Radiation and divergence in the Rhagoletis Pomonella species complex: inferences from DNA sequence data X. XIE, * A. P. MICHEL,* D. SCHWARZ, J. RULL, à S. VELEZ,* A. A. FORBES,* M. ALUJA à & J. L. FEDER* *Department of Biological Sciences, University of Notre Dame, Notre Dame, IN, USA Department of Entomology, Pennsylvania State University, University Park, PA, USA àInstituto de Ecologı´a, Asociacio ´n Civil, Xalapa, Veracruz, Mexico Introduction Understanding what a species is and the processes responsible for their formation are two central issues in evolutionary biology. The standard answer to the first question has been the biological species concept (BSC), which defines species on the basis of intrinsic barriers to gene flow (Mayr, 1963). Geographic isolation (allopatry) has been argued to be the common denominator with respect to answering the second question of speciation mode (Mayr, 1963). In recent years, a number of alternative, genetically based species concepts have been proposed to the BSC. These concepts include defining species based on the existence of genotypic clusters in sympatry (Mallet, 1995; Feder, 1998), on the presence of diagnostic autapomor- phies distinguishing populations (Cracraft, 1989) and on reciprocal monophyly between evolutionary lineages (de Queiroz, 1998). Although debate continues on the merits, practicality and relationship of these genetic concepts relative to the BSC (Coyne & Orr, 2004), the discussion has served to highlight that the genomes of diverging populations, particularly taxa undergoing divergence-with-gene-flow speciation, can be mosaic. Regions containing loci under selection contributing to gene flow barriers (islands of speciation; Turner et al., 2005) can display greater differentiation than regions not involved in reproductive isolation. A single value of reproductive isolation between taxa may therefore not apply uniformly across the genome (Mallet, 1995; Feder, 1998; Wu, 2001), as whole genomes need not be isolated (Mayr, 2001) or monophyletic (Hudson & Coyne, 2002) to have good species. Correspondence: Jeffrey L. Feder, Department of Biological Sciences, PO Box 369, Galvin Life Science Center, University of Notre Dame, Notre Dame, IN 46556-0369, USA. Tel.: +1 574 631 4159; fax: +1 574 631 7413; e-mail: [email protected] Present addresses: A. P. Michel, Department of Entomology, Ohio Agricultural Research and Development Center, Ohio State University, 210 Thorne Hall, 1680 Madison Ave., Wooster, OH 44691, USA. D. Schwarz, Department of Entomology, University of Illinois at Urbana- Champaign, Morrill Hall, 505 S. Goodwin Ave., Urbana, IL 61801, USA. S. Velez, Museum of Comparative Zoology, Harvard University, 26 Oxford St, Cambridge, MA 02138, USA. ª 2008 THE AUTHORS. J. EVOL. BIOL. 21 (2008) 900–913 900 JOURNAL COMPILATION ª 2008 EUROPEAN SOCIETY FOR EVOLUTIONARY BIOLOGY Keywords: biogeography; differential introgression; genetic divergence; host races; inversions; sibling species; speciation mode plurality; sympatric speciation. Abstract Here, we investigate the evolutionary history and pattern of genetic divergence in the Rhagoletis pomonella (Diptera: Tephritidae) sibling species complex, a model for sympatric speciation via host plant shifting, using 11 anonymous nuclear genes and mtDNA. We report that DNA sequence results largely coincide with those of previous allozyme studies. Rhagoletis cornivora was basal in the complex, distinguished by fixed substitutions at all loci. Gene trees did not provide reciprocally monophyletic relationships among US populations of R. pomonella, R. mendax, R. zephyria and the undescribed flowering dogwood fly. However, private alleles were found for these taxa for certain loci. We discuss the implications of the results with respect to identifiable genetic signposts (stages) of speciation, the mosaic nature of genomic differentiation distinguishing formative species and a concept of speciation mode plurality involving a biogeographic contribution to sympatric speciation in the R. pomonella complex. doi: 10.1111/j.1420-9101.2008.01507.x

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Radiation and divergence in the Rhagoletis Pomonella speciescomplex: inferences from DNA sequence data
X. XIE,* A. P. MICHEL,* D. SCHWARZ,� J. RULL,� S. VELEZ,* A. A. FORBES,* M. ALUJA�& J. L. FEDER*
*Department of Biological Sciences, University of Notre Dame, Notre Dame, IN, USA
�Department of Entomology, Pennsylvania State University, University Park, PA, USA
�Instituto de Ecologıa, Asociacion Civil, Xalapa, Veracruz, Mexico
Introduction
Understanding what a species is and the processes
responsible for their formation are two central issues in
evolutionary biology. The standard answer to the first
question has been the biological species concept (BSC),
which defines species on the basis of intrinsic barriers to
gene flow (Mayr, 1963). Geographic isolation (allopatry)
has been argued to be the common denominator with
respect to answering the second question of speciation
mode (Mayr, 1963).
In recent years, a number of alternative, genetically
based species concepts have been proposed to the BSC.
These concepts include defining species based on the
existence of genotypic clusters in sympatry (Mallet, 1995;
Feder, 1998), on the presence of diagnostic autapomor-
phies distinguishing populations (Cracraft, 1989) and on
reciprocal monophyly between evolutionary lineages (de
Queiroz, 1998). Although debate continues on the
merits, practicality and relationship of these genetic
concepts relative to the BSC (Coyne & Orr, 2004), the
discussion has served to highlight that the genomes of
diverging populations, particularly taxa undergoing
divergence-with-gene-flow speciation, can be mosaic.
Regions containing loci under selection contributing to
gene flow barriers (islands of speciation; Turner et al.,
2005) can display greater differentiation than regions not
involved in reproductive isolation. A single value of
reproductive isolation between taxa may therefore not
apply uniformly across the genome (Mallet, 1995; Feder,
1998; Wu, 2001), as whole genomes need not be isolated
(Mayr, 2001) or monophyletic (Hudson & Coyne, 2002)
to have good species.
Correspondence: Jeffrey L. Feder, Department of Biological Sciences, PO
Box 369, Galvin Life Science Center, University of Notre Dame, Notre
Dame, IN 46556-0369, USA.
Tel.: +1 574 631 4159; fax: +1 574 631 7413;
e-mail: [email protected]
Present addresses: A. P. Michel, Department of Entomology, Ohio
Agricultural Research and Development Center, Ohio State University,
210 Thorne Hall, 1680 Madison Ave., Wooster, OH 44691, USA.
D. Schwarz, Department of Entomology, University of Illinois at Urbana-
Champaign, Morrill Hall, 505 S. Goodwin Ave., Urbana, IL 61801, USA.
S. Velez, Museum of Comparative Zoology, Harvard University, 26
Oxford St, Cambridge, MA 02138, USA.
ª 2 0 0 8 T H E A U T H O R S . J . E V O L . B I O L . 2 1 ( 2 0 0 8 ) 9 0 0 – 9 1 3
900 J O U R N A L C O M P I L A T I O N ª 2 0 0 8 E U R O P E A N S O C I E T Y F O R E V O L U T I O N A R Y B I O L O G Y
Keywords:
biogeography;
differential introgression;
genetic divergence;
host races;
inversions;
sibling species;
speciation mode plurality;
sympatric speciation.
Abstract
Here, we investigate the evolutionary history and pattern of genetic
divergence in the Rhagoletis pomonella (Diptera: Tephritidae) sibling species
complex, a model for sympatric speciation via host plant shifting, using 11
anonymous nuclear genes and mtDNA. We report that DNA sequence results
largely coincide with those of previous allozyme studies. Rhagoletis cornivora
was basal in the complex, distinguished by fixed substitutions at all loci. Gene
trees did not provide reciprocally monophyletic relationships among US
populations of R. pomonella, R. mendax, R. zephyria and the undescribed
flowering dogwood fly. However, private alleles were found for these taxa
for certain loci. We discuss the implications of the results with respect to
identifiable genetic signposts (stages) of speciation, the mosaic nature of
genomic differentiation distinguishing formative species and a concept of
speciation mode plurality involving a biogeographic contribution to sympatric
speciation in the R. pomonella complex.
doi: 10.1111/j.1420-9101.2008.01507.x

The issue of speciation process has also been conten-
tious. Recently, however, a growing consensus has
emerged that allopatry is not the exclusive mode of
speciation, even for animals; divergence-with-gene-flow
speciation can and does occur, although under more
restrictive circumstances than speciation in allopatry
(Coyne & Orr, 2004). An important accompanying
development in this discussion has been an increasing
awareness of the possibility of speciation mode ‘plural-
ity’; that categorizing speciation as strictly allopatric vs.
nonallopatric may not adequately describe the geo-
graphic context of divergence for many taxa. If, for
example, some genetic changes leading to reproductive
isolation occur in allopatry and others in sympatry, then
it would seem appropriate for the geographic mode of
speciation to be mixed (Mallet, 2005). In this regard,
Coyne & Orr (2004) have proposed several new terms
describing situations of mixed geographic mode.
The Rhagoletis pomonella (Diptera: Tephritidae) complex
provides an opportunity to investigate questions con-
cerning speciation pattern and process. The complex
contains a number of host races (e.g. apple and haw-
thorn-infesting populations of R. pomonella) and sibling
species (e.g. R. mendax, R. zephyria, R. cornivora and the
undescribed flowering dogwood fly) at varying stages of
divergence. The close morphological similarity, distinct
host plant affiliations and broadly overlapping geographic
ranges of R. pomonella complex flies led Bush (1966,
1969) to propose that its members all speciated sympat-
rically via host plant shifting in North America. The
central premise of the sympatric hypothesis is that
differential adaptation to alternative host plants gener-
ates ecological barriers to gene flow that initiate specia-
tion. For R. pomonella, each fly in the complex infests a
unique, nonoverlapping set of host plants (see supple-
mentary Table S1) which tend to fruit at different times
of the field season (Smith, 1988; Feder et al., 1993;
Berlocher, 2000; Dambroski & Feder, 2007). Because
Rhagoletis is univoltine, differences in the depth of the
over-wintering diapause differentially adapts flies to
variation in host fruiting phenology, generating allo-
chronic mating isolation between R. pomonella taxa
(Smith, 1988; Feder et al., 1993, 1994; Dambroski &
Feder, 2007). Rhagoletis flies also mate almost exclusively
on or near the fruit of their respective host plants
(Prokopy et al. 1971, 1972). Traits related to host plant
discrimination, including differences in fruit volatile
preference, therefore also directly affect mate choice
and represent another important ecological barrier to
gene flow among R. pomonella flies (Feder et al., 1994;
Linn et al., 2003, 2004).
Here, we investigate patterns of DNA sequence diver-
gence for 11 anonymous nuclear genes and mtDNA in
the R. pomonella complex. The objectives of the study are
threefold. Our first goal is to test whether any ‘genetic
signposts’ of divergence exist for R. pomonella flies. By
genetic signpost, we are referring to identifiable patterns
of differentiation, such as degree of genotypic clustering
or monophyly, in gene trees indicative of stages (the
extent of ecological and intrinsic isolation) in the speci-
ation continuum, rather than specific conditions that
have to be met to satisfy a particular species concept.
With the exception of R. cornivora, previous allozyme and
mtDNA work has failed to uncover diagnostically fixed
allele differences distinguishing any of the other taxa
(McPheron et al., 1988; Berlocher et al., 1993; Feder,
1998; Feder et al., 1999; Berlocher, 2000). Some taxa,
such as R. mendax and R. zephyria, do possess high-
frequency allozyme variants that are rare in other
populations (Berlocher et al., 1993; Feder, 1998; Feder
et al., 1999; Berlocher, 2000), but they are not diagnostic.
This has lead to speculation that certain R. pomonella taxa
may represent ‘quantitative genetic’ species; species that
display marked phenotypic differences in host-related
ecological adaptations causing substantial reproductive
isolation because of the cumulative effects of significant
allele frequency, but not fixed differences, across con-
tributing loci (Berlocher & Feder, 2002). If more sensitive
sequence analysis reveals the same pattern, then we
must begin to ask why fixed differences are absent; are
most of the sibling species too young for the lineage
sorting of neutral variation to be complete or is persis-
tent, low-level gene flow among taxa sufficient to
prevent alleles from becoming fixed?
Our second goal was to determine whether gene trees
for the nuclear loci are consistent with a hypothesis of
speciation mode plurality for R. pomonella. Earlier work
has suggested that introgression from Mexico helped
facilitate the sympatric radiation of the R. pomonella
complex in the USA (Feder et al., 2003a, 2005; Xie et al.,
2007). Sometime during a period of past geographic
separation �1.57 Ma, inversions appeared to have arisen
and fixed in an isolated hawthorn fly population located
in the Eje Volcanico Trans Mexicano (EVTM; Fig. 1) for
three different genomic regions on chromosomes 1, 2
and 3 (haploid n = 6 for R. pomonella; Bush, 1966).
Following secondary contact, gene flow from the EVTM
into the USA probably through the conduit of the Sierra
Madre Oriental Mountains (SMO in Fig. 1) created
adaptive inversion clines for diapause life-history traits
in the USA (Xie et al., 2007). This latitudinal diapause
variation in conjunction with additional changes in host
discrimination aided the US hawthorn fly population in
sympatrically shifting and adapting to a variety of new
plants with differing fruiting times and other physical
and chemical characteristics, generating R. mendax,
R. zephyria and the flowering dogwood fly in the process.
In historical time (< 150 years), R. pomonella also shifted
from hawthorn to introduced, domesticated apple (Malus
pumila) resulting in the formation of the apple fly race in
the north-eastern and mid-western USA (Bush, 1966).
Therefore, a portion of the diapause variation contribut-
ing to the radiation of the R. pomonella complex in the
USA originated at an earlier time and in a different
Radiation of Rhagoletis pomonella complex 901
ª 2 0 0 8 T H E A U T H O R S . J . E V O L . B I O L . 2 1 ( 2 0 0 8 ) 9 0 0 – 9 1 3
J O U R N A L C O M P I L A T I O N ª 2 0 0 8 E U R O P E A N S O C I E T Y F O R E V O L U T I O N A R Y B I O L O G Y

location than the sympatric host shifts triggering speci-
ation. In this study, we increase the sampling of nuclear
loci and taxa to confirm the chronology of the isolation of
Mexican EVTM flies relative to the radiation of the
R. pomonella complex in the USA.
Our third goal is to determine whether patterns of
genetic divergence vary across the R. pomonella genome.
Previous work implied that the inverted regions of
chromosomes 1–3 display greater levels of divergence
between Mexican and US hawthorn flies than loci
mapping elsewhere on chromosomes 4 and 5 (Feder
et al., 2005; Xie et al., 2007). From these data, we inferred
that after the initial period of introgression from Mexico,
the rearranged regions of chromosomes 1–3 may have
evolved to become more impervious to gene flow than
loci residing on other chromosomes. Through increased
sampling of sibling taxa, we investigate whether a similar
pattern is evident for the other US members of the
R. pomonella complex as well.
Methods
Flies and collection sites
Rhagoletis pomonella complex flies were genetically anal-
ysed from 14 different sites ⁄ populations, 10 from the
USA and four from Mexico (Fig. 1; see supplementary
Table S1 for complete details of the sites, including host
species and collecting dates). Two of the four Mexican
hawthorn sites [Coajomulco, Morelos (Evtm CJ) and
Tancitaro, Michoacan (Evtm MC)] reside in the Eje
Volcanico Trans Mexicano plateau (green outlined area
in Fig. 1). The other two Mexican hawthorn sites [San
Joaquin, Queretaro (Smo SJ) and Piletas, Veracruz
(Smo PL)] reside in the Sierra Madre Oriental Moun-
tains, and are located along the hypothesized conduit
for introgression from Mexico into the USA (orange
area in Fig. 1). We also sequenced Rhagoletis electromor-
pha from Dowagiac, MI, USA. to serve as an outgroup
(R. electromorpha belongs to the tabellaria species com-
plex, the sister group to R. pomonella; Bush, 1966). Flies
were sampled as larvae in infested fruit at all sites and
either immediately dissected from the fruit and frozen
for later genetic analysis or reared to adulthood in the
laboratory.
Genes sequenced
Sequence data were generated for 11 anonymous
nuclear loci and a 626-bp fragment of the mitochondrial
cytochrome oxidase II gene (see Table 1 and supple-
mentary information for details; accession numbers in
GenBank for DNA sequences are AY152477–AY152526,
AY930466–AY931013, DQ812553–DQ812885 and
EU108879–EU109174). Six of the 11 loci (P181,
P3072, P2956, P667, P7 and P22) map to chromosomes
1–3 and are subsumed by inversions (Roethele et al.,
2001; Feder et al., 2003b). Four of the remaining five
loci analysed in the study (P661, P2963, P1700 and
P309) map to chromosomes 4 and 5 (Table 1). The
exact map position of P3060 is not known, but it is not
located on chromosomes 1–3. The 11 loci analysed in
the current study are therefore broadly representative of
R. pomonella genome with respect to map position (all
five major autosomes were sequenced for at least two
genes each), location within rearranged vs. collinear
regions and the potential for host-related selection and
introgression. Consequently, if fixed genetic differences
do exist among R. pomonella complex flies, there are
reasonable expectations that they should be revealed by
the current survey.
Fig. 1 Collection sites for fly populations analysed in the study. See
supplementary Table S1 for full site information. The Rhagoletis
pomonella hawthorn fly sites Evtm CJ and Evtm MC (Coajomulco,
Morelos and Tancitaro, Michoacan, Mexico respectively) are located
in the Eje Volcanico Trans Mexicano (area outlined in green). Sites
Smo SJ and Smo PL (San Joaquin, Queretaro and Piletas, Veracruz)
are in the Sierra Madre Oriental Mountains of Mexico (area outlined
in orange). Sites Pom NY (Geneva, NY), Pom MI (Grant, MI) and
Pom TX (Brazos Bend, TX) are R. pomonella populations in the US
Sites Dog IN (Granger, IN) and Dog GA (Byron, GA) are flowering
dogwood populations. Sites Zeph WA (Dixie, Washington) and Zeph
PA (Munson, PA) are R. zephyria populations. Sites Mend MI
(Sawyer, MI) and Mend PA (Middleburg, PA) are R. mendax sites.
Site R. corn (Urbana, IL) is an R. cornivora population, whereas site
R. elect. (Dowagiac, MI) is a population of the outgroup Rhagoletis
electromorpha. Ranges for R. pomonella (red line), R. mendax (purple
line), R. zephyria (light blue line), R. cornivora (black line) and the
flowering dogwood fly (pink line) in the USA are also shown. The
majority of R. pomonella taxa are broadly sympatric in the eastern
USA. The exception is R. zephyria, which is parapatric and exten-
sively distributed in the western USA (solid light blue line), as well as
the East (stippled light blue line). Given their pattern of host plant
usage, eastern populations of R. zephyria appear to be endemic and
not introduced (Gavrilovic et al., 2007), but this hypothesis requires
further phylogeographic analysis.
902 X. XIE ET AL.
ª 2 0 0 8 T H E A U T H O R S . J . E V O L . B I O L . 2 1 ( 2 0 0 8 ) 9 0 0 – 9 1 3
J O U R N A L C O M P I L A T I O N ª 2 0 0 8 E U R O P E A N S O C I E T Y F O R E V O L U T I O N A R Y B I O L O G Y
DNA cloning and sequencing
Genomic DNA were isolated from individual flies and
PCR amplified for 35 cycles (94 �C, 30 s; 52 �C, 1 min;
72 �C, 1.5 min) using locus-specific primers for the 11
nuclear and mtDNA fragments as described in Roethele
et al. (2001). Products were TA cloned into pCR II vectors
(Invitrogen, Calsbad, CA, USA). PCR amplification prod-
ucts were initially cloned separately for a minimum of
two flies from each study site, with an attempt made to
sequence four to six clones per locus per fly in both the
5¢- and 3¢-directions on an ABI 3700 sequencer using the
ABI Prism� BigDyeTM Terminator v3.0 system (Applied
Biosystems, Foster City, CA, USA). To try and increase
sample sizes for certain sites, we also separately amplified
genomic DNA for four to eight flies from the site, and TA
cloned the pooled amplification products for sequencing.
To avoid analysis of identical alleles from the same
individual, sequences generated from the pooled library
were not included unless they differed from each other.
We were unable to generate reliable sequence data for
the Brazos Bend, Texas (Pom TX) and Geneva, New York
(Pom NY) hawthorn-infesting R. pomonella population
for the locus P1700. In addition, we only sequenced
R. mendax and R. zephyria flies from the Pennsylvania
(PA) sites for P1700 and mtDNA.
Gene tree construction and analysis
Parsimony and maximum-likelihood gene trees were
constructed using PAUP*b10 (Swofford, 2002). For the
parsimony analysis, gaps were treated as a fifth base pair,
with indels of identical length and sequence position
recoded to count as single mutational steps. Rhagoletis
electromorpha was used as an out-group taxon to root
trees. Parsimony and maximum-likelihood trees were
very similar and so we report the results for only the
parsimony trees here. Intragenic recombination was
statistically tested using the methods of Hudson & Kaplan
(1985). The molecular clock was tested for each locus for
R. pomonella and R. electromorpha sequences by comparing
log-likelihood scores enforcing vs. relaxing the clock
hypothesis for the best supported DNA substitution model
identified using MODELTEST (Posada & Crandall, 1998).
Neighbour-joining trees (Saitou & Nei, 1987) summa-
rizing the overall genetic relatedness of populations were
constructed using PHYLIP v 3.66 (Felsenstein, 1989).
Trees were constructed separately for the six loci map-
ping to chromosomes 1–3 plus P1700 and for the
remaining four loci residing elsewhere in the genome.
To construct the neighbour-joining trees, mean pairwise
uncorrected genetic distances were first computed sepa-
rately for each locus between each pair of populations
(except the PA sites), as well as between these popula-
tions and the outgroup R. electromorpha using Mega v3.1
(Kumar et al., 2004). The pairwise distance between two
populations for a locus was then divided by the average
distance of all R. pomonella populations to R. electromorpha
to standardize for sequence length and substitution rate
differences among loci. For the two SMO populations,
two diverged haplotypes could generally be identified at
loci, one more closely related to Mexican EVTM alleles
and the other to SN alleles from the USA (see Figs 2–4
and supplementary Figs S1–S4). We considered these
haplotype classes as separate SMO populations in the
calculations of genetic distance. Similarly, for the chro-
mosome 1–3 loci and P1700, taxa possessing North (N)
and South–North (SN) haplotypes at a locus were
considered different populations in genetic distance
calculations. Standardized distances were then averaged
across P181, P3072, P2956, P667, P7, P22 and P1700 and
across P661, P2963, P309 and P3060 to give the overall
pairwise distances used for tree construction. Because of
the variable presence of N haplotypes in taxa other than
R. pomonella, we only included the N haplotype popula-
tion from NY as a general representative of the N clade in
the network.
Unfortunately, other analytical approaches, such as the
isolation model with migration (IM or IMa; Nielsen &
Wakeley, 2001; Hey & Nielsen, 2007), are not readily
applicable to the R. pomonella data set because of viola-
tions of underlying assumptions of the model, including
that there be no unsampled populations exchanging
genes with the sampled populations or their ancestor, no
directional or balancing selection acting on sites (selec-
tive neutrality), no recombination within loci and free
recombination between loci.
Analysis of population differentiation and structure
A hierarchical analysis of molecular variance (AMOVAAMOVA)
was performed using Arlequin 2.0 (Schneider et al.,
Table 1 Loci sequenced in the study.
Locus Chr. Seq. Ln P Model Rec.
P181 1 288 1.000 TrN + I 2
P3072 1 465 0.377 TIM 5
P2956 2 510 0.244 TrN + I 5
P667 2 583 0.866 TVM + G 5
P22 3 593 0.757 HKY 2
P7 3 736 0.576 TrN + I 2
P2963 4 377 0.756 GTR + G 3
P661 4 436 0.088 TVM + G 6
P1700 5 484 0.997 HKY + G 6
P309 5 482 1.000 K81uf + G 3
P3060 ? 562 0.999 HKY 0
mtDNA – 626 0.990 TrN + I 0
Given are chromosome map positions (Chr.), the length of the
sequence in base pairs (Seq. Ln), the probability level (P) for
conformation to a molecular clock, the ML substitution model
(model) determined by MODELTEST using the AIC criterion (Posada
& Crandall, 1998), and the minimum number of recombination
events (Rec.) estimated by the method of Hudson & Kaplan (1985)
for each locus.
Radiation of Rhagoletis pomonella complex 903
ª 2 0 0 8 T H E A U T H O R S . J . E V O L . B I O L . 2 1 ( 2 0 0 8 ) 9 0 0 – 9 1 3
J O U R N A L C O M P I L A T I O N ª 2 0 0 8 E U R O P E A N S O C I E T Y F O R E V O L U T I O N A R Y B I O L O G Y
2000) to test for genetic structuring among populations.
For the AMOVAAMOVAs, populations were divided into six
groups: (1) EVTM R. pomonella sites (MC and CJ); (2)
SMO R. pomonella sites (SJ and PL); (3) US R. pomonella
sites (Pom MI, NY and TX); (4) flowering dogwood fly
sites (Dog IN and GA); (5) R. mendax (Mend MI site); and
(a) (b)
(c) (d)
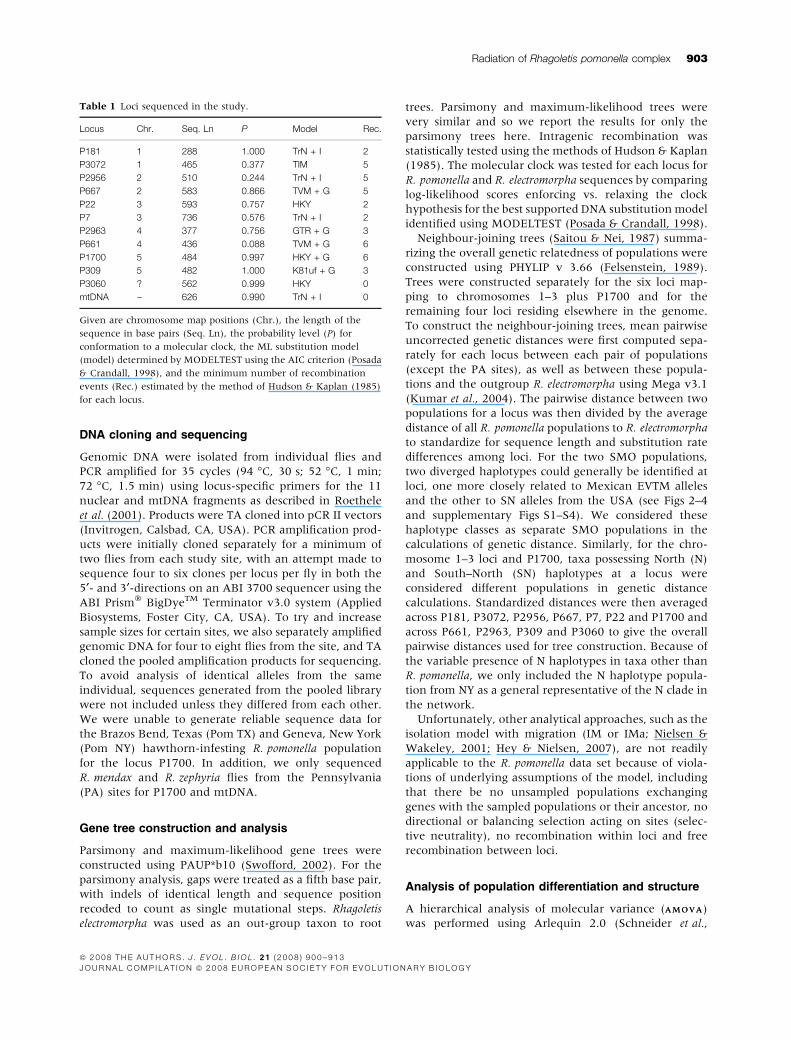
Fig. 2 Most parsimonious gene trees for (a) P3072, (b) P22, (c) P2956 and (d) mtDNA. Species and site designations are given in Fig. 1
legend and supplementary Table S1. Trees are scaled so that the longest distance from an allele to the outgroup Rhagoletis electromorpha
(R. elect.) are relatively the same across loci. Chromosome position for loci, sequence lengths (no. of bp), branch lengths (no. of steps), and
bootstrap support for nodes (10 000 reps.) are given. The three numbers in the arrow brackets (< >) following site designation for an allele
indicate the number of identical alleles sequenced for the population, the number of alleles sequenced that differ by a single substitution
from the haplotype and the number of alleles that differ by two substitutions respectively. N (blue coloured) designates North US alleles,
SN (red) = South ⁄ North alleles, SMO (orange) = Mexican Sierra Madre Oriental alleles and EVTM (green) = Mexican Eje Volcanico Trans
Mexicano. Yellow-coloured circles demark the relative depth of the node connecting N alleles for nuclear loci and SN alleles for mtDNA
with EVTM haplotypes. The congruence of relative N vs. SN ⁄ SMO ⁄ EVTM node depths among chromosomes 1–3 loci and mtDNA (compare
yellow-coloured nodes) is hypothesized to be a consequence of the past geographic separation of EVTM and US hawthorn fly populations
�1.57 Ma (Feder et al., 2003a). Black-coloured nodes represent the shallowest relative node depth connecting SN and EVTM haplotypes.
Comparisons of these nodes to those in Fig. 4a–d highlight the deeper coalescence times of SN and EVTM haplotypes between chromosome
1–3 loci, hypothesized to be because of the greater permeability and more recent gene flow of collinear loci outside of the rearrangements on
chromosomes 1–3.
904 X. XIE ET AL.
ª 2 0 0 8 T H E A U T H O R S . J . E V O L . B I O L . 2 1 ( 2 0 0 8 ) 9 0 0 – 9 1 3
J O U R N A L C O M P I L A T I O N ª 2 0 0 8 E U R O P E A N S O C I E T Y F O R E V O L U T I O N A R Y B I O L O G Y

(6) R. zephyria (Zeph WA site). Separate AMOVAAMOVAs were
performed for the six chromosome 1–3 loci and P1700
with N haplotypes both excluded ()N) and included
(+N). Neither R. cornivora nor R. electromorpha were
included in the AMOVAAMOVAs, as gene trees showed these
taxa to be basal and their inclusion would have inflated
the per cent variation explained by among-group differ-
ences for the other ‘in-group’ taxa in the analysis.
Tamura & Nei (1993) genetic distances between in-group
taxa and R. electromorpha for loci were calculated using
Mega v3.1 (Kumar et al., 2004).
Results
None of the 11 sequenced nuclear genes or mtDNA
deviated significantly from a molecular clock (Table 1).
All nuclear loci except P3060 displayed evidence for
possible recombination, as implied by the method of
Hudson and Kaplan (Table 1). Inferred recombination
was generally limited, however, to alleles within the
same haplotype class for loci on chromosomes 1–3 or
same geographic population ⁄ taxa. There was no evidence
for recombination among mtDNA sequences (Table 1).
Confirmation of clock-like evolution and intra-haplo-
type-limited recombination allowed for inferences to be
drawn concerning the three goals of the study based on
comparisons of branching topologies and node depths
among gene trees. We organize the presentation of these
results below into three subsections highlighting the
similarities and differences in patterns of genetic differ-
entiation for: (1) the six loci on chromosomes 1–3 (genes
residing in inverted regions of the genome); (2) the five
loci mapping to chromosomes 4 and 5 (genes in putative
co-linear regions of the genome); and (3) mtDNA.
Genes on chromosomes 1–3
Several features characterized the gene trees for the six
loci mapping to chromosomes 1–3 (Fig. 2a–c and sup-
plementary Figs S1–S3). First, R. cornivora (designated by
R. corn. in the figures) was clearly basal to the other
R. pomonella populations, displaying fixed, autapomor-
phic substitutions for P181, P3072, P2956, P667, P7 and
P22.
Second, with the exception of P7, gene trees for
chromosome 1–3 loci resolved US and Mexican R. pomo-
nella, R. zephyria, R. mendax and the flowering dogwood
fly as a distinct monophyletic ‘in-group’ clade from
R. cornivora (Fig. 2a–c and supplementary Figs S1–S3).
Third, within the ‘in-group’ taxa of US and Mexican
R. pomonella populations, R. zephyria, R. mendax and the
flowering dogwood fly, two distinct clades ⁄ classes of
haplotypes could be identified for each locus. One clade
consisted of North haplotypes (designated by the blue-
coloured letter ‘N’ in Fig. 2a–c and supplementary
Figs S1–S3) found in northern populations of R. pomo-
nella (NY and MI), and to varying degrees, depending on
the locus, in R. mendax, R. zephyria and the flowering
dogwood fly as well. The other clade was comprised of a
trinity of haplotypes present: (1) in northern and
southern populations of US R. pomonella, R. mendax,
R. zephyria and the flowering dogwood fly (designated
by the red-coloured letters ‘SN’ in the figures); (2) in the
Sierra Madre Oriental population of Mexican hawthorn
flies (designated by the orange-coloured letters ‘SMO’);
and (3) in the Eje Volcanico Trans Mexicano region
(designated by the green-coloured letters ‘EVTM’).
Fourth, the depths of the nodes uniting the N clade and
the triad of SN ⁄ SMO ⁄ EVTM haplotypes relative to the
(a)
(b)
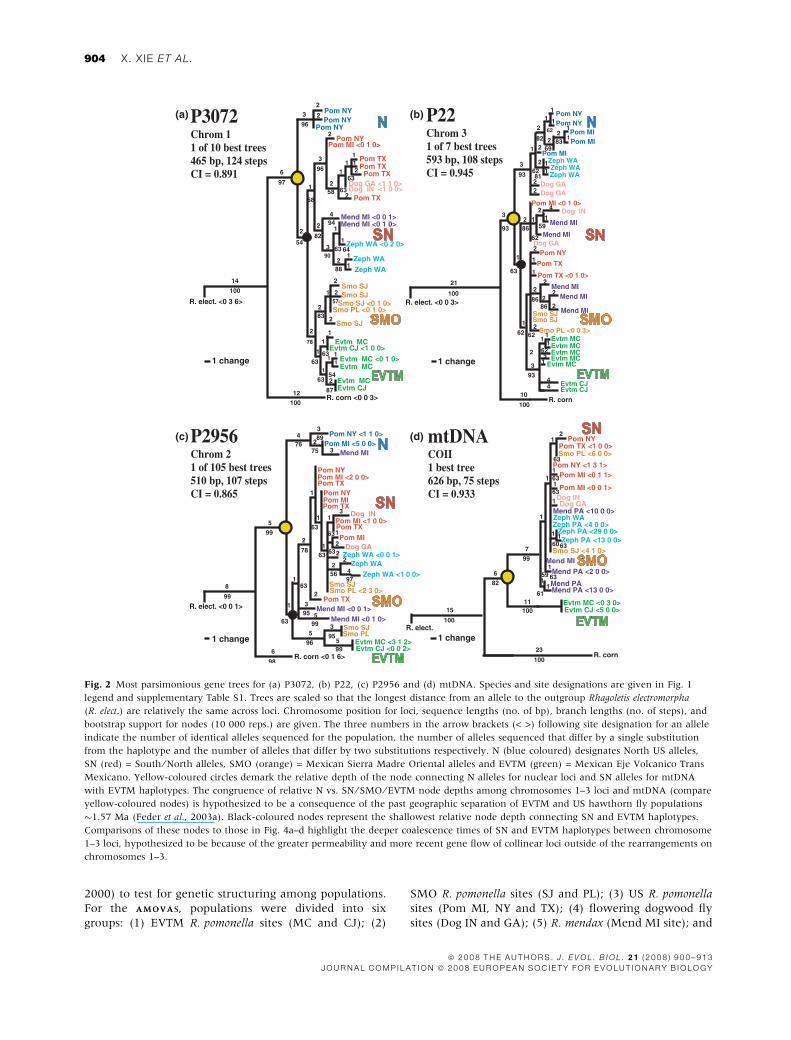
Fig. 3 Neighbour-joining trees based on overall genetic distances
among Rhagoletis pomonella taxa relative to the outgroup taxon
Rhagoletis electromorpha for (a) chromosome 1–3 loci and P1700 and
(b) loci not residing on chromosomes 1–3. Trees are scaled so that the
lengths of the branch from the R. electromorpha ⁄ R. pomonella node to
terminal populations are the same in (a) and (b). For chromosome
1–3 loci, the N and SN clades of haplotypes present were considered as
separate populations. Only the NY N haplotypes were included in the
network to serve as a general indicated of the divergence of this clade
of alleles in the USA. For the SMO, alleles displaying affinity to SN
haplotypes and EVTM haplotypes were also treated as separate
populations. The N vs. SN ⁄ SMO ⁄ EVTM haplotype node is designated
by the compare yellow-coloured circle in (a) and the SN vs. EVTM
haplotype nodes by the black-coloured circle in (a) and (b).
Radiation of Rhagoletis pomonella complex 905
ª 2 0 0 8 T H E A U T H O R S . J . E V O L . B I O L . 2 1 ( 2 0 0 8 ) 9 0 0 – 9 1 3
J O U R N A L C O M P I L A T I O N ª 2 0 0 8 E U R O P E A N S O C I E T Y F O R E V O L U T I O N A R Y B I O L O G Y
outgroup R. electromorpha were similar among loci and
concurred with a deep division seen for mtDNA between
EVTM flies in Mexico and the remainder of the in-group
taxa (compare yellow-coloured nodes in Fig. 2a–c and
supplementary Figs S1–S3 among loci). The congruence
of these nodes among gene trees is hypothesized to
reflect the initial isolation of the Mexican EVTM popu-
lation that predated the secondary contact and introgres-
sion of nuclear SN haplotypes into the USA.
Fifth, aside from P7, the Mexican population of
hawthorn-infesting R. pomonella in the EVTM designated
by the green-coloured haplotypes was monophyletic and
displayed fixed, autapomorphic differences from the
other taxa for the five other nuclear loci on chromosomes
1–3 (Fig. 2a–c and supplementary Figs S1–S3).
Sixth, the hawthorn-infesting populations of R. pomo-
nella in the SMO generally contained a mixture of alleles
that showed affinity to both the EVTM and the USA. For
P3072 and P667, SMO flies possessed EVTM haplotypes,
forming a monophyletic clade (Fig. 2a and supplemen-
tary Fig. S2). For P22 and P181, SMO flies had alleles
embedded within the SN class of haplotypes present in
US R. pomonella, R. mendax, R. zephyria and flowering
dogwood fly populations (Fig. 2b and supplementary
Fig. S1). The locus 2956 had haplotypes related to both
EVTM and the SN clades of alleles (Fig. 2b). Finally, P7
(supplementary Fig. S1) contrasted with the other chro-
mosome 1–3 genes in that SMO alleles formed a distinct
clade genealogically unrelated to either EVTM or SN
haplotypes.
Seventh, no fixed diagnostic substitution distinguished
R. pomonella, R. mendax, R. zephyria and the flowering
dogwood fly in the USA from each other for P3072, P181,
P2956, P667 or P7, although an autapomorphy possibly
(a) (b)
(c) (d)
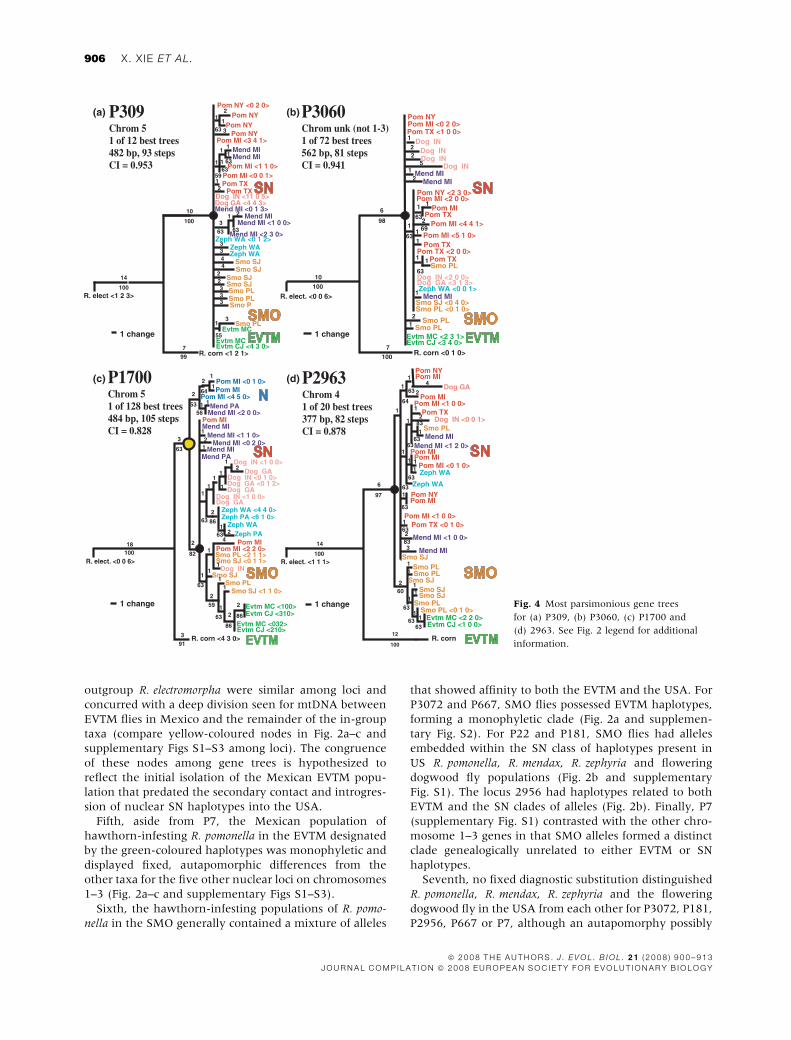
Fig. 4 Most parsimonious gene trees
for (a) P309, (b) P3060, (c) P1700 and
(d) 2963. See Fig. 2 legend for additional
information.
906 X. XIE ET AL.
ª 2 0 0 8 T H E A U T H O R S . J . E V O L . B I O L . 2 1 ( 2 0 0 8 ) 9 0 0 – 9 1 3
J O U R N A L C O M P I L A T I O N ª 2 0 0 8 E U R O P E A N S O C I E T Y F O R E V O L U T I O N A R Y B I O L O G Y
exists for R. zephyria at P22 (Fig. 2a–c and supplementary
Figs S1–S4). However, private alleles were observed at
several genes for R. mendax and R. zephyria, and much
less so for the flowering dogwood fly.
The neighbour-joining network based on overall rel-
ative genetic distance measures among populations for
chromosome 1–3 loci and P1700 (Fig. 3a) highlighted the
general patterns discerned from the individual gene trees,
including: (1) the basal position for R. cornivora in the
pomonella complex; (2) the presence of a deep northern
clade of alleles (designated by Pom-NY) within the
in-group taxa in the USA; (3) the existence of a distinct,
monophyletic clade of Mexican hawthorn fly haplotypes
in the EVTM; and (4) the composite nature of the
Mexican hawthorn fly population in the SMO, contain-
ing genetic elements with affinity to both EVTM and US
haplotypes. The network also inferred that R. pomonella
flies in the USA are overall genetically most similar to
flowering dogwood flies, followed by R. zephyria, and
then R. mendax. In addition, the class of alleles within the
SMO showing affinity to SN haplotypes in the USA is
most closely related to R. pomonella, and, in particular,
the TX population.
Genes mapping outside chromosomes 1–3
The five loci mapping outside chromosomes 1–3 were
similar to the six chromosome 1–3 loci in several
respects. Gene trees for P661, P2963, P1700, P309 and
P3060 clearly positioned R. cornivora basal to the other
members of the R. pomonella complex (Fig. 4a–d and
supplementary Fig. S4). The in-group taxa comprised of
US and Mexican R. pomonella, R. zephyria, R. mendax and
the flowering dogwood fly were also monophyletic for
P661, P2963, P1700, P309 and P3060, sharing several
synapomorphic substitutions in common. In addition,
P661, P2963, P1700, P309 and P3060 showed substantial
genetic differentiation among R. pomonella populations,
as indicated by significant FCT values in a hierarchical
AMOVAAMOVA, similar to the six chromosome 1–3 loci (Table 2).
Indeed, P1700 displayed two potentially fixed autapo-
morphic substitutions diagnostically distinguishing
R. zephyria from all other R. pomonella complex flies
(Fig. 4c). The flowering dogwood fly also possessed a
clade of private haplotypes for P1700, as did R. mendax for
the loci P309 and P1700 (Fig. 4a) and R. zephyria for P661
(supplementary Fig. S4).
There were several pronounced differences, however,
between loci residing on chromosomes 1–3 and most
genes mapping elsewhere in the genome. First, except for
P1700, no locus mapping outside of chromosomes 1–3
possessed a highly diverged N haplotype class of alleles
(Fig. 4b–d and supplementary Fig. S4). Second, no
monophyletic clade of Mexican EVTM haplotypes was
present for P309, P3060 or P661 (Fig. 4a,b and supple-
mentary Fig. S4). Third, Mexican SMO alleles for P309,
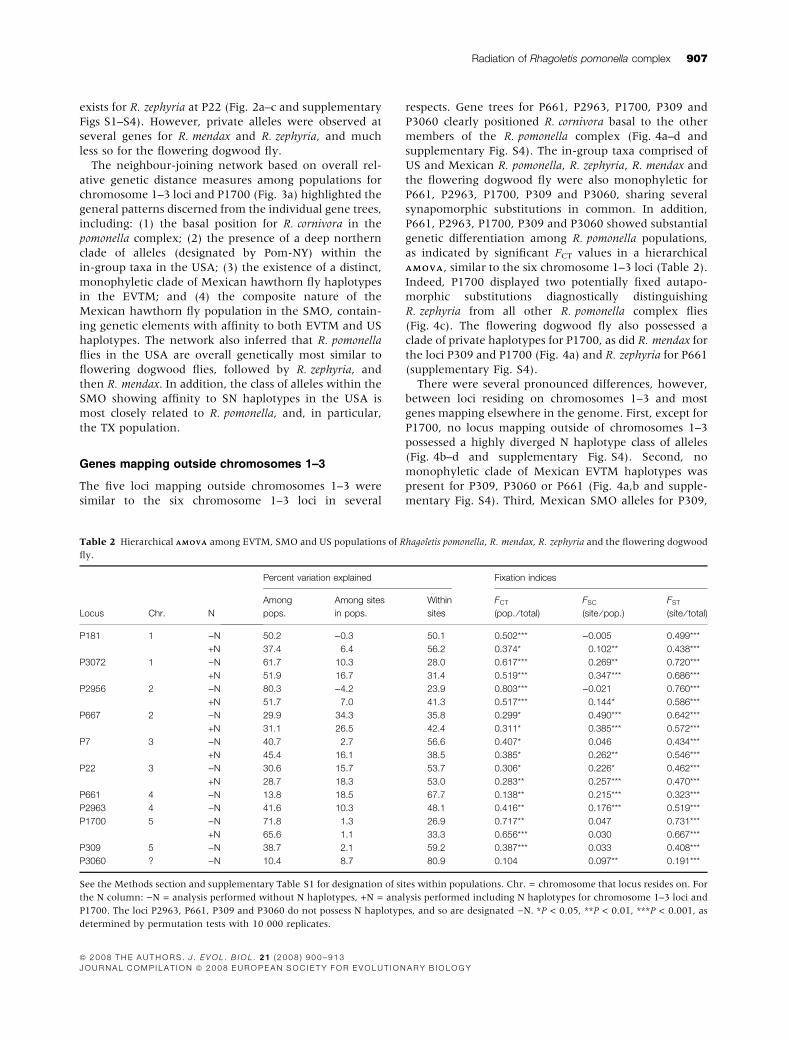
Table 2 Hierarchical AMOVAAMOVA among EVTM, SMO and US populations of Rhagoletis pomonella, R. mendax, R. zephyria and the flowering dogwood
fly.
Locus Chr. N
Percent variation explained Fixation indices
Among
pops.
Among sites
in pops.
Within
sites
FCT
(pop. ⁄ total)
FSC
(site ⁄ pop.)
FST
(site ⁄ total)
P181 1 )N 50.2 )0.3 50.1 0.502*** )0.005 0.499***
+N 37.4 6.4 56.2 0.374* 0.102** 0.438***
P3072 1 )N 61.7 10.3 28.0 0.617*** 0.269** 0.720***
+N 51.9 16.7 31.4 0.519*** 0.347*** 0.686***
P2956 2 )N 80.3 )4.2 23.9 0.803*** )0.021 0.760***
+N 51.7 7.0 41.3 0.517*** 0.144* 0.586***
P667 2 )N 29.9 34.3 35.8 0.299* 0.490*** 0.642***
+N 31.1 26.5 42.4 0.311* 0.385*** 0.572***
P7 3 )N 40.7 2.7 56.6 0.407* 0.046 0.434***
+N 45.4 16.1 38.5 0.385* 0.262** 0.546***
P22 3 )N 30.6 15.7 53.7 0.306* 0.226* 0.462***
+N 28.7 18.3 53.0 0.283** 0.257*** 0.470***
P661 4 )N 13.8 18.5 67.7 0.138** 0.215*** 0.323***
P2963 4 )N 41.6 10.3 48.1 0.416** 0.176*** 0.519***
P1700 5 )N 71.8 1.3 26.9 0.717** 0.047 0.731***
+N 65.6 1.1 33.3 0.656*** 0.030 0.667***
P309 5 )N 38.7 2.1 59.2 0.387*** 0.033 0.408***
P3060 ? )N 10.4 8.7 80.9 0.104 0.097** 0.191***
See the Methods section and supplementary Table S1 for designation of sites within populations. Chr. = chromosome that locus resides on. For
the N column: )N = analysis performed without N haplotypes, +N = analysis performed including N haplotypes for chromosome 1–3 loci and
P1700. The loci P2963, P661, P309 and P3060 do not possess N haplotypes, and so are designated )N. *P < 0.05, **P < 0.01, ***P < 0.001, as
determined by permutation tests with 10 000 replicates.
Radiation of Rhagoletis pomonella complex 907
ª 2 0 0 8 T H E A U T H O R S . J . E V O L . B I O L . 2 1 ( 2 0 0 8 ) 9 0 0 – 9 1 3
J O U R N A L C O M P I L A T I O N ª 2 0 0 8 E U R O P E A N S O C I E T Y F O R E V O L U T I O N A R Y B I O L O G Y

P3060 and P661 were not distinct and were embedded in
the gene tree with SN alleles from the USA; these three
loci provided no evidence for the SMO having genetic
affinity to the EVTM. Finally, although a significant
proportion of the genetic variation present for P309,
P3060, P2963 and P661 was partitioned among taxa, the
relative degree of genetic divergence separating the
alleles displaying variation was less for these loci than
for the six loci residing on chromosomes 1–3 (compare
the depths of the black-coloured circles signifying the
nodes uniting EVTM and SN between gene tree in Fig. 4
and supplementary Fig. S4 vs. Fig. 2 and supplementary
Figs S1–S3, as well as between the neighbour-joining
networks in Fig. 3a,b; note: P1700 was included with the
six chromosome 1–3 loci for network construction
because of the observation of distinct N and SN haplo-
types at the locus). The shallower divergence among
alleles for nonchromosomes 1–3 loci was not because
of a slower rate of evolution for these genes (mean
Tamura–Nei distance for in-group R. pomonella taxa to
R. electromorpha for loci not on chromosomes
1–3 = 0.053 ± 0.0152, n = 4, range 0.032–0.066; mean
for chromosome 1–3 loci = 0.046 ± 0.0222, n = 6, range
0.020–0.085; distance for P1700 = 0.056 ± 0.0104).
Moreover, the gene trees in Figs 2 and 4 are all scaled
relative to R. electromorpha and the networks in Fig. 3 are
based on relative, not raw, genetic distances to R. electro-
morpha. Rather, the difference reflects the greater genetic
similarity of P661, P2963, P309 and P3060 alleles among
the in-group R. pomonella taxa than P181, P3072, P2956,
P667, P7, P22 and P1700 haplotypes. As a consequence,
the neighbour-joining network for P661, P2963, P309
and P3060 (Fig. 3b) was less well resolved than that for
the chromosome 1–3 loci and P1700 (Fig. 3a), with US
R. pomonella and flowering dogwood fly populations
appearing paraphyletic and R. zephyria being embedded
within the Mexican SMO populations.
MtDNA
MtDNA displayed both similarities and differences from
the patterns of genetic variation observed for the nuclear
loci. Rhagoletis cornivora was again basal in the R. pomo-
nella complex. Like P661, P2963, P309 and P3060,
mtDNA did not display a highly diverged N haplotype
confined to northern US fly populations (Fig. 2d). How-
ever, the EVTM and the in-group R. pomonella taxa
showed a deep genetic divergence for mtDNA that was
congruent with the relative node depths between N and
SN ⁄ SMO ⁄ EVTM clades of haplotypes for chromosomes
1–3 loci (Fig. 2). However, there was no signature for
subsequent introgression in the mtDNA between the
EVTM and US or SMO fly populations. Moreover, unlike
the chromosome 1–3 loci, but like P661, P309 and P3600,
mtDNA, haplotypes for SMO flies were not a mixture of
EVTM and US types. Instead, they were all embedded
within and very similar to SN haplotypes found in the
USA (Fig. 2d). There was no diagnostic mtDNA autapo-
morphy distinguishing R. pomonella, R. mendax, R. zephy-
ria, the flowering dogwood fly or SMO flies for COII
(Fig. 2d). However, R. mendax and R. zephyria appeared
to possess potential private substitutions and, on this
basis, were the most genetically differentiated of the
in-group taxa for mtDNA.
Discussion
Genetic signposts for speciation
The first goal of the current sequence study was to
determine whether genetically distinguishable stages
of population divergence could be identified in the
R. pomonella complex. Our results suggest that on a
coarse, qualitative scale such stages may be recognized
with respect to: (1) R. cornivora (fixed, reciprocal mono-
phyly for all loci); (2) EVTM hawthorn flies (fixed
substitutions and monophyly for several, but not all
loci); (3) SMO hawthorn flies (fixed differences for a
couple of loci and some private alleles for a handful of
genes); (4) the in-group sibling taxa in the USA (possible
autapomorphies and private alleles at a few loci); and (5)
the apple and hawthorn host races (no fixed substitution
or private allele difference). The sequence data did not
reveal any finer genetic resolution than previous allo-
zymes studies, however (Feder & Bush, 1989; Berlocher
et al., 1993; Feder, 1998; Feder et al., 1999; Berlocher,
2000). Moreover, neither nuclear or mtDNA loci clearly
resolved the phylogenetic relationships (i.e. a bifurcating
gene tree) among the in-group populations of R. pomo-
nella, R. mendax, R. zephyria and flowering dogwood fly in
the USA. Reciprocal monophyly was not observed for
any locus among these four in-group taxa and it is
conceivable that with increased sampling the potential
private and autapomorphic variants seen for R. mendax
and R. zephyria for a couple of genes in the current study
will prove to be shared among taxa, albeit at low
frequencies, as is the case for allozymes. Such an
outcome is suggested by RFLP data for P1700 for
R. mendax and R. zephyria (Schwarz et al., 2005). The
sequence analysis for R. pomonella flies therefore did not
codify a clear demarcation of taxonomic status for US
populations based on phylogenetic or lineage concepts of
species, although the neighbour-joining network based
on overall relative genetic distances measures for chro-
mosome 1–3 loci and P1700 did imply that R. pomonella
and the flowering dogwood fly are sister taxa, followed
by R. zephyria and R. mendax (Fig. 3a).
There are two interpretations for the general lack of
phylogenetic resolution of the DNA data for the USA in-
group taxa. The first explanation is that diagnostic
differences exist, but that the sequencing of 11 nuclear
loci and mtDNA was not an extensive enough survey of
the genome to detect these differences. The current data
for the in-group taxa therefore mainly reflect the
908 X. XIE ET AL.
ª 2 0 0 8 T H E A U T H O R S . J . E V O L . B I O L . 2 1 ( 2 0 0 8 ) 9 0 0 – 9 1 3
J O U R N A L C O M P I L A T I O N ª 2 0 0 8 E U R O P E A N S O C I E T Y F O R E V O L U T I O N A R Y B I O L O G Y

consequences of incomplete lineage sorting among the
relatively recently diverged in-group fly populations in
the USA. The second interpretation is that the over 30
allozymes and 11 nuclear loci analysed to date actually
do represent an accurate snapshot of the state of genetic
differentiation among the in-group R. pomonella taxa. If
true, then these taxa are probably best characterized as a
continuum of quantitative genetic sibling species and
host races distinguishable as sympatric genotypic clusters
(Mallet, 1995) on the basis of near private mutations
and ⁄ or allele frequency differences (Feder, 1998). Mem-
bers of the group would therefore reflect a dynamic
equilibrium between low-level gene flow and the
strength of differential host selection, forming a complex
analogous to the syngameons of plants. The recent
findings of Schwarz et al. (2005) of an historical hybrid-
ization event between R. mendax and R. zephyria poten-
tially giving rise to a new race ⁄ species of fly in the eastern
USA infesting introduced honeysuckle from the Lonicera
tatarica complex are consistent with this dynamic equi-
librium view for the R. pomonella group.
Although the current data failed to reveal reciprocal
monophyly among the R. pomonella in-group taxa in the
USA, there were several a priori reasons to believe that
the 11 anonymous nuclear loci sequenced in the study
should have been sufficient to uncover fixed, diagnostic
differences (autapomorphies) provided they exist. First,
the in-group taxa R. pomonella, R. mendax, R. zephyria and
the flowering dogwood fly possess differential host-
associated adaptations related to diapause and host
discrimination that serve as significant ecological barriers
to gene flow (Smith, 1986; Bierbaum & Bush, 1990;
Feder et al., 1993, 1994; Linn et al., 2003; Schwarz et al.,
2007). Second, R. pomonella · R. mendax and R. men-
dax · R. zephyria display a degree of nonhost-related
prezygotic isolation. In particular, male R. mendax have
difficulty mating with female R. pomonella because of a
difference in adult body size between the species (Smith,
1986; J. L. Feder, pers. obs.) and even the similar-sized
R. mendax and R. zephyria show significant sexual isola-
tion (Schwarz & McPheron, 2007). Third, R. pomonella
and R. mendax also exhibit post-zygotic isolation (over
50%) that could be because of intrinsic genomic incom-
patibility and ⁄ or host fruit-related larval survivorship
differences (Smith, 1986; J. L. Feder, pers. obs.). No
prezygotic isolation appears to exist between R. pomonella
and the flowering dogwood fly. However, there is subtle
evidence for possible low-level post-zygotic isolation.
Although there is no reduction in fecundity in apple
race · dogwood fly matings, backcrosses of F1 hybrids to
parental taxa suggest that egg hatch may be reduced by
10% for second generation flies of mixed apple ⁄ dogwood
ancestry (Smith, 1986). Fourth, although there is also no
evidence for reduced fecundity or fertility in hybrid
crosses between the apple and hawthorn host races of
R. pomonella (Reissig & Smith, 1978; Smith, 1986), F1
apple · hawthorn flies fail to orient to host fruit volatiles
in flight tunnel assays (Linn et al., 2004; Dambroski et al.,
2005) indicative of an impaired olfactory system that
could have significant fitness consequences in the field.
Fifth, the 11 nuclear loci we sequenced in the study
representatively cover five of the six chromosomes
constituting the R. pomonella genome (we had no marker
on the small dot sixth chromosome). Moreover, six of
these loci reside in inverted regions on chromosomes 1–3
that are in linkage disequilibrium with allozymes show-
ing host-related differences among R. pomonella taxa
(Roethele et al., 2001; Feder et al., 2003b). These rear-
ranged regions contain genes affecting diapause life-
history variation that are responsible for generating
reproductive isolation between the sibling species and
host races (Feder et al., 1997a, b; Filchak et al., 2000). As
such, these six sequenced loci are located in what could
be considered ‘islands of speciation’ for R. pomonella and
are prime targets for reflecting population divergence.
Nevertheless, moving from the temporally flat, infinite-
allele framework of the allozymes to the genealogically
deeper, infinite-site perspective provided by the current
DNA sequence data did not appreciably change the
resulting view of the R. pomonella in-group taxa. It is
conceivable that advances in molecular techniques will
soon make it possible to cost effectively sequence a large
portion of the genome of R. pomonella for variation and
reveal an essential core of fixed, host-related substitu-
tions defining the different species. However, we must
also entertain the possibility that even more exhaustive
searching of the in-group taxa will not greatly change the
implications of this study; R. pomonella host races and
sibling species represent a seamless transition that form
without the complete closure of the genome to gene flow
and in the absence of the evolution of fixed genetic
differences. Only when we move to the level of R. corn-
ivora at the base of the R. pomonella complex where
nearly complete pre- and post-zygotic isolations are
present (Berlocher, 2000) do we observe reciprocally
discreet genetic differences throughout the genome.
Speciation mode plurality
With respect to the second issue of speciation mode
plurality, the sequence data are consistent with the
hypothesis that past geographic isolation and subsequent
introgression of inversion polymorphism from an isolated
hawthorn-infesting fly population of R. pomonella in the
EVTM of Mexico played a role in the adaptive radiation
of the USA in-group taxa (Feder et al., 2003a). From this
study, we could infer that the initial isolation of the
EVTM population in Mexico (estimated at �1.57 Ma
based on a mtDNA insect molecular clock; Feder et al.,
2003a) occurred after the divergence of R. cornivora from
the rest of the complex. Moreover, the introgression of
inversions from the EVTM into the USA appears to have
preceded the genesis of R. mendax, R. zephyria and the
flowering dogwood fly, as well as the apple race.
Radiation of Rhagoletis pomonella complex 909
ª 2 0 0 8 T H E A U T H O R S . J . E V O L . B I O L . 2 1 ( 2 0 0 8 ) 9 0 0 – 9 1 3
J O U R N A L C O M P I L A T I O N ª 2 0 0 8 E U R O P E A N S O C I E T Y F O R E V O L U T I O N A R Y B I O L O G Y

Additional information is needed, however, to assess the
current taxonomic status of EVTM flies. Hawthorn-
infesting populations of R. pomonella in the EVTM of
Mexico clearly formed a monophyletic clade from the
rest of the group for several loci. The EVTM population
also does not presently appear to be in contact with
hawthorn-infesting populations in the SMO or USA (Rull
et al., 2006). The genetics and biogeography of EVTM
population are therefore consistent with these flies
currently being completely reproductively isolated from
the other R. pomonella taxa and potentially having been
formed via an allopatric mode of speciation not involving
host shifting. However, further studies involving crosses
of EVTM to SMO and US flies are needed to test for
nonhost-related reproductive isolation to confirm this
hypothesis.
The status of hawthorn flies in the SMO of Mexico is
also not definitive. The SMO population contains genetic
elements from both the EVTM and the USA, consistent
with it having served as a conduit for past gene flow
connecting the EVTM with an ancestral US population of
hawthorn-infesting flies. Incomplete lineage sorting in
combination with low levels of recombination in inverted
regions of chromosomes 1–3 could contribute to the
mosaic nature of the SMO gene pool, but this explanation
alone is insufficient to account for the totality of the
pattern. In the neighbour-joining networks (Fig. 3), the
alleles in the SMO showing affinity to SN haplotypes in
the USA are more closely related to each other than are
those genes in the SMO and EVTM showing affinity. In
addition, mtDNA indicates a close genetic relationship
between SMO and US R. pomonella populations (Fig. 2d).
These data imply a temporal dynamic of differential
introgression, with more extensive and recent gene flow
between the USA and the SMO than between the SMO
and EVTM. Indeed, it is not obvious whether the
differences seen between SMO and US hawthorn fly
populations are indicative of interspecific divergence or
conspecific geographic variation. Tentative support for
the latter hypothesis comes from a recent survey of
microsatellites in SMO and US hawthorn flies revealing
clinal variation for several loci between the regions
(Michel et al., 2007). More extensive genetic surveys
and tests for nonhost-related reproductive isolation are
needed to clarify the taxonomic status of SMO flies.
Patterns of genetic differentiation across the genome
Our third goal was to assess patterns of genetic differ-
entiation across the R. pomonella genome. In this regard,
with the exception of P1700, there was a general trend
for loci residing on chromosomes 1–3 to display more
pronounced levels of allelic divergence within and
among populations than genes mapping to chromo-
somes 4 and 5. Previous studies have documented this
pattern among EVTM, SMO and US hawthorn-infesting
populations of R. pomonella (Feder et al., 2005; Xie et al.,
2007). Here, we observe that the same pattern holds for
the in-group taxa R. mendax, R. zephyria and the flow-
ering dogwood fly. The result implies that the rear-
ranged regions of chromosomes 1–3 have become less
permeable to gene flow among fly taxa than loci in
putative co-linear regions of the genome, consistent
with recent inversion models for speciation (Noor et al.,
2001a; Rieseberg, 2001; Navarro & Barton, 2003;
Kirkpatrick & Barton, 2006). Against this backdrop,
the lack of a signature of mtDNA introgression between
the EVTM and SMO ⁄ US populations since the initial
isolation of Mexican highland flies �1.57 Ma is puz-
zling. Possible explanations for the disjunct nature of
mtDNA variation between the EVTM and SMO ⁄ USA
include male-driven gene flow, cytonuclear gene
incompatibilities, and ⁄ or a factor directly under differ-
ential selection in the mtDNA.
A delta of life?
In conclusion, our studies of the R. pomonella complex
suggest that many of the dichotomies we impose
concerning geographic modes ⁄ mechanisms of diver-
gence, the cladistic splitting of taxa and systematic
categories of organisms may blur during speciation.
Rather than the analogy of a branching ‘tree of life’, a
‘delta of life’ comprised of many inter-tangled channels
may be more appropriate for describing the formative
stages of R. pomonella speciation. Thus, a plurality of
mechanisms and processes including host-associated
selection, sympatric host shifts, biogeography, secondary
contact, past differential introgression, inversions, clines
and ongoing and low-level hybridization probably inter-
act to generate what we call species in the R. pomonella
complex. These species do not necessarily display
reciprocal monophyly at their inception. Rather, they
exist as genotypic clusters that can be statistically iden-
tified on the basis of near private alleles and frequency
differences at a subset of loci resulting from differential
selection and gene flow barriers across the genome. Thus,
we see no obvious qualitative genetic difference between
the apple and hawthorn host races of R. pomonella,
R. mendax, R. zephyria or the flowering dogwood fly in
the current sequence-based analysis or past allozyme
studies. Eventually, some of these species may continue
to be channelled by divergent ecological selection alone
or in combination with processes acting during periods of
geographic isolation to consolidate into the types of
discreet operational taxonomic units (OTUs) favoured by
systematists for classification. Such a scenario may help
explain the monophyly of R. cornivora and the EVTM
population of hawthorn flies. However, care must be
taken in interpreting phylogenies of such OTUs, as during
the formative period of population divergence, there may
be no one single bifurcating population history encapsu-
lating the genetic and biogeographic ⁄ ecological processes
that acted across the genome to generate these ‘species’.
910 X. XIE ET AL.
ª 2 0 0 8 T H E A U T H O R S . J . E V O L . B I O L . 2 1 ( 2 0 0 8 ) 9 0 0 – 9 1 3
J O U R N A L C O M P I L A T I O N ª 2 0 0 8 E U R O P E A N S O C I E T Y F O R E V O L U T I O N A R Y B I O L O G Y

By contrast, other taxa may persist for extended periods
of time in a dynamic equilibrium of semi-permanent
divergence dictated by the balance between disruptive
selection and introgression, whereas still others may fuse
if environmental ⁄ ecological circumstances were to
change. This view of life and the genesis of biodiversity
is by no means unique to Rhagoletis (e.g. Wang et al.,
1997; Rieseberg et al., 1999; Noor et al., 2001b; Beltran
et al., 2002; Machado & Hey, 2003; Stump et al., 2005;
Turner et al., 2005; Mallet et al., 2007). It will be
interesting to see whether and how this view changes
as the entire genome sequences of more and more of
life’s diversity become known.
Acknowledgments
The authors thank the following individuals for their
assistance: S. Berlocher, G. Bush, B. Matta, B. McPheron,
J.J Smith, F. Wang, F. Wang Jr, J. Wise and the Trevor
Nichols Research Station of Michigan State University at
Fennville, MI. This research was supported, in part, by
grants from the National Science Foundation, a National
Research Initiative grant from the United States Depart-
ment of Agriculture, and the 21st Century Fund of the
state of Indiana to JLF.
References
Beltran, M., Jiggins, C.D., Bull, V., Linares, M., Mallet, J.,
McMillan, W.O. & Bermingham, E. 2002. Phylogenetic
discordance at the species boundary: comparative gene gene-
alogies among rapidly radiating Heliconius butterflies. Mol. Biol.
Evol. 19: 2176–2190.
Berlocher, S.H. 2000. Radiation and divergence in the Rhagoletis
pomonella species group: inferences from allozymes. Evolution
54: 543–557.
Berlocher, S.H. & Feder, J.L. 2002. Sympatric speciation in
phytophagous insects: moving beyond controversy? Annu.
Rev. Entomol. 47: 773–815.
Berlocher, S.H., McPheron, B.A., Feder, J.L. & Bush, G.L. 1993. A
revised phylogeny of the Rhagoletis pomonella (Diptera: Tephri-
tidae) sibling species group. Ann. Entomol. Soc. Am. 86: 716–727.
Bierbaum, T.J. & Bush, G.L. 1990. Genetic differentiation in the
viability of sibling species of Rhagoletis fruit flies on host plants,
and the influence of reduced hybrid viability on reproductive
isolation. Entomol. Exp. Appl., 55: 105–118.
Bush, G.L. 1966. The taxonomy, cytology, and evolution of the
genus Rhagoletis in North America (Diptera: Tephritidae). Bull.
Mus. Comp. Zool. 134: 431–562.
Bush, G.L. 1969. Sympatric host race formation and speciation
in frugivorous flies of the genus Rhagoletis (Diptera: Tephriti-
dae). Evolution 23: 237–251.
Coyne, J.A. & Orr, H.A. 2004. Speciation. Sinauer Associates,
Sunderland, MA, 545 pp.
Cracraft, J. 1989. Speciation and its ontology: the empirical
consequences of alternative species concepts for understand-
ing patterns and processes of differentiation. In: Speciation and
its Consequences (D. Otte & J. A. Endler, eds), pp. 28–59.
Sinauer Associates Inc., Sunderland, MA.
Dambroski, H.R. & Feder, J.L. 2007. Host plant and latitude-
related diapause variation in Rhagoletis pomonella: a test for
multifaceted life history adaptation on different stages of
diapause development. J. Evol. Biol. 20: 2101–2112.
Dambroski, H.R., Linn, C. Jr, Berlocher, S.H., Forbes, A.A.,
Roelofs, W. & Feder, J.L.. 2005. The genetic basis for fruit odor
discrimination in Rhagoletis flies and its significance for
sympatric host shifts. Evolution 59: 1953–1964.
Feder, J.L. 1998. The apple maggot fly, Rhagoletis pomonella: flies
in the face of conventional wisdom about speciation? In:
Endless Forms: Species and Speciation (D. Howard & S. H.
Berlocher, eds), pp. 130–144. Oxford University Press, Oxford.
Feder, J.L. & Bush, G.L. 1989. A field test of differential host
plant usage between two sibling species Rhagoletis pomonella
fruit flies (Diptera: Tephritidae) and its consequences for
sympatric models of speciation. Evolution 43: 1813–1819.
Feder, J.L., Hunt, T.A. & Bush, G.L. 1993. The effects of climate,
host plant phenology and host fidelity on the genetics of apple
and hawthorn infesting races of Rhagoletis pomonella. Entomol.
Exp. Appl. 69: 117–135.
Feder, J.L., Opp, S., Wlazlo, B., Reynolds, K., Go, W. & Spisak, S.
1994. Host fidelity is an effective pre-mating barrier between
sympatric races of the Apple Maggot Fly. Proc. Natl Acad. Sci.
USA 91: 7990–7994.
Feder, J.L., Roethele, J.B., Wlazlo, B. & Berlocher, S.H. 1997a.
Selective maintenance of allozyme differences between sym-
patric host races of the apple maggot fly. Proc. Natl Acad. Sci.
USA, 94: 11417–11421.
Feder, J.L., Stolz, U., Lewis, K.M., Perry, W., Roethele, J.B. &
Rogers, A. 1997b. The effects of winter length on the genetics
of apple and hawthorn races of Rhagoletis pomonella (Diptera:
Tephritidae). Evolution 51: 1862–1876.
Feder, J.L., Williams, S.M., Berlocher, S.H., McPheron, B.A. &
Bush, G.L. 1999. The population genetics of the apple maggot
fly and the snowberry maggot: implications for models of
sympatric speciation. Entomol. Exp. Appl. 90: 9–24.
Feder, J.L., Berlocher, S.H., Roethele, J.B., Dambroski, H.R.,
Smith, J.J., Perry, W.L., Gavrilovic, V., Filchak, K.E., Rull, J. &
Aluja, M.. 2003a. Allopatric genetic origins for sympatric host-
plant shifts and race formation in Rhagoletis. Proc. Natl Acad. Sci.
USA 100: 10314–10319.
Feder, J.L., Roethele, J.B., Filchak, K.E., Niedbalski, J. &
Romero-Severson, J. 2003b. Evidence for inversion polymor-
phism related to sympatric host race formation in the apple
maggot fly, Rhagoletis pomonella. Genetics 163: 939–953.
Feder, J.L., Xie, X., Rull, J., Velez, S., Forbes, A., Dambroski, H.,
Filchak, K. & Aluja, M. 2005. Mayr, Dobzhansky, Bush and
the complexities of sympatric speciation in Rhagoletis. Proc.
Natl Acad. Sci. 102: 6573–6580.
Felsenstein, J. 1989. PHYLIP – phylogeny inference package.
Cladistics 5: 164–166.
Filchak, K.E., Roethele, J.B. & Feder, J.L. 2000. Natural selection
and sympatric divergence in the apple maggot Rhagoletis
pomonella. Nature 407: 739–742.
Gavrilovic, V., Bush, G.L., Schwarz, D., Crossno, J.E. & Smith,
J.J. 2007. Rhagoletis zephyria (Diptera: Tephritidae) in the Great
Lakes basin: a native insect on a native host? Ann. Entomol. Soc.
Amer. 100: 474–482.
Hey, J. & Nielsen, R. 2007. Integration within the Felsenstein
equation for improved Markov chain Monte Carlo methods in
population genetics. PNAS 104: 2785–2790.
Radiation of Rhagoletis pomonella complex 911
ª 2 0 0 8 T H E A U T H O R S . J . E V O L . B I O L . 2 1 ( 2 0 0 8 ) 9 0 0 – 9 1 3
J O U R N A L C O M P I L A T I O N ª 2 0 0 8 E U R O P E A N S O C I E T Y F O R E V O L U T I O N A R Y B I O L O G Y

Hudson, R.D. & Coyne, J.A. 2002. Mathematical conse-
quences of the genealogical species concept. Evolution 56:
1557–1565.
Hudson, R.R. & Kaplan, N.L. 1985. Statistical properties of the
number of recombination events in the history of a sample of
DNA sequences. Genetics 111: 147–164.
Kirkpatrick, M. & Barton, N. 2006. Chromosomal inversions,
local adaptation, and speciation. Genetics 173: 419–434.
Kumar, S., Tamura, K. & Nei, M. 2004. MEGA3: integrated
software for molecular evolutionary genetics analysis and
sequence alignment. Brief. Bioinform. 5: 150–163.
Linn, C., Feder, J.L., Nojima, S., Dambroski, H.R., Berlocher,
S.H. & Roelofs, W. 2003. Fruit odor discrimination and
sympatric host race formation in Rhagoletis. Proc. Natl Acad.
Sci. USA 100: 11490–11493.
Linn, C.E., Dambroski, H.R., Feder, J.L., Berlocher, S.H., Nojima,
S. & Roelofs, W.L. 2004. Postzygotic isolating factor in
sympatric speciation in Rhagoletis flies: reduced response of
hybrids to parental host-fruit odors. Proc. Natl Acad. Sci. USA
101: 17753–17758.
Machado, C.A. & Hey, J. 2003. The causes of phylogenetic
conflict in a classic Drosophila species group. Proc. R. Soc. Lond.
B 270: 1193–1202.
Mallet, J. 1995. A species definition for the modern synthesis.
Trends Ecol. Evol. 10: 294–299.
Mallet, J. 2005. Speciation in the 21st Century. Review of
‘‘Speciation’’, by Jerry A. Coyne and H. Allen Orr. Heredity 95:
105–109.
Mallet, J., Beltran, M., Neukirchen, W. & Linares, M. 2007.
Natural hybridization in heliconiine butterflies: the species
boundary as a continuum. BMC Evol. Biol. 7: 28.
Mayr, E. 1963. Animal Species and Evolution. Harvard University
Press, Cambridge, MA, 797 pp.
Mayr, E. 2001. Wu’s genic view of species. J. Evol. Biol. 14: 866–
867.
McPheron, B.A., Smith, D.C. & Berlocher, S.H. 1988. Genetic
differences between host races of Rhagoletis pomonella. Nature
336: 64–66.
Michel, A.P., Rull, J., Aluja, M. & Feder, J.L. 2007. The genetic
structure of hawthorn-infesting Rhagoletis pomonella popula-
tions in Mexico: implications for sympatric host race forma-
tion. Mol. Ecol. 16: 2867–2878.
Navarro, A. & Barton, N.H. 2003. Chromosomal speciation and
molecular divergence-accelerated evolution in rearranged
chromosomes. Science 300: 321–324.
Nielsen, R. & Wakeley, J. 2001. Distinguishing migration from
isolation. A Markov chain Monte Carlo approach. Genetics 158:
885–896.
Noor, M.A.F., Grams, K.L., Bertucci, L.A. & Reiland, J. 2001a.
Chromosomal inversions and the reproductive isolation of
species. Proc. Natl Acad. Sci. USA 98: 12084–12088.
Noor, M.A.F., Grams, K.L., Bertucci, L.A., Almendarez, Y.,
Reiland, J. & Smith, K.R. 2001b. The genetics of reproductive
isolation and the potential for gene exchange between
Drosophila pseudoobscura and D. persimilis via backcross hybrid
males. Evolution 55: 512–521.
Posada, D. & Crandall, K.A. 1998. MODELTEST: testing the
model of DNA substitution. Bioinformatics 14: 817–818.
Prokopy, R.J., Bennett, E.W. & Bush, G.L. 1971. Mating
behavior in Rhagoletis pomonella (Diptera: Tephritidae). I. Site
of assembly. Canadian Entomologist 103: 1405–1409.
Prokopy, R.J., Bennett, E.W. & Bush, G.L. 1972. Mating
behavior in Rhagoletis pomonella. II. Temporal organization.
Canadian Entomologist 104: 97–104.
de Queiroz, K. 1998. The general lineage concept of species.
Species criteria, and the process of speciation: a conceptual
unification and terminological recommendations. In: Endless
Forms: Species and Speciation (D. Howard & S. H. Berlocher,
eds), pp. 57–78. Oxford University Press, Oxford.
Reissig, W.H. & Smith, D.C. 1978. Bionomics of Rhagoletis
pornonella in Crataegus. Ann. Entomol. Soc. Am. 71: 155–159.
Rieseberg, L.H. 2001. Chromosomal rearrangements and speci-
ation. Trends Ecol. Evol. 16: 351–358.
Rieseberg, L.H., Whitton, J. & Gardner, K. 1999. Hybrid zones
and the genetic architecture of a barrier to gene flow between
two sunflower species. Genetics 152: 713–727.
Roethele, J.B., Romero-Severson, J. & Feder, J.L. 2001. Evi-
dence for broad-scale conservation of linkage map relation-
ships between Rhagoletis pomonella and Drosophila melanogaster.
Ann. Entomol. Soc. Am. 94: 936–947.
Rull, J., Aluja, M., Feder, J.L. & Berlocher, S.H. 2006. The
distribution and host range of hawthorn-infesting Rhagoletis
pomonella (Diptera: Tephritidae) in Mexico. Ann. Entomol. Soc.
Am. 99: 662–672.
Saitou, N. & Nei, M. 1987. The neighbor-joining method: a new
method for reconstructing phylogenetic trees. Mol. Biol. Evol.
4: 406–425.
Schneider, S., Roessli, D. & Excoffier, L. 2000. Arlequin ver. 2.000:
A Software for Population Genetics Data Analysis.
Schwarz, D. & McPheron, B.A. 2007. When ecological isolation
breaks down: sexual isolation is an incomplete barrier to
hybridization between Rhagoletis species. Evol. Ecol. Res. 9:
829–841.
Schwarz, D., Matta, B.M., Shakir-Botteri, N.L. & McPheron, B.A.
2005. Host shift to an invasive plant triggers rapid animal
hybrid speciation. Nature 436: 546–549.
Schwarz, D., Shoemaker, K.D., Shakir, N.L. & McPheron, B.A.
2007. A novel preference for an invasive plant as a mechanism
for animal hybrid speciation. Evolution 61: 245–256.
Smith, D.C. 1986. Genetics and Reproductive Isolation of Rhagoletis
flies. PhD dissertation. University of Illinois at Urbana, 97 pp.
Smith, D.C. 1988. Heritable divergence of Rhagoletis pomonella
host races by seasonal asynchrony. Nature 336: 66–68.
Stump, A.D., Fitzpatrick, M.C., Lobo, N.F., Traore, S., Sagnon,
N.F., Costantini, C., Collins, F.H. & Besansky, N.J. 2005.
Centromere-proximal differentiation and speciation in Anoph-
eles gambiae. Proc. Natl Acad. Sci. 102: 15930–15935.
Swofford, D.L. 2002. PAUP*: Phylogenetic Analysis Using Parsimony
(*and Other Methods), Version 4.0b10. Sinauer Associates,
Sunderland, MA.
Tamura, K. & Nei, M. 1993. Model selection in the estimation of
the number of nucleotide substitutions. Mol. Biol. Evol. 10:
512–526.
Turner, T.L., Hahn, M.W. & Nuzhdin, S.V. 2005. Genomic islands
of speciation in Anopheles gambiae. PLoS Biol. 3: 1572–1578.
Wang, R.L., Wakeley, J. & Hey, J. 1997. Gene flow and natural
selection in the origin of Drosophila pseudoobscura and close
relatives. Genetics 147: 1091–1106.
Wu, C.-I. 2001. The genic view of the process of speciation.
J. Evol. Biol. 14: 851–865.
Xie, X., Rull, J., Michel, A.P., Velez, S., Forbes, A.A., Lobo, N.F.,
Aluja, M. & Feder, J.L. 2007. Hawthorn-infesting populations
912 X. XIE ET AL.
ª 2 0 0 8 T H E A U T H O R S . J . E V O L . B I O L . 2 1 ( 2 0 0 8 ) 9 0 0 – 9 1 3
J O U R N A L C O M P I L A T I O N ª 2 0 0 8 E U R O P E A N S O C I E T Y F O R E V O L U T I O N A R Y B I O L O G Y

of Rhagoletis pomonella in Mexico and speciation mode plural-
ity. Evolution 61: 1091–1105.
Supplementary material
The following supplementary material is available for this
article:
Table S1 Collecting sites for flies genetically analysed in
the study.
Figure S1 Parsimony gene tree for locus P181.
Figure S2 Parsimony gene tree for locus P667.
Figure S3 Parsimony gene tree for locus P7.
Figure S4 Parsimony gene tree for locus P661.
This material is available as part of the online article
from: http://www.blackwell-synergy.com/doi/abs/10.1111/
j.1420-9101.2008.01507.x.
Please note: Blackwell Publishing are not responsible
for the content or functionality of any supplementary
materials supplied by the authors. Any queries (other
than missing material) should be directed to the corres-
ponding author for the article.
Received 1 November 2007; revised 19 December 2007; accepted 19
December 2007
Radiation of Rhagoletis pomonella complex 913
ª 2 0 0 8 T H E A U T H O R S . J . E V O L . B I O L . 2 1 ( 2 0 0 8 ) 9 0 0 – 9 1 3
J O U R N A L C O M P I L A T I O N ª 2 0 0 8 E U R O P E A N S O C I E T Y F O R E V O L U T I O N A R Y B I O L O G Y
Related Documents











