors. In A/E pathogens, the expression of LEE-carriedgenes is tightly regulated by both global and A/E-specific reg-ulatory factors (reviewed in reference 45). The LEE encodesthree A/E-specific regulators. Ler (LEE-encoded regulator)has an essential role in the positive regulation of all the geneswithin the LEE (5, 28, 46, 54), as well as of others locatedoutside the LEE such as espC in EPEC (20) and lpf in EHEC(62). Ler positively regulates LEE gene expression by counter-acting the repression exerted by the global regulator H-NS (5,28, 63). The first gene of the LEE1 operon encodes Ler, whoseexpression is negatively and positively regulated by a myriad ofglobal regulatory proteins (reviewed in reference 45). In addi-tion, A/E-specific regulatory proteins such as the EAF plas-mid-encoded PerC in EPEC (5, 26, 50) or Pch in EHEC (32),as well as GrlA, which is also encoded within the LEE (3, 15),play critical roles in ler positive regulation, while GrlR, whichis coexpressed with GrlA, has a negative role (15, 40). Inter-estingly, Ler and GrlA establish a positive regulatory loop that,together with GrlR, seems to modulate Ler levels in the cell(3). Ler and GrlA have been shown to play an essential role invivo (15, 68). Furthermore, the regulatory mechanisms associ-ated with the expression of effector genes located outside theLEE have just started to be elucidated (47, 51, 53).
Due to its association with the gene coding for the previouslyidentified non-LEE-encoded effector NleF in C. rodentium andits similarity with OspG of Shigella flexneri, we hypothesizedthat the upstream gene, herein designated nleH, was likelyto encode an effector protein. Using double-hemagglutinin(2HA)- and CyaA-tagged versions of NleH, we showed that itssecretion and translocation to epithelial cells depend on afunctional T3SS and that upon translocation it localizes to thecell membranes. Regulation of nleH was analyzed in differentregulatory and secretory mutants by using an operon fusionbetween the entire nleH gene and the cat reporter gene. NleHis regulated at the posttranslational level by a novel mechanismthat couples protein stability with Ler-GrlA-dependent regu-lation of LEE gene expression. Its role in colonization wasassessed using the murine model of infection.
MATERIALS AND METHODS
Bacterial strains, plasmids, cell lines, and culture conditions. The bacterialstrains and plasmids used in this study are described in Table 1. Bacteria weregrown at 37°C in Luria-Bertani (LB) broth or agar or in Dulbecco’s modifiedEagle’s medium (DMEM). When required, the medium was supplemented withampicillin (100 �g/ml) and kanamycin (30 �g/ml). Growth of C. rodentium inDMEM was carried out at 37°C in a 5% (vol/vol) CO2 atmosphere without
shaking. When EPEC was grown in DMEM, media were supplemented with 1%LB and incubated at 37°C with shaking (220 rpm). Caco-2 (ATCC HTB-37) cellswere cultivated in minimum essential medium buffered with 2.4 g/liter HEPESand supplemented with 15% (vol/vol) fetal bovine serum and 200 mML-glutamine at 37°C in a humidified 5% (vol/vol) CO2 atmosphere.
Construction of deletion mutants and plasmids. Deletion of the nleH genefrom C. rodentium DBS100 was performed by the one-step mutagenesis proce-dure developed by Datsenko and Wanner (12). The mutation eliminated thestructural region of nleH and replaced it with a Km resistance marker. For theconstruction of plasmid pTnleH-HA expressing NleH tagged with a 2HAepitope, the DNA sequence containing the nleH putative ribosome binding siteand the NleH-coding region without its stop codon was amplified by PCR fromthe C. rodentium DBS100 genome using primers NleHCF (5�-GCAAAAAGCTTCCGGTTTTTGTTGTCATGTCAGGG-3�) and NleHCR (5�-GCCACTCGAGAATTCTACTTAATACCACTCTGATAAG-3�) containing HindIII andXhoI sites, respectively. The resulting PCR product was digested with HindIII-XhoI and ligated into pTOPO-2HA (Table 1) digested with the same restrictionenzymes. EPEC �lon �ler and �lon �grlA mutants were generated by the sacBgene-based allelic exchange method as described previously (15) using suicideclones carrying in-frame deletions of grlA and ler (Table 1).
To generate pTnleH20-293HA and pTnleH1-133HA expressing truncated ver-sions of NleH-2HA, pTnleH-HA DNA was used as the template for inverse PCRwith primers RecHA (5�-AGCTCCGGTACCCATACATTTCAACCTTCAAAATAAGAC-3�) and RecH20B (5�-AGAAACGGTACCTCGCCTGATAATGCCGTCTTATCC-3�), which were designed to eliminate nleH codons 2 to 19 andto introduce KpnI sites. The resulting PCR products were digested with KpnIand self-ligated. To generate pTnleH1-133HA, pTnleH-HA DNA was used asthe template for inverse PCR with primers NleHHAF (5�-AGTCGCTCGAGTATCCGTATGATG-3�) and NleH2AC160 (5�-ATTACCCTCGAGGGGTGACTTGTTATAGTCCAC-3�), which were designed to eliminate nleH codons 134to 293 and to introduce XhoI sites. The resulting PCR products were digestedwith XhoI and self-ligated.
To generate a plasmid containing the 2HA-tagged nleH gene and the catreporter gene forming an operon under the control of the nleH regulatory region,the nleH coding region without its stop codon plus 1,334 bp upstream of thetranslational start codon was amplified by PCR from the C. rodentium genomeusing primers NleHReg (5�-AAGTGCAAGCTTCCTCATAAGATGGAAAACTGC-3�) and NleHCR. The resulting PCR product was cloned into the HindIII-XhoI sites of pTOPO-2HA to generate pTregNleH-HA (Table 1). Next, PCRswere performed to amplify the whole insert from pTregNleH-HA, including thesequence corresponding to the 2HA epitope. The resulting PCR product wascloned into the BamHI-HindIII sites of pKK232-8 to generate pKKnleH-HA(Table 1). Likewise, pKKespG-HA was constructed using primers EspGHibF(5�-GTCCTAGGATCCCCGGGGCGGGGTCAGTCC-3�) and EspGHibR (5�-GTAATACTCGAGAGCATTGTTCAGATATGTTTCAG-3�), which were de-signed to amplify espG from the C. rodentium chromosome, including 353 bpupstream of the start codon. The PCR product was cloned into the BamHI-XhoIsites of pKKnleH-HA.
To create a vector suitable for the construction of CyaA reporter fusions, thesequence corresponding to codons 2 to 401 of the cyaA sequence contained inpOG-ATC�304 was amplified by PCR and cloned into the EcoRI-NotI sites ofpMPM-K3, to generate pK3cyaA (Table 1). pK3cyaA possesses unique PstI andBglII sites at the cyaA 5� end. To generate pK3nleHcyaA, the nleH coding regionwithout its stop codon, plus 73 bp upstream of the start codon, was amplifiedfrom the C. rodentium chromosome and cloned into the HindIII-PstI sites ofpK3cyaA. To construct pK3nleH1-19cyaA, pK3nleHcyaA DNA was used as thetemplate for inverse PCR with primers containing PstI restriction sites that weredesigned to amplify nleH without codons 20 to 293. The resulting PCR productwas PstI digested and self-ligated.
Protein secretion assays. Fifty milliliters of DMEM with the appropriateantibiotic was inoculated with 500 �l of an LB broth overnight culture (opticaldensity at 600 nm � 2) of the indicated strains, incubated for 1 h at 37°C in a 5%CO2 atmosphere without shaking, and then induced with 0.5 mM or 5 �M IPTG(isopropyl-�-D-thiogalactopyranoside) and incubated for another 5 h. Bacteriawere harvested by centrifugation at 17,900 � g for 10 min. Secreted proteins wereprecipitated from 1,350 �l of culture supernatants of each strain with 10%trichloroacetic acid at 4°C overnight and concentrated by centrifugation at17,900 � g for 30 min at 4°C. The resulting fractions were subjected to sodiumdodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and Westernblot analysis.
Cell fractionation. Mechanical fractionation of infected epithelial cells wasperformed as previously described (23), with slight modifications. Caco-2 cellmonolayers (approximately 2 � 106 cells) in 75-cm2 flasks were inoculated with
VOL. 190, 2008 NleH OF CITROBACTER RODENTIUM 2389
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
25
Oct
ober
202
1 by
119
.237
.38.
221.

200 �l of a 3-h-preinduced DMEM culture of EPEC strains (multiplicity ofinfection � 25) carrying different plasmids. Infection proceeded for 1.5 h, andthen the expression of NleH and its derivatives was induced by adding 0.5 mMIPTG and incubating for another 1.5 h. Media were discarded from monolayers,and cells were washed three times with phosphate-buffered saline (PBS) andharvested after treatment with 0.025% trypsin–0.2-g/liter EDTA in PBS. Afterthe cells were washed three times with ice-cold PBS, they were resuspended in300 �l of homogenization buffer (3 mM imidazole [pH 7.4], 250 mM sucrose, 0.5mM EDTA) plus a protease inhibitor cocktail (Roche). Cells were mechanicallydisrupted by vigorous passage two times through a 22-gauge needle and six timesthrough a 27-gauge needle. Low-speed centrifugation (3,000 � g) for 15 min wasapplied to this homogenate to pellet bacteria, unbroken Caco-2 cells, host nuclei,and cytoskeleton components (low-speed pellet). The supernatant was thensubjected to ultracentrifugation at 33,000 � g for 60 min to separate the insoluble
fraction (host cell membranes) from the soluble fraction (host cytosol). Theresulting fractions were resolved by SDS-PAGE (10% polyacrylamide) andtransferred to nitrocellulose for Western blot analysis.
Western blot assays. Nitrocellulose membranes containing transferred pro-teins were blocked in 5% nonfat milk for 1 h. Membranes were incubated witheither a 1:5,000 dilution of an anti-HA monoclonal (MAb) (Covance), 1:15,000dilution of an anti-DnaK MAb (Invitrogen), a 1:5,000 dilution of an anti-CyaAMAb (Cedarlane), a 1:10,000 dilution of an anti-�-tubulin MAb (Sigma), a1:10,000 dilution of an anticalnexin rabbit polyclonal antibody (Sigma), or a1:20,000 dilution of an anti-EspA rabbit polyclonal antibody (a kind gift of V.Sperandio and J. B. Kaper). Secondary antibodies included goat anti-rabbitimmunoglobulin G (Pierce) at a 1:10,000 dilution and rabbit anti-mouse immu-noglobulin G (Pierce) at a 1:10,000 dilution. Positive signals were visualized withWestern Lightning Chemiluminescence Reagent Plus (Perkin-Elmer).
TABLE 1. Strains and plasmids used in this study
Strain, strain genotype,or plasmid Description Reference or source
C. rodentium strainsDBS100 WT (ATCC 51459) 55�ler DBS100 carrying an in-frame deletion of ler 15�grlA DBS100 carrying an in-frame deletion of grlA 15�grlR DBS100 carrying an in-frame deletion of grlR 15�sepD DBS100 carrying an in-frame deletion of sepD 15�sepD �escN DBS100 carrying an in-frame deletion of sepD and an in-frame deletion of escN 15�escN DBS100 carrying an in-frame deletion of escN 15�cesT DBS100 carrying an in-frame deletion of cesT 15�nleH DBS100 �nleH::Km This study�nleF DBS100 �nleF::Km 66
EPEC strainsE2348/69 WT 38�escN E2348/69 carrying an in-frame deletion of escN 24�lon E2348/69 carrying an in-frame deletion of lon W. Deng, unpublished�cesT �lon E2348/69 carrying an in-frame deletion of cesT and an in-frame deletion of lon W. Deng, unpublished�clpP E2348/69 carrying an in-frame deletion of clpP W. Deng, unpublished�cesT �clpP E2348/69 carrying an in-frame deletion of cesT and an in-frame deletion of clpP W. Deng, unpublished�ler E2348/69 carrying an in-frame deletion of ler V. H. Bustamante, unpublished�grlA E2348/69 carrying an in-frame deletion of grlA A. Huerta, unpublished�ler �lon E2348/69 carrying an in-frame deletion of ler and an in-frame deletion of lon This study�grlA �lon E2348/69 carrying an in-frame deletion of grlA and an in-frame deletion of lon This study
PlasmidspKD46 Red recombinase system under control of araB promoter; Apr 12pKD4 Template plasmid containing the Km cassette for lambda Red recombinase 12pTOPO-2HA pCR2.1-TOPO derivative carrying C. rodentium espG coding region fused to
two HA epitopes at the carboxyl terminus15
pTnleH-HA pTOPO-2HA derivative carrying C. rodentium nleH coding region fused to twoHA epitopes at the caboxyl terminus
This study
pTnleH20-293HA pTnleH-HA with nleH coding region deleted from codon 2 to 19 This studypTnleH1-133HA pTnleH-HA with nleH coding region deleted from codon 134 to 293 This studypMPM-K3 Low-copy-no. cloning vector; p15A derivative; Kmr 44pOG-ATC�304 pACYC184 derivative expressing amino acids 1–32 of Salmonella enterica SseK1
fused to CyaA37
pK3cyaA pMPM-K3 derivative containing cyaA residues 2–401 This studypK3nleHcyaA pK3cyaA containing nleH coding region fused to cyaA This studypK3nleH1-19cyaA pK3nleHcyaA with nleH coding region deleted (codons 20–293) This studypKK232-8 pBR322 derivative containing a promoterless cat gene Pharmacia LKB BiotechnologypTregNleH-HA pTOPO-2HA derivative containing C. rodentium nleH fused to two HA
epitopes at the carboxyl terminus, including 1,749 bp of the regulatory regionThis study
pKKnleH-HA pKK232-8 derivative containing nleH-cat transcriptional fusion from nucleotide1741 to �876 with respect to the start codon; nleH fused to two HAepitopes
This study
pKKespG-HA pKKnleH-HA derivative containing espG-cat transcriptional fusion fromnucleotide 363 to �1182 with respect to the start codon; espG fused to twoHA epitopes
This study
pRE112 Suicide vector; sacB Cmr 18pRE-�ler Suicide construct carrying an in-frame deletion of EPEC ler in pRE112 V. H. Bustamante, unpublishedpRE-�grlA Suicide construct carrying an in frame deletion of EPEC grlA in pRE112 A. Huerta, unpublished
2390 GARCIA-ANGULO ET AL. J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
25
Oct
ober
202
1 by
119
.237
.38.
221.

Infection analysis of C. rodentium in mice. Three- to 4-week-old C57BL/6 micewere obtained from the Jackson Laboratory (Bar Harbor, ME). Wild-type (WT)C. rodentium and its nleH mutant derivative were grown in LB broth overnight at37°C at 200 rpm. Mice were inoculated by oral gavage with 100 �l of thesecultures. The inoculum was titrated by serial dilution and plating and was cal-culated to be approximately 2.5 � 108 CFU per mouse for all groups. To assaybacterial colonization, mice were sacrificed and the first 4 cm of the distal colonstarting from the anal verge was collected. Colonic tissue and fecal pellets werehomogenized using a Polytron tissue homogenizer and serially diluted beforebeing plated on MacConkey agar (Difco Laboratories) to determine the totalbacterial burden in the mouse at the time of sacrifice.
CAT assay. Determination of chloramphenicol acetyltransferase (CAT) activ-ity and protein quantification to calculate CAT specific activities were performedas previously described (42).
Statistical analysis. One-way analysis of variance, using SPSS software, wasused for statistical analysis.
RESULTS
Identification of nleH in the genome of C. rodentium. Wepreviously reported the identification, by proteomic analysis, of
a new set of putative effector proteins secreted by a sepDmutant strain of C. rodentium in a T3S-dependent manner(15). Genome sequence analysis revealed that these proteinswere encoded not by genes within the LEE but by genes scat-tered throughout the genome that were located in crypticprophages or putative pathogenicity islands, and they thereforewere designated non-LEE-encoded effectors (Nle) (15). Anal-ysis of the chromosomal location of nle genes in the partiallysequenced C. rodentium genome (http://www.sanger.ac.uk)showed that nleF and a nleG homologue were located in achromosomal locus inserted between the yjjG and prfC genes,which are contiguous in the EHEC EDL933 and nonpatho-genic E. coli K-12 chromosomes (Fig. 1A). This locus spans12.6 kb and also contains the gene coding for the T3SS-trans-located effector EspJ, which is found in a different location inEHEC and EPEC (11). Besides the described effector genes, inthis locus we found an open reading frame of 293 codonsimmediately upstream of nleF, whose product shares 15%
FIG. 1. NleH is conserved in A/E pathogens. (A) Schematic representation of the loci containing nleH in C. rodentium DBS100 and the twohomologues of nleH in EPEC E2348/69 and EHEC EDL933. NleH1 and NleH2 correspond, respectively, to Z6021 and Z0989 in the EHECEDL933 genome sequence, to ECs1814 and ECs0848 in the EHEC Sakai genome sequence, and to NleH1-2 and NleH1-1 according to a recentlysuggested nomenclature (64). Flanking genes correspond to those conserved in E. coli K-12 and are considered the boundaries of the insertion sitesof this putative horizontally acquired set of genes. (B) Multiple alignment of NleH homologues from C. rodentium, EPEC E2348/69, and EHECEDL933 and OspG from Shigella flexneri M90T. Sequences of C. rodentium and EPEC E2348/69 were obtained from the sequencing genomeproject at the Sanger Institute (www.sanger.ac.uk). Conserved amino acid residues involved in NF-B signaling as described for OspG (36) areindicated by arrowheads.
VOL. 190, 2008 NleH OF CITROBACTER RODENTIUM 2391
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
25
Oct
ober
202
1 by
119
.237
.38.
221.
identity with OspG, an effector of Shigella flexneri (36) (Fig.1B). Because of its location in the C. rodentium genome andthe fact that its predicted amino acid sequence shares identitywith a type III secreted effector in S. flexneri, we speculatedthat this gene codes for an effector protein, herein namedNleH. In addition, we had also found that an NleH homologuepresent in the secreted proteins of EPEC �sepD interacts withCesT (60).
NleH shares a high degree of identity (83%) with the pre-dicted protein products of two open reading frames of un-known function found in the genome of EHEC EDL933, des-ignated Z6021 and Z0989, herein called NleH1 and NleH2,respectively (Fig. 1B). In the partially sequenced genome ofEPEC strain E2348/69 (http://www.sanger.ac.uk), we alsofound two NleH homologues sharing around 83% identity withC. rodentium NleH. The loci containing the EPEC and EHECnleH homologues do not share the chromosomal insertion sitewith C. rodentium nleH. The nleH1 gene in EHEC is part of a58.6-kb chromosomal locus containing prophage genes and thegenes coding for NleA, NleF, and a NleG-like protein, whichinterrupt yciD, between E. coli backbone genes yciE and ynfA.
In EPEC, the 55.5-kb locus containing nleH1 is inserted be-tween yciD and trpA and also contains nleA and nleF but notnleG (Fig. 1A). nleH2 genes in EPEC and EHEC are alsofound in regions containing prophage structural genes. In bothcases the region containing nleH2 is inserted between the E.coli K-12 backbone genes ybhC and ybhB, though the size ofthe locus in each case is different (Fig. 1A). In EPEC the nleH2locus spans 52.3 kb and in EHEC 38.1 kb. In EPEC, the nleH2locus contains a cif homologue, while in EHEC the nleH2 locuscontains homologues of three nle genes: nleD, nleC, and nleB2.The gene organization of the nleH locus in C. rodentium issimilar to that of the nleH1 loci of both EHEC and EPEC;however, the predicted protein shares higher identity withNleH2, including the lack of a 10-amino-acid indel betweenresidues 29 and 30 (Fig. 1B).
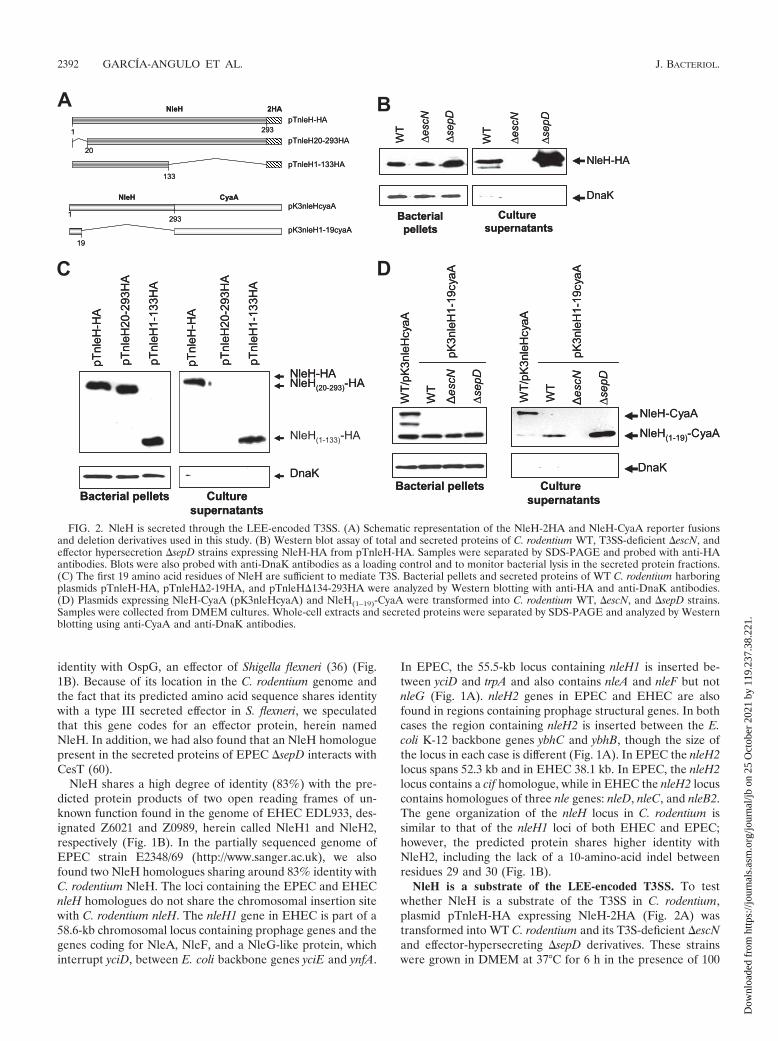
NleH is a substrate of the LEE-encoded T3SS. To testwhether NleH is a substrate of the T3SS in C. rodentium,plasmid pTnleH-HA expressing NleH-2HA (Fig. 2A) wastransformed into WT C. rodentium and its T3S-deficient �escNand effector-hypersecreting �sepD derivatives. These strainswere grown in DMEM at 37°C for 6 h in the presence of 100
FIG. 2. NleH is secreted through the LEE-encoded T3SS. (A) Schematic representation of the NleH-2HA and NleH-CyaA reporter fusionsand deletion derivatives used in this study. (B) Western blot assay of total and secreted proteins of C. rodentium WT, T3SS-deficient �escN, andeffector hypersecretion �sepD strains expressing NleH-HA from pTnleH-HA. Samples were separated by SDS-PAGE and probed with anti-HAantibodies. Blots were also probed with anti-DnaK antibodies as a loading control and to monitor bacterial lysis in the secreted protein fractions.(C) The first 19 amino acid residues of NleH are sufficient to mediate T3S. Bacterial pellets and secreted proteins of WT C. rodentium harboringplasmids pTnleH-HA, pTnleH�2-19HA, and pTnleH�134-293HA were analyzed by Western blotting with anti-HA and anti-DnaK antibodies.(D) Plasmids expressing NleH-CyaA (pK3nleHcyaA) and NleH(1–19)-CyaA were transformed into C. rodentium WT, �escN, and �sepD strains.Samples were collected from DMEM cultures. Whole-cell extracts and secreted proteins were separated by SDS-PAGE and analyzed by Westernblotting using anti-CyaA and anti-DnaK antibodies.
2392 GARCIA-ANGULO ET AL. J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
25
Oct
ober
202
1 by
119
.237
.38.
221.

�M IPTG to induce the expression of NleH-2HA. The se-creted proteins were recovered from culture supernatants bytrichloroacetic acid precipitation and subjected to Westernblot analysis using anti-HA antibodies (Fig. 2B). NleH-2HAwas expressed in all the strains tested but was detected only inthe culture supernatants of the WT and �sepD strains, asexpected for a T3S effector (Fig. 2B). Together, these resultsdemonstrate that NleH is secreted through the LEE-encodedT3SS in C. rodentium.
Although a signature secretion signal has not been describedfor T3S effectors, it has been determined to reside within thefirst 10 to 20 N-terminal amino acid residues (8, 58). To de-termine whether the NleH secretion signal was located at theN or C terminus, two NleH-2HA derivatives were generated bydeleting codons 2 to 19 and 134 to 293, obtaining plasmidspTnleH20-293-HA and pTnleH1-133HA, respectively (Fig.2A). The secretion of these NleH derivatives was tested in C.rodentium, where, as shown in Fig. 2C, the construct lackingthe first 19 amino acids was expressed but not detected inculture supernatants. In contrast, deletion of the C terminusdid not affect expression or secretion, indicating that, as forother T3S effectors, the signal required for NleH secretionresides at the amino-terminal domain.
To further support this observation, two CyaA reporter fu-sions were constructed, with the entire NleH amino acid se-quence (pK3nleHcyaA) and with only the first 19 amino-ter-minal residues (pK3nleH1-19cyaA) (Fig. 2A). In accordancewith our previous results, the complete NleH-CyaA fusion wassecreted by WT C. rodentium, as it was detected by Westernblotting with anti-CyaA antibodies in the culture supernatant(Fig. 2D). The presence of two NleH-CyaA processing prod-ucts indicated that this construct was somehow unstable, but itdid not interfere with the secretion of the entire fusion. TheCyaA fusion containing NleH residues 2 to 19 was detected inthe culture supernatants of the WT and �sepD strains but notin the supernatant of the �escN strain, demonstrating that thefirst 19 residues of NleH are sufficient to confer type III-dependent secretion (Fig. 2D).
NleH associates with host cell membranes upon transloca-tion. In order to determine whether NleH is also translocatedinto epithelial cells through the LEE-encoded T3SS, we usedEPEC E2348/69 and its �escN derivative expressing the NleH-2HA and NleH-CyaA fusions depicted in Fig. 2A to infectCaco-2 cells and perform fractionation assays. It has beenshown that EPEC adheres more efficiently to epithelial cells invitro, thus providing a more sensitive way to evaluate theT3SS-dependent translocation of effector proteins by A/Epathogens into epithelial cells (11, 41, 64). Infected cells weresubjected to mechanical fractionation and the fractions ob-tained analyzed by Western blotting using anti-HA and anti-CyaA antibodies (Fig. 3). These results showed that upontranslocation, full-length NleH-2HA and NleH-CyaA associatemainly with the host membrane fraction and that this associa-tion requires a functional T3SS, as NleH was not found inmembrane fractions of cells infected with EPEC �escN (Fig.3). NleH(1–133)-HA was also found to be translocated andassociated with the membrane fraction, indicating that thecarboxyl terminus does not contain the translocation or mem-brane association signal. In contrast, NleH(1–19)-CyaA was nottranslocated, suggesting that even when sufficient for secretion,
the first 19 amino acids are not enough for translocation. Col-lectively, these results indicate that while secretion is achievedwith only the first 19 amino acids, the signals sustaining trans-location and membrane association of NleH are located withinits first 133 amino acids.
Regulation of NleH expression by LEE-encoded regulators.Despite the fact that the number of identified non-LEE-en-coded effectors in A/E pathogens has grown significantly inrecent years, little is known about their regulation. As NleH issecreted and translocated in a T3S-dependent manner, weinvestigated whether its expression is coregulated with the ex-pression of LEE-encoded genes. To monitor the expression ofboth the protein and a reporter gene under the control of thenleH regulatory region, we constructed an operon fusion be-tween the nleH regulatory and structural regions double taggedwith the HA epitope and the CAT gene in plasmid pKK232-8,generating plasmid pKKnleH-HA (Fig. 4A). To assess the roleof LEE-encoded regulators in NleH expression, we introducedpKKnleH-HA in C. rodentium WT, �ler, �grlA, and �grlRstrains. Strains were grown in DMEM at 37°C without shakingunder a 5% CO2 atmosphere for 6 h, which are conditionsknown to induce LEE gene expression in C. rodentium (3, 15).Culture samples were used to determine CAT activity andanalyze NleH-2HA expression and secretion by Western blot-ting. As shown in Fig. 4B, the CAT activity directed by thefusion contained in pKKnleH-HA was not significantly re-duced in the �ler and �grlA strains with respect to the WTstrain, suggesting that nleH transcription is not part of theLer-GrlA regulon. In contrast, NleH-HA was undetectable byWestern blotting in both whole cells and culture supernatants ofthe �ler and �grlA mutants (Fig. 4C). Although not statisticallysignificant, the CAT activities directed by the fusion contained inpKKnleH-HA showed opposite trends in the grlA (reduced ex-pression) and grlR (enhanced expression) mutants, suggestingthat GrlA and GrlR may modulate NleH expression positively
FIG. 3. NleH translocation into Caco-2 cells. Caco-2 cells wereinfected with WT and �escN EPEC E2348/69 strains harboring plas-mids expressing the NleH fusions described in Fig. 2A. Upon IPTGinduction, cells were subjected to fractionation by mechanical lysis asdescribed in Materials and Methods. The low-speed pellet fractioncontains intact Caco-2 cells, complete organelles, cytoskeleton compo-nents, and intact bacteria. As fractionation controls, all blots wereanalyzed using anti-HA, anti-CyaA, anti-DnaK, anticalnexin, and an-titubulin antibodies. Each fractionation was done twice with identicalresults.
VOL. 190, 2008 NleH OF CITROBACTER RODENTIUM 2393
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
25
Oct
ober
202
1 by
119
.237
.38.
221.
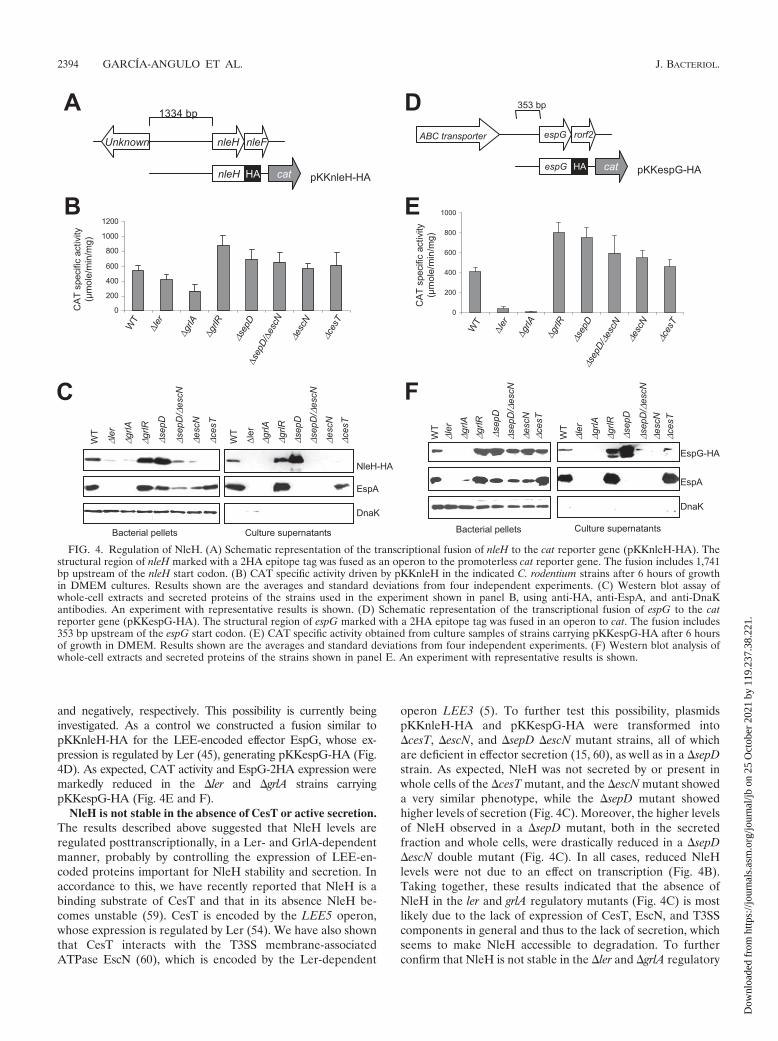
and negatively, respectively. This possibility is currently beinginvestigated. As a control we constructed a fusion similar topKKnleH-HA for the LEE-encoded effector EspG, whose ex-pression is regulated by Ler (45), generating pKKespG-HA (Fig.4D). As expected, CAT activity and EspG-2HA expression weremarkedly reduced in the �ler and �grlA strains carryingpKKespG-HA (Fig. 4E and F).
NleH is not stable in the absence of CesT or active secretion.The results described above suggested that NleH levels areregulated posttranscriptionally, in a Ler- and GrlA-dependentmanner, probably by controlling the expression of LEE-en-coded proteins important for NleH stability and secretion. Inaccordance to this, we have recently reported that NleH is abinding substrate of CesT and that in its absence NleH be-comes unstable (59). CesT is encoded by the LEE5 operon,whose expression is regulated by Ler (54). We have also shownthat CesT interacts with the T3SS membrane-associatedATPase EscN (60), which is encoded by the Ler-dependent
operon LEE3 (5). To further test this possibility, plasmidspKKnleH-HA and pKKespG-HA were transformed into�cesT, �escN, and �sepD �escN mutant strains, all of whichare deficient in effector secretion (15, 60), as well as in a �sepDstrain. As expected, NleH was not secreted by or present inwhole cells of the �cesT mutant, and the �escN mutant showeda very similar phenotype, while the �sepD mutant showedhigher levels of secretion (Fig. 4C). Moreover, the higher levelsof NleH observed in a �sepD mutant, both in the secretedfraction and whole cells, were drastically reduced in a �sepD�escN double mutant (Fig. 4C). In all cases, reduced NleHlevels were not due to an effect on transcription (Fig. 4B).Taking together, these results indicated that the absence ofNleH in the ler and grlA regulatory mutants (Fig. 4C) is mostlikely due to the lack of expression of CesT, EscN, and T3SScomponents in general and thus to the lack of secretion, whichseems to make NleH accessible to degradation. To furtherconfirm that NleH is not stable in the �ler and �grlA regulatory
FIG. 4. Regulation of NleH. (A) Schematic representation of the transcriptional fusion of nleH to the cat reporter gene (pKKnleH-HA). Thestructural region of nleH marked with a 2HA epitope tag was fused as an operon to the promoterless cat reporter gene. The fusion includes 1,741bp upstream of the nleH start codon. (B) CAT specific activity driven by pKKnleH in the indicated C. rodentium strains after 6 hours of growthin DMEM cultures. Results shown are the averages and standard deviations from four independent experiments. (C) Western blot assay ofwhole-cell extracts and secreted proteins of the strains used in the experiment shown in panel B, using anti-HA, anti-EspA, and anti-DnaKantibodies. An experiment with representative results is shown. (D) Schematic representation of the transcriptional fusion of espG to the catreporter gene (pKKespG-HA). The structural region of espG marked with a 2HA epitope tag was fused in an operon to cat. The fusion includes353 bp upstream of the espG start codon. (E) CAT specific activity obtained from culture samples of strains carrying pKKespG-HA after 6 hoursof growth in DMEM. Results shown are the averages and standard deviations from four independent experiments. (F) Western blot analysis ofwhole-cell extracts and secreted proteins of the strains shown in panel E. An experiment with representative results is shown.
2394 GARCIA-ANGULO ET AL. J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
25
Oct
ober
202
1 by
119
.237
.38.
221.

mutants, these strains were transformed with pTnleH-HA,which expresses NleH-2HA from an IPTG-inducible promoter(Table 1). In contrast to the case for the WT and �grlR strains,NleH-2HA was not seen in whole cells or culture supernatantsof the �ler and �grlA strains (Fig. 5A). Together, these resultsindicated that even when NleH was expressed from an induc-ible promoter, its stability was dependent not only on thepresence of its chaperone but also on active secretion. Inter-estingly, the EspG effector protein was not subjected to thesame posttranslational control, as EspG-2HA could be de-tected in whole cells of all strains, except for the �ler and �grlAstrains, and as a secreted protein in the �cesT strain whenexpressed from its own promoter (Fig. 4F). When expressedfrom an inducible promoter, EspG-2HA could be detectedeven in whole cells of the regulatory mutants but not, as ex-pected, in their secreted proteins (Fig. 5B).
Lon, but not ClpP, is involved in the degradation of NleH inthe absence of CesT. Energy-dependent proteases such as Lonand ClpP have been involved in the expression or stability ofT3S effectors in different pathogenic bacteria (reviewed in ref-erence 6). To explore whether these proteases are responsiblefor the rapid turnover of NleH observed in the absence ofCesT, NleH-2HA was expressed in WT EPEC and its �cesT,�cesT �lon, and �cesT �clpP derivatives. As shown in Fig. 6A,NleH-2HA was readily detected in whole cells of the �cesTstrain in the absence of the Lon protease but was still degradedin the absence of ClpP, suggesting that when NleH is not beingrecruited by CesT and actively secreted, this effector protein isbeing specifically targeted for degradation by Lon. We thenaddressed whether the lon mutation was able to restore NleHstability in the �ler and �grlA backgrounds. Interestingly, NleHstability was restored in the �grlA �lon but not in the �ler �londouble mutant (Fig. 6A), revealing that the posttranslationalregulation of NleH may involve two different pathways andthat more than one Ler-regulated component, in addition toCesT, may be involved in NleH protein stability. This resultprompted an additional experiment in which the roles in pro-
tein instability of the N- and C-terminal domains of NleH weretested by expressing the N-terminal [NleH(20–293)-HA] andC-terminal [NleH(1–133)-HA] deletions in �ler and �grlA mu-tants. Consistent with the notion that two different pathwaysare involved in NleH degradation, only the N-terminal deletionwas stable in the �grlA mutant, while none of the NleH dele-tion derivatives was stable in the �ler mutant (Fig. 6B). Theseobservations are in line with our recent observation that theCesT binding domain of NleH is contained within the first 40amino acids (59).
Lack of NleH delays colonization of C57BL/6 mice by C.rodentium. Considering that NleH displays different features ofa T3S effector, its role as a virulence factor for C. rodentiumwas assessed by infecting C57BL/6 mice with C. rodentiumDBS100 and its nleH null mutant. Infected animals were sac-rificed at days 6 and 10 postinfection (p.i.), and the total bac-terial burden in the mouse colon was calculated. The nleHmutant showed a moderate, statistically significant (P � 0.01)reduction in its capacity to colonize the mouse colon at day6 p.i. (Fig. 7A). However, no difference in colonization be-tween the WT and �nleH strains was observed at day 10 p.i.(Fig. 7B). In contrast, a strong attenuation was observed forthe nleB and escN mutants (data not shown), as previouslyreported (15, 35, 65). Overall, these results indicate that NleHhas a role in promoting efficient colonization of the mousecolon by C. rodentium during early stages of infection.
FIG. 5. NleH expressed from Plac is coordinately regulated with theLEE-encoded T3SS. Western blot assays of the bacterial pellet andsecreted proteins of the indicated C. rodentium strains carryingpTnleH-HA expressing NleH-HA from Plac (A) and pTOPO-2HAexpressing EspG-HA from Plac (B), using anti-HA and anti-DnaKantibodies, are shown. Expression was induced with 5 �M IPTG for5 h. Assays were done twice with identical results.
FIG. 6. Lon protease degrades NleH in the absence of CesT.(A) Western blot assay of bacterial pellets of EPEC strains (as indi-cated) transformed with pTnleH-HA, which expresses an NleH-HAfusion from Plac, using anti-HA and anti-DnaK antibodies. Expressionof the fusion was induced with 5 �M IPTG for 5 h. (B) Analysis of thestability of full-length NleH-HA and of its amino- and carboxy-termi-nal deletions NleH(20–293)-HA and NleH(1–133)-HA (illustrated in Fig.2A). Western blot assays of WT, �ler, and �grlA strains of C. rodentiumcontaining pTnleH-HA, pTnleH20-293HA, and pTnleH1-133HA plas-mids are shown. Strains were grown in DMEM, and the expression offusions was induced with 5 �M IPTG for 5 h. Assays were done twicewith identical results.
VOL. 190, 2008 NleH OF CITROBACTER RODENTIUM 2395
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
25
Oct
ober
202
1 by
119
.237
.38.
221.

DISCUSSION
In recent years, the development of effector hypersecretionmutants combined with the use of proteomics and bioinfor-matics has allowed the identification of a significant number ofnew T3S effector proteins in A/E pathogens, which are en-coded outside the LEE pathogenicity island (15, 61). In thiswork, we describe different features of NleH, a non-LEE-encoded effector that we originally identified in C. rodentiumbased on its homology to S. flexneri OspG and its geneticlinkage to other previously characterized T3S effectors (Fig.1A) (15), as well as in EPEC as a CesT binding substrate (60).
In C. rodentium, nleH is located in a 12.6-kb locus insertedbetween the yjjG and prfC genes that has no apparent struc-tural phage genes. In EPEC E2348/69 and EHEC EDL933there are two loci carrying nleH homologues (nleH1 andnleH2), but their site of insertion is different with respect tothat in C. rodentium (Fig. 1A). The fact that both nleH copiesin EHEC and EPEC are prophage carried (61) is not unex-pected given the importance that phages have acquired asvirulence gene dissemination vectors, in particular T3S effec-tors (19, 61). Furthermore, for EPEC serogroups O86a, O127,and O142, isolated from patients with acute diarrhea, it hasrecently been reported that the locus carrying nleH2 is aninducible lambdoid prophage that also encodes the cytolethal-distending toxin (Cdt-I) (2), an observation that highlights thepotential mobility of these putative virulence factors. Differ-ences in the genomic context of the nleH homologues, as forother T3SS effectors, support the notion that the acquisition of
effector genes by A/E pathogens has been the result of multipleand independent horizontal transfer events.
Using NleH-2HA- and NleH-CyaA-tagged proteins, weshowed that the first 19 N-terminal amino acid residues of C.rodentium NleH are essential and sufficient for its secretionthrough the LEE-encoded T3SS but not for its translocationinto epithelial cells. In contrast, it has been reported that thefirst 20 amino acids are sufficient to mediate both secretion andtranslocation of other A/E effectors, such as Tir, EspF, Map,Cif, and NleA/EspI (8). However, our results are consistentwith the notion that the secretion signal resides within the first15 to 20 amino acids of effector proteins of different T3SSs,while translocation signals are located further down the N-terminal domain (25). In fact, the modular organization of Ntermini consisting of separated secretion and translocation do-mains has been reported for other effectors, such as YersiniaYopE and YopH (57). Secretion and translocation of NleHhomologues in EPEC and EHEC have also been recently re-ported (59, 61). In addition, we show that upon translocationNleH associates with the membrane fraction, likely corre-sponding to the cytoplasmic membrane instead of organellemembranes, considering that upon translocation EPEC NleHlocalizes underneath adherent bacteria, as we have recentlyreported (59).
Another distinctive feature of all LEE-encoded effectors, aswell as of T3SS components, is that their expression is coordi-nated at the transcriptional level by Ler (45). As non-LEE-encoded effectors are secreted by the LEE-encoded T3SS, ithas been tempting to speculate that their expression is coregu-lated with the LEE. In this regard, it has been recently re-ported that nleA, nleB, nleD, and nleE are transcribed in EHECunder secretion-permissive conditions and that the expressionof nleA::gfp, nleA::bla, and nleD::bla translational reporter fu-sions showed a reduction in the absence of Ler (53). Thisresult, however, did not distinguish whether Ler-mediated reg-ulation occurs at the transcriptional or posttranslational level.Interestingly, we found here that even when Ler and GrlA donot seem to play a significant role in nleH transcription, theLer-mediated regulation of LEE expression influences post-translationally the expression of NleH, as even when NleH wasexpressed from an inducible promoter, its levels in the cellwere drastically reduced in the absence of these regulatoryproteins. In addition, NleH protein levels were greatly dimin-ished in the absence of CesT, as well as in secretion-deficientstrains such as the escN mutant, although no significant effectswere seen for nleH transcription in these strains.
We have recently shown that CesT is a multieffector chap-erone that recruits different LEE- and non-LEE-encoded ef-fectors for their efficient T3S (59, 60). These observations ledto us to speculate that when NleH in not actively engaged forsecretion, its protein levels may be controlled by the competingaction of proteases to prevent its potentially toxic accumula-tion within the cell. One would expect that it would be moreenergetically favorable to regulate the expression of these non-LEE-encoded effector proteins at the transcriptional level,through the same mechanisms that regulate the expression ofLEE genes. However, we still have to determine when andunder which conditions NleH is expressed, as we cannot ruleout the possibility that the spatio-temporal expression of NleHdiffers from that of LEE-encoded genes. This would imply that
FIG. 7. NleH is required at early stages during colonization in themouse model of infection. Groups of C57BL/6 mice were orally in-fected with equal numbers of WT or �nleH bacteria. Mice were sac-rificed at day 6 (A) or 10 (B) p.i., and the total burden of C. rodentiumin the distal colon was assessed by MacConkey serial dilution platting.*, statistically significant difference with respect to the WT strain(P � 0.01).
2396 GARCIA-ANGULO ET AL. J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
25
Oct
ober
202
1 by
119
.237
.38.
221.

its Ler-independent transcriptional regulation is required toensure its expression at a different stage of the infection or ata different niche, once the T3SS components are all in place.Thus, if T3S does not occur, NleH expression would be post-transcriptionally regulated, by controlling either mRNA stabil-ity, translation, or protein stability. The first two options do notseem to be the case, as no polar effect is seen in the expressionof the cat reporter gene when in an operon with nleH.
Regarding protein stability, recent reports have establishedthat energy-dependent proteases, such as Lon and ClpP, con-trol the intracellular concentration of effector proteins in dif-ferent bacterial pathogens expressing T3SS effectors (reviewedin reference 6). Here, we show that NleH is fully stable in theabsence of CesT when Lon, but not ClpP, is inactive, suggest-ing that when NleH is not bound by CesT and/or engaged forsecretion, Lon-specific proteolysis degrades the accumulatedprotein to prevent undesired effects. Interestingly, the muta-tion of lon restored NleH levels in the �grlA mutant but not inthe �ler mutant, suggesting that NleH stability may rely on, inaddition to CesT, another NleH-stabilizing factor regulated byLer that prevents NleH degradation by a Lon-independentpathway. Consistently, we also observed differential stabilitybetween the N-terminal and C-terminal deletion derivatives ofNleH in the �ler and �grlA mutants. In line with these obser-vations, even in the presence of CesT, lack of secretion ofNleH in the �escN mutant also prompts its degradation, whilehigher levels of NleH are secreted when actively engaged insecretion in the �sepD hypersecretion mutant. Thus, in theabsence of GrlA, although the expression of Ler is not com-pletely abolished, it is nevertheless highly reduced (3, 15). Thislow level of Ler expression may not support the expression ofCesT to sustain NleH stability but may be enough to activatethe expression of an additional NleH-stabilizing factor that isneeded to prevent NleH degradation from the C terminus.This model may explain why the stability of NleH and itsderivatives was not restored in the �ler�lon mutant, where thelack of Ler has a profound effect on the expression of the Lerregulon (reviewed in reference 45). In this way, under envi-ronmental conditions that are not permissive for Ler-depen-dent expression of LEE genes, the lack of CesT, as well as ofT3SS and other Ler-regulated elements, will prompt the deg-radation of NleH by endogenous proteases and probably thedegradation of other non-LEE-encoded effectors, whose ex-pression is not directly controlled by Ler. Of note, ClpP hasinstead been shown to play a positive role in controlling theexpression of LEE genes by regulating RpoS and GrlR levelsin EHEC (31). However, the slight increase of nleH transcrip-tion seen in the �grlR strain does not seem to account for thehigher levels of accumulated and secreted NleH seen in the C.rodentium grlR mutant. This suggests that GrlR could also belinked to the posttranslational control of NleH expression,since when expressed under the control of an inducible pro-moter, higher levels of NleH are still observed in the �grlRmutant than in the WT. It is interesting to note that thisputative posttranslational control does not apply for the Ler-regulated LEE-encoded effector EspG, whose protein levelsare not reduced in regulatory mutants when expressed from aninducible promoter.
Overall, these observations confirmed that NleH behaves asa T3S effector and prompted the experiments to address its
potential role during infection. Initial experiments to look atpedestal formation and actin accumulation underneath adher-ent bacteria on epithelial cells revealed that A/E lesion forma-tion was not affected in the absence of NleH (data not shown).This was not completely unexpected, as in addition to Tir onlyone effector encoded outside the LEE, EspFu/TccP, has beenshown to be required for pedestal formation (7, 22). We thentested the ability of the C. rodentium nleH mutant to colonizeC57BL/6 mice, which have become an accepted model for thestudy of the disease caused by A/E pathogens (48). In thisassay, the nleH mutant showed a moderate, but statisticallysignificant, reduction of about 1 log in its ability to colonize themouse colon at day 6 p.i.; however, this defect was overcome atday 10 p.i. This result suggests that NleH may play a role atearly stages of infection by C. rodentium, as has been suggestedfor the LEE-encoded effector EspG (29). It is worth noticingthat among all the effectors described to date, only Tir, EspZ/SepZ, NleA/EspI, and NleB have been shown to play a criticalrole in the colonization of the mouse colon by C. rodentium(15, 16, 27, 35, 49, 65). The other tested effector proteins havebeen shown to have only moderate effects on colonization oron the outcome of the disease (see the introduction). Consid-ering the diverse distribution of genetic variability that is beingobserved among different human and animal isolates of EHECO157:H7, non-O157 Shiga toxin-producing E. coli, EPEC, andatypical EPEC strains, it is tempting to speculate that thepotential redundancy between different effector proteins, dueeither to the existence of genes encoding highly similar pro-teins, as in the case of NleH in EPEC and EHEC, or tofunctional mimicry despite not sharing sequence similarity,may obscure the roles of some of them during infection.
Functional redundancy has been reported for EspG andEspG2, two effector proteins that share 43.5% identity inEPEC. Both EspG and EspG2 can independently activateRhoA in the host cell and increase paracellular permeability(43), besides being able to induce microtubule elimination(56). Map and EspF, two effector proteins that are not relatedat the sequence level, participate in EPEC-mediated disrup-tion of epithelial barrier function, which can occur in the ab-sence of Map or EspF but completely disappears in a map espFdouble mutant (13). Furthermore, the results obtained duringthe in vivo evaluation of effector gene mutants may vary de-pending on the animal model employed. For example, a C.rodentium espH mutant colonizes and causes hyperplasia aswell as the WT strain in mice (49); however, the inactivation ofespH in EHEC affects its capacity to colonize the intestinaltracts of baby rabbits and reduces the severity of diarrheacaused in this model (52). Due to differences among A/Epathogens and specificity for their hosts, the analysis of differ-ent models may be necessary to identify the function of aparticular effector protein. Moreover, some effectors havebeen more frequently associated with A/E strains derived fromsevere cases of the disease. For example, NleE, which plays amoderate role in colonization and hyperplasia in the mousemodel of infection (35, 66), has been linked to diarrhea pro-duction by atypical EPEC strains (1) and to hemolytic-uremicsyndrome development and outbreaks by non-O157 EHECstrains (65). In a recent work, a microarray-based comparativegenomic hybridization technique was used to detect differencesin the genomic contents of 31 0157:H7 EHEC strains. Inter-
VOL. 190, 2008 NleH OF CITROBACTER RODENTIUM 2397
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
25
Oct
ober
202
1 by
119
.237
.38.
221.

estingly, the presence of nleH1, nleH2, and nleF was found tobe associated with lineage I strains, which are more frequentlylinked to human disease (67).
The role in disease of the ample collection of Nle proteins,either individually or in concert, in A/E pathogens may dependon several, but at least two, different aspects: the genetic rep-ertoire of each strain and the host’s characteristics. Theseelements may influence the pathogenic potential of the strains,including their ability to colonize different hosts. In addition,A/E pathogens may have implemented different mechanismsto coordinate and control the expression of these effector pro-teins. The tremendous challenge we face in future studies is toelucidate the molecular mechanisms underlying the coordi-nated expression and function in pathogenesis and disease ofsuch an extended repertoire of effector proteins.
ACKNOWLEDGMENTS
We thank F. J. Santana, A. Huerta-Saquero, and R. Banos-Lara fortechnical assistance and V. H. Bustamante for helpful discussions andcritical reading of the manuscript.
V.A.G.-A. was supported by a Ph.D. fellowship from CONACyT(157392) and a Hugo Arechiga Urtuzuastegui fellowship from theColegio de Sinaloa. This research was supported by grants from theCanadian Institutes of Health Research (CIHR) and the HowardHughes Medical Institute (HHMI) to B.B.F. and from the DireccionGeneral de Asuntos del Personal Academico (DGAPA-PAPIITIN224107 and IN201703-3), Consejo Nacional de Ciencia y Tecnologıa(CONACyT 42918Q), and HHMI to J.L.P.
REFERENCES
1. Afset, J. E., G. Bruant, R. Brousseau, J. Harel, E. Anderssen, L. Bevanger,and K. Bergh. 2006. Identification of virulence genes linked with diarrheadue to atypical enteropathogenic Escherichia coli by DNA microarray anal-ysis and PCR. J. Clin. Microbiol. 44:3703–3711.
2. Asakura, M., A. Hinenoya, M. S. Alam, K. Shima, S. H. Zahid, L. Shi, N.Sugimoto, A. N. Ghosh, T. Ramamurthy, S. M. Faruque, G. B. Nair, and S.Yamasaki. 2007. An inducible lambdoid prophage encoding cytolethal dis-tending toxin (Cdt-I) and a type III effector protein in enteropathogenicEscherichia coli. Proc. Natl. Acad. Sci. USA 104:14483–14488.
3. Barba, J., V. H. Bustamante, M. A. Flores-Valdez, W. Deng, B. B. Finlay, andJ. L. Puente. 2005. A positive regulatory loop controls expression of the locusof enterocyte effacement-encoded regulators Ler and GrlA. J. Bacteriol.187:7918–7930.
4. Bielaszewska, M., A. K. Sonntag, M. A. Schmidt, and H. Karch. 2007.Presence of virulence and fitness gene modules of enterohemorrhagic Esch-erichia coli in atypical enteropathogenic Escherichia coli O26. MicrobesInfect. 9:891–897.
5. Bustamante, V. H., F. J. Santana, E. Calva, and J. L. Puente. 2001. Tran-scriptional regulation of type III secretion genes in enteropathogenic Esch-erichia coli: Ler antagonizes H-NS-dependent repression. Mol. Microbiol.39:664–678.
6. Butler, S. M., R. A. Festa, M. J. Pearce, and K. H. Darwin. 2006. Self-compartmentalized bacterial proteases and pathogenesis. Mol. Microbiol.60:553–562.
7. Campellone, K. G., D. Robbins, and J. M. Leong. 2004. EspFU is a trans-located EHEC effector that interacts with Tir and N-WASP and promotesNck-independent actin assembly. Dev. Cell 7:217–228.
8. Charpentier, X., and E. Oswald. 2004. Identification of the secretion andtranslocation domain of the enteropathogenic and enterohemorrhagic Esch-erichia coli effector Cif, using TEM-1 beta-lactamase as a new fluorescence-based reporter. J. Bacteriol. 186:5486–5495.
9. Creuzburg, K., J. Recktenwald, V. Kuhle, S. Herold, M. Hensel, and H.Schmidt. 2005. The Shiga toxin 1-converting bacteriophage BP-4795 encodesan NleA-like type III effector protein. J. Bacteriol. 187:8494–8498.
10. Creuzburg, K., and H. Schmidt. 2007. Molecular characterization and dis-tribution of genes encoding members of the type III effector NleA familyamong pathogenic Escherichia coli strains. J. Clin. Microbiol. 45:2498–2507.
11. Dahan, S., S. Wiles, R. M. La Ragione, A. Best, M. J. Woodward, M. P.Stevens, R. K. Shaw, Y. Chong, S. Knutton, A. Phillips, and G. Frankel.2005. EspJ is a prophage-carried type III effector protein of attaching andeffacing pathogens that modulates infection dynamics. Infect. Immun. 73:679–686.
12. Datsenko, K. A., and B. L. Wanner. 2000. One-step inactivation of chromo-
somal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad.Sci. USA 97:6640–6645.
13. Dean, P., and B. Kenny. 2004. Intestinal barrier dysfunction by enteropatho-genic Escherichia coli is mediated by two effector molecules and a bacterialsurface protein. Mol. Microbiol. 54:665–675.
14. Deng, W., Y. Li, P. R. Hardwidge, E. A. Frey, R. A. Pfuetzner, S. Lee, S.Gruenheid, N. C. Strynakda, J. L. Puente, and B. B. Finlay. 2005. Regulationof type III secretion hierarchy of translocators and effectors in attaching andeffacing bacterial pathogens. Infect. Immun. 73:2135–2146.
15. Deng, W., J. L. Puente, S. Gruenheid, Y. Li, B. A. Vallance, A. Vazquez, J.Barba, J. A. Ibarra, P. O’Donnell, P. Metalnikov, K. Ashman, S. Lee, D.Goode, T. Pawson, and B. B. Finlay. 2004. Dissecting virulence: systematicand functional analyses of a pathogenicity island. Proc. Natl. Acad. Sci. USA101:3597–3602.
16. Deng, W., B. A. Vallance, Y. Li, J. L. Puente, and B. B. Finlay. 2003.Citrobacter rodentium translocated intimin receptor (Tir) is an essential vir-ulence factor needed for actin condensation, intestinal colonization andcolonic hyperplasia in mice. Mol. Microbiol. 48:95–115.
17. Dziva, F., P. M. van Diemen, M. P. Stevens, A. J. Smith, and T. S. Wallis.2004. Identification of Escherichia coli O157:H7 genes influencing coloniza-tion of the bovine gastrointestinal tract using signature-tagged mutagenesis.Microbiology 150:3631–3645.
18. Edwards, R. A., L. H. Keller, and D. M. Schifferli. 1998. Improved allelicexchange vectors and their use to analyze 987P fimbria gene expression.Gene 207:149–157.
19. Ehrbar, K., and W. D. Hardt. 2005. Bacteriophage-encoded type III effectorsin Salmonella enterica subspecies 1 serovar Typhimurium. Infect. Genet.Evol. 5:1–9.
20. Elliott, S. J., V. Sperandio, J. A. Giron, S. Shin, J. L. Mellies, L. Wainwright,S. W. Hutcheson, T. K. McDaniel, and J. B. Kaper. 2000. The locus ofenterocyte effacement (LEE)-encoded regulator controls expression of bothLEE- and non-LEE-encoded virulence factors in enteropathogenic and en-terohemorrhagic Escherichia coli. Infect. Immun. 68:6115–6126.
21. Garmendia, J., G. Frankel, and V. F. Crepin. 2005. Enteropathogenic andenterohemorrhagic Escherichia coli infections: translocation, translocation,translocation. Infect. Immun. 73:2573–2585.
22. Garmendia, J., A. D. Phillips, M. F. Carlier, Y. Chong, S. Schuller, O.Marches, S. Dahan, E. Oswald, R. K. Shaw, S. Knutton, and G. Frankel.2004. TccP is an enterohaemorrhagic Escherichia coli O157:H7 type IIIeffector protein that couples Tir to the actin-cytoskeleton. Cell. Microbiol.6:1167–1183.
23. Gauthier, A., M. de Grado, and B. B. Finlay. 2000. Mechanical fractionationreveals structural requirements for enteropathogenic Escherichia coli Tirinsertion into host membranes. Infect. Immun. 68:4344–4348.
24. Gauthier, A., J. L. Puente, and B. B. Finlay. 2003. Secretin of the entero-pathogenic Escherichia coli type III secretion system requires components ofthe type III apparatus for assembly and localization. Infect. Immun. 71:3310–3319.
25. Ghosh, P. 2004. Process of protein transport by the type III secretion system.Microbiol. Mol. Biol. Rev. 68:771–795.
26. Gomez-Duarte, O. G., and J. B. Kaper. 1995. A plasmid-encoded regulatoryregion activates chromosomal eaeA expression in enteropathogenic Esche-richia coli. Infect. Immun. 63:1767–1776.
27. Gruenheid, S., I. Sekirov, N. A. Thomas, W. Deng, P. O’Donnell, D. Goode,Y. Li, E. A. Frey, N. F. Brown, P. Metalnikov, T. Pawson, K. Ashman, andB. B. Finlay. 2004. Identification and characterization of NleA, a non-LEE-encoded type III translocated virulence factor of enterohaemorrhagic Esch-erichia coli O157:H7. Mol. Microbiol. 51:1233–1249.
28. Haack, K. R., C. L. Robinson, K. J. Miller, J. W. Fowlkes, and J. L. Mellies.2003. Interaction of Ler at the LEE5 (tir) operon of enteropathogenic Esch-erichia coli. Infect. Immun. 71:384–392.
29. Hardwidge, P. R., W. Deng, B. A. Vallance, I. Rodriguez-Escudero, V. J. Cid,M. Molina, and B. B. Finlay. 2005. Modulation of host cytoskeleton functionby the enteropathogenic Escherichia coli and Citrobacter rodentium effectorprotein EspG. Infect. Immun. 73:2586–2594.
30. Hayward, R. D., J. M. Leong, V. Koronakis, and K. G. Campellone. 2006.Exploiting pathogenic Escherichia coli to model transmembrane receptorsignalling. Nat. Rev. Microbiol. 4:358–370.
31. Iyoda, S., and H. Watanabe. 2005. ClpXP protease controls expression of thetype III protein secretion system through regulation of RpoS and GrlR levelsin enterohemorrhagic Escherichia coli. J. Bacteriol. 187:4086–4094.
32. Iyoda, S., and H. Watanabe. 2004. Positive effects of multiple pch genes onexpression of the locus of enterocyte effacement genes and adherence ofenterohaemorrhagic Escherichia coli O157:H7 to HEp-2 cells. Microbiology150:2357–2571.
33. Kaper, J. B., J. P. Nataro, and H. L. Mobley. 2004. Pathogenic Escherichiacoli. Nat. Rev. Microbiol. 2:123–140.
34. Karmali, M. A., M. Mascarenhas, S. Shen, K. Ziebell, S. Johnson, R. Reid-Smith, J. Isaac-Renton, C. Clark, K. Rahn, and J. B. Kaper. 2003. Associ-ation of genomic O island 122 of Escherichia coli EDL 933 with verocyto-toxin-producing Escherichia coli seropathotypes that are linked to epidemicand/or serious disease. J. Clin. Microbiol. 41:4930–4940.
2398 GARCIA-ANGULO ET AL. J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
25
Oct
ober
202
1 by
119
.237
.38.
221.

35. Kelly, M., E. Hart, R. Mundy, O. Marches, S. Wiles, L. Badea, S. Luck, M.Tauschek, G. Frankel, R. M. Robins-Browne, and E. L. Hartland. 2006.Essential role of the type III secretion system effector NleB in colonizationof mice by Citrobacter rodentium. Infect. Immun. 74:2328–2337.
36. Kim, D. W., G. Lenzen, A. L. Page, P. Legrain, P. J. Sansonetti, and C.Parsot. 2005. The Shigella flexneri effector OspG interferes with innate im-mune responses by targeting ubiquitin-conjugating enzymes. Proc. Natl.Acad. Sci. USA 102:14046–14051.
37. Kujat Choy, S. L., E. C. Boyle, O. Gal-Mor, D. L. Goode, Y. Valdez, B. A.Vallance, and B. B. Finlay. 2004. SseK1 and SseK2 are novel translocatedproteins of Salmonella enterica serovar Typhimurium. Infect. Immun. 72:5115–5125.
38. Levine, M. M., E. J. Bergquist, D. R. Nalin, D. H. Waterman, R. B. Hornick,C. R. Young, and S. Sotman. 1978. Escherichia coli strains that cause diar-rhoea but do not produce heat-labile or heat-stable enterotoxins and arenon-invasive. Lancet i:1119–1122.
39. Li, M., I. Rosenshine, H. B. Yu, C. Nadler, E. Mills, C. L. Hew, and K. Y.Leung. 2006. Identification and characterization of NleI, a new non-LEE-encoded effector of enteropathogenic Escherichia coli (EPEC). MicrobesInfect. 8:2890–2898.
40. Lio, J. C., and W. J. Syu. 2004. Identification of a negative regulator for thepathogenicity island of enterohemorrhagic Escherichia coli O157:H7.J. Biomed. Sci. 11:855–863.
41. Marches, O., S. Wiles, F. Dziva, R. M. La Ragione, S. Schuller, A. Best, A. D.Phillips, E. L. Hartland, M. J. Woodward, M. P. Stevens, and G. Frankel.2005. Characterization of two non-locus of enterocyte effacement-encodedtype III-translocated effectors, NleC and NleD, in attaching and effacingpathogens. Infect. Immun. 73:8411–8417.
42. Martınez-Laguna, Y., E. Calva, and J. L. Puente. 1999. Autoactivation andenvironmental regulation of bfpT expression, the gene coding for the tran-scriptional activator of bfpA in enteropathogenic Escherichia coli. Mol. Mi-crobiol. 33:153–166.
43. Matsuzawa, T., A. Kuwae, and A. Abe. 2005. Enteropathogenic Escherichiacoli type III effectors EspG and EspG2 alter epithelial paracellular perme-ability. Infect. Immun. 73:6283–6289.
44. Mayer, M. P. 1995. A new set of useful cloning and expression vectorsderived from pBlueScript. Gene 163:41–46.
45. Mellies, J. L., A. M. Barron, and A. M. Carmona. 2007. Enteropathogenicand enterohemorrhagic Escherichia coli virulence gene regulation. Infect.Immun. 75:4199–4210.
46. Mellies, J. L., S. J. Elliott, V. Sperandio, M. S. Donnenberg, and J. B. Kaper.1999. The Per regulon of enteropathogenic Escherichia coli: identification ofa regulatory cascade and a novel transcriptional activator, the locus of en-terocyte effacement (LEE)-encoded regulator (Ler). Mol. Microbiol. 33:296–306.
47. Mellies, J. L., K. R. Haack, and D. C. Galligan. 2007. SOS regulation of thetype III secretion system of enteropathogenic Escherichia coli. J. Bacteriol.189:2863–2872.
48. Mundy, R., T. T. MacDonald, G. Dougan, G. Frankel, and S. Wiles. 2005.Citrobacter rodentium of mice and man. Cell. Microbiol. 7:1697–1706.
49. Mundy, R., L. Petrovska, K. Smollett, N. Simpson, R. K. Wilson, J. Yu, X.Tu, I. Rosenshine, S. Clare, G. Dougan, and G. Frankel. 2004. Identificationof a novel Citrobacter rodentium type III secreted protein, EspI, and roles ofthis and other secreted proteins in infection. Infect. Immun. 72:2288–2302.
50. Porter, M. E., P. Mitchell, A. J. Roe, A. Free, D. G. Smith, and D. L. Gally.2004. Direct and indirect transcriptional activation of virulence genes by anAraC-like protein, PerA from enteropathogenic Escherichia coli. Mol. Mi-crobiol. 54:1117–1133.
51. Reading, N. C., A. G. Torres, M. M. Kendall, D. T. Hughes, K. Yamamoto,and V. Sperandio. 2007. A novel two-component signaling system that acti-vates transcription of an enterohemorrhagic Escherichia coli effector in-volved in remodeling of host actin. J. Bacteriol. 189:2468–2476.
52. Ritchie, J. M., and M. K. Waldor. 2005. The locus of enterocyte effacement-
encoded effector proteins all promote enterohemorrhagic Escherichia colipathogenicity in infant rabbits. Infect. Immun. 73:1466–1474.
53. Roe, A. J., L. Tysall, T. Dransfield, D. Wang, D. Fraser-Pitt, A. Mahajan, C.Constandinou, N. Inglis, A. Downing, R. Talbot, D. G. Smith, and D. L.Gally. 2007. Analysis of the expression, regulation and export of NleA-E inEscherichia coli O157:H7. Microbiology 153:1350–1360.
54. Sanchez-SanMartin, C., V. H. Bustamante, E. Calva, and J. L. Puente. 2001.Transcriptional regulation of the orf19 gene and the tir-cesT-eae operon ofenteropathogenic Escherichia coli. J. Bacteriol. 183:2823–2833.
55. Schauer, D. B., and S. Falkow. 1993. Attaching and effacing locus of aCitrobacter freundii biotype that causes transmissible murine colonic hyper-plasia. Infect. Immun. 61:2486–2492.
56. Shaw, R. K., K. Smollett, J. Cleary, J. Garmendia, A. Straatman-Iwanowska,G. Frankel, and S. Knutton. 2005. Enteropathogenic Escherichia coli type IIIeffectors EspG and EspG2 disrupt the microtubule network of intestinalepithelial cells. Infect. Immun. 73:4385–4390.
57. Sory, M. P., A. Boland, I. Lambermont, and G. R. Cornelis. 1995. Identifi-cation of the YopE and YopH domains required for secretion and internal-ization into the cytosol of macrophages, using the cyaA gene fusion ap-proach. Proc. Natl. Acad. Sci. USA 92:11998–12002.
58. Tampakaki, A. P., V. E. Fadouloglou, A. D. Gazi, N. J. Panopoulos, and M.Kokkinidis. 2004. Conserved features of type III secretion. Cell. Microbiol.6:805–816.
59. Thomas, N. A., W. Deng, N. T. Baker, J. L. Puente, and B. B. Finlay. 2007.Hierarchical delivery of an essential host colonization factor in enteropatho-genic Escherichia coli. J. Biol. Chem. 282:29634–29645.
60. Thomas, N. A., W. Deng, J. L. Puente, E. A. Frey, C. K. Yip, N. C. Strynadka,and B. B. Finlay. 2005. CesT is a multi-effector chaperone and recruitmentfactor required for the efficient type III secretion of both LEE- and non-LEE-encoded effectors of enteropathogenic Escherichia coli. Mol. Microbiol.57:1762–1779.
61. Tobe, T., S. A. Beatson, H. Taniguchi, H. Abe, C. M. Bailey, A. Fivian, R.Younis, S. Matthews, O. Marches, G. Frankel, T. Hayashi, and M. J. Pallen.2006. An extensive repertoire of type III secretion effectors in Escherichiacoli O157 and the role of lambdoid phages in their dissemination. Proc. Natl.Acad. Sci. USA 103:14941–14946.
62. Torres, A. G., L. Milflores-Flores, J. G. Garcia-Gallegos, S. D. Patel, A. Best,R. M. La Ragione, Y. Martinez-Laguna, and M. J. Woodward. 2007. Envi-ronmental regulation and colonization attributes of the long polar fimbriae(LPF) of Escherichia coli O157:H7. Int. J. Med. Microbiol. 297:177–185.
63. Umanski, T., I. Rosenshine, and D. Friedberg. 2002. Thermoregulated ex-pression of virulence genes in enteropathogenic Escherichia coli. Microbiol-ogy 148:2735–2744.
64. Vlisidou, I., O. Marches, F. Dziva, R. Mundy, G. Frankel, and M. P. Stevens.2006. Identification and characterization of EspK, a type III secreted effectorprotein of enterohaemorrhagic Escherichia coli O157:H7. FEMS Microbiol.Lett. 263:32–40.
65. Wickham, M. E., C. Lupp, M. Mascarenhas, A. Vazquez, B. K. Coombes,N. F. Brown, B. A. Coburn, W. Deng, J. L. Puente, M. A. Karmali, and B. B.Finlay. 2006. Bacterial genetic determinants of non-O157 STEC outbreaksand hemolytic-uremic syndrome after infection. J. Infect. Dis. 194:819–827.
66. Wickham, M. E., C. Lupp, A. Vazquez, M. Mascarenhas, B. Coburn, B. K.Coombes, M. A. Karmali, J. L. Puente, W. Deng, and B. B. Finlay. 2007.Citrobacter rodentium virulence in mice associates with bacterial load and thetype III effector NleE. Microbes Infect. 9:400–407.
67. Zhang, Y., C. Laing, M. Steele, K. Ziebell, R. Johnson, A. K. Benson, E.Taboada, and V. P. Gannon. 2007. Genome evolution in major Escherichiacoli O157:H7 lineages. BMC Genomics 8:121–136.
68. Zhu, C., S. Feng, T. E. Thate, J. B. Kaper, and E. C. Boedeker. 2006. Towardsa vaccine for attaching/effacing Escherichia coli: a LEE encoded regulator(ler) mutant of rabbit enteropathogenic Escherichia coli is attenuated, im-munogenic, and protects rabbits from lethal challenge with the wild-typevirulent strain. Vaccine 24:3845–3855.
VOL. 190, 2008 NleH OF CITROBACTER RODENTIUM 2399
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
25
Oct
ober
202
1 by
119
.237
.38.
221.
Related Documents











