Regulation of ciliary motility: conserved protein kinases and phosphatases are targeted and anchored in the ciliary axoneme Maureen Wirschell a , Ryosuke Yamamoto a , Lea Alford a , Avanti Gokhale a , Anne Gaillard b , and Winfield S. Sale a,c Maureen Wirschell: [email protected]; Ryosuke Yamamoto: [email protected]; Lea Alford: [email protected]; Avanti Gokhale: [email protected]; Anne Gaillard: [email protected] a Emory University School of Medicine, Department of Cell Biology, 615 Michael St. Atlanta, GA USA 30322 b Sam Houston State University, Department of Biological Sciences, 1900 Ave. I, P.O Box 2116, Huntsville, TX USA 77341 Abstract Recent evidence has revealed that the dynein motors and highly conserved signaling proteins are localized within the ciliary 9 + 2 axoneme. One key mechanism for regulation of motility is phosphorylation. Here, we review diverse evidence, from multiple experimental organisms, that ciliary motility is regulated by phosphorylation / dephosphorylation of the dynein arms through kinases and phosphatases that are anchored immediately adjacent to their axonemal substrates. 1. Introduction Understanding the mechanism of ciliary / flagellar movement is crucial: motile cilia play essential roles in human development, male and female fertility, protection and function of the airway and circulation of cerebral spinal fluid [1, 2]. Defects in ciliary motility lead to a variety of “ciliopathies” that can result in an especially wide range of diseases and syndromes including left-right pattern defects in early development, hydrocephaly, infertility and Primary Ciliary Dyskinesia (PCD) [3–9]. Furthermore, defective assembly of the “primary” cilium, an immotile, sensory and signal transduction organelle found on all differentiated cells, can lead to additional problems including failure in normal patterning, polycystic kidney disease, retinal degeneration, obesity, skeletal abnormalities and a growing list of syndromes [10–13]. Importantly, the cytoskeletal structure of the motile cilium, the axoneme, is a highly conserved 9 + 2 arrangement of microtubules, associated dynein motors and structures and signaling molecules which contribute to control of ciliary movement [14, 15]. Normal ciliary motility is a consequence of precisely controlled dynein c corresponding author: Winfield S. Sale, Emory University School of Medicine, Whitehead Research Building, Ste 465, 615 Michael St., Atlanta, GA USA, (404)-727-6265 Phone, (404)-727-6256 Fax, [email protected], [email protected]. Suggested Reviewers: Susan K. Dutcher, Professor, Department of Genetics, Department of Cell Biology and Physiology, Washington University School of Medicine, Box 8232, 660 S. Euclid Avenue, St. Louis, MO 63110, [email protected], 314-362-2765 Stephen M. King, Professor, Molecular, Microbial & Structural Biology, University of Connecticut Health Center, 263 Farmington Avenue, Farmington, Connecticut 06030-3305, Tel: (860) 679 3347, Fax: (860) 679 3408, [email protected] or [email protected] Elizabeth F. Smith, Professor, 7 Lucent Drive, Room 106, Dartmouth, Biology, Centerra BioLabs, Lebanon, NH 03766, [email protected], 603-646-1129 Greg Pazour, Associate Professor, Program in Molecular Medicine, UMass Medical School, Suite 213, Biotech I, 373 Plantation Street, Worcester, MA 01605, [email protected], (T) 508 856 8078: (F) 508 856 2950 Dr. Peter Satir, Department of Anatomy & Structural Biology, Forchheimer 602, Morris Park Avenue, Bronx NY 10461 – USA, [email protected], phone: (718)-430-4061, fax: (718)-430-8964 NIH Public Access Author Manuscript Arch Biochem Biophys. Author manuscript; available in PMC 2012 June 15. Published in final edited form as: Arch Biochem Biophys. 2011 June 15; 510(2): 93–100. doi:10.1016/j.abb.2011.04.003. NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Regulation of ciliary motility: conserved protein kinases andphosphatases are targeted and anchored in the ciliary axoneme
Maureen Wirschella, Ryosuke Yamamotoa, Lea Alforda, Avanti Gokhalea, Anne Gaillardb,and Winfield S. Salea,c
Maureen Wirschell: [email protected]; Ryosuke Yamamoto: [email protected]; Lea Alford:[email protected]; Avanti Gokhale: [email protected]; Anne Gaillard: [email protected] University School of Medicine, Department of Cell Biology, 615 Michael St. Atlanta, GAUSA 30322bSam Houston State University, Department of Biological Sciences, 1900 Ave. I, P.O Box 2116,Huntsville, TX USA 77341
AbstractRecent evidence has revealed that the dynein motors and highly conserved signaling proteins arelocalized within the ciliary 9 + 2 axoneme. One key mechanism for regulation of motility isphosphorylation. Here, we review diverse evidence, from multiple experimental organisms, thatciliary motility is regulated by phosphorylation / dephosphorylation of the dynein arms throughkinases and phosphatases that are anchored immediately adjacent to their axonemal substrates.
1. IntroductionUnderstanding the mechanism of ciliary / flagellar movement is crucial: motile cilia playessential roles in human development, male and female fertility, protection and function ofthe airway and circulation of cerebral spinal fluid [1, 2]. Defects in ciliary motility lead to avariety of “ciliopathies” that can result in an especially wide range of diseases andsyndromes including left-right pattern defects in early development, hydrocephaly, infertilityand Primary Ciliary Dyskinesia (PCD) [3–9]. Furthermore, defective assembly of the“primary” cilium, an immotile, sensory and signal transduction organelle found on alldifferentiated cells, can lead to additional problems including failure in normal patterning,polycystic kidney disease, retinal degeneration, obesity, skeletal abnormalities and agrowing list of syndromes [10–13]. Importantly, the cytoskeletal structure of the motilecilium, the axoneme, is a highly conserved 9 + 2 arrangement of microtubules, associateddynein motors and structures and signaling molecules which contribute to control of ciliarymovement [14, 15]. Normal ciliary motility is a consequence of precisely controlled dynein
ccorresponding author: Winfield S. Sale, Emory University School of Medicine, Whitehead Research Building, Ste 465, 615 MichaelSt., Atlanta, GA USA, (404)-727-6265 Phone, (404)-727-6256 Fax, [email protected], [email protected] Reviewers:Susan K. Dutcher, Professor, Department of Genetics, Department of Cell Biology and Physiology, Washington University School ofMedicine, Box 8232, 660 S. Euclid Avenue, St. Louis, MO 63110, [email protected], 314-362-2765Stephen M. King, Professor, Molecular, Microbial & Structural Biology, University of Connecticut Health Center, 263 FarmingtonAvenue, Farmington, Connecticut 06030-3305, Tel: (860) 679 3347, Fax: (860) 679 3408, [email protected] [email protected] F. Smith, Professor, 7 Lucent Drive, Room 106, Dartmouth, Biology, Centerra BioLabs, Lebanon, NH 03766,[email protected], 603-646-1129Greg Pazour, Associate Professor, Program in Molecular Medicine, UMass Medical School, Suite 213, Biotech I, 373 PlantationStreet, Worcester, MA 01605, [email protected], (T) 508 856 8078: (F) 508 856 2950Dr. Peter Satir, Department of Anatomy & Structural Biology, Forchheimer 602, Morris Park Avenue, Bronx NY 10461 – USA,[email protected], phone: (718)-430-4061, fax: (718)-430-8964
NIH Public AccessAuthor ManuscriptArch Biochem Biophys. Author manuscript; available in PMC 2012 June 15.
Published in final edited form as:Arch Biochem Biophys. 2011 June 15; 510(2): 93–100. doi:10.1016/j.abb.2011.04.003.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

motors and the mechanical and chemical interplay of axonemal structural and enzymaticcomponents that regulate beating of the cilium. Thus, discovery of the conservedmechanisms that regulate the dynein motors is essential for understanding ciliary bending.
Here, we focus, specifically, on the key ciliary axonemal structures and conserved axonemalprotein kinases and phosphatases that regulate ciliary movement. We review diverseevidence from multiple experimental systems revealing that ciliary motility is regulated byphosphorylation and that the key kinases and phosphatases are conserved, targeted to andanchored in the axoneme in positions to control motility [16]. We feature studies using themodel genetic organism Chlamydomonas, that has not only revealed conserved genes thatencode proteins essential for ciliary assembly and function, but that has also provided a verypowerful experimental system for study of the mechanism and regulation of the dyneinmotors that control motility [17]. For example, taking advantage of isolated axonemes frominformative Chlamydomonas ciliary mutants, a microtubule sliding assay has helpeddemonstrate that protein kinases and phosphatases in the axoneme control dynein activitythrough phosphorylation [16].
Also supporting regulation of ciliary motility by phosphorylation is the finding that kinasesand phosphatases are a physical part of the axonemal structure [14, 18]. We focus on thepharmacological, biochemical and molecular approaches that revealed the proteinphosphatases, PP1 and PP2A, and kinases, CK1 and PKA, are anchored in the axoneme andcontrol movement [19–24]. We summarize how new proteomic analyses of cilia andaxonemes have revealed additional conserved kinases, phosphatases, calcium-bindingproteins and specific phosphoproteins in the motile cilia [25, 26] see also(http://v3.ciliaproteome.org/cgi-bin/index.php) and discuss opportunities for new researchcharacterizing the role of kinases and phosphatases in regulation of motility.
2. Conserved axonemal structures and regulation of motilityCilia, sometimes called flagella, are microtubule-based structures that are highly conservedin protein composition as well as structural organization [2, 9]. Indeed, recent studies haveled to a revolution in our understanding of the diverse, vital roles of cilia and revealed thepresence of cilia on nearly all differentiated cells in vertebrates [8, 11, 27–29]. In addition tomotility, cilia display an extraordinary range of vital signaling roles required for normalcontrol of the cell division cycle, sensory transduction required for vertebrate developmentand adult functions [10, 12, 30–32]. We focus on motile cilia and the 9+2 axoneme that notonly localizes a number of distinct dynein motors required for motility, but also localizesubiquitous kinases and phosphatases required for regulation of ciliary motility [14, 16, 19,33]. As indicated in the Introduction, defects in either assembly or function of cilia canresult in a wide spectrum of diseases, termed “ciliopathies” [8, 10, 34–38]. Despite theconservation, we are only beginning to understand how dyneins and ciliary motility areregulated.
Motile cilia contain a bundle of nine outer doublet microtubules and two centralmicrotubules called the 9+2 axoneme (Fig. 1). The ciliary axoneme originates from the basalbody which provides a template for microtubule growth and is required for assembly of allcilia, anchoring the 9+2 axoneme to the apical end of the cell [39–42]. The central pair andthe nine outer doublet microtubules are continuous throughout the length of the axoneme.Each outer doublet appears to be associated with the adjacent outer doublet by the dyneinarms and structures called nexins recently determined to be part of the dynein regulatorycomplex – (DRC) [43]. The 9+2 structural configuration is highly conserved among allciliated eukaryotes [25, 44–46] and the basis of this conservation appears to be related to theconservation of basal body organization.
Wirschell et al. Page 2
Arch Biochem Biophys. Author manuscript; available in PMC 2012 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Ultrastructural analyses have revealed axonemal structures in fine detail (Fig. 1). Whenviewed along the longitudinal axis, the complexity of the axonemal structures is appreciatedin greater detail (Fig. 2). Each component associated with the outer doublet microtubule isorganized in regular, repeating 96-nm units that are best illustrated using cryoEMtomography of intact axonemes [43, 47–52] which revealed unprecedented fine detail inaxonemal morphology. In each 96-nm axonemal repeat, there are four outer arm dyneinsthat repeat at 24 nm and a surprisingly complex collection of at least seven different innerdynein arm structures, including the inner dynein arm called “I1 dynein” thought to regulatebending by a mechanism that includes phosphorylation and associated kinases andphosphatases [14, 19]. Although the 96-nm repeat organization is highly conserved, themolecular basis that establishes this organization is unknown. Understanding the basis forthe 96 nm repeat structure is a high priority since it not only establishes the precise locationof dynein motors, radial spokes and DRC, it must also provide the foundation for targetingand anchoring of the protein kinases, phosphatases and calmodulin complexes localized tothe outer doublet microtubule (Fig. 3).
The central pair and radial spokes are essential for normal control of motility. The centralpair apparatus is composed of the “C1” and “C2” microtubules and associated structures[47, 50, 51, 53–60]. The asymmetric structure of the central pair is founded on the distinctcomposition of components associated with the C1 and C2 microtubules includingubiquitous and conserved kinases and phosphatases, such as PKA (and associated A-kinaseanchoring proteins-AKAPs), PP1, as well as calmodulin-containing complexes important forcalcium-based regulatory pathways (Fig. 3; [60–63]). The asymmetric nature of the centralpair apparatus is a fundamental feature for control of ciliary movement; in some motile cilia,the central pair rotates, directing signals through the radial spokes to specific outer doubletson one side of the axoneme (Fig. 1). Signaling proteins and complexes, localized to the outerdoublets (Fig. 3), relay the signals from the central pair-radial spokes to the dynein motors,ultimately impacting dynein motor function [14, 19, 60]. The signaling molecules localizedto the outer doublets include kinases (CK1 and PKA), phosphatases (PP1 and PP2A) and theDRC—an axonemal structure also important for control of dynein activity [33]
The radial spokes are T-shaped structures anchored to the A microtubule of the outerdoublets that project towards the central pair. There are two radial spokes, termed S1 andS2, in each 96-nm repeat (spoke S1 is most proximal). Radial spokes are a highly conservedcollection of at least 23 proteins found in all ciliated organisms and required for motility andregulation of the ciliary dyneins [64–70]. One radial spoke protein (RSP3) anchors the radialspokes to the A-microtubule [71, 72]. Moreover, RSP3 has been identified as an AKAP [22,23]. Therefore, as discussed below, one predicted function of RSP3 is to anchor the proteinkinase, PKA, in the axoneme, in position to regulate the kinase activity by physicallylocalizing it near its substrate including the dynein motors [23]. The radial spokes alsocontain calmodulin as part of their structure [69, 73] and interact with a novel calmodulincomplex located at the base of the radial spokes on the outer doublet microtubule (Fig. 3;[62]).
The DRC is a part of a mechano-chemical pathway that regulates dynein activity and acts asthe nexin interdoublet linker (Fig. 2; [43, 74–80]. Given its location, one prediction is thatthe DRC plays a role in mediating signals between spoke S2 and the dynein motors. Thenature of such signals and the exact function of each of the DRC subunit are only justbeginning to be defined [33, 77, 78, 80–84]. Challenges include identification and functionalcharacterization of individual DRC subunits and testing the idea that the DRC, in part,localizes kinases and phosphatases required for regulation of dynein activity.
Wirschell et al. Page 3
Arch Biochem Biophys. Author manuscript; available in PMC 2012 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

The axoneme bears at least eight different axonemal dynein motors, each localized to aunique position in the 96 nm repeat and apparently responsible for distinct features of ciliarymovement (Fig. 3; [85]). Most generally, the dynein arm structures are categorized in tworows – the outer dynein arms (ODA) and the inner dynein arms (IDA; Fig. 1 and 2). Theouter and inner rows of dyneins are structurally and functionally distinct: the outer dyneinarms are homogenous in composition and structure, whereas, the inner dynein arms arecomplex, composed of at least seven distinct dynein isoforms each localized in a fixedpattern in the 96-nm repeat (Fig 2). Studies in several experimental systems indicate ciliarybeat frequency is regulated by the ODA and that beat frequency can be regulated byphosphorylation [18]. In addition, phosphorylation plays a role in control of the inner dyneinarms and regulation of axonemal bending. In this article, we feature the inner arm dyneincalled I1dynein: data has revealed I1 dynein displays unusual properties and is required fornormal control of the size and shape of the ciliary/ flagellar bend (i.e. the amplitude and thedegree of curvature of flagellar bending) [19, 86]. Genetic and biochemical analysis revealsthat the key I1 dynein regulatory phospho-protein is IC138 [21, 87–90]. Notably, the keykinases and phosphatases that control I1 dynein are physically localized to the axoneme.Therefore, we argue these enzymes must be relatively abundant, targeted and anchored nearI1 dynein, regularly repeating every 96 nm on the doublet microtubules (Fig. 3).Alternatively, and of equal interest, the kinases and phosphatases that regulate IC138 may belocalized to subsets of doublet microtubules. This result would be quite important since itwould further address a functional axis of the axoneme (defined in Fig. 1) important formodulation of bending movement.
3. A “sliding microtubule-switching” model for oscillatory ciliary bendingBefore proceeding to a discussion of the axonemal kinases and phosphatases and how theymight regulate motility, it is important to describe a working model for ciliary bending (Fig1 and 4). Seminal work, using multiple experimental systems including isolated motileaxonemes, has revealed that the basis for ciliary motility lies in controlled dynein-drivenmicrotubule sliding- a “sliding microtubule” model for ciliary bending [91–96]. In addition,it was determined that the dynein motors generate force in one direction (minus-enddirection) relative to microtubule polarity [97, 98]. Thus, since all dyneins are minus-endmotors, this observation directly led to a “switching” model for alternating effective(principal/forward) and recovery (reverse) bending: dyneins on one side of a structural andfunctional axis (Fig. 1) of the axoneme are active for bending in one direction. Dyneinactivity and associated microtubule sliding are regulated, in part, by a phospho-regulatorymechanism (described below) that includes the central pair and radial spoke structures. Onemodel indicates that the central pair and its associated appendages acts as a distributor [60,99], selectively stimulating specific radial spokes to alter dynein activity on specific outerdoublets.
Predictably, when the direction of bending changes or “switches”, there is a switch inactivity where the previously inactive dyneins, on one side of the axis, are activated, and theformerly active dyneins on the opposite side, are turned off [100, 101]. The precisemechanism for sensing the end point of each bend direction is not understood, but clearly foroscillatory bending, dynein motors must be regulated. Detailed experimental evidenceindicates the basic oscillatory movement and “switches” in bend direction are inherentproperties of the dynein motors involving microtubule curvature and a mechanical feedbackcontrol [102, 103]. The switching sensor is not known, but Patel-King and King (2009)propose a model in which the outer dynein arm plays a role as a sensor of microtubulecurvature [104]. Thus, the phospho-regulatory mechanism discussed in this chapter is notresponsible for control of the basic oscillatory bending in ciliary axonemes, a mechanismthat does not appear to require any form of alternating posttranslational modification.
Wirschell et al. Page 4
Arch Biochem Biophys. Author manuscript; available in PMC 2012 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Rather, the phospho-regulatory mechanism discussed here is superimposed on the basicdynein – driven mechanism to modulate beat frequency or the size and shape of the bend,parameters we will refer to as ciliary waveform.
4. Kinases targeted and localized to the 9+2 axonemePKA, PKG and A-kinase anchoring proteins (AKAPs)
Both direct and indirect approaches have revealed that PKA and PKG are associated withand regulate axonemal motility by phosphorylation [18]. This includes a wide range ofstudies indicating that, for example, sperm motility is activated or altered by cyclicnucleotides [105–112]. However, only a few studies have focused on the kinases directlylocalized to the axoneme for control of motility. In most cases studied to date, the upstreamregulator of phosphorylation is calcium [18, 63, 113–115].
The focus on kinases associated with the axoneme began with biochemical andphysiological assays using isolated ciliary axonemes or detergent extracted “models” [116,117]. For example, classical studies using ciliary axonemes or detergent extractedTetrahymena and Paramecium cells revealed that cAMP and cGMP alter ATP –inducedciliary beating [114, 118, 119]. Consistent with the functional and pharmacological analysis,PKA has been isolated and characterized from axonemes [120] and may alter dynein activityin cilia from Paramecium [121]. Additional biochemical and functional evidence indicatedone of the axonemal substrates is a 29-kDa outer dynein arm light chain [122–124] and thatphosphorylation leads to increased ciliary beat frequency consistent with the known role ofthe outer dynein arms for control of beat frequency. In studies of mammalian respiratoryairways, cGMP and PKG have been shown to modulate ciliary beat frequency [115, 125–128], while, in Paramecium, cGMP alters the direction of ciliary beating and PKG–dependent phosphorylation of ciliary targets is required for normal ciliary motility [114,129, 130]. Furthermore, in studies of Corbicula fluminea, a freshwater clam, sperm motilityis reduced upon the addition of a pharmacological inhibitor of PKG [131].
Similar studies with ciliary axonemes from mammalian epithelia also reveal that outerdynein arm-mediated changes in beat frequency correlate with increased cyclic nucleotidesecond messengers [18, 127, 132–135] and that the ciliary axoneme bears AKAPs (seebelow [136]), PKA and PKG. However, to date, the regulatory phosphoprotein substratesare not defined with certainty in the mammalian axonemes, but the substrates must be partof a regulatory pathway that mediates outer dynein arm activity for control of ciliary beatfrequency and could include dynein subunits such as the 29 kDa subunit cited above or theouter arm dynein heavy chains [137]. Since these studies make use of the intact, isolated 9+2axoneme, predictably the kinases are targeted and anchored in the axoneme near thesubstrates by specialized scaffold proteins such as the AKAPs (discussed below).
Genetic and in vitro studies using isolated axonemes from Chlamydomonas have alsorevealed that inner arm dynein activity is regulated by phosphorylation and that the kinasesare located in the axoneme. Based upon functional assays and the use of the PKA inhibitor,PKI, one of the kinases is PKA [138], and, as discussed further below, another axonemalkinase that controls inner arm dynein phosphorylation is CK1 [21, 24]. Genetic andbiochemical evidence reveals that the dynein target is I1 dynein and its regulatory subunitIC138 (Fig. 5) and much experimentation has been devoted to their study [21, 87–90, 139,140]. Notably, this mechanism was revealed in axonemes defective in control of kinaseactivity. Mutations that result in failure of assembly of the central pair or radial spokes resultin ciliary paralysis. Part of the reason for paralysis is a failure in normal control of theaxonemal kinases that appear to become constitutively active in these mutants leading to aglobal inhibition of the dynein motors.
Wirschell et al. Page 5
Arch Biochem Biophys. Author manuscript; available in PMC 2012 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Chlamydomonas mutants that are defective in I1 dynein or exhibit hyperphosphorylatedIC138 display abnormal flagellar waveform, defective dynein – driven microtubule slidingand failure in cellular phototaxis [19]. As indicated in Figure 5, there appears to be a tightcorrelation between IC138 phosphorylation and regulation of microtubule sliding: inhibitionof microtubule sliding correlates with phosphorylation of IC138; active or rescuedmicrotubule sliding correlates with de-phosphorylation of IC138. We do not yet know howphosphorylation alters dynein activity. Much research in several labs is now focused on thedownstream kinases and phosphatases that regulate IC138 and I1 dynein. One of the kinasesis CK1 (see below), and to date, the only mechanism for localizing PKA in the axoneme isthrough AKAPs [141–143].
AKAPs are proteins that interact with the regulatory subunits of PKA and confer sub-cellular localization (134, 135) and have been identified in the axoneme [22, 23, 136]. Basedon biochemical analysis, the Chlamydomonas flagellar axoneme has at least two AKAPs:one located in the central pair apparatus (AKAP240) and the other located at the base of theradial spokes (RSP3) (Fig. 3; [22]). To further test the hypothesis that RSP3 is an AKAP andresponsible for regulation of flagellar motility, an RSP3 gene containing a mutation in thePKA-binding domain was transformed into a null mutant of RSP3 (pf14) and the motility ofthe transformants was analyzed [23]. The results indicated that disruption of the PKA-binding domain results in failure of motility in a manner that is consistent with a role forPKA, and precise localization of PKA, in control of flagellar motility. RSP3 is a highlyconserved protein of all motile cilia [72]: thus, predictably, the RSP3 AKAP plays aconserved regulatory role in all cilia and flagella. Further tests of these ideas now require theidentification and characterization of PKA in Chlamydomonas flagella and analysis of RSP3in cilia from other organisms. Moreover, these AKAPs are predicted to localize PKA in theaxonemal structure, placing PKA in position near its substrates, including IC138 of I1dynein.
An axonemal CK1 regulates dynein – driven microtubule slidingExperimental analysis of isolated Chlamydomonas ciliary axonemes has implicated theprotein kinase, CK1, in regulation of dynein-driven motility [21, 24]. CK1 is a highlyconserved serine/threonine kinase that has multiple cellular functions including regulation ofthe cell cycle, apoptosis, motility, organelle transport, circadian rhythms and regulation ofdevelopmental pathways such as the Wnt pathway [144–146]. CK1 kinases are monomericenzymes with constitutively active enzymatic activity, utilizing ATP as an exclusivephosphate donor (139). Several CK1 isoforms have been described in yeast and mammaliansystems. They share common signature domains including the ATP- and substrate-bindingdomains, the catalytic triad, nuclear localization signal and a kinesin-homology domain[146].
Using CK1-specific inhibitors, it was shown that the characteristic slow-sliding velocities ofparalyzed radial spoke mutant Chlamydomonas axonemes were restored to wild-type levelsand that IC138 phosphorylation / dephosphorylation correlates with control of microtubulesliding [21, 24]. In vitro assays using recombinant CK1 proteins demonstrate that anenzymatically functional CK1 is required to inhibit I1 dynein-dependent microtubule sliding[24]. CK1 is located in the flagellar axoneme in Chlamydomonas [21, 24, 25] where itfunctions as a downstream component of the central pair-radial spoke mechanism andtogether with I1 dynein, controls flagellar waveform.
Several functions of CK1 involve interaction with the cytoskeleton, presumably forlocalization of CK1 near its substrates [145, 147–150]. However the molecular mechanismsfor targeting CK1 within the cell are not well understood. It is likely that CK1 is targeted tothe outer doublet microtubules by a type of CK1-anchoring protein (CKAP) analogous to
Wirschell et al. Page 6
Arch Biochem Biophys. Author manuscript; available in PMC 2012 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

AKAPs. This model could define a basis for regulating CK1 activity and thereby ciliarymotility. Identification of such a putative CKAP could define a general class of proteins thatlocalize CK1 in the cell to direct and regulate CK1 function.
5. Axonemal PhosphatasesBiochemical and pharmacological analyses in diverse experimental systems have revealedthat protein phosphatases are also localized to the cilium [20, 25, 151–159]. In addition, therole of PKA and CK1 in regulation of microtubule sliding in axonemes fromChlamydomonas was first defined by taking advantage of mutant cells defective in normalcontrol of the kinases and resulting in constitutively active axonemal kinase activity [21, 60,87, 138, 160]. A consequence of the mutations and the constitutively active axonemalkinases was paralyzed flagellar axonemes and inhibition of dynein. Consistent with thisinterpretation, and illustrated in Figure 5, kinase inhibitors “rescued” microtubule sliding inisolated axonemes [21, 138]. Therefore, since protein kinase inhibitors rescued microtubulesliding in isolated axonemes, predictably the axonemes must contain protein phosphatasesrequired for de-phosphorylation of phospho-proteins and increased microtubule sliding [20,160, 161].
Consistent with this prediction, Habermacher and Sale (1996), determined that phosphataseinhibitors, including microcystin LR and okadaic acid, when applied before or with thekinase inhibitors, block kinase inhibitor-dependent rescue of microtubule sliding [161].Subsequent biochemical, molecular and proteomic analyses revealed the axoneme containsseveral phosphatases including the highly conserved Protein Phosphatase 1 (PP1) andProtein Phosphatase 2A (PP2A) [25, 162–164]. PP1 is a highly conserved serine/threoninephosphatase comprising one catalytic subunit and a range of regulatory mechanisms [165–167]. PP1 has been shown to be involved in various cellular processes such as mitosis,meiosis, transcription, apoptosis, cell cycle progression, and cell division [168, 169]. PP1was shown to exist in flagella using microcystin-sepharose affinity purification [20].Analyses of flagella from Chlamydomonas mutants revealed that PP1 is primarily, but notexclusively, anchored in the central pair apparatus, and is associated with the C1microtubule [20]. Thus, PP1 is thought to be part of the central pair-radial spoke signalingmechanism that controls axonemal dynein activity. A small fraction of PP1 is also presenton the outer doublet microtubules, possibly in close association with the dyneins.
PP2A is a highly conserved serine/threonine phosphatase important for normal development,cell cycle, and other many cell processes [169]. The PP2A heterodimeric core enzymecomprises one scaffold subunit (A-subunit) and one catalytic subunit (C-subunit), while thePP2A heterotrimeric holoenzyme comprises the A- and C- heterodimer and any one of afamily of regulatory subunits called B - subunits (also known as B55 or PR55), B′ (B56 orPR61), B″ (PR48/PR72/PR130), or B‴ (PR93/PR110). By using microcystin-sepharoseaffinity purification, a scaffold subunit and a catalytic subunit of PP2A were identified inChlamydomonas flagellar axonemes [20]. In Chlamydomonas, all three subunits (Scaffold,Regulatory, and Catalytic) of PP2A have been identified biochemically or in the flagellarproteome [20, 25]. Moreover, analysis of structural mutants revealed the axonemal PP2A islocalized to the outer doublet microtubules, presumably in position to regulate the dyneinmotors.
Based on proteomic (see below) and biochemical analyses, additional protein phosphataseshave been identified in ciliary and flagellar fractions, including recent analysis of primarycilia [151–153, 155, 164, 170]. In many cases, the phosphatases are localized to themembrane – matrix fractions, where they likely play important roles in control of ionchannels, ATPases or receptors [156, 159]. Consistent with these ideas, in Paramecium,
Wirschell et al. Page 7
Arch Biochem Biophys. Author manuscript; available in PMC 2012 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

PP1is involved in a sustained backward swimming response to depolarizing stimuli,suggesting PP1 functions on the ciliary/flagellar membrane [157]. However, to date, otherthan PP1 and PP2A, we do not have an idea for the functional roles of other axonemalphosphatases. The mechanism for anchoring of such a diverse pool of phosphatases in theaxoneme is largely unknown; presumably, they are anchored by specialized proteins similarto the axonemal kinases or by the same kinase-anchoring proteins [171–173]. In summary,PP1 is primarily localized to the central pair, possibly acting upstream in a pathway thatregulates the ciliary dynein motors and PP2A and a small fraction of PP1 are localized to theouter doublet microtubules, presumably near the dynein substrates, as downstreamphosphatases (Fig. 3).
6. Proteomic studies—and future opportunitiesSeveral studies have recently focused on identifying all ciliary proteins. These studiesinclude proteomic analyses of isolated motile and sensory cilia and ciliary fractions(axonemes, membranes and matrix proteins) as well as comparative genomic approaches(http://v3.ciliaproteome.org/cgi-bin/index.php) [25, 44, 45, 162–164, 174–177]. Theseapproaches contributed to a revolution in our understanding of cilia, revealing conservedgenes that, when defective, result in diseases and syndromes in the human. For example, thegenomic analysis of Li et al., (2004) contributed to discovery of a link between cilia andBardet-Biedel Syndrome [178] and helped lead investigators to discovery of the “BBSome”and mechanisms for trafficking ciliary membrane proteins [179–181].
The comparative genomic approaches omitted the conserved kinases and phosphatases thatfunction in cilia because they also have non-ciliary roles within the cell. The proteomicstudies, using tandem mass spectrometry coupled with interrogation of available genomedatabases, have identified several classes of signaling proteins including kinases,phosphatases, calcium-binding proteins, AKAPs and others [25]. A comprehensiveproteomic analysis of the Chlamydomonas cilium and ciliary fractions identified 20 kinases,11 phosphatases, 9 GTP-binding proteins, 27 EF hand calcium-binding proteins, 5 ionchannels, 1 IP3 receptor and 2 14-3-3 proteins indicating that the Chlamydomonas cilium isrich in signaling proteins and that motile cilia and flagella also function as sensory/signalingorganelles. The challenge is to determine the functional role of each ciliary component. Inaddition, genes predicted to encode candidate cAMP and cGMP dependent protein kinaseswere identified. These studies have provided the foundation for characterization of specificsignaling proteins and their role in regulation of ciliary motility.
Clearly, reversible phosphorylation is a critical feature of control of ciliary movement. Bothbiochemical and proteomic analyses demonstrate that the axoneme houses a large repertoireof phosphoproteins [26, 68]. Using 32P labeling of axonemal proteins and 2D SDS-PAGEanalysis, more than 80 axonemal phosphoproteins were identified in vivo and at steady state,five of which are known radial spoke stalk components [68], providing a preview that theradial spoke is an exceptionally complex axonemal signaling structure [70]. Given the largenumber of axonemal kinases, phosphatases and phosphoproteins, and the important role ofphosphorylation in ciliary function, it is critical to identify the exact substrates in thephosphoregulatory pathways. To begin to address this question, Boesger and colleagues tooka proteomics approach to identify ciliary phosphoproteins [26]. Using isolated cilia fromChlamydomonas and immobilized metal affinity chromatography, this study identified 141phosphopeptides with 126 phosphorylation sites belonging to 32 ciliary proteins includingstructural and motor proteins, kinases, EF hand proteins, FAPs (Flagellar Associated Protein—proteins identified in the Chlamydomonas flagellar proteome (25)), and manyuncharacterized ciliary proteins. Importantly, phosphorylation sites were determined inIC138 and radial spoke protein 17 (RSP17). This important study has provided a new and
Wirschell et al. Page 8
Arch Biochem Biophys. Author manuscript; available in PMC 2012 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

important database of candidate substrates with specific phosphorylation sites which can betested for their role in motility. One opportunity is to couple this sensitive method foridentifying the phospho-proteins in the axoneme with a useful selection of Chlamydomonasmutant axonemes defective in control of kinase or phosphatase activities.
Important questions for the future include (1) how does phosphorylation of dynein subunitsalter dynein activity? (2) How do the kinases and phosphatases, and yet to be determinedsubstrates located in the axoneme, modulate the dynein motors? (3) How do signals,initiated in the central pair and transmitted through the radial spokes, affect dynein activity?(4) What is the link between calcium signaling and axonemal protein phosphorylation?These questions, and others, will be best addressed using a combination of model organismsand useful, informative functional assays for ciliary motility.
References1. Satir P, Christensen ST. Annu Rev Physiol. 2007; 69:377–400. [PubMed: 17009929]2. Satir P, Christensen ST. Histochem Cell Biol. 2008; 129:687–693. [PubMed: 18365235]3. Badano JL, Mitsuma N, Beales PL, Katsanis N. Annu Rev Genomics Hum Genet. 2006; 7:125–148.
[PubMed: 16722803]4. Zariwala MA, Knowles MR, Omran H. Annu Rev Physiol. 2007; 69:423–450. [PubMed: 17059358]5. Sutherland MJ, Ware SM. Am J Med Genet C Semin Med Genet. 2009; 151C:307–317. [PubMed:
19876930]6. Hashimoto M, Hamada H. Curr Opin Genet Dev. 2010; 20:433–437. [PubMed: 20439159]7. Eggenschwiler JT, Anderson KV. Annu Rev Cell Dev Biol. 2007; 23:345–373. [PubMed:
17506691]8. Wallingford JB. Curr Opin Cell Biol. 2010; 22:597–604. [PubMed: 20817501]9. Basu B, Brueckner M. Curr Top Dev Biol. 2008; 85:151–174. [PubMed: 19147005]10. Gerdes JM, Davis EE, Katsanis N. Cell. 2009; 137:32–45. [PubMed: 19345185]11. Berbari NF, O'Connor AK, Haycraft CJ, Yoder BK. Curr Biol. 2009; 19:R526–R535. [PubMed:
19602418]12. Goetz SC, Anderson KV. Nat Rev Genet. 2010; 11:331–344. [PubMed: 20395968]13. Green JA, Mykytyn K. Cell Mol Life Sci. 2010; 67:3287–3297. [PubMed: 20544253]14. Porter ME, Sale WS. J Cell Biol. 2000; 151:F37–F42. [PubMed: 11086017]15. Gokhale, A.; Wirschell, M.; Sale, WS.; Mitchell, DM., editors. Regulation of ciliary and flagellar
dyneins. Singapore: Pan Stanford Publishing; 2010.16. Elam, CA.; Sale, WS.; Wirschell, M. Methods in Cell Biology. Stephen, MK.; Gregory, JP.,
editors. Academic Press; 2009. p. 133-151.17. Witman, GB. The Chlamydomons Sourcebook: Cell Motility and Behavior. Kidlington, Oxford:
Academic Press; 2009.18. Salathe M. Annu Rev Physiol. 2007; 69:401–422. [PubMed: 16945069]19. Wirschell M, Hendrickson T, Sale WS. Cell Motil Cytoskeleton. 2007; 64:569–579. [PubMed:
17549744]20. Yang P, Fox L, Colbran RJ, Sale WS. J Cell Sci. 2000; 113(Pt 1):91–102. [PubMed: 10591628]21. Yang P, Sale WS. J Biol Chem. 2000; 275:18905–18912. [PubMed: 10858448]22. Gaillard AR, Diener DR, Rosenbaum JL, Sale WS. J Cell Biol. 2001; 153:443–448. [PubMed:
11309423]23. Gaillard AR, Fox LA, Rhea JM, Craige B, Sale WS. Mol Biol Cell. 2006; 17:2626–2635.
[PubMed: 16571668]24. Gokhale A, Wirschell M, Sale WS. J Cell Biol. 2009; 186:817–824. [PubMed: 19752022]25. Pazour GJ, Agrin N, Leszyk J, Witman GB. J Cell Biol. 2005; 170:103–113. [PubMed: 15998802]26. Boesger J, Wagner V, Weisheit W, Mittag M. Eukaryot Cell. 2009; 8:922–932. [PubMed:
19429781]
Wirschell et al. Page 9
Arch Biochem Biophys. Author manuscript; available in PMC 2012 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

27. Christensen ST, Pedersen SF, Satir P, Veland IR, Schneider L. Curr Top Dev Biol. 2008; 85:261–301. [PubMed: 19147009]
28. Marshall WF, Kintner C. Curr Opin Cell Biol. 2008; 20:48–52. [PubMed: 18194854]29. Satir P, Pedersen LB, Christensen ST. J Cell Sci. 2010; 123:499–503. [PubMed: 20144997]30. Jones C, Chen P. Curr Top Dev Biol. 2008; 85:197–224. [PubMed: 19147007]31. Wong SY, Reiter JF. Curr Top Dev Biol. 2008; 85:225–260. [PubMed: 19147008]32. Tobin JL, Beales PL. Genet Med. 2009; 11:386–402. [PubMed: 19421068]33. Wirschell, M.; Nicastro, D.; Porter, ME.; Sale, WS. The Chlamydomonas Sourcebook. Witman,
GB., editor. Kidlington, Oxford: Academic Press; 2009. p. 253-282.34. Snell WJ, Pan J, Wang Q. Cell. 2004; 117:693–697. [PubMed: 15186771]35. Christensen ST, Pedersen LB, Schneider L, Satir P. Traffic. 2007; 8:97–109. [PubMed: 17241444]36. Marshall WF, Nonaka S. Curr Biol. 2006; 16:R604–R614. [PubMed: 16890522]37. Marshall WF. J Cell Biol. 2008; 180:17–21. [PubMed: 18180369]38. Sharma N, Berbari NF, Yoder BK. Curr Top Dev Biol. 2008; 85:371–427. [PubMed: 19147012]39. Dutcher SK. Traffic. 2003; 4:443–451. [PubMed: 12795689]40. Hiraki M, Nakazawa Y, Kamiya R, Hirono M. Curr Biol. 2007; 17:1778–1783. [PubMed:
17900905]41. Marshall WF. Curr Top Dev Biol. 2008; 85:1–22. [PubMed: 19147000]42. Azimzadeh J, Marshall WF. Curr Biol. 2010; 20:R816–R825. [PubMed: 20869612]43. Heuser T, Raytchev M, Krell J, Porter ME, Nicastro D. J Cell Biol. 2009; 187:921–933. [PubMed:
20008568]44. Avidor-Reiss T, Maer AM, Koundakjian E, Polyanovsky A, Keil T, Subramaniam S, Zuker CS.
Cell. 2004; 117:527–539. [PubMed: 15137945]45. Li JB, Gerdes JM, Haycraft CJ, Fan Y, Teslovich TM, May-Simera H, Li H, Blacque OE, Li L,
Leitch CC, Lewis RA, Green JS, Parfrey PS, Leroux MR, Davidson WS, Beales PL, Guay-Woodford LM, Yoder BK, Stormo GD, Katsanis N, Dutcher SK. Cell. 2004; 117:541–552.[PubMed: 15137946]
46. Li JB, Lin S, Jia H, Wu H, Roe BA, Kulp D, Stormo GD, Dutcher SK. J Eukaryot Microbiol. 2003;50:145–155. [PubMed: 12836870]
47. Nicastro D, Schwartz C, Pierson J, Gaudette R, Porter ME. McIntosh, Science. 2006; 313:944–948.
48. Nicastro D. Methods Cell Biol. 2009; 91:1–39. [PubMed: 20409778]49. Nicastro D, McIntosh JR, Baumeister W. Proc Natl Acad Sci U S A. 2005; 102:15889–15894.
[PubMed: 16246999]50. Bui KH, Sakakibara H, Movassagh T, Oiwa K, Ishikawa T. J Cell Biol. 2008; 183:923–932.
[PubMed: 19029338]51. Bui KH, Sakakibara H, Movassagh T, Oiwa K, Ishikawa T. J Cell Biol. 2009; 186:437–446.
[PubMed: 19667131]52. Oda T, Hirokawa N, Kikkawa M. J Cell Biol. 2007; 177:243–252. [PubMed: 17438074]53. Adams GM, Huang B, Piperno G, Luck DJ. J Cell Biol. 1981; 91:69–76. [PubMed: 7028763]54. Dutcher SK, Huang B, Luck DJ. J Cell Biol. 1984; 98:229–236. [PubMed: 6707088]55. Rupp G, O'Toole E, Porter ME. Mol Biol Cell. 2001; 12:739–751. [PubMed: 11251084]56. Mitchell DR, Sale WS. J Cell Biol. 1999; 144:293–304. [PubMed: 9922455]57. Wargo MJ, Dymek EE, Smith EF. J Cell Sci. 2005; 118:4655–4665. [PubMed: 16188941]58. Lechtreck KF, Delmotte P, Robinson ML, Sanderson MJ, Witman GB. J Cell Biol. 2008; 180:633–
643. [PubMed: 18250199]59. Lechtreck KF, Witman GB. J Cell Biol. 2007; 176:473–482. [PubMed: 17296796]60. Smith EF, Yang P. Cell Motil Cytoskeleton. 2004; 57:8–17. [PubMed: 14648553]61. Mitchell DR, Smith B. Methods Cell Biol. 2009; 92:197–213. [PubMed: 20409807]62. DiPetrillo CG, Smith EF. J Cell Biol. 2010; 189:601–612. [PubMed: 20421426]63. Dymek EE, Smith EF. J Cell Biol. 2007; 179:515–526. [PubMed: 17967944]
Wirschell et al. Page 10
Arch Biochem Biophys. Author manuscript; available in PMC 2012 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

64. Luck D, Piperno G, Ramanis Z, Huang B. Proc Natl Acad Sci U S A. 1977; 74:3456–3460.[PubMed: 269405]
65. Witman GB, Plummer J, Sander G. J Cell Biol. 1978; 76:729–747. [PubMed: 632325]66. Sturgess JM, Chao J, Wong J, Aspin N, Turner JA. N Engl J Med. 1979; 300:53–56. [PubMed:
152870]67. Huang B, Piperno G, Ramanis Z, Luck DJ. J Cell Biol. 1981; 88:80–88. [PubMed: 7204490]68. Piperno G, Huang B, Ramanis Z, Luck DJ. J Cell Biol. 1981; 88:73–79. [PubMed: 6451632]69. Yang P, Diener DR, Rosenbaum JL, Sale WS. J Cell Biol. 2001; 153:1315–1326. [PubMed:
11402073]70. Yang P, Diener DR, Yang C, Kohno T, Pazour GJ, Dienes JM, Agrin NS, King SM, Sale WS,
Kamiya R, Rosenbaum JL, Witman GB. J Cell Sci. 2006; 119:1165–1174. [PubMed: 16507594]71. Diener DR, Ang LH, Rosenbaum JL. J Cell Biol. 1993; 123:183–190. [PubMed: 8408197]72. Wirschell M, Zhao F, Yang C, Yang P, Diener D, Gaillard A, Rosenbaum JL, Sale WS. Cell Motil
Cytoskeleton. 2008; 65:238–248. [PubMed: 18157907]73. Patel-King RS, Gorbatyuk O, Takebe S, King SM. Mol Biol Cell. 2004; 15:3891–3902. [PubMed:
15194815]74. Brokaw CJ, Luck DJ, Huang B. J Cell Biol. 1982; 92:722–732. [PubMed: 7085755]75. Huang B, Ramanis Z, Luck DJ. Cell. 1982; 28:115–124. [PubMed: 6461414]76. Gardner LC, O'Toole E, Perrone CA, Giddings T, Porter ME. J Cell Biol. 1994; 127:1311–1325.
[PubMed: 7962092]77. Piperno G, Mead K, LeDizet M, Moscatelli A. J Cell Biol. 1994; 125:1109–1117. [PubMed:
8195292]78. Piperno G, Mead K, Shestak W. J Cell Biol. 1992; 118:1455–1463. [PubMed: 1387875]79. Porter ME, Power J, Dutcher SK. J Cell Biol. 1992; 118:1163–1176. [PubMed: 1387404]80. Rupp G, Porter ME. J Cell Biol. 2003; 162:47–57. [PubMed: 12847082]81. Kabututu ZP, Thayer M, Melehani JH, Hill KL. J Cell Sci. 2010; 123:3587–3595. [PubMed:
20876659]82. Bekker JM, Colantonio JR, Stephens AD, Clarke WT, King SJ, Hill KL, Crosbie RH. Cell Motil
Cytoskeleton. 2007; 64:461–473. [PubMed: 17366626]83. Colantonio JR, Bekker JM, Kim SJ, Morrissey KM, Crosbie RH, Hill KL. Traffic. 2006; 7:538–
548. [PubMed: 16643277]84. Ralston KS, Lerner AG, Diener DR, Hill KL. Eukaryot Cell. 2006; 5:696–711. [PubMed:
16607017]85. King, SM.; Kamiya, R. The Chlamydomonas Sourcebook. Witman, GB., editor. Kidlington,
Oxford: Academic Press; 2009. p. 131-208.86. Brokaw CJ, Kamiya R. Cell Motil Cytoskeleton. 1987; 8:68–75. [PubMed: 2958145]87. Habermacher G, Sale WS. J Cell Biol. 1997; 136:167–176. [PubMed: 9008711]88. King SJ, Dutcher SK. J Cell Biol. 1997; 136:177–191. [PubMed: 9008712]89. Bower R, VanderWaal K, O'Toole E, Fox L, Perrone C, Mueller J, Wirschell M, Kamiya R, Sale
WS, Porter ME. Mol Biol Cell. 2009; 20:3055–3063. [PubMed: 19420135]90. Hendrickson TW, Perrone CA, Griffin P, Wuichet K, Mueller J, Yang P, Porter ME, Sale WS. Mol
Biol Cell. 2004; 15:5431–5442. [PubMed: 15469982]91. Holwill ME, Cohen HJ, Satir P. J Exp Biol. 1979; 78:265–280. [PubMed: 438720]92. Satir P. J Cell Biol. 1968; 39:77–94. [PubMed: 5678451]93. Brokaw CJ. Science. 1972; 178:455–462. [PubMed: 4673044]94. Summers KE, Gibbons IR. Proc Natl Acad Sci U S A. 1971; 68:3092–3096. [PubMed: 5289252]95. Shingyoji C, Murakami A, Takahashi K. Nature. 1977; 265:269–270. [PubMed: 834273]96. Brokaw CJ. J Cell Biol. 1991; 114:1201–1215. [PubMed: 1894694]97. Sale WS, Satir P. Proc Natl Acad Sci U S A. 1977; 74:2045–2049. [PubMed: 266725]98. Fox LA, Sale WS. J Cell Biol. 1987; 105:1781–1787. [PubMed: 2959667]99. Woolley DM. Biological Reviews. 2010; 85:453–470. [PubMed: 20002389]
Wirschell et al. Page 11
Arch Biochem Biophys. Author manuscript; available in PMC 2012 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

100. Brokaw CJ. Cell Motil Cytoskeleton. 2009; 66:425–436. [PubMed: 18828155]101. Smith EF. J Cell Biol. 2007; 176:403–404. [PubMed: 17296793]102. Morita Y, Shingyoji C. Curr Biol. 2004; 14:2113–2118. [PubMed: 15589153]103. Hayashi S, Shingyoji C. Cell Motil Cytoskeleton. 2009; 66:292–301. [PubMed: 19343792]104. Hayashibe K, Shingyoji C, Kamiya R. Cell Motil Cytoskeleton. 1997; 37:232–239. [PubMed:
9227853]105. Inaba K. Zoolog Sci. 2003; 20:1043–1056. [PubMed: 14578564]106. Brokaw CJ. J Cell Biochem. 1987; 35:175–184. [PubMed: 2826504]107. Harrison RA. Reprod Domest Anim. 2003; 38:102–110. [PubMed: 12654020]108. Tash JS, Means AR. Biol Reprod. 1983; 28:75–104. [PubMed: 6299416]109. Tash JS, Means AR. Prog Clin Biol Res. 1988; 267:335–355. [PubMed: 2853368]110. Yoshida M, Kawano N, Yoshida K. Cell Mol Life Sci. 2008; 65:3446–3457. [PubMed:
18622578]111. Fraser LR, Monks NJ. J Reprod Fertil Suppl. 1990; 42:9–21. [PubMed: 1963902]112. Garbers DL, Kopf GS. Adv Cyclic Nucleotide Res. 1980; 13:251–306. [PubMed: 6106348]113. DiPetrillo C, Smith E. Methods Cell Biol. 2009; 92:163–180. [PubMed: 20409805]114. Bonini NM, Nelson DL. J Cell Biol. 1988; 106:1615–1623. [PubMed: 2836435]115. Gertsberg I, Hellman V, Fainshtein M, Weil S, Silberberg SD, Danilenko M, Priel Z. J Gen
Physiol. 2004; 124:527–540. [PubMed: 15477378]116. Lindemann CB, Lesich KA. Methods Mol Biol. 2009; 586:337–353. [PubMed: 19768440]117. Lindemann CB, Schmitz KA. Methods Mol Biol. 2001; 161:241–252. [PubMed: 11190510]118. Bonini NM, Gustin MC, Nelson DL. Cell Motil Cytoskeleton. 1986; 6:256–272. [PubMed:
2427226]119. Hamasaki T, Murtaugh TJ, Satir BH, Satir P. Cell Motil Cytoskeleton. 1989; 12:1–11. [PubMed:
2539909]120. Hochstrasser M, Carlson GL, Walczak CE, Nelson DL. J Eukaryot Microbiol. 1996; 43:356–362.
[PubMed: 8768441]121. Walczak CE, Anderson RA, Nelson DL. Biochem J. 1993; 296(Pt 3):729–735. [PubMed:
8280070]122. Hamasaki T, Barkalow K, Richmond J, Satir P. Proc Natl Acad Sci U S A. 1991; 88:7918–7922.
[PubMed: 1654550]123. Bonini NM, Evans TC, Miglietta LA, Nelson DL. Adv Second Messenger Phosphoprotein Res.
1991; 23:227–272. [PubMed: 1997042]124. Christensen ST, Guerra C, Wada Y, Valentin T, Angeletti RH, Satir P, Hamasaki T. J Biol Chem.
2001; 276:20048–20054. [PubMed: 11274140]125. Geary CA, Davis CW, Paradiso AM, Boucher RC. Am J Physiol. 1995; 268:L1021–L1028.
[PubMed: 7611424]126. Zhang L, Sanderson MJ. J Physiol. 2003; 551:765–776. [PubMed: 12819300]127. Wyatt TA, Forget MA, Adams JM, Sisson JH. Am J Physiol Lung Cell Mol Physiol. 2005;
288:L546–L551. [PubMed: 15542545]128. Wyatt TA, Forget MA, Sisson JH. Am J Pathol. 2003; 163:1157–1166. [PubMed: 12937157]129. Miglietta LA, Nelson DL. J Biol Chem. 1988; 263:16096–16105. [PubMed: 3182784]130. Ann KS, Nelson DL. Cell Motil Cytoskeleton. 1995; 30:252–260. [PubMed: 7796456]131. Howard DR, Trantow CM, Thaler CD. Cell Motil Cytoskeleton. 2004; 59:120–130. [PubMed:
15362116]132. Salathe M, Pratt MM, Wanner A. Am J Respir Cell Mol Biol. 1993; 9:306–314. [PubMed:
8398168]133. Schmid A, Sutto Z, Nlend MC, Horvath G, Schmid N, Buck J, Levin LR, Conner GE, Fregien N,
Salathe M. J Gen Physiol. 2007; 130:99–109. [PubMed: 17591988]134. Schmid A, Bai G, Schmid N, Zaccolo M, Ostrowski LE, Conner GE, Fregien N, Salathe M. J Cell
Sci. 2006; 119:4176–4186. [PubMed: 16984973]
Wirschell et al. Page 12
Arch Biochem Biophys. Author manuscript; available in PMC 2012 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

135. Sisson JH, May K, Wyatt TA. Alcohol Clin Exp Res. 1999; 23:1528–1533. [PubMed: 10512320]136. Kultgen PL, Byrd SK, Ostrowski LE, Milgram SL. Mol Biol Cell. 2002; 13:4156–4166.
[PubMed: 12475942]137. King SM, Witman GB. J Biol Chem. 1994; 269:5452–5457. [PubMed: 7508939]138. Howard DR, Habermacher G, Glass DB, Smith EF, Sale WS. J Cell Biol. 1994; 127:1683–1692.
[PubMed: 7798320]139. Toba S, Fox LA, Sakakibara H, Porter ME, Oiwa K, Sale WS. Mol Biol Cell. 2011; 22:342–353.
[PubMed: 21148301]140. Okita N, Isogai N, Hirono M, Kamiya R, Yoshimura K. J Cell Sci. 2005; 118:529–537. [PubMed:
15657081]141. Scott JD. Biochem Soc Trans. 2003; 31:87–89. [PubMed: 12546660]142. Beene DL, Scott JD. Curr Opin Cell Biol. 2007; 19:192–198. [PubMed: 17317140]143. Carr DW, Newell AE. Soc Reprod Fertil Suppl. 2007; 63:135–141. [PubMed: 17566268]144. Bryja V, Schulte G, Rawal N, Grahn A, Arenas E. J Cell Sci. 2007; 120:586–595. [PubMed:
17244647]145. Gross SD, Anderson RA. Cell Signal. 1998; 10:699–711. [PubMed: 9884021]146. Knippschild U, Gocht A, Wolff S, Huber N, Lohler J, Stoter M. Cell Signal. 2005; 17:675–689.
[PubMed: 15722192]147. Sillibourne JE, Milne DM, Takahashi M, Ono Y, Meek DW. J Mol Biol. 2002; 322:785–797.
[PubMed: 12270714]148. Behrend L, Stoter M, Kurth M, Rutter G, Heukeshoven J, Deppert W, Knippschild U. Eur J Cell
Biol. 2000; 79:240–251. [PubMed: 10826492]149. Ben-Nissan G, Cui W, Kim DJ, Yang Y, Yoo BC, Lee JY. Plant Physiol. 2008; 148:1897–1907.
[PubMed: 18945931]150. Li G, Yin H, Kuret J. J Biol Chem. 2004; 279:15938–15945. [PubMed: 14761950]151. Kroner C, Boekhoff I, Breer H. Biochim Biophys Acta. 1996; 1312:169–175. [PubMed:
8672541]152. Clement A, Solnica-Krezel L, Gould KL. Development. 2011; 138:291–302. [PubMed:
21177342]153. Jacoby M, Cox JJ, Gayral S, Hampshire DJ, Ayub M, Blockmans M, Pernot E, Kisseleva MV,
Compere P, Schiffmann SN, Gergely F, Riley JH, Perez-Morga D, Woods CG, Schurmans S. NatGenet. 2009; 41:1027–1031. [PubMed: 19668215]
154. Wang W, Brautigan DL. BMC Cell Biol. 2008; 9:62. [PubMed: 19036150]155. Boucher CA, Ward HH, Case RL, Thurston KS, Li X, Needham A, Romero E, Hyink D, Qamar
S, Roitbak T, Powell S, Ward C, Wilson PD, Wandinger-Ness A, Sandford RN. BiochimBiophys Acta. 2010
156. Klumpp S, Cohen P, Schultz JE. J Chromatogr. 1990; 521:179–186. [PubMed: 1962786]157. Klumpp S, Cohen P, Schultz JE. EMBO J. 1990; 9:685–689. [PubMed: 2155778]158. Klumpp S, Schultz JE. Biochim Biophys Acta. 1990; 1037:233–239. [PubMed: 2155027]159. Friderich G, Klumpp S, Russell CB, Hinrichsen RD, Kellner R, Schultz JE. Eur J Biochem. 1992;
209:43–49. [PubMed: 1327778]160. Habermacher G, Sale WS. Cell Motil Cytoskeleton. 1995; 32:106–109. [PubMed: 8681389]161. Habermacher G, Sale WS. J Cell Sci. 1996; 109(Pt 7):1899–1907. [PubMed: 8832412]162. Ostrowski LE, Blackburn K, Radde KM, Moyer MB, Schlatzer DM, Moseley A, Boucher RC.
Mol Cell Proteomics. 2002; 1:451–465. [PubMed: 12169685]163. Inglis PN, Boroevich KA, Leroux MR. Trends Genet. 2006; 22:491–500. [PubMed: 16860433]164. Liu Q, Tan G, Levenkova N, Li T, Pugh EN Jr, Rux JJ, Speicher DW, Pierce EA. Mol Cell
Proteomics. 2007; 6:1299–1317. [PubMed: 17494944]165. Roy J, Cyert MS. Sci Signal. 2009; 2:re9. [PubMed: 19996458]166. Bollen M, Peti W, Ragusa MJ, Beullens M. Trends Biochem Sci. 2010; 35:450–458. [PubMed:
20399103]167. Virshup DM, Shenolikar S. Mol Cell. 2009; 33:537–545. [PubMed: 19285938]
Wirschell et al. Page 13
Arch Biochem Biophys. Author manuscript; available in PMC 2012 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

168. Wang B, Xie X, Wei Q. Wei Sheng Wu Xue Bao. 2008; 48:269–273. [PubMed: 18438014]169. Shi Y. Cell. 2009; 139:468–484. [PubMed: 19879837]170. Bielas SL, Silhavy JL, Brancati F, Kisseleva MV, Al-Gazali L, Sztriha L, Bayoumi RA, Zaki MS,
Abdel-Aleem A, Rosti RO, Kayserili H, Swistun D, Scott LC, Bertini E, Boltshauser E, Fazzi E,Travaglini L, Field SJ, Gayral S, Jacoby M, Schurmans S, Dallapiccola B, Majerus PW, ValenteEM, Gleeson JG. Nat Genet. 2009; 41:1032–1036. [PubMed: 19668216]
171. Scott JD. Soc Gen Physiol Ser. 1997; 52:227–239. [PubMed: 9210233]172. Coghlan VM, Perrino BA, Howard M, Langeberg LK, Hicks JB, Gallatin WM, Scott JD. Science.
1995; 267:108–111. [PubMed: 7528941]173. Dodge K, Scott JD. FEBS Lett. 2000; 476:58–61. [PubMed: 10878251]174. Gherman A, Davis EE, Katsanis N. Nat Genet. 2006; 38:961–962. [PubMed: 16940995]175. Hart SR, Lau KW, Hao Z, Broadhead R, Portman N, Huhmer A, Gull K, McKean PG, Hubbard
SJ, Gaskell SJ. J Am Soc Mass Spectrom. 2009; 20:167–175. [PubMed: 18930411]176. Broadhead R, Dawe HR, Farr H, Griffiths S, Hart SR, Portman N, Shaw MK, Ginger ML,
Gaskell SJ, McKean PG, Gull K. Nature. 2006; 440:224–227. [PubMed: 16525475]177. Stolc V, Samanta MP, Tongprasit W, Marshall WF. Proc Natl Acad Sci U S A. 2005; 102:3703–
3707. [PubMed: 15738400]178. Tobin JL, Beales PL. Pediatr Nephrol. 2007; 22:926–936. [PubMed: 17357787]179. Nachury MV, Loktev AV, Zhang Q, Westlake CJ, Peranen J, Merdes A, Slusarski DC, Scheller
RH, Bazan JF, Sheffield VC, Jackson PK. Cell. 2007; 129:1201–1213. [PubMed: 17574030]180. Nachury MV, Seeley ES, Jin H. Annu Rev Cell Dev Biol. 2010; 26:59–87. [PubMed: 19575670]181. Jin H, White SR, Shida T, Schulz S, Aguiar M, Gygi SP, Bazan JF, Nachury MV. Cell. 2010;
141:1208–1219. [PubMed: 20603001]
Wirschell et al. Page 14
Arch Biochem Biophys. Author manuscript; available in PMC 2012 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
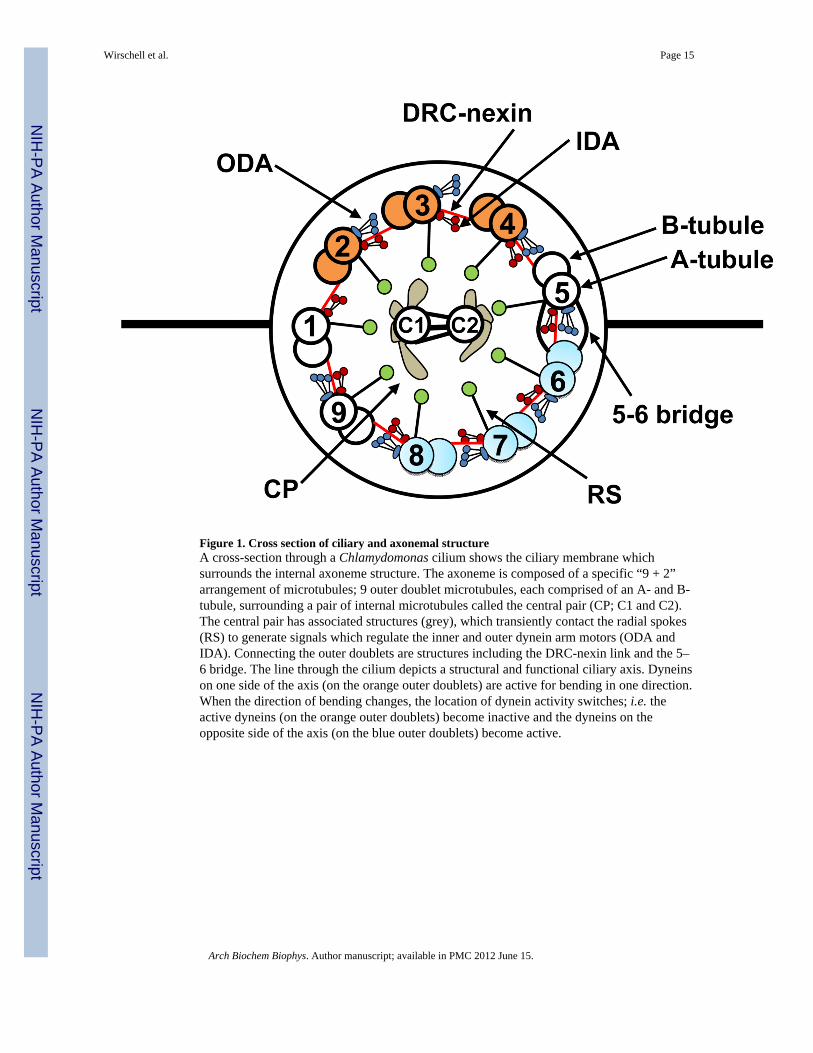
Figure 1. Cross section of ciliary and axonemal structureA cross-section through a Chlamydomonas cilium shows the ciliary membrane whichsurrounds the internal axoneme structure. The axoneme is composed of a specific “9 + 2”arrangement of microtubules; 9 outer doublet microtubules, each comprised of an A- and B-tubule, surrounding a pair of internal microtubules called the central pair (CP; C1 and C2).The central pair has associated structures (grey), which transiently contact the radial spokes(RS) to generate signals which regulate the inner and outer dynein arm motors (ODA andIDA). Connecting the outer doublets are structures including the DRC-nexin link and the 5–6 bridge. The line through the cilium depicts a structural and functional ciliary axis. Dyneinson one side of the axis (on the orange outer doublets) are active for bending in one direction.When the direction of bending changes, the location of dynein activity switches; i.e. theactive dyneins (on the orange outer doublets) become inactive and the dyneins on theopposite side of the axis (on the blue outer doublets) become active.
Wirschell et al. Page 15
Arch Biochem Biophys. Author manuscript; available in PMC 2012 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 2. Predicted location of axonemal signaling proteinsThe diagram depicts a cross-section view of the central pair, one radial spoke and one outerdoublet microtubule. The outer and inner dynein arms (ODA and IDA respectively) areanchored to the outer doublet microtubules. Regulatory kinases and phosphatases includePP1 (located primarily in the central pair with a small amount on the outer doublets,possibly near the ODA), PP2A (located on the outer doublets, possibly near I1 dynein), CK1(located on the outer doublets, possibly near I1 dynein), and PKA (predictably located at thebase of the radial spoke and in the central pair by the RSP3-AKAP and AKAP240respectively). Also shown are the predicted locations of calcium-sensitive regulatorycomplexes that contain calmodulin.
Wirschell et al. Page 16
Arch Biochem Biophys. Author manuscript; available in PMC 2012 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 3. Schematic of a longitudinal view of the 96-nm repeatA longitudinal view of an outer doublet microtubule with the dynein motors (ODA andIDA), radial spokes (S1 and S2), and the DRC-nexin link. These structures are arranged in aprecise manner which forms a 96-nm structural element that repeats along the entire lengthof the axoneme. Within each 96-nm repeat, there are four ODA, two RS, one DRC-nexinlink (DRC), and one copy each of several different types of IDA motors. The IDA shown inred is called I1 dynein (also known as dynein-f). This particular motor is regulated by aphosphorylation by a mechano-chemical mechanism that involves the central pair and radialspoke structures.
Wirschell et al. Page 17
Arch Biochem Biophys. Author manuscript; available in PMC 2012 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
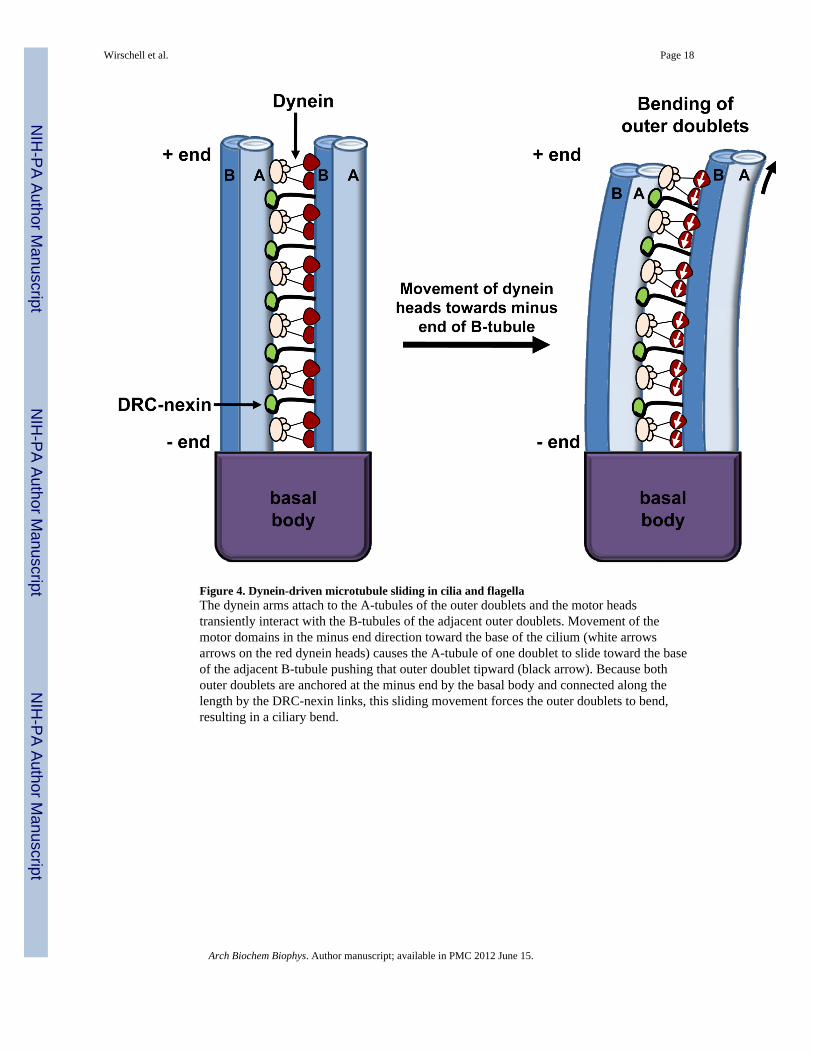
Figure 4. Dynein-driven microtubule sliding in cilia and flagellaThe dynein arms attach to the A-tubules of the outer doublets and the motor headstransiently interact with the B-tubules of the adjacent outer doublets. Movement of themotor domains in the minus end direction toward the base of the cilium (white arrowsarrows on the red dynein heads) causes the A-tubule of one doublet to slide toward the baseof the adjacent B-tubule pushing that outer doublet tipward (black arrow). Because bothouter doublets are anchored at the minus end by the basal body and connected along thelength by the DRC-nexin links, this sliding movement forces the outer doublets to bend,resulting in a ciliary bend.
Wirschell et al. Page 18
Arch Biochem Biophys. Author manuscript; available in PMC 2012 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 5. Model for regulation of I1 dynein by phosphorylationI1 dynein activity is regulated by phosphorylation of IC138. When phosphorylated, I1dynein is inactive; upon dephosphorylation, I1 dynein is active. The kinases andphosphatases involved were identified based on pharmacological data using inhibitors andinclude CK1, PKA, PP1 and PP2A. The central pair (CP) and radial spokes (RS) function, inpart, to regulate the kinases in vivo; when mutated, the kinases are constitutively active,resulting in uniform phosphorylation of I1 dynein and global inactivation of dynein activity.This inactivation can be relieved, in vitro, by the addition of kinase inhibitors. Consequently,in vitro inhibition of phosphatase activity prevents dynein activation.
Wirschell et al. Page 19
Arch Biochem Biophys. Author manuscript; available in PMC 2012 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Related Documents