ARTICLE Purification and Characterization of a 28 kDa Cytosolic Inhibitor of Cholesteryl Ester Hydrolases in Rat Testis Denise S. Hines, Siowfong Wee 1, and W. McLean Grogan* Department of Biochemistry and Molecular Biophysics, Virginia Commonwealth University, Richmond, Virginia 23298-0614 I~STRACT: A 28 kDa inhibitory protein was purified from rat lestis cytosol by sequential 40-65% ammonium sulfate precipi- ~tion, cation exchange chromatography, anion exchange chro- matography, and preparative SDS-polyacrylamide gel elec- ~ophoresis. The heat-stable, trypsin-labile protein exhibited ~nenzymatic, concentration-dependent inhibition of testicular and pancreatic cholesteryl ester hydrolases at all stages of pu- blication. Copurifying at each stage was a 26.5 kDa protein which comprised 25% of the mass of the two proteins. Poly- donal antibodies raised to either or both 28 kDa and 26.5 kDa proteins by direct injection of excised electrophoretic bands cross-reacted with both proteins on western blots, immunopre- cipitated both proteins, and neutralized inhibitory activity. Amino acid compositions of the individual proteins electroe- luted from SDS-polyacrylamide gels were different from those of other surface-active proteins of similar molecular weights. 8oth proteins exhibited identical pl of 4.8 on chromatofocusing columns and two-dimensional gel electrophoresis. Although the subcellular distribution of the 28 kDa protein is unknown, its testicular cytosolic concentration, calculated from the purified protein mass, was 8 x 10-9 mols/L, which probably underesti- mates the actual concentration by an order of magnitude. This is greater than the minimum concentration required for in vitro inhibition (10-9 mols/L), consistent with a physiological role for this protein. Lipids 31, 1233-1243 (1996). Catalytic activities of surface-active enzymes are known to be influenced by the presence of other proteins associated with the substrate surface. Well-characterized, physiologi- cally relevant examples of such interactions include the acti- vation of pancreatic lipase by colipase (1,2); activation of lipoprotein lipase by apolipoprotein C-II (3,4); activation of lecithin/cholesterol-acyltransferase by apolipoprotein A-I (5-7), and activation of bee venom phospholipase A 2 by mel- litin (8). Other proteins are known to influence activity of hy- drolytic enzymes by modifying the surface of lipophilic sub- tCurrent address:Immunex Corp.,51 University St., Seattle,WA 98101. *Towhom correspondence shouldbe addressed. Abbreviations: ASP,ammonium sulfateprecipitate;CEH,cholesteryl ester hydrolase; CIP-28,28 kDaCEH inhibitory protein;HPLC, high-performance liquid chromatgraphy; I50, amount of proteininhibiting 50% under assay con- ditions, evaluated froma proteinconcentration curve;PLAP, phospholipase- activating protein;PNP,p-nitrophenol;SDS-PAGE,SDS-polyacrylamide gel electrophoresis. strates (9-11). The pathophysiological significance of such proteins is manifest in the cytolytic potential of intracellular hydrolytic enzymes, which are generally characterized by broad substrate specificities, and in the general lack of under- standing of mechanisms for regulating their activity in vivo. It has been suggested that accessory proteins may be involved in regulation of cholesteryl ester hydrolase (CEH) in various organs (12-18). However, there has been no report of the pu- rification or specific characterization of such a putative regu- latory protein. Several investigators in this laboratory have noted unex- pectedly high recoveries of catalytic activity or loss of activ- ity at specific stages of purification of CEH isoenzymes from rat testis (19) or liver (20). These findings were suggestive of differential loss of inhibitory or stimulatory accessory factors during the course of purification. In this report, we describe the purification and characterization of a specific testicular cytosolic protein with properties consistent with a physiolog- ical role in regulation of CEH activity. MATERIALS AND METHODS Chemicals and supplies. Cholesterol [ 1-14C]oleate (59.4 mCi/mmol) and Aquasol-2 were purchased from DuPont- New England Nuclear (Wilmington, DE); heptane [high-per- formance liquid chromatography (HPLC) grade], from Bax- ter (Columbia, MD); dialysis tubing, Gelman acrodiscs (0,22 IJM) and other solvents, from Fisher Scientific (Norcross, GA); Immobilon-P Transfer Membranes (PVDF), from Mil- lipore (Bedford, MA); SDS, acrylamide, bis-acrylamide, TEMED, ammonium persulfate, silver stain kit, elec- trophoretic molecular weight markers, gelatin, blotting grade GAR-HRP and blotting grade DAB, from BioRad (Melville, NY); BCA protein assay kit, from Pierce Chemical Co. (Rockford, IL); ultrapure (NH4)2SO 4 and Coomassie R-250, from ICN (Irvine, CA); porcine pancreatic CEH, trypsin- linked agarose beads and other chemicals and reagents, from Sigma (St. Louis, MO); electrophoretic equipment, from Hoeffer (San Francisco, CA); frozen rat testes (250-350 g Sprague-Dawley rats), from Zivic-Miller Laboratories (Zelienople, PA) and Hilltop Laboratories (Scottsdale, PA); FPLC Mono S (HR 5/5 and HR 10/10) cation exchange col- umn, FPLC Mono Q (HR 5/5) anion exchange column, FPLC Copyright© 1996 by AOCS Press t 233 Lipids, Vol. 31, no. 12 (1996)

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ARTICLE
Purification and Characterization of a 28 kDa Cytosolic Inhibitor of Cholesteryl Ester Hydrolases in Rat Testis
Denise S. Hines, Siowfong Wee 1, and W. McLean Grogan* Department of Biochemistry and Molecular Biophysics, Virginia Commonwealth University, Richmond, Virginia 23298-0614
I~STRACT: A 28 kDa inhibitory protein was purified from rat lestis cytosol by sequential 40-65% ammonium sulfate precipi- ~tion, cation exchange chromatography, anion exchange chro- matography, and preparative SDS-polyacrylamide gel elec- ~ophoresis. The heat-stable, trypsin-labile protein exhibited ~nenzymatic, concentration-dependent inhibition of testicular and pancreatic cholesteryl ester hydrolases at all stages of pu- blication. Copurifying at each stage was a 26.5 kDa protein which comprised 25% of the mass of the two proteins. Poly- donal antibodies raised to either or both 28 kDa and 26.5 kDa proteins by direct injection of excised electrophoretic bands cross-reacted with both proteins on western blots, immunopre- cipitated both proteins, and neutralized inhibitory activity. Amino acid compositions of the individual proteins electroe- luted from SDS-polyacrylamide gels were different from those of other surface-active proteins of similar molecular weights. 8oth proteins exhibited identical pl of 4.8 on chromatofocusing columns and two-dimensional gel electrophoresis. Although the subcellular distribution of the 28 kDa protein is unknown, its testicular cytosolic concentration, calculated from the purified protein mass, was 8 x 10 -9 mols/L, which probably underesti- mates the actual concentration by an order of magnitude. This is greater than the minimum concentration required for in vitro inhibition (10 -9 mols/L), consistent with a physiological role for this protein. Lipids 31, 1233-1243 (1996).
Catalytic activities of surface-active enzymes are known to be influenced by the presence of other proteins associated with the substrate surface. Well-characterized, physiologi- cally relevant examples of such interactions include the acti- vation of pancreatic lipase by colipase (1,2); activation of lipoprotein lipase by apolipoprotein C-II (3,4); activation of lecithin/cholesterol-acyltransferase by apolipoprotein A-I (5-7), and activation of bee venom phospholipase A 2 by mel- litin (8). Other proteins are known to influence activity of hy- drolytic enzymes by modifying the surface of lipophilic sub-
tCurrent address: Immunex Corp., 51 University St., Seattle, WA 98101. *To whom correspondence should be addressed. Abbreviations: ASP, ammonium sulfate precipitate; CEH, cholesteryl ester hydrolase; CIP-28, 28 kDa CEH inhibitory protein; HPLC, high-performance liquid chromatgraphy; I50, amount of protein inhibiting 50% under assay con- ditions, evaluated from a protein concentration curve; PLAP, phospholipase- activating protein; PNP, p-nitrophenol; SDS-PAGE, SDS-polyacrylamide gel electrophoresis.
strates (9-11). The pathophysiological significance of such proteins is manifest in the cytolytic potential of intracellular hydrolytic enzymes, which are generally characterized by broad substrate specificities, and in the general lack of under- standing of mechanisms for regulating their activity in vivo. It has been suggested that accessory proteins may be involved in regulation of cholesteryl ester hydrolase (CEH) in various organs (12-18). However, there has been no report of the pu- rification or specific characterization of such a putative regu- latory protein.
Several investigators in this laboratory have noted unex- pectedly high recoveries of catalytic activity or loss of activ- ity at specific stages of purification of CEH isoenzymes from rat testis (19) or liver (20). These findings were suggestive of differential loss of inhibitory or stimulatory accessory factors during the course of purification. In this report, we describe the purification and characterization of a specific testicular cytosolic protein with properties consistent with a physiolog- ical role in regulation of CEH activity.
MATERIALS AND METHODS
Chemicals and supplies. Cholesterol [ 1-14C]oleate (59.4 mCi/mmol) and Aquasol-2 were purchased from DuPont- New England Nuclear (Wilmington, DE); heptane [high-per- formance liquid chromatography (HPLC) grade], from Bax- ter (Columbia, MD); dialysis tubing, Gelman acrodiscs (0,22 IJM) and other solvents, from Fisher Scientific (Norcross, GA); Immobilon-P Transfer Membranes (PVDF), from Mil- lipore (Bedford, MA); SDS, acrylamide, bis-acrylamide, TEMED, ammonium persulfate, silver stain kit, elec- trophoretic molecular weight markers, gelatin, blotting grade GAR-HRP and blotting grade DAB, from BioRad (Melville, NY); BCA protein assay kit, from Pierce Chemical Co. (Rockford, IL); ultrapure (NH4)2SO 4 and Coomassie R-250, from ICN (Irvine, CA); porcine pancreatic CEH, trypsin- linked agarose beads and other chemicals and reagents, from Sigma (St. Louis, MO); electrophoretic equipment, from Hoeffer (San Francisco, CA); frozen rat testes (250-350 g Sprague-Dawley rats), from Zivic-Miller Laboratories (Zelienople, PA) and Hilltop Laboratories (Scottsdale, PA); FPLC Mono S (HR 5/5 and HR 10/10) cation exchange col- umn, FPLC Mono Q (HR 5/5) anion exchange column, FPLC
Copyright © 1996 by AOCS Press t 233 Lipids, Vol. 31, no. 12 (1996)

1234 D.S. HINES ETAL.
Superose 6 gel filtration column, FPLC Mono P chromatofo- cusing column (HR 5/20) and FPLC supplies, from Pharma- cia (Piscataway, NJ); Bio-Gel TSK Phenyl-5PW HPLC col- umn, from BioRad (Melville, NY).
Purification of CEH. Rat testicular cytosolic CEH was pre- pared according to Wee and Grogan (19). Testes were homog- enized in buffer (! :2 wt/vol) containing: 20 mM Tris-HC1, pH 7.5; 250 mM sucrose; 80 mM KCI; 0.05% sodium cholate; 10 mM EDTA and 0.1 mM thioglycolate. Cytosol (S 104) was prepared by differential centrifugation at 2000 x g (30 min), 10,000 × g (30 min), and 104,000 × g (90 min); brought to 40% saturation with (NH4)2SO4; stirred for 30 min at 4°C and centrifuged at 10,000 x g for 30 min. Resulting ammonium sulfate precipitate (ASP) pellets were stored a t -20°C until use. ASP was redissolved in 500 laL buffer (20 mM Tris-HCl, pH 7.5; 10 mM EDTA, 0.1 mM thioglycolate, 1 mM sodium cholate), desalted over Sephadex G-25 and used as a CEH source for inhibitor assays or further purified by cation ex- change chromatography.
For further purification, the desalted sample (10-30 mg) was chromatographed on a Pharmacia Mono S HR 10/10 FPLC cation exchange column (4 mL/min), eluting with 40 mL of Buffer A (20 mM Tris-HCl, pH 7.5; 0.05% sodium cholate; 0.1 mM EDTA; 0.1 mM thioglycolate); 72 mL linear gradient, 0-14% Buffer B (Buffer A + 2 M NaC1); 24 mL 14% Buffer B; 1 mL linear gradient 14-100% Buffer B; 33 mL 100% Buffer B. Fractions (8 mL) were assayed for CEH activity. Fractions 13-15 were pooled and used as a CEH source for assays. EDTA (10 mM) was added to stabilize ac- tivity. Protein was estimated by the Pierce Bicinchoninic Acid method (21).
Purification of cytosolic CEH inhibitor. Supernatant from 0-40% ASP was adjusted to 65% saturation with (NH4)2SO 4 and stirred for 30 min at 4°C. ASP (40-65%) was collected by centrifugation at 10,000 x g for 30 min and stored at -20°C until needed. Each ASP pellet, corresponding to ten testes, was dissolved in 1 mL 20 mM Tris-HC1 (pH 7.5, 0. t mM thioglycotate, 10 mM EDTA), desalted on a preequili- brated Sephadex G-25 column, and used for inhibitor assays or further purified.
For further purification, desalted 40-65% ASP (50-70 mg protein) was centrifuged at 10,000 × g for 15 min at 4°C. Su- pernatant was chromatographed on a Pharmacia Mono S HR 5/5 FPLC cation exchange column, eluted at 1 mL/min with 5 mL buffer C (20 mM Tris-HCl, pH 7.5, 0.1 mM EDTA and 0. ! mM thioglycolate); 3 mL linear gradient, 0-16% buffer D (buffer C + 2 M NaCI); 10 mL 16% buffer D; 3 mL linear gra- dient, 16-40% buffer D; 7 mL 40% buffer D, 2 mL linear gra- dient 40-100% buffer D; 15 mL 100% buffer D. One mL fractions were desatted and assayed for inhibitory activity. Fractions 21-24 were pooled, desalted (Centriprep-10 or Centricon- 10 concentrators; Amicon Corp., Danvers, MA) and used as a source of cation exchange purified inhibitor or for further purification. For anion exchange chromatography, cation exchange purified fractions 21-24 (50-300 lag protein) were chromatographed on a Pharmacia Mono Q HR 5/5 anion
exchange column (1 mL/min), eluted with 5 mL buffer (2; 3 mL linear gradient, 0-16% buffer D; 10 mL 16% buffer D; 3 mL linear gradient, 16--40% buffer D; 7 mL 40% buffer D; 2 mL linear gradient, 40-100% buffer D; 15 mL buffer 13. One mL fractions were collected, desalted, and assayed for inhibitory activity.
Active fractions were concentrated by TCA precipitation or Centricon-10 microconcentrator, electrophoresed on 5-20% or 9-12% SDS-polyacrylamide gradient gels (22), and silver stained to assess purity, using BioRad low molecu- lar weight standards for calibration.
CEH activity. Rat testicular neutral cytosolic CEH activity was assayed by the modified method of Wee and Grogan (19). Each assay contained 20 mM Tris-HCl, pH 7.5, 1 mM sodium cholate, 10 mM EDTA, 0.1 mM thioglycolate, and 10--25 IJg 0-40% ASP protein or 5-10 lag cation exchange purified CEH in 500 pL total volume. Tubes were preincubated 5 rain at the assay temperature (32 or 37°C). Cholesterol oleate (35 laM) labeled with 2~4 x 10 4 dpm cholesterol [l-lnc]oleate was added in 12.5 laL acetone, and incubation was continued for 20 min, unless otherwise stated. The reaction was termi- nated by adding 3.25 mL methanol/chloroform/heptane (1.4:1.25:1, by vol) and 50 pL of 1 N NaOH. Product oleate was measured in the aqueous phase by liquid scintillation counting as described by Ghosh and Grogan (23).
Porcine pancreatic CEH (Sigma) was assayed as described above by the modified method of Ghosh and Grogan (20), using 1 pg CEH and 5 mM sodium taurocholate (24). Activ- ity of all enzymes was linear with protein concentration and time of incubation.
Measurement ofinhibitoo' activity. CEH activity was mea- sured with and without 40-65% ASP, cation exchange puri- fied fractions 21-24, anion exchange purified fractions 21-24, or other inhibitor sources. Inhibitor was added just prior to addition of substrate. Inhibitory activity is expressed as %in- hibition = 100 × (activity without inhibitor - activity with in- hibitor)/activity without inhibitor; or units (1 unit = 150, amount of protein inhibiting 50% under assay conditions, evaluated from a protein concentration curve).
Elecovelution. Proteins in inhibitory fractions (1.7-2 mg cation exchange purified protein or 100-150 lag anion ex- change purified protein) were electrophoresed on a 9-12% SDS-polyacrylamide gradient gel. Bands corresponding to Coomassie stained 25-31 kDa markers were cut from the gel and protein electroeluted into 0.04 M Tris-Acetate, 0.002 M EDTA, pH 8.6, using a Shandon (Cheshire, England) elec- troeluter for 3 h at 50 mA. Electrode buffer was 0.4 M Tris- Acetate. Excess SDS was removed by extensive dialysis against 0.04 M Tris-HCI, pH 7.5. Prior to assay, sample was adjusted to pH 7.5 with dilute acetic acid. For individual bands, 28 or 26.5 kDa proteins were prestained with CuCI2 (25) and destained before electroelution (26).
Phosphatase assa~ Hydrolysis ofp-nitrophenylphosphate (20 mg/mL) was measured spectrophotometricaUy at 410 nm (27) using assay conditions described for testicular and pancreatic CEH. The reaction was terminated by addition
Lipids, Vol. 31, no. 12 (1996)

CHOLESTERYL ESTER HYDROLASE INHIBITORY PROTEIN 1235
of 600 laL of 0.5 M phosphate buffer, pH 7.5 and 10 mM EDTA.
Hydrophobic chromatography. Pooled cation exchange fractions 21-24 (800 pg protein) were concentrated by 65% (NH4)2SO 4 precipitation and resuspended in 1 mL 0.1 M sodium phosphate, pH 7.0, 2 M (NH4)2SO 4. The 100 laL was chromatographed on a Biogel TSK-phenyl 5PW HPLC col- umn pre-equilibrated with the same buffer and eluted (1 mL/min) by a 60 min decreasing linear (NH4)2SO 4 gradient (2 to 0 M); followed by 20 min elution with distilled H20.
Production of polyclonal antibody. For primary immuniza- tion, 650 pg cation exchange purified protein (pooled and concentrated fractions 21-28) was electrophoresed on a 9-12% SDS-polyacrylamide gradient gel. The gel was stained briefly with Coomassie R-250, to visualize bands cor- responding to 26.5, 27, and 28 kDa and washed to remove ex- cess stain. Selected bands were excised, homogenized in phosphate buffered saline and Freund's complete adjuvant (1:2:2, gel-saline-adjuvant). An emulsion was generated by repeated passage between two ground-glass barreled syringes and injected intradermally at 10-15 sites on the back of a New Zealand rabbit on day 0. On days 25 and 72, booster im- munizations of 300-500 pg anion exchange purified protein (fractions 21-28) were injected with Freund's incomplete ad- juvant. Ten days after the third injection, serum was tested for antibodies by Western blot analysis as described below. Ad- ditional boosters were given in incomplete adjuvant 7-12 d prior to subsequent bleedings. Individual rabbits were in- jected with all three bands, the 28 kDa band alone, or the 26.5 kDa band alone. Preimmune serum was collected 7 d before primary immunization. IgG was prepared from immune and preimmune serums by PEG precipitation (28).
Western blot analysis. Proteins separated by SDS-PAGE (9-12% gradient) were transferred to Millipore Immobilon- PVDF membrane by semi-dry blot transfer (Integrated Sepa- ration System, Natick, MA) for I h at 2 mA/cm 2, according to the manufacturer's instructions. Western blot analysis was carried out as described previously (29) using 1:300 dilution of IgG (1.3 mg/mL).
Immunoprecipitation. Inhibitory proteins were immuno- precipitated from cation exchange purified inhibitor (fractions 21-28) by the modified procedure of Kessler (30,31) using antibody complexed with purified protein-A conjugated poly- acrylic beads (Sigma). Beads were precoated with gelatin to decrease nonspecific binding to beads, washed to remove ex- cess gelatin, incubated for 1 h with immune IgG or preim- mune IgG, centrifuged to remove excess IgG and incubated for 1 h with inhibitor-containing fractions or buffer. Beads were then centrifuged and the supernatants collected and as- sayed for inhibitory activity against pancreatic CEH.
Amino acid analysis. Cation exchange purified protein (25 lag) was electrophoresed on a 9-12% SDS-polyacry- lamide gradient gel, electroblotted to Millipore PVDF mem- brane (32), and stained with Coomassie blue R-250. Stained proteins corresponding to the two major bands (28 and 26.5 kDa) and the minor 27 kDa were excised. Quantitative amino
acid analysis was carried out by Hewlett-Packard Amino- quant Methodology in the Protein Core Facility at Virginia Commonwealth University. The amino acid compositions for mature rat serum apoA-I and PLAP were translated from the reported cDNA sequences (33,34). The amino acid composi- tion of the 28 kDa estrogen-regulated protein of Leydig cells is published (35,36,37).
Two-dimensional gel electrophoresis. Using the method of O'Farrell (38), the cation exchange purified inhibitor 200-300 pg fractions 21-24) was separated by nonequilib- rium pH gradient gel electrophoresis (5.5 h at 550v) in the first dimension and 10% SDS-PAGE in the second dimen- sion. Gels were then silver stained or transferred to PVDF membrane for Western blot analysis. Protein molecular weight and pI were compared to the migration of two-dimen- sional electrophoresis markers (Sigma) (pI range 7.6-3.8; MW range 17,000-89,000).
Chromatofocusing. The 200-300 lag cation exchange frac- tions 21-24 were chromatographed on a Pharmacia FPLC Mono P HR 5/20 column eluted (1 mL/min) with a pH 6-4 linear gradient, consisting of 3 mL 0.025 M bis-Tris-HCl, pH 6.3, 0.1 mM EDTA, 0.1 mM thioglycolate, followed by 39 mL 1:10 Pharmacia Polybuffer 74-HC1, pH 4.0, 0.1 mM EDTA, 0.1 mM thioglycolate. Inhibitory activity was assayed in 1 mL fractions.
Gel permeation chromatography. Cation exchange puri- fied protein (200 lag) was chromatographed on an FPLC Su- perose 6 gel permeation column eluted (0.2 mL/min) with 2 0 mM Tris-HCt, pH 7.5, 0.1 mM thioglycotate, 10 mM EDTA, 1 mM sodium cholate, and 150 mM NaCI. Inhibitory activity was assayed in 0.5 mL fractions.
RESULTS
Testis contains a temperature-labile CEH, which is active at 32°C and inactive at 37°C, and a temperature stable enzyme, which is active at both temperatures (39,40). Both activities precipitate from rat testis cytosol in 40% ammonium sulfate (41). As seen in Figure 1 and Table 1, the 40-65% ASP in- hibits both forms of testicular CEH. Although the I5o for inhi- bition of the temperature-stable component (72 lag in Fig. IA), which persists at 37°C, is lower than that for inhibition at 32°C (580 lag in Fig. IA), this difference may not reflect differential sensitivities of the enzymes to inhibition (see the Discussion section). The inhibition is concentration-depen- dent, saturable (Fig. 1A), and reversible by dilution (data not shown), suggesting a reversible binding phenomenon. The principal inhibitory activity is trypsin-labile (Fig. 2B) and re- mined by dialysis membrane (Fig. 2C), indicative of a protein with molecular weight > 12,000. However, the inhibition is heat-stable (Fig. 2A), independent of incubation time (Fig. 3), and independent of preincubation time with CEH (data not shown), indicating a nonenzymatic mechanism. Moreover, porcine pancreatic CEH was also inhibited by this factor (Fig. 1B), suggesting a general mechanism for inhibition. Pancre- atic CEH provided a more homogeneous and stable enzyme
Lipids, Vol. 31, no. 12 (1996)

t236 D.S. HINES ETAL.
; : 80 "
60
~ o -
-20 ~
-40 0 . 0 0.2 0.4 0.6 0.8 1.0 1.2
mg 40-65% ASP
o
o ¢ - ¢
t ~ l t
100
80
60
40
20
0
-20 0
B ~---------~
- - A
32°C
[] 37°C
1 t I ~ I l I r I ! 1 ~ t t I0 20 30 40 50 60 70 80
lag 40-65% ASP
FIG. 1. Concentration-dependent inhibition of rat testicular cholesteryl ester hydrolase (CEH) (A) and pancreatic CEH (B) at 32°C (,~) and 37°C (U]) by the 40-65% ammonium sulfate precipikate from rat testis cytosol. Values are mean percentage inhibition of CEll control _+ SEM (n = 3). Error bars not seen are contained within symbols.
model for mechanistic studies than the testicular CEH. The 150 for pancreatic CEH (10 pg in Fig. 1B) was somewhat lower than that for the temperature-stable testicular CEH and
did not differ at 32 and 37°C, indicating that the inhibition~i not intrinsically temperature-dependent.
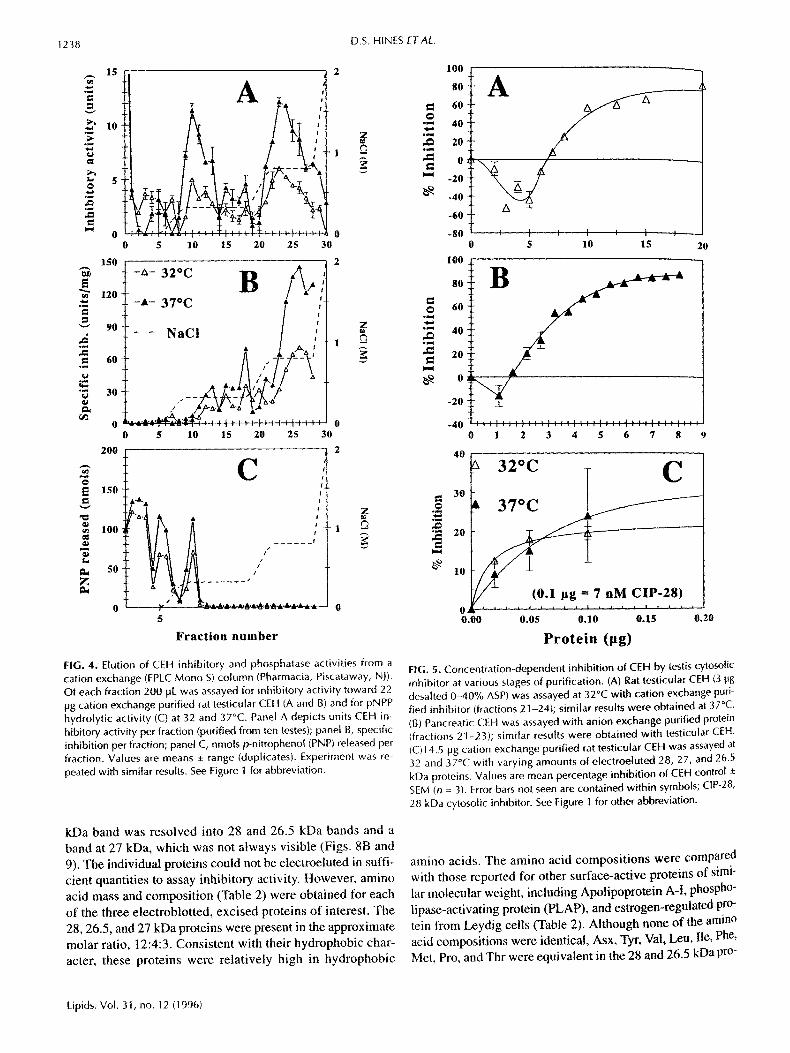
The 40-65% ASP was further fractionated by cation ex- change chromatography, yielding two major peaks of in~ hibitory activity in fractions 9-12 and fractions 21-28, re, spectivety (Fig. 4A). Fractions 21-28 not only contained most of the inhibitory activity but were also much higher in spe- cific inhibitory activity than any other fractions (Fig. 4B). AS with 40-65% ASP (cf. Fig. 1A), cation exchange purified in- hibitor was somewhat less effective in inhibiting testicular CEH at 32°C than at 37°C (Table !). Although inhibition showed similar concentration dependency to that seen with 40-65% ASP (Fig. 5A), specific inhibitory activity was sub- stantially increased ( I50 = I0 pg in Fig. 5A; cf. 5.4 lag for 1 pg pancreatic CEH, not shown). Negative inhibition at very low inhibitor concentrations reflects activation, which per- sisted at every stage of purification, although it was not al- ways apparent in the range of concentrations depicted.
Inasmuch as the testicular CEH is inhibited by phos- phatase (41,42), general phosphatase activity was measured in cation exchange fractions (Fig. 4C). Coelution of the first peak of inhibitory activity with a major peak of phosphatase activity suggested that the inhibitory activity of interest was contained in the second, larger peak, which had no apprecia- ble phosphatase activity. Thus, fractions 21-28 with 41-56% of the ASP inhibitory activity (Table 1) were used in subse- quent purification steps.
Cation exchange fractions 21-28 were pooled and further fractionated by anion exchange chromatography. As seen in Figure 6, most of the inhibitory activity eluted in fractions 21-24. Although activity was assayed with the pancreatic CEH in the typical elution profile shown i n Figure 6, a simi- lar pattern was seen with testis CEH (data not shown). More- over, specific inhibitory activities did not differ significantly between pancreatic (typically, 900 units/mg) and testicular CEH (Table 1) or between temperature-stable and tempera- ture-labile CEH (Table 1) at this stage of purification. Inhibi- tion by pooled anion exchange fractions (Fig. 5B) was also
TABLE 1 Purification of 28 kDa CEH Inhibitory Protein (CIP-28) from Rat Testis Cytosol (S-104) a
32 °C 37°C
Total Specific Fractional Total Specific Fractional Purification recovery activity recovery recovery activity recovery step (units) (units/rag) (%) Fold (units) (units/rag) (%) Fold
40-65% (NH4)2SO 4 precipitate (ASP) 56 0.75 100 1 124 16 [ 00 1
Cation exchange FPLC (fractions 21-28) 23 56 41 75 70 215 56 13
Anion exchange FPLC (fractions 21-28) 9 847 18 1129 9 821 8 51
SDS-PAGE of pooled cation exchange fractions (electroelution) 6 1898 11 2531 7 1866 5 ] 17
alnhibitory activity was measured with partially purified testicular cholesteryl ester hydrolase (CEH), as described in the Materials and Methods section. Inhibition of pancreatic CEH was similar to that of testicular CEH at each stage of purifica- tion. Total recovery is referenced to 10 testes; SDS-PAGE, SDS-polyacrylamide gel electrophoresis.
Lipids, Vol. 31, no. 12 (1996)

CHOLESTERYL ESTER HYDROLASE INHIBITORY PROTEIN 1237
U
'otein filing
Y l control
[ ~ +inhibitor
32°C 37°C
A s s a y t e m p e r a t u r e
S
u
2.0
1.6
1.2-
0.8-
0.4
0.0
2.0
-B
no 3 h 3 h 3 h, 37°C inh ib i tor 4°C 370C tryps in
Inhibitor pretreatment
1.5 O
E
1.0
r~ 0.5
0.0
C
None + ASP + dialyzed ASP
A d d i t i o n s to a s s a y
FIG. 2. Characterization of the inhibitory factor as a heat-stable protein. (A) Cation exchange purified inhibitor was boiled for 10 rain and pre- cipitated protein removed by centrifugation at 10,000 x g for 30 rain (see Fig. I0 for recovery of protein after boiling). Rat testicular CEH was assayed at 32 or 37°C with buffer (control), with inhibitor or with equiv- alent supernatant from boiled inhibitor. (B) Cation exchange purified inhibitor was incubated for 3 h at 4°C or at 37°C in presence or absence of trypsin-linked agarose beads. Samples were then centrifuged at 12,000 x gfor 5 min. Pancreatic CEH was assayed with or without 300 pL of each supernatant. (C) 40-65% ammonium sulfate precipitate (ASP) Was dialyzed (M r cutoff = 12 kDa) against 6 × 2 L assay buffer. Rat tes- [icular CEH was assayed with or without 520 pg ASP, dialyzed or undi- alyzed. Values are mean percentage inhibit ion of CEH control +_ SEM (n= 3). See Figure 1 for other abbreviation.
testis 370C t3 pancreas z~ testis 320C
100 @
4020 A
0 ~ ~ 4 ~ 0 10 20 30 40 50 60
I n c u b a t i o n t i m e ( m i n ) ~.~.
FIG. 3. Time-dependent inhibition of rat testicular CEH or pancreatic CEH by testicular cytosolic CEH inhibitor. Cation exchange purified tes- ticular CEH or pancreatic CEH were assayed with and without cation exchange purified inhibitor (fractions 21 ~24) for the times indicated. Values are mean percentage inhibition of CEH control ± SEM (n = 3). Error bars not seen are contained within symbols. See Figures 1 and 2 for abbreviations.
concentration-dependent and produced a saturation curve similar to that of the 40-65% ASP and cation exchange puri- fied fractions, whereas specific inhibitory activity was further increased (I50 = 3.3 lag in Fig. 5B). As in earlier stages of pu- rification, very low inhibitor concentrations activated CEH.
Homogeneity was assessed by SDS-PAGE at each stage of purification (e.g., Figs. 7-8). However, none of the proce- dures resulted in a completely homogeneous protein. More- over, purification steps beyond cation exchange gave poor re- covery of total activity (see Table 1). Efforts to further purify the inhibitor by gel filtration chromatography produced a dif- fuse pattern of elution, indicating aggregation, even in the presence of bile salts, with no increase in specific activity (not shown). Hydrophobic interaction chromatography of ion-ex- change fractions indicated that the inhibitor was hydrophobic but produced no appreciable increase in homogeneity (not shown).
Although none of the above purification procedures yielded a completely homogeneous protein, silver staining of SDS-polyacrylamide gels revealed a strong band at M r 28 kDa in active fractions at each stage of purification (Figs. 7A and 8A). The electroeluted proteins from this band exhibited concentration-dependent inhibition of testis CEH (Fig. 5C), indicating presence of the inhibitory factor. Although the in- hibition appeared to saturate at lower concentrations with the electroeluted inhibitor (by extrapolation: 40%, at 37°C; 23%, at 32°C) than with less homogenous preparations (typically 80-100%), I20 values were consistent with a high degree of enrichment (74 ng, 2, 8, and 20 lag, for etectroelution, anion exchange, cation exchange, and ASP-purified inhibitor, re- spectively, assayed at 37°C). Moreover, very low concentra- tions of the less highly purified inhibitor also saturated at lower levels of activity (data not shown).
On narrow gradient (9-12%) SDS-PAGE, the broad 28
Lipids, Vol. 31, no. 12 (1996)
1238 D.S. HINES ETAL.
° ~
A=
15 . . . . . . . . . . .
,o A
5 ~ T " ~ T --
0 5 10 15 20 25 30
150 ~ - - g - 32oc 1 1 / \
90 ~__ _ NaCl [ : I • I A. !
60 A ~
, \: .,.--
o ~ . . . . . . " , ~ , ~ ; ~ ' " ' : . . . . ; . . . . I . . . .
0 5 10 15 20 25 30
2 0 0
O 15o
100
1~ 5 0 z
,-%
C k I
. . . . . . I
/
A.~ &±_._±.±.._.~.~.~...*...... 0 5
Fraction number
z
FIG. 4. Elution of CEH inhibitory and phosphatase activities from a cation exchange (FPLC Mono S) column (Pharmacia, Piscataway, NJ). Of each fraction 200 pL was assayed for inhibitory activity toward 22 Hg cation exchange purified rat testicular CEH (A and B) and for pNPP hydrolytic activity (C) at 32 and 37°C. Panel A depicts units CEH in- hibitory activity per fraction (purified from ten testes); panel B, specific inhibition per fraction; panel C, nmols p-nitrophenol (PNP) released per fraction. Values are means :!: range (duplicates). Experiment was re- peated with similar results. See Figure I for abbreviation.
O . ¢ 4
a=
.c:
100
8 0 -
60 40 - ZO -
OZ
- 2 0 -
- 4 0 -
-6o ; - 8 0
0
A
I I I I I t J 5 10 15 20
'° I 40
- 2 0 _L
-4 0 . . . . . : I ~ : , : : : , : : : I : : . . . . . . . . : ' , : : 0 l 2 3 4 5 6 7 8 9
A= .=
4 0
3 0
2 0
10
3 oc [ C
37°C
( ~ 0 , 1 ~ - ? nM CIP-28) O i l I , , , , i , t i t ! t i i
0 . 0 0 0 . 0 5 0 . 1 0 0 . 1 5 0 .20
Prote in (IJg)
FIG. 5. Concentration-dependent inhibition of CEH by testis cytosolic inhibitor at various stages of purification• (A) Rat testicular CEH (3 pg desalted 0-40% ASP) was assayed at 32°C with cation exchange puri- fied inhibitor (fractions 21-24); similar results were obtained at 37°C. (B) Pancreatic CEH was assayed with anion exchange purified protein (fractions 21-23); similar results were obtained with testicular CEH. iC)l 4.5 Hg cation exchange purified rat testicular CEH was assayed at 32 and 37°C with varying amounts of electroeluted 28, 27, and 26.5 kDa proteins. Values are mean percentage inhibition of CEH control -+ SEM (n = 3). Error bars not seen are contained within symbols; CIP-28, 28 kDa cytosolic inhibitor. See Figure 1 for other abbreviation.
kDa band was resolved into 28 and 26.5 kDa bands and a band at 27 kDa, which was not always visible (Figs. 8B and 9). The individual proteins could not be electroeluted in suffi- cient quantities to assay inhibitory activity. However, amino acid mass and composition (Table 2) were obtained for each of the three electroblotted, excised proteins of interest. The 28, 26.5, and 27 kDa proteins were present in the approximate molar ratio, 12:4:3. Consistent with their hydrophobic char- acter, these proteins were relatively high in hydrophobic
amino acids. The amino acid compositions were compared with those reported for other surface-active proteins of simi- lar molecular weight, including Apolipoprotein A-I, phospho- lipase-activating protein (PLAP), and estrogen-regulated pro- tein from Leydig cells (Table 2). Although none of the amino acid compositions were identical, Asx, Tyr, Val, Leu, Ile, Phe, Met, Pro, and Thr were equivalent in the 28 and 26.5 kDa pro-
Lipids, Vol. 31, no. 12 (1996)

CHOLESTERYL ESTER HYDROLASE INHIBITORY PROTEIN 1239
~ 3
2
o
~ 0
t - A - i n h i b i t o r y
_ , N:tCil v i t y
0 5 10 15 20
T
o 25 30
F r a c t i o n n u m b e r
7
28~
B
21 2 2 2 3 2 4 2 5 2 6 2 7 2 8
C a t i o n c o l u m n frac t ion #
C
OG. 6. Elution of cation exchange purified CEH inhibitor from an anion ~change column. Fractions 21-24 of cation exchange purified in- @bitor were refractionated by anion exchange (FPLC Mono Q; Pharma- da). Of each fraction 400 pL was desalted and assayed for inhibitory ~ivity against pancreatic CEH. Activity is reported as total units in- hibitory activity per fraction (purified from 20 testes); values are mean ~0ercentage inhibition of CEH control _+ SEM (n = 3). Error bars not seen .~ contained within symbols. See Figure 1 for other abbreviation.
teins. Attempts to obtain primary sequences for these proteins ~ere unsuccessful. Both proteins appeared to be N-terminally blocked.
Rabbit antibodies were raised by direct injection of the bands from SDS-PAGE, in combination or separately. As seen in Figure 10, strongest antibody titer was achieved with the combined proteins (lane 1), and antibodies raised to the individual 26.5 kDa (lane 2) or 28 kDa (lane 3) proteins ex- hibited mutual cross-immunoreactivity, further evidence that these proteins are closely related. In contrast, immunoreactiv- ity to the 27 kDa band was not apparent with any of the antis- era. The 26.5 and 28 kDa proteins were immunoreactive on ~westem blots (Figs. 7-10 and 12) of inhibitory fractions at all :~tages of purification, and absent from fractions lacking ac- tivity, establishing a persistent correlation between the in- hibitory activity and these proteins. Moreover, the intensity of western blot reactivity was consistent with the amount of hlhibitory activity measured in various fractions (cf. Figs. 4,6 Vs. Figs. 7,8).
To confirm the identity of the inhibitor, the partially puri- fied inhibitor was precipitated with protein A-bead com- plexed antibodies to the 28/26.5 kDa proteins (Fig. 11). tmmunoprecipitation abolished inhibitory activity in a con- eentration-dependent manner, whereas preimmune serum had no effect. SDS-PAGE analysis confirmed precipitation of the 26.5/28 kDa proteins with the antibody-complexed beads and ~emoval of both proteins from the resulting supernatant (not ~hown).
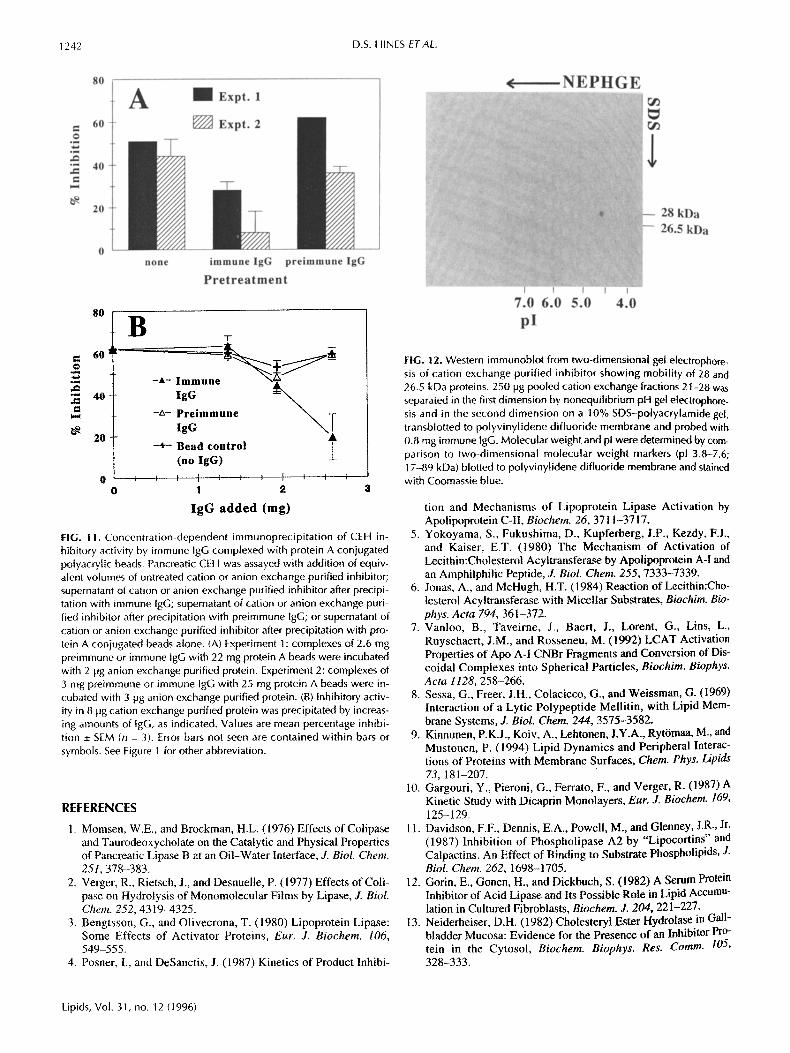
The relationship between the 26.5 and 28 kDa proteins was ~rther examined by two-dimensional (nonequilibrium pH gel plectrophoresis/SDS-PAGE) electrophoresis. As seen in Fig- are 12, the pI of these proteins are identical and no additional proteins are resolved. Both proteins also eluted from an
~" ~ ~ ~ ~ 21 2 3 2 5 ~ ~ ~., ~ ~ ~ ~ ? 2 2 2 4 2 6
P o o l e d f r a c t i o n s ~ Frac t ion #
¢ b
FIG. 7. Analysis of cation exchange fractions by SDS-polyacrylamide gel electrophoresis. Panel A: Silver stained 5-20% SDS-polyacrylamide gradient gel showing prominent 28 kDa band in fractions 21-28, corre- sponding to elution of inhibitory activity from cation exchange column. Positions of BioRad molecular weight markers are provided for compar- ison. Panel B: Western immunoblot of pooled cation exchange frac- tions, electrophoresed on 9-12% SDS-polyacrylamide gradient gel, transb[otted to PVDF membrane and probed with antibody to 28 and 26.5 kDa proteins. Immunoreactive proteins at 26.5 and 28 kDa are re- solved by the shallow gradient and coelute with inhibitory activity (see Fig. 4). Panel C: Western immunoblot of individual cation exchange fractions showing elution of immunoreactive 26.5 and 28 kDa proteins in fractions containing inhibitory activity (see Fig. 4). Other cross-react- ing bands were also immunoreactive with preimmune sera, which did not react with 28 and 26.5 kDa proteins.
HPLC chromatofocusing column, with pl 4.8 (not shown). Consistent with this pI, the inhibitory protein was insoluble in buffers at pH 4.5-5.5 (data not shown).
Association of these proteins with the testis was confirmed by Western blotting after organ perfusion to remove blood. The 26.5/28 kDa immunoreactive proteins were undimin- ished in the perfused testis (data not shown).
DISCUSSION
The major CEH inhibitory activity in the 40--65% ASP of tes- ticular cytosol is associated with unique 28 and 26.5 kDa hy- drophobic proteins which have been purified to apparent ho- mogeneity by sequential cation exchange chromatography, anion exchange chromatography, and SDS-PAGE. Homo-
Lipids, Vol. 31, no. 12 (1996)

1240 D.S. HINES ETAL.
28
C o o m a s s i e W e s t e r n
s t a i n e d b l o t
21 22 23 24 25 26 27 28
Anion column fraction #
B C
22 23 24 ~ ,~ u~ ~ ~ I
Frac t ion# ~ ~ ~ ~ ~ Pooled fractions
FIG. 8. Analysis of anion exchange fractions by SDS-polyacrylamide gel electrophoresis. Panel A: Silver stained 5-20% SDS-polyacry- lamide gradient gel showing prominent 28 kDa band in fractions 22-28, corresponding to elution of inhibitory activity from anion ex- change column. Positions of BioRad molecular weight markers are pro- vided for comparison. Panel B: Silver-stained 9-12% SDS-polyacry- lamide gradient gel showing resolution of 27 and 26.5 kDa proteins from the major 28 kDa band which co-purified with inhibitory activity (see Fig. 6). Panel C: Western immunoblot of pooled anion exchange fractions showing elution of immunoreactive 28 and 26.5 kDa in frac- tions 22-24 and 25-30, which contain inhibitory activity (see Fig. 6).
geneity was confirmed by two-dimensional gel electrophore- sis. Inasmuch as these proteins copurify with virtually all fractionation methods, share the same pI and elicit cross-re- active antibodies, they may share surface domains and both may be inhibitory. Although neither could be purified in suf- ficient quantity to test for individual activity, Coomassie staining, western blotting, and quantitative amino acid analy- sis indicate that the 28 kDa protein (CIP-28) is 3-fold higher in concentration than the 26.5 kDa protein, in inhibitory frac- tions at all levels of purification. Moreover, inhibitory activ- ity is immunoprecipitated by antibodies to the 28 and 26.5 kDa proteins. Thus, it is likely that the inhibitory activity is predominantly associated with CIP-28.
Specific inhibitory activity toward testicular CEH was much lower and saturated at a lower inhibitor concentration
b e f o r e b o i l i n g a f t e r b o i l i n g
(5 ~g protein) (1 ~g protein)
FIG. 9. SDS-polyacrylamide electrophoresis of cation exchange puri- fied CEH inhibitor, showing bands used for immunization and electroe- lution of purified proteins. Cation exchange fractions 21-24 were pooled and electrophoresed on a 9-12% SDS-polyacrylamide gradient gel. Gels were stained with Coomassie blue to identify 28, 27, and 26.5 kDa bands for excision. Western blot shows immunoreactivity with an- tibody to 26.5/28 kDa proteins, both before and after boiling of pooled cation exchange fractions. See Figure 1 for other abbreviation.
at 32 than at 37°C, possibly reflecting lower susceptibility of the temperature-labile CEH, which contributes most of the activity at 32°C (39,40). However, this may also reflect the higher concentration of active enzymes at 32°C, whereas only the temperature-stable enzyme is active at 37°C. In support of this view, additional experiments with pancreatic CEH have shown that both the I50 and maximum inhibition (Ima x) decrease as the CEH concentration increases (unpublished data). Inhibition of pancreatic CEH was not different at 32 and 37°C (Fig. IB), ruling out an intrinsic temperature effect on inhibitory activity.
Although inhibitory activity is stable, even after boiling (Figs. 2A and 9), recoveries are unacceptably low in more highly purified preparations, apparently due to aggregation and adherence to columns, filters, and vessels. This problem was partially solved by excising individual bands from poly- acrylamide gels and electroblots for immunization and amino acid analysis, respectively. Although this property prevented most studies with the pure protein in solution under condi- tions appropriate for CEH activity, it is consistent with a role in regulating the activity of an enzyme which acts at the hy- drophilic/hydrophobic interface.
Inasmuch as C1P-28 is hydrophobic and loses its solubility as its homogeneity increases during purification, this protein is apparently stabilized in the native state by the presence of other cellular components, perhaps by association with other cytosolic proteins and with the substrate itself. This is consis-
Lipids, Vol. 31, no. 12 (1996)

CHOLESTERYL ESTER HYDROLASE INHIBITORY PROTEIN 1241
TABLE 2 Amino Acid Compositions (moi/moi protein) of CEH Inhibitory Proteins and Other Proteins of Similar Molecular Weight
Estrogen- Rat serum regulated
Amino Rat testicular proteins Apo-AI protein PLAP
acid 28 kDa 26.5 kDa 27 kDa (27 kDa) a (28 kDa) b (28kDa) c
Asx d 17.1 18.4 30.0 31 16.2 22 GIx e 18.8 24.8 34,2 40 30.7 23 Ser 27.3 19.1 13.9 8 22.6 36 His 1.9 22.0 5.0 6 6.3 11 Gly 23.0 5.8 13.1 10 20,5 20 Thr 14.0 10.7 1t.1 t l 15,8 32 Ala 34.5 29.2 25.8 17 22.1 19 Arg 18.3 10.5 10,3 14 21.4 16 Tyr 3.2 3.1 2.3 5 4,0 7 Val 15.7 14.4 19.2 13 13.9 18 Met 3.0 ] .8 0.0 8 2.3 7 Phe 5.3 6.9 9.8 6 9.2 10 fie 6.2 7.6 10.4 4 9.6 26 Leu 23.1 20.2 3t .3 28 22.9 32 Lys 36.9 21.6 19.4 21 11.4 13 Pro 16.8 19.8 18.8 7 22.9 12 Trp ND ND ND 6 ND 7 Cys ND ND ND 0 1.4 14
aAmino acid composition translated from cDNA sequence (Ref. 33). t~Amino acid composition of MCF-7 Estrogen-regulated breast cancer pro- tein (37). Monoclonal antibody to this protein recognizes a 28 kDa cytosolic estrogen-regulated rat Leydig cell protein postulated to be responsible for the early steroidogenic lesion occurring during gonadotropin desensitization (Refs. 35,36). CAm]no acid composition translated from cDNA sequence of phospholipase A2-activating protein from BC3H 1 murine smooth muscle cells (Ref. 34). d,~sp +Asn residues. eGlu + Gin residues; PLAP, phospholipase-activating protein. See Table 1 for other abbreviation.
tent with a role in regulation of CEH by reversible associa- tion with the substrate surface. Indeed, a substantial fraction of this protein is recovered from the floating fat layer after differential centrifugation (data not shown). This interaction may alter CEH activity by occluding the surface or displac- ing bound CEH. Although this mechanism lacks the speci- ficity of classical competitive inhibition, it is consistent with the broad substrate specific]ties of hydrolytic enzymes and may play an important rote in their regulation. Evidence will be presented elsewhere that this protein does in fact inhibit CEH by this mechanism.
CIP-28 can be distinguished from other reported CEH in- hibitors, which are heat-labile and soluble in aqueous solu- tion at pH 5-5.2 (13-16,43). The monomeric molecular weight of CIP-28 is much lower than those reported for the other inhibitors (typically, =50 kDa). Like the other inhibitors, CIP-28 is highly hydrophobic and elutes diffusely from gel permeation columns. CIP-28 shares similar properties with apolipoprotein A-I (43), including heat stability, molecular weight, and ability to inhibit CEH activity in vitro. However, CIP-28 and apoA-I have different amino acid compositions (Table 2), and polyclonal antibodies to CIP-28 do not recog- nize rat apoA-I on western blots (data not shown). CIP-28 also exhibited important differences in amino acid composi-
immune pre-immune
28 kDa
26~ kDa
97.4 66.2
42.7
31
21.5
14.4
1 2 3 1 2 3
FIG. I0. Specific]ties of polyclonal antibodies raised to 28 kDa and 26.5 kDa proteins combined (lanes 1 ), the 26.5 kDa protein alone (lanes 2) or the 28 kDa protein alone (lanes 3). Each lane contains 5 pg cation exchange purified protein (fractions 2]-24) electrophoresed on a 9-12% SDS-polyacrylamide gradient gel, transbiotted to polyvinyli- dene difluoride membrane and probed with equivalent amounts of im- mune or pre-immune IgG. Specific 28 and 26.5 kDa immunoreactive bands are located relative to BioRad (Melville, NY) molecular weight markers.
tion from other surface-active proteins, including PLAP (34) and estrogen-regulated protein (35,36). Thus, CIP-28 appears to be a unique protein.
The physical properties of CIP-28 which hinder recovery also make it difficult to determine the total cellular content. However, a minimum testicular content can be estimated from the amino acid analysis. Correcting for dilution factors, the lower limiting concentrations of the 28, 26.5, and 27 kDa proteins are 8.1 x 10 -9, 2.6 x 10 -9, and 2.2 x 10 -9 mols/L, re- spectively. Although losses during purification would dictate large positive correction factors to obtain accurate values, the calculated values are comparable to minimum effective in- hibitor concentrations seen in these studies ( - 10 -9 M; see Fig. 5C). Moreover, the activation of CEH seen at very low con- centrations (see Fig. 5), which has been reproduced with elec- troeluted CIP-28 (not shown), suggests that this protein may also be a positive modulator of CEH at concentrations too low to produce inhibition. Clarification of the physiological role of CIP-28 would require knowledge of localized concentra- tions and interactions with other cellular components. Never- theless, it is clear that this protein is present in the testis at sufficient levels to be a potentially important regulator of CEH activity.
A C K N O W L E D G M E N T
This work was supported by NIH grant DK44613.
Lipids, Vol. 31, no. 12 (1996)
1242 D.S. HINES ETAL.
80
O
° ~
60
40
20
A I I E x i t . 1
none immune IgG preimmune IgG
Pretreatment
o . M
* N d~ ° N
80
60
40
20
0
0
B T
- t - I ig :Une
-ZX- PgGmmune ~
-+ - B e a d contro l (no l g G )
1 2
IgG added (rag)
FIG. 11. Concentration-dependent immunoprecipitation of CEH in- hibitory activity by immune IgG complexed with protein A conjugated polyacrylic beads. Pancreatic CEH was assayed with addition of equiv- alent volumes of untreated cation or anion exchange purified inhibitor; supernatant of cation or anion exchange purified inhibitor after precipi- tation with immune IgG; supernatant of cation or anion exchange puri- fied inhibitor after precipitation with preimmune IgG; or supernatant of cation or anion exchange purified inhibitor after precipitation with pro- tein A coniugated beads alone. (A) Experiment I : complexes of 2.6 mg preimmune or immune IgG with 22 mg protein A beads were incubated with 2 lag anion exchange purified protein. Experiment 2: complexes of 3 mg preimmune or immune [gG with 25 mg protein A beads were in- cubated with 3 lag anion exchange purified protein. (B) Inhibitory activ- ity in 8 lag cation exchange purified protein was precipitated by increas- ing amounts of lgG, as indicated. Values are mean percentage inhibi- tion _+ SEM (n = 3). Error bars not seen are contained within bars or symbols. See Figure 1 for other abbreviation.
REFERENCES
1. Momsen, W.E., and Brockman, H.U (1976) Effects of Colipase and Taurodeoxycholate on the Catalytic and Physical Properties of Pancreatic Lipase B at an Oil-Water Interface, J. BioL Chem. 251,378-383.
2. Verger, R., Rietsch, J., and Desnuelle, P. (1977) Effects of Coli- pase on Hydrolysis of Monomolecular Films by Lipase, J. BioL Chem. 252, 4319-4325.
3. Bengtsson, G., and Olivecrona, T. (1980) Lipoprotein Lipase: Some Effects of Activator Proteins, Eur. J. Biochem. 106, 549-555.
4. Posner, I., and DeSanctis, J. (1987) Kinetics of Product Inhibi-
¢ NEPHGE
28 kDa 26.5 kDa
7.0 6.0 5.0 4.0 pl
FIG. 12. Western immunob}ot from two-dimensional gel electrophore- sis of cation exchange purified inhibitor showing mobility of 28 and 26.5 kDa proteins. 250 lag pooled cation exchange fractions 21-28 was separated in the first dimension by nonequilibrium pH gel electrophore- sis and in the second dimension on a 10% SDS-polyacrylamide gel, transblotted to polyvinylidene difluoride membrane and probed with 0.8 mg immune tgG. Molecular weight and pl were determined by com- parison to two-dimensional molecular weight markers (pl 3,8-7.6; 17-89 kDa) blotted to polyvinylidene difluoride membrane and stained with Coomassie blue.
tion and Mechanisms of Lipoprotein Lipase Activation by Apolipoprotein C-II, Biochem. 26, 3711-37 i 7.
5. Yokoyama, S., Fukushima, D., Kupferberg, J.P., Kezdy, F.J., and Kaiser, E.T. (1980) The Mechanism of Activation of Lecithin:Cholesterol Acyltransferase by Apolipoprotein A-I and an Amphilphilic Peptide, J. Biol. Chem. 255, 7333-7339.
6. Jonas, A., and McHugh, H.T. (1984) Reaction of Lecithin:Cho- lesterol Acyltransferase with Micellar Substrates, Biochim. Bio- phys. Acta 794, 361-372.
7. Vanloo, B., Taveirne, J., Baert, J., Lorent, G., Lins, L., Ruyschaert, J.M., and Rosseneu, M. (1992) LCAT Activation Properties of Apo A-I CNBr Fragments and Conversion of Dis- coidal Complexes into Spherical Particles, Biochim. Biophys. Acta 1128, 258-266.
8. Sessa, G., Freer, J.H., Colacicco, G., and Weissman, G. (1969) Interaction of a Lytic Polypeptide Mellitin, with Lipid Mem- brane Systems, J. BioL Chem. 244, 3575-3582.
9. Kinnunen, P.K.J., Koiv, A., Lehtonen, J.Y.A., Ryt6maa, M., and Mustonen, P. (1994) Lipid Dynamics and Peripheral Interac- tions of Proteins with Membrane Surfaces, Chem. Phys. Lipids 73, 181-207.
10. Gargouri, Y., Pieroni, G., Ferrato, F., and Verger, R. (1987) A Kinetic Study with Dicaprin Monolayers, Eur. J. Biochem. 169, 125-129.
11. Davidson, F.F., Dennis, E.A., Powell, M., and Glenney, J.R., Jr. (1987) Inhibition of Phospholipase A2 by "Lipocortins" and Calpactins. An Effect of Binding to Substrate Phospbolipids, J. Biol. Chem. 262, 1698-1705.
12. Gorin, E., Gonen, H., and Dickbuch, S. (1982) A Serum Protein Inhibitor of Acid Lipase and Its Possible Rote in Lipid Accumu- lation in Cultured Fibroblasts, Biochem. J. 204, 221-227.
13. Neiderheiser, D.H. (1982) Cholesteryl Ester Hydrolase in Gall- bladder Mucosa: Evidence for the Presence of an Inhibitor Pro- tein in the Cytosol, Biochem. Biophys. Res. Comm. 105, 328-333.
Lipids, Vol. 31, no. 12 (1996)

CHOLESTERYE ESTER HYDROLASE INHIBITORY PROTEIN 1243
14. Tanaka, M., Yonekura, R., lio, T., and Tabata, T. (1984) Char- aderization of a Cytosolic Protein Inhibiting Lysosomal Acid Cholesteryl Ester Hydrolase, LipMs 19. 714--720.
15. Shand, J.H., and West, D.W. (1992)Characterization of a Cy- tosolic Protein in Rat Liver Inhibiting Neutral Cholesteryl Ester Hyd olase. Lipids 27, 406-4 12.
16. Shand..I.H., and West, D.W. (1993) Inhibition of Neutral Cho- lesteryl Ester Hydrolase by a Cytosolic Protein in Adipose Tis- sue. Biochem. Soc. Trans. 21,457S.
17. Shand, J.H.. Crilly, P.J., and West, D.W. (1993) Inhibition of yeulral Cholesteryl Ester Hydrolase by a Naturally Occurring Cytosolic Protein in Macrophages, FEBS Lett. 321, 132-134.
18. Shand. J.H., West, D.W., McCartney, R.J., Noble, R.C., and Speakc. B.K. (1993) The Esterification of Cholesterol in the Yolk Sac Membrane of the Chick Embryo, Lipids 28, 621 625.
19. Wee, S.F, and Grogan, W.M. (1993)Testicular Temperature- Labile Cholesteryl Ester Hydrolase. Relationship to Isoenzymes from ()ther Tissues, CoJTelation with Spermatogenesis and Inhi- bition by Physiological Concentrations of Divalent Cations, J. BioL Chem. 268, 8158-8163.
20. Ghosh. S., and Grogan, W. M. ( 1991 ) Rapid Three-Step Purifi cation of a Hepatic Neutral Cholesteryl Ester Hydrolase Which ls Not the Pancreatic Enzyme, Lipids 26, 793-798.
2t. Smith. P.K.. Krohn, R.I., Hermanson. G.T., Mallia, A.K., Oart- her, F.H., Provenzano, M.D.. Fujimoto, E.K., Goeke, N.M., OL, on, B.J., and Klenk. D.C. (1985)Measurement of Protein Using Bicmchoninic Acid, Anal Biochem. 150, 76-85.
22. Laemmli. U.K. (1970) Cleavage of Structural Proteins During the Asseinbly of the Head of Bacteriophage T4, Nature 227, 680-685.
23. Ghosh. S.. and Grogan, W.M. (1989) Activation of Rat Liver Cholesterol Ester Hydrolase by cAMP-Dependent Protein Ki- nasc and Protein Kinase C, Lipids 24, 733-736.
24. Momsen. W.E.. and Brockman. H.L. (1977) Purification and Characterization of Cholesten~l Esterase from Porcine Pancreas, Biochim. l~iophvs. Acre 4&'6, 103-113.
25. Lee. C., Levin. A.. and Branton, I). (1987) Copper Staining: A Five-Minute Protein Stain for Sodium Dodecyl Sulfate-Poly- acrvlamide Gels, Attal. Biochem. 166, 308-312.
26. Gaff in, D.E. (1990) One-Dimensional Gel Electropboresis, Meth. E1~:wmfl. 182, 425-441.
27+ Plocke. D J., Levinthal, C.. and Vallee, B.L. (1962) Alkaline Phosphatase of Escherichia colt: A Zinc Metallo Enzyme, Biochem. 1. 373-378.
28. Carter, R.J., and Boyd, N.D. (1979) A Comparison of Methods for Obtaining High Yields of Pure hnmunoglobulin from Se- ¥e.rely Haemolysed Plasma. J. lmmm+ot. Methods 26, 213-222.
29. Ghosh. S.. and Grogan, W. M. (1992) Immunological Charac- terization of Neutral Cholesteryl Ester Hydrolase from Rat Liver Cytosol. Biochem. Cell Biol. 70, 800-803.
30. Kessler, S.W. (1975) Rapid Isolation of Antigens from Cells with a Staphylococcal Protein A-Antibody Adsorbent: Parame- ters of the Interaction of Antibody-Antigen Complexes with Protein A, J. lmmunol. 115, 1617-1623.
31. Kessler, S.W. ( 1981 ) Use of Protein-A Bearing Staphylococci for the Immunoprecipitation and Isolation of Antigens from Cells, Meth. Enzymol. 73, 442-458.
32. Matsudaria, P. (1987) Sequence from Picomole Quantities of Proteins Electroblotted onto Polyvinylidene Difluoride Mem- branes, J. Biol. Chem. 262, 10035-10038.
33. Poncin, J.E., Martial, J.A., and Gielen, J.E. (1984) Cloning and Structure Analysis of the Rat Apolipoprotein A I eDNA, Eur. J. Biochem. 140, 493-498.
34. Clark, M.A., Ozgiir, L.E., Conway, T.M., Dispoto, J., Crooke, S.T. and Bomalaski, J.S. (1991 ) Cloning of a Phospholipase A2- Activating Protein, Proc. Natl. Acad. Sci. 88, 5418-5422.
35. Ciocca, D.R., and Dufau, M.L (1984) Estrogen-dependent Ley- dig Cell Protein Recognized by Monoclonal Antibodies to NCF- 7 Cell Line, Science 226, 445~-46.
36. Ciocca, D.R., Winters, C.A., and Dufau. M.L. (1986) Expres- sion of an Estrogen-Regulated Protein in Rat Testis Leydig Ceils, J. Steroid Biochem. 24, 219-229.
37. Adams, D.J., Hajj, H., Bitar, K.G., Edwards, D.P., and McGuire, W.L. (1983) Purification of an Estrogen-Regnlated Breast Can- cer Protein by Monoclonal Antibody Affinity Chromatography, Endocrinol. 113, 415-4 17.
38. O'Farrell, P.Z., Goodman, H.M., and O'Farrell, P.H. (1977) High-Resolution Two-Dimensional Electrophoresis of Proteins, Cell I2, 1133-1142.
39. Durham, L.A., and Grogan, W.M. (1982) Temperature Sensitiv- ity of Cholesteryl Ester Hydrolases in the Rat Testis, LipMs 17, 970-975.
40. Durham, L.A., and Grogan, W.M. (1984) Characterization of Multiple Forms of Cholesteryl Ester Hydrolase in the Rat Testis, J. Biol. Chem. 259, 7433-7438.
41. Wee, S.F., and Grogan, W.M. (1989) Temperature Lability and cAMP-Dependent Protein Kinase Activation of Cholesteryl Ester Hydrolase as a Function of Age in Developing Rat Testis, Lipids 24, 824-828.
42. Bailey, M.L., and Grogan, W.M. (1986) Protein Kinase-Medi ated Activation of Temperature-Labile and Temperature-Stable Cholesteryl Ester Hydrolases in the Rat Testis, J. Biol. Chem. 261, 7717-7722.
43. Tanaka. M., Iio, T., and Tabata, T. (1990) Properties of an Acid Cholesteryl Ester Hydrolase Inhibitor from Rat Serum, Lipids 25, 775-778.
[Received July 12, 1996, and in final revised form October 11, 1996; Revision accepted October 22, 1996]
Lipids, Vol. 31, no. 12 i1996;
Related Documents











